
Adenylate cyclase in the rat pancreas properties and stimulation by hormones

Adenylate cyclase toxin translocates across target cellmembrane without forming a poremmi_7077 1550..1562
Adriana Osickova,1† Jiri Masin,1†
Catherine Fayolle,2,3† Jan Krusek,4 Marek Basler,1
Eva Pospisilova,1 Claude Leclerc,2,3 Radim Osicka1
and Peter Sebo1,5*1Institute of Microbiology AS CR v.v.i., Prague2Institut Pasteur, Unité de Régulation Immunitaire etVaccinologie, Paris, France3INSERM U883, Paris, France4Institute of Physiology AS CR, v.v.i., Prague5Institute of Biotechnology AS CR v.v.i., Academy ofSciences of the Czech Republic, Prague, CzechRepublic
Summary
The adenylate cyclase toxin-haemolysin of Borde-tella (CyaA) targets CD11b+ myeloid phagocytes andtranslocates across their cytoplasmic membrane anadenylate cyclase (AC) enzyme that catalyses con-version of cytosolic ATP into cAMP. In parallel, CyaAacts as a cytolysin forming cation-selective pores,which permeabilize cell membrane and eventuallyprovoke cell lysis. Using cytolytic activity, potas-sium efflux and patch-clamp assays, we show that acombination of substitutions within the pore-forming(E570Q) and acylation-bearing domain (K860R)ablates selectively the cell-permeabilizing activity ofCyaA. At the same time, however, the capacity ofsuch mutant CyaA to translocate the AC domainacross cytoplasmic membrane into cytosol of mac-rophage cells and to elevate cellular cAMP concen-trations remained intact. Moreover, the combinationof E570Q+K860R substitutions suppressed theresidual cytolytic activity of the enzymatically inac-tive CyaA/OVA/AC- toxoid on CD11b-expressingmonocytes, while leaving unaffected the capacity ofthe mutant toxoid to deliver in vitro a reporter CD8+
T cell epitope from ovalbumin (OVA) to the cytosolicpathway of dendritic cells for MHC class I-restrictedpresentation and induce in vivo an OVA-specificcytotoxic T cell response. CyaA, hence, employs a
mechanism of AC enzyme domain translocationacross cellular membrane that avoids passageacross the cytolytic pore formed by toxin oligomers.
Introduction
The bi-functional RTX adenylate cyclase toxin-haemolysin (CyaA, ACT or AC-Hly) is a key virulencefactor of Bordetella pertussis (Vojtova et al., 2006). Its1706-residue-long polypeptide is a fusion of an N-terminaladenylate cyclase (AC) enzyme domain (~400 residues)to a pore-forming RTX haemolysin (Repeat in ToXin cytol-ysin) of ~1300 residues (Glaser et al., 1988). The latterharbours the sites of activation of proCyaA to CyaA bycovalent post-translational palmitoylation of e-aminogroups of Lys860 and Lys983, as well as the numerous RTXrepeats. These form ~40 calcium binding sites, theloading of which is required for cytotoxic activity of CyaA(Rose et al., 1995; Masin et al., 2005).
The toxin primarily targets host myeloid phagocytesexpressing the aMb2 integrin known as CD11b/CD18, CR3or Mac-1 (Guermonprez et al., 2001). CyaA inserts intocytoplasmic membrane of cells and translocates the ACenzyme domain into their cytosol without the need forendocytosis (Gordon et al., 1988; Schlecht et al., 2004).Inside cells, the AC is activated by calmodulin and cataly-ses uncontrolled conversion of cellular ATP to cAMP, akey second messenger molecule provoking impairment ofbactericidal functions of phagocytes (Vojtova et al., 2006;Kamanova et al., 2008). At high CyaA doses(> 100 ng ml-1) substantial enzymatic dissipation of ATPinto cAMP by the enzymatic activity of CyaA becomescytotoxic and promotes rapid apoptosis, or necrotic deathand subsequent lysis of CD11b+ monocytes (Khelef et al.,1993; Basler et al., 2006).
Recently, we showed that CyaA binds N-linked oli-gosaccharides of its receptor CD11b/CD18 (Morova et al.,2008). This suggests that low specificity interactions withglycans of ubiquitous cell surface proteins or glycolipidsmay account for the about two orders of magnitudereduced, but readily detectable capacity of CyaA to pen-etrate also cells lacking CD11b/CD18. Due to theextremely high specific catalytic activity of the AC domain,CyaA was indeed found to substantially elevate cAMPalso in mammalian and avian erythrocytes, lymphocytes,
Accepted 25 January, 2010. *For correspondence. E-mail [email protected]; Tel. (+420) 241 062 762; Fax (+420) 241 062 152.†These authors contributed equally to this work.
Molecular Microbiology (2010) 75(6), 1550–1562 � doi:10.1111/j.1365-2958.2010.07077.xFirst published online 23 February 2010
© 2010 Blackwell Publishing Ltd

lymphoma, neuroblastoma, CHO or tracheal epithelialcells (Vojtova et al., 2006; Paccani et al., 2008).
Besides elevating cAMP, the toxin exhibits also a mod-erate haemolytic activity on mammalian and avianerythrocytes. This is due to its capacity to form smallcation-selective pores, of an estimated diameter of0.6–0.8 nm, which permeabilize cellular membrane andeventually provoke colloid-osmotic cell lysis (Bellalouet al., 1990; Ehrmann et al., 1991; Benz et al., 1994).Recently, we and others have shown that the pore-forming activity of CyaA synergizes with its cell-invasiveAC enzyme activity and contributes to the overall cytolyticpotency of CyaA on CD11b+ cells (Hewlett et al., 2006;Basler et al., 2006). Due to its intact pore-forming(haemolytic) capacity, the enzymatically inactive CyaA/AC- toxoid (Fayolle et al., 1996) still exhibits a fullhaemolytic activity on erythrocytes and in the absence ofosmoprotectants such as serum, also a residual about10-fold reduced cytolytic potency on CD11b-expressingmonocytes (Basler et al., 2006).
The haemolytic (pore-forming) and AC membrane trans-location (cell-invasive) activities of CyaA were found to bedissociable by low calcium concentration, low temperature(Rogel and Hanski, 1992) and by the extent and nature ofacylation of CyaA (Benz et al., 1994; Hackett et al., 1995;Havlicek et al., 2001; Masin et al., 2005). Moreover, thetwo activities differ substantially in sensitivity to charge-reversing or neutral substitutions of glutamates at positions509, 516, 570 and 581 within the hydrophobic domain(Osickova et al., 1999; Basler et al., 2006; Basler et al.,2007). The cell-invasive and pore-forming activities ofCyaA were thus proposed to be mutually independent andoperating in parallel in target cell membrane. The modelpredicts that two distinct CyaAconformers insert into targetcell membrane in parallel, one being the translocationprecursor, accounting for delivery of the AC domain acrosscellular membrane, with concomitant influx of calcium ionsinto cells, the other being a pore precursor eventuallyforming oligomeric pores and provoking potassium effluxfrom cells (Gray et al., 1998; Osickova et al., 1999; Basleret al., 2007; Fiser et al., 2007).
Here, we tested this model and show, by using specificCyaA mutants, that the pore-forming activity is notinvolved in translocation of the AC domain across targetcell membrane.
Results
Combined elimination of negative charge of glutamate570 and of acylation of lysine 860 ablatescell-permeabilizing capacity of CyaA
The working model of CyaA action predicts that it shouldbe possible to generate CyaA variants with selectivelyablated pore-forming (haemolytic) activity but preserving
the capacity to deliver the AC domain into cytosol of targetcells. We previously observed that the capacity of enzy-matically inactive CyaA/AC- toxoids to lyse cells can beup- or down-modulated by substitutions within the pore-forming domain (Benz et al., 1994; Osickova et al., 1999;Basler et al., 2006; Hewlett et al., 2006; Basler et al.,2007). Therefore, we sought to generate CyaA/AC- vari-ants exhibiting as low haemolytic and cytolytic (pore-forming) activities as possible. These toxoids were furthertagged at residue 233 by insertion of the Kb-restrictedCD8+ T-cell epitope SIINFEKL from ovalbumin (OVA), toenable assessment of the capacity to translocate theOVA/AC- domain polypeptide into cytosol of CD11b+
antigen presenting cells (APCs). Tagging with OVA was,indeed, previously shown to have no impact on the cell-invasive capacity of the AC domain polypeptide. It allowsdelivery and processing of the OVA/AC- by cytosolic pro-teasome and the presentation of the excised OVA epitopeon MHC Class I glycoproteins on cell surface. This, inturn, enables CyaA/OVA/AC--treated APCs to measurablystimulate OVA-specific CD8+ T cells in vitro and in vivo,providing an assay for OVA/AC- domain delivery into cells(Osicka et al., 2000).
To generate toxoids possibly lacking the cytolytic activ-ity, we combined the E570Q and K860R substitutionsshown to reduce the specific haemolytic activity of CyaAon sheep erythrocytes, where the E570Q substitution wasalso shown to reduce the cytolytic activity of CyaA/AC- onCD11b+ J774A.1 monocytes (Basler et al., 2006; Basleret al., 2007). The substitutions were engineered intoCyaA/OVA/AC- individually and in combination, and thespecific haemolytic and cytolytic activities of the con-structs were compared using sheep erythrocytes, usedhere as model CD11b- targets, and J774A.1 monocytesused as model CD11b+ targets respectively (Table 1). Inagreement with results obtained previously with CyaA/AC- lacking the OVA epitope (Basar et al., 1999; Masinet al., 2005; Basler et al., 2006; Basler et al., 2007), thetoxoids carrying individually the E570Q and K860R sub-stitutions exhibited a twofold reduced (55 � 8%) and nil(1 � 1%) relative haemolytic activity on erythrocytes,when compared with the activity of CyaA/OVA/AC- (OVA/AC-) taken as 100%. Moreover, the relative cytolytic activ-ity of the E570Q toxoid towards CD11b-expressingJ774A.1 cells was significantly (P < 0.001) reduced by afactor of about three (37 � 10%). In turn, as expectedfrom results obtained with an enzymatically active K860Rconstruct, despite its low haemolytic activity on CD11b-
erythrocytes, the K860R toxoid exhibited only a slightlyreduced relative cytolytic activity on CD11b-expressingJ774A.1 cells (72 � 22%). This confirmed that the defectcaused by the K860R substitution was rescued by inter-action with the CD11b/CD18 receptor (Masin et al., 2005).When combined with E570Q, however, the K860R sub-
Adenylate cyclase translocation: avoiding the pore 1551
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

stitution exhibited a clear additive effect in further reduc-ing the relative cytolytic activity of the E570Q+K860Rconstruct on J774A.1 cells down to 14 � 7% of the rela-tive activity of intact CyaA/AC-.
Pore-forming activity of CyaA is dispensable for deliveryof AC domain into cells
To allow quantification of the capacity of the AC domain totranslocate across the cytoplasmic membrane into thecytosol of cells, the E570Q and K860R substitutions werenext transferred into the enzymatically active (AC+) con-structs derived from CyaA233/OVA (CyaA/OVA). Theseconstructs were produced and purified in the same way asthe AC- toxoids (not shown) and were first characterizedon CD11b- sheep erythrocytes for cell binding, haemolyticand AC translocation capacities. The latter was assessedas cell-invasive AC activity and corresponds to theamount of adenylate cyclase enzyme that becomes inac-cessible to inactivation by extracellularly added trypsinfollowing translocation into cytosol of erythrocytes. ThisAC enzyme activity, protected from tryptic digestion bycellular membrane, is measured upon cell washing andlysis in the AC assay buffer (Bellalou et al., 1990). Asshown in Fig. 1A, in agreement with results obtained fortoxins lacking the OVA epitope (Masin et al., 2005; Basleret al., 2007), the E570Q substitution selectively reducedonly the relative haemolytic activity and had no impact onerythrocyte binding or the capacity of CyaA/OVA to deliver
the AC domain into erythrocyte cytosol (invasive ACactivity). As further expected on the basis of previous work(Basar et al., 1999), the K860R substitution significantlyreduced the capacity of CyaA/OVA to bind and penetrateerythrocytes, yielding a sharp reduction of the relativehaemolytic and cell-invasive AC activities of the K860Rand E570Q+K860R mutants on erythrocytes.
It has to be noted that the haemolytic activity of CyaA isa highly cooperative function of the amount of cell-associated CyaA (Szabo et al., 1994), in line with therecent demonstration that oligomerization is a prerequi-site for CyaA pore formation (Vojtova-Vodolanova et al.,2009). Therefore, the specific haemolytic activities ofintact and mutated CyaAs had to be compared at equalamounts of cell-associated toxin per erythrocyte. Asshown in Fig. 2B, this was achieved by increasing theinput concentration of proteins bearing the K860R substi-tution fivefold over that of the intact toxin (25 and 5 mg ml-1
respectively). This allowed determination of the specifichaemolytic activity of all tested proteins under conditionsof ~23 ng of CyaA bound per 5 ¥ 108 erythrocytes. Asindeed shown in Fig. 1C, under these conditions theE570Q and K860R substitutions still exhibited a clearadditiveness in further reducing, by a factor of two, thealready impaired haemolytic activities of the singlymutated constructs. As further reported previously, theK860R substitution reduced by a factor of 2 to 2.5 also thespecific capacity of CyaA to translocate the AC domainacross the membrane of erythrocytes (Basar et al., 1999).
In contrast to the severe impact of the K860R substitu-tion on toxin activity towards CD11b- erythrocytes,however, both the E570Q and K860R substitutions werepreviously found to have no effect on the capacity of CyaAto bind and penetrate J774A.1 monocytes expressing theCD11b/CD18 receptor of CyaA (Masin et al., 2005; Basleret al., 2006). As indeed documented in Fig. 2A, when thetwo substitutions were combined in the same toxin mol-ecule, the enzymatically active CyaA/OVA/E570Q+K860Rconstruct, as well as the single mutants bearing theE570Q and K860R substitutions individually, exhibited anequal capacity to bind J774A.1 cells over the range oftoxin concentrations up to 2.5 mg ml-1. Moreover, asshown in Fig. 2B, the single and double mutated con-structs exhibited an identical capacity to deliver the ACdomain into cytosol of J774A.1 cells and to elevate cellu-lar cAMP concentrations as the intact CyaA. In turn, nocAMP elevation was observed with the CyaA/E570K+E581P protein that lacks the capacity to translo-cate the AC domain across target cytoplasmic membrane(Vojtova-Vodolanova et al., 2009) and served as controlfor CyaA leakage from endosomes upon receptor-mediated uptake.
As however, shown in Fig. 2C, when compared withintact CyaA, the singly mutated E570Q and the doubly
Table 1. Cytolytic activities of OVA/AC- and its derivatives on sheeperythrocytes and J774A.1 macrophages.
Protein
Haemolytic activityon erythrocytes(% of AC-)a
Lysis of J774A.1cells (% of AC-)b
AC- 100 � 5 100 � 10OVA/AC- 93 � 4 93 � 12OVA/E570Q/AC- 55 � 8** 37 � 10**OVA/K860R/AC- 1 � 1** 72 � 22*OVA/E570Q+K860R/AC- 1 � 1** 14 � 7**
a. Lysis of sheep erythrocytes was determined after 4.5 h as theamount of haemoglobin released upon incubation of 5 ¥ 108 RBC at37°C in the presence of 2 mM Ca2+ with 5 mg ml-1 of the given protein(Bellalou et al., 1990). The haemolytic activity of CyaA/AC-, whichyielded on average the lysis of 58% of erythrocytes under the givenconditions, was taken as 100% of relative haemolytic activity. Theresults represent the average of values obtained in four independentexperiments performed in duplicates � SD with two different proteinpreparations.b. Lysis of J774A.1 cells was determined as the amount of releasedlactate dehydrogenase from 105 cells upon 3 h of incubation with10 mg ml-1 of the appropriate protein at 37°C in DMEM. Under theseconditions CyaA/AC- provoked on average the lysis of 47.8% ofJ774A.1 cells, which was taken as 100% of relative cytolytic activity.The results represent the average of values � SD from four separateexperiments performed in duplicates and using two different proteinpreparations (*P < 0.05; **P < 0.001).
1552 A. Osickova et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

mutated E570Q+K860R constructs were importantlyimpaired in the capacity to elicit decrease of intracellularconcentration of potassium ions ([K+]i) in toxin-treatedJ774A.1 cells. While no cell lysis was detected over20 min by the assay for release of lactate dehydrogenase(data not shown), the [K+]i of J774A.1 cells exposed to3 mg ml-1 of intact CyaA decreased from 140 mM to wellbelow 30 mM already in 10 min upon toxin addition. Inturn, when the same amounts of the ER570Q+K860Rconstructs were used (3 mg ml-1), the [K+]i levels in cellsdid not decrease below 100 mM (Fig. 2C). Indeed, effluxof potassium from cells was previously shown to be thehallmark of insertion of the CyaA pore precursors into cellmembrane (Gray et al., 1998). These results, hence,suggest that the E570Q and E570Q+K860R substitutionsselectively impaired the capacity of CyaA to form oligo-meric pores and permeabilize cellular membrane, but didnot affect the capacity of the mutants to translocate theAC domain across cellular membrane.
This conclusion was further supported by an importantlyreduced relative cytolytic activity of the correspondingE570Q/AC- and E570Q+K860R/AC- toxoids, as dis-cussed above (Table 1) and documented in detail inFig. 3A. The doubly mutated E570Q+K860R toxoid at3 mg ml-1 was essentially unable to provoke any detect-able release of lactate dehydrogenase from J774A.1 cellsin 3 h of incubation, while 20% of cells lysed in the pres-ence of equal amounts of intact toxoid.
AC
om
pensate
dactivity o
n R
BC
s (
%)
C
0
20
40
60
80
100
120A
ctivity
on R
BC
s (
%)
Binding
Hemolysis
Invasive AC activity
Cya
A
Cya
A/O
VA
Cya
A/O
VA
/E570Q
Cya
A/O
VA
/K860R
Cya
A/O
VA
/E570Q
+K
860R
Cya
A/O
VA
Cya
A/O
VA
/E570Q
Cya
A/O
VA
/K860R
Cya
A/O
VA
/E570Q
+K
860R
**
****
0
5
10
15
20
25
30
35
5 15 25 35
Input CyaA (µg/ml)
CyaA
bound
to R
BC
s (
ng/m
l)
CyaA/OVACyaA/OVA/E570Q
CyaA/OVA/K860RCyaA/OVA/E570Q+K860R
B
0
20
40
60
80
100
120
140
Compensated RBC binding
Binding-compensated hemolysis
**
**
**
Binding-compensated invasive AC activity
** **
Fig. 1. Substitutions in the pore-forming and acylation domainsadditively decrease the specific haemolytic activity of CyaA.A. Sheep erythrocytes (5 ¥ 108 ml-1) in TNC buffer were incubatedwith 5 mg ml-1 of enzymatically active CyaA-derived proteins at37°C. After 30 min, aliquots of cells suspensions were washedrepeatedly to remove unbound CyaA and used to determine theamount of cell-associated and cell-invasive AC enzyme activities.Haemolytic activity was measured after 5 h of incubation as theamount of released haemoglobin by photometric determination(A541). Activities of intact CyaA were taken as 100% of relativeactivities. Under the given conditions, the 100% activitycorresponded on average to binding of 23.4 ng CyaA per ml oferythrocytes (2.033 nmol cAMP min-1 ml-1), of which 14.25 ng ml-1
of CyaA (1239 nmol cAMP min-1 ml-1) had translocated the ACdomain into eryhtrocyte cytosol, so that this was protected againstinactivation by extracellularly added trypsin (invasive AC activity).Lysis of 66.2% of erythrocytes in 5 h was taken as 100% activity.B. Erythrocytes were incubated as above with the indicatedconcentrations of the enzymatically active CyaA-derived proteinsfor 30 min, washed, and the amount of cell-associated AC enzymeactivity was determined.C. To compensate for the reduced cell binding activity, theconcentration of proteins with the K860R substitution wasincreased to 25 mg ml-1, as compared with 5 mg ml-1 for CyaA/OVAand CyaA/OVA/E570Q, allowing to achieve binding of equalamounts of each protein per ml of erythrocytes (~23 ng ml-1).Activities of CyaA/OVA were taken as 100%. The results representaverage values from at least three independent experimentsperformed in duplicates. The asterisks indicate statisticallysignificant differences (**P < 0.001) from activities of CyaA (A) orCyaA/OVA (C).
Adenylate cyclase translocation: avoiding the pore 1553
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

To corroborate on this, we analysed the cell-permeabilizing capacity of the CyaA/OVA/E570Q+K860R/AC- toxoid construct in patch-clamp experiments on wholecells. As documented in Fig. 3B by a representative indi-vidual recording of ion currents across the membrane of asingle patch-clamped J774A.1 cell at 1 mg ml-1 of toxoid,following an initial lag of about 3 min, the J774A.1 cellswere progressively and massively permeabilized by CyaA/OVA/AC-, with currents across cell membrane reaching-3000 pA within 10 min. In contrast, exposure to doublymutated CyaA/OVA/E570Q+K860R/AC- toxoid reproduc-ibly caused only a transient and minimal initial permeabili-zation of the cells, with currents across cell membrane notexceeding -200 pA and returning close to zero within10 min after toxoid addition. Quite similar picture wasobserved when toxoid concentrations were elevated to10 mg ml-1, which was the concentration used for compari-sons of relative cytolytic activity of toxoids summarized inTable 1. The 10-fold increase of concentration from 1 to10 mg ml-1 resulted in about twofold increase of the cur-rents produced across cell membrane by OVA/AC-, whileessentially no enhancement of cell permeabilization wasobserved even at the increased concentration of OVA/E570Q+K860R/AC- (note the expanded scale of y-axis formeasurements at 10 mg ml-1). These results, representa-
ACyaA
CyaA/OVA
CyaA/OVA/E570Q
CyaA/OVA/K860R
CyaA/OVA/E570Q+K860R
CyaA+M1/70CyaA/E570K+E581P
0
2
4
6
8
10
12
14
0 1 2 3
Input CyaA (µg/ml)
CyaA
bound
to 1
06
J7
74
A.1
ce
lls(n
g/m
l)
0.1
1
10
100
0 10 20 30 40 50
Toxin concentration (ng/ml)
pm
olcA
MP
/10
5J774A
.1cells
CyaA
CyaA/OVA
CyaA/OVA/E570Q
CyaA/OVA/K860R
CyaA/OVA/E570Q+K860R
CyaA+M1/70
CyaA/E570K+E581PB
[K+]i in
J774A
.1 c
ells
(mM
)
Time (min)
Inte
nsity r
atio
(450/5
10 n
m)
0.85
0.90
0.95
1.00
1.05
0 2 4 6 8 10
CyaA/OVA/E570Q
CyaA/OVA
CyaA/OVA/E570Q+K860R
CyaA/OVA/K860R
Buffer
140
60
30
C
Fig. 2. CyaA/OVA/E570Q+K860R binds and translocates intoCD11b+ monocytes without provoking leakage of intracellularpotassium ions.A. J774A.1 cells (106 ml-1) were incubated in DMEM for 30 min at4°C with CyaA-derived constructs, washed repeatedly and theamount of cell-associated AC activity was determined in celllysates. To block the CD11b/CD18 receptor, cells were incubatedfor 30 min with the CD11b-specific antibody M1/70 (Exbio, Vestec,Czech Republic) at a final concentration of 10 mg ml-1 prior toaddition of CyaA.B. The capacity of constructs to translocate the AC domain acrosscytoplasmic membrane was assessed as the capacity to penetratecells and convert cytosolic ATP to cAMP. J774A.1 cells wereincubated with 0–50 ng ml-1 of the various CyaA-derived constructsfor 30 min at 37°C and the amounts of accumulated cAMP weredetermined in cell lysates. As a control, the CD11b/CD18 receptorwas specifically blocked with the anti-CD11b antibody M1/70.Membrane penetration of CD11b/CD18-bound and endocytosedtoxin was controlled by using the doubly mutatedCyaA/E570K+E581P variant, which is intact for receptor binding butfails to translocate the AC domain across cell membrane andelevate cytosolic cAMP concentrations (Vojtova-Vodolanova et al.,2009).C. J774A.1 cells were loaded with the K+ sensitive fluorescentprobe PBFI/AM at 9.5 mM final external concentration and 25°C for45 min in the presence of Pluronic F-127 [0.05% (w/w)]. Cells werewashed in HBSS before 3 mg ml-1 of the indicated toxins wereadded. Fluorescence intensity ratio of PBFI (excitation wavelength340, emission wavelengths 450 and 510 nm) reflecting theintracellular K+ concentration was recorded every 15 s. The rightscale shows intracellular [K+]I values derived from calibrationexperiments (see Experimental procedures). No cell lysis, assessedas lactate dehydrogenase release, was observed within the timeinterval of the assay (data not shown). Results representative ofthree independent determinations performed in duplicates areshown.
1554 A. Osickova et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

tive of at least six measurements in three independentexperiments, demonstrate that combination of the E570Qand K860R substitutions had a major impact on the capac-ity of CyaA/OVA/AC- to permeabilize the membrane ofCD11b+ J774A.1 cells. At the same time, however, theenzymatically active version of the same construct wasfully capable to translocate the AC domain into J774A.1cells (cf. Fig. 2B). These results strongly suggest that thecell-permeabilizing (pore-forming) activity of CyaA was notrequired for AC domain translocation across the mem-brane of J774A.1 monocytes.
Membrane-permeabilizing activity of CyaA isdispensable for delivery of passenger antigens to thecytosolic MHC class I pathway
To corroborate these observations and assess the cellpenetration capacity of the AC- toxoids, we used a surro-gate assay to measure delivery of the reporter OVAepitope into the cytosolic processing compartment of theMHC class I antigen presentation pathway (Guermonprezet al., 1999; Schlecht et al., 2004). Towards this end, wedetermined the capacity of C57BL/6 mouse bone marrow-derived dendritic cells (BMDCs), used as APCs loadedwith the toxoids, to stimulate IL-2 release by B3Z T cellsthat selectively recognize on APC surface the complex ofKb MHC class I molecules with the SIINFEKL (OVA)peptide. It has, indeed, been previously shown that thecapacity of CyaA/AC- toxoids to translocate the AC-
domain across the cytoplasmic membrane into cytosol ofBMDCs is essential for the capacity of the toxoids topromote presentation of the delivered OVA epitope incomplex with the H-2Kb MHC class I molecules. This, inturn, is essential for specific in vitro stimulation of cytotoxicT cells to occur (Karimova et al., 1998a). Nevertheless, itwas important to confirm here that delivery of the OVAepitope for proteasome processing and subsequentpresentation was due to AC domain translocation intocytosol of BMDCs across their cytoplasmic membrane,and was not due to sampling of the added antigen by fluidphase uptake, or endocytosis. For this purpose, a doublymutated non-translocating OVA/E570K+E581P/AC-
toxoid variant was used, which bears a combination ofcharge-reversing and a-helix-breaking substitutions of
A
B
IN 0
(p
A)
0
20
40
60
80
100
120
0 10 20 30 40
Toxoid concentration (µg/ml)
LH
D r
ele
ase
fro
m10
5J774A
.1 c
ells
(%)
AC-
OVA/AC-
OVA/E570Q/AC-
OVA/K860R/AC -
OVA/E570Q+K860R/AC-
Time (min)
-3000
-2000
-1000
0
OVA/AC- 1 µg/ml
0 10.05.02.5 7.5
-8000
-6000
-4000
-2000
0
5.02.5 7.5 10.00
Time (min)
OVA/AC- 10 µg/ml
IN 0
(pA
)
0Time (min)
-3000
-2000
-1000
0
OVA/E570Q+K860R/AC- 1 µg/ml
10.05.02.5 7.5
IN 0
(pA
)IN
0 (
pA
)
Time (min)10.00
-8000
-6000
-4000
-2000
0
5.02.5 7.5
OVA/E570Q+K860R/AC- 10 µg/ml
Fig. 3. The E570Q+K860R toxoid fails to permeabilize J774A.1cells.A. Lysis of J774A.1 cells was determined as the amount of lactatedehydrogenase released from 105 cells upon 3 h of incubation with3, 10 and 30 mg ml-1 of the indicated protein at 37°C in DMEMwithout serum. The results represent the average of valuesobtained in two experiments performed in duplicates.B. Whole-cell patch-clamp measurements were performed at roomtemperature on individual J774A.1 cells exposed to 1 or 10 mg ml-1
of toxoids, as described in Experimental procedures. The showncurves are representative of six determinations from threeindependent experiments. Arrows indicate time of toxoid addition.
Adenylate cyclase translocation: avoiding the pore 1555
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

glutamates 570 and 581 in the transmembrane domain ofCyaA (Vojtova-Vodolanova et al., 2009). This constructwas previously found to exhibit a full capacity to bindCD11b/CD18-expressing cells (cf. Fig. 2A), while itscapacity to translocate the AC domain across target cellmembrane was ablated by the combination of E570K andE581P substitutions (cf. Fig. 2B).
As shown in Fig. 4A, the B3Z hybridoma cells were,indeed, effectively stimulated upon co-incubation withBMDCs loaded with the OVA/E570Q/AC- and OVA/E570Q+K860R/AC- toxoids. In contrast, no B3Z stimula-tion was observed upon co-incubation with BMDCsloaded with the OVA/E570K+E581P/AC- toxoid defectivein AC domain translocation across cell membrane. More-over, the OVA/E570Q/AC- and OVA/E570Q+K860R/AC-
toxoids induced B3Z lymphocyte stimulation by APCs withas high efficiency as the intact OVA/AC- toxoid. Theseresults confirm that the E570Q+K860R double mutantwas fully capable to translocate its OVA/AC- domain intoBMDC cytosol for processing of the delivered OVAepitope, while being essentially unable to permeabilizethe J774A.1 cells. These results suggest that the cell-permeabilizing (pore-forming) activity of CyaA was neitherrequired for AC domain translocation across cellular mem-brane, nor did it play a role in the capacity of CyaA todeliver passenger epitopes into APC cytosol.
To corroborate the in vitro antigen delivery assays, wefurther assessed the capacity of the toxoids to primeOVA-specific cytotoxic CD8+ T lymphocytes (CTL) in vivo.Some 50 mg of the various OVA-toxoids was injectedintravenously into C57BL/6 mice and 1 week later theOVA-specific CTL responses were assessed in immu-nized mice by an in vivo killing assay. C57BL/6 micereceived i.v. injection of a mixture (1:1) of OVA (SIINFEKL)peptide-loaded CFSEhigh and unloaded CFSElow spleno-cytes, followed 1 day later by FACS analysis of CFSE-labelled cells. As shown in Fig. 4B, immunization of micewith the mock AC- toxoid did not induce any SIINFEKL-specific CTL activity in vivo. In turn, immunization with thenon-cytolytic OVA/E570Q+K860R/AC- toxoid induced thesame OVA-specific in vivo CTL killing response, as thecytolytic OVA/AC- toxoid without substitutions. Theseresults show that the cell-permeabilizing activity of CyaAwas dispensable also for the in vivo capacity of the CyaA/OVA/AC- toxoids to deliver an AC-inserted passengerantigen into cytosol of APCs.
Discussion
We demonstrate here that translocation of the AC domainof CyaA across membrane of CD11b/CD18 receptor-expressing myeloid target cells does not depend on thecapacity of the toxin to form pores and permeabilize thecellular membrane.
As summarized in the model proposed in Fig. 5, wehave previously reported that the balance between thetwo activities of CyaA can be shifted by mutations, or byalternative acylation of CyaA. Enhancement of the pore-forming (haemolytic) activity at the expense of the capac-ity to deliver AC into cells was, indeed, observed uponlysine substitutions of glutamates 509, 516 and 581(Osickova et al., 1999; Basler et al., 2007), or upon block-ing of AC translocation by the 3D1 monoclonal antibody(Gray et al., 2001). In turn, a shift in the opposite directionwas observed for the recombinant r-Ec-CyaA, acylated inEscherichia coli by palmitoleic (C16:1) residues, as com-pared with the native (C16:0) palmitylated Bp-CyaA pro-duced by B. pertussis. The r-Ec-CyaA was found to exhibitabout fourfold reduced haemolytic activity and about10-fold lower pore-forming activity in planar lipid bilayersthan Bp-CyaA (Benz et al., 1994). Both CyaA forms were,however, equally active in penetrating cellular membraneand in translocation of the AC domain into erythrocytes(Hackett et al., 1995; Havlicek et al., 2001). Moreover, theCyaA/E570Q construct was recently found to exhibit a fullcapacity to deliver the AC domain into both erythrocytesand J774A.1 macrophages, while exhibiting a reducedhaemolytic activity and a lower specific pore-formingcapacity in planar lipid bilayers than intact CyaA. Conse-quently, the CyaA/E570Q/AC- toxoid exhibited a twofoldreduced cytolytic activity on J774A.1 cells (Basler et al.,2006; Basler et al., 2007).
Despite the above mentioned and the many mutantCyaAs characterized, the question remained whether for-mation of a membrane pore by CyaA can be avoidedduring translocation of the AC domain across the mem-brane of CD11b-expressing cells. It is worth mentioningthat, based on previous comparisons of haemolyticpotency of the intact r-Ec-CyaA with that of the nativeBp-CyaA purified from B. pertussis, the specifichaemolytic activity of the here described r-Ec-CyaA/OVA/E570Q+K860R/AC- toxoid on sheep erythrocytes wouldbe reduced by about three orders of magnitude. Theresidual specific cytolytic activity of r-Ec-CyaA/OVA/E570Q+K860R/AC- on CD11b-expressing cells wouldthen be estimated to be about 50-fold lower than that ofBp-CyaA, while the specific capacity of both proteins todeliver the AC domain into these cells would be the same(using intact r-Ec-CyaA as 100% invasive AC activity stan-dard for comparisons).
The CyaA/OVA/E570Q+K860R mutant described hereis the first construct that is essentially unable to elicitpotassium efflux and to permeabilize cells by formation ofCyaA pores, while retaining a full capacity to translocatethe AC domain across cellular membrane. This resultanswers the question on the role of CyaA pores in ACtranslocation. It shows that on its way into cell cytosol, thetranslocating AC domain is bypassing the cation-selective
1556 A. Osickova et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

Fig. 4. Toxoid with E570Q+K860Rsubstitutions delivers the OVA T-cell epitopefor presentation by MHC class I moleculesand induction of CD8+ CTLs.A. BMDC (3 ¥ 105 cells/well) used as APCswere incubated in the presence of indicatedconcentrations of the toxoids harbouring theOVA epitope or with mock CyaA/AC-. Uponco-culture for 24 h with B3Z T cells (1 ¥ 105
cells/well), IL-2 secretion by the stimulatedB3Z cells was determined by the CTLLproliferation method. Results are expressedas Dcpm of incorporated [3H]thymidine (cpmin the presence of toxoid - cpm in theabsence of toxoid) � SD and arerepresentative of five independentexperiments.B. Analysis of the induction of OVA(SIINFEKL)-specific CTL responses by in vivokilling assay. On day 0, mice received 50 mgi.v. of mock AC- or OVA/AC- toxoids and onday 7, they were i.v. injected with a mixture(1:1) of OVA (SIINFEKL) peptide-loadedCFSEhigh and unloaded CFSElow splenocytes.The number of CFSE-positive cells remainingin the spleen after 20 h was determined byFACS analysis, as documented for onerepresentative in vivo killing assay in theupper panel assembly of plots, wherepercentages of cells in the gates areindicated. The lower panel shows pooledresults of in vivo killing assays for threeindependent experiments. Statisticalsignificance was determined by the Student’st-test (P = 0.75 for OVA/AC- versusOVA/E570Q+K860R/AC-).
A
B
CyaA (µg/ml)
3H
incorp
ora
tion (
cpm
)
-
% o
fin
viv
o kill
ing
AC
-
OVA
/AC
-
OVA
/E570Q
+K
860R
/AC
0.0018 0.018 0.18 1.8 180
0
250000
500000
750000
1000000
1250000
0
10
20
30
40
50
AC- OVA/AC-OVA/E570Q+K860R/AC-
100
101
102
103
104
0
50
100
150
# C
ells
53 47
100
101
102
103
104
CFSE
0
50
100
150
200 71 29
100
101
102
103
104
0
50
100
150
200 67 33
AC -
OVA/AC-
OVA/E570Q+K860R/AC -
OVA/E570Q/AC -
OVA/K860R/AC -
OVA/E570K+E581P/AC -
Adenylate cyclase translocation: avoiding the pore 1557
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562
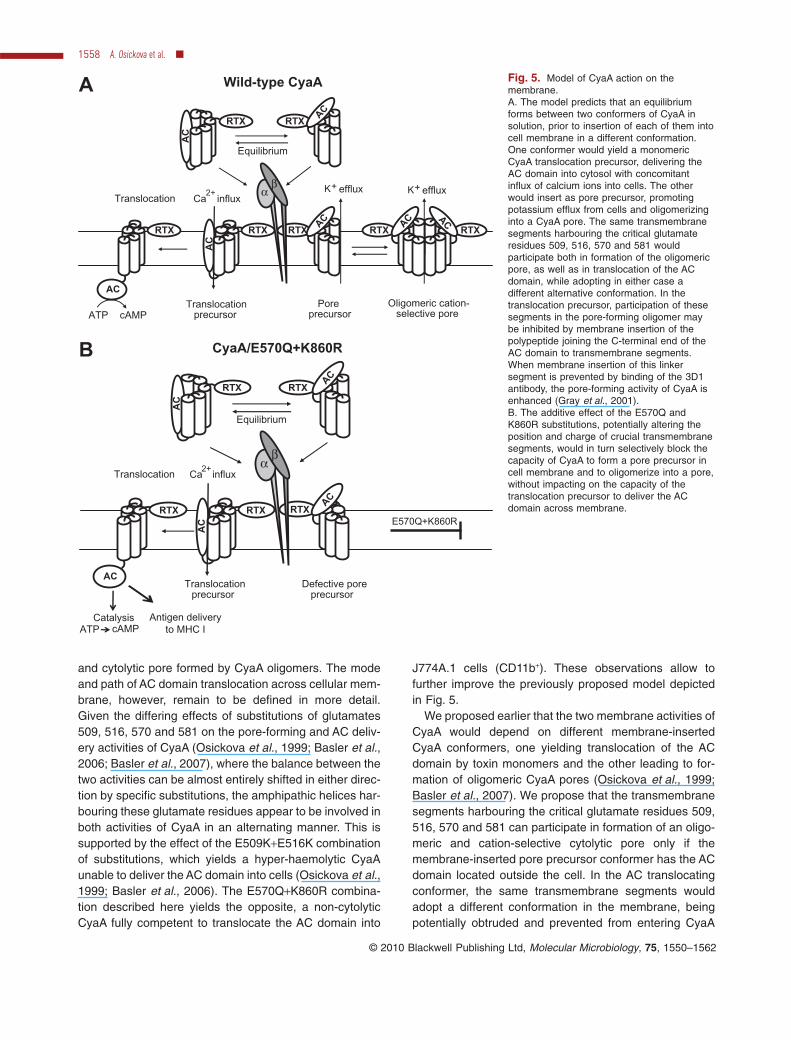
and cytolytic pore formed by CyaA oligomers. The modeand path of AC domain translocation across cellular mem-brane, however, remain to be defined in more detail.Given the differing effects of substitutions of glutamates509, 516, 570 and 581 on the pore-forming and AC deliv-ery activities of CyaA (Osickova et al., 1999; Basler et al.,2006; Basler et al., 2007), where the balance between thetwo activities can be almost entirely shifted in either direc-tion by specific substitutions, the amphipathic helices har-bouring these glutamate residues appear to be involved inboth activities of CyaA in an alternating manner. This issupported by the effect of the E509K+E516K combinationof substitutions, which yields a hyper-haemolytic CyaAunable to deliver the AC domain into cells (Osickova et al.,1999; Basler et al., 2006). The E570Q+K860R combina-tion described here yields the opposite, a non-cytolyticCyaA fully competent to translocate the AC domain into
J774A.1 cells (CD11b+). These observations allow tofurther improve the previously proposed model depictedin Fig. 5.
We proposed earlier that the two membrane activities ofCyaA would depend on different membrane-insertedCyaA conformers, one yielding translocation of the ACdomain by toxin monomers and the other leading to for-mation of oligomeric CyaA pores (Osickova et al., 1999;Basler et al., 2007). We propose that the transmembranesegments harbouring the critical glutamate residues 509,516, 570 and 581 can participate in formation of an oligo-meric and cation-selective cytolytic pore only if themembrane-inserted pore precursor conformer has the ACdomain located outside the cell. In the AC translocatingconformer, the same transmembrane segments wouldadopt a different conformation in the membrane, beingpotentially obtruded and prevented from entering CyaA
Fig. 5. Model of CyaA action on themembrane.A. The model predicts that an equilibriumforms between two conformers of CyaA insolution, prior to insertion of each of them intocell membrane in a different conformation.One conformer would yield a monomericCyaA translocation precursor, delivering theAC domain into cytosol with concomitantinflux of calcium ions into cells. The otherwould insert as pore precursor, promotingpotassium efflux from cells and oligomerizinginto a CyaA pore. The same transmembranesegments harbouring the critical glutamateresidues 509, 516, 570 and 581 wouldparticipate both in formation of the oligomericpore, as well as in translocation of the ACdomain, while adopting in either case adifferent alternative conformation. In thetranslocation precursor, participation of thesesegments in the pore-forming oligomer maybe inhibited by membrane insertion of thepolypeptide joining the C-terminal end of theAC domain to transmembrane segments.When membrane insertion of this linkersegment is prevented by binding of the 3D1antibody, the pore-forming activity of CyaA isenhanced (Gray et al., 2001).B. The additive effect of the E570Q andK860R substitutions, potentially altering theposition and charge of crucial transmembranesegments, would in turn selectively block thecapacity of CyaA to form a pore precursor incell membrane and to oligomerize into a pore,without impacting on the capacity of thetranslocation precursor to deliver the ACdomain across membrane.
ATP cAMPTranslocation
precursorPore
precursorOligomeric cation-
selective pore
Wild-type CyaAA
Equilibrium
Translocation
Translocationprecursor
Defective poreprecursor
CyaA/E570Q+K860RB
E570Q+K860R
cAMPATPCatalysis Antigen delivery
to MHC I
Ca2+
influx
RTX AC
RTX
AC
RTX AC
RTX
AC
RTX AC
αβ
RTX
AC
RTX
AC
K+
Equilibrium
Translocation Ca2+
influx
RTX
AC
RTX
AC
RTX
AC
αβ
RTX
AC
RTX
AC
efflux K+ efflux
1558 A. Osickova et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

oligomers by the polypeptide linking the C-terminal end ofthe AC domain to transmembrane segments. Support forthis interpretation can be deduced from results obtainedby Gray and co-workers (Gray et al., 2001). Theseauthors showed that deletion of the AC domain togetherwith the segment linking it to the pore-forming domain (upto residue 489), or binding of the 3D1 antibody that blocksmembrane translocation of the terminal AC domainsegment located between residues 373 and 399, impor-tantly enhances the pore-forming (haemolytic) activity ofthe toxin. This is likely to be due to imposing a conforma-tion on the transmembrane segments of CyaA that isfavourable for formation of the oligomeric pores.
It remains, however, to be defined what CyaA segmentsoutside the pore-forming domain are involved in ACdomain translocation across membrane. Given therequirement for structural integrity of CyaA (Iwaki et al.,1995), its large RTX repeat domain (residues 1006–1706)is likely to be taking part in AC domain translocation intocells. It would be sized enough (700 residues) to form ahydrophilic translocation interface within cellular mem-brane that might allow passage of an unfolded AC domainacross the membrane without a concomitant formation ofa real cell-permeabilizing pore.
CyaA was, indeed, previously shown to promote forma-tion of inverted non-lamellar (inverted hexagonal phase)lipid structures (Martin et al., 2004). These might poten-tially take part in formation of a well-sealed protein-lipidinterface, thus allowing translocation of the AC domainacross membrane in the absence of cell permeabilization.Formation of non-lamellar lipid structures is favoured incholesterol-enriched lipid rafts and CyaA was, indeed,recently found to mobilize into rafts in complex with itsreceptor CD11b/CD18. Moreover, we could recently showthat AC domain translocation across membrane is accom-plished only upon relocation of CyaA into rafts (L. Bumba,J. Masin, R. Fiser and P. Sebo, submitted). Intriguingly,translocation of the catalytic subunit of diphtheria toxin(DT) across the cell cytoplasmic membrane was also pre-viously found to occur without detectable cell permeabili-zation, when DT was pulsed into cells by low pH uponbinding to a truncated GPI-anchored DT receptor (Lan-zrein et al., 1996). The authors did not examine whetherthe GPI-anchored DT receptor localized into lipid rafts, butthis is highly likely. It is, hence, tempting to speculate thatthe specific lipid composition of the raft membrane maysupport translocation of different protein toxins into targetcells without the need for formation of a true protein con-ducting pore permeabilizing the cell.
Last but not least, a practical discovery reported hereinis that the CyaA/E570Q+K860R/AC- toxoid, with the muchreduced cell-permeabilizing (cytolytic) activity, remainsfully active in antigen delivery into CD11b+ APCs. This is ofimportance in the light of its potential use as a tool for
delivery of tumor-specific antigens in second generationof CyaA/AC--derived vaccines for immunotherapy ofcancer.
Experimental procedures
Construction, production and purification ofCyaA proteins
The mutations yielding CyaA/AC-, CyaA/OVA, CyaA/E570Qand CyaA/K860R constructs were previously described(Basar et al., 1999; Osicka et al., 2000; Basler et al., 2007),and were introduced into CyaA/OVA/AC- individually or incombination. The CyaA-derived proteins were produced in E.coli XL-1 Blue and were purified close to homogeneity aspreviously described (Karimova et al., 1998b). During thehydrophobic chromatography, the Phenyl Sepharose resinwith bound toxin was repeatedly washed with 60% isopro-panol (Franken et al., 2000) to reduce the endotoxin contentof CyaA samples below 100 IU mg-1 of protein, as determinedby the LAL assay QCl-1000 (Cambrex, Charles City, USA).
Cell binding and haemolytic activities onsheep erythrocytes
Aliquots of 5 ¥ 108 washed sheep erythrocytes in 50 mM TrispH 7.4, 150 mM NaCl and 2 mM CaCl2 (TNC buffer) wereincubated at 37°C with 5 mg ml-1 of CyaA proteins and cellbinding, cell-invasive AC and haemolytic activities of CyaAwere determined as described in detail previously (Iwakiet al., 1995; Basler et al., 2007). Significance of differences inactivity values was analysed using a one-way analysis ofvariance (ANOVA) with Bonferroni post-test (SigmaStat v. 3.11,Systat, San Jose, CA, USA).
Binding, cAMP elevation and cytolytic capacity of CyaAon J774A.1 cells
J774.A1 murine monocytes/macrophages (ATCC, numberTIB-67) were cultured at 37°C in a humidified air/CO2 (19:1)atmosphere in RPMI medium supplemented with 10% (v/v)heat-inactivated fetal bovine serum, penicillin (100 IU ml-1),streptomycin (100 mg ml-1) and amphotericin B (250 ng ml-1).Prior to assays, RPMI was replaced with Dulbecco’s modifiedEagle’s medium (DMEM) (1.9 mM Ca2+) without FCS and thecells were allowed to rest in DMEM for 1 h at 37°C in ahumidified 5% CO2 atmosphere (Basler et al., 2006). J774A.1cells (106) were incubated in DMEM with CyaA variants for30 min at 4°C, prior to removal of unbound toxin by threewashes in DMEM. Cells were lysed with 0.1% Triton X-100for determination of cell-bound AC enzyme activity. For intra-cellular cAMP assays, 105 cells were incubated with CyaA for30 min in DMEM with 100 mM IBMX (3-isobutyl-1-methylxanthin), the reaction was stopped by addition of 0.2%Tween-20 in 100 mM HCl, samples were boiled for 15 min at100°C, neutralized by addition of 150 mM unbuffered imida-zol and cAMP was measured as described (Karimova et al.,1998b). Toxin-induced lysis of J774A.1 cells was determinedusing the CytoTox 96 kit assay (Promega, Madison, USA) as
Adenylate cyclase translocation: avoiding the pore 1559
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

the amount of lactate dehydrogenase released from 105 cellsin 3 h of incubation at 37°C in DMEM without serum asdescribed (Basler et al., 2006). Significance of differences inactivity values was analysed as above.
Patch clamp measurements of cell permeabilization
Whole-cell patch-clamp measurements were performed onsingle J774A.1 cells bathing in modified Hanks’ balanced saltsolution (HBSS: 140 mM NaCl, 5 mM KCl, 2 mM CaCl2,3 mM MgCl2, 10 mM HEPES-Na, 50 mM glucose; pH 7.4).Fire-polished glass micropipettes with an outer diameter ofabout 3 mm were filled with the solution of 125 mM potassiumgluconate, 15 mM KCl, 0.5 mM CaCl2, 1 mM MgCl2, 5 mMEGTA, 10 mM HEPES-KOH pH 7.2. The resulting resis-tances of the microelectrodes were 3–5 MW. Cells wereclamped at -40 mV, the data were filtered at 1 kHz and digi-tized at 2 kHz using Axopatch 200A amplifier, Digidata 1320Adigitizer and PClamp-9 software package (Axon Instruments,Foster City, CA, USA).
Determination of decrease of cytosolic K+ concentration
Cells grown on glass coverslips were washed in HBSS andloaded with 9.5 mM PBFI acetoxymethyl ester (PBFI/AM,Molecular Probes, Eugene, OR, USA) for 30 min at 25°C inthe presence of 0.05% (w/w) Pluronic F-127 (Sigma, St.Louis, MO), in the dark. Ratiometric measurement was per-formed at 25°C using a FluoroMax-3 spectrofluorimeter(Jobin Yvon Horriba, France) equipped with DataMaxsoftware. Fluorescence intensity of PBFI (excitation wave-length 340, emission wavelengths 450 and 510 nm) wasrecorded every 15 s and the integration time for each wave-length was 3 s. The ratio of 450/510 nm wavelengths isshown. The observed area of coverslip mounted in the 1 cmcuvette was about 10 mm2, corresponding to approximately104 cells. Calibration experiments were performed in 50 mMHEPES, pH 7.2, with varying concentrations of PotassiumAcetate (10, 30, 60 or 140 mM) and Sodium Acetate (135,115, 85 or 5 mM), respectively, on cells permeabilized for30 min with 3 mM valinomycin or nigericin. Final intensity ratio(450/510 nm) is shown on right vertical axis of the plots.
Mice and cell lines
Animal studies were approved by the Institut Pasteur SafetyCommittee and were conducted in accordance with French(Law n°88–1138 of 20 Decembre 1988, application decreen°90–872 of 27 September 1990) and European guidelines.Female C57BL/6 mice obtained from Charles River Labora-tories were kept under specific pathogen-free conditions andmanipulated according to institutional guidelines. CTLL-2cells were obtained from ATCC. B3Z, a CD8+ specific T cellhybridoma specific for the Kb restricted OVA (SIINFEKL)epitope, was provided by N. Shastri (University of California,Berkeley) and maintained in the presence of 1 mg ml-1 G418and 400 mg ml-1 hygromycin B in complete RPMI 1640medium (Invitrogen Life Technologies, Carlsbad, USA) with10% heat-inactivated FCS, 100 U ml-1 penicillin, 100 mg ml-1
streptomycin and 5 ¥ 10-5 M 2-ME.
Antigen presentation studies
Bone Marrow Dendritic Cells (BMDC, 3 ¥ 105 per well) usedas APCs were incubated in the presence of various concen-trations (0–60 nM) of the recombinant CyaA/OVA/AC- toxoidcarrying the OVA (SIINFEKL) epitope, or with mock CyaA/AC-, and were co-cultured for 24 h with B3Z T cells (1 ¥ 105
per well), which selectively recognize the OVA SIINFEKL/H-2Kb MHC class I complexes. After 18 h of culture, superna-tants were frozen for at least 2 h at -80°C. The amount of IL-2produced by the stimulated B3Z cells was then determined bythe CTLL proliferation method. Briefly, 104 cells of the IL-2-dependent CTLL line per well were cultured with 100 ml ofsupernatant in 200 ml of final volume. Twenty-four hours later,[3H]-thymidine (50 mCi per well) was added and cells wereharvested 6 h later with an automated cell harvester. Incor-porated [3H]-thymidine was detected by scintillation counting.Each point was done in duplicate and the experiment wasrepeated five times. Results are expressed in Dcpm of incor-porated [3H]-thymidine (cpm in the presence of toxoid - cpmin the absence of toxoid).
In vivo killing assay
For CTL priming, mice were immunized by i.v. injection with50 mg of recombinant CyaA/OVA/AC- carrying the OVA (SIIN-FEKL) epitope or with mock CyaA/AC-. Seven days afterimmunization, naive syngenic splenocytes were pulsed withOVA (SIINFEKL) peptide (10 mg ml-1, 30 min, 37°C), washedextensively and labelled with a high concentration (1.25 mM)of carboxyfluorescein succinimidyl ester (CFSE; MolecularProbes, Eugene, OR, USA). The non-pulsed control popula-tion was labelled with a low concentration (0.125 mM) ofCFSE. CFSEhigh- and CFSElow-labelled cells were mixed in a1:1 ratio (5 ¥ 106 cells of each population) and injected i.v.into mice. Spleen cells were collected 20 h after, washed andresuspended in FACS buffer (PBS supplemented with 1%BSA and 0.1% NaN3). The number of CFSE-positive cellsremaining in the spleen after 20 h was determined by FACS.The percentage of specific lysis was calculated as follows: %specific lysis = 100 - [100 ¥ (% CFSEhigh immunized mice/%CFSElow immunized mice)/(% CFSEhigh naive mouse/%CFSElow naive mouse)].
Acknowledgements
This work was supported by Research ConceptsAV0Z50200510 (P.S.) and AV0Z50110509 (J.K.), the LigueNationale Contre le Cancer (C.L., Equipe Labellisée) andgrants IAA500200914 and GA310/08/0447 (P.S.), 1M0506(J.M.) and 2B06161 (R.O.). The excellent technical help of H.Kubinova and S. Charvatova is gratefully acknowledged.
References
Basar, T., Havlicek, V., Bezouskova, S., Halada, P., Hackett,M., and Sebo, P. (1999) The conserved lysine 860 in theadditional fatty-acylation site of Bordetella pertussis ade-nylate cyclase is crucial for toxin function independently ofits acylation status. J Biol Chem 274: 10777–10783.
1560 A. Osickova et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

Basler, M., Masin, J., Osicka, R., and Sebo, P. (2006) Pore-forming and enzymatic activities of Bordetella pertussisadenylate cyclase toxin synergize in promoting lysis ofmonocytes. Infect Immun 74: 2207–2214.
Basler, M., Knapp, O., Masin, J., Fiser, R., Maier, E., Benz,R., et al. (2007) Segments crucial for membrane translo-cation and pore-forming activity of Bordetella adenylatecyclase toxin. J Biol Chem 282: 12419–12429.
Bellalou, J., Sakamoto, H., Ladant, D., Geoffroy, C., andUllmann, A. (1990) Deletions affecting hemolytic and toxinactivities of Bordetella pertussis adenylate cyclase. InfectImmun 58: 3242–3247.
Benz, R., Maier, E., Ladant, D., Ullmann, A., and Sebo, P.(1994) Adenylate cyclase toxin (CyaA) of Bordetellapertussis. Evidence for the formation of small ion-permeable channels and comparison with HlyA of Escheri-chia coli. J Biol Chem 269: 27231–27239.
Ehrmann, I.E., Gray, M.C., Gordon, V.M., Gray, L.S., andHewlett, E.L. (1991) Hemolytic activity of adenylate cyclasetoxin from Bordetella pertussis. FEBS Lett 278: 79–83.
Fayolle, C., Sebo, P., Ladant, D., Ullmann, A., and Leclerc, C.(1996) In vivo induction of CTL responses by recombinantadenylate cyclase of Bordetella pertussis carrying viralCD8+ T cell epitopes. J Immunol 156: 4697–4706.
Fiser, R., Masin, J., Basler, M., Krusek, J., Spulakova, V.,Konopasek, I., and Sebo, P. (2007) Third activity of Bor-detella adenylate cyclase (AC) toxin-hemolysin. Mem-brane translocation of AC domain polypeptide promotescalcium influx into CD11b+ monocytes independently ofthe catalytic and hemolytic activities. J Biol Chem 282:2808–2820.
Franken, K.L., Hiemstra, H.S., van Meijgaarden, K.E., Sub-ronto, Y., Hartigh, J., Ottenhoff, T.H., and Drijfhout, J.W.(2000) Purification of his-tagged proteins by immobilizedchelate affinity chromatography: the benefits from the useof organic solvent. Protein Expr Purif 18: 95–99.
Glaser, P., Sakamoto, H., Bellalou, J., Ullmann, A., andDanchin, A. (1988) Secretion of cyclolysin, the calmodulin-sensitive adenylate cyclase-haemolysin bifunctionalprotein of Bordetella pertussis. EMBO J 7: 3997–4004.
Gordon, V.M., Leppla, S.H., and Hewlett, E.L. (1988) Inhibi-tors of receptor-mediated endocytosis block the entry ofBacillus anthracis adenylate cyclase toxin but not that ofBordetella pertussis adenylate cyclase toxin. Infect Immun56: 1066–1069.
Gray, M., Szabo, G., Otero, A.S., Gray, L., and Hewlett, E.(1998) Distinct mechanisms for K+ efflux, intoxication, andhemolysis by Bordetella pertussis AC toxin. J Biol Chem273: 18260–18267.
Gray, M.C., Lee, S.J., Gray, L.S., Zaretzky, F.R., Otero, A.S.,Szabo, G., and Hewlett, E.L. (2001) Translocation-specificconformation of adenylate cyclase toxin from Bordetellapertussis inhibits toxin-mediated hemolysis. J Bacteriol183: 5904–5910.
Guermonprez, P., Ladant, D., Karimova, G., Ullmann, A., andLeclerc, C. (1999) Direct delivery of the Bordetella pertus-sis adenylate cyclase toxin to the MHC class I antigenpresentation pathway. J Immunol 162: 1910–1916.
Guermonprez, P., Khelef, N., Blouin, E., Rieu, P., Ricciardi-Castagnoli, P., Guiso, N., et al. (2001) The adenylatecyclase toxin of Bordetella pertussis binds to target cells
via the alpha(M)beta(2) integrin (CD11b/CD18). J Exp Med193: 1035–1044.
Hackett, M., Walker, C.B., Guo, L., Gray, M.C., Van Cuyk, S.,Ullmann, A., et al. (1995) Hemolytic, but not cell-invasiveactivity, of adenylate cyclase toxin is selectively affected bydifferential fatty-acylation in Escherichia coli. J Biol Chem270: 20250–20253.
Havlicek, V., Higgins, L., Chen, W., Halada, P., Sebo, P.,Sakamoto, H., and Hackett, M. (2001) Mass spectrometricanalysis of recombinant adenylate cyclase toxin from Bor-detella pertussis strain 18323/pHSP9. J Mass Spectrom36: 384–391.
Hewlett, E.L., Donato, G.M., and Gray, M.C. (2006) Macroph-age cytotoxicity produced by adenylate cyclase toxin fromBordetella pertussis: more than just making cyclic AMP!Mol Microbiol 59: 447–459.
Iwaki, M., Ullmann, A., and Sebo, P. (1995) Identification by invitro complementation of regions required for cell-invasiveactivity of Bordetella pertussis adenylate cyclase toxin. MolMicrobiol 17: 1015–1024.
Kamanova, J., Kofronova, O., Masin, J., Genth, H., Vojtova,J., Linhartova, I., et al. (2008) Adenylate cyclase toxin sub-verts phagocyte function by RhoA inhibition and unproduc-tive ruffling. J Immunol 181: 5587–5597.
Karimova, G., Fayolle, C., Gmira, S., Ullmann, A., Leclerc, C.,and Ladant, D. (1998a) Charge-dependent translocation ofBordetella pertussis adenylate cyclase toxin into eukaryoticcells: implication for the in vivo delivery of CD8(+) T cellepitopes into antigen-presenting cells. Proc Natl Acad SciUSA 95: 12532–12537.
Karimova, G., Pidoux, J., Ullmann, A., and Ladant, D.(1998b) A bacterial two-hybrid system based on a recon-stituted signal transduction pathway. Proc Natl Acad SciUSA 95: 5752–5756.
Khelef, N., Zychlinsky, A., and Guiso, N. (1993) Bordetellapertussis induces apoptosis in macrophages: role of ade-nylate cyclase-hemolysin. Infect Immun 61: 4064–4071.
Lanzrein, M., Sand, O., and Olsnes, S. (1996) GPI-anchoreddiphtheria toxin receptor allows membrane translocation ofthe toxin without detectable ion channel activity. EMBO J15: 725–734.
Martin, C., Requero, M.A., Masin, J., Konopasek, I., Goni,F.M., Sebo, P., and Ostolaza, H. (2004) Membrane restruc-turing by Bordetella pertussis adenylate cyclase toxin, amember of the RTX toxin family. J Bacteriol 186: 3760–3765.
Masin, J., Basler, M., Knapp, O., El-Azami-El-Idrissi, M.,Maier, E., Konopasek, I., et al. (2005) Acylation of lysine860 allows tight binding and cytotoxicity of Bordetella ade-nylate cyclase on CD11b-expressing cells. Biochemistry44: 12759–12766.
Morova, J., Osicka, R., Masin, J., and Sebo, P. (2008) RTXcytotoxins recognize {beta}2 integrin receptors throughN-linked oligosaccharides. Proc Natl Acad Sci USA 105:5355–5360.
Osicka, R., Osickova, A., Basar, T., Guermonprez, P., Rojas,M., Leclerc, C., and Sebo, P. (2000) Delivery of CD8(+)T-cell epitopes into major histocompatibility complex classI antigen presentation pathway by Bordetella pertussisadenylate cyclase: delineation of cell invasive structuresand permissive insertion sites. Infect Immun 68: 247–256.
Adenylate cyclase translocation: avoiding the pore 1561
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562

Osickova, A., Osicka, R., Maier, E., Benz, R., and Sebo, P.(1999) An amphipathic alpha-helix including glutamates509 and 516 is crucial for membrane translocation of ade-nylate cyclase toxin and modulates formation and cationselectivity of its membrane channels. J Biol Chem 274:37644–37650.
Paccani, S.R., Dal Molin, F., Benagiano, M., Ladant, D.,D’Elios, M.M., Montecucco, C., and Baldari, C.T. (2008)Suppression of T-lymphocyte activation and chemotaxis bythe adenylate cyclase toxin of Bordetella pertussis. InfectImmun 76: 2822–2832.
Rogel, A., and Hanski, E. (1992) Distinct steps in the pen-etration of adenylate cyclase toxin of Bordetella pertussisinto sheep erythrocytes. Translocation of the toxin acrossthe membrane. J Biol Chem 267: 22599–22605.
Rose, T., Sebo, P., Bellalou, J., and Ladant, D. (1995) Inter-action of calcium with Bordetella pertussis adenylatecyclase toxin. Characterization of multiple calcium-binding
sites and calcium-induced conformational changes. J BiolChem 270: 26370–26376.
Schlecht, G., Loucka, J., Najar, H., Sebo, P., and Leclerc, C.(2004) Antigen targeting to CD11b allows efficient presen-tation of CD4+ and CD8+ T cell epitopes and in vivo Th1-polarized T cell priming. J Immunol 173: 6089–6097.
Szabo, G., Gray, M.C., and Hewlett, E.L. (1994) Adenylatecyclase toxin from Bordetella pertussis produces ion con-ductance across artificial lipid bilayers in a calcium- andpolarity-dependent manner. J Biol Chem 269: 22496–22499.
Vojtova, J., Kamanova, J., and Sebo, P. (2006) Bordetellaadenylate cyclase toxin: a swift saboteur of host defense.Curr Opin Microbiol 9: 69–75.
Vojtova-Vodolanova, J., Basler, M., Osicka, R., Knapp, O.,Maier, E., Cerny, J., et al. (2009) Oligomerization isinvolved in pore formation by Bordetella adenylate cyclasetoxin. FASEB J 23: 2381–2843.
1562 A. Osickova et al. �
© 2010 Blackwell Publishing Ltd, Molecular Microbiology, 75, 1550–1562
Copyright © 2022 FDOKUMEN




















