
governor's sfy2023 budget testimony - Disabilities, Aging and ...
Upload
independentCategory
view
1download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/authorsrights

Author's personal copy
Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208
Contents lists available at ScienceDirect
Neuroscience and Biobehavioral Reviews
jou rn al h om epage: www.elsev ier .com/ locate /neubiorev
Neuromodulation and developmental contextual influences on neuraland cognitive plasticity across the lifespan
Shu-Chen Lia,b,∗
a Department of Psychology, Chair of Lifespan Developmental Neuroscience, TU Dresden, Dresden, Germanyb Center for Lifespan Psychology, Max Planck Institute for Human Development, Berlin, Germany
a r t i c l e i n f o
Article history:Received 5 December 2012Received in revised form 11 July 2013Accepted 12 July 2013
Keywords:Developmental plasticityConstructivistNeuromodulationRewardMotivationGoal directed behaviorStress
a b s t r a c t
Behavioral, cognitive, and motivational development entails co-constructive interactions between theenvironmental and social influences from the developmental context, on the one hand, and the individ-ual’s neurobiological inheritance, on the other hand. Key brain networks underlying cognition, emotion,and motivation are innervated by major transmitter systems (e.g., the catecholamines and acetylcholine).Thus, the maturation and senescence of neurotransmitter systems have direct implications for lifespandevelopment. In addition to reviewing evidence on life age differences in dopaminergic modulationand cognitive development, this brief review selectively highlights recent findings on how importantinfluences from the developmental context, such as reward-mediated motivational processes, transgen-erational stress transmission, psychosocial stress, and cognitive interventions, may, in part, exert theireffects on brain and behavioral development through their effects on neuromodulatory mechanisms.
© 2013 Elsevier Ltd. All rights reserved.
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22011.1. The development of embodied and situated adaptive neurocognitive representations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2201
2. The maturation and senescence of neuromodulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22023. Lifespan changes in neuromodulation may affect genotype-phenotype relations . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22024. Contextual influences on development operated through neuromodulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2203
4.1. Cognitive training and dopamine modulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22034.2. Reward, motivational incentives, and dopamine modulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22034.3. Stress and dopamine modulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2204
5. Summary and future directions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2206References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2207
1. Introduction
Development entails cumulative reciprocal interactionsbetween the individual’s neurobiological faculties (genetic pre-dispositions and brain mechanisms) and the social as wellas environmental influences from the developmental context(Baltes et al., 2006; Bronfenbrenner, 1979; Gottlieb, 1998; Li,2003; Vygotsky, 1978). Moreover, it also needs to be under-scored that individuals are not mere passive recipients of their
∗ Correspondence to: Chair of Lifespan Developmental Neuroscience, Departmentof Psychology, TU Dresden, Zellescher Weg 17, D-01062 Dresden, Germany.Tel.: +49 0351 46334162; fax: +49 0351 46342194.
E-mail address: [email protected]
neurobiological, social and cultural inheritances, the person’s voli-tional choices and actions (e.g., Heckhausen et al., 2010; Gollwitzeret al., 2011; Goschke, 2004) play critical roles in regulating howenvironmental and socio-contextual influences may affect thecourse of development across the lifespan (e.g., Baltes and Baltes,1990; Freund, 2008).
1.1. The development of embodied and situated adaptiveneurocognitive representations
To better capture the interactive dynamics between (i) theindividual’s self regulatory behavior and (ii) the resources andconstraints on individual ontogeny that arise from the develop-ment context as well as (iii) mechanisms of brain maturation andsenescence, a perhaps fruitful conceptualization is to view lifespan
0149-7634/$ – see front matter © 2013 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.neubiorev.2013.07.019

Author's personal copy
2202 S.-C. Li / Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208
development as the development of adaptive neurocognitive rep-resentations that are “embodied” in motor, sensory, and perceptualprocesses and “situated” in social and environmental contexts (cf.Clark, 2001; Robbins and Aydede, 2008). The development of suchadaptive neurocognitive representations entails multiple levels ofplasticity, ranging from the individual’s genetic, neuronal, brainand behavioral plasticity to large-scale adaptions in social and cul-tural practices that are relevant for individual development at themacro level (Li, 2003). Being self-organizing systems (Ford, 1987),brains optimize the levels of matches and mismatches betweenenvironmental states and action outcomes (Friston, 2010).
One of the most striking features of brains is that neuronscontain and release a large number of neurotransmitters, whichplay important roles in regulating signal transmissions betweenneurons (see Vizi and Lajtha, 2008, for an overview). Several trans-mitter systems, such as the catecholamines (dopamine, serotonin,and norepinephrine), broadly innervate various neural circuitriesthroughout the brain to modulate key aspects of cognition andbehavior. Taking the dopamine system as an example, other thaneffects of the prefrontal dopamine system, given that multiple cir-cuits connect striatal regions with regions in the frontal cortex(Pennartz et al., 2009), recent human research has begun to investi-gate the role of striatal dopamine in working memory and attention(e.g., Cools et al., 2004; Landau et al., 2005; Li et al., 2013a; McNabet al., 2009) as well as in reward-mediated motivational influenceson behavior control (see Kalivas and Volkow, 2005; Noudoost andMoore, 2011; Shohamy and Adcock, 2010; Volkow et al., 2011 forrecent reviews).
Indeed, neuromodulatory systems have been considered as keyneural substrates for adaptive value-dependent selection in thebrain (Friston et al., 1994). Depending on situational demands andthe integrity of brain functions, neurotransmitters modulate task-relevant brain circuitries, so that individuals can adapt their behav-ior, action, and goals. Throughout development, the embodied andsituated neurocognitive representations are modulated by variousneurotransmitters that implicate executive control, memory andmotivational processes. Therefore, the maturation and senescenceof neurotransmitter systems have direct implications for behav-ioral and cognitive development across the lifespan. This reviewfocuses on the maturation and senescence of neurotransmitter sys-tems, with a focus on highlighting research questions that may shedlight on how lifespan age differences in dopaminergic neuromod-ulation may contribute to co-constructive interactions betweenbrain, behavior, and influences of the developmental context.
2. The maturation and senescence of neuromodulation
Currently, there is the consensus that the integrities of thedopamine, serotonin, and acetylcholine systems decline duringthe course of usual aging. For instance, cross-sectional estimatesshowed that in various extrastriatal and striatal regions the pre-and postsynaptic markers of the dopamine system decline about10% per decade starting around the begin of the second decade oflife (e.g., Kaasinen et al., 2000; Inoue et al., 2001; see also Bäckmanet al., 2006 for review). Similarly, cross-sectional estimates of aging-related declines in the availability of serotonin receptors invarious brain regions also range from 3% to 10% per decade (e.g.,Pirker et al., 2000; Yamamoto et al., 2002; see also Eppinger et al.,2011, for review). As for the cholinergic system, which is known toimplicate neurodegenerative processes associated with dementa(see Court et al., 2001, for review), results from a recent receptorimaging study also showed that, on average, there is about 5% perdecade decline of the nicotinic acetylcholine receptor in eight brainregions, including the frontal cortex and striatum (Mitsis et al.,2009). Empirical evidence and computational studies have related
aging-related declines in dopamine modulation to age-relateddeficits in processing speed, processing fluctuations, episodic mem-ory, working memory, and cognitive control (e.g., Bäckman et al.,2011; Erixon-Lindroth et al., 2005; see Bäckman et al., 2006 for anempirical review; see Li et al., 2001 for theoretical integration).
As for the maturation of neuromodulatory systems during childand adolescent development, the evidence is much scarcer dueto practical limitations of applying invasive methods, such as PETreceptor imaging, in these age groups. Therefore, comparisonsof dopamine functions across the lifespan have, so far, based onresults from animal studies (e.g., Tarazi and Baldessarini, 2000;see Herlenius and Lagercrantz, 2004 for review) or from humanpost-mortem studies (e.g., Haycock et al., 2003; Tunbridge et al.,2007). For instance, postnatally dopamine level in the striatumincreases 2–3 folds through adolescence and then decreases dur-ing aging (Haycock et al., 2003). Consider also the evidence fromanother study (Tunbridge et al., 2007), which shows that the activ-ity of an enzyme that regulates extracellular dopamine levels in theprefrontal cortex (i.e., the catechol-o-methyltransferase enzyme)increases about 2-folds from neonate to adulthood, and declineslightly afterwards. Evidence from animal studies also suggests thatthe efficacies of both the subcortical and cortical dopamine systems(e.g., the density of different receptor types) increase continuouslyand steadily during the postnatal period. Furthermore, the subcor-tical dopamine system reaches its peak already in peri-adolescence,whereas the development of the cortical system is slower andreaches its peak level only in early adulthood. Specifically, Taraziand Baldessarini (2000) found that the density of dopamine D1,D2, and D4 receptors in the caudate and nucleus accumbens of ratsincreased to a peak at about postnatal day 28 (approximately equiv-alent to late childhood and peri-adolescence in humans), but thendeclined significantly (from postnatal 35 to 60 days) to adult level.Interestingly, the density of these three receptor types in the pre-frontal cortex, however, rose steadily throughout the childhood andadolescence and only reached a maximum of the adult level by 60postnatal day 60.
A recent positron emission tomography (PET) study with sam-ples of adolescents (n = 12) and younger adults (n = 18) providedthe rare human in vivo data and showed that dopamine D1 bindingpotential in the prefrontal cortex continued to show age-dependentdifferences until adulthood, whereas no such age-dependent differ-ences was observed in the dorsal or ventral striatum (Jucaite et al.,2010). The more protracted maturation of the cortical dopaminesystem may constraint the development of attention and otherfrontal executive functions during childhood and adolescence (e.g.,Diamond, 1996; Diamond et al., 2004; Liotti et al., 2005). Moreover,the discrepancy in the maturational trajectories of the subcorticaland cortical dopamine systems in adolescence (see Andersen, 2003;Crews et al., 2007; Luciana et al., 2012 for reviews) may make thislife period particularly malleable by positive (e.g., social rewards)or negative contextual influences (e.g., extreme stress or addiction).
3. Lifespan changes in neuromodulation may affectgenotype-phenotype relations
Progresses in molecular genetics have opened new avenuesfor investigating neuromodulation of behavioral and cognitivedevelopment (e.g., Diamond et al., 2004; Lindenberger et al., 2008;Posner et al., 2007; Spangler et al., 2009). Of particular interestin this context, changes in brain resources at the anatomicalor neurochemical levels during maturation or senescence maymodulate genotype-phenotype relations in different life periods,as brain mechanisms are the “intermediate phenotypes” (Meyer-Lindenberg and Weinberger, 2006) between genetic expressionsin the central nervous system and behavioral phenotypes. Genes

Author's personal copy
S.-C. Li / Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208 2203
related to the neurotransmitter dopamine (DA) is a good exam-ple. Evidence from clinical (Mattay et al., 2003) and animal(Vijayraghavan et al., 2007) studies as well as neurocomputationalsimulations (Li and Sikström, 2002) suggests that the relationbetween DA signaling and cognitive performance follows aninverted-U function (see Cools and D’Esposito, 2011, for review).The nonlinear function relating DA modulation to cognitive per-formance predicts that genotype effects on cognition may not bestatic across the lifespan. Instead, the patterns and strengths ofgenotype effects on cognitive and brain phenotypes may inter-act with age. For instance, the resource-modulation hypothesissuggests that genotype effects would be more apparent when DAsignaling recedes from an optimal level, such as in developing andaging individuals or in conductions when the nature dopaminelevel is perturbed by excessive stress or stimulants that affectneuromodulation (Li et al., 2010; Lindenberger et al., 2008).
Findings from recent studies on aging lend preliminary supportto this resource-modulation hypothesis. Specifically, Older adults’spatial working memory and executive functioning were associ-ated with individual differences in genetic predispositions of theCOMT gene, which affects dopamine signaling in the prefrontalcortex. Specifically, older Met homozygotes of the COMT gene,presumably operating with higher prefrontal dopamine levels, per-formed better than COMT Val carries on spatial working memoryand the Wisconsin Card Sorting task (Nagel et al., 2008; Störmeret al., 2011). In younger adults no such relation was observed,however. In addition, two studies involving child developmen-tal samples also showed that the effects of COMT genotype onprefrontal functions interacted with age. Wahlstrom et al. (2007)observed an advantage of COMT met homozygotes in prefrontalcognition over val homozygotes during childhood but not in adoles-cence. In line with the inverted-U dopamine modulation function,this finding suggests that during adolescence when prefrontaldopamine level is already relatively high, the lower degradationrate of extracellular dopamine associated with carrying two metalleles of the COMT gene may render the level of extracellulardopamine too high and hampers performance. In another study,Dumontheil et al. (2011) observed a positive correlation betweenage (6–20 yeas) and working-memory related activation in theprefrontal cortex only in COMT val homozygotes, indicating thatCOMT genotype effects on prefrontal cognition is not static duringdevelopment.
Other than working memory and executive control functions,genotype effects of dopamine transporter (DAT) and receptor(DRD2) genes on serial order memory (Lewandowsky and Murdock,1989) – an aspect of episodic memory that places high encodingand retrieval operational demands on the frontal-hippocampal net-work (e.g., DeVito and Eichenbaum, 2011) – were also found tobe larger in older than in younger adults. In older adults, carry-ing of genotypes associated with higher levels of striatal synapticdopamine (DAT 9/9) and higher density of extrastriatal D2 recep-tors (C957T CC) was associated with better backward serial recall.Critically, the gene–gene interaction effect was reliably strongerin older than in younger adults (Li et al., 2013b; Papenberg et al.,2013). Beyond memory functions, in a lifespan sample Hämmereret al. (2013) recently observed larger effects of variations in anothergene (PPP1R1B), relevant for dopamine D1 and D2 receptor func-tions, on feedback-related brain evoked potentials in children andin older adults as compared to the effect in younger adults. Theseresults from candidate gene association studies need to be repli-cated in independent samples in order for the observed relations tobe considered as established. Nonetheless, in the search for associa-tions between behavioral phenotypes and genotypes, the potentialinfluences of lifespan changes in the intermediate brain phenotypes(e.g., neuromodulation) on such relations in different life periodsneed to be considered.
4. Contextual influences on development operated throughneuromodulation
In order to better understand co-constructive interactionsbetween the individual’s self-regulatory actions, brain functions,and the impacts of the developmental context, mechanisms ofaction monitoring and other influences from the developmen-tal context need to be brought to the foreground, in addition toconsidering the unidirectional effects from neuromodulation tobehavior. Many approaches can be taken to study the effects ofdevelopmental contexts on developmental plasticity. For instance,physical fitness training and cognitive interventions have beenshown to exert reciprocal influences on brain structure and func-tion (see Hertzog et al., 2009; Lövdén et al., 2010, for reviews).However, to specifically investigate the co-constructive dynamicsbetween neuromodulation and contextual influences on develop-ment, contextual factors that operated on or through the varioustransmitter systems would be of particular interest (see schematicillustration in Fig. 1), as they serve as a case in point of contex-tual influences on the neural substrates of brain’s value-dependentadaptation. Below three such contextual influences affecting brainand cognition and outstanding questions lifespan research arehighlighted.
4.1. Cognitive training and dopamine modulation
In terms of cognitive interventions, training paradigms thattargeting functional brain circuitries (e.g., the working memorynetwork) that are innervated by the major transmitter pathwaysseem to be a promising avenue. In younger adults, working memorytraining across five weeks was found to be associated with changesin cortical dopamine D1 receptor binding potential in the prefrontaland parietal cortices (McNab et al., 2009) and in striatal dopamineD2 receptor binding potential (Bäckman et al., 2011). Togetherthese findings demonstrate a reciprocal co-constructive interplaybetween cognitive intervention and dopamine modulation in vivo.It remain to be seen how effects of cognitive intervention on neu-romodulation may differ between adults and other age groupsin whom the efficacy of the dopamine systems are either stillundergoing maturation or have started to decline. For instance,two recent behavioral genetic studies reported an associationbetween dopamine transporter (DAT) genotype on working mem-ory plasticity in children (Söderqvist et al., 2011) and in youngeradults (Brehmer et al., 2009). To understand the epigenetic effects,future studies need to investigate interactions between individualdifferences in dopamine relevant genotypes and cognitive inter-vention effects on training-related dopamine release in differentage groups.
4.2. Reward, motivational incentives, and dopamine modulation
Adaptive behavioral control is a crucial mean with which theindividuals can actively shape their own development. People areoften confronted with complex, uncertain situations that nonethe-less require decisive actions that would facilitate the pursuits ofshort-term or long-term goals. On the one hand, achieving goalsare rewarding, and, on the other hand, performance incentives canfacilitate goal attainment (Jimura et al., 2010; Niv et al., 2006).Adaptive behavior or action as such entail online interactionsbetween processes that monitor the choice-outcome relations aswell as evaluate these relations with respect to goal relevanceand modified future actions based on the valuations (Rangel et al.,2008). These dynamics require close interplays between atten-tion and motivation, which are subserved by the frontal-striatalnetworks (Berridge and Robinson, 2003).
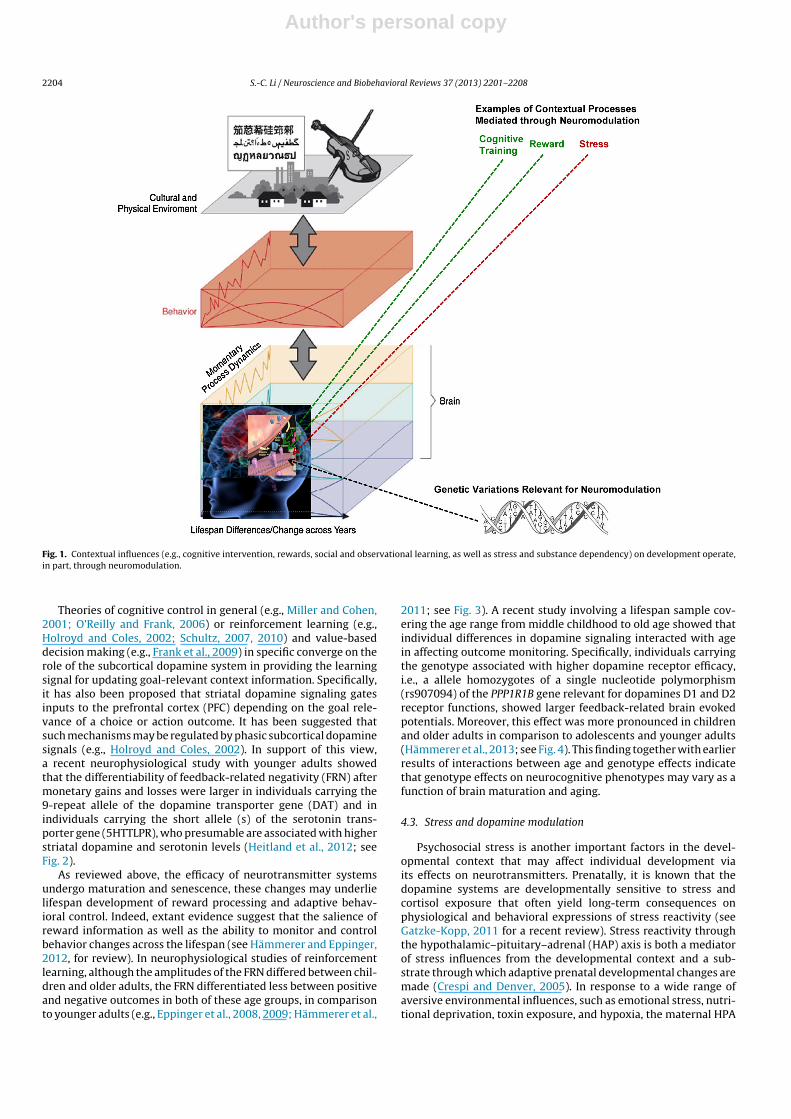
Author's personal copy
2204 S.-C. Li / Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208
Fig. 1. Contextual influences (e.g., cognitive intervention, rewards, social and observational learning, as well as stress and substance dependency) on development operate,in part, through neuromodulation.
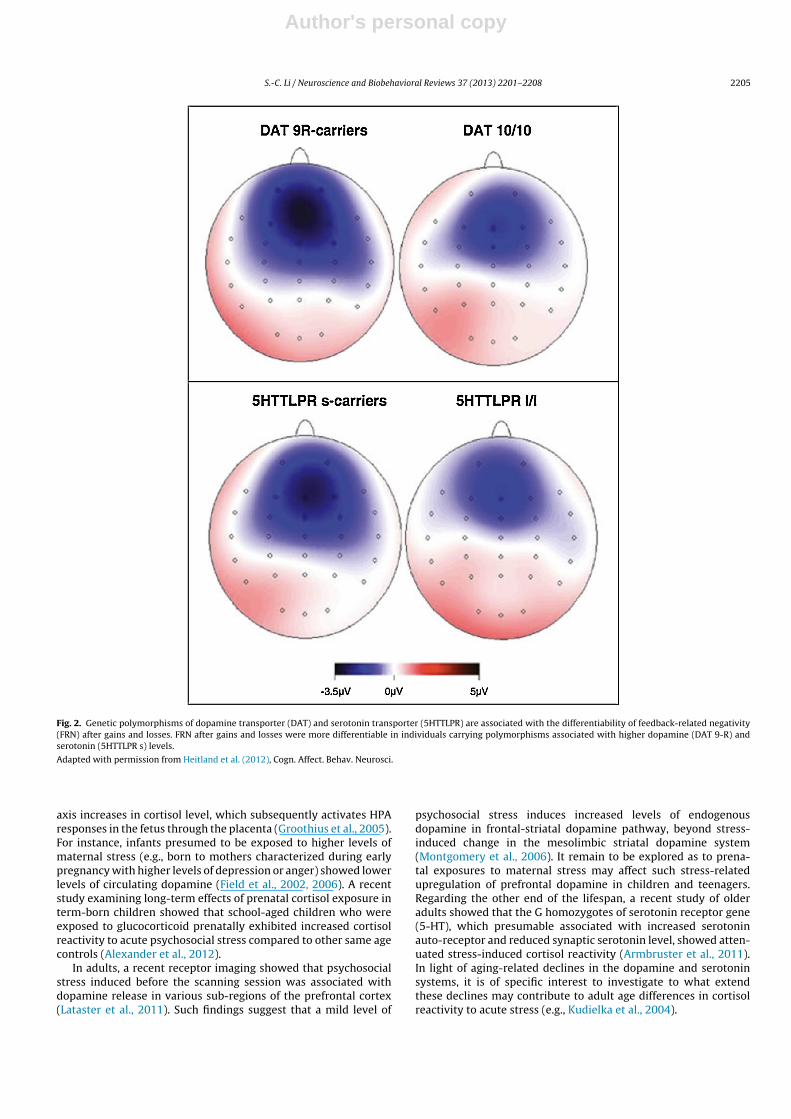
Theories of cognitive control in general (e.g., Miller and Cohen,2001; O’Reilly and Frank, 2006) or reinforcement learning (e.g.,Holroyd and Coles, 2002; Schultz, 2007, 2010) and value-baseddecision making (e.g., Frank et al., 2009) in specific converge on therole of the subcortical dopamine system in providing the learningsignal for updating goal-relevant context information. Specifically,it has also been proposed that striatal dopamine signaling gatesinputs to the prefrontal cortex (PFC) depending on the goal rele-vance of a choice or action outcome. It has been suggested thatsuch mechanisms may be regulated by phasic subcortical dopaminesignals (e.g., Holroyd and Coles, 2002). In support of this view,a recent neurophysiological study with younger adults showedthat the differentiability of feedback-related negativity (FRN) aftermonetary gains and losses were larger in individuals carrying the9-repeat allele of the dopamine transporter gene (DAT) and inindividuals carrying the short allele (s) of the serotonin trans-porter gene (5HTTLPR), who presumable are associated with higherstriatal dopamine and serotonin levels (Heitland et al., 2012; seeFig. 2).
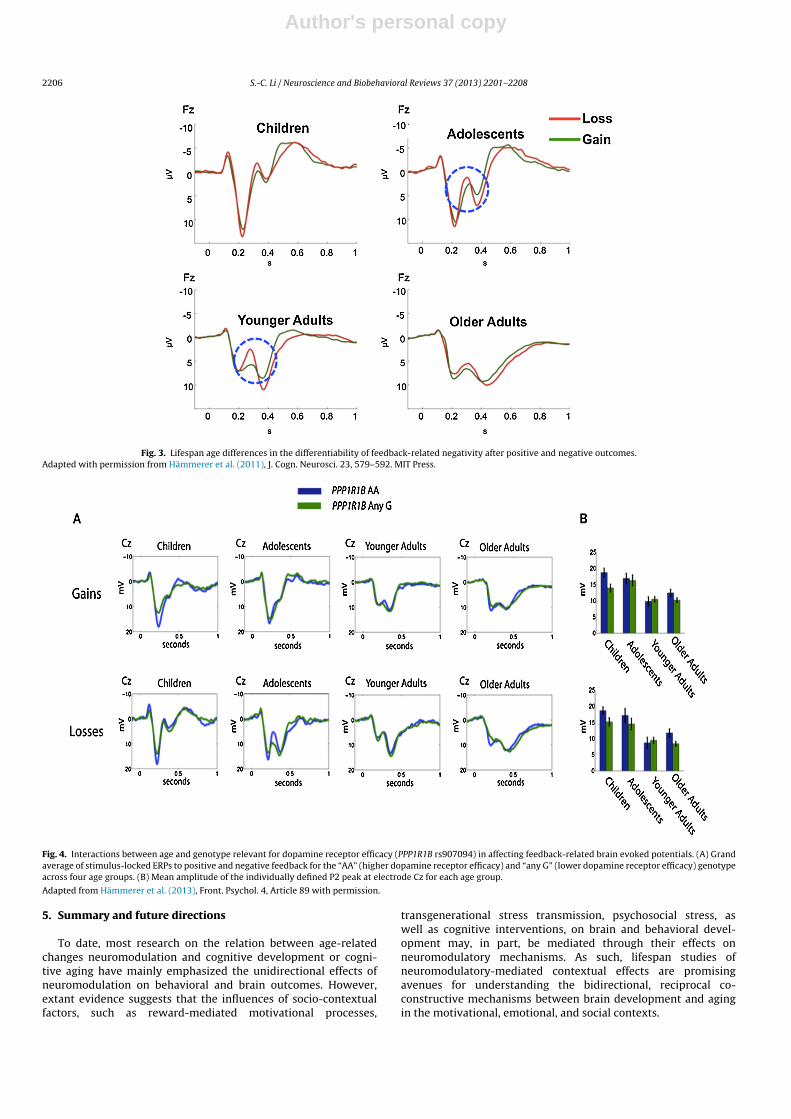
As reviewed above, the efficacy of neurotransmitter systemsundergo maturation and senescence, these changes may underlielifespan development of reward processing and adaptive behav-ioral control. Indeed, extant evidence suggest that the salience ofreward information as well as the ability to monitor and controlbehavior changes across the lifespan (see Hämmerer and Eppinger,2012, for review). In neurophysiological studies of reinforcementlearning, although the amplitudes of the FRN differed between chil-dren and older adults, the FRN differentiated less between positiveand negative outcomes in both of these age groups, in comparisonto younger adults (e.g., Eppinger et al., 2008, 2009; Hämmerer et al.,
2011; see Fig. 3). A recent study involving a lifespan sample cov-ering the age range from middle childhood to old age showed thatindividual differences in dopamine signaling interacted with agein affecting outcome monitoring. Specifically, individuals carryingthe genotype associated with higher dopamine receptor efficacy,i.e., a allele homozygotes of a single nucleotide polymorphism(rs907094) of the PPP1R1B gene relevant for dopamines D1 and D2receptor functions, showed larger feedback-related brain evokedpotentials. Moreover, this effect was more pronounced in childrenand older adults in comparison to adolescents and younger adults(Hämmerer et al., 2013; see Fig. 4). This finding together with earlierresults of interactions between age and genotype effects indicatethat genotype effects on neurocognitive phenotypes may vary as afunction of brain maturation and aging.
4.3. Stress and dopamine modulation
Psychosocial stress is another important factors in the devel-opmental context that may affect individual development viaits effects on neurotransmitters. Prenatally, it is known that thedopamine systems are developmentally sensitive to stress andcortisol exposure that often yield long-term consequences onphysiological and behavioral expressions of stress reactivity (seeGatzke-Kopp, 2011 for a recent review). Stress reactivity throughthe hypothalamic–pituitary–adrenal (HAP) axis is both a mediatorof stress influences from the developmental context and a sub-strate through which adaptive prenatal developmental changes aremade (Crespi and Denver, 2005). In response to a wide range ofaversive environmental influences, such as emotional stress, nutri-tional deprivation, toxin exposure, and hypoxia, the maternal HPA
Author's personal copy
S.-C. Li / Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208 2205
Fig. 2. Genetic polymorphisms of dopamine transporter (DAT) and serotonin transporter (5HTTLPR) are associated with the differentiability of feedback-related negativity(FRN) after gains and losses. FRN after gains and losses were more differentiable in individuals carrying polymorphisms associated with higher dopamine (DAT 9-R) andserotonin (5HTTLPR s) levels.
Adapted with permission from Heitland et al. (2012), Cogn. Affect. Behav. Neurosci.
axis increases in cortisol level, which subsequently activates HPAresponses in the fetus through the placenta (Groothius et al., 2005).For instance, infants presumed to be exposed to higher levels ofmaternal stress (e.g., born to mothers characterized during earlypregnancy with higher levels of depression or anger) showed lowerlevels of circulating dopamine (Field et al., 2002, 2006). A recentstudy examining long-term effects of prenatal cortisol exposure interm-born children showed that school-aged children who wereexposed to glucocorticoid prenatally exhibited increased cortisolreactivity to acute psychosocial stress compared to other same agecontrols (Alexander et al., 2012).
In adults, a recent receptor imaging showed that psychosocialstress induced before the scanning session was associated withdopamine release in various sub-regions of the prefrontal cortex(Lataster et al., 2011). Such findings suggest that a mild level of
psychosocial stress induces increased levels of endogenousdopamine in frontal-striatal dopamine pathway, beyond stress-induced change in the mesolimbic striatal dopamine system(Montgomery et al., 2006). It remain to be explored as to prena-tal exposures to maternal stress may affect such stress-relatedupregulation of prefrontal dopamine in children and teenagers.Regarding the other end of the lifespan, a recent study of olderadults showed that the G homozygotes of serotonin receptor gene(5-HT), which presumable associated with increased serotoninauto-receptor and reduced synaptic serotonin level, showed atten-uated stress-induced cortisol reactivity (Armbruster et al., 2011).In light of aging-related declines in the dopamine and serotoninsystems, it is of specific interest to investigate to what extendthese declines may contribute to adult age differences in cortisolreactivity to acute stress (e.g., Kudielka et al., 2004).
Author's personal copy
2206 S.-C. Li / Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208
Fig. 3. Lifespan age differences in the differentiability of feedback-related negativity after positive and negative outcomes.Adapted with permission from Hämmerer et al. (2011), J. Cogn. Neurosci. 23, 579–592. MIT Press.
Fig. 4. Interactions between age and genotype relevant for dopamine receptor efficacy (PPP1R1B rs907094) in affecting feedback-related brain evoked potentials. (A) Grandaverage of stimulus-locked ERPs to positive and negative feedback for the “AA” (higher dopamine receptor efficacy) and “any G” (lower dopamine receptor efficacy) genotypeacross four age groups. (B) Mean amplitude of the individually defined P2 peak at electrode Cz for each age group.
Adapted from Hämmerer et al. (2013), Front. Psychol. 4, Article 89 with permission.
5. Summary and future directions
To date, most research on the relation between age-relatedchanges neuromodulation and cognitive development or cogni-tive aging have mainly emphasized the unidirectional effects ofneuromodulation on behavioral and brain outcomes. However,extant evidence suggests that the influences of socio-contextualfactors, such as reward-mediated motivational processes,
transgenerational stress transmission, psychosocial stress, aswell as cognitive interventions, on brain and behavioral devel-opment may, in part, be mediated through their effects onneuromodulatory mechanisms. As such, lifespan studies ofneuromodulatory-mediated contextual effects are promisingavenues for understanding the bidirectional, reciprocal co-constructive mechanisms between brain development and agingin the motivational, emotional, and social contexts.

Author's personal copy
S.-C. Li / Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208 2207
References
Alexander, N., Rosenlöcher, F., Stalder, T., Linke, J., Distler, W., Morgner, J.,Kirschbaum, C., 2012. Impact of antenatal synthetic glucocorticoid exposure onendocrine stress reactivity in term born children. J. Clin. Endocrinol. Metab. 97,3538–3544.
Andersen, S.L., 2003. Trajectories of brain development: point of vulner-ability or window of opportunity. Neurosci. Biobehav. Rev. 27, 3–18,http://dx.doi.org/10.1016/S0149-7634(03)00005-8.
Armbruster, D., Mueller, A., Strobel, A., Lesch, K.-P., Brocke, B., Kirschbaum, C., 2011.Predicting cortisol stress responses in older individuals: influence of serotoninreceptor 1 A gene (HTR1A) and stressful life events. Horm. Behav. 60, 105–111,http://dx.doi.org/10.1016/j.yhbeh.2011.03.010.
Bäckman, L., Nyberg, L., Lindenberger, U., Li, S.-C., Farde, L., 2006. Thecorrelative triad among aging, dopamine, and cognition: Currentstatus and future prospects. Neurosci. Biobehav. Rev. 30, 791–807,http://dx.doi.org/10.1016/j.neubiorev.2006.06.005.
Bäckman, L., et al., 2011. Dopamine D1 receptors and age differences in brain acti-vation during working memory. Neurobiol. Aging 32, 1849–1856.
Baltes, P.B., Baltes, M.M., 1990. Psychological perspectives on successful aging: themodel of selective optimization with compensation. In: Baltes, P.B., Baltes, M.M.(Eds.), Successful Aging: Perspectives from the Behavioral Sciences. CambridgeUniversity Press, New York, pp. 1–34.
Baltes, P.B., Reuter-Lorenz, P.A., Rösler, F., 2006. Lifespan Development and the Brain:The Perspective of Biocultural Co-Constructivism. Cambridge University Press,Cambridge, UK, http://dx.doi.org/10.1017/CBO9780511499722.
Berridge, K.C., Robinson, T.E., 2003. Parsing reward. Trends Neurosci. 26, 507–513.Brehmer, Y., Westerberg, H., Bellander, M., Fürth, D., Karlsson, S., Bäckman, L., 2009.
Working memory plasticity modulated by dopamine transporter genotype. Neu-rosci. Lett. 467, 117–120.
Bronfenbrenner, U., 1979. The Ecology of Human Development. Harvard UniversityPress, Cambridge, MA.
Clark, A., 2001. Where brain, body, and world collide. In: Edelman, G.M., Changeux,J.-P. (Eds.), The brain. Transaction, London, pp. 257–380.
Cools, R., Clark, L., Robbins, T.W., 2004. Differential responses in human striatumand prefrontal cortex to changes in object and rule relevance. J. Neurosci. 24,1129–1135, http://dx.doi.org/10.1523/JNEUROSCI.4312-03.2004.
Cools, R., D’Esposito, M., 2011. Inverted U-shaped dopamine action on humanworking memory and cognitive control. Biol. Psychiatry 69, e113–e125,http://dx.doi.org/10.1016/j.biopsych.2011.03.028.
Court, J., Martin-Ruiz, C., Piggott, M., Spurden, D., Griffiths, M., Perry, E., 2001. Nico-tinic receptor abnormalities in Alzheimer’s disease. Biol. Psychiatry 49, 175–184,http://dx.doi.org/10.1016/S0006-3223(00)01116-1.
Crespi, E.J., Denver, R.J., 2005. Ancient origins of human developmental plasticity.Am. J. Hum. Biol. 17, 44–54.
Crews, F., He, J., Hodge, C., 2007. Adolescent cortical development: a criticalperiod of vulnerability for addiction. Pharmacol. Biochem. Behav. 86, 189–199,http://dx.doi.org/10.1016/j.pbb.2006.12.001.
DeVito, L.M., Eichenbaum, H., 2011. Memory for the order of events in specificsequences: contribution of the hippocampus and medial prefrontal cortex. J.Neurosci. 31, 3169–3175, http://dx.doi.org/10.1523/JNEUROSCI. 4202-10.2011.
Diamond, A., 1996. Evidence for the importance of dopamine for prefrontal cor-tex functions early in life. Philos. Trans. R. Soc. London Ser. B: Biol. Sci. 351,1483–1493, http://dx.doi.org/10.1098/rstb.1996.0134.
Diamond, A., Briand, L., Fossella, J., Gehlbach, L., 2004. Genetic and neurochemicalmodulation of prefrontal cognitive functions in children. Am. J. Psychiatry 161,125–132, http://dx.doi.org/10.1176/appi.ajp.161.1.125.
Dumontheil, I., Roggeman, C., Ziermans, T., Peyrard-Janvid, M., Matsson, H., Kere, J.,Klingberg, T., 2011. Influence of the COMT genotype on working memory andbran activity changes during development. Biol. Psychiatry 70, 222–229.
Eppinger, B., et al., 2008. Better or worse than expected? Aging, learning, and theERN. Neuropsychologia 46, 521–538.
Eppinger, B., Hämmerer, D., Li, S.-C., 2011. Neuromodulation of reward-based learn-ing and decision making in human aging. Ann. N. Y. Acad. Sci. 1235, 1–17,http://dx.doi.org/10.1111/j. 1749-6632.2011.06230.x.
Eppinger, B., Mock, B., Kray, J., 2009. Developmental differences in learning and errorprocessing: evidence from ERPs. Psychophysiology 46, 1043–1053.
Erixon-Lindroth, N., Farde, L., Wahlin, T.B., Sovago, J., Halldin, C., Bäckman, L., 2005.The role of the striatal dopamine transporter in cognitive aging. Psychiatry Res.138, 1–12.
Field, T., Diego, M., Hernandez-Reif, M., Salman, F., Schanberg, S., Kuhn, C., et al.,2002. Prenatal anger effects on the fetus and neonate. J. Obstet. Gynaecol. 22,260–266.
Field, T., Hernandez-Reif, M., Diego, M., Figueiredo, B., Schanberg, S., Kuhn, C., et al.,2006. Prenatal cortisol, prematurity and low birth weight. Infant Behav. Dev. 29,268–275.
Friston, K.J., 2010. The free energy principle: a unified brain theory? Nat. Rev. Neu-rosci. 11, 127–138.
Friston, K.J., Tononi, G., Reeke, G.N., Sporns, O., Edelman, G.M., 1994. Value-dependent selection in the brain: simulation in a synthetic neural model.Neuroscience 59, 229–243.
Frank, M.J., Doll, B.B., Oas-Terpstra, J., Moreno, F., 2009. Prefrontal and striataldopaminergic genes predict individual differences in exploration and exploita-tion. Nat. Neurosci. 12, 1062–1068.
Freund, A.M., 2008. Successful aging as management of resources: the role of selec-tion, optimization, and compensation. Res. Hum. Dev. 5, 94–106.
Ford, D.H., 1987. Humans as Self-Constructing Living Systems: A DevelopmentalPerspective on Behavior and Personality. Erlbaum, Hillsdale.
Gatzke-Kopp, L.M., 2011. The canary in the coalmine: the sensitivity of mesolimbicdopamine to environmental adversity during development. Neurosci. Biobehav.Rev. 35, 794–803.
Gollwitzer, P.M., Sheeran, P., Trötschel, R., Webb, T.L., 2011. Self-regulation of prim-ing effects on behavior. Psychol. Sci. 22, 901–907.
Goschke, T., 2004. From free will to self-determination. Cognitive and volitionalmechanisms of intentional action control. Psychol. Rundsch. 55, 186–197,http://dx.doi.org/10.1026/0033-3042.55.4.186.
Gottlieb, G., 1998. Normally occurring environmental and behavioral influences ongene activity: from central dogma to probabilistic epigenesis. Psychol. Rev. 109,211–218, http://dx.doi.org/10.1037/0033-295X.109.2.211.
Groothius, T.G.G., Muller, W., von Engelhardt, N., Carere, C., Eising, C., 2005. Maternalhormones as a tool to adjust offspring phenotype in avian species. Neurosci.Biobehav. Rev. 29, 329–352.
Hämmerer, D., Li, S.-C., Müller, V., Lindenberger, U., 2011. Life span differences inelectrophysiological correlates of monitoring gains and losses during probabilis-tic reinforcement learning. J. Cogn. Neurosci. 23, 579–592.
Hämmerer, D., Eppinger, B., 2012. Dopaminergic and prefrontal contributions toreward-based learning and outcome monitoring during child development andaging. Dev. Psychol., http://dx.doi.org/10.1037/a0027342 (advance online pub-lication).
Hämmerer, D., Biele, G., Müller, V., Thiele, H., Nürnberg, P., Heekeren, H.R., Li, S.-C.,2013. Effects of PPP1R1B (DARPP-32) polymorphism on feedback-related brainpotentials across the life span. Front. Psychol., 4 (Article 89).
Haycock, J.W., Becker, L., Ang, L., Furukawa, Y., Hornykiewick, O., Kish, S.J.,2003. Marked disparity between age-related changes in dopamine and otherpresynaptic dopaminergic markers in the human striatum. J. Neurochem. 87,574–585, http://dx.doi.org/10.1046/j. 1471-4159.2003.02017.x.
Heckhausen, J., Wrosch, C., Schulz, R., 2010. A motivational theory of life-span devel-opment. Psychol. Rev. 117, 32–60, http://dx.doi.org/10.1037/a0017668.
Heitland, I., Oosting, R.S., Baas, J.M.P., Massar, S.A.A., Kenemans, J.L., Böcker, K.E.B.,2012. Genetic polymorphisms of the dopamine and serotonin systems modulateneurophysiological response to feedback and risk-tasking in health humans.Cogn. Affect. Behav. Neurosci. 12, 678–691.
Herlenius, E., Lagercrantz, H., 2004. Development of neurotrans-mitters during critical periods. Exp. Neurol. 190, S8–S21,http://dx.doi.org/10.1016/j.expneurol.2004.03.027.
Hertzog, C., Kramer, A.F., Wilson, R.S., Lindenberger, U., 2009. Enrichment effectson adult cognitive development. Can the functional capacity of older adults bepreserved and enhanced. Psychol. Sci. Public Interest 9, 1–65.
Holroyd, C.B., Coles, M.G.H., 2002. The neural basis of human error processing: rein-forcement learning, dopamine, and the error-related negativity. Psychol. Rev.109, 679–709 http://dx.doi.org/10.1037/0033-295X.109.4.679
Inoue, M., et al., 2001. Age-related reduction of extrastriataldopamine D2 receptor measured by PET. Life Sci. 20, 1079–1084,http://dx.doi.org/10.1016/S0024-3205(01)01205-X.
Jimura, K., Locke, H.S., Braver, T.S., 2010. Prefrontal cortex mediation of cognitiveenhancement in rewarding motivational contexts. Proc. Natl. Acad. Sci. U.S.A.107, 8871–8887.
Jucaite, A., Forssberg, H., Karlsson, P., Halldin, C., Farde, L., 2010. Age-relatedreduction in dopamine D1 receptors in the human brain: from late childhoodto adulthood, a positronemission tomography study. Neuroscience 167 (1),104–110.
Kaasinen, V., et al., 2000. Age-related dopamine D2/D3 receptor loss inextrastriatal regions of the human brain. Neurobiol. Aging 21, 563–568,http://dx.doi.org/10.1016/S0197-4580(00)00149-4.
Kalivas, P.W., Volkow, N.D., 2005. The neural basis of addiction: a pathol-ogy of motivation and choice. Am. J. Psychiatry 162, 1403–1413,http://dx.doi.org/10.1176/appi.ajp.162.8.1403.
Kudielka, B.M., Buske-Kirschbaum, A., Hellhammer, H.D., Kirschbaum, C., 2004.HAP axis responses to laboratory psychosocial stress in healthy elderly adults,younger adults, and children: impact of age and gender. Psychoneuroen-docrinology 29, 83–98.
Landau, S.M., Lal, R., O’Neil, J.P., Baker, S., Jagust, W.J., 2005. Stri-atal dopamine and working memory. Cereb. Cortex 19, 445–454,http://dx.doi.org/10.1093/cercor/bhn095.
Lataster, J., Collip, D., Ceccarini, J., Haas, D., Booij, L., Os, J.V., et al., 2011. Psychoso-cial stress is associated with in vivo dopamine release in human ventromedialprefrontal cortex: a positron emission tomography study using [18F]fallypride.Neuroimage 58, 1081–1089.
Lewandowsky, S., Murdock Jr., B.B., 1989. Memory for serial order. Psychol. Rev. 96,25–57.
Li, S.-C., 2003. Biocultural orchestration of developmental plasticityacross levels: the interplay of biology and culture in shaping themind and behavior across the lifespan. Psychol. Bull. 129, 171–194,http://dx.doi.org/10.1037/0033-2909.129.2.171.
Li, S.-C., Lindenberger, U., Bäckman, L., 2010. Dopaminergic modulation ofcognition across the lifespan. Neurosci. Biobehav. Rev. 34, 625–638,http://dx.doi.org/10.1016/j.neubiorev.2010.02.003.
Li, S.-C., Lindenberger, U., Sikström, S., 2001. Aging cognition: fromneuromodulation to representation. Trends Cogn. Sci. 5, 479–486,http://dx.doi.org/10.1016/S1364-6613(00)01769-1.

Author's personal copy
2208 S.-C. Li / Neuroscience and Biobehavioral Reviews 37 (2013) 2201–2208
Li, S.-C., Passow, S., Nietfeld, W., Schröder, J., Bertram, L., Heekeren, H.R.,et al., 2013a. Striatal dopamine modulates attentional control of auditoryperception. DARPP-32 genotype effects on behavior and cortical evoked poten-tials. Neuropsychologia 51, 1649–1661.
Li, S.-C., Papenberg, G., Nagel, I.E., Preuschhof, C., Schröder, J., Nietfeld, W., et al.,2013b. Aging magnifies effects of dopamine transporter and D2 receptor geneson backward serial memory. Neurobiol. Aging 34, 358e1–358e10.
Li, S.-C., Sikström, S., 2002. Integrative neurocomputational perspectives on cogni-tive aging, neuromodulation, and representation. Neurosci. Biobehav. Rev. 26,795–808, http://dx.doi.org/10.1016/S0149-7634(02)00066-0.
Lindenberger, U., Nagel, I.E., Chicherio, C., Li, S.-C., Heekeren, H.R., Bäck-man, L., 2008. Age-related decline in brain resources modulatesgenetic effects on cognitive functioning. Front. Neurosci. 2, 234–244,http://dx.doi.org/10.3389/neuro.01.039.2008.
Liotti, M., Pliszka, S.R., Perez, R., Kothmann, D., Woldorff, M.G., 2005. Cortex 41,377–388, http://dx.doi.org/10.1016/S0010-9452(08)70274-0.
Lövdén, M., Bäckman, L., Lindenberger, U., Schaefer, S., Schmiedek, F., 2010. A theo-retical framework for the study of adult cognitive plasticity. Psychol. Bull. 136,659–676.
Luciana, M., Wahlstrom, D., Porter, J.M., Collins, P.F., 2012. Dopaminergic mod-ulation of incentive motivation in adolescence: age-related changes insignaling, individual differences, and implications for the development of self-regulation. Dev. Psychol., http://dx.doi.org/10.1037/a0027432 (advance onlinepublication).
Mattay, V.S., Goldberg, T.E., Fera, F., Hariri, A.R., Tessitore, A., Egan, M.F., et al., 2003.Catechol-o-methyltransferase val158-met genotype and individual variationin the brain response to amphetamine. Proc. Natl. Acad. Sci. 100, 6186–6191,http://dx.doi.org/10.1073/pnas.0931309100.
McNab, F., Varrone, A., Farde, L., Lucaite, A., Bystritsky, P., Forssberg, H., et al., 2009.Science 323, 800–802.
Meyer-Lindenberg, A., Weinberger, D.R., 2006. Intermediate phenotypes and genet-ics of psychiatric disorders. Nat. Rev. Neurosci. 7, 818–827.
Miller, E.K., Cohen, J.D., 2001. An integrative theory of pre-frontal cor-tex function. Annu. Rev. Neurosci. 24, 167–202, http://dx.doi.org/10.1146/annurev.neuro.24.1.167.
Mitsis, E.M., et al., 2009. Age-related decline in nicotinic receptor avail-ability with [123I]5-IA-85380 SPECT. Neurobiol. Aging 30, 1490–1497,http://dx.doi.org/10.1016/j.neurobiolaging.2007.12.008.
Montgomery, J.D., Mehta, M.A., Grasby, P.M., 2006. Is psychological stress in humansassociated with increased striatal dopamine levels?: a [11C]raclopride PET study.Synapse 60, 124–131.
Nagel, I.E., Chicherio, C., Li, S.-C., Oertzen, T.v., Sander, T., Villringer, A., Heekeren,H.R., Bäckman, L., Lindenberger, U., 2008. Human aging magnifies genetic effectson executive functioning and working memory. Front. Hum. Neurosci. 2, 1–8,http://dx.doi.org/10.3389/neuro.09.001.2008.
Niv, Y., Joel, D., Dayan, P., 2006. A normative perspective on motivation. Trends Cogn.Sci. 10, 375–381.
Noudoost, B., Moore, T., 2011. The role of neuromodulators in selective attention.Trends Cogn. Sci. 15, 585–591, http://dx.doi.org/10.1016/j.tics.2011.10.006.
O’Reilly, R.C., Frank, M.J., 2006. Making working memory work: a computationalmodel of learning in the prefrontal cortex and basal ganglia. Neural Comput. 18,283–328 http://dx.doi.org/10.1162/089976606775093909
Papenberg, G., Bäckman, L., Nagel, I.E., Nietfled, W., Schröder, J., Bertram, L., et al.,2013. Dopaminergic gene polymorphisms affect long-term forgetting in oldage: further support for the magnification hypothesis. J. Cogn. Neurosci. 25,571–579.
Pennartz, C.M.A., Berke, J.D., Graybiel, A.M., Ito, R., Lansink, C.S., Voorn, P.,2009. Corticostriatal interactions during learning, memory processing, anddecision making. J. Neurosci. 14, 12831–12838, http://dx.doi.org/10.1523/JNEUROSCI.3177-09.2009.
Pirker, W., et al., 2000. Imaging serotonin and dopamine transporters with 123I-beta-CIT SPECT: binding kinetics and effects of normal aging. J. Nucl. Med. 41,36–44.
Posner, M.I., Rothbart, M.K., Sheese, B.E., Tang, Y., 2007. The anterior cingulate gyrusand the mechanism of self-regulation. Cogn. Affect. Behav. Neurosci. 7, 391–395.
Rangel, A., Camerer, C., Montague, R.P., 2008. A framework for studying the neuro-biology of value-based decision making. Nat. Rev. Neurosci. 9, 545–556.
Robbins, P., Aydede, M., 2008. The Cambridge Handbook of Situated Cognition.Cambridge University Press, Cambridge, UK.
Schultz, W., 2007. Behavioral dopamine signals. Trends Neurosci. 30, 203–209,http://dx.doi.org/10.1016/j.tins.2007.03.007.
Schultz, W., 2010. Dopamine signals for reward value and risk: basic and recent data.Behav. Brain Funct. 6, 1–9, doi 10.1186/1744-9081-6-24.
Shohamy, D., Adcock, R.A., 2010. Dopamine and adaptive memory. Trends Cogn. Sci.14, 464–472, http://dx.doi.org/10.1016/j.tics.2010.08.002.
Söderqvist, S., Nutley, S.B., Peyrard-Janvid, M., Matsson, H., Humphreys, K., Kere, J.,Klingberg, T., 2011. Dopamine, working memory, and training-induced plas-ticity: implications for developmental research. Dev. Psychol. 48, 836–843,http://dx.doi.org/10.1037/a0026179.
Spangler, G., Johann, M., Ronai, Z., Zimmermann, P., 2009. Genetic and environmentalinfluences on attachment disorganization. J. Psychol. Psychiatry 50, 952–961,http://dx.doi.org/10.1111/j. 1469-7610.2008.02054.x.
Störmer, S., Passow, V.S., Biesenack, J., Li, S.-C., 2011. Dopaminergic and cholin-ergic modulations of visual-spatial attention and working memory. Insightsfrom molecular genetic research and implications for adult cognitive devel-opment. Dev. Psychol., http://dx.doi.org/10.1037/a0026198 (advance onlinepublication).
Tarazi, F.I., Baldessarini, R.J., 2000. Comparative postnatal development of dopamineD1, D2, and D4, receptors in rat’s forebrain. Int. J. Dev. Neurosci. 18, 29–37,http://dx.doi.org/10.1016/S0736-5748(99)00108-2.
Tunbridge, E.M., Weickert, C.S., Kleinman, J.E., Herman, M.M., Chen, J., Kolachana,B.S., et al., 2007. Catechol-o-Methyltransferase enzyme activity and proteinexpression in human prefrontal cortex across the postnatal lifespan. Cereb.Cortex 17, 1206–1212, http://dx.doi.org/10.1093/cercor/bhl032.
Vijayraghavan, S., et al., 2007. Inverted-U dopamine D1 receptor actions on pre-frontal neurons engaged in working memory. Nat. Neurosci. 10, 376–384,http://dx.doi.org/10.1038/nn1846.
Vizi, S.E., Lajtha, A., 2008. Handbook of Neurochemistry and Molecular Neurobio-logy: Neurotransmitter Systems, 3rd ed. Springer, New York.
Volkow, N.D., Wang, G.-J., Baler, R.D., 2011. Reward, dopamine and the con-trol of food intake: implications for obesity. Trends Cogn. Sci. 15, 37–46,http://dx.doi.org/10.1016/j.tics.2010.11.001.
Vygotsky, L.S., 1978. Mind in Society. Harvard University Press, Cambridge. MA.Yamamoto, M., et al., 2002. Age-related decline for serotonin trans-
porters in living human brain of healthy males. Life Sci. 71, 751–757,http://dx.doi.org/10.1016/S0024-3205(02)01745-9.
Wahlstrom, D., White, T., Hooper, C.J., Vrshek-Schallhorn, S., Oetting, W.S., Brott,M.J., et al., 2007. Association of the catecholamine-O-methyltranferase (COMT)gene to prefrontally-mediated cognitions in adolescents. Biol. Psychiatry 6 (61),626–632.
Copyright © 2022 FDOKUMEN




















