
Amyloid Peptides Decrease Soluble Guanylyl Cyclase Expression in Astroglial Cells
11
-Amyloid Peptides Decrease Soluble Guanylyl Cyclase Expression in Astroglial Cells Marı´a Antonia Baltrons,* Carlos E. Pedraza,* Michael T. Heneka, † and Agustina Garcı´a* *Instituto de Biotecnologı ´a y Biomedicina V. Villar Palasi and Departamento de Bioquı ´mica Biologı ´a Molecular, Universidad Auto ´noma de Barcelona, 08193 Bellaterra, Spain; and † Department of Neurology, University of Bonn, 53105 Bonn, Germany Received June 21, 2001; revised March 7, 2002; accepted for publication March 28, 2002 In astroglial cells -amyloid peptides (A) induce a reactive phenotype and increase expression of NO synthase. Here we show that treatment of rat brain astrocytes with A decreases their capacity to accumulate cyclic GMP (cGMP) in response to NO as a result of a decreased expression of soluble guanylyl cyclase (sGC) at the protein and mRNA levels. Potentiation of A-induced NO formation by interferon- did not result in a larger decrease in cGMP formation and inhibition of NO synthase failed to reverse down-regulation of sGC, indicating that NO is not involved. The A effect was prevented by the protein synthesis inhibitor cycloheximide. Intracerebral A injection also decreased sGC 1 subunit mRNA levels in adult rat hippocampus and cerebellum. A loss of sGC in reactive astrocytes surrounding -amyloid plaques could be a mechanism to prevent excess signalling via cGMP at sites of high NO production. © 2002 Elsevier Science (USA) Key Words: astroglial; -amyloid peptides; Cyclic GMP; soluble guanylyl cyclase; nitric oxide. INTRODUCTION Reactive astroglial and microglial cells are key cel- lular elements in the neuroinflammation observed in Alzheimer’s disease (AD) and other neurodegenera- tive conditions (Eddleston and Mucke, 1993; Gonza ´- lez-Scarano and Baltuch, 1999). In AD brains, reactive astrocytes are associated with amyloid plaques at early stages of the disease (Pike et al., 1995) and have been shown to express nitric oxide synthase type 2 (NOS-2) (Wallace et al., 1997). The highly reactive de- rivative of NO peroxynitrite is formed at these sites and appears to be an important source of oxidative stress leading to neuronal death (Smith et al., 1997; Pappolla et al., 1998). -Amyloid peptides (A) are capable of activating rodent glial cells in culture leading to induction of NOS-2 expression in both astroglia and microglia and release of inflammatory cytokines mostly from micro- glia (Pike et al., 1994; Meda et al., 1995; Goodwin et al., 1995; Rossi et al., 1996; Hu et al., 1998). In rat cortical glial cultures, microglial derived interleukin-1 (IL- 1) appears to be responsible for NOS-2 induction in astrocytes (Akama and Van Eldik, 2000). In the normal CNS moderate amounts of NO are generated in neurones and in astrocytes by constitu- tive nitric oxide synthases (NOS-1 and NOS-3) stimu- lated by neurotransmitters that increase intracellular Ca 2 (reviews in Zhang and Synder, 1995; Fo ¨ rster- mann et al., 1998; Baltrons and Garcı ´a, 2001). Soluble guanylyl cyclase (sGC; GTP pyrophosphate-lyase (cy- clizing), EC 4.6.1.2) is the major physiological target for NO and cyclic GMP (cGMP) the mediator of im- portant neuromodulatory actions of NO in synaptic plasticity, brain development, neuroendocrine secre- tion, etc. (reviews in Garthwaite and Boulton, 1995; Zhang and Synder, 1995). Very little is known about the regulation of cGMP formation under neuropatho- logical conditions associated with chronic inflamma- tion and excess NO production by NOS-2 in activated glial cells and/or infiltrated macrophages. We have recently demonstrated that long-term treatment of rat brain astroglial cells with lipopolysaccharide (LPS), condition that induces astroglial reactivity and NOS-2 Neurobiology of Disease 10, 139 –149 (2002) doi:10.1006/nbdi.2002.0492 0969-9961/02 $35.00 © 2002 Elsevier Science (USA) All rights reserved. 139
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Amyloid Peptides Decrease Soluble Guanylyl Cyclase Expression in Astroglial Cells

Neurobiology of Disease 10, 139–149 (2002)doi:10.1006/nbdi.2002.0492
�-Amyloid Peptides Decrease Soluble GuanylylCyclase Expression in Astroglial Cells
Marıa Antonia Baltrons,* Carlos E. Pedraza,*Michael T. Heneka,† and Agustina Garcıa**Instituto de Biotecnologıa y Biomedicina V. Villar Palasi and Departamento de BioquımicaBiologıa Molecular, Universidad Autonoma de Barcelona, 08193 Bellaterra, Spain; and†Department of Neurology, University of Bonn, 53105 Bonn, Germany
Received June 21, 2001; revised March 7, 2002; accepted for publication March 28, 2002
In astroglial cells �-amyloid peptides (�A) induce a reactive phenotype and increase expression of NOsynthase. Here we show that treatment of rat brain astrocytes with �A decreases their capacity toaccumulate cyclic GMP (cGMP) in response to NO as a result of a decreased expression of solubleguanylyl cyclase (sGC) at the protein and mRNA levels. Potentiation of �A-induced NO formation byinterferon-� did not result in a larger decrease in cGMP formation and inhibition of NO synthase failedto reverse down-regulation of sGC, indicating that NO is not involved. The �A effect was prevented bythe protein synthesis inhibitor cycloheximide. Intracerebral �A injection also decreased sGC �1subunit mRNA levels in adult rat hippocampus and cerebellum. A loss of sGC in reactive astrocytessurrounding �-amyloid plaques could be a mechanism to prevent excess signalling via cGMP at sitesof high NO production. © 2002 Elsevier Science (USA)
Key Words: astroglial; �-amyloid peptides; Cyclic GMP; soluble guanylyl cyclase; nitric oxide.
INTRODUCTION
Reactive astroglial and microglial cells are key cel-lular elements in the neuroinflammation observed inAlzheimer’s disease (AD) and other neurodegenera-tive conditions (Eddleston and Mucke, 1993; Gonza-lez-Scarano and Baltuch, 1999). In AD brains, reactiveastrocytes are associated with amyloid plaques atearly stages of the disease (Pike et al., 1995) and havebeen shown to express nitric oxide synthase type 2(NOS-2) (Wallace et al., 1997). The highly reactive de-rivative of NO peroxynitrite is formed at these sitesand appears to be an important source of oxidativestress leading to neuronal death (Smith et al., 1997;Pappolla et al., 1998).
�-Amyloid peptides (�A) are capable of activatingrodent glial cells in culture leading to induction ofNOS-2 expression in both astroglia and microglia andrelease of inflammatory cytokines mostly from micro-glia (Pike et al., 1994; Meda et al., 1995; Goodwin et al.,1995; Rossi et al., 1996; Hu et al., 1998). In rat corticalglial cultures, microglial derived interleukin-1� (IL-
0969-9961/02 $35.00© 2002 Elsevier Science (USA)All rights reserved. 139
1�) appears to be responsible for NOS-2 induction inastrocytes (Akama and Van Eldik, 2000).
In the normal CNS moderate amounts of NO aregenerated in neurones and in astrocytes by constitu-tive nitric oxide synthases (NOS-1 and NOS-3) stimu-lated by neurotransmitters that increase intracellularCa2� (reviews in Zhang and Synder, 1995; Forster-mann et al., 1998; Baltrons and Garcıa, 2001). Solubleguanylyl cyclase (sGC; GTP pyrophosphate-lyase (cy-clizing), EC 4.6.1.2) is the major physiological targetfor NO and cyclic GMP (cGMP) the mediator of im-portant neuromodulatory actions of NO in synapticplasticity, brain development, neuroendocrine secre-tion, etc. (reviews in Garthwaite and Boulton, 1995;Zhang and Synder, 1995). Very little is known aboutthe regulation of cGMP formation under neuropatho-logical conditions associated with chronic inflamma-tion and excess NO production by NOS-2 in activatedglial cells and/or infiltrated macrophages. We haverecently demonstrated that long-term treatment of ratbrain astroglial cells with lipopolysaccharide (LPS),condition that induces astroglial reactivity and NOS-2

expression, down-regulates sGC (Baltrons and Garcıa,1999). Chronic infusion of LPS in the brain of youngrats has been reported to reproduce several features ofAD, such as chronic neuroinflammation associatedwith glial activation and increased expression of IL-1,tumor necrosis factor-� and �-amyloid precursor pro-tein (Hauss-Wegrzyniak et al., 1998). Considering thisand the reduction in sGC activity reported in brainextracts from subjects with AD (Bonkale et al., 1995),we investigated the effect of �A in NO-dependentcGMP formation in neural cells. Here we show for thefirst time that �A decrease the capacity of rat brainastroglial cells to generate cGMP by decreasing theexpression of sGC at the protein and mRNA levels bya mechanism independent of NO. The decrease in sGCmRNAs is also observed in brain tissue after focalinjection of the peptides.
EXPERIMENTAL PROCEDURES
Materials
Dulbecco’s modified Eagle’s medium (DMEM) wasobtained from Flow Laboratories and foetal calf serum(FCS) from Biowhittaker; [3H]cGMP (34.5 Ci/mmol)from New England Nuclear; Dowex 50W-X4 (200–400mesh, H� form) from Bio-Rad; benzamidine fromMerck; sodium nitroprusside (SNP), GTP, 3-isobutyl-1-methylxanthine (IBMX), cicloheximide, NG-monometh-yl-l-arginine (l-NMMA), creatine kinase, creatine phos-phate, trypsin inhibitor, phenylmethylsulfonyl fluoride(PMSF), pepstatin A, leupeptin, dithiothreitol (DTT),snake venom (Crotalus atrox) and urea from Sigma. Pen-tobarbital was purchased from Byk Gulden (Konstanz,Germany). TRIzol reagent, Superscript-II reverse tran-scriptase, taq DNA polymerase, forward and reverseprimers and deoxinucleotides (dNTPs), phosphate buff-ered saline (PBS) were obtained from Gibco-BRL. Re-combinant rat interferon-� (INF-�) was from PeproTech;rabbit polyclonal antibody to glial fibrillary acidic pro-tein (anti-GFAP) from Dako; polyclonal antibody againstthe �1 subunit of sGC was from Cayman Chemical;horseradish peroxidase coupled to rabbit IgG from Am-ersham Life Science; Supersignal Ultra Chemiluminines-cent Substrate from Pierce. Anti-cGMP antiserum wasprepared in our laboratory as described by Brooker et al.(1979).
�A were obtained from Neosystem (Strasbourg,France), American Peptide Company (CA) and Sigma(MO). For stock solutions �A(1–40) was dissolved at
500 �M and �A(25–35) and SCR �A(25–35) at 2.5 mMin sterile double-deionized water and were stored at�80°C until use. Before use �A(1–40) was aged byincubation for 1 week a 37°C. Stock solutions werediluted to the desired concentration and added toculture medium containing serum. To examine thestructure of the peptides, 10 �l 500 �M of the peptidesolutions were applied to a 400-mesh carbon coatedgrids. After 2 min excess fluid was blotted, grids air-dried and the samples were stained with 1% (w/v)phosphotungstic acid. Images were acquired on a Hi-tachi H-7000 (Fig. 1).
Cell Cultures
Astrocyte-enriched primary cultures were preparedfrom newborn (cortex and hippocampus) and 7-day-old (cerebellum) Sprague–Dawley rats as described(Agullo et al., 1995). Dissociated cells suspended inDMEM-10 % FCS, 20 U/ml penicillin and 20 �g/mlstreptomycin were seeded in 35-, 60-, or 100-mm di-ameter plastic petri dishes (1.25 � 105 cells/cm2). Me-dium was changed once a week and cells were usedafter 14 days in culture. As previously reported(Agullo et al., 1995), the majority of cells in the cultureswere glial fibrillary acidic protein positive and themain contaminating cell type was microglia. To de-crease microglial contamination, in some experimentscultures of cerebellar astroglia were treated with 10�M cytosine arabinoside from day 10 until the day ofuse (14th). This treatment decreased microglial con-tamination to around 2% as judged by the histochem-ical reaction for nucleoside diphosphatase (Agullo etal., 1995). Cell viability of treated cells was assessed bynigrosine exclusion and by lactate dehydrogenase(LDH) release to the medium (Klegeris and McGeer,1997). Cell morphology was examined by immuno-staining with a rabbit polyclonal antibody to glialfibrillary acidic protein (anti-GFAP; 1:300 dilution,Dako) (Agullo et al., 1995).
cGMP and Nitrite Accumulation
For determination of cGMP, cell monolayers pre-treated with different agents, were washed and prein-cubated for 10 min at 37°C in Hepes-buffered saline(118 mM NaCl, 4.7 mM KCl, 2.5 mM CaCl2, 1.2 mMMgSO4, 1.2 mM KH2PO4, 10 mM glucose and 20 mMHepes, pH 7.4) before adding 1 mM 3-isobutyl-1-methylxanthine (IBMX) alone or with sodium nitro-prusside (SNP, 100 �M). After a 3-min incubation,reactions were terminated by aspirating the medium
140 Baltrons et al.
© 2002 Elsevier Science (USA)All rights reserved.

and adding ice-cold ethanol. Ethanol extracts wereevaporated, resuspended in 5 mM sodium acetate pH4.8 and cGMP quantified by radioimmunoassay usingacetylated [3H]cGMP (Agullo and Garcıa, 1992). Ni-trite accumulation was assayed in the media by theGreiss reaction as described (Baltrons and Garcıa,1999).
Enzyme Assays
sGC and phosphodiesterase (PDE) activities weredetermined in cytosolic fractions as described (Bal-trons and Garcıa, 1999). Briefly, cultured cells werehomogenised with a glass-Teflon Potter-Elvehjeim ho-mogenizer (20 hand strokes) in ice-cold 50 mM Tris/HCl buffer (pH 7.4 at 37°C), containing 1 mM dithio-threitol (DTT), 1 mM EDTA, 0.2 mM benzamidine, 10mg/l trypsin inhibitor, 100 mg/l phenylmethylsulfo-
nyl fluoride (PMSF), 1 mg/l pepstatin A and 0.1 mg/lleupeptin. Homogenates were centrifuged at 100,000gfor 1 h at 4°C and supernatants stored at �80°C. ForsGC activity supernatant fractions (30–40 �g of pro-tein) were incubated for 10 min at 37°C with 1 mMGTP, 1 mM DTT, 4 mM MgCl2, 15 mM creatine phos-phate, 200 mg/l creatine kinase, 1 mM IBMX, 1 mMEGTA in 50 mM Tris/HCl, in the presence or absenceof 100 �M SNP. The cGMP product was determinedby radioimmunoassay as described above. For PDEactivity supernatant fractions (15–20 �g of protein)were incubated for 5–30 min at 37°C in 50 mM Tris/HCl (pH 7.4) with 10 �M [3H]cGMP (0.075 �Ci), 1 mMMgCl2, 0.5 mM EGTA in the absence (less than 1 nM)or presence of 25 �M free Ca2� concentration. Afterterminating the reaction by boiling, samples were in-cubated with 50 �g of snake venom (Crotalux atrox) totransform [3H]GMP into [3H]guanosine and then ap-
FIG. 1. Morphology of �A peptides examined by electron microscopy. �A (1–40) nonaged (A) or aged (B), �A (25–35) (C), and SCR �A(25–35)(D) were applied (10 �l) to 400-mesh carbon coated grids. After drying, the samples were stained with 1% (w/v) phosphotungstic acid.Negative stain electron microscopy shows the presence of fibrillary aggregates in the aged but not in the non-aged peptide �A (1–40).�A(25–35) and SCR �A(25–35) aggregate into shorter fibrils. Bars, 100 nm.
141�-Amyloids Decrease Soluble Guanylyl Cyclase
© 2002 Elsevier Science (USA)All rights reserved.

plied to cation exchange resin columns (Dowex-X4,200–400 mesh). [3H]Guanosine was eluted with 3 Mammonium hydroxide.
Western Blotting
Western blots were performed as described (Bal-trons and Garcıa, 1999). Cytosolic fractions (20–40 �gof protein) were subjected to 7.5% sodium dodecylsulfate–polyacrylamide gel electrophoresis (SDS–PAGE) followed by transfer to nitrocellulose filters at100 V for 1.5 h at 4°C. Membranes were blocked at 4°Covernight in phosphate-buffered saline (pH 7.4) con-taining 0.05% Tween 20 and 5% nonfat dry milk, andthen incubated at room temperature for 2 h with apolyclonal anti-serum (1:1000) that preferentially rec-ognizes the �1 subunit of sGC. Bound antibody wasdetected with a horseradish peroxidase conjugatedanti-rabbit IgG (1:4000) visualised by chemiluminis-cence.
Reverse Transcription-Polymerase ChainReaction (RT-PCR) Analysis
Total RNA from astrocyte primary cultures wereextracted with TRIzol reagent and quantified sprecto-photometrically. Integrity of RNAs was checked byethidium bromide staining in agarose gels (1% aga-rose). Total RNA (1 �g) was reverse transcribed (RT)into cDNA using 1 U of Superscript-II reverse tran-scriptase and 3 �g random hexamer primers in a finalreaction volume of 20 �l. Amplification was per-formed in a 50-�l reaction volume containing 10 mMTris–HCl, pH 8.3, 50 mM KCl, 1.75 mM MgCl2, 200�M each dNTPs, 0.5 U of Taq DNA polymerase, 0.4�M of each primer and 5 �l of RT product. Primersequences for sGC �1 subunit were: forward, 5�-ATAC-GGGTGAGGAGATGGGATAACT-3�; reverse, 5�-CCT-CATTGAACTTCTTGGCTTGC-3�; PCR reaction per-formed at 36 cycles of amplification (95°C for 30 s; 59°Cfor 25 s, 72°C for 25 s). Primer sequences for sGC �1
subunit were: forward, 5�-GCCAAAAGATACGACA-ATGTGACCA-3� and reverse, 5�-TTGCCATCTACTT-GAACTTGACCAG-3�; PCR was performed at 34 cy-cles of amplification (95°C for 30 s; 61°C for 25 s; 72°Cfor 25 s). Amplification of GAPDH was used to controlfor differences in cDNA synthesis efficiency betweensamples. Primers for GAPDH were: forward, 5�-GC-CAAGTATGATGACATCAAGAAG-3� and reverse,5�-TCCAGGGGTTTCTTACTCCTTGGA-3�; PCR wasperformed at 24 cycles of amplification (95°C for 30 s;at 61°C for 25 s, 72°C for 25 s). PCR products were
analysed by electrophoresis through 2% agarose gelscontaining 0.5 �g/ml ethidium bromide. The amplifi-cation was found to be linear between 33 and 37 cyclesfor sGC �1, 32 and 36 cycles for sGC �1 and 20 to 26cycles for GAPDH.
Animal Experiments
Sprague-Dawley rats (Charles River Laboratories,Wilmington, MA) weighing 250–300 g, were housedin groups of 3 under standard conditions at 22°C anda 12 h light-dark cycle, with free access to food andwater. Rats were anesthetized with pentobarbital (50mg/kg i.p.) and placed in a stereotaxic frame (Stoelt-ing, U.S.A.) on a heating blanket. Body temperaturewas maintained at 37 � 0.5°C for the time of surgery.After exposure of the skull, holes were drilled at therespective injection sites and 2 �l of aggregated �A(1–40) (0.5 �g/�l) in PBS was injected bilaterally over aperiod of 120 s into the hippocampus at anteroposte-rior (AP) �3.0 mm, lateral (L) �/� 2.0 mm and ven-tral (V) 3.0 mm, or into the cerebellar vermis at AP�12,5 mm, L �/� 0.0 mm and V 5.0 mm relative tobregma (Paxinos et al., 1985) using a 2 �l Hamiltonsyringe, as previously reported (Heneka et al., 2000).Controls received injections with 2 �l of PBS. At 24and 48 h, animals were killed by an overdose of pen-tobarbital and brains were removed. All experimentswere carried out in accordance with the declaration ofHelsinski and the animal welfare guidelines and lawsof the Federal Republic of Germany and were ap-proved by the local ethical committee for animal ex-periments.
Protein was determined by the method of Lowry etal. (1951) or with the Bradford reagent (Bradford, 1976)using bovine serum albumin as standard. Experimentswith cultured cells were always performed in tripli-cate plates and were replicated the indicated numberof times in different culture preparations. Statisticalsignificance of differences (P � 0.05) was calculated byStudent’s t test.
RESULTS AND DISCUSSION
Exposure of rat cerebellar astrocyte-enriched cul-tures to aggregated �A(1–40) (20 �M, 72 h) or to thehighly hydrophobic fragment �A(25–35) (50 �M, 48 h)induced a marked morphological change in the cul-tured astrocytes from the typical flat and polygonal-shaped appearance (Figs. 2A and 2C) to a more stellatemorphology (Figs. 2B and 2D). This change has been
142 Baltrons et al.
© 2002 Elsevier Science (USA)All rights reserved.

previously reported for cortical astrocytes (Pike et al.,1994; Hu et al., 1998) and taken as an indication of theacquisition of a reactive phenotype. In contrast, non-aggregated �A(1–40) (not shown) and scrambled(SCR) �A(25–35) (Fig. 2E) did not affect cell shape.Neither peptide affected cell viability as assessed byvital dye exclusion or by measuring LDH release tothe medium (not shown), in agreement with otherworks performed in astrocyte cultures exposed to �Ain medium containing high serum concentration (Sa-linero et al., 1997).
Incubation of astrocyte cultures for 72 h with aggre-gated �A(1–40) had no effect on cGMP accumulationbut decreased the capacity of the cells to generatecGMP in response to the NO donor SNP (100 �M, 3min) in a concentration-dependent manner (Fig. 3).Aggregation of the peptide is necessary to exert thiseffect since the nonaged �A(1–40) (20 �M) did notaffect SNP-stimulated cGMP formation. The effect ofaggregated �A(1–40) was mimicked by �A(25–35) butapparently with lower potency. Whereas �A(1–40) at
FIG. 2. Effect of �A on the morphology of rat cerebellar astrocyte cultures. Cell cultures were incubated in the absence (A, C) or presence of20 �M �A(1–40) for 72 h (B) or 50 �M �A(25–35) (D) or SCR �A(25–35) (E) for 48 h. Cells were fixed and immunostained with polyclonalantibody against GFAP (1:300) as described under Experimental Procedures. Note the change from polygonal (A, C, E) to stellate morphology(B, D). 600�.
FIG. 3. Effect of �A on SNP-stimulated cGMP formation in cere-bellar astrocytes. Cerebellar astrocyte cultures were treated withaggregated and nonaggregated �A (1–40) for 72 h or with �A(25–35) or SCR �A(25–35) for 48 h at the indicated concentrations.After treatment, cells were washed, and cGMP formation in re-sponse to 100 �M SNP (3 min in the presence of 1 mM IBMX) wasmeasured, as indicated under Experimental Procedures. Results,expressed as percentage of the SNP effect in untreated cells (98 � 5pmol/mg protein), are means � SEM of three experiments per-formed in different culture preparations.
143�-Amyloids Decrease Soluble Guanylyl Cyclase
© 2002 Elsevier Science (USA)All rights reserved.
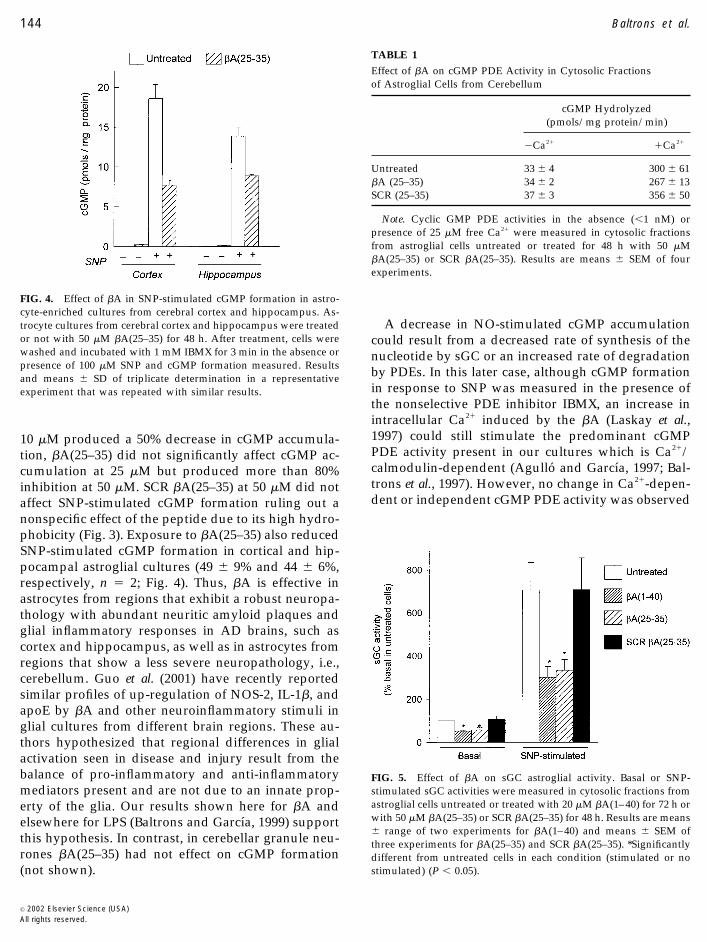
10 �M produced a 50% decrease in cGMP accumula-tion, �A(25–35) did not significantly affect cGMP ac-cumulation at 25 �M but produced more than 80%inhibition at 50 �M. SCR �A(25–35) at 50 �M did notaffect SNP-stimulated cGMP formation ruling out anonspecific effect of the peptide due to its high hydro-phobicity (Fig. 3). Exposure to �A(25–35) also reducedSNP-stimulated cGMP formation in cortical and hip-pocampal astroglial cultures (49 � 9% and 44 � 6%,respectively, n � 2; Fig. 4). Thus, �A is effective inastrocytes from regions that exhibit a robust neuropa-thology with abundant neuritic amyloid plaques andglial inflammatory responses in AD brains, such ascortex and hippocampus, as well as in astrocytes fromregions that show a less severe neuropathology, i.e.,cerebellum. Guo et al. (2001) have recently reportedsimilar profiles of up-regulation of NOS-2, IL-1�, andapoE by �A and other neuroinflammatory stimuli inglial cultures from different brain regions. These au-thors hypothesized that regional differences in glialactivation seen in disease and injury result from thebalance of pro-inflammatory and anti-inflammatorymediators present and are not due to an innate prop-erty of the glia. Our results shown here for �A andelsewhere for LPS (Baltrons and Garcıa, 1999) supportthis hypothesis. In contrast, in cerebellar granule neu-rones �A(25–35) had not effect on cGMP formation(not shown).
A decrease in NO-stimulated cGMP accumulationcould result from a decreased rate of synthesis of thenucleotide by sGC or an increased rate of degradationby PDEs. In this later case, although cGMP formationin response to SNP was measured in the presence ofthe nonselective PDE inhibitor IBMX, an increase inintracellular Ca2� induced by the �A (Laskay et al.,1997) could still stimulate the predominant cGMPPDE activity present in our cultures which is Ca2�/calmodulin-dependent (Agullo and Garcıa, 1997; Bal-trons et al., 1997). However, no change in Ca2�-depen-dent or independent cGMP PDE activity was observed
TABLE 1
Effect of �A on cGMP PDE Activity in Cytosolic Fractionsof Astroglial Cells from Cerebellum
cGMP Hydrolyzed(pmols/mg protein/min)
�Ca2� �Ca2�
Untreated 33 � 4 300 � 61�A (25–35) 34 � 2 267 � 13SCR (25–35) 37 � 3 356 � 50
Note. Cyclic GMP PDE activities in the absence (�1 nM) orpresence of 25 �M free Ca2� were measured in cytosolic fractionsfrom astroglial cells untreated or treated for 48 h with 50 �M�A(25–35) or SCR �A(25–35). Results are means � SEM of fourexperiments.
FIG. 5. Effect of �A on sGC astroglial activity. Basal or SNP-stimulated sGC activities were measured in cytosolic fractions fromastroglial cells untreated or treated with 20 �M �A(1–40) for 72 h orwith 50 �M �A(25–35) or SCR �A(25–35) for 48 h. Results are means� range of two experiments for �A(1–40) and means � SEM ofthree experiments for �A(25–35) and SCR �A(25–35). *Significantlydifferent from untreated cells in each condition (stimulated or nostimulated) (P � 0.05).
FIG. 4. Effect of �A in SNP-stimulated cGMP formation in astro-cyte-enriched cultures from cerebral cortex and hippocampus. As-trocyte cultures from cerebral cortex and hippocampus were treatedor not with 50 �M �A(25–35) for 48 h. After treatment, cells werewashed and incubated with 1 mM IBMX for 3 min in the absence orpresence of 100 �M SNP and cGMP formation measured. Resultsand means � SD of triplicate determination in a representativeexperiment that was repeated with similar results.
144 Baltrons et al.
© 2002 Elsevier Science (USA)All rights reserved.

in cytosolic fractions from �A(25–35)-treated cells (Ta-ble 1). In contrast, basal and SNP-stimulated GC ac-tivity were significantly decreased in cytosolic frac-tions from �A(1–40) and �A(25–35)-treated cells butnot from SCR �A(25–35)-treated cells (Fig. 5). Thedecrease in sGC activity (around 50%) was accompa-nied by a similar decrease in sGC �1 subunit levels asdetermined on immunoblots (Figs. 6A and 6B). Wecould not clearly detect the �1 subunit in the cellsextracts with the antibody used in this study but sincesGC appears to be an obligate heterodimer composedof � and � subunits (Hartenek et al., 1990; Buechler etal., 1991) a lower �1 expression would cause a decreasein functional sGC. Nevertheless, a decrease in �1 is
probably occurring because, as shown by RT-PCR,both �1 and �1 subunit mRNA levels were decreasedin �A-treated cells (Figs. 6C and 6D). Both �1 and �1
subunits have been detected by in situ hybridization inglial elements in the rat cerebellum (Gibb and Garth-waite, 2001). Studies are underway to elucidatewhether the decrease in sGC levels is the result of adecreased transcription or a decreased mRNA and/orprotein stability. Whatever is the mechanism operat-ing it requires protein synthesis. Treatment of the cellswith the protein synthesis inhibitor cicloheximide (20�M, 48 h) that decreases sGC �1 subunit levels by30–40% (not shown), does not significantly decreaseSNP-stimulated cGMP formation on a per mg protein
FIG. 6. �A decrease sGC subunit protein (A, B) and mRNA (C, D) levels in cultured astroglia. Proteins (20–40 �g) from soluble fractions ofastroglial cells untreated or treated with 20 �M �A(1–40) for 72 h or with 50 �M �A(25–35) or 50 �M SCR �A(25–35) for 48 h were fractionatedby SDS–PAGE and transferred to nitrocellulose membranes. Immunoreactive sGC �1 subunit was detected using 1:2000 dilution of acommercial rabbit anti-serum (A). RNA samples of astroglial cells untreated or treated with 20 �M �A(1–40) for 72 h were analysed byRT-PCR, using primers specific for sGC �1 or �1 subunits and GAPDH (C), as described under Experimental Procedures. Data was analyzedby densitometry: (B) sGC �1 subunit protein levels expressed relative to levels in untreated cultures; (D) and sGC �1 or �1 subunits mRNAlevels normalised to GAPDH mRNA levels and expressed relative to the levels in untreated cells. Results are means � SEM of threeindependent experiments.
145�-Amyloids Decrease Soluble Guanylyl Cyclase
© 2002 Elsevier Science (USA)All rights reserved.

basis, but prevents the �A(25–35) inhibitory effect(89 � 20% and 78 � 11% of SNP effect in the absenceand presence of �A, respectively, n � 2). The need forprotein synthesis to induce sGC down-regulation hasbeen also observed by us for the effect of LPS inastrocytes (Baltrons and Garcıa, 1999) and by othersfor the effect of NO donors in smooth muscle cells(Filippov et al., 1997) and of nerve growth factor in ratpheochromocytoma PC12 cells (Liu et al., 1997). How-ever, nothing is known about the role of newly syn-thesised proteins in down-regulating sGC, although arole in sGC mRNA destabilization has been proposed(Filippov et al., 1997). In the case of �A the need forprotein synthesis to observe sGC down-regulationcould arise from the implication of inflammatory cy-tokines synthesised and released from the glial cells inresponse to the peptides (Araujo and Cotman, 1992;Hu et al., 1998; Akama and Van Eldik, 2000). In fact, incortical glial cultures inflammatory cytokines releasedby microglia have been implicated in �A induction ofNOS-2 in astrocytes (Akama and Van Eldik, 2000). Toinvestigate if factors released from contaminating mi-croglia were contributing to the �A-induced decreasein SNP-stimulated cGMP formation, we examined thisresponse in cultures where the presence of microgliawas effectively decreased by treatment with cytosinearabinoside when the astrocyte monolayer reachesconfluence, as we previously described (Agullo et al.,1995). As shown in Fig 7, SNP-stimulated cGMP ac-cumulation was similar in cytosine arabinosidetreated and untreated cultures, in agreement with thelack of sGC in microglial cells (Agullo et al., 1995).Also of the same magnitude was the decrease in SNP-stimulated cGMP accumulation produced by �A(25–35), indicating that agents released by microglia arenot responsible for this effect and that �A are actingdirectly on the astrocytes. Weather an inflammatoryagent released by the astrocytes activated by �A isacting in an autocrine or paracrine way needs furtherinvestigation.
Activation of astroglial and microglial cultures with�A alone or in combination with INF-� has beenshown to induce NOS-2 expression and NO release(Meda et al., 1995; Goodwin et al., 1995; Rossi et al.,1996; Hu et al., 1998). Down-regulation of sGC bylong-term exposure to NO donors has been reportedin vascular smooth muscle cells and other cell types(Ujiie et al., 1994; Papapetropoulos et al., 1996a; Filip-pov et al., 1997) and proposed as a mechanism under-lying tolerance to nitrovasodilators. Thus, we investi-gated if �A-induced NO release was involved in thedecrease in SNP-dependent cGMP formation. Results
indicate that this is not the case. As shown in Fig. 8A,�A(25–35)-induced NO formation determined by ni-trite accumulation in the media was greatly enhancedby INF-� (three- to fourfold), but the decrease in SNP-stimulated cGMP formation was not affected (Fig. 8B).More definitive, coincubation of the cells with theNOS inhibitor NG-monomethyl-l-arginine (l-NMMA;1 mM) that totally prevented NO formation in re-sponse to �A(25–35) alone or in combination withINF-�, was not able to prevent the decrease in cGMPformation (Figs. 8A and 8B). Thus, �A down-regulatesGC in astrocytes in a NO-independent manner, sim-ilarly to what we previously observed in the same cellstreated with LPS (Baltrons and Garcıa, 1999) and incontrast to reports in smooth muscle cells exposed toLPS or inflammatory cytokines where the effect ap-pears to be totally or partially NO-dependent (Papa-petropoulos et al., 1996b; Tsuchida et al., 1994; Takataet al., 2001). NO endogenously generated in astrocytesin response to �A or LPS may not be sufficient todecrease sGC expression. Alternatively, these cellsmay have an efficient NO scavenging system. Re-cently, a potent NO inactivation mechanism that canmaintain low steady-state NO concentrations at con-stant rates of NO formation has been described incerebellar and other cells (Griffiths and Garthwaite,2001). Additionally, reduced glutathione levels areknown to be high in astrocytes (Dringen et al., 2000)
FIG. 7. Effect of �A on SNP-stimulated cGMP formation in cere-bellar astrocytes treated with cytosine-arabinoside. Cultures treatedor not with 10 �M cytosine-arabinoside (Cyt-ara) from day 10 inculture to day 14, were incubated with �A(25–35) for 48 h andcGMP formation in response to 100 �M SNP in the presence of 1mM IBMX was measured. Results are means � SD of triplicatedeterminations in a representative experiment that was replicatedwith similar results.
146 Baltrons et al.
© 2002 Elsevier Science (USA)All rights reserved.

and expression of the cystein-rich protein methallo-thionein, that can efficiently bind NO (Zangger et al.,2001), is increased in astrocytes of LPS treated rats(Itano et al., 1991) and in AD brains (Zambenedetti etal., 1998; Adlard et al., 1998).
To test if the effect of �A on sGC expression ob-served in cultured cells also occurred in vivo we in-jected 0.5 �g/�l of �A(1–40) into the cerebellar vermisand hippocampus of adult rats and analyzed the lev-els of sGC �1 and �1 subunit mRNA 24 and 48 h after
the treatment. As shown in Fig. 9, the mRNA of the �1
subunit was decreased in the cerebellar vermis by 45%respect to vehicle-treated animals 24 h after the treat-ment and remained low after 48 h. However, only asight tendency to decrease was observed in the mRNAof the �1 subunit. Similarly, in hippocampus there wasa 30–35% decrease in �1 subunit mRNA but no signif-icant decrease in �1 subunit mRNA 48 h after thetreatment. This suggests that the two subunits may bedifferentially regulated in the adult animal. Neverthe-less, the observed decrease in �1 mRNA indicate thatin vivo �A deposition can decrease the levels of sGCheterodimers containing the �1 subunit and thus limit
FIG. 9. Effect of �A focal injection on sGC subunit mRNA levels inadult rat hippocampus and cerebellum. PBS or �A(1–40) (0.5 �g/�lin PBS) were microinjected (2 �l) into the hippocampus and cere-bellar vermis of adult rats as described under Experimental Proce-dures. After 24 or 48 h, the brains were removed and the brainregions dissected. (A) RNA samples analysed by RT-PC using prim-ers specific for sGC �1 or �1 subunits and GAPDH. (B) RT-PCR dataanalysed by densitometry. sGC �1 or �1 subunit mRNA levels werenormalised to GAPDH mRNA levels and expressed relative to thelevels in vehicle-treated animals. Results are means � SEM of threeanimals per experimental group. *Significantly different from un-treated cells in each condition (P � 0.05).
FIG. 8. Effect of INF-� and l-NMMA on �A induced accumulationof nitrites (A) and cGMP (B) in cerebellar astrocytes. Cell cultureswere treated or not with 50 �M �A (25–35) for 48 h in the absenceor presence of 10 ng/ml INF-�, 1 mM l-NMMA, or both as indi-cated. l-NMMA was added 30 min before other agents. (A) Nitritelevels determined with the Greiss reagent in the culture media; (B)intracellular cGMP accumulated in response to 100 �M SNP in thepresence of 1 mM IBMX. Results are means � SD of triplicatedeterminations in a representative experiment that was replicatedwith similar results.
147�-Amyloids Decrease Soluble Guanylyl Cyclase
© 2002 Elsevier Science (USA)All rights reserved.

the formation of cGMP in response to NO. A reductionin sGC activity has been reported in the brain ofsubjects with AD (Bonkale et al., 1995). In light of ourresults this reduction may reflect the loss of sGC inreactive astrocytes surrounding �-amyloid plaques.Down-regulation of sGC could prevent excess signal-ling via cGMP under conditions of reactive gliosisassociated with high levels of NO production that mayoccur in senile plaques (Pike et al., 1995; Wallace et al.,1997). Alternatively, a decreased capacity of astroglialcells to generate cGMP may aggravate neuroinflam-mation since cGMP has been implicated in the inhibi-tion of �A-induced vasoactivity and microglial proin-flammatory response (Paris et al., 1999) and to antago-nise apoptotic death in cells of neural origin (Kim etal., 1999).
ACKNOWLEDGMENTS
We thank Francisca Garcıa for assistance in cell culture prepara-tions and Annabel Segura for technical assistance. This work wassupported in part by DGICYT (PB97-0201), Fundacio La Marato TV3(1008/97), and DGR (SGR99-00123) grants. Carlos E. Pedraza is therecipient of a predoctoral fellowship from Ministerio de Educaciony Cultura (Spain).
REFERENCES
Adlar, P. A., West, A. K., & Vickers, J. C. (1998) Increased density ofmetallothionein I/II-immunopositive cortical glial cells in theearly stages of Alzheimer’s disease. Neurobiol. Dis. 5, 349–356.
Akama, K. T., & Van Eldik, L. J. (2000) beta-Amyloid stimulation ofinducible nitric oxide synthase in astrocytes is interleukin-1�- andtumor necrosis factor-� (TNF-�)-dependent, and involves aTNF-� receptor-associated factor-and NFkappaB-inducing ki-nase-dependent signalling mechanism. J. Biol. Chem. 275, 7918–7924.
Araujo, D. M., & Cotman, C. W. (1992) �-Amyloid stimulates glialcells in vitro to produce growth factors that accumulate in senileplaques in Alzheimer’s disease. Brain Res. 569, 141–145.
Agullo, L., & Garcıa, A. (1992) Characterization of noradrenaline-stimulated cyclic GMP formation in brain astrocytes in culture.Biochem. J. 288, 619–624.
Agullo, L., Baltrons, M. A., & Garcıa, A. (1995) Calcium-dependentnitric oxide formation in glial cells. Brain Res. 686, 160–168.
Agullo, L., & Garcıa, A. (1997) Ca2�/calmodulin dependent cyclicGMP phosphodiesterase activity in granule neurons and astro-cytes from cerebellum. Eur. J. Pharmacol. 323, 119–125.
Baltrons, M. A., Saadoun, S., Agullo, L., & Garcıa, A. (1997) Regu-lation by calcium of the nitric oxide/cyclic GMP system in cere-bellar granule cells and astroglia in culture. J. Neurosci. Res. 49,333–341.
Baltrons, M. A., & Garcıa, A. (1999) Nitric oxide-independent down-regulation of soluble guanylyl cyclase by bacterial endotoxin inastroglial cells. J. Neurochem. 73, 2149–2157.
Baltrons, M. A., & Garcıa, A. (2001) The nitric oxide/cyclic GMPsystem in astroglial cells. In Progress in Brain Research (B. Castel-lano and M. Nieto-Sampedro, Eds.), Vol. 132, pp. 335–347.Elsevier Science, SV.
Bonkale, W. L., Winblad, B., Ravid, R., & Cowbum, R. F. (1995)Reduced nitric oxide responsive soluble guanylyl cyclase activityin the superior temporal cortex of patients with Alzheimer’sdisease. Neurosci. Lett. 187, 5–8.
Bradford, M. M. (1976) A rapid and sensitive method for the quan-titation of microgram quantities of protein utilizing the principleof protein-dye binding. Anal Biochem. 72, 248–254.
Brooker, G., Harper, J. F., Terasaki, W. L., & Moylan, R. D. (1979)Radioimmunoassay of cyclic AMP and cyclic GMP. Adv. CyclicNucleotide Res. 10, 1–33.
Buechler, W. A., Nakane, M., & Murad, F. (1991) Expression ofsoluble guanylate cyclase activity requires both enzyme subunits.Biochem. Biophys. Res. Commun. 174, 351–357.
Dringen, R., Gutterer, J. M., & Hirrlinger, J. (2000) Gluthationemetabolism in brain. Eur. J. Biochem. 267, 4912–4916.
Eddleston, M., & Mucke, L. (1993) Molecular profile of reactiveastrocytes-implications for their role in neurologic disease. Neu-roscience 54, 15–36.
Filippov, G., Bloch, D. B., & Bloch, K. D. (1997) Nitric oxide de-creases stability of mRNAs encoding soluble guanylate cyclasesubunits in rat pulmonary artery smooth muscle cells. J. Clin.Invest. 100, 942–948.
Forstermann, U., Boissel, J.-P., & Kleinert, H. (1998) Expressionalcontrol of the ‘constitutive’ isoforms of nitric oxide synthase (NOSI and NOS III). FASEB J. 12, 773–790.
Garthwaite, J., & Boulton, C. L. (1995) Nitric oxide signalling in thecentral nervous system. Annu. Rev. Physiol. 57, 683–706.
Gibb, B. J., & Garthwaite, J., (2001) Subunits of the nitric oxidereceptor, soluble guanylyl cyclase, expressed in rat brain. Eur.J. Neurosci. 13, 539–544.
Gonzalez-Scarano, F., & Baltuch, G. (1999) Microglia as mediators ofinflammatory and degenerative diseases. Annu. Rev. Neurosci. 22,219–240.
Goodwin, J. L., Uemura, E., & Cunnick, J. E. (1995) Microglialrelease of nitric oxide by the synergistic action of beta-amyloidand INF-gamma. Brain Res. 692, 207–214.
Griffiths, C., & Garthwaite, J. (2001) The shaping of nitric oxidesignals by a cellular sink. J. Physiol. 536, 855–862.
Guo, L., Sawkar, A., Zasadzki, M., Watterson, D. M., & Van Eldik,L. J. (2001) Similar activation of glial cultures from different ratbrain regions by neuroinflammatory stimuli and downregulationof the activation by a new class of small molecule ligands. Neu-robiol. Aging 22, 975–981.
Harteneck, C., Koesling, D., Soling, A., Schultz, G., & Bohme, E.(1990) Expression of soluble guanylyl cyclase. Catalytic activityrequires two enzyme subunits. FEBS Lett. 272, 221–223.
Hauss-Wegrzyniak, B., Dobrzanski, P., Stoehr, J. D., & Wenk, G. L.(1998) Chronic neuroinflammation in rats reproduces compo-nents of the neurobiology of Alzheimer’s disease. Brain Res. 780,294–303.
Heneka, M. T., Klockgether, T., & Feinstein, D. L. (2000) Peroxixomeproliferator-activated receptor-� ligands reduce neuronal induc-ible nitric oxide synthase expression and cell death in vivo. J. Neu-rosci. 20, 6862–6867.
Hu, J., Akama, K. T., Krafft, G. A., Chromy, B. A., & Van Eldik, L. J.(1998) Amyloid-� peptide activates cultured astrocytes: Morpho-logical alterations, cytokine induction and nitric oxide release.Brain Res. 785, 195–206.
148 Baltrons et al.
© 2002 Elsevier Science (USA)All rights reserved.

Itano, Y., Noji, S., Koyama, E., Taniguchi, S., Taga, N., Takahashi, T.,Ono, K., & Kosaka, F. (1991) Bacterial endotoxin-induced expres-sion of metallothionein genes in rat brain, as revealed by in situhybridization. Neurosci. Lett. 124, 13–16.
Klegeris, A., & McGeer, P. L. (1997) �-Amyloid protein enhancesmacrophage production of oxygen free radicals and glutamate.J. Neurosci. Res. 49, 229–235.
Kim, Y.-M., Chung, H.-T., Kim, S.-S., Han, J.-A., Yoo, Y.-M., Kim,K.-M., Lee, G.-H., Yun, H.-Y., Green, A., Li, J., Simmons, R. L., &Billiar, T. R. (1999) Nitric oxide protects PC12 cells from serumdeprivation-induced apoptosis by cGMP-dependent inhibition ofcaspase signalling. J. Neurosci. 19, 6740–6747.
Laskay, G., Zarandi, M., Varga, J., Jost, K., Fonagy, A., Torday, C.,Latzkovits, L., & Penke, B. (1997) A putative tetrapeptide antag-onist prevents �-amyloid-induced long-term elevation of [Ca2�]i
in rat astrocytes. Biochem. Biophys. Res. Commun. 235, 479–481.Liu, H., Force, T., & Bloch, K. D. (1997) Nerve growth factor de-
creases soluble guanylate cyclase in rat pheochromocytoma PC12cells. J. Biol. Chem. 272, 6038–6043.
Lowry, O. M., Rosebrough, N. J., Farr, A. L., & Randall, R. J. (1951)Protein measurements with the folin phenol reagent. J. Biol. Chem.193, 265–275.
Meda, L., Cassatella, M. A., Szendrei, G. I., Otvos, Jr. L., Baron, P.,Villalba, M., Ferrari, D., & Rossi, F. (1995) Activation of microglialcells by �-amyloide protein and interferon-�. Nature 374, 647–650.
Papapetropoulos, A., Go, C., Murad, F., & Catravas, J. D. (1996a)Mechanisms of tolerance to sodium nitroprusside in cultured rataortic smooth muscle cells. Br. J. Pharmacol. 117, 147–155.
Papapetropoulos, A., Abou-Mohamed, G., Marczin, N., Murad, F.,Cadwell, W., & Catravas, J. D. (1996b) Downregulation of nitro-vasodilator-induced cyclic GMP accumulation in cells exposed toendotoxin or interleukin-1�. Br. J. Pharmacol. 118, 1359–1366.
Pappolla, M. A., Chyan, Y. J., Omar, R. A., Hsiao, K., Perry, G.Smith, M. A., & Bozner, P. (1998) Evidence of oxidative stress andin vivo neurotoxicity of beta-amyloid in a transgenic mouse modelof Alzheimer’s disease: A chronic oxidative paradigm for testingantioxidant therapies in vivo. Am. J. Pathol. 152, 871–877.
Paris, D., Town, T., Parker, T. A., Tan, J., Humphrey, J., Crawford,F., & Mullan, M. (1999) Inhibition of Alzheimer’s �-Amyloidinduced vasoactivity and proinflammatory response in microgliaby a cGMP-dependent mechanism. Exp. Neurol. 157, 211–221.
Paxinos, G., Watson, C., Pennisi, M., & Topple, A. (1985) Bregma,lambda and the interaural midpoint in stereotaxic surgery with
rats of different sex, strain and weight. J. Neurosci. Methods 13,139–143.
Pike, C. J., Cummings, B. J., Monzavi, R., & Cotman, C. W. (1994)�-Amyloide-induced changes in cultured astrocytes parallel reac-tive astrocytosis associated with senile plaques in Alzheimer’sdisease. Neuroscience 63, 517–531.
Pike, C. J., Cummings, B. J., & Cotman, C. W. (1995) Early associa-tion of reactive astrocytes with senile plaques in Alzheimer’sdisease. Exp. Neurol. 132, 172–179.
Rossi, F., & Bianchini, E. (1996) Synergistic induction of nitric oxideby �-amyloid and cytokines in astrocytes. Biochem. Biophys. Res.Commun. 225, 474–478.
Salinero, O., Moreno-Flores, M. T., de Ceballos, M. L., & Wandosell,F. (1997) �-Amyloid peptide induced cytoskeletal reorganizationin cultured astrocytes. J. Neurosci. Res. 47, 216–223.
Smith, M. A., Richey Harris, P. L., Sayre, L. M., Beckman, J. S., &Perry, G. (1997) Widespread peroxynitrite-mediated damage inAlzheimer’s disease. J. Neurosci. 17, 2653–2657.
Takata, M., Filipopov, G., Liu, H., Ichinose, F., Janssens, St., Bloch,D. B., & Bloch, K. D. (2001) Cytokines decrease sGC in pulmonaryartery smooth muscle cells via NO-dependent and NO-indepen-dent mechanisms. Am. J. Physiol. Lung Cell Mol. Physiol. 280,L272–L278.
Tsuchida, S., Hiraoka, M., Sudo, M., Kigoshi, S., & Muramatsu, I.(1994) Attenuation of sodium nitroprusside responses after pro-longed incubation of rat aorta with endotoxin. Am. J. Physiol. 267,H2305–H2310.
Ujiie, K., Hogarth, L., Danziger, R., Drewett, J. G., Yuen, P. S. T.,Pang, I.-H., & Star R. A. (1994) Homologous and heterologousdesensitization of a guanylyl cyclase-linked nitric oxide receptorin cultured rat medullar interstitial cells. J. Pharmacol. Exp. Ther.270, 761–767.
Wallace, M. N., Geddes, J. G., Farquhar, D. A., & Masson, M. R.(1997) Nitric oxide synthase in reactive astrocytes adjacent to�-amyloid plaques. Exp. Neurol. 144, 266–272.
Zambenedetti, P., Girodano, R., & Zatta, P. (1998) Metallothioneinsare highly expressed in astrocytes and micropillaries in Alzhei-mer’s disease. J. Chem. Neuroanat. 15, 21–26.
Zangger, K., Oz, G., Haslinger, E., Kunert, O., & Armitage, I. M.(2001) Nitric oxide selectively release metals from the N-terminaldomain of metallothioneins: Potential role at inflammatory sites.FASEB J. 10. 1096/fj.00-o641fje.
Zhang, J., & Snyder, S. H. (1995) Nitric oxide in the nervous system.Annu. Rev. Pharmacol. Toxicol. 35, 213–233.
149�-Amyloids Decrease Soluble Guanylyl Cyclase
© 2002 Elsevier Science (USA)All rights reserved.












![Acute ethanol exposure induces [Ca 2+ ] i transients, cell swelling and transformation of actin cytoskeleton in astroglial primary cultures](https://static.fdokumen.com/doc/165x107/63221b7a61d7e169b00c78d0/acute-ethanol-exposure-induces-ca-2-i-transients-cell-swelling-and-transformation.jpg)







