
Altered emotional behaviors in the diabetes mellitus OLETF type 1 congenic rat
11
Research Report Altered emotional behaviors in the diabetes mellitus OLETF type 1 congenic rat Akihito Watanabe a,b, ⁎ , Shiro Okuno c , Mai Okano b , Shaun Jordan e , Koutoku Aihara c , Takeshi K. Watanabe d , Yuki Yamasaki b , Hisashi Kitagawa b , Kiyoshi Sugawara a , Satoru Kato a a Department of Molecular Neurobiology, Graduate School of Medicine, Kanazawa University, Kanazawa, 920-8640, Japan b Second Institute of New Drug Discovery, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, Japan c Quests Research Institute, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, Japan d Otsuka GEN Research Institute, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, Japan e Department of Neuroscience Research, Otsuka Maryland Medicinal Laboratories, 9900 Medical Center Drive, Rockville, Maryland, USA ARTICLE INFO ABSTRACT Article history: Accepted 28 July 2007 Available online 15 August 2007 GPR10 is a G-protein-coupled receptor expressed in thalamic and hypothalamic brain regions, including the reticular thalamic nucleus (RTN) and periventricular nucleus (Pev), and the endogenous ligand for this receptor, prolactin-releasing peptide (PrRP), has demonstrated regulatory effects on the stress response. We produced a congenic rat by introducing the Dmo1 allele from the OLETF rat which encodes the amino acid sequences of GPR10 with a truncated NH 2 -terminus, into the Brown–Norway background. Using receptor autoradiography, we determined a lack of specific [ 125 I]PrRP binding in the RTN and Pev of these mutant rats compared to the control rats. Furthermore, intracerebroventricular injection of PrRP did not induce a significant increase of c-fos-like immunoreactivity in the paraventricular nucleus of the mutant rats compared to the control rats. The mutant rats also displayed a less anxious-like phenotype in three behavioral-based models of anxiety-like behavior (open field, elevated plus maze and defensive withdrawal test). These data show the mutant congenic rat, of which GPR10 neither binds nor responds to PrRP, expresses less anxious-like phenotypes. On the basis of these observations, the GPR10 might be a novel target for the developing new drugs against anxiety and/or other stress-related diseases. © 2007 Elsevier B.V. All rights reserved. Keywords: Diabetes mellitus OLETF type 1 (Dmo1) G-protein-coupled receptor 10 (GPR10) Otsuka Long–Evans Tokushima Fatty (OLETF) rat Prolactin-releasing peptide Less anxious behavior BRAIN RESEARCH 1178 (2007) 114 – 124 ⁎ Corresponding author. Second Institute of New Drug Discovery, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, Japan. Fax: +81 88 665 6106. E-mail address: [email protected] (A. Watanabe). Abbreviations: BN, Brown–Norway; BST, bed nucleus of stria terminalis; CCK, cholecystokinin; CCKAR, CCK-A receptor; CeA, central nucleus of amygdala; CRH, corticotropin releasing hormone; Dmo1, diabetes mellitus OLETF type 1; F344, Fischer 344; HPA axis, hypothalamic–pituitary–adrenal axis; i.c.v., intracerebroventricular; IR, immunoreactivity; KO, knockout; NTS, nucleus of the solitary tract; OLETF, Otsuka Long–Evans Tokushima Fatty; Pev, periventricular nucleus; PrRP, prolactin-releasing peptide; PVN, paraventricular nucleus; QTL, quantitative trait loci 0006-8993/$ – see front matter © 2007 Elsevier B.V. All rights reserved. doi:10.1016/j.brainres.2007.07.075 available at www.sciencedirect.com www.elsevier.com/locate/brainres
Transcript of Altered emotional behaviors in the diabetes mellitus OLETF type 1 congenic rat

B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
ava i l ab l e a t www.sc i enced i rec t . com
www.e l sev i e r. com/ loca te /b ra in res
Research Report
Altered emotional behaviors in the diabetes mellitusOLETF type 1 congenic rat
Akihito Watanabea,b,⁎, Shiro Okunoc, Mai Okanob, Shaun Jordane,Koutoku Aiharac, Takeshi K. Watanabed, Yuki Yamasakib,Hisashi Kitagawab, Kiyoshi Sugawaraa, Satoru Katoa
aDepartment of Molecular Neurobiology, Graduate School of Medicine, Kanazawa University, Kanazawa, 920-8640, JapanbSecond Institute of New Drug Discovery, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, JapancQuests Research Institute, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, JapandOtsuka GEN Research Institute, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, JapaneDepartment of Neuroscience Research, Otsuka Maryland Medicinal Laboratories, 9900 Medical Center Drive, Rockville, Maryland, USA
A R T I C L E I N F O
⁎ Corresponding author. Second Institute of Ne665 6106.
E-mail address: a_watanabe@otsuka-euroAbbreviations: BN, Brown–Norway; BST, be
nucleus of amygdala; CRH, corticotropinhypothalamic–pituitary–adrenal axis; i.c.v.,tract; OLETF, Otsuka Long–Evans Tokushimanucleus; QTL, quantitative trait loci
0006-8993/$ – see front matter © 2007 Elsevdoi:10.1016/j.brainres.2007.07.075
A B S T R A C T
Article history:Accepted 28 July 2007Available online 15 August 2007
GPR10 is a G-protein-coupled receptor expressed in thalamic and hypothalamic brain regions,including the reticular thalamic nucleus (RTN) and periventricular nucleus (Pev), and theendogenous ligand for this receptor, prolactin-releasing peptide (PrRP), has demonstratedregulatory effects on the stress response. We produced a congenic rat by introducing the Dmo1allele from the OLETF rat which encodes the amino acid sequences of GPR10 with a truncatedNH2-terminus, into the Brown–Norway background. Using receptor autoradiography, wedetermined a lack of specific [125I]PrRP binding in the RTN and Pev of these mutant ratscompared to the control rats. Furthermore, intracerebroventricular injection of PrRP did notinducea significant increaseof c-fos-like immunoreactivity in theparaventricularnucleus of themutant rats compared to the control rats. The mutant rats also displayed a less anxious-likephenotype in three behavioral-basedmodels of anxiety-like behavior (open field, elevated plusmazeanddefensivewithdrawal test). These datashow themutant congenic rat, ofwhichGPR10neither binds nor responds to PrRP, expresses less anxious-like phenotypes. On the basis ofthese observations, the GPR10 might be a novel target for the developing new drugs againstanxiety and/or other stress-related diseases.
© 2007 Elsevier B.V. All rights reserved.
Keywords:Diabetes mellitus OLETFtype 1 (Dmo1)G-protein-coupled receptor10 (GPR10)Otsuka Long–Evans TokushimaFatty (OLETF) ratProlactin-releasing peptideLess anxious behavior
wDrug Discovery, Otsuka Pharmaceutical Co., Ltd., Tokushima, 771-0192, Japan. Fax: +81 88
pe.com (A. Watanabe).d nucleus of stria terminalis; CCK, cholecystokinin; CCKAR, CCK-A receptor; CeA, centralreleasing hormone; Dmo1, diabetes mellitus OLETF type 1; F344, Fischer 344; HPA axis,intracerebroventricular; IR, immunoreactivity; KO, knockout; NTS, nucleus of the solitaryFatty; Pev, periventricular nucleus; PrRP, prolactin-releasing peptide; PVN, paraventricular
ier B.V. All rights reserved.

115B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
1. Introduction
The Otsuka Long–Evans Tokushima Fatty (OLETF) rat strain is auseful model of obese type II diabetes, which is hypertriglycae-mic, hyperinsulinaemic, hyperglycaemic, insulin resistant andhas abundant visceral fat (Kawano et al., 1992). Genome-widequantitative trait loci (QTL) analyses have identified 12 statisti-cally significantQTL, termedDmo1 (The rat genomedata base ID:71112) through Dmo12, which were associated with the type IIdiabetes, obesity and dyslipidaemia phenotypes of the maleOLETF rat (Kanemoto et al., 1998; Okuno et al., 1999; Watanabeet al., 1999; Yamasaki et al., 2000). Diabetes mellitus OLETF type 1(Dmo1) on the long arm of rat chromosome 1 was a major QTL,exerting significant effects on body weight, visceral fat deposi-tion, and plasma triglyceride levels. Congenic lines for the Dmo1locus have been established by introducing nondiabetic Dmo1alleles from the Brown–Norway (BN) (Watanabe et al., 2001) andFischer 344 (F344) rats (Okuno et al., 2001) into the OLETF ratbackground. The established congenic lines, which are hetero-zygous for the nondiabetic Dmo1 allele, showed significantimprovements in obesity, dyslipidemia, hyperinsulinaemia,hyperglycaemia and glucose-intolerance phenotypes in eachgeneration, and these data have been confirmed with homozy-gous congenic rats (Okuno et al., 2002; Watanabe et al., 2005).
In an attempt to identify the gene(s) responsible for theDmo1phenotype, we generated congenic sub-lines, analyzed thecorrelation between their genotypes and obesity phenotypes,and further narrowed down the responsible region. Extensiveinvestigation, using positional cloning and the positionalcandidate approach, discovered a single nucleotide change atthe start codon of the GPR10 gene (Watanabe et al., 2005). The
Fig. 1 – Inheritance of chromosome and GPR10 genotype on chromshowing allelic inheritance at Dmo1 locus in mutant rats. Black rhomozygous OLETF allele, and gray represents the recombinatioanalyzed. (B) Inheritance of the parental chromosome in the conrepresents the OLETF chromosome.
ATG translation initiation codon of theGPR10 genewasmutatedtoATA in theOLETF strain. Thismutationwould encode aGPR10amino acid sequences with an NH2-terminus truncation, sug-gesting a functional impairment of the GPR10 in the OLETF rat.We previously reported that the OLETF rat strain possessed thehomozygous Dmo1 gene mutation, and the congenic rats whichwere replaced Dmo1 locus from OLETF allele to F344 or BN wild-type allele resulted in a decrease ofmanifestation of obesity anddiabetes in the OLETF genetic background (Okuno et al., 2001,2002;Watanabe et al., 2001). To explore another function ofDmo1locus, we tried to generate complementary congenic BN rats, toenable us to observe behavioral changes in different paradigmsby removing confounding factors of obesity- and diabetes-related phenotypes. Similarly to the report of GPR10 knockout(KO)mice (Guetal., 2004), the congenic ratsof theOLETFmutatedGPR10gene in theBNbackgroundhadnoeffect onbodyweight inyounger animals studied at 6 to 10 weeks old.
The GPR10 is a G-protein-coupled receptor, which wasidentified by screening a genomic DNA library (Marchese et al.,1995). In the rat, GPR10 is expressed abundantly in the thalamus,hypothalamus, anterior pituitary and adrenal medulla, whereasits endogenous ligand, prolactin-releasing peptide (PrRP)(Hinuma et al., 1998), is expressed mainly in the hypothalamus,medulla oblongata and intestine (Fujii et al., 1999; Roland et al.,1999). The distribution of PrRP nerve fiber is widespread in thebrain including theparaventricularnucleus (PVN)andsupraopticnucleus (SON), paratenial thalamic nucleus, basolateral amyg-daloid nucleus and bed nucleus of the stria terminalis (BST)(Maruyama et al., 1999a). The distribution profile indicates thatPrRP plays a wide range of roles in the brain. Intracerebroven-tricular (i.c.v.) injection of PrRP in rats reduces food intake and
osome 1 inmutant rats. (A) Representation of chromosome 1epresents the homozygous BN allele, white represents then regions. Each box indicates the representative SSLP markergenic rat. Black represents the BN chromosome and white

Fig. 2 – Binding of [125I]PrRP-20 in coronal brain sections ofcontrol rat andmutant rat. Autoradiographs are at the levelof reticular thalamic neucleus (RT). Sections were incubated inbinding buffer containing 200 pM [125I] PrRP-20 in the absence(A, C) or presence (B, D) of 200 nM unlabeled PrRP-31. (A) Strongbindings were observed in RT, the peri-third ventricular areaand optic chiasm in the section of control rat. (B) Specificbindings were reduced in the presence of excess of unlabeledPrRP-31. (C, D) Strong biding was not observed in the sectionof the mutant rat. Similar results were obtained in 3 othercontrol and mutant rats, respectively. Scale bar: 3 mm.
116 B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
body weight (Lawrence et al., 2000, 2002), increases plasmaoxytocin (Maruyama et al., 1999b) and ACTH (Matsumoto et al.,2000; Seal et al., 2002), and promotes wakefulness (Zhang et al.,2000). Moreover, PrRP has been found to control the secretion ofACTH, throughcorticotropin releasinghormone (CRH), andPrRP-containing neurons are known to be activated by severalstressful conditions in rats, indicating a likely role of PrRP onregulating the stress axis (Maruyamaet al., 2001). PrRPmight alsobe involved in autonomic control of stress/fear conditioning,because GPR10 is found in the BST, the central nucleus ofamygdala (CeA) and the PVN, and the PrRP cell bodies are foundin the catecholaminergic neurons of the nucleus of the solitarytract (NTS) which projects to most of these regions.
In the present study, a homozygous congenic rat wasgenerated by introducing the OLETF Dmo1 allele (mutatedGPR10 gene) into the BN rat background. To confirm loss ofligand binding to mutated GPR10, if expressed, we determinedloss of signals in receptor autoradiography using [125I]PrRP in themutant Dmo1 congenic rat. For the further confirmation of thefunctional loss of mutant GPR10, we also i.c.v. injected PrRP andobserved reduced c-fos activation of neurons in the congenic ratswith mutant Dmo1. In addition, we evaluated measures ofanxiety in the mutant Dmo1 congenic rat in the open field test,elevated plus maze test and defensive withdrawal test. Suchtests are based on thenatural neophobia of rodents, andused forthe evaluation of the anxiolytic activity (Heinrichs et al., 1997;Liebsch et al., 1995, 1999; Skutella et al., 1998).We found that thismutant Dmo1 congenic rats had a significantly less anxiousphenotype than control rats with the wild-type Dmo1.
2. Results
2.1. Preparation of congenic rat
We generated congenic rats for the Dmo1 locus, a 28.8 cM regionfrom the marker D1Rat169 to D1Rat90 that contains the GPR10gene locus (Fig. 1A). Female OLETF rats were crossed with BNstrain. Subsequently, male offspring was backcrossed to the BNrats. In each generation, offspring that inherited the OLETFDmo1allelewas selectedand further genotypedover thewholegenometo select rats with high replacement of the BN background. Thismarker-assisted speed congenic procedure resulted in 97–99%replacement rates out of Dmo1 region for the BN backgroundgenome at the fourth generation (BC4). Selected BC4 offsprings,heterozygote for theDmo1 region,were intercrossed to eachotherand obtained homozygosity for the Dmo1 locus. The first orsubsequent generation of progenies of the Dmo1 homozygoteswere checked for the GPR10 SNP (single nucleotide polymor-
Table 1 – Body weight change for control and mutant rats
Rats 6 weeks 7 weeks 8 weeks 9 weeks 10 weeks
Control(N=10)
115.8±3.7 140.8±5.2 152.4±4.1 198.4±4.4 223.1±5.3
Mutant(N=8)
105.7±5.6 133.8±7.5 142.6±8.1 182.8±8.3 203.9±8.6
Values represent means±S.E.M. for 10 control rats and 8 mutantrats, respectively.
phism) genotype and used in this study. Hereafter the congenicrats with wild-type Dmo1 are termed control rats, and thecongenic ratswithmutantDmo1 are termedmutant rats (Fig. 1B).
2.2. Body weight change of the mutant rat
Themutant rat did not show thedifference in bodyweight at 6 to10weeks old comparedwith the control rat (Table 1). In addition,recent generation of mutant rat neither showed difference inbody weight at 8 to 18 weeks old (at 18 weeks; control (N=14):334.7±4.8 g, mutant (N=14): 326.7±4.3 g [mean±S.E.M.]).
2.3. Lack of [125I]PrRP binding in the mutant rat brain
Anautoradiographicanalysis revealeddense [125I]PrRPbinding inthe reticular thalamic nucleus (RT), periventricular nucleus (Pev)and optic chiasm in control (Fig. 2A). This [125I]PrRP binding wasspecificandreversible, as itwasdisplacedbycoldPrRP (Fig. 2B). Incontrast, no specific [125I]PrRP binding was detected in the samebrain regionsofmutant rats (Fig. 2C), asno change indensitywasobserved by adding cold PrRP in the brain ofmutant rats (Fig. 2D).
2.4. Reduction of c-fos signals by PrRP in themutant rat brain
c-Fos immunoreactivity (IR) was increased (P<0.01) in controlrats after i.c.v. injection of 40 nmol PrRP (Figs. 3B and E). Incontrast, mutant rats exhibit a small number of c-fos-positivecells after injection of 40 nmol PrRP (Figs. 3D and E). Fig. 3E showsthenumber of c-fos IR cells in theparvocellular regionof the PVN.Therewas a significant interaction anda significant difference ofoverall means between control and mutant rats (F(1,16)=20.73;P<0.01,F(1,16)=17.93;P<0.01). Inaddition, therewasa significant
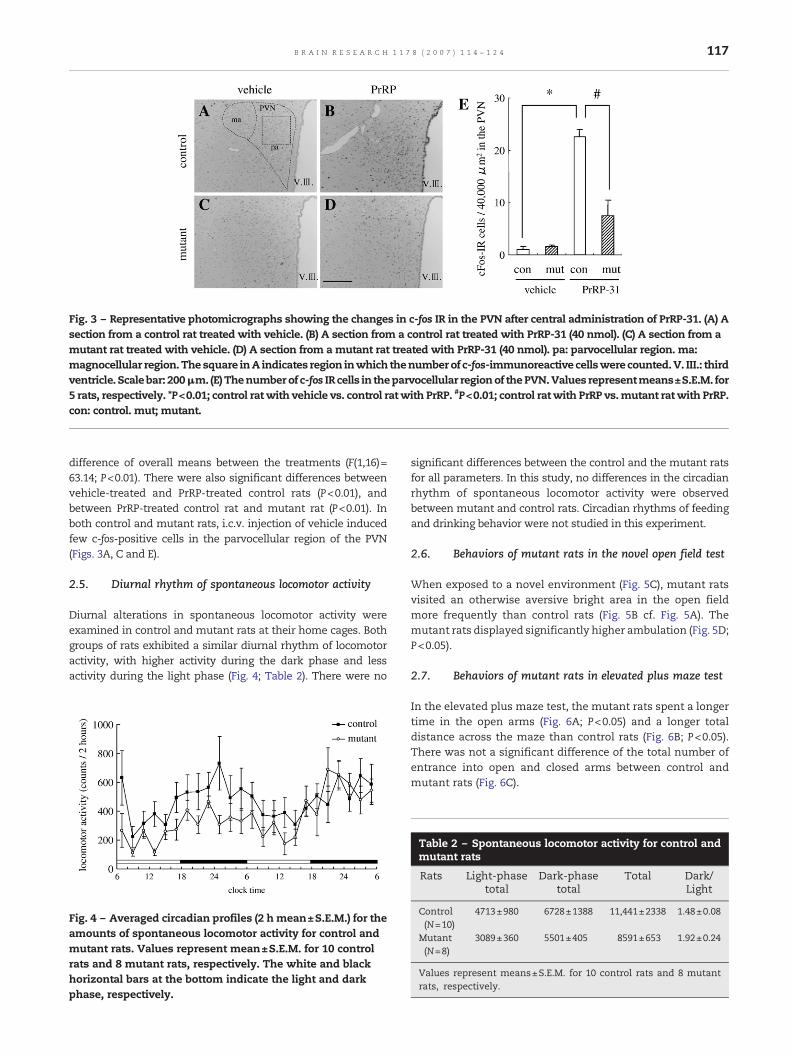
Fig. 3 – Representative photomicrographs showing the changes in c-fos IR in the PVN after central administration of PrRP-31. (A) Asection from a control rat treated with vehicle. (B) A section from a control rat treated with PrRP-31 (40 nmol). (C) A section from amutant rat treated with vehicle. (D) A section from amutant rat treated with PrRP-31 (40 nmol). pa: parvocellular region. ma:magnocellular region.Thesquare inA indicates region inwhich thenumberof c-fos-immunoreactive cellswere counted.V. III.: thirdventricle. Scalebar: 200μm. (E) Thenumberof c-fos IR cells in theparvocellular regionof thePVN.Values representmeans±S.E.M. for5 rats, respectively. *P<0.01; control rat with vehicle vs. control ratwith PrRP. #P<0.01; control ratwith PrRP vs.mutant rat with PrRP.con: control. mut; mutant.
117B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
difference of overall means between the treatments (F(1,16)=63.14; P<0.01). There were also significant differences betweenvehicle-treated and PrRP-treated control rats (P<0.01), andbetween PrRP-treated control rat and mutant rat (P<0.01). Inboth control and mutant rats, i.c.v. injection of vehicle inducedfew c-fos-positive cells in the parvocellular region of the PVN(Figs. 3A, C and E).
2.5. Diurnal rhythm of spontaneous locomotor activity
Diurnal alterations in spontaneous locomotor activity wereexamined in control and mutant rats at their home cages. Bothgroups of rats exhibited a similar diurnal rhythm of locomotoractivity, with higher activity during the dark phase and lessactivity during the light phase (Fig. 4; Table 2). There were no
Fig. 4 – Averaged circadian profiles (2 h mean±S.E.M.) for theamounts of spontaneous locomotor activity for control andmutant rats. Values represent mean±S.E.M. for 10 controlrats and 8 mutant rats, respectively. The white and blackhorizontal bars at the bottom indicate the light and darkphase, respectively.
significant differences between the control and the mutant ratsfor all parameters. In this study, no differences in the circadianrhythm of spontaneous locomotor activity were observedbetween mutant and control rats. Circadian rhythms of feedingand drinking behavior were not studied in this experiment.
2.6. Behaviors of mutant rats in the novel open field test
When exposed to a novel environment (Fig. 5C), mutant ratsvisited an otherwise aversive bright area in the open fieldmore frequently than control rats (Fig. 5B cf. Fig. 5A). Themutant rats displayed significantly higher ambulation (Fig. 5D;P<0.05).
2.7. Behaviors of mutant rats in elevated plus maze test
In the elevated plus maze test, the mutant rats spent a longertime in the open arms (Fig. 6A; P<0.05) and a longer totaldistance across the maze than control rats (Fig. 6B; P<0.05).There was not a significant difference of the total number ofentrance into open and closed arms between control andmutant rats (Fig. 6C).
Table 2 – Spontaneous locomotor activity for control andmutant rats
Rats Light-phasetotal
Dark-phasetotal
Total Dark/Light
Control(N=10)
4713±980 6728±1388 11,441±2338 1.48±0.08
Mutant(N=8)
3089±360 5501±405 8591±653 1.92±0.24
Values represent means±S.E.M. for 10 control rats and 8 mutantrats, respectively.
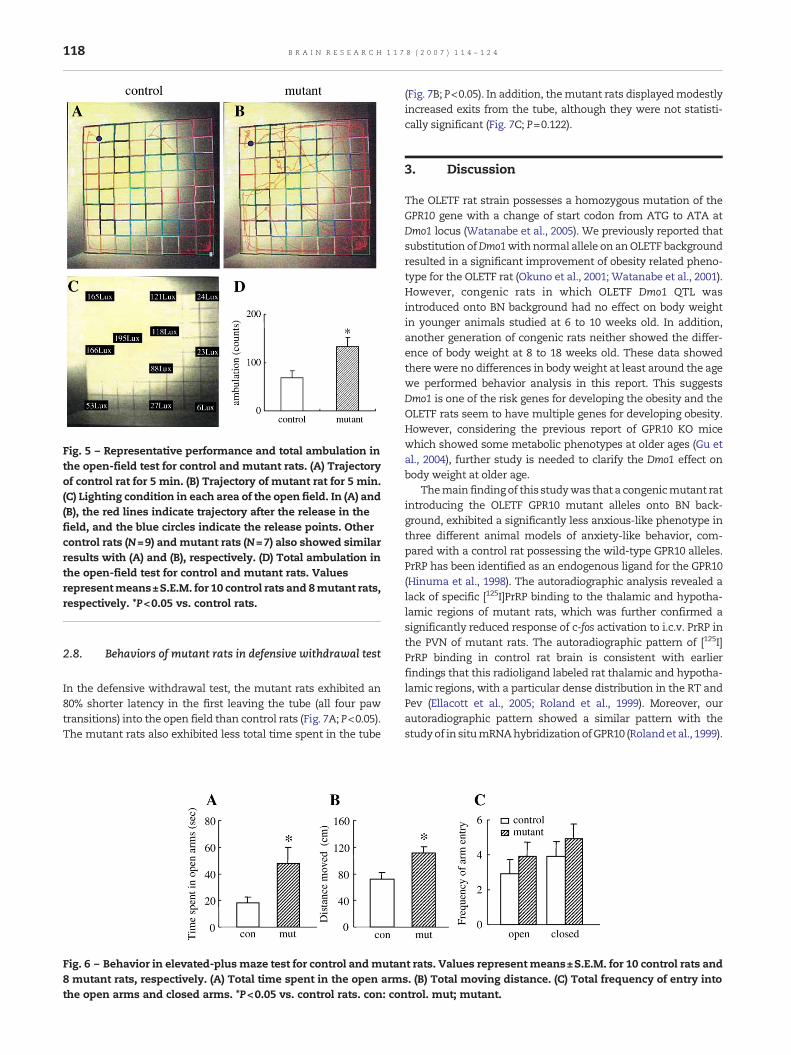
Fig. 5 – Representative performance and total ambulation inthe open-field test for control and mutant rats. (A) Trajectoryof control rat for 5 min. (B) Trajectory of mutant rat for 5 min.(C) Lighting condition in each area of the open field. In (A) and(B), the red lines indicate trajectory after the release in thefield, and the blue circles indicate the release points. Othercontrol rats (N=9) andmutant rats (N=7) also showed similarresults with (A) and (B), respectively. (D) Total ambulation inthe open-field test for control and mutant rats. Valuesrepresentmeans±S.E.M. for 10 control rats and 8mutant rats,respectively. *P<0.05 vs. control rats.
118 B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
2.8. Behaviors of mutant rats in defensive withdrawal test
In the defensive withdrawal test, the mutant rats exhibited an80% shorter latency in the first leaving the tube (all four pawtransitions) into the open field than control rats (Fig. 7A; P<0.05).The mutant rats also exhibited less total time spent in the tube
Fig. 6 – Behavior in elevated-plus maze test for control andmutan8 mutant rats, respectively. (A) Total time spent in the open armthe open arms and closed arms. *P<0.05 vs. control rats. con: co
(Fig. 7B; P<0.05). In addition, themutant rats displayedmodestlyincreased exits from the tube, although they were not statisti-cally significant (Fig. 7C; P=0.122).
3. Discussion
The OLETF rat strain possesses a homozygous mutation of theGPR10 gene with a change of start codon from ATG to ATA atDmo1 locus (Watanabe et al., 2005). We previously reported thatsubstitution ofDmo1withnormal allele on anOLETF backgroundresulted in a significant improvement of obesity related pheno-type for the OLETF rat (Okuno et al., 2001; Watanabe et al., 2001).However, congenic rats in which OLETF Dmo1 QTL wasintroduced onto BN background had no effect on body weightin younger animals studied at 6 to 10 weeks old. In addition,another generation of congenic rats neither showed the differ-ence of body weight at 8 to 18 weeks old. These data showedthere were no differences in bodyweight at least around the agewe performed behavior analysis in this report. This suggestsDmo1 is one of the risk genes for developing the obesity and theOLETF rats seem to have multiple genes for developing obesity.However, considering the previous report of GPR10 KO micewhich showed some metabolic phenotypes at older ages (Gu etal., 2004), further study is needed to clarify the Dmo1 effect onbody weight at older age.
Themain findingof this studywas that a congenicmutant ratintroducing the OLETF GPR10 mutant alleles onto BN back-ground, exhibited a significantly less anxious-like phenotype inthree different animal models of anxiety-like behavior, com-pared with a control rat possessing the wild-type GPR10 alleles.PrRP has been identified as an endogenous ligand for the GPR10(Hinuma et al., 1998). The autoradiographic analysis revealed alack of specific [125I]PrRP binding to the thalamic and hypotha-lamic regions of mutant rats, which was further confirmed asignificantly reduced response of c-fos activation to i.c.v. PrRP inthe PVN of mutant rats. The autoradiographic pattern of [125I]PrRP binding in control rat brain is consistent with earlierfindings that this radioligand labeled rat thalamic and hypotha-lamic regions, with a particular dense distribution in the RT andPev (Ellacott et al., 2005; Roland et al., 1999). Moreover, ourautoradiographic pattern showed a similar pattern with thestudyof in situmRNAhybridizationofGPR10 (Rolandet al., 1999).
t rats. Values represent means±S.E.M. for 10 control rats ands. (B) Total moving distance. (C) Total frequency of entry intontrol. mut; mutant.
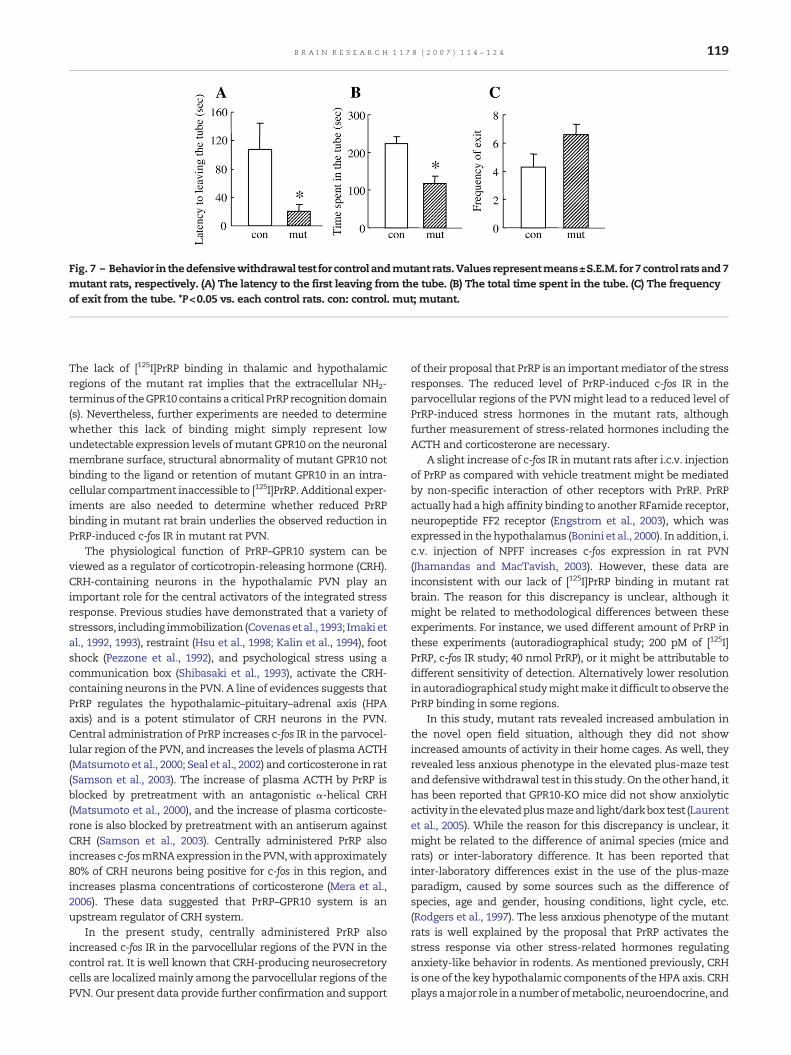
Fig. 7 – Behavior in thedefensivewithdrawal test for control andmutant rats.Values representmeans±S.E.M. for 7control ratsand7mutant rats, respectively. (A) The latency to the first leaving from the tube. (B) The total time spent in the tube. (C) The frequencyof exit from the tube. *P<0.05 vs. each control rats. con: control. mut; mutant.
119B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
The lack of [125I]PrRP binding in thalamic and hypothalamicregions of the mutant rat implies that the extracellular NH2-terminusof theGPR10containsacritical PrRP recognitiondomain(s). Nevertheless, further experiments are needed to determinewhether this lack of binding might simply represent lowundetectable expression levels ofmutant GPR10 on the neuronalmembrane surface, structural abnormality of mutant GPR10 notbinding to the ligand or retention of mutant GPR10 in an intra-cellular compartment inaccessible to [125I]PrRP. Additional exper-iments are also needed to determine whether reduced PrRPbinding in mutant rat brain underlies the observed reduction inPrRP-induced c-fos IR in mutant rat PVN.
The physiological function of PrRP–GPR10 system can beviewed as a regulator of corticotropin-releasing hormone (CRH).CRH-containing neurons in the hypothalamic PVN play animportant role for the central activators of the integrated stressresponse. Previous studies have demonstrated that a variety ofstressors, including immobilization (Covenasetal., 1993; Imaki etal., 1992, 1993), restraint (Hsu et al., 1998; Kalin et al., 1994), footshock (Pezzone et al., 1992), and psychological stress using acommunication box (Shibasaki et al., 1993), activate the CRH-containing neurons in the PVN. A line of evidences suggests thatPrRP regulates the hypothalamic–pituitary–adrenal axis (HPAaxis) and is a potent stimulator of CRH neurons in the PVN.Central administration of PrRP increases c-fos IR in the parvocel-lular region of the PVN, and increases the levels of plasma ACTH(Matsumoto et al., 2000; Seal et al., 2002) and corticosterone in rat(Samson et al., 2003). The increase of plasma ACTH by PrRP isblocked by pretreatment with an antagonistic α-helical CRH(Matsumoto et al., 2000), and the increase of plasma corticoste-rone is also blocked by pretreatment with an antiserum againstCRH (Samson et al., 2003). Centrally administered PrRP alsoincreases c-fosmRNAexpression in the PVN,with approximately80% of CRH neurons being positive for c-fos in this region, andincreases plasma concentrations of corticosterone (Mera et al.,2006). These data suggested that PrRP–GPR10 system is anupstream regulator of CRH system.
In the present study, centrally administered PrRP alsoincreased c-fos IR in the parvocellular regions of the PVN in thecontrol rat. It is well known that CRH-producing neurosecretorycells are localizedmainly among the parvocellular regions of thePVN. Our present data provide further confirmation and support
of their proposal that PrRP is an importantmediator of the stressresponses. The reduced level of PrRP-induced c-fos IR in theparvocellular regions of the PVNmight lead to a reduced level ofPrRP-induced stress hormones in the mutant rats, althoughfurther measurement of stress-related hormones including theACTH and corticosterone are necessary.
A slight increase of c-fos IR inmutant rats after i.c.v. injectionof PrRP as compared with vehicle treatment might be mediatedby non-specific interaction of other receptors with PrRP. PrRPactually had a high affinity binding to another RFamide receptor,neuropeptide FF2 receptor (Engstrom et al., 2003), which wasexpressed in thehypothalamus (Bonini et al., 2000). In addition, i.c.v. injection of NPFF increases c-fos expression in rat PVN(Jhamandas and MacTavish, 2003). However, these data areinconsistent with our lack of [125I]PrRP binding in mutant ratbrain. The reason for this discrepancy is unclear, although itmight be related to methodological differences between theseexperiments. For instance, we used different amount of PrRP inthese experiments (autoradiographical study; 200 pM of [125I]PrRP, c-fos IR study; 40 nmol PrRP), or it might be attributable todifferent sensitivity of detection. Alternatively lower resolutioninautoradiographical studymightmake it difficult toobserve thePrRP binding in some regions.
In this study, mutant rats revealed increased ambulation inthe novel open field situation, although they did not showincreased amounts of activity in their home cages. As well, theyrevealed less anxious phenotype in the elevated plus-maze testanddefensivewithdrawal test in this study.On theother hand, ithas been reported that GPR10-KO mice did not show anxiolyticactivity in theelevatedplusmazeand light/darkbox test (Laurentet al., 2005). While the reason for this discrepancy is unclear, itmight be related to the difference of animal species (mice andrats) or inter-laboratory difference. It has been reported thatinter-laboratory differences exist in the use of the plus-mazeparadigm, caused by some sources such as the difference ofspecies, age and gender, housing conditions, light cycle, etc.(Rodgers et al., 1997). The less anxious phenotype of the mutantrats is well explained by the proposal that PrRP activates thestress response via other stress-related hormones regulatinganxiety-like behavior in rodents. As mentioned previously, CRHis one of the keyhypothalamic components of theHPAaxis. CRHplaysamajor role inanumberofmetabolic, neuroendocrine, and

120 B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
autonomic adaptations to stress (de Kloet et al., 2005; Stecklerand Holsboer, 1999). The CRF-R1 receptor subtype is highlyexpressed in the anterior pituitary, neocortex, hippocampus,amygdala, and cerebellum (Chalmers et al., 1995; De Souza, 1995;Van Pett et al., 2000). This receptor takes part in the activation ofHPA system and a major mediator of emotionality in the limbicsystem (Preil et al., 2001; Smith et al., 1998; Timpl et al., 1998).Numerous investigations in animals have described anxiogenic-like effects after central administration of CRF via CRF-R1 (Keckand Holsboer, 2001). For example, inhibition of CRF-R1 expres-sion, by central administration of CRF-R1 antisense oligo deoxy-nucleotides, has an anxiolytic outcome in rodents (Heinrichset al., 1997; Liebsch et al., 1995, 1999; Skutella et al., 1998). Simi-larly, CRF-R1 antagonists are anxiolytic in most but not all an-xiety paradigms of rodent (Griebel et al., 1998; Keck et al., 2001),and the human, CRF-R1 receptor has been proposed as apromising target for the development of antidepressant oranxiolytic drugs (Saunders and Williams, 2003; Zorrilla andKoob, 2004).
PrRP-producing catecholaminergic neurons of the NTS proj-ect to the BST, CeA and the PVN, where GPR10 is highlyexpressed. Whereas only small percentage of cells in theseareas are found to co-express CRH andGPR10 (Lin et al., 2002; Linand Civelli, 2004), approximately 80% of CRH neurons in the PVNwere activated after i.c.v. injection of PrRP as mentioned above(Mera et al., 2006). The data together suggest that a loss of GPR10functionmodulates the sensitivity of CRH neurons to emotionalstress, which in turn results in less-anxious behavior in rat. Thebehavioral data present herein are in this line.
We conclude, in this report, that Dmo1 locus confers lessanxiety-like behavior whereas it is reported that OLETF strainshowed anxiogenic-like behavior compared with LETO rats (Liet al., 2002; Yamamoto et al., 2000). We believe those previousreports cannot attribute the observed increased anxiety-likebehaviors of OLETF strain to any specific genetic components.Since the genetic difference between OLETF and LETO strainsare as much as 38% (Watanabe et al., 2000), any observeddifferences between these distant two strains should not beassigned to any specific genes including currently knownmutations in cholecystokinin(CCK)-A receptor and GPR10genes instantly (Takiguchi et al., 1997; Watanabe et al., 2005).It was reported that a potent and selective CCK-A receptor(CCKAR) antagonist of IQM-95,333 showed anxiolytic-likephenotype both in vitro and in vivo in two different animalmodels (Ballaz et al., 1997). In addition, other reports showedthat CCKAR antagonists are anxiolytic (Bickerdike et al., 1994;Crawley, 1991; Hendrie et al., 1993). These data suggested thatusing such distant strains were not useful to identify thegenetic components for observed strain differences withoutpharmacological studies. In our experiments according to BN-based congenics, 97% to 99% of genomic regions are the samebetween mutants and controls, which would allow us toassign the observed anxiolytic-like effect to Dmo1 locus. Thesedata suggested that GPR10 mutation was likely to explain theless anxiolytic-like phenotypes. However, we still must becareful of the possibility that residual genetic factors contrib-ute to the observed phenotypes. Further study with specificGPR10 antagonist would help clarify this point. The presentdata collectively indicate that Dmo1 locus plays an importantrole in the stress response.
Finally, it is interesting to see anxiogenic phenotypes ofOLETF strain in light of genetic components. OLETF strain lacksfunctions of two relatedG-protein-coupled receptor expressed inthe central nervous system: CCKAR and GPR10 (Takiguchi et al.,1997;Watanabeet al., 2005). BothCCKandPrRPhavebeenshownto increase plasma levels of ACTH and corticosterone in rats(Kamilaris et al., 1992; Katsuura et al., 1992), and the effect wasinhibited by the CCKAR antagonist (Katsuura et al., 1992). Itsuggests that CCKAR and GPR10 are functionally related on thesame pathway. Moreover, CCK was known as upstream regula-tors of PrRP production (Lawrence et al., 2002). These findings arealso supported by the data that CCK failed to increase plasmacorticosterone levels in GPR10−/− animals (Bechtold and Luck-man, 2006). From the anatomical view points, these twoneuroendocrine systems are also interconnected. CCKARs arelocalized in NTS (Hill et al., 1987; Honda et al., 1993), where themajority of PrRP-expressing neurons are localized. In addition,administrationofCCK tomice leads to c-fos induction in thePVN,and the increase was inhibited by the CCKAR antagonist (Wanget al., 1998).These report indicate that PrRP neuronsmay serve torelay CCK-initiated signals to higher brain centers including thePVN and amygdala, and CCKAR could be involved in the signaltransduction.
Considering CCKAR antagonist showed anxiolytic-like phe-notypes in two different animal models (Ballaz et al., 1997) andDmo1 locus represented by GPR10 mutation confers anxiolytic-like phenotypes, these two mutations might cooperativelyproduce more anxiolytic-like phenotypes. In this viewpoint, it isinteresting that OLETF strain showed anxiogenic-like behaviorcompared with LETO rats (Li et al., 2002; Yamamoto et al., 2000)with the genetic difference as much as 38% (Watanabe et al.,2000). Simply, these data suggested that quite strong anxiogenicgenetic components are in the genomic regions of OLETF strainand surpass the anxiolytic effects of CCKAR mutation and Dmo1locus, although the data with distant strains must be carefullyexamined.
In conclusion, the present data indicate that Dmo1 locusrepresented by GPR10 mutation plays an important role in thestress response, and it leads to the anti stress/anxiety likebehavior. Therefore Dmo1 allele, especially GPR10, might be anovel target for the developing newdrugs against anxiety and/orother stress-related diseases.
4. Experimental procedures
4.1. Preparation of mutant and control congenic rats
BN male rats were purchased from Charles River Japan andOLETF female rats were obtained from Laboratory of AnimalResources, Otsuka Pharmaceutical. For congenic breeding,female OLETF rats were crossed with male BN rats reciprocallyto produce an F1 population. This F1 population was subse-quently back-crossed to the BN strain (BN×F1) to make a BC1population, which was successively backcrossed in the samemanner. Genomic DNAwas isolated from the tail tip of BC1, andsubsequent BCn populations, and was analyzed using micro-satellitemarkers that distinguish the BN andOLETF alleles at theDmo1 locus (D1Rat169–D1Rat90) (Watanabe et al., 2000). Animalsheterozygous at theDmo1 locus were selected and subsequently

121B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
genotypedover thewholegenome to select individualswithhighreplacement rates to the BN background. This marker-assistedspeed congenic procedure resulted in 97% to 99% replacementout of the Dmo1 region to the BN background genome at thefourth generation (BC4). After four generations of backcrossing, apair of homozygotes for theDmo1 locuswas generated bymatingheterozygotes (BC4:F1). The allelic inheritance of congenic ratson chromosome 1 is presented in Fig. 1A. Subsequently, animalsweremaintainedby intercrossing.At this timepoint,wedetectedthe SNP in the putative start codon of GPR10 gene within theDmo1 locus (Watanabe et al., 2005). Heterozygous animals attheGPR10genewere intercrossed toobtainmutant (homozygousmutated GPR10) and control (homozygous wild-type GPR10) rats(Fig. 1B). First or subsequent generations of progeny of the Dmo1homozygotewere checked for theGPR10 SNP genotype and usedfor subsequent experimentations. In addition, CCKAR genotypeswere examined using internal and external primers of 6.9 kbdeletion region according to the previous report (Takiguchi et al.,1998), and found that all animals used in this study hadhomozygously BN-type normal allele. In all experiments, ratswere used at 6–10 weeks old.
All rats were kept in individual cages before use in eachexperiment. Ratsweremaintained in a 12 h:12 h light/dark cycle,humidity- (40–70%) and temperature- (20–26 °C) controlledfacilities with standard rodent chow and water ad libitum. Allof the experiments described in the present study wereperformed using male rats during the light period according to“the Guidelines for Animal Care and Use at Otsuka Pharmaceu-tical October 1, 1994”.
4.2. Autoradiography of [125I]PrRP in rat brain sections
Theautoradiographic procedurewasdescribedpreviouslywith aslight modification (Kato et al., 1993). Animals were decapitatedby guillotine, and their brains were collected and immediatelyfrozenusingdry ice. A cryostatwasused to prepare coronal brainslices (20μm),whichweremountedontoAPS-coatedglass slides,dried for 1 h at 4 °C and stored dry at −20 °C until use. On the dayof the experiment, the sectionswere thawedanddried for 10minat room temperature, re-hydrated 15 min in washing buffer (1×PBS (pH 7.4) containing 0.1% BSA), and then incubated for 15mininbindingbuffer (50mMHEPES, 2mMMgCl2, 1mMEDTA,1μg/mlpepstatin A and 0.1% BSA). These sections were then incubatedfor 90min in binding buffer containing 200 pM [125I]human PrRP-20 (Amersham Biosciences UK limited., England) in the absenceor presence of 200 nM unlabeled human PrRP-31 (PeptideInstitute, Inc., Osaka, Japan). Following this incubation, thesections were washed three times for 5 min in ice-cold washingbuffer, rinsed briefly in distilled water, dried at room tempera-ture, and then exposed to an imaging plate (Fujifilm, Japan).Radioactivity bound to each section was visualized using a Bio-Imaging Analyzer BAS 5000 system (Fujifilm, Japan).
4.3. Surgical procedure of i.c.v. injection
Rats were injectedwith sodiumpentobarbital (50mg/kg i.p.) andplaced in a stereotaxic frame. A stainless steel guide cannula(22 gauge, Plastics One, USA)was stereotaxically implanted,withits tip projecting into the right lateral cerebral ventricle (internalcannula extends 1 mm below the guide cannula), using the
stereotaxic coordinates 0.8mmcaudal to bregma, 1.6mm lateralto the midline and 3.5 mm below the skull. The cannula wassecured to the skull with dental cement and each animal wasreturned to itshomecageafter surgery andallowed to recover forat least 7 days. The animals were individually housed in theirhome cage before the start of each experiment. Cannulaplacement was verified by a histological examination of eachanimal brain at the end of each study.
4.4. c-fos immunohistochemistry
Rats received an i.c.v. injection of rat PrRP-31 (40 nmol per rat,Peptide Institute, Inc., Osaka, Japan) or vehicle (0.1% BSA in PBS),were anesthetized 2 h later with sodiumpentobarbital (50mg/kgi.p.) and thenperfused transcardiallywith isotonic saline (150ml)followed by a fixative (150 ml, 10% formalin in PBS). Each brainwas postfixed in fixative for 72 h at room temperature and thendehydrated and embedded in paraffin. Coronal 4.5-μm-thicksectionswere cut at 90-μmintervals andmountedonAPS-coatedglass slides. Immunohistochemistry procedureswere performedwith the Discovery HX system (Ventana Medical Systems, Inc.,AZ, USA). Briefly, the slides were automatically subjected toantigen retrieval using EDTA buffer-based CC1 reagent (VentanaMedical Systems, Inc., AZ,USA) at setting 100 for 30minand thenincubatedwith anti-c-fos antibody (1:100; Santa Cruz) at 37 °C for30 min. Each slide was then incubated for 20 min at 37 °C insecondary antibody (anti-goat biotinylated IgG antibody, 1:250;Vector Laboratories, Inc., CA, USA) and then in horseradishperoxidase-conjugated streptavidin. Hydrogen peroxidase and3,3′-diaminobenzidine (DAB) were added to visualize the signalsand the cell nuclei were counterstained with hematoxylin.
Microscopic image capture and cell counting was performedby a blinded observer. Brain sectionswere viewedunder amicro-scope (Olympus, Tokyo, Japan) using aCCD camera (HamamatsuPhotonics K.K., Shizuoka, Japan) and photomicrographs werecaptured from anatomicallymatched sections usingMac SCOPCsoftware (MITANICORP. Japan) onanAppleMacintosh computersystem. For the quantitative analysis of c-fos-immunoreactivecells in the PVN, a previously reported method was used with aslight modification (Tobe et al., 2005). In the PVN, the parvocel-lular regionwasdeterminedaccording to theatlasof PaxinosandWatson (1986). In the parvocellular region of the PVN, c-fos-positive cells were counted bilaterally from two sections,separated by 90 μm and anatomically matched across animals.These were then averaged to obtain a single value, which wasexpressed as the mean number of c-fos IR cells/40,000 μm2.
4.5. Diurnal rhythm of spontaneous locomotor activity
The diurnal rhythm of spontaneous locomotor activity wasmeasured in thehomecage of each rat using an infrareddetector(Neuroscience Inc., Tokyo, Japan) located in a soundproof roomand spontaneous locomotor activity was measured over2 consecutive days.
4.6. Novel open field tests
The novel open field test was conducted as previously described(Bruhwyler etal., 1990)withaslightmodifications.Theapparatusconsisted of a gray plastic open field (120×120 cm2 arena;

122 B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
surrounded by a wall 60-cm high). The field was demarcated by15-cm squares, with 3-mm-wide black lines, and was lit by a 60-W incandescent lamp suspended 100 cm above the field. Theanimal was gently placed in a square just under the lamp, andallowed towalkabout andexplore freelyduringa 5-minperiod inthe open field. The ambulation (the number of times the animalcrossed the demarcation line) scores were recorded during thesubsequent 5-min period.
4.7. Elevated plus maze
The elevated plus maze test was conducted as previouslyreported (Pellow and File, 1986) with modifications. The appara-tus consisted of a plus-shaped platform elevated 60 cm from thefloor with two opposing closed arms (42×15 cm2) with 30-cm-high side and endwalls (5–16 lx) and two opposing armswith nowalls (open arms, 17–30 lx). Each animal was placed onto thecentral area (15×15 cm2) of themaze facing one of the open armsand monitored for 5 min. During the 5-min testing period, thenumber of entries into each arm, time spent in the open armandthe total path length were recorded and analyzed using SMARTsoftware (Bio Research Center, Japan).
4.8. Defensive withdrawal test
The defensive withdrawal test was conducted as previouslydescribed (Takahashi et al., 1989) with a slightmodification. Thistest was performed in a 35×35 cm2 black acrylic arena with 30-cm-high walls, and on the testing day the arena was lit to auniform intensity (700 lx). To begin the trial, each rat was placedwithin a small black-colored acrylic tube (2 lx, 10 cm in diam-eter×20 cm in length, closed at one end). The tube was thenplaced in the open fieldwith the open end facing the corner, andservedas thewithdrawal chamber. Each test lasted for5min,andan observer, blind to the rat strain being studied, recorded thelatency to leave the tube (first all four-paw transition from thetube into the arena), the frequency of exit from the tube and thetotal amount of time spent inside the tube.
4.9. Statistical analysis
All results were expressed as mean±S.E.M. for each group.Student's t-test or Aspin–Welch's t-test was used to detectstatistically significant differences between control and mutantrats studied in the spontaneous locomotor activity and elevatedplus maze studies. The data recorded in the open field test wereanalyzed using Student's t-test. The data recorded in thedefensive withdrawal test were analyzed using Wilcoxon'srank sum test. The results of the c-fos IR study were analyzedby two-way analysis of variance. Independent Student's t-test orAspin–Welch's t-test were also used to make specific groupcomparisons. A probability of less than 0.05 was consideredstatistically significant in every statistical analysis.
Acknowledgments
The authors appreciate Takahiro Sato and Syuichi Iwashita forgenotyping of congenic rat, andMari Kondo for the discussion.The authors also thank Tami Urano for her secretarial work.
R E F E R E N C E S
Ballaz, S., Barber, A., Fortuno, A., Del Rio, J., Martin-Martinez, M.,Gomez-Monterrey, I., Herranz, R., Gonzalez-Muniz, R.,Garcia-Lopez, M.T., 1997. Pharmacological evaluation ofIQM-95,333, a highly selective CCKA receptor antagonist withanxiolytic-like activity in animal models. Br. J. Pharmacol. 121,759–767.
Bechtold, D.A., Luckman, S.M., 2006. Prolactin-releasing peptidemediates cholecystokinin-induced satiety inmice. Endocrinology147, 4723–4729.
Bickerdike, M.J., Marsden, C.A., Dourish, C.T., Fletcher, A., 1994.The influence of 5-hydroxytryptamine re-uptake blockade onCCK receptor antagonist effects in the rat elevated zero-maze.Eur. J. Pharmacol. 271, 403–411.
Bonini, J.A., Jones, K.A., Adham, N., Forray, C., Artymyshyn, R.,Durkin,M.M., Smith, K.E., Tamm, J.A., Boteju, L.W., Lakhlani, P.P.,Raddatz, R., Yao, W.J., Ogozalek, K.L., Boyle, N., Kouranova, E.V.,Quan, Y., Vaysse, P.J., Wetzel, J.M., Branchek, T.A., Gerald, C.,Borowsky, B., 2000. Identification and characterization of two Gprotein-coupled receptors for neuropeptide FF. J. Biol. Chem. 275,39324–39331.
Bruhwyler, J., Chleide, E., Liegeois, J.F., Delarge, J., Mercier, M., 1990.Anxiolytic potential of sulpiride, clozapine and derivatives inthe open-field test. Pharmacol. Biochem. Behav. 36, 57–61.
Chalmers, D.T., Lovenberg, T.W., De Souza, E.B., 1995. Localizationof novel corticotropin-releasing factor receptor (CRF2) mRNAexpression to specific subcortical nuclei in rat brain: comparisonwith CRF1 receptor mRNA expression. J. Neurosci. 15, 6340–6350.
Covenas, R., de Leon, M., Cintra, A., Bjelke, B., Gustafsson, J.A.,Fuxe, K., 1993. Coexistence of c-Fos and glucocorticoid receptorimmunoreactivities in the CRF immunoreactive neurons of theparaventricular hypothalamic nucleus of the rat after acuteimmobilization stress. Neurosci. Lett. 149, 149–152.
Crawley, J.N., 1991. Cholecystokinin–dopamine interactions.Trends Pharmacol. Sci. 12, 232–236.
de Kloet, E.R., Joels, M., Holsboer, F., 2005. Stress and the brain:from adaptation to disease. Nat. Rev., Neurosci. 6, 463–475.
De Souza, E.B., 1995. Corticotropin-releasing factor receptors:physiology, pharmacology, biochemistry and role incentral nervous system and immune disorders.Psychoneuroendocrinology 20, 789–819.
Ellacott, K.L., Donald, E.L., Clarkson, P., Morten, J., Masters, D.,Brennand, J., Luckman, S.M., 2005. Characterization of anaturally-occurring polymorphism in the UHR-1 geneencoding the putative rat prolactin-releasing peptide receptor.Peptides 26, 675–681.
Engstrom, M., Brandt, A., Wurster, S., Savola, J.M., Panula, P., 2003.Prolactin releasing peptide has high affinity and efficacy atneuropeptide FF2 receptors. J. Pharmacol. Exp. Ther. 305, 825–832.
Fujii, R., Fukusumi, S., Hosoya, M., Kawamata, Y., Habata, Y.,Hinuma, S., Sekiguchi, M., Kitada, C., Kurokawa, T., Nishimura,O., Onda, H., Sumino, Y., Fujino, M., 1999. Tissue distribution ofprolactin-releasing peptide (PrRP) and its receptor. Regul. Pept.83, 1–10.
Griebel, G., Perrault, G., Sanger, D.J., 1998. Characterization of thebehavioral profile of the non-peptide CRF receptor antagonistCP-154,526 in anxiety models in rodents. Comparison withdiazepam and buspirone. Psychopharmacology (Berl) 138,55–66.
Gu, W., Geddes, B.J., Zhang, C., Foley, K.P., Stricker-Krongrad, A.,2004. The prolactin-releasing peptide receptor (GPR10)regulates body weight homeostasis in mice. J. Mol. Neurosci.22, 93–103.
Heinrichs, S.C., Lapsansky, J., Lovenberg, T.W., De Souza, E.B.,Chalmers, D.T., 1997. Corticotropin-releasing factor CRF1, butnot CRF2, receptors mediate anxiogenic-like behavior. Regul.Pept. 71, 15–21.

123B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
Hendrie, C.A., Neill, J.C., Shepherd, J.K., Dourish, C.T., 1993. Theeffects of CCKA and CCKB antagonists on activity in theblack/white exploration model of anxiety in mice. Physiol.Behav. 54, 689–693.
Hill, D.R., Campbell, N.J., Shaw, T.M., Woodruff, G.N., 1987.Autoradiographic localization and biochemicalcharacterization of peripheral type CCK receptors in rat CNSusing highly selective nonpeptide CCK antagonists. J. Neurosci.7, 2967–2976.
Hinuma, S., Habata, Y., Fujii, R., Kawamata, Y., Hosoya, M.,Fukusumi, S., Kitada, C., Masuo, Y., Asano, T., Matsumoto, H.,Sekiguchi, M., Kurokawa, T., Nishimura, O., Onda, H., Fujino,M., 1998. A prolactin-releasing peptide in the brain. Nature 393,272–276.
Honda, T., Wada, E., Battey, J.F., Wank, S.A., 1993. Differential geneexpression of CCKA and CCKB receptors in the rat brain. Mol.Cell. Neurosci. 4, 143–154.
Hsu, D.T., Chen, F.L., Takahashi, L.K., Kalin, N.H., 1998. Rapidstress-induced elevations in corticotropin-releasing hormonemRNA in rat central amygdala nucleus and hypothalamicparaventricular nucleus: an in situ hybridization analysis.Brain Res. 788, 305–310.
Imaki, T., Shibasaki, T., Hotta, M., Demura, H., 1992. Earlyinduction of c-fos precedes increased expression ofcorticotropin-releasing factor messenger ribonucleic acid inthe paraventricular nucleus after immobilization stress.Endocrinology 131, 240–246.
Imaki, T., Shibasaki, T., Hotta, M., Demura, H., 1993.Intracerebroventricular administration of corticotropin-releasingfactor induces c-fos mRNA expression in brain regions related tostress responses: comparison with pattern of c-fosmRNAinduction after stress. Brain Res. 616, 114–125.
Jhamandas, J.H., MacTavish, D., 2003. Central administration ofneuropeptide FF causes activation of oxytocin paraventricularhypothalamic neurones that project to the brainstem.J. Neuroendocrinol. 15, 24–32.
Kalin, N.H., Takahashi, L.K., Chen, F.L., 1994. Restraint stressincreases corticotropin-releasing hormone mRNA content inthe amygdala and paraventricular nucleus. Brain Res. 656,182–186.
Kamilaris, T.C., Johnson, E.O., Calogero, A.E., Kalogeras, K.T.,Bernardini, R., Chrousos, G.P., Gold, P.W., 1992.Cholecystokinin-octapeptide stimulateshypothalamic–pituitary–adrenal function in rats: role ofcorticotropin-releasing hormone. Endocrinology 130,1764–1774.
Kanemoto, N., Hishigaki, H., Miyakita, A., Oga, K., Okuno, S., Tsuji,A., Takagi, T., Takahashi, E., Nakamura, Y., Watanabe, T.K.,1998. Genetic dissection of “OLETF”, a rat model fornon-insulin-dependent diabetes mellitus. Mamm. Genome 9,419–425.
Kato, S., Ishita, S., Sugawara, K., Mawatari, K., 1993.Cystine/glutamate antiporter expression in retinal Muller glialcells: implications for DL-alpha-aminoadipate toxicity. Neuro-science 57, 473–482.
Katsuura, G., Ibii, N., Matsushita, A., 1992. Activation of CCK-Areceptors induces elevation of plasma corticosterone in rats.Peptides 13, 203–205.
Kawano, K., Hirashima, T., Mori, S., Saitoh, Y., Kurosumi, M.,Natori, T., 1992. Spontaneous long-term hyperglycemic ratwith diabetic complications. Otsuka Long–Evans TokushimaFatty (OLETF) strain. Diabetes 41, 1422–1428.
Keck, M.E., Holsboer, F., 2001. Hyperactivity of CRH neuronalcircuits as a target for therapeutic interventions in affectivedisorders. Peptides 22, 835–844.
Keck, M.E., Welt, T., Wigger, A., Renner, U., Engelmann, M.,Holsboer, F., Landgraf, R., 2001. The anxiolytic effect of theCRH(1) receptor antagonist R121919 depends on innateemotionality in rats. Eur. J. Neurosci. 13, 373–380.
Laurent, P., Becker, J.A., Valverde, O., Ledent, C., de Kerchoved'Exaerde, A., Schiffmann, S.N., Maldonado, R., Vassart, G.,Parmentier, M., 2005. The prolactin-releasing peptideantagonizes the opioid system through its receptor GPR10. Nat.Neurosci. 8, 1735–1741.
Lawrence, C.B., Celsi, F., Brennand, J., Luckman, S.M., 2000.Alternative role for prolactin-releasing peptide in theregulation of food intake. Nat. Neurosci. 3, 645–646.
Lawrence, C.B., Ellacott, K.L., Luckman, S.M., 2002. PRL-releasingpeptide reduces food intake andmaymediate satiety signaling.Endocrinology 143, 360–367.
Li, X.L., Aou, S., Hori, T., Oomura, Y., 2002. Spatial memory deficitand emotional abnormality in OLETF rats. Physiol. Behav. 75,15–23.
Liebsch, G., Landgraf, R., Gerstberger, R., Probst, J.C., Wotjak, C.T.,Engelmann, M., Holsboer, F., Montkowski, A., 1995. Chronicinfusion of a CRH1 receptor antisense oligodeoxynucleotideinto the central nucleus of the amygdala reducedanxiety-related behavior in socially defeated rats. Regul.Pept. 59, 229–239.
Liebsch, G., Landgraf, R., Engelmann, M., Lorscher, P., Holsboer, F.,1999. Differential behavioural effects of chronic infusion ofCRH 1 and CRH 2 receptor antisense oligonucleotides into therat brain. J. Psychiatr. Res. 33, 153–163.
Lin, S.H., Leslie, F.M., Civelli, O., 2002. Neurochemical properties ofthe prolactin releasing peptide (PrRP) receptor expressingneurons: evidence for a role of PrRP as a regulator of stress andnociception. Brain Res. 952, 15–30.
Lin, S.H., Civelli, O., 2004. Orphan G protein-coupled receptors:targets for new therapeutic interventions. Ann. Med. 36,204–214.
Marchese, A., Heiber, M., Nguyen, T., Heng, H.H., Saldivia, V.R.,Cheng, R., Murphy, P.M., Tsui, L.C., Shi, X., Gregor, P., et al.,1995. Cloning and chromosomal mapping of three novel genes,GPR9, GPR10, and GPR14, encoding receptors related tointerleukin 8, neuropeptide Y, and somatostatin receptors.Genomics 29, 335–344.
Maruyama, M., Matsumoto, H., Fujiwara, K., Kitada, C., Hinuma, S.,Onda, H., Fujino, M., Inoue, K., 1999a. Immunocytochemicallocalization of prolactin-releasing peptide in the rat brain.Endocrinology 140, 2326–2333.
Maruyama, M., Matsumoto, H., Fujiwara, K., Noguchi, J., Kitada, C.,Hinuma, S., Onda, H., Nishimura, O., Fujino, M., Higuchi, T.,Inoue, K., 1999b. Central administration of prolactin-releasingpeptide stimulates oxytocin release in rats. Neurosci. Lett. 276,193–196.
Maruyama, M., Matsumoto, H., Fujiwara, K., Noguchi, J., Kitada, C.,Fujino, M., Inoue, K., 2001. Prolactin-releasing peptide as anovel stress mediator in the central nervous system.Endocrinology 142, 2032–2038.
Matsumoto, H., Maruyama, M., Noguchi, J., Horikoshi, Y., Fujiwara,K., Kitada, C., Hinuma, S., Onda, H., Nishimura, O., Inoue, K.,Fujino, M., 2000. Stimulation of corticotropin-releasinghormone-mediated adrenocorticotropin secretion by centraladministration of prolactin-releasing peptide in rats. Neurosci.Lett. 285, 234–238.
Mera, T., Fujihara, H., Kawasaki, M., Hashimoto, H., Saito, T.,Shibata, M., Saito, J., Oka, T., Tsuji, S., Onaka, T., Ueta, Y., 2006.Prolactin-releasing peptide is a potent mediator of stressresponses in the brain through the hypothalamicparaventricular nucleus. Neuroscience 141, 1069–1086.
Okuno, S., Watanabe, T.K., Ono, T., Yamasaki, Y., Goto, Y., Miyao,H., Asai, T., Kanemoto, N., Oga, K., Mizoguchi-Miyakita, A.,Takagi, T., Takahashi, E., Nakamura, Y., Tanigami, A., 1999.Genetic determinants of plasma triglyceride levels in(OLETF×BN)×OLETF backcross rats. Genomics 62, 350–355.
Okuno, S.,Watanabe, T.K., Ono, T., Oga, K., Mizoguchi-Miyakita, A.,Yamasaki, Y., Goto, Y., Shinomiya, H., Momota, H., Miyao, H.,Hayashi, I., Asai, T., Suzuki, M., Harada, Y., Hishigaki, H.,

124 B R A I N R E S E A R C H 1 1 7 8 ( 2 0 0 7 ) 1 1 4 – 1 2 4
Wakitani, S., Takagi, T., Nakamura, Y., Tanigami, A., 2001.Effects of Dmo1 on obesity, dyslipidaemia and hyperglycaemiain the Otsuka Long Evans Tokushima Fatty strain. Genet. Res.77, 183–190.
Okuno, S., Kondo, M., Yamasaki, Y., Miyao, H., Ono, T., Iwanaga, T.,Omori, K., Okano, M., Suzuki, M., Momota, H., Hishigaki, H.,Hayashi, I., Goto, Y., Shinomiya, H., Harada, Y., Hirashima, T.,Kanemoto, N., Asai, T., Wakitani, S., Takagi, T., Nakamura, Y.,Tanigami, A., Watanabe, T.K., 2002. Substitution of Dmo1 withnormal alleles results in decreased manifestation of diabetesin OLETF rats. Diabetes Obes. Metab. 4, 309–318.
Paxinos, G., Watson, C., 1986. The Rat Brain in StereotaxicCoordinates. Academic Press, New York.
Pellow, S., File, S.E., 1986. Anxiolytic and anxiogenic drugeffects on exploratory activity in an elevated plus-maze: anovel test of anxiety in the rat. Pharmacol. Biochem. Behav.24, 525–529.
Pezzone, M.A., Lee,W.S., Hoffman, G.E., Rabin, B.S., 1992. Inductionof c-Fos immunoreactivity in the rat forebrain by conditionedand unconditioned aversive stimuli. Brain Res. 597, 41–50.
Preil, J., Muller, M.B., Gesing, A., Reul, J.M., Sillaber, I., van Gaalen,M.M., Landgrebe, J., Holsboer, F., Stenzel-Poore, M., Wurst, W.,2001. Regulation of the hypothalamic–pituitary–adrenocorticalsystem in mice deficient for CRH receptors 1 and 2.Endocrinology 142, 4946–4955.
Rodgers, R.J., Cao, B.J., Dalvi, A., Holmes, A., 1997. Animal modelsof anxiety: an ethological perspective. Braz. J. Med. Biol. Res. 30,289–304.
Roland, B.L., Sutton, S.W., Wilson, S.J., Luo, L., Pyati, J., Huvar, R.,Erlander, M.G., Lovenberg, T.W., 1999. Anatomical distributionof prolactin-releasing peptide and its receptor suggestsadditional functions in the central nervous system andperiphery. Endocrinology 140, 5736–5745.
Samson, W.K., Keown, C., Samson, C.K., Samson, H.W., Lane, B.,Baker, J.R., Taylor, M.M., 2003. Prolactin-releasing peptide andits homolog RFRP-1 act in hypothalamus but not in anteriorpituitary gland to stimulate stress hormone secretion.Endocrine 20, 59–66.
Saunders, J., Williams, J., 2003. Antagonists of the corticotropinreleasing factor receptor. Prog. Med. Chem. 41, 195–247.
Seal, L.J., Small, C.J., Dhillo, W.S., Kennedy, A.R., Ghatei, M.A.,Bloom, S.R., 2002. Prolactin-releasing peptide releasescorticotropin-releasing hormone and increases plasmaadrenocorticotropin via the paraventricular nucleus of thehypothalamus. Neuroendocrinology 76, 70–78.
Shibasaki, T., Imaki, T., Hotta, M., Ling, N., Demura, H., 1993.Psychological stress increases arousal through braincorticotropin-releasing hormone without significant increasein adrenocorticotropin and catecholamine secretion. Brain Res.618, 71–75.
Skutella, T., Probst, J.C., Renner, U., Holsboer, F., Behl, C., 1998.Corticotropin-releasing hormone receptor (type I) antisensetargeting reduces anxiety. Neuroscience 85, 795–805.
Smith, G.W., Aubry, J.M., Dellu, F., Contarino, A., Bilezikjian, L.M.,Gold, L.H., Chen, R., Marchuk, Y., Hauser, C., Bentley, C.A.,Sawchenko, P.E., Koob, G.F., Vale, W., Lee, K.F., 1998.Corticotropin releasing factor receptor 1-deficient mice displaydecreased anxiety, impaired stress response, and aberrantneuroendocrine development. Neuron 20, 1093–1102.
Steckler, T., Holsboer, F., 1999. Corticotropin-releasing hormonereceptor subtypes and emotion. Biol. Psychiatry 46,1480–1508.
Takahashi, L.K., Kalin, N.H., Vanden Burgt, J.A., Sherman, J.E.,1989. Corticotropin-releasing factor modulates defensive-withdrawal and exploratory behavior in rats. Behav. Neurosci.103, 648–654.
Takiguchi, S., Takata, Y., Funakoshi, A., Miyasaka, K., Kataoka, K.,Fujimura, Y., Goto, T., Kono, A., 1997. Disrupted cholecystokinintype-A receptor (CCKAR) gene in OLETF rats. Gene 197, 169–175.
Takiguchi, S., Takata, Y., Takahashi, N., Kataoka, K., Hirashima, T.,Kawano, K., Miyasaka, K., Funakoshi, A., Kono, A., 1998. Adisrupted cholecystokinin A receptor gene induces diabetes inobese rats synergistically with ODB1 gene. Am. J. Physiol. 274,E265–E270.
Timpl, P., Spanagel, R., Sillaber, I., Kresse, A., Reul, J.M., Stalla, G.K.,Blanquet, V., Steckler, T., Holsboer, F., Wurst, W., 1998.Impaired stress response and reduced anxiety in mice lackinga functional corticotropin-releasing hormone receptor 1. Nat.Genet. 19, 162–166.
Tobe, I., Ishida, Y., Tanaka, M., Endoh, H., Fujioka, T., Nakamura, S.,2005. Effects of repeated maternal stress on FOS expression inthe hypothalamic paraventricular nucleus of fetal rats.Neuroscience 134, 387–395.
Van Pett, K., Viau, V., Bittencourt, J.C., Chan, R.K., Li, H.Y., Arias, C.,Prins, G.S., Perrin, M., Vale, W., Sawchenko, P.E., 2000.Distribution of mRNAs encoding CRF receptors in brain andpituitary of rat and mouse. J. Comp. Neurol. 428, 191–212.
Wang, L., Martinez, V., Barrachina, M.D., Tache, Y., 1998. Fosexpression in the brain induced by peripheral injection of CCKor leptin plus CCK in fasted lean mice. Brain Res. 791, 157–166.
Watanabe, T.K., Okuno, S., Oga, K., Mizoguchi-Miyakita, A., Tsuji,A., Yamasaki, Y., Hishigaki, H., Kanemoto, N., Takagi, T.,Takahashi, E., Irie, Y., Nakamura, Y., Tanigami, A., 1999.Genetic dissection of “OLETF”, a rat model fornon-insulin-dependent diabetes mellitus: quantitative traitlocus analysis of (OLETF×BN)×OLETF. Genomics 58, 233–239.
Watanabe, T.K., Ono, T., Okuno, S., Mizoguchi-Miyakita, A.,Yamasaki, Y., Kanemoto, N., Hishigaki, H., Oga, K., Takahashi,E., Irie, Y., Bihoreau, M.T., James, M.R., Lathrop, G.M., Takagi, T.,Nakamura, Y., Tanigami, A., 2000. Characterization of newlydeveloped SSLP markers for the rat. Mamm. Genome 11,300–305.
Watanabe, T.K., Okuno, S., Ono, T., Yamasaki, Y., Oga, K.,Mizoguchi-Miyakita, A., Miyao, H., Suzuki, M., Momota, H.,Goto, Y., Shinomiya, H., Hishigaki, H., Hayashi, I., Asai, T.,Wakitani, S., Takagi, T., Nakamura, Y., Tanigami, A., 2001.Single-allele correction of the Dmo1 locus in congenic animalssubstantially attenuates obesity, dyslipidaemia and diabetesphenotypes of the OLETF rat. Clin. Exp. Pharmacol. Physiol. 28,28–42.
Watanabe, T.K., Suzuki, M., Yamasaki, Y., Okuno, S., Hishigaki, H.,Ono, T., Oga, K., Mizoguchi-Miyakita, A., Tsuji, A., Kanemoto, N.,Wakitani, S., Takagi, T., Nakamura, Y., Tanigami, A., 2005.Mutated G-protein-coupled receptor GPR10 is responsible for thehyperphagia/dyslipidaemia/obesity locus of Dmo1 in the OLETFrat. Clin. Exp. Pharmacol. Physiol. 32, 355–366.
Yamamoto, Y., Akiyoshi, J., Kiyota, A., Katsuragi, S., Tsutsumi, T.,Isogawa, K., Nagayama, H., 2000. Increased anxiety behavior inOLETF rats without cholecystokinin-A receptor. Brain Res. Bull.53, 789–792.
Yamasaki, Y., Watanabe, T.K., Okuno, S., Ono, T., Oga, K.,Mizoguchi-Miyakita, A., Goto, Y., Shinomiya, H., Momota, H.,Miyao, H., Hayashi, I., Asai, T., Suzuki, M., Harada, Y., Hishigaki,H., Wakitani, S., Takagi, T., Nakamura, Y., Tanigami, A., 2000.Quantitative trait loci for lipid metabolism in the study ofOLETF×(OLETF×Fischer 344) backcross rats. Clin. Exp.Pharmacol. Physiol. 27, 881–886.
Zhang, S.Q., Kimura, M., Inoue, S., 2000. Effects of prolactin-releasingpeptide (PrRP) on sleep regulation in rats. Psychiatry Clin.Neurosci. 54, 262–264.
Zorrilla, E.P., Koob, G.F., 2004. The therapeutic potential of CRF1antagonists for anxiety. Expert Opin. Investig. Drugs 13, 799–828.




















