
AMPA induced Ca 2+ influx in motor neurons occurs through voltage gated Ca 2+ channel and Ca 2+...
9
AMPA induced Ca 2+ influx in motor neurons occurs through voltage gated Ca 2+ channel and Ca 2+ permeable AMPA receptor Dinesh C. Joshi, Mahendra Singh, Karthik Krishnamurthy, Preeti G. Joshi, Nanda B. Joshi ⇑ Department of Biophysics, National Institute of Mental Health and Neuro Sciences, Bangalore 560 029, India article info Article history: Received 22 March 2011 Received in revised form 27 June 2011 Accepted 29 June 2011 Available online xxxx Keywords: Motor neurons Amyotrophic lateral sclerosis AMPA receptors Excitotoxicity Voltage gated calcium channels Calcium signaling Mitochondrial calcium abstract The rise in intracellular Ca 2+ mediated by AMPA subtype of glutamate receptors has been implicated in the pathogenesis of motor neuron disease, but the exact route of Ca 2+ entry into motor neurons is not clearly known. In the present study, we examined the role of voltage gated calcium channels (VGCCs) in AMPA induced Ca 2+ influx and subsequent intracellular signaling events responsible for motor neuron degeneration. AMPA stimulation caused sodium influx in spinal neurons that would depolarize the plasma membrane. The AMPA induced [Ca 2+ ] i rise in motor neurons as well as other spinal neurons was drastically reduced when extracellular sodium was replaced with NMDG, suggesting the involve- ment of voltage gated calcium channels. AMPA mediated rise in [Ca 2+ ] i was significantly inhibited by L-type VGCC blocker nifedipine, whereas x-agatoxin-IVA and x-conotoxin-GVIA, specific blockers of P/Q type and N-type VGCC were not effective. 1-Napthyl-acetyl spermine (NAS), an antagonist of Ca 2+ permeable AMPA receptors partially inhibited the AMPA induced [Ca 2+ ] i rise but selectively in motor neu- rons. Measurement of AMPA induced currents in whole cell voltage clamp mode suggests that a moderate amount of Ca 2+ influx occurs through Ca 2+ permeable AMPA receptors in a subpopulation of motor neu- rons. The AMPA induced mitochondrial calcium loading [Ca 2+ ] m , mitochondrial depolarization and neuro- toxicity were also significantly reduced in presence of nifedipine. Activation of VGCCs by depolarizing concentration of KCl (30 mM) in extracellular medium increased the [Ca 2+ ] i but no change was observed in mitochondrial Ca 2+ and membrane potential. Our results demonstrate that a subpopulation of motor neurons express Ca 2+ permeable AMPA receptors, however the larger part of Ca 2+ influx occurs through L-type VGCCs subsequent to AMPA receptor activation and consequent mitochondrial dysfunction is the trigger for motor neuron degeneration. Nifedipine is an effective protective agent against AMPA induced mitochondrial stress and degeneration of motor neurons. Ó 2011 Elsevier B.V. All rights reserved. 1. Introduction Excitotoxicity mediated by AMPA subtypes of glutamate recep- tors has been implicated in the selective vulnerability of motor neurons in amyotrophic lateral sclerosis (ALS) (Carriedo et al., 1996; Cleveland and Rothstein, 2001; Sen et al., 2005; Van Damme et al., 2002; Corona and Tapia, 2007; Duncan, 2009). Excessive in- crease in intracellular calcium on AMPA receptor stimulation has been considered as the primary trigger for motor neuron degener- ation. The Ca 2+ loading is largely mediated by AMPA receptors (Sen et al., 2008). AMPARs are cation conducting channels composed of four subunits GluR1 to GluR4 and their functional properties de- pend on various factors such as subunit composition, alternative splicing and RNA editing (Hollmann et al., 1991; Palmer et al., 2005). The AMPA receptors are permeable to Na + and K + but their Ca 2+ permeability is variable. GluR2 subunit of AMPARs undergoes RNA editing at Q/R site in second transmembrane domain replac- ing glutamine to arginine during post transcriptional modification (Duncan, 2009; Van Den Bosch et al., 2000; Van Damme et al., 2002). GluR2 subunit is considered as the determinant of Ca 2+ per- meability of AMPA receptors (Van Damme et al., 2002; Duncan, 2009; Kawahara and Kwak, 2005). It was suggested that the lack of GluR2 subunits or unedited form of GluR2 makes the AMPA receptors permeable to Ca 2+ ions. Some earlier studies have re- ported that the motor neurons possess unedited form of GluR2 or completely lack GluR2 subunit which makes the AMPA receptor highly permeable to Ca 2+ ions thus increasing their vulnerability to excitotoxicity (Williams et al., 1997; Bar-Peled et al., 1999). In contrast another study on single cell RT-PCR showed that the mo- 0197-0186/$ - see front matter Ó 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.neuint.2011.06.023 Abbreviations: AMPA, a-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid; EMEM, Eagle’s minimal essential medium; Hepes, N-2-hydroxyethyl piperazine-n-2 ethanesulphonic acid; CCCP, carbonyl cynide m-chlorophenyl hydrazone; TMRM, tetramethylrhodamine methyl ester; NAS, 1-napthyl acetyl spermine; VGCCs, voltage gated calcium channels. ⇑ Corresponding author. Tel.: +91 80 2699 5101/5106; fax: +91 80 2656 4830. E-mail addresses: [email protected], [email protected] (N.B. Joshi). Neurochemistry International xxx (2011) xxx–xxx Contents lists available at ScienceDirect Neurochemistry International journal homepage: www.elsevier.com/locate/nci Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca 2+ influx in motor neurons occurs through voltage gated Ca 2+ channel and Ca 2+ per- meable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.023
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of AMPA induced Ca 2+ influx in motor neurons occurs through voltage gated Ca 2+ channel and Ca 2+...

Neurochemistry International xxx (2011) xxx–xxx
Contents lists available at ScienceDirect
Neurochemistry International
journal homepage: www.elsevier .com/locate /nci
AMPA induced Ca2+ influx in motor neurons occurs through voltage gatedCa2+ channel and Ca2+ permeable AMPA receptor
Dinesh C. Joshi, Mahendra Singh, Karthik Krishnamurthy, Preeti G. Joshi, Nanda B. Joshi ⇑Department of Biophysics, National Institute of Mental Health and Neuro Sciences, Bangalore 560 029, India
a r t i c l e i n f o
Article history:Received 22 March 2011Received in revised form 27 June 2011Accepted 29 June 2011Available online xxxx
Keywords:Motor neuronsAmyotrophic lateral sclerosisAMPA receptorsExcitotoxicityVoltage gated calcium channelsCalcium signalingMitochondrial calcium
0197-0186/$ - see front matter � 2011 Elsevier B.V. Adoi:10.1016/j.neuint.2011.06.023
Abbreviations: AMPA, a-amino-3-hydroxy-5-methEMEM, Eagle’s minimal essential medium; Hepes, N-2ethanesulphonic acid; CCCP, carbonyl cynide m-chlotetramethylrhodamine methyl ester; NAS, 1-napthvoltage gated calcium channels.⇑ Corresponding author. Tel.: +91 80 2699 5101/51
E-mail addresses: [email protected], nbjoJoshi).
Please cite this article in press as: Joshi, D.C., etmeable AMPA receptor. Neurochem. Int. (2011)
a b s t r a c t
The rise in intracellular Ca2+ mediated by AMPA subtype of glutamate receptors has been implicated inthe pathogenesis of motor neuron disease, but the exact route of Ca2+ entry into motor neurons is notclearly known. In the present study, we examined the role of voltage gated calcium channels (VGCCs)in AMPA induced Ca2+ influx and subsequent intracellular signaling events responsible for motor neurondegeneration. AMPA stimulation caused sodium influx in spinal neurons that would depolarize theplasma membrane. The AMPA induced [Ca2+]i rise in motor neurons as well as other spinal neuronswas drastically reduced when extracellular sodium was replaced with NMDG, suggesting the involve-ment of voltage gated calcium channels. AMPA mediated rise in [Ca2+]i was significantly inhibited byL-type VGCC blocker nifedipine, whereas x-agatoxin-IVA and x-conotoxin-GVIA, specific blockers ofP/Q type and N-type VGCC were not effective. 1-Napthyl-acetyl spermine (NAS), an antagonist of Ca2+
permeable AMPA receptors partially inhibited the AMPA induced [Ca2+]i rise but selectively in motor neu-rons. Measurement of AMPA induced currents in whole cell voltage clamp mode suggests that a moderateamount of Ca2+ influx occurs through Ca2+ permeable AMPA receptors in a subpopulation of motor neu-rons. The AMPA induced mitochondrial calcium loading [Ca2+]m, mitochondrial depolarization and neuro-toxicity were also significantly reduced in presence of nifedipine. Activation of VGCCs by depolarizingconcentration of KCl (30 mM) in extracellular medium increased the [Ca2+]i but no change was observedin mitochondrial Ca2+ and membrane potential. Our results demonstrate that a subpopulation of motorneurons express Ca2+ permeable AMPA receptors, however the larger part of Ca2+ influx occurs throughL-type VGCCs subsequent to AMPA receptor activation and consequent mitochondrial dysfunction is thetrigger for motor neuron degeneration. Nifedipine is an effective protective agent against AMPA inducedmitochondrial stress and degeneration of motor neurons.
� 2011 Elsevier B.V. All rights reserved.
1. Introduction et al., 2008). AMPARs are cation conducting channels composed of
Excitotoxicity mediated by AMPA subtypes of glutamate recep-tors has been implicated in the selective vulnerability of motorneurons in amyotrophic lateral sclerosis (ALS) (Carriedo et al.,1996; Cleveland and Rothstein, 2001; Sen et al., 2005; Van Dammeet al., 2002; Corona and Tapia, 2007; Duncan, 2009). Excessive in-crease in intracellular calcium on AMPA receptor stimulation hasbeen considered as the primary trigger for motor neuron degener-ation. The Ca2+ loading is largely mediated by AMPA receptors (Sen
ll rights reserved.
yl-4-isoxazole propionic acid;-hydroxyethyl piperazine-n-2rophenyl hydrazone; TMRM,yl acetyl spermine; VGCCs,
06; fax: +91 80 2656 [email protected] (N.B.
al. AMPA induced Ca2+ influx i, doi:10.1016/j.neuint.2011.06.
four subunits GluR1 to GluR4 and their functional properties de-pend on various factors such as subunit composition, alternativesplicing and RNA editing (Hollmann et al., 1991; Palmer et al.,2005). The AMPA receptors are permeable to Na+ and K+ but theirCa2+ permeability is variable. GluR2 subunit of AMPARs undergoesRNA editing at Q/R site in second transmembrane domain replac-ing glutamine to arginine during post transcriptional modification(Duncan, 2009; Van Den Bosch et al., 2000; Van Damme et al.,2002). GluR2 subunit is considered as the determinant of Ca2+ per-meability of AMPA receptors (Van Damme et al., 2002; Duncan,2009; Kawahara and Kwak, 2005). It was suggested that the lackof GluR2 subunits or unedited form of GluR2 makes the AMPAreceptors permeable to Ca2+ ions. Some earlier studies have re-ported that the motor neurons possess unedited form of GluR2or completely lack GluR2 subunit which makes the AMPA receptorhighly permeable to Ca2+ ions thus increasing their vulnerability toexcitotoxicity (Williams et al., 1997; Bar-Peled et al., 1999). Incontrast another study on single cell RT-PCR showed that the mo-
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023

2 D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx
tor neurons express mRNAs for all four GluR subunits with GluR2being most abundant and edited at Q/R site (Greig et al., 2000;Vandenberghe et al., 2000a,b). It has been reported that GluR2 con-taining and GluR2 lacking AMPA receptors coexist in motor neu-rons (Vandenberghe et al., 2001). Also the whole cell currentmeasurements showed that the relative Ca2+ permeability of AMPAreceptors in motor neurons and dorsal horn neurons is comparable,nevertheless the current density was 2- to 3-folds higher in motorneurons than in dorsal horn neurons (Vandenberghe et al.,2000a,b). Such conflicting reports warrant further investigationon expression of GluR2 subunits and high Ca2+ permeability ofAMPA receptors in motor neurons.
Some earlier studies have shown that nifedipine, the voltagegated calcium channel (VGCC) blocker protects the AMPA inducedtoxicity in motor neurons (Fryer et al., 1999; Arakawa et al., 2002).This raises the possibility that the AMPA induced Ca2+ influx in mo-tor neurons could occur indirectly through VGCC. Owing to theirpermeability to Na+ and K+ the AMPA receptors depolarize the neu-ronal membrane which may lead to activation of VGCC. The pres-ent study was undertaken to elucidate the route of Ca2+ entry inspinal neurons during AMPA receptor activation and the down-stream events responsible for the selective vulnerability of motorneurons to excitotoxicity.
2. Experimental procedure
2.1. Materials
a-Amino-3-hydroxy-5-methyl-4-isoxazole propionic acid(AMPA), nifedipine, tissue culture grade N-2-hydroxyethylpiperazi-nen-2-ethanesulfonic acid (Hepes), glucose, sodium bicarbonate,poly-L-lysine and Eagle’s minimal essential medium (EMEM) werepurchased from Sigma–Aldrich India. x-Agatoxin-IVA, x-conotox-in-GVIA and polyclonal anti-Cav1.2 were purchased from AlomoneLabs, (Jerusalem, Israel). Horse serum and fetal bovine serum werepurchased from Gibco (Grand Island, NY). The fluorescent probesFura-2 acetoxymethyl ester (AM), SBFI-AM, Rhod-2AM, tetrameth-ylrhodamine methyl ester (TMRM), pluronic, goat-anti-mouse andgoat-anti-rabbit Alexafluor 488 and Alexafluor 633 labeled second-ary antibodies and Zenon antibody labeling kit were purchased fromMolecular Probes Inc. (Eugene, OR, USA). SMI-32 antibody was pur-chased from Sternberger Monoclonal Inc., USA. Monoclonal mouseanti-MAP-2 and anti-GluR2 were purchased from BD PharmingenSingapore. All other chemicals were analytical grade and were ob-tained from commercial sources.
2.2. Spinal cord neuronal cultures
Studies were conducted using primary spinal cord cultures from13- to 15-day-old embryonic Sprague Dawley rats. Cultures weregrown as described earlier with minor modifications (Sen et al.,2008). The spinal cords were dissected and cell suspension wasprepared by trituration using a pasture pipette. Cells were thenplated on 24 mm poly-L-lysine coated coverslips placed in 35 mmpetri dishes. Cells were plated at a density of 2.5 � 105 cells/ml.Plating medium consisted of EMEM with 2 mM glutamine, supple-mented with 10% horse serum, 10% fetal bovine serum, 25 mM glu-cose and muscle extract from 2-day-old rats. Cultures weremaintained at 37 �C in 5% CO2 atmosphere. After 4–6 days, cultureswere treated with 10 mM cytosine arabinoside for 24 h. Subse-quently, the cultures were maintained in a medium similar tothe plating medium without fetal bovine serum. Medium waschanged twice a week and experiments were performed after13–15 day old cultures.
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx imeable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.
2.3. Immunocytochemistry
Cultures were fixed with 4% paraformaldehyde for 15 min,washed three times with PBS and permeabilized with 0.1% TritonX-100 in PBS for 10 min. Cells were then washed three times withPBS, incubated with blocking solution (10% bovine serum albuminin PBS) for 30 min and washed with PBS. Subsequently, cells wereincubated with respective primary and secondary antibodies.
For identification of motor neurons and other spinal neuronsthe cultures were immunostained with SMI-32 antibody that spe-cifically labels non-phosphorylated neurofilaments in motor neu-rons and MAP-2 antibody which stains all types of neurons (Senet al., 2008). The primary antibodies anti-SMI-32 and anti-MAP-2were labeled with Alexafluor 488 and Alexafluor 633 respectivelyusing Zenon antibody labeling kit as per the method describedby the manufacturer (Molecular Probes, USA). The pre-labeledSMI-32 and MAP-2 antibodies were simultaneously added to theculture (at 1:500 dilution) and incubated for 30 min at room tem-perature. Cells were then washed with PBS three times.
The expression of GluR2 and L-type voltage gated Ca2+ channelswas examined using immunostaining with monoclonal anti-GluR2and anti-Cav1.2 antibodies respectively. Cultures were incubatedwith anti-GluR2 (at 1:200 dilution) for 2 h at room temperature,washed with PBS three time and incubated with goat-anti mouseAlexafluor 633 secondary antibody for 1 h. Subsequently cultureswere immunostained with anti-Cav1.2 (1:200 dilution) and Alexa-fluor 488 following the same procedure.
The immunostained cells were visualized with Olympus FV-1000 confocal microscope. The image acquisition and analysiswas performed using FV10-ASW1.66b software.
2.4. Live cell Ca2+ imaging
Intracellular Ca2+ was measured using the fluorescent indicatorfura-2AM as described earlier (Sen et al., 2005, 2008). The cultureswere loaded with 5 lM Fura-2AM in Hepes-buffered salt solution(containing in mM: Na+ 130, K+ 5.0, Mg2+ 0.8, Ca2+ 1.8, Cl� 130,Hepes 20 and glucose 15, pH 7.4) along with 0.08% pluronic acidfor 45 min at 37 �C, washed three times with HBSS and kept inthe dark for an additional 30 min to allow for complete deesterifi-cation of the dye. The cultures were then maintained at 25 �C inHBSS and mounted on a perfusion chamber and the same wasplaced on the stage of an inverted microscope (Olympus IX70, Ja-pan). The bath volume was 0.5 ml and images were acquired usingan apo 20� or 40� objective (Olympus).
Ratiometric fluorescence imaging was performed with a fluo-rescence imaging system (TILL Photonics, Germany). After loadingwith Fura-2 AM, the cells were alternately excited at 340 and380 nm with polychrome IV, a compact xenon arc lamp sourcecoupled to a high-speed monochromator. The emitted fluorescencewas selected with an appropriate band pass filter. Fluorescenceimages were acquired with a 12-bit Peltier cooled CCD camera.The data acquisition and analysis was done with TILLVision soft-ware. Fields containing at least one morphologically identified mo-tor neuron and 3–5 other spinal neurons were selected for imaging.[Ca2+]i was estimated by the method of Grynkiewicz et al., using Kd
value equal to 225 nM (Grynkiewiicz et al., 1985). Calibration wasperformed by measuring Rmax and Rmin in cells permeabilized with10 lM ionomycin in presence of 10 mM CaCl2 or 2 mM EGTArespectively.
Intracellular Na+ was measured using the fluorescent indicatorSBFI-AM (Minta and Tsien, 1989; Kiedrowski et al., 1994). Spinalcord cultures grown on cover slips were loaded with SBFI-AM(10 lM) and 0.08% (W/V) pluronic in HBSS at 37 �C for 90 minand [Na+]i was measured by ratiometric fluorescence imaging sim-ilarly as for live cell calcium imaging.
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023

D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx 3
2.5. Electrophysiology
Whole cell voltage-clamp recordings were performed on neu-rons using EPC-10 patch-clamp amplifier (HEKA, Germany). Patchpipettes fabricated from borosilicate glass capillaries (1.5 mm ODand 0.86 mm ID) had a resistance of 3–5 MO when filled withinternal solution containing (in mM): 120 CsCl. 3 MgCl2, 5 EGTA,20 Hepes, and 10 glucose, pH 7.2 adjusted with CsOH. The extracel-lular solution contained (in mM): 145 NaCl, 3 KCl, 2 CaCl2, 1 MgCl2,20 Hepes and 10 glucose pH 7.4 adjusted with NaOH. To assess theCa2+ permeability of AMPA receptors whole cell currents were re-corded in Na+ free extracellular solution in which NaCl and KClwere replaced by equimolar concentration of N-methyl-D-gluc-amine (NMDG) and CaCl2 concentration was increased to 10 mM.Extracellular solutions were supplemented with, APV (500 lM),tetrodotoxin (200 nM) and CdCl2 (100 lM) to block the NMDAreceptors, and voltage gated Na+ and Ca2+ channels respectively.All recordings were performed at room temperature. Cells held ata voltage of �60 mV were stimulated with 50 lM AMPA in pres-ence or absence of CTZ and currents were recorded in the wholecell mode. I–V curves were obtained by applying a voltage rampfrom �80 to +60 mV with the steps of 5 mV. Current recordingand analysis were done using Pulse and Pulsefit softwares (HekaInstruments, Germany) respectively.
2.6. Measurement of mitochondrial Ca2+
Mitochondrial Ca2+ was measured using mitochondrial Ca2+
sensitive fluorescent probe Rhod-2AM (Peng et al., 1998). The cellsgrown on coverslips were incubated with 500 nM of Rhod-2AM for10 min in Hepes buffer at room temperature. Subsequently, cellswere washed three times with Hepes buffer and incubated for20 min at room temperature to allow complete de-esterificationof the dye. The coverslip was placed on the perfusion chamberand mounted on the stage of confocal microscope (OlympusFV1000). Fluorescence was visualized with apo 40� water immer-sion objective and Rhod-2 fluorescence was imaged using 543 nmHe–Ne laser. The emitted fluorescence was monitored at 580 nm.As shown in Supplementary Fig. S1, punctate fluorescence ofRhod-2 was observed in the mitochondria with no appreciablefluorescence in the cytosol. AMPA and other agents were addedin the static bath. For quantitation of mitochondrial Ca2+ the re-gions of interest were taken from the perinuclear mitochondriarich regions that include either a single mitochondrion or a bunchof mitochondria. The change in mitochondrial Ca2+ was repre-sented by [(F/F0) � 100] where F is the fluorescence intensity at agiven time point and F0 is the initial fluorescence intensity (beforeaddition of agonist).
2.7. Measurement of mitochondrial membrane potential
Change in mitochondrial membrane potential was evaluatedusing lipophilic cationic fluorescent probe tetramethyl rhodaminemethyl ester (TMRM) that localizes within polarized mitochondriaas a function of membrane potential (Solenski et al., 2003). Cellsgrown on coverslips were loaded with 200 nM TMRM for 45 minat 37 �C in Hepes buffer. The coverslip containing TMRM loadedcells was placed onto mounting chamber on the stage of invertedmicroscope (Olympus IX70). Cells were visualized with apo 40�objective and fluorescence images were recorded at 537 nm withexcitation at 520 nm. The punctate loading of TMRM was observedin the mitochondria. TMRM (50 nM) was constantly present in thebath during experiment. The TMRM fluorescence of mitochondriarich regions in individual neurons was recorded as a function oftime. The fluorescence intensity (F) at a given time point was nor-malized to the initial fluorescence intensity (F0) before addition of
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx imeable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.
the AMPA or KCl and data are represented as [(F/F0) � 100]. At theend of each experiment protonophore CCCP (750 nM) was added tothe bath. CCCP collapses the mitochondrial membrane potentialand decreases the TMRM fluorescence corresponding to zero mito-chondrial membrane potential. The difference in TMRM fluores-cence intensity before and after CCCP addition was used for thequantitation of change in mitochondrial membrane potential.
2.8. Measurement of cell viability
Motor neuron specific marker SMI-32 and all neuronal markerMAP-2 were used to assess the percentage of viable motor neuronsand other spinal neurons. The viability of spinal neurons in culturewas determined by immunostaining method. Cultures grown oncoverslips, were divided into four groups (i) untreated control (ii)treated with 50 lM AMPA, (iii) treated with 50 lM AMPA in pres-ence of 10 lM nifedipine and (iv) treated with 10 lM nifedipinealone. Given concentration of AMPA was added to the coverslipsmaintained in 3 ml of EMEM and incubated for 1 h at room tem-perature. Cells in group (i) and (iv) were also incubated in parallel.Subsequently cells were washed with fresh EMEM and maintainedin the EMEM at 37 �C in 5% CO2 atmosphere. After 24 h cells werefixed with 4% paraformaldehyde and immunostained with SMI-32and MAP-2 antibodies as described above. The immunostained cul-tures were visualized with confocal microscope using 40� objec-tive and the number of viable neurons were counted. Theneurons with intact cell soma and neurites were considered as via-ble neurons. The cells immunopositive for both SMI-32 and MAP-2were considered as motor neurons whereas the cells immunopos-itive for only MAP-2 were considered as other spinal neurons. Thequantitation of viable neurons was performed by counting the via-ble motor neurons and other spinal neurons in 12 randomly se-lected fields on the coverslips. The average number of viable cellsin untreated control cultures was taken as 100% and the cell viabil-ity in treated groups is given as percentage of control. The datashown are from 5 experiments containing triplicate cultures ineach group.
2.9. Statistical analysis
Data for the number of experiments mentioned in the legendsare presented as mean ± SEM. Statistical significance was achievedby Student’s t-test and p < 0.05 was considered as significant.
3. Results and discussion
We examined the possibility of Ca2+ entry through voltagegated Ca2+ channels as well as Ca2+ permeable AMPA receptors.The AMPA induced Ca2+ current and change in [Ca2+]i were mea-sured in motor neurons and other spinal neurons by whole cellpatch-clamp recording and fluorescence imaging respectively. Forthese experiments, the motor neurons and other spinal neuronswere identified by morphological criteria. Cells with soma diame-ter P20 lm and extensive arborization were considered as motorneurons (Schaffner et al., 1987). At the end of some randomly se-lected Ca2+ imaging and electrophysiology experiments cultureswere fixed and the identification of motor neurons was confirmedby SMI-32 and MAP-2 immunostaining. The large neurons showedimmunofluorescence for motor neuron marker SMI-32 and neuro-nal marker MAP-2 and the smaller neurons in the field showedimmunofluorescence only for MAP-2 (Supplementary Fig. S2).
Exposure of spinal neurons to AMPA (50 lM) caused a transientrise in [Ca2+]i, however, replacement of extracellular Na+ withequimolar concentration of N-methyl-D-glucamine (NMDG) drasti-cally reduced the [Ca2+]i rise (Fig. 1A). AMPA receptor stimulation
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023

Fig. 1. [Ca2+]i and [Na+]i rise in motor neurons and other spinal neurons upon AMPAreceptor stimulation and KCl depolarization. AMPA induced changes in (A) Fura-2fluorescence intensity ratio 340/380 nm in presence or absence of Na+ in theextracellular buffer and (B) SBFI fluorescence intensity ratio 340/380 nm in motorneurons. (C) Change in Fura-2 fluorescence intensity ratio on KCl (30 mM)depolarization in presence and absence of 10 lM nifedipine. The data representmean ± SEM for 8–10 motor neurons and 10–12 other spinal neurons from 3 to 5different experiments.
4 D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx imeable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.
caused a rise in intracellular Na+ as indicated by a sustained in-crease in the fluorescence intensity of SBFI loaded into the cells(Fig. 1B). These data suggest that depolarization of neuronal mem-brane consequent to AMPA receptor mediated Na+ influx leads toCa2+ entry through VGCC. Depolarization of neuronal membranesby increasing the extracellular K+ concentration also caused greater[Ca2+]i increase in motor neurons as compared to other spinal neu-rons (Fig. 1C). Nifedipine (10 lM) caused about 95% inhibition inKCl induced peak [Ca2+]i, suggesting that the L-type VGCC are themajor contributors to depolarization induced Ca2+ influx in bothtypes of spinal neurons. In motor neurons the peak [Ca2+]i observedupon depolarization with buffer containing 30 mM KCl was com-parable to that caused by 50 lM AMPA but the rate of decay to ba-sal level was much slower in the latter case (Fig. 1A and C).
We further investigated the subtype of VGCCs involved in AMPAinduced Ca2+ influx by using specific blockers. x-Agatoxin-IVA andx-conotoxin-GVIA, the respective blockers of P/Q and N type chan-nels did not show any appreciable effect on AMPA-induced [Ca2+]i
but it was drastically reduced in presence of nifedipine, the specificblocker of L-type Ca2+ channel (Fig. 2A–C). In both types of neuronsa substantial inhibition in [Ca2+]i was observed in presence of10 lM nifedipine and no further inhibition was observed at higherconcentrations of the blocker. In presence of combination of thesethree blockers, the magnitude of [Ca2+]i rise was similar to that ob-served in presence of nifedipine alone (Data not shown).
The contribution of Ca2+ permeable AMPA receptor was alsoexamined. These receptors can be specifically blocked by 1-napthylacetyl spermine (NAS) (Van Damme et al., 2002). To assess the con-tribution of Ca2+ permeable AMPA receptors the spinal neurons,the cells were stimulated with AMPA (50 lM) in the absence andpresence of NAS (50 lM). As described earlier (Sen et al., 2008)there was considerable variability in AMPA induced [Ca2+]i risewithin the motor neurons as well as within other spinal neurons.Fig. 3 shows the time course of AMPA induced [Ca2+]i rise in motorneuron from a typical experiment and an average change in peak[Ca2+]i rise induced by 50 lM AMPA in absence and presence ofNAS (50 lM). NAS caused partial but a significant reduction inAMPA induced [Ca2+]i rise suggesting that the Ca2+ permeableAMPA receptors also contribute to Ca2+ influx in motor neurons.
To confirm these results we directly measured AMPA inducedcurrents in whole cell voltage clamp experiments. In Na+ enriched(145 mM) solution and at a holding potential of �60 mV, applica-tion of AMPA (50 lM) induced a fast decaying inward current inall the tested motor neurons (n = 9) as well as in other spinal neu-rons (n = 7) (data not shown). To measure the current mediated byCa2+ permeable AMPA receptors, recordings were performed in Na+
free buffer in which extracellular Na+ was replaced with NMDG,Ca2+ concentration raised to 10 mM and supplemented with100 lM Cd2+ to block the voltage gated Ca2+ channels. No apprecia-ble AMPA induced current was observed in other spinal neurons(n = 7), however, some motor neurons showed small fast decayinginward current (Fig. 4C and D). When AMPA was applied in pres-ence of CTZ (100 lM) to avoid the fast desensitization of AMPAreceptors, modest inward current was observed in some motorneurons (in 4 out of 9 tested motor neurons) whereas no apprecia-ble current was seen in other spinal neurons. These results suggestthat a moderate amount of Ca2+ influx occurs through Ca2+ perme-able AMPA receptors in a subpopulation of motor neurons. Thesedata are in agreement with earlier reports (Vandenberghe et al.,2000a,b; Greig et al., 2000). I–V curves were generated to furthercharacterize the Ca2+ permeable AMPA receptors in motor neurons.Fig. 5 shows the I–V relationship in a representative motor neuronand other spinal neuron in Na+ enriched and Na+ free solutions. InNa+ free buffer the motor neurons eliciting Ca2+ current (n = 4)
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023

Fig. 2. Effect of VGCC blocker on AMPA induced [Ca2+]i rise in motor neurons andother spinal neurons. AMPA induced change in Fura-2 fluorescence intensity ratio340/380 nm in presence and absence of 10 lM nifedipine in (A) motor neurons and(B) other spinal neurons. (C) Peak [Ca2+]i rise in presence and absence 10 lMnifedipine, 50 nM x-agatoxin or 50 nM x-conotoxin. The data represent mean ± SEMof 8–10 motor neurons and 12–15 other spinal neurons from 5 different experiments.
Fig. 3. Effect of 1-napthyl acetyl spermine (NAS) on AMPA induced [Ca2+]i rise inspinal neurons. (A) Representative traces of Fura-2 fluorescence intensity ratio 340/380 nm in motor neurons in presence and absence of 25 lM NAS. (B) The peak[Ca2+]i in presence or absence of NAS. The data represent mean ± SEM from 8 to 10motor neurons and 10–12 other spinal neurons from 5 experiments.
Fig. 4. AMPA induced whole cell current in motor neuron (A and C) and other spinalneuron (B and D). Recordings were done in Na+ free extracellular buffer containing10 mM Ca2+. AMPA (50 lM) was applied with and without 200 lM cyclothiazide(CTZ) at a holding potential of �60 mV. Traces A and C show AMPA induced currentin motor neuron in presence and absence of CTZ respectively. (B) Other spinalneuron does not show AMPA induced current in absence of CTZ, D, however a smallcurrent was observed in presence of CTZ.
D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx 5
showed outward rectification with rectification index ranging from1.6 to 4.7 (mean ± SEM: 3.37 ± 0.77). The relative Ca2+ permeabilityof AMPA receptor was estimated using the constant field equation(Lino et al., 1990; Greig et al., 2000).
PCa=PCs ¼ ½Csþ�i=½Ca2þ�0 � expðErvF=RTÞ � ½expðErvF=RTÞ þ 1�� 0:25
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx imeable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.
where PCa and PCs represent the permeability coefficients to Ca2+
and Cs+, respectively, Erv is the reversal potential in Na+ free/Ca2+
enriched buffer, R, T and F have their usual meaning.
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023

Fig. 5. I–V relationship for AMPA induced current in motor neuron (A) and other spinal neuron (B) in presence of 200 lM CTZ and 100 lM Cd2+. I–V curves were obtained as2 s voltage ramps from �80 to +60 mV in, (1) Na+ rich and (2) Na+ free extracellular buffer. In Na+ free extracellular buffer Ca2+ concentration was increased to 10 mM.
6 D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx
For motor neurons showing Ca2+ current the PCa2+/PCs+ was inthe range of 0.8–1.7 (mean ± SEM: 1.05 ± 0.22). These values aremuch lower than the reported PCa2+/PCs+ for GluR2 lacking AMPAreceptors (Vandenberghe et al., 2000a,b).
The expression of GluR2 subunit of AMPA receptor in spinalneurons was examined by immunocytochemistry. As shown inFig. 6A, GluR2 subunit is abundantly expressed in motor neuronsand other spinal neurons. These neurons also showed abundantexpression of L-type VGCC (Fig. 6B). Taken together these resultsdemonstrate that motor neurons express both Ca2+ permeableand impermeable AMPA receptors but the AMPA induced rise in[Ca2+]i predominantly occurs through L-type VGCC.
Mitochondria play an important role in Ca2+ homeostasis bysequestering Ca2+ when its level in the cytosol rises beyond a par-ticular concentration (Szabadkai and Duchen, 2008). Excessive Ca2+
loading in mitochondria is known to trigger apoptotic signaling.We probed the mitochondrial calcium [Ca2+]m with Rhod-2 whichpredominantly localizes in mitochondria. Localization of Rhod-2 incells was critically assessed by taking high resolution confocalimages. Punctate fluorescence of Rhod-2 was observed in the mito-chondria with no appreciable fluorescence in the cytosol (Supple-mentary Fig. S2). Stimulation of spinal neurons with AMPA(50 lM) caused an increase in Rhod-2 fluorescence intensity inboth type of neurons but the increase was significantly higher inmotor neurons than in other spinal neurons (Fig. 7A and B). Unlike
Fig. 6. Confocal images of spinal cord neurons immunostained with (A) anti-GluR2 andsecondary antibodies conjugated with Alexafluor 633 (red) and Alexafluor 488 (green) rneurons as well as other spinal neurons. (For interpretation of the references to colour
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx imeable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.
the transient rise in [Ca2+]i the [Ca2+]m remained high for the entireduration of recording (10 min).
The potential across the mitochondrial membrane is crucial forits function and it alters upon Ca2+ loading. We monitored themitochondrial membrane potential with potential sensitive fluo-rescent probe TMRM. Under normal conditions TMRM localizesin polarized mitochondria and shows punctate staining. As themitochondrial membrane is depolarized, it diffuses out into thecytoplasm and subsequently moves out of the plasma membrane.As depicted in Fig. 8, AMPA stimulation caused a decrease in thefluorescence intensity of TMRM, indicating depolarization of mito-chondrial membrane. The AMPA-induced decrease in TMRM fluo-rescence was much more prominent and faster in motor neuronsas compared to other spinal neurons. After 20 min recording, pro-tonophore carbonyl cyanide m-chlorophenyl hydrazone (CCCP)was added to dissipate the mitochondrial membrane potentialand the residual TMRM fluorescence corresponding to zero mem-brane potential was recorded. The AMPA induced depolarizationof mitochondrial membrane potential is represented as the differ-ence in normalized TMRM fluorescence before and after addition ofCCCP (Fig. 8C). The AMPA-induced changes in Rhod-2 and TMRMfluorescence were potently inhibited by nifedipine (Figs. 7A and8A). However, activation of VGCC by increasing the extracellularKCl concentration (30 mM) did not cause any appreciable changein mitochondrial Ca2+ level and membrane potential. Although
(B) anti-CaV1.2. Cells were immunostained with anti-GluR2 and anti-CaV1.2 usingespectively. Abundant immunofluorescence is seen for GluR2 and CaV1.2 in motor
in this figure legend, the reader is referred to the web version of this article.)
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023

Fig. 7. AMPA induced mitochondrial Ca2+ loading in spinal neurons. Rhod-2 loadedspinal cord cultures were stimulated with 50 lM AMPA or depolarized with 30 mMKCl. AMPA induced changes in Rhod-2 fluorescence in (A) motor neurons and (B)other spinal neurons in presence or absence of 10 lM nifedipine. (C) Increase inpeak fluorescence intensity on AMPA receptor stimulation in presence and absenceof nifedipine and upon KCl depolarization. The data shown are mean ± SEM from 6to 8 motor neurons and 8–10 other spinal neurons from 3 to 5 experiments.
Fig. 8. Change in mitochondrial membrane potential during AMPA receptorstimulation and KCl depolarization. The spinal cord cultures were loaded withTMRM were stimulated with 50 lM AMPA or depolarized with 30 mM KCl and750 nM CCCP was applied at time points shown by arrows. Time course of AMPAinduced changes in TMRM fluorescence in (A) motor neurons and (B) other spinalneurons, (a) unstimulated control, (b) in presence of nifedipine and (c) in absence ofnifedipine. (C) Difference in normalized TMRM fluorescence before and afteraddition of CCCP. The data shown are mean ± SEM from 6 to 8 motor neurons and8–10 other spinal neurons from 3 to 5 experiments.
D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx 7
the KCl-induced peak [Ca2+]i was similar to that observed withAMPA (50 lM) stimulation, there was no appreciable change in
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx in motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-meable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.023
8 D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx
Rhod-2 fluorescence (Fig. 7C). Similarly the TMRM fluorescencealso did not change upon KCl exposure, nevertheless, subsequentaddition of CCCP rapidly decreased the TMRM fluorescence in bothtype of neurons indicating that the mitochondrial membrane po-tential is maintained during KCl depolarization (Fig. 8C).
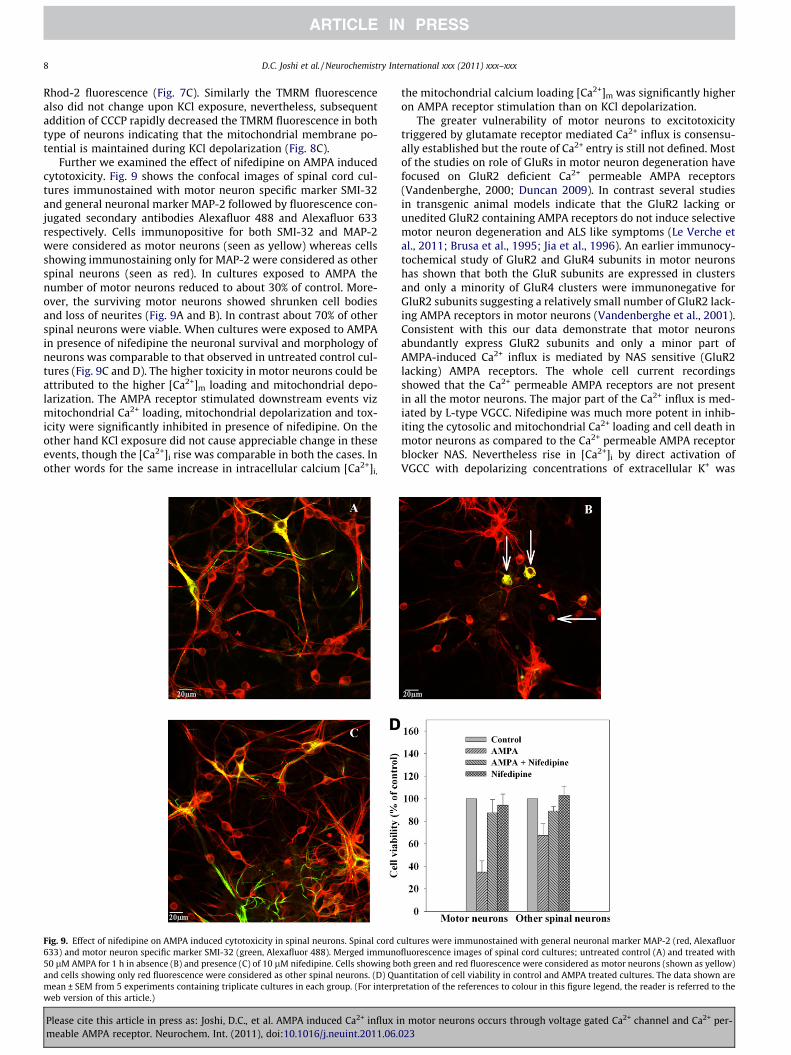
Further we examined the effect of nifedipine on AMPA inducedcytotoxicity. Fig. 9 shows the confocal images of spinal cord cul-tures immunostained with motor neuron specific marker SMI-32and general neuronal marker MAP-2 followed by fluorescence con-jugated secondary antibodies Alexafluor 488 and Alexafluor 633respectively. Cells immunopositive for both SMI-32 and MAP-2were considered as motor neurons (seen as yellow) whereas cellsshowing immunostaining only for MAP-2 were considered as otherspinal neurons (seen as red). In cultures exposed to AMPA thenumber of motor neurons reduced to about 30% of control. More-over, the surviving motor neurons showed shrunken cell bodiesand loss of neurites (Fig. 9A and B). In contrast about 70% of otherspinal neurons were viable. When cultures were exposed to AMPAin presence of nifedipine the neuronal survival and morphology ofneurons was comparable to that observed in untreated control cul-tures (Fig. 9C and D). The higher toxicity in motor neurons could beattributed to the higher [Ca2+]m loading and mitochondrial depo-larization. The AMPA receptor stimulated downstream events vizmitochondrial Ca2+ loading, mitochondrial depolarization and tox-icity were significantly inhibited in presence of nifedipine. On theother hand KCl exposure did not cause appreciable change in theseevents, though the [Ca2+]i rise was comparable in both the cases. Inother words for the same increase in intracellular calcium [Ca2+]i,
Fig. 9. Effect of nifedipine on AMPA induced cytotoxicity in spinal neurons. Spinal cord c633) and motor neuron specific marker SMI-32 (green, Alexafluor 488). Merged immuno50 lM AMPA for 1 h in absence (B) and presence (C) of 10 lM nifedipine. Cells showing band cells showing only red fluorescence were considered as other spinal neurons. (D) Qumean ± SEM from 5 experiments containing triplicate cultures in each group. (For interprweb version of this article.)
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx imeable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.
the mitochondrial calcium loading [Ca2+]m was significantly higheron AMPA receptor stimulation than on KCl depolarization.
The greater vulnerability of motor neurons to excitotoxicitytriggered by glutamate receptor mediated Ca2+ influx is consensu-ally established but the route of Ca2+ entry is still not defined. Mostof the studies on role of GluRs in motor neuron degeneration havefocused on GluR2 deficient Ca2+ permeable AMPA receptors(Vandenberghe, 2000; Duncan 2009). In contrast several studiesin transgenic animal models indicate that the GluR2 lacking orunedited GluR2 containing AMPA receptors do not induce selectivemotor neuron degeneration and ALS like symptoms (Le Verche etal., 2011; Brusa et al., 1995; Jia et al., 1996). An earlier immunocy-tochemical study of GluR2 and GluR4 subunits in motor neuronshas shown that both the GluR subunits are expressed in clustersand only a minority of GluR4 clusters were immunonegative forGluR2 subunits suggesting a relatively small number of GluR2 lack-ing AMPA receptors in motor neurons (Vandenberghe et al., 2001).Consistent with this our data demonstrate that motor neuronsabundantly express GluR2 subunits and only a minor part ofAMPA-induced Ca2+ influx is mediated by NAS sensitive (GluR2lacking) AMPA receptors. The whole cell current recordingsshowed that the Ca2+ permeable AMPA receptors are not presentin all the motor neurons. The major part of the Ca2+ influx is med-iated by L-type VGCC. Nifedipine was much more potent in inhib-iting the cytosolic and mitochondrial Ca2+ loading and cell death inmotor neurons as compared to the Ca2+ permeable AMPA receptorblocker NAS. Nevertheless rise in [Ca2+]i by direct activation ofVGCC with depolarizing concentrations of extracellular K+ was
ultures were immunostained with general neuronal marker MAP-2 (red, Alexafluorfluorescence images of spinal cord cultures; untreated control (A) and treated with
oth green and red fluorescence were considered as motor neurons (shown as yellow)antitation of cell viability in control and AMPA treated cultures. The data shown areetation of the references to colour in this figure legend, the reader is referred to the
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023

D.C. Joshi et al. / Neurochemistry International xxx (2011) xxx–xxx 9
not sufficient to cause cell death. Similar results have been re-ported earlier by other groups (Tymianski et al., 1993; Fryer etal., 1999). This could be due to the fact that the AMPA receptoractivation causes an increase in [Na+]i, in addition to an increasein [Ca2+]i, which could play a crucial role in mitochondrial Ca2+
loading (Mc Cormack et al., 1990; Kiedrowski et al., 1994). Negligi-bly small [Ca2+]i rise was seen when AMPA stimulation was done inthe absence of extracellular sodium, suggesting that even the NASsensitive AMPA receptors are affected by cytosolic Na+ concentra-tion. It was observed that with brief (100 s) exposure of neuronsto AMPA, the cytosolic Na+ (Fig. 1) and mitochondrial Ca2+
(Fig. 7) levels remained high during the recording time of 600 s,whereas the cytosolic Ca2+ recovered to basal level within 200 s.Due to sustained high level of Na+ in cytosol the plasma membranewould be in depolarized state causing continuous influx of Ca2+
through VGCC. The cytosolic Ca2+ level recovers to basal valuedue to buffering mechanisms including sequestration by mito-chondria and this was reflected as sustained increase in mitochon-drial Ca2+. We further demonstrate that Ca2+ influx consequent toAMPA receptor stimulation causes mitochondrial stress as evi-denced by rise in mitochondrial Ca2+ and its depolarization thatis not seen in case of K+-induced depolarization. Mitochondrialstress is considered as a crucial factor in inducing apoptotic signal-ing events. As suggested earlier possibly focal increase in [Ca2+]i inthe vicinity of AMPA receptors could be the determinant of downstream excitotoxic signaling events via locally expressed adaptorproteins and enzymes (Tymianski et al., 1993; Sattler et al.,1998). Together our data show that the selective vulnerability ofmotor neurons to excitotoxicity cannot be attributed only to Ca2+
permeable AMPA receptors but the Ca2+ impermeable AMPAreceptors play a major role via co-activation of L-type VGCC andimpairment of mitochondrial function. Most importantly theAMPA-induced mitochondrial stress and motor neuron death wasprevented by nifedipine.
Acknowledgement
This work is supported by Council of Scientific and IndustrialResearch, New Delhi, India.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.neuint.2011.06.023.
References
Arakawa, Y., Nishijima, C., Shimizu, N., Urushidani, T., 2002. Survival promotingactivity of nimodipine and nifedipine in rat motoneurons: implications of anintrinsic calcium toxicity in motoneurons. J. Neurochem. 83, 150–156.
Bar-Peled, O., O’Brien, R.J., Morrison, J.H., Rothstein, J.D., 1999. Cultured motorneurons possess calcium-permeable AMPA/kainate receptors. NeuroReport 10,855–859.
Brusa, R., Zimmerman, F., Koh, D.S., Feldmeyer, D., Gass, P., Seeburg, P.H., Sprengel,R., 1995. Early onset epilepsy and postnatal lethality associated with an editingdeficient GluR-B allele in mice. Science 270, 1677–1680.
Carriedo, S.G., Yin, H.Z., Weiss, J.H., 1996. Motor neurons are selectivelyvulnerable to AMPA/kainate receptor-mediated injury in vitro. J. Neurosci.16, 4069–4079.
Corona, J.C., Tapia, R., 2007. Ca2+ permeable AMPA receptors and intracellular Ca2+
determine motoneuron vulnerability in rat spinal cord in vivo.Neuropharmacology 52, 1219–1228.
Cleveland, D.W., Rothstein, J.D., 2001. From Charcot to Lou Gehrig: decipheringselective motor neuron death in ALS. Nat. Rev. Neurosci. 2, 806–819.
Please cite this article in press as: Joshi, D.C., et al. AMPA induced Ca2+ influx imeable AMPA receptor. Neurochem. Int. (2011), doi:10.1016/j.neuint.2011.06.
Duncan, K., 2009. The role of AMPA-receptor mediated excitotoxicity in ALS: isdeficient RNA editing to blame? Curr. Anaesth. Crit. Care 20, 227–235.
Fryer, H.J., Knox, R.J., Strittmatter, S.M., Kalb, R.G., 1999. Excitotoxic death of asubset of embryonic rat motor neurons in vitro. J. Neurochem. 72, 500–513.
Greig, A., Donevan, S.D., Mujtaba, T.J., Parks, T.N., Rao, M.S., 2000. Characterization ofthe AMPA-activated receptors present on motoneurons. J. Neurochem. 74, 179–191.
Grynkiewiicz, G., Poenie, M., Tsien, R.Y., 1985. A new generation of Ca2+
indicators with greatly improved fluorescent properties. J. Biol. Chem. 260,3440–3450.
Hollmann, M., Hartley, M., Heinemann, S., 1991. Ca2+ permeability of KA-AMPA-gated glutamate receptor channels depends on subunit composition. Science252, 851–853.
Jia, Z., Agopyan, N., Xiongz, M.P., Henderson, J., Gerlai, R., Taverna, F.A., Velumian, A.,MacDonald, J., Carlen, P., Abramov-Newerly, W., Roger, J., 1996. Enhanced LTP inmice deficient in AMPA receptorGluR2. Neuron 34, 759–772.
Kawahara, Y., Kwak, S., 2005. Excototoxicity and ALS: what is unique about theAMPA receptors expressed on spinal neurons? Amyotroph. Lateral Scler. 6, 131–144.
Kiedrowski, L., Wroblewski, J.T., Coast, E., 1994. Intracellular sodium concentrationin cultured cerebellar granule cells challenged with glutamate. Mol. Pharm. 45,1050–1054.
Le Verche, V., Ikiz, B., Jacquier, A., Przedborski, S., Re D, B., 2011. Glutamatepathways implication in amyotrophic lateral sclerosis: what is the signal in thenoise? J. Recept. Ligang Channel Res. 4, 1–22.
Lino, M., Ozawa, S., Tsuzuki, K., 1990. Permeation of calcium through excitatoryamino acid receptor channels in cultured rat hippocampal neurons. J. Physiol.424, 151–165.
Mc Cormack, J.G., Halestrap, A.P., Denton, R.M., 1990. Role of calcium ions inregulation of mammalian intramitochondrial metabolism. Physiol. Rev. 70,391–425.
Minta, A.K., Tsien, R., 1989. Fluorescent indicators for cytosolic sodium. J. Biol.Chem. 264, 19449–19457.
Palmer, C.L., Cotton, L., Heneley, J.M., 2005. The molecular pharmacology and cellbiology of a-amoni-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptors.Pharmacol. Rev. 57, 253–277.
Peng, T.I., Jou, M.J., Greenamare, T., 1998. Privileged access to mitochondria ofcalcium influx through NMDA receptors. Mol. Pharma. 53, 974–980.
Sattler, R., Charlton, M.P., Hafner, M., Tymianski, M., 1998. Distinct influx pathways,not calcium load, determine neuronal vulnerability to calcium neurotoxicity. J.Neurochem. 7, 2349–2364.
Schaffner, A.E., St. John, P.A., Barker, J.L., 1987. Fluorescence activated cell sorting ofembryonic mouse and rat motor neurons and their long term survival in vitro. J.Neurosci. 7, 3088–3104.
Sen, I., Nalini, A., Joshi, N.B., Joshi, P.G., 2005. Cerebrospinal fluid fromamyotrophic lateral sclerosis patients preferentially elevates intracellularcalcium and toxicity in motor neurons via AMPA/kainate receptor. J. Neurol.Sci. 235, 45–54.
Sen, I., Joshi, D.C., Joshi, P.G., Joshi, N.B., 2008. NMDA and non-NMDA receptormediated differential Ca2+ load and greater vulnerability of motor neurons inspinal cord cultures. Neurochem. Int. 52, 247–255.
Solenski, N.J., Kostecki, V.K., Dovey, S., Periasamy, A., 2003. Nitric-oxide-induceddepolarization of neuronal mitochondria: implications for neuronal cell death.Mol. Cell. Neurosci. 24, 1151–1169.
Szabadkai, G., Duchen, M.R., 2008. Mitochondria: The hub of Ca2+ signaling.Physiology 23, 84–94.
Tymianski, M., Charlton, M.P., Carlen, P.L., Tator, C.H., 1993. Source specificity ofearly calcium neurotoxicity in cultured embryonic spinal neurons. J. Neurosci.13, 2085–2104.
Van Den Bosch, L., Vandenberghr, W., Klaassen, H., Van Houtte, E., Robberecht, W.,2000. Ca2+ – permeable AMPA receptors and selective vulnerability of motorneurons. J. Neurological. Sci. 180, 29–34.
Van Damme, P., Dewil, M., Van Den Bosch, L., Van Houtte, E., Callewaert, G.,Robberecht, W., 2002. GluR2 dependent properties of AMPA receptorsdetermine the selective vulnerability of motor neurons to excitotoxicity. J.Neurophysiol. 88, 1279–1287.
Vandenberghe, W., Robberecht, W., Brorson, J.R., 2000a. AMPA receptor calciumpermeability, GluR2 expression, and selective motor neuron vulnerability. J.Neurosci. 20, 123–132.
Vandenberghe, W., Ihle, E.C., Patneau, D.K., Robberecht, W., Brorson, J.R., 2000b.AMPA receptor current density, not desensitization, predicts selectivemotoneuron vulnerability. J. Neurosci. 20, 7158–7166.
Vandenberghe, W., Bindokas, V.P., Miller, R.J., Robberecht, W., Brorson, J.R., 2001.Subcellular localization of calcium permeable AMPA receptors in spinalmotoneurons. Euro. J. Neurosci. 14, 305–314.
Williams, T.L., Day, N.C., Ince, P.G., Kamboj, R.K., Shaw, P.J., 1997. Calcium-permeable alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acidreceptors: a molecular determinant of selective vulnerability in amyotrophiclateral sclerosis. Ann. Neurol. 42, 200–207.
n motor neurons occurs through voltage gated Ca2+ channel and Ca2+ per-023




















