
Acidic mist and nitrogen fertilization effects on growth, nitrate reductase activity, gas exchange,...
13
New Phytol. (1992), 121, 611-622 Acidic mist and nitrogen fertilization effects on growth, nitrate reductase activity, gas exchange, and frost hardiness of red spruce seedhngs BY S. J. L'HIRONDELLE^*, J. S.JACOB SO N^ AND J. P. LAS SO IE^ ^ Department of Natural Resources, Cornell University, Ithaca, NY 14853, USA ^ Boyce Thompson Institute, Cornell University, Ithaca, NY 14853, USA {Received 12 August 1991; accepted 9 April 1992) SUMMARY Acidic deposition may contribute to the decline of red spruce (Picea rubens Sarg.) in eastern North America; response to acidic mist may depend on mist content and plant nutrient status. We grew red spruce seedlings with high or low nitrogen fertilization to roots (other elements in constant proportion to N), exposed only shoots to pH 30, high sulphate (10:1, S:N) or high nitrate (1 :10, S:N) mist for 5 wk, then moved seedlings outdoors for cold acclimation. Effects of mist content were found until the third week of cold acclimation. Foliar necrosis was higher after exposure to sulphate than to nitrate mist and greater in older than new needles. Sulphate-misted seedlings had lower new-shoot biomass, shoot water content, and nitrate reductase activity, and higher foliar sulphur levels than nitrate-misted seedlings. They also had greater needle frost hardiness (estimated by electrolyte leakage) at the start of cold acclimation. Effects of fertilization were found throughout the experiment. High-N fertilization increased biomass, tissue N level, and shoot water content, and decreased foliar necrosis from mist. During cold acclimation, high-N seedlings had lower transpiration and photosynthetic rates and higher water use efficiency than low-N seedlings. High-N fertilization increased needle frost hardiness after 5 wk of acclimation and decreased visible needle injury after an outdoor frost. However, high-N seedlings had fewer needle primordia in terminal buds and more bud frost-damage than low-N seedlings. Fertilization, mist content, and their interaction strongly affected seedlings, showing that nutrient status can influence the response of red spruce to acidic deposition and frost. Key words: Acidic mist, frost hardiness, nitrate reductase, nitrogen fertilization, Picea rubens (red spruce). sition could be influenced by these anion ratios, INTRODUCTION ^.^j^ ^^^j^ ^^^^^^ reduced or mitigated by tree Acidic deposition has been suggested as a contributor physiological condition during exposure, to increased mortality and decreased growth of high There have been several recent reports on the elevation red spruce (Picea rubens Sarg.) (Pitelka & effects of acidic deposition on red spruce seedlings. Raynal, 1989; LeBlanc & Raynal, 1990). Red spruce Exposure to acidic mist or fog has altered epi- stands in eastern North America are exposed to wet cuticular wax structure on elongating needles (Percy, deposition containing considerable amounts of both Krause & Jensen, 1990), resulted in foliar uptake of sulphate and nitrate at pH values that are sometimes both sulphur (S) and nitrogen (N) from mist less than 3 (Kimball et al., 1988; Mohnen, 1988; (Bowden, Geballe & Bowden, 1989; Jacobson e^ a/., Mueller & Weatherford, 1988). There is often at 1989) and cation loss from foliage (Joslin, McDuffie least twice as much sulphate as nitrate in cloud water & Brewer, 1988), increased (Roberts & Cannon, and precipitation, with cloud water having much 1989) or decreased (Eamus, Leith & Fowler, 1989) higher levels of both (Kimball et al., 1988; Mohnen, seedling water potential, and decreased needle frost 1988). The response of red spruce to acidic depo- hardiness (Laurence ef a/., 1988; Fowler ef a/., 1989; Cape et al., 1991). Anion content of mist is important; red spruce seedlings were more injured * Current address and to whom correspondence should be ,^ sulphate than nitrate in acidic mist, especially addressed • British Columbia Ministry of Forests, Research Lab, \ '^ , t 1320 Glyn Road, Victoria B.C. V8W 3E7 Canada. when mist exposure was mtermittent (Jacobson et al. ^j ANP 121
Transcript of Acidic mist and nitrogen fertilization effects on growth, nitrate reductase activity, gas exchange,...

New Phytol. (1992), 121, 611-622
Acidic mist and nitrogen fertilizationeffects on growth, nitrate reductase activity,gas exchange, and frost hardiness of redspruce seedhngs
BY S. J. L ' H I R O N D E L L E ^ * , J. S.JACOB SO N AND J. P. LAS SO IE^
^ Department of Natural Resources, Cornell University, Ithaca, NY 14853, USA^ Boyce Thompson Institute, Cornell University, Ithaca, NY 14853, USA
{Received 12 August 1991; accepted 9 April 1992)
SUMMARY
Acidic deposition may contribute to the decline of red spruce (Picea rubens Sarg.) in eastern North America;response to acidic mist may depend on mist content and plant nutrient status. We grew red spruce seedlings withhigh or low nitrogen fertilization to roots (other elements in constant proportion to N), exposed only shoots topH 30, high sulphate (10:1, S:N) or high nitrate (1 :10, S:N) mist for 5 wk, then moved seedlings outdoors forcold acclimation. Effects of mist content were found until the third week of cold acclimation. Foliar necrosis washigher after exposure to sulphate than to nitrate mist and greater in older than new needles. Sulphate-mistedseedlings had lower new-shoot biomass, shoot water content, and nitrate reductase activity, and higher foliarsulphur levels than nitrate-misted seedlings. They also had greater needle frost hardiness (estimated by electrolyteleakage) at the start of cold acclimation. Effects of fertilization were found throughout the experiment. High-Nfertilization increased biomass, tissue N level, and shoot water content, and decreased foliar necrosis from mist.During cold acclimation, high-N seedlings had lower transpiration and photosynthetic rates and higher water useefficiency than low-N seedlings. High-N fertilization increased needle frost hardiness after 5 wk of acclimation anddecreased visible needle injury after an outdoor frost. However, high-N seedlings had fewer needle primordia interminal buds and more bud frost-damage than low-N seedlings. Fertilization, mist content, and their interactionstrongly affected seedlings, showing that nutrient status can influence the response of red spruce to acidicdeposition and frost.
Key words: Acidic mist, frost hardiness, nitrate reductase, nitrogen fertilization, Picea rubens (red spruce).
sition could be influenced by these anion ratios,INTRODUCTION ^ . ^ j ^ ^ ^ ^ j ^ ^^^^^^ reduced or mitigated by tree
Acidic deposition has been suggested as a contributor physiological condition during exposure,to increased mortality and decreased growth of high There have been several recent reports on theelevation red spruce (Picea rubens Sarg.) (Pitelka & effects of acidic deposition on red spruce seedlings.Raynal, 1989; LeBlanc & Raynal, 1990). Red spruce Exposure to acidic mist or fog has altered epi-stands in eastern North America are exposed to wet cuticular wax structure on elongating needles (Percy,deposition containing considerable amounts of both Krause & Jensen, 1990), resulted in foliar uptake ofsulphate and nitrate at pH values that are sometimes both sulphur (S) and nitrogen (N) from mistless than 3 (Kimball et al., 1988; Mohnen, 1988; (Bowden, Geballe & Bowden, 1989; Jacobson e a/.,Mueller & Weatherford, 1988). There is often at 1989) and cation loss from foliage (Joslin, McDuffieleast twice as much sulphate as nitrate in cloud water & Brewer, 1988), increased (Roberts & Cannon,and precipitation, with cloud water having much 1989) or decreased (Eamus, Leith & Fowler, 1989)higher levels of both (Kimball et al., 1988; Mohnen, seedling water potential, and decreased needle frost1988). The response of red spruce to acidic depo- hardiness (Laurence ef a/., 1988; Fowler ef a/., 1989;
Cape et al., 1991). Anion content of mist isimportant; red spruce seedlings were more injured
* Current address and to whom correspondence should be , sulphate than nitrate in acidic mist, especiallyaddressed • British Columbia Ministry of Forests, Research Lab, \ '^ , t1320 Glyn Road, Victoria B.C. V8W 3E7 Canada. when mist exposure was mtermi t ten t (Jacobson et al.
^j ANP 121

612 S.y. L'Hirondelle, jf. S. Jacobson and J. P. Lassoie
1990a, b). Both sulphate and ammonium ions inacidic mist decreased needle frost hardiness (relativeto controls) in seedlings, while nitrate ions did not(Cape et al., 1991).
These studies have exposed seedlings to nearambient levels of acidity and anion content, showingthe potential vulnerability of red spruce to currentwet deposition, but most have not quantified orcontrolled the nutrient status of the seedlings used.Interactions between mineral nutrient status anddamage from air pollutants such as sulphur dioxideand ozone are known to occur (Darrall, 1989), andsuch interactions should be considered when lookingat the effects of wet deposition on seedling growthand physiology.
Knowing that sulphate can be more damagingthan nitrate mist, and that nutrient status affectsplant response to stress, we chose to study the effectsof fertilization and mist anion content on growth andphysiology of red spruce seedlings. We used seed-lings in controlled growing conditions to increase theprobability of detecting effects and establishing causeand effect relationships; we did not intend to directlyextrapolate our results to mature trees in the field.We looked for differences in visible injury, biomass,tissue N and S concentrations, nitrate reductaseactivity, gas exchange, water content, frost hardiness,and needle primordia number during and afterexposure of seedlings to mist.
MATERIALS AND METHODS
Growing conditions
On 8 June 1988, 256 red spruce seedlings wereselected from a 19-wk-old set grown by the USForest Service (Burlington, VT) from a Nova Scotiaseed source. Seedlings were removed from paperpots, rinsed free of potting mix, and transplantedinto 150-cm^ Ray~Leach containers filled with base-acid-washed, medium-grade, quartz sand with alayer of base-acid-washed aquarium gravel on top.Seedlings were kept in a light room with 16 h days(under high intensity discharge sodium and multi-vapour lamps) with photosynthetic photon fluxdensity (PPFD, waveband 400-700 nm) at plantheight of 570 fimo\ n\~^ s~^ Average conditions were20/15 °C day/night temperature, 50% relative hu-midity, and 360/^1 CO. 1" The eight racks (blocks)of seedlings were randomly assigned positions on thelight room bench. From age 19 to 33 wk, seedlingswere in the light room, except during acidic mistapplication (age 24 5-29-5 wk), when they were in agreenhouse at night with temperatures from 20 to27 °C. Seedlings were moved to a south-facing out-door location in Ithaca, NY (42° 27' N, 76° 27' W)for cold acclimation (age 33-41 wk) in ambientconditions (mean maximum temperature 19 °C,mean minimum AS °C; photoperiod 123 h day atstart on 22 Sept., 10 h d at end on 8 Nov.).
Nitrogen fertilization
There were two levels of N fertilization: 100 (highN) or 20 mg N T (low N), with other elements heldin constant proportion to N (100 N:16 P:40 K:9 S:5 Ca:5 Mg:l Fe:0-2 B:04 Mn:0-03 Zn:0-03 Cu:0-007 Mo; L'Hirondelle 1990). Nutrientsolutions were applied to roots by drip irrigationthree times a day, with one spaghetti tube perseedling supplying a daily total of 75-90 ml ofsolution. Fresh solutions were prepared three timesa week. Solution pH, which was not adjusted, rangedfrom 4-9 to 5-5.
Acidic mist treatments
Exposure to acidic mist began on 18 July 1988 whenseedlings were 24-5 wk old. Each seedling had aplastic rainshield around the lower stem to excludemist from the roots. Seedlings were placed in thegreenhouse on one of eight 1-m diameter rotating(3 rpm) turntables, each enclosed in a plastic-walledchamber 120 X 105 x 105 cm (Jacobson e< a/., 19906).There were four turntables for each of the two misttypes. Acidic solution flowed through a solenoidvalve and plastic tubing to a spinning disk mistnozzle (Mini-ULVA, Micron Corp.) 75 cm aboveeach turntable. Mist, with a droplet diameter ofabout 20 fim, was applied at a rate of 1 mm h~ (4 1per turntable per event). There were five 1-h-longmist applications alternating with five 2-h-long dryperiods (total of 15 h = 1 mist event) during whichfans circulated air at about 100 cm s" to increaseevaporation of droplets from foliage. Mist appli-cation began at 17.00 h; seedlings were returned tothe light room at 08.00 h. From 18 July to 19 Augustthere were 25 mist events, for a total of 125 h of mistapplication. Assignment of treatment and rack toturntable was randomized daily.
Two types of pH 3-0 acidic mist with NOg andSO4 were used. The high nitrate mist (NO3:SO4 =10:1 by weight) contained 54900//gl"^ (886/tM) ofNO3 and 5470/igl"' (57/IM) of SO^ in deionizedwater. The high sulphate mist (NO3: SO4 = 1 :10 byweight) contained 4460 /ig T (72 //M) of NO3 and44500/^gT^ (464/^M) of SO4 in deionized water.The pH of solutions was always within +0-1 unit ofpH 3-0.
Visible injury rating
Each seedling (48 or 56 per treatment) was rated forpercent of needles showing necrotic lesions (i.e.incidence of foliar injury) with the following scale: 0= 0%, l = l e s s than 10%, 2 = 10-25%, 3 =25-50%, 4 = 50-75 % , 5 = 75-90%, 6 = more than90%, and 7 = 100% (Jacobson et al, 19906). Oneobserver perfornned all ratings.

Acidic mist and fertilization effects on red spruce 613
025 15
Number of mist events25
Figure 1. Incidence of visible injury to red spruce needlesafter exposure to pH 3-0 mist. Seedlings were 28 wk oldafter 15 events, 30 wk old after 25 events. New needles =current growth; Old = 6 months old. NO3 mist = 10:1,NO3.SO, by weight; SO, mist = 1.10, NOjiSO,.Hi(gh N) = 100 mg N 1' , Lo(w N) = 20 mg N 1 ' in nu-trient solution. • , Hi N, NO3 mist; H, Hi N, SO^ mist;0 , LoN, NO3 mist; D, Lo N, SO^ mist. Bars aremeans + 1 SE of 56 or 48 seedlings.
Biomass and physiology measurements
There were eight destructive harvests between July(25 wk old) and November (41 wk old) 1988: onebefore the mist exposure period, two during mistexposure, and five after misting ended. Severalvariables were measured each time on one or twoseedlings per treatment from each block (eight blocksfrom week 25 to 32, four blocks from week 33 to 41).Before harvest, seedlings were sprayed with de-ionized water to remove surface deposits. Shootswere severed at the root collar, weighed for freshweight (f. wt), and separated into older (first flush,older than 6 months) and new (second Hush, currentneedles) growth. Roots were washed in tap water,rinsed in deionized water, and weighed for f. wt. Dryweight (d. wt) was recorded after oven drying for atleast 48 h at 80 °C. Shoot (SWC) and root (RWC)water content were calculated from the entire abo\'e-or below-ground portions from:
SWC or RWC = (f. w t - d . wt) x 100/f. wt,
with f. wt and d. wt in g.Total organic N in dry tissue was determined by a
micro-Kjeldahl procedure. Samples were digested insulphuric acid-hydrogen peroxide (Thomas, Sheard& Moyer, 1967), and colour was developed withsalicylate-dichloroiso-cyanurate reagent (Crooke &Simpson 1971). Samples less than 0-2 g d. wt weredigested whole; larger samples were ground in liquidnitrogen. Total sulphur in dry samples was measuredby inductively-coupled argon plasma atomicemission spectroscopy (Isaac & Johnson, 1985)after digestion of tissue in nitric-perchloric acid.
Nitrate reductase activity (NRA) was assayed invivo with a method based on that used by Norby,Weerasuriya & Hanson (1989). Twenty non-necroticneedles (about 50 mg f. wt) or about 15 root tip
sections (about 200 mg f. wt) were weighed. Needles(cut into four sections) or roots (cut into 4- to 5-mmlengths) were placed in 14-ml polyethylene tubesand 2-4 ml of potassium phosphate buffer (pH 8,with 30 jUg chloramphenicol Tn\~\ 0-5% v/v n-pro-panol, and 0-02 % w/w Tween 80) and 0-6 ml of 0-1 MKNO3 were added and mixed. Samples were evacu-ated with a vacuum pump for 5 min to infiltratetissues with solution. Tubes were incubated for atleast 2 h at about 20 °C in a dark box on a rotaryshaker. After incubation, 2 ml of 1 % (w/v) sulpha-nilamide in 2 M HCl was added to 1 ml aliquotsof samples or standard solutions in 10-ml test tubes.After 5 min, 2 ml of 0-02 % (w/v) iV-(l-napthyl)-ethylenediamine dihydrochloride was added andmixed. Absorbance at 540 nm was read with aspectrophotometer. Tissue was dried at 80 °C for48 h and weighed. Nitrite concentration in testsamples was calculated from a standard curve ofsamples with 2 7-30 nmol NaNO.^ m r \ giving NRAas nmol NOj g d. wt~' h~\
Net CO2 assimilation, transpiration, and darkrespiration rates were measured with a calibrated Li-Cor LI-6200 portable gas exchange system in thelight room. Leader or top lateral shoots were sealedin a 250 cm^ transparent chamber. Average condi-tions for measurements ranged from 500-630//mol m"^ s" of PPFD, 19-23 °C, 370-410/^1
I , and 40-55 "o relative humidity. Beforerespiration measurements, seedlings were left in thedark for at least 20 min. Water-use efficiency wascalculated as the ratio of photosynthetic COg uptake{jumo\ m"^ s~') to water loss by transpiration(mmol m~' s~^).
Total needle surface area was determined with theglass bead technique (Thompson & Leyton 1971).Area was calculated from bead weight after dippingneedles or shoots (or wires of known surface area) inrubber cement mixed with chloroform (1:7 byweight) then in 0-11 mm diameter glass beads. Areawas regressed on needle length, needle d. wt, orshoot volume determined by Archimedes principle;the resulting regression equations (R^ = 0-91-0-94)were used to predict areas for the remaining samples.
The number of needle primordia was determinedfor excised leader buds. Bud scales were removedunder a dissecting microscope; the number ofcolumns in a longitudinal half of the bud wasmultiplied by the average number of rows in threeadjacent columns of primordia to give the totalnumber of needle primordia in a half bud.
Frost hardiness was estimated from the relativeconductivity of diffusates of needles exposed tosubfreezing temperatures, with a modification of theelectrolyte leakage technique described by Colombo,Webb & Glerum (1984). Six sets (one for eachtemperature) of three fully-expanded needles wereexcised from each leader shoot, placed in six 7-mlpolyethylene scintillation vials, and capped. Racks of
41..2

614 S. J. L'Hirondelle, Jf. S. Jacobson and J. P. Lassoie
Table 1. Probability values for injury and biomass variables for red spruceseedlings grown at two levels of Nfertilization and exposed to pH 3-0 nitrateor sulphate acidic mist
Variable (seedlings/trt)*
New-needle injury (56)• (48)
Older-needle (56)injury (48)
New shoot d. wt (8)(8)(4)(4)(4)(4)(4)
Root d. wt (8)(8)(8)(4)(4)(4)(4)(4)
Shoot/root ratio (8)(8)(4)(4)(4)(4)(4)
Age (wk)
2830
2830
28-5303435-53739-541
2528-5303435-53739-541
28-5303435-53739-541
P values from
N fertilizer
000000000000
0-000000-08000000000000
0-850-390000-360-920-320-540-41
0 0 00 0 00-080 0 00 0 50-14001
ANOVA
Mist anion
000000000000
0-130 0 00-580-230-460-610-26
N x M
0-280-61
0 0 00 0 0
0-170-980-260 0 00-080-920-57
Not misted0-560-740-190-260-170-920-15
0-760-120-010-110-670-400-25
0-750 0 20-120-590-780-400-610-190-200-170-930-770-400-64
There were 21 (weeks 25-32) or 9 (weeks 33-41) degrees of freedom for error.Analyses were run on block means (see methods).
* Seedlings/trt = number of seedlings per treatment.
vials (including blanks without needles) were put ina 4 °C cold room (controls) or in a programmablefreezer at 4 °C. Freezer temperature was lowered nomore than 4 °C h ' to the first set point and held for1 h, then the first rack was removed. This continuedfor a total of five subfreezing temperatures. Sampleswere thawed in insulated boxes at 4 °C for at least2 h. After addition of 3 ml of 0-02 % Triton-X(surfactant) in 0-01 M H3BO3, vials were shaken for19 h at about 20 °C. Electrical conductivity ofsolutions was measured at about 20 °C with acalibrated conductivity meter after shaking, andafter autoclaving at 121 °C for 40 min. Relativeconductivity (RC) was calculated from:
RC = (Ci-Bi)xl00/(C.,-B2),
where Cj and Cj are conductivity of the needlediffusate before and after autoclaving, respectively,and Bj and Bg are mean conductivity of blanks beforeand after autoclaving, respectively.
Visible frost damage was recorded after a frostoutdoors ( —5 °C on 31 October). The total numberof branches and the number with visible browning ofthe youngest needles were recorded for eight seed-
lings per treatment, and percent injury was calcu-
lated.
Experimental design and statistical tests
Seedlings were assigned to a randomized completeblocks design. For 19- to 32-wk-old seedlings therewere two N fertilization levels, two acidic mist types,and eight blocks; for 33- to 41-wk-old seedlingsthere were two N fertilization levels, two acidic misttypes, and four blocks. Block was the experimentalunit. Because daily randomization minimized mist-chamber effect, chamber was not included as afactor. Analysis of variance was used to calculate Fand P values, following the model:
Y,^, = / / . . . + a, + /^, + S, + {y.^),^ + e,,.,,
where / / . . . is the grand mean, ol.^ is the fixed effect ofN fertilization, / ^ is the fixed effect of acidic mist, 8^is the random effect of block, e j, is the random errorterm, and z = 1, 2;;" = 1, 2; /e = 1, 2,... 4 or 8. Whenmore than one seedling was sampled per block,analyses were run on block means. There were 21(19- to 32-wk-old) or 9 (33- to 41-wk-old) degrees of
Acidic mist and fertilization effects on red spruce 615
freedom for error. Scores for visible injury fromacidic mist were converted to class midpoints(percent) before analysis. Analyses of percent datawere run on both transformed (arcsin square root)and untransformed data; because there were nodifferences in interpretation, untransformed resultsare presented.
RESULTS
Visible injury from mist
Both fertilization and mist content affected visibleneedle injury with different patterns for older andnew needles (Fig. 1, Table 1). Sulphate mist causedtwo to eight times as much injury as nitrate mist. Forolder needles, injury ranged from 5 to 40%, withmuch more injury shown by low- than high-Nseedlings. There was a strong interaction betweenfertilizer and mist owing to the high injury in thelow-N, sulphate-mist treatment. For new needles,injury levels were all less than 7%, but there was
O)
25 30 35Seedling age (wk)
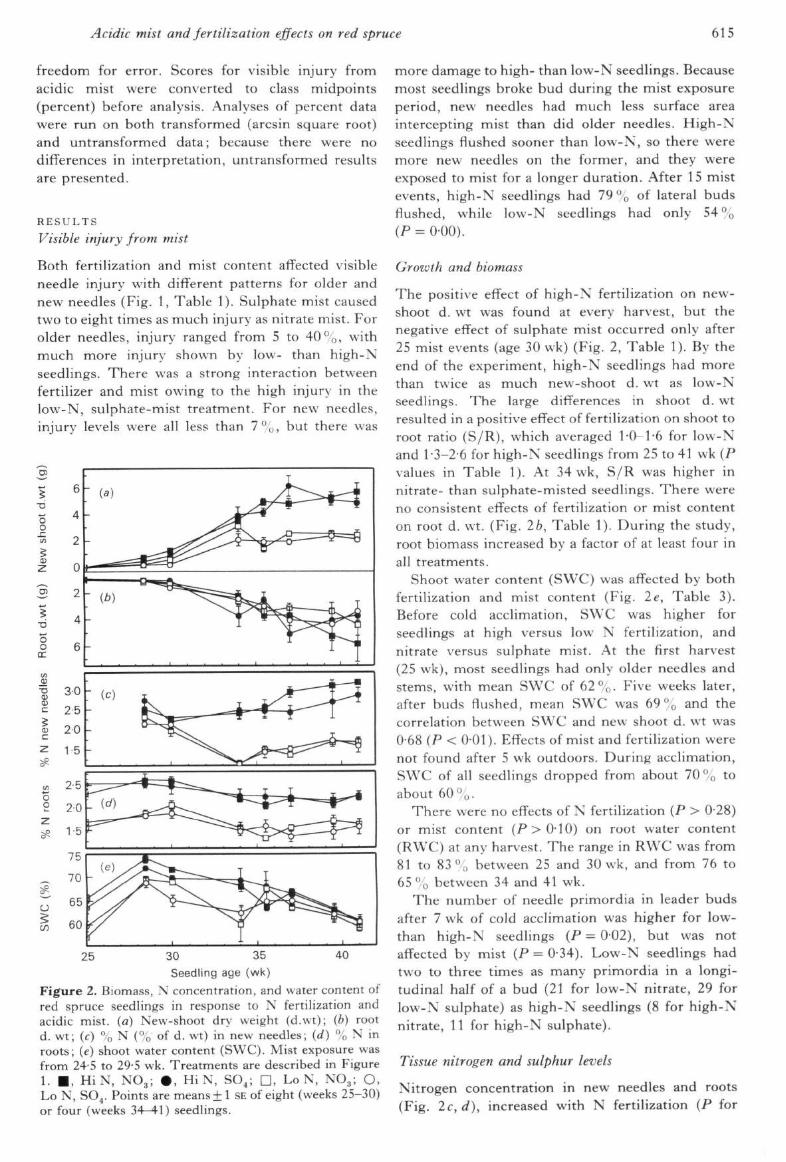
Figure 2. Biomass, N concentration, and water content ofred spruce seedlings in response to N fertilization andacidic mist, (a) New-shoot dry weight (d.wt); (b) rootd. wt; (c) % N (% of d. wt) in new needles; (d) % N inroots; (e) shoot water content (SWC). Mist exposure wasfrom 24-5 to 29-5 wk. Treatments are described in Figure1. • , HiN, NO3; • , HiN, SO,; D, Lo N, NO3; O,Lo N, SO4. Points are means + 1 SE of eight (weeks 25-30)or four (weeks 34-^1) seedlings.
more damage to high- than low-N seedlings. Becausemost seedlings broke bud during the mist exposureperiod, new needles had much less surface areaintercepting mist than did older needles. High-Nseedlings flushed sooner than low-N, so there weremore new needles on the former, and they wereexposed to mist for a longer duration. After 15 mistevents, high-N seedlings had 79% of lateral budsflushed, while low-N seedlings had only 54%(P = 0-00).
Growth and biomass
The positive effect of high-N fertilization on new-shoot d. wt was found at every harvest, but thenegative effect of sulphate mist occurred only after25 mist events (age 30 wk) (Fig. 2, Table 1), By theend of the experiment, high-N seedlings had morethan twice as much new-shoot d. wt as low-Nseedlings. The large differences in shoot d. wtresulted in a positive effect of fertilization on shoot toroot ratio (S/R), which averaged 1-0-1-6 for low-Nand 1 •3-2-6 for high-N seedlings from 25 to 41 vvk (Pvalues in Table 1). At 34 wk, S/R was higher innitrate- than sulphate-misted seedlings. There wereno consistent effects of fertilization or mist contenton root d. wt. (Fig. 2b, Table 1). During the study,root biomass increased by a factor of at least four inall treatments.
Shoot water content (SWC) was affected by bothfertilization and mist content (Fig. 2e, Table 3).Before cold acclimation, SWC was higher forseedlings at high versus low N fertilization, andnitrate versus sulphate mist. At the first harvest(25 wk), most seedlings had only older needles andstems, with mean SWC of 62%. Five weeks later,after buds flushed, mean SWC was 69 "0 and thecorrelation between SWC and new shoot d. wt was0-68 (P < 0-01). Effects of mist and fertilization werenot found after 5 wk outdoors. During acclimation,SWC of all seedlings dropped from about 70 % toabout 60 °0'
There were no effects of N fertilization (P > 028)or mist content (P>0-10) on root water content(RWC) at any harvest. The range in RWC was from81 to 83% between 25 and 30 wk, and from 76 to65 % between 34 and 41 wk.
The number of needle primordia in leader budsafter 7 wk of cold acclimation was higher for low-than high-N seedlings (P = 0-02), but was notaffected by mist (P = 0-34). Low-N seedlings hadtwo to three times as many primordia in a longi-tudinal half of a bud (21 for low-N nitrate, 29 forlow-N sulphate) as high-N seedlings (8 for high-Nnitrate, 11 for high-N sulphate).
Tissue nitrogen and sulphur levels
Nitrogen concentration in new needles and roots(Fig. 2c, d), increased with N fertilization (P for

616 S. y. UHirondelle, J. S. Jacobson and J. P. Lassoie
Table 2. Sulphur level and nitrate reductase activity {NRA) of red spruce seedlings grown at two levels of Nfertilization and exposed to pH 3-0 high nitrate or sulphate mist
Wk evt
Mean %S in tissue ANOVA P values
Hi N, NO3 Hi N, SO, Lo N, NO.. Lo N, SO, N Mist N x M
2528-53034
28-5303441
2528-5303441
0182525
18252525
018252525
0-3220-3410-2730-315
0-1440-1520-1800-212
0-1910-2070-1740-1410-152
0-3270-4120-5210-434
0-1740-2330-2190-167
0-1870-1960-1630-1420-170
Older needles
0-1390-2020-2240-153
New needles
0-1040-1260-0640-083
Roots
0-1240-1480-1310-1050-102
0-1370-3200-4170-296
0-1730-2090-0860-092
0-1280-1520-1340-0980-108
0 000000-090 00
0-060-16000000
0000000000000-00
0-95000000001
0000000-120-31
0-900-700-980-710-23
md0-31md0-78
mdmd0-640-15
0-750-450-810-640-57
Older needlesNew needlesRoots
Sulphur to nitrogen ratio at 30 wk of age (25 evt)
0-0990-0660-067
0-1710-1060-068
0-0920-0710-064
0-1790-1040-072
P valuesfor NRA
Older needlesNew needlesRoots
After
N
0-630-300-85
18 evt (28-5
Mist
0-060-12005
wk)
N x M
0-390-280-96
After
N
0-440-930-22
25 evt (30
Mist
0010-680-89
wk)
NxM
0-700-180-59
Treatments are described in Eigure 1. Wk, seedling age in weeks; evt, number of mist events; md, missing data(ANOVA run without interaction). One seedling was harvested per treatment per block. Erom weeks 25 to 32 there were8 blocks (21 d.f for error), from weeks 33-41 there were 4 blocks (9 d.f.)
effect of N < 0-03 for needles, < 0-01 for roots atseven harvests), but not with nitrate mist (P foreffect of mist > 010 for needles and roots). Differ-ences between high- and low-N seedlings in needleN concentration increased after 30 wk of age.
Foliar S concentration increased in response tosulphate mist. Sulphate-misted seedlings had abouttwice as much S in older needles and 1-5 times asmuch in new needles as nitrate-misted seedlings after25 mist events (Table 2). Low-N seedlings showedthe largest relative change, with a three-fold increasein older-needle S from the start to the end of mistexposure. After misting ended, foliar S decreased asmuch as 50 "o in sulphate-misted seedlings.Sulphur-to-nitrogen ratios were much higher forseedlings exposed to sulphate than nitrate mist, butwere almost identical for high- and low-N seedlings.High-N fertilization increased foliar S at mostharvests.
Root S concentration did not respond to mistcontent, but was affected by fertilization (Table 2).
High-N seedlings had about 50 °o rnore sulphur inthe roots than low-N seedlings for the duration ofthe experiment. Sulphur-to-nitrogen ratios in rootsof all treatments were similar to those in new needlesexposed to nitrate mist.
Nitrate reductase activity
Nitrogen fertilization did not affect nitrate reductaseactivity (NRA) in needles or roots, but nitrate mistincreased NRA as much as three-fold after 18 mistevents (Fig. 3, Table 2). The mist effect persisted inolder needles after seven more mist events. Highwithin-treatment variation (coefficients of variationup to 138%) made detection of treatment effectsdifficult.
Most nitrate reductase activity was in the roots forall treatments (Fig. 3). High-N seedlings, which hadat least twice as much new-needle d. wt as low-N,had an average of 30 % of total NRA in needles,while low-N seedlings had less than 20%. For low-

Acidic mist and fertilization effects on red spruce 617
18 25 18 25 18
Number of mist events25
Figure 3. Nitrate reductase activity (NRA) in new needles,old needles, and roots of red spruce seedlings in responseto N fertilization and acidic mist. Treatments are describedin Figure 1. Bars are means+ 1 SE of eight seedlings.
N seedlings, nitrate and sulphate-misted treatmentshad the same relative proportions of NRA amongneedle and root tissues; for high-N seedlings, the
Table 3. Probability values of physiological variablesmeasured on red spruce seedlings in response to Nfertilization and acidic mist
Variable Age (wk)
A 3435-53739-541
K 3435-53739-541
E 3435-53739-541
WUE 3435-53739-541
Shoot water 25content 28 5
303435-5373 9 541
P values from ANOVA
N fertilizer
0-500 8 00 0 10-070010030-680-760-890-74
0030020000040010010000000 9 30-22
0000000000020 8 90-090 5 20-46
Mist anion
0-350-950-120-890-62
0-280-920-400-560 2 9
0-130-360-290 8 90-48
0-420-180-230 8 90-87
0-900-100 0 00-520 7 60-710 9 20-29
N x M
0-500-770-920-430-52
0-730-270-300 0 00-53
0-540-740-800-640 0 5
0-730-590-680-480-23
0-180-440-190-870-200 4 40 8 30-56
A, net CO2 assimilation rate; R^, dark respiration rate,E, transpiration rate; WUE, water use efficiency. Numbersof seedlings and degrees of freedom are as shown in Figure2.
20
E 1-5du"0
E 10
05
_ 05
J" 04Eo 0-3E^ 02
q 7
o 6
u
Eo 4
34 36 38Seedling age (wk)
40 42
Figure 4. Gas exchange of red spruce seedlings in responseto N fertilization and acidic mist, (a) Net COj assimilationrate (A); (b) dark respiration rate (RJ; (c) transpirationrate (E); (d) water-use efficiency (WUE). Treatments andsymbols are described in Figures 1 and 2. Points aremeans + 1 SE of four seedlings.
proportion of NRA in needles was almost double innitrate versus sulphate-misted seedlings.
Gas exchange
Gas exchange, which was measured after the mistexposure period, was affected by N fertilization butnot mist content (Fig. 4, Table 3). When seedlingswere first moved outdoors, net COg assimilation rate(A) of new needles was similar in all treatments.During cold acclimation, A fell in all seedlings; in7 wk, rates dropped 55% for high-N and 35% forlow-N seedlings. For the last three harvests, A wasabout 30% lower in high- than in low-N seedlings.Dark respiration rate {R^) was initially higher inhigh- than low-N seedlings, but treatment effectswere not found after 34 wk.
Water use was affected by N fertilization but notmist content (Fig. 4c, d; Table 3). Transpirationrates {E) were up to 90% higher in low- than high-N seedlings; water use efficiency (WUE) showed theopposite pattern. Transpiration rates in all treat-tnents decreased as much as 38 % in the last 5 wk ofcold acclimation; WUE decreased as much as 48 % asa result of bigger decreases in A than in E.

618 . y. L'Hirondelle, J. S. Jacobson and J. P. Lassoie
100
80
60
40
20E 0(J 100DC
80
60
40
20
34 wk old(19 Sept.)
J39-5 wk old(26 Oct.)
s—<1
^ ^
1
1
37 wk o\6 P^(11 Oct.) Y
41 wk old(7 Nov.)
in o m o m« - CM CN
I I I
LO O LD O o in oCN CM 0 0
I I ITemperature (°C)
Figure 5. Relative conductivity (RC) of needle diffusatesof red spruce after exposure to low temperatures. Treat-ments and symbols are described in Figures 1 and 2.Outdoor cold acclimation started at 34 wk. Points aremeans+1 SE of four seedlings.
Table 4. Probability values for relative conductivity ofred spruce needles exposed to cold temperatures
P values from ANOVATemperature(°C) Age (v
+ 4- 4- 8
- 1 2- 1 6- 2 0
+ 4- 8
- 1 2- 1 6- 2 0- 2 4
+ 4- 1 2- 1 6- 2 0- 2 4- 2 8
+ 4- 1 0- 1 4- 1 8- 2 2- 2 6
34
37
39-5
41
N fertilizer
0150-540-960-530-27008
0290-400-870-24060046
0000-120-130160-15018
005003003004001004
Mist anion
0-480010040180-160040-380-740150070020040-570-36052065089081
0250-130-570-890-770-57
N x M
0330-460-940-470-640-59
0-670-670010-160-12020
0060-960-850980-89089
0-230-770-370-720-65038
One seedling per treatment was harvested from each offour blocks. There were 9 d.f. for error.
Visible injury from frost
A cold night ( — 5 °C) on 31 October resulted in thedeath of many terminal buds and visible injury to theyoungest needles. Needle injury ranged from 0-5 %
for high-N sulphate and 1"5 % for high-N nitrate to20 and 18% for low-N sulphate and nitrate treat-ments. The difference between N treatments waslarge, but the low number of degrees of freedom forerror gave a P value of 0'12 for the N effect. Mistcontent had no efTect on needle injury (P — 0-96).Although low-N seedlings had more needle damagethan high-N seedlings, they had less bud death.Terminal buds on five of eight high-N seedlingswere killed, while only one bud of eight low-Nseedlings died.
Electrolyte leakage
Relative conductivity (RC) (Fig. 5, Table 4) wasaffected by mist content at the start of coldacclimation, and later by N fertilization. RC washigher for nitrate- than sulphate-misted needles atseveral freezing temperatures on 19 Sept. and 11Oct. (34- to 37-wk-old). After 5 wk of acclimation,low-N needles had higher RC after freezing thanhigh-N needles (39- to 41-wk-old, 26 Oct.-7 Nov.).This agreed in direction with the frost-inducedvisible injury described above. However, low-Nneedles had about 3% RC at -10°C, but almost20% visible injury at —5 °C. The RC of unfrozenneedles was not affected by mist but increased withN fertilization.
Treatment effects on RC could result fromdifferences in conductivities before (Cj) or afterkilling (Cj), or both. There were mostly smalldifferences in C^ among treatments. The higher RCfor nitrate versus sulphate mist was not due to C^,because nitrate-misted needles had higher (34 wk) oralmost identical (37 wk) Cg compared with sulphate-misted needles. Likewise, differences in C^ betweenhigh- and low-N seedlings were too small to accountfor treatment effects on RC.
DISCUSSION
Effects before cold acclimation
Treatment with high-sulphate acidic mist resulted inmuch more injury and lower new-shoot biomassthan nitrate mist, as reported in other studies(Jacobson et al, 1990a, b; Cape et al, 1991). Thedifferent effects of these anions may be due todifferences in absorption (see below) and subsequentmetabolism. Sulphate and nitrate may influence thedamage caused by hydrogen ions according to theway in which the two anions are metabolized.Sulphur requirements are lower than for nitrogen, sosulphate from wet deposition may be stored ininorganic form, while nitrate is likely incorporatedimmediately into organic compounds. Hydrogenions are used in nitrogen reduction, which wouldincrease buffering capacity and counter the effects ofacidity.
Older (6-7-month-old) needles were more sus-ceptible to injury from mist than current, elongating

Acidic mist and fertilization effects on red spruce 619
needles, as also found by Jacobson et al. (19906), andbad niucb bigber sulphur levels after mist exposuretban new foliage. Tbese differences could be relatedto tbe smaller surface area of new needles exposed tomist or to cbanges in cuticular properties witb needleage. Red spruce needles bave a tbin layer ofepicuticular wax tbat can be altered during bio-syntbesis (Percy et al., 1990), or could be eroded byacidic mist as in Norway spruce {Picea abies (L.)Karst.] (Rinallo et al., 1986). Permeability of K"'tbrougb isolated cuticles of conifer needles is bigberfor young tban old cuticles (Huttunen, Turunen &Reinikainen, 1989). However, contact angles ofdroplets on red spruce needles decrease (i.e. greaterarea of droplet in contact witb needle surface) greatlywitb age (Boyce & Berlyn, 1988), wbicb couldincrease mist penetration in older needles. Incontrast to our findings, Leitb et al. (1989) reportedtbat on 80 % of red spruce seedlings exposed tosimulated acidic mist, current year needles bad tbemost (or only) necrosis.
Tbe strong interaction of nutrient supply and mistcontent points out tbe need to consider nutrientstatus wben interpreting results of mist exposurestudies. Older needles of low-N seedlings bad twiceas mucb injury from sulpbate mist as tbose grown atbigb-N, even tbougb S concentrations were 25 %bigher in bigb-N needles. Higb-N seedlings appar-ently were able to mitigate tbe damage caused byS uptake. Altbougb direct extrapolation from seed-lings to mature foliage is not possible, higb nutrientstatus would likely decrease injury from acidicdeposition in tbe field.
Sboot to root ratio (S/R) was bigber for seedlingsgrown at higb tban low N, largely because ofincreased sboot biomass witb increased nutrientsupply. Tbere was no indication tbat mist contentaffected carbon allocation patterns; root d. wt andS/R did not respond to mist anion. Tbis could bebecause tbe most active period of root growtboccurred well after mist exposure ended (Fig. 2b),and because roots were not directly exposed to mist.In actively growing deciduous tree seedlings exposedto pH 2-0 acid rain, S/R increased because ofdecreased root biomass (Neufeld, Jernstedt &Haines, 1985).
As expected, foliar and root N increased witbincreasing N fertilization. Tbe biggest difference wasfound at tbe start of cold acclimation, when foliar Nwas M % in low- and 2 4 % in bigb-N seedlings. Asuggested optimum for red spruce seedlings is 2-6 %(Swan, 1971). Effects of mist content on tissue Nwere not detected after mist exposure, but tbis wasnot surprising given tbe low N concentration innitrate mist (12 mg N \~^), tbe duration of exposureof new needles (less tban 125 b), exclusion of rootsfrom mist exposure, and tbe difficulty of detectingsmall cbanges in tissue macronutrient levels. HigberN in nitrate- (2-1 % N) versus sulpbate-misted
(1'8 %) needles was reported for red spruce seedlingsexposed to mist for 1040 b outdoors (Jacobson et al.1989).
Altbougb nitrate mist did not detectably increasefoliar N, increased S concentration in sboots exposedto sulpbate mist was readily detected, probablybecause of tbe 10-fold lower background levels of Scompared witb N. Tbere may also be big differencesin tbe amount of nitrate versus sulpbate adsorbed andabsorbed by needles, owing to bigber volatilizationof nitrate from mist drying on foliage (Jacobson et al.1990c). Older needles bad S concentrations up to0-5%, wbicb is bigber tban tbe 0-3 "o reported forred spruce needles exposed to pH 2-6 sulpbate mistfor 17 wk (Jacobson et al., 1989), and more tbantwice as higb as tbe bigbest levels found in coniferneedles exposed to sulpbur pollution in Great Britain(Innes & Boswell, 1989).
Critical sulpbur levels for toxicity bave beensuggested to range from 0-12 to 0-21 % for Norwayspruce (Landolt, Guecbeva & Bucber, 1989). Tbesevalues are at tbe higb end of tbe range reported for Sin mature red spruce; foliar S of 0-11 to 0-22% wasfound in Vermont and New Hampsbire (Lord,1982), and from 0-04 to 0-10% in eastern Maine(Fernandez & Strucbtemeyer, 1984). Because N andS status in conifer foliage are closely correlated(Clement & Gessel, 1985), foliar S/N ratios are ofinterest. Tbe bigbest ratio we found was 0-18 in low-N, sulpbate-misted, older needles, wbicb is bigbertban tbe ratio of 0-04 in Norway spruce (Clement &Gessel, 1985), but mucb lower tban tbe S/N ratio of0-4 reported by Leitb et al. (1989) for red spruceexposed to pH 2-5 mist. Because low-N seedlingsbad lower foliar S and almost identical S/N ratioscompared witb bigb-N seedlings, but still sufferedmore damage, it seems tbat injury may depend onthe status of otber foliar elements ratber tban just Sand N concentrations and ratios.
Nitrate reductase activity in our seedlings wasbigber in roots than shoots, as is common intemperate, perennial plants (Andrews, 1986), es-pecially in gymnosperms (Smirnoff, Todd &Stewart, 1984). Nitrate reductase activity (NRA) didnot increase with increased nitrate supply in thenutrient solution. Because N was provided asNH4NO3, NH4 was likely preferentially absorbedfrom the high-N nutrient solution, as reported forNorway spruce (Marschner, Haussling & George,1991). Conifers typically grow better with NH4 thanNO3 (van den Driessche, 1971) because the carboncosts of assimilating NO3 are higher (Margolis,Vezina & Ouimet, 1988).
There are two possible explanations for why bothfoliar and root NRA were higher in nitrate- thansulphate-misted seedlings. In nitrate-misted seed-lings, nitrate may have been absorbed by needles andtransported to the roots, stimulating NRA in bothtissues; or, in sulphate-misted seedlings, absorbed

620 5 . y. L'Hirondelle, y. S. yacobson and y. P. Lassoie
sulphate may have decreased NRA. The first optionis supported by the observation that the biggesteffect of mist on NRA was in the high-N seedlings,which had the greatest foliar biomass and surfacearea exposed to mist. Norby et al (1989) did notdetect increases in needle NRA of red spruceseedlings exposed to acidic mist, but they usedpH 3-5 and 5-0, and exposed seedlings only fourtimes.
Shoot water content increased with N fertilizationand N in mist. Both forms of N supply increasednew shoot biomass, so higher SWC reflected in-creased tissue expansion. Shoot water content wasalso highest during shoot elongation in white spruce[Picea glauca var. albertiana (S. Brown) Sarg.]seedlings (Grossnickle, 1989).
Effects during cold acclimation
Mist content did not affect gas exchange, shoot watercontent, or needle primordia number after mistexposure ended. Mean photosynthetic rates at thestart of acclimation were lower than for seedlings notexposed to acidic mist (unpublished data; Eamus &Fowler, 1990), so nitrate and sulphate mist may havereduced gas exchange equally.
Fertilization, which continued to the end of ourstudy, affected physiology during acclimation. Gasexchange decreased faster in high- than low-Nseedlings, and high-N needles eventually had thegreatest frost hardiness. Development of frost hardi-ness in conifers is associated with reduced photo-synthesis due to chloroplast and stomatal changes(Teskey, Grier & Hinckley, 1984; Oquist & Strand,1986). Our results show that neither high-N fertil-ization nor high N in acidic mist prolonged activefoliar growth in autumn.
Electrolyte leakage is influenced by many factorssuch as tissue morphology and ion concentrationgradients, and controUed-freezing data should not bedirectly extrapolated to field conditions. However,leakage results provide a useful ranking of frosthardiness among treatments, and have been shownto follow changes in frost hardiness measured bywhole plant freezing injury (Burr et al, 1990). In ourstudy, relative conductivity of excised needles was arelatively insensitive indicator of frost hardinesscompared with frost-induced visible injury to foliageoutdoors. Glerum (1976) stated that excised twigs orneedles always appear to be hardier than those stillattached to trees. This has also been observed incomparing excised needles with intact shoots orseedlings of red spruce (unpublished data). Conse-quently, treatment effects on visible injury wouldlikely be more pronounced than those reported forRC.
Increased frost hardiness of high-N needles hasbeen reported in other studies (DeHayes, Ingle &Waite, 1989; L'Hirondelle, 1990). In contrast to its
beneficial effects on needle frost hardiness, high Nfertilization decreased the number of needle pri-mordia and made buds more sensitive to frost, asfound in other studies with red spruce seedlings(L'Hirondelle, 1990). However, DeHayes et al.(1990) reported that current needles were moresensitive to frost than buds for 30-yr-old red sprucegrowing in a plantation.
Although needle frost hardiness increased with Nfertilization, it decreased with high nitrate in mist.This mist effect disappeared after 3 wk of acclim-ation, perhaps because seedlings had recovered after7 wk without mist. Red spruce seedlings exposedfrom May to October to synthetic cloud watercontaining nitrate and sulphate showed no effects ofmist on frost hardiness measured in March (Klein,Perkins & Myers, 1989). Two studies that exposedred spruce seedlings to mist until November orDecember reported effects on frost hardiness late inacclimation; frost hardiness decreased with increased(NH4).jSO4 and HNO3 until the end of November(Fowler et al, 1989), and decreased with increasedammonium and sulphate, but not nitrate, after 31October (Cape et al, 1991). There are severaldifferences among these studies that might explainthe variation in results; these include seedling ageand seed source, duration and conditions of mistexposure, foliar nutrient status, timing of coldacclimation, and frost hardiness methods.
There are several possible reasons why nitratemist could result in higher needle relative con-ductivity (RC) after freezing than sulphate mist, inour study. Total electrolyte concentration was13—15% higher in nitrate- than sulphate-mistedneedles, but this is not a large enough difference inconcentration gradient (from inside needles to bath-ing solution) to explain the 300 % higher electrolyteleakage from nitrate-misted needles after freezing.Differences in morphology or area of cut surfaces arelikely not large enough to account for the bigdifference in leakage. Mist content had no effect onRC of unfrozen needles, so there were probably nodifferences in electrolytes adsorbed to the cuticle orin the apoplast. These observations suggest thatdifferences in RC values after freezing were fromdiffering effects of nitrate and sulphate on mem-branes.
In summary, fertilization applied to roots affectedthe response of red spruce seedlings to acidic mist,with seedlings of low nutrient status more sensitivethan well-fertilized ones. Sulphate mist led to higherfoliar necrosis and decreased growth compared withnitrate mist. Increased foliar S, but not N, levelswere detected after mist exposure. Nitrate reductaseactivity in foliage was higher in nitrate-mistedseedlings, showing that more nitrate was assimilatedthan in sulphate-misted seedlings. Gas exchange andrelative conductivity during cold acclimation showedthat high-N needles increased frost hardiness faster

Acidic mist and fertilization effects on red spruce 621
than low-N needles, but sensitivity of buds to frostwas increased by high N. Long-term exposure of redspruce seedlings to acidic mist with typical ambientratios of sulphate to nitrate could cause needleinjury, particularly in seedlings of low nutrientstatus, or seedlings with unbalanced nutrition.Susceptibility to early autumn frosts could beincreased for buds by high N fertilization and forneedles by high nitrate mist, particularly for seed-lings with indeterminate growth such as we used.Could mature trees show similar responses ? Wecannot be sure yet; we believe that the infiuence ofacidic mist on red spruce at different times in itsgrowth cycle is likely a complex function of com-position of mist, phenology, and tree nutrition.
ACKNOWLEDGEMENTS
We thank Elizabeth Allis and Larry Heller for excellenttechnical assistance, Mike Rutzke for tissue analyses, andKen Gerow for statistical advice. This research wassupported with funds provided by the Northeastern EorestExperiment Station, Spruce-Fir Research Cooperativewithin the joint U.S. Environmental Protection Agency-USDA Forest Service Eorest Response Program of theNational Acid Precipitation Assessment Program; theAndrew W. Mellon Foundation; and the New York StateAgricultural Experiment Station Regional Hatch ProjectNE-121. This publication has not been subject to policyreview, nor does it necessarily represent the policies of thesupporting agencies.
REFERENCES
ANDREWS, M . (1986). The partitioning of nitrate assimilationbetween root and shoot of higher plants. Plant, Cell andEnvironment 9. 511-519.
BowDEN, R. D., GEBALLE, G . T . & BOWDEN, W . B. (1989). Foliaruptake of '°N from simulated cloud water by red spruce {Picearubens) seedlings. Canadian Journal of Forest Research 19,382-386.
BoYCE, R. L. & BERLYN, G . P. (1988). Measuring the contactangle of water droplets on foliar surfaces. Canadian Journal ofBotany 66, 2599-2602.
BURR, K . E . , TINUS, R. W . , W.M.I.NER, S . J . & KING, R. M .
(1990). Comparison of three cold hardiness tests for coniferseedlings. Tree Physiology 6, 351-369.
CAPE, J. X., LEITH, I. D., FOWLER, D . , MURRAY, .M. B., SHEP-
PARD, L. J., EAMUS, D . & WILSON, R. H . F . (1991). Sulphateand ammonium in nnist impair the frost hardening of red spruceseedlings. New Phytologist 188, 119-126.
CLEMENT, .'\. & GESSEL, S. P. (1985). N, S, P status and proteinsynthesis in the foliage of Norway spruce [Picea abies (L.)Karst.] and .Austrian black pine {Pinus nigra var. nigra). Plantand Soil 85, 345-359.
COLOMBO, S. J., WEBB, D . P. & GLERUM, C . (1984). Frosthardiness testing : An operational manual for use with extendedgreenhouse culture. Ontario Ministry of Natural Resources,Forest Research Report No. 110.
CROOKE, W . M . & SIMPSON, W . E . (1971). Determination ofammonium in Kjeldahl digests of crops by an automatedprocedure. Journal of Science and Food in Agriculture 22, 9-10.
DARRALL, N . M . (1989). The effects of air pollutants onphysiological processes in plants. Plant, Cell and Environment12,' 1-30.
DEHAYES, D . H . , INGLE, M . A. & WAITE, C . E . (1989). Nitrogen
fertilization enhances cold tolerance of red spruce seedlings.Canadian Journal of Forest Research 19, 1037-1043.
DEHAYES, D . H . , WAITE, C . E. , INGLE, M . A. & WILLIAMS,
M. W. (1990). Winter injury susceptibility and cold tolerance ofcurrent and year-old needles of red spruce trees from severalprovenances. Forest Science 36, 982-994.
EAMUS, D . & FOWLER, D . (1990). Photosynthetic and stomatalconductance responses to acid mist of red spruce seedlings.Plant, Cell and Environment 13, 349-357.
EAMUS, D . , LEITH, I. & FOWLER, D . (1989). Water relations of redspruce seedlings treated with acid mist. Tree Physiology 5,387-397.
FERNANDEZ, I. J. & STRUCHTEMEYER, R. A. (1984). Correlationsbetween element concentrations in spruce foliage and forestsoils. Communications in Soil Science and Plant Analysis 15,1243-1255.
FOWLER, D . , CAPE, J. N., DEANS, J. D., LEITH, 1. D., MURRAY,
M. B., SMITH, R. I., SHEPPARD, L . J. & LTNSWORTH, M . H .
(1989). EfTects of acid mist on the frost hardiness of red spruceseedlings. New Phytologist 113, 321-335.
GLERUM, C . (1976). Frost hardiness of forest trees. In: TreePhysiology and Yield Improvement (Ed. by M. G. R. Cannelland F. T. Last), pp. 403^20. Academic Press, New York.
GROSSNICKLE, S. C. (1989). Shoot phenology and water relationsof Picea glauca. Canadian Journal of Forest Research 19,1287-1290.
HuTTUNEN, S., TuRUNEN, M. & REINIKAINEN, J. (1989). Studieson Scots pine {Pinus sylvestris L.) and Norway spruce {Piceaabies (L.) Karst.) needle cuticles. Annales des Sciences Forestieres46, Supplement, 553s-556s.
INNES, J. L. & BOSWELL, R. C . (1989). Sulphur contents of coniferneedles in Great Britain. Geojournal 19, 63-66.
ISAAC, R. A. & JOHNSON, W . C. (1985). Elemental analysis of planttissue by plasma emission spectroscopy: collaborative study.Journal Association Official Analytical Chemists 68, 499-505.
JACOBSON, J. S., BETHARD, T . , HELLER, L . I. & LASSOIE, J. P.
(1990a). Response oi Picea rubens seedlings to varying concen-trations of acidity, sulfur-, and nitrogen-containing pollutantsin mist. Physiologia Plantarum 78, 595-601.
JACOBSON, J. S., HELLER, L . I., GREENWOOD, F . & GOLD, J.
(1990f). Volatilization of nitric acid from foliage of red spruceas an explanation for the greater toxicity of sulfuric than nitricacid. In: Abstracts of International Conference on AcidicDeposition : Its Nature and Impacts, p. 301. Glasgow, Scotland,Royal Society of Edinburgh.
JACOBSON, J. S., HELLER, L . I., YAMADA, K . E. , OSMELOSKI, J. F.,
BETHARD, T . & LASSOIE, J. P. (19906). Foliar injury andgrowth response of red spruce to sulfate and nitrate acidic mist.Canadian Journal of Forest Research 20, 58-65.
JACOBSON, J. S., LASSOIE, J. P., OSMELOSKI, J. & YAMADA, K .
(1989). Changes in foliar elements in red spruce seedlings afterexposure to sulfuric and nitric acid mist. Water, Air, and SoilPotlution 48, 141 159.
JosLiN, J. D., McDuEFlE, C. & BREWER, P . F . (1988). Acidiccloud water and cation loss from red spruce foliage. Water, Air,and Soil Pollution 39, 355-363.
KiMBALL, K. D., JAGELS, R . , GORDON, G . A., WEATHERS, K . C . &
CARLISLE, J. (1988). Differences between New England coastalfog and mountain cloud water chemistry. Water, Air, and SoilPollution 39, 383-393.
KLEIN, R. M . , PERKINS, T . D . & MYERS, H . L . (1989). Nutrientstatus and winter hardiness of red spruce foliage. CanadianJournal of Forest Research 19, 754-758.
LANDOLT, W., GUECHEVA, M . & BUCHER, J. B. (1989). The spatialdistribution of different elements in and on the foliage ofNorway spruce growing in Switzerland. Environmental Pol-lution 56, 15.5-167.
LAURENCE, J. A., KOHUT, R. J., AMUNDSON, R. G . , FAHEY, T . J.,
WEINSTEIN, D . A. , MELKONIAN, J. J., LAUVER, T . L . , BELOIN,
R., SHERMAN, R. , SAYRE, R. & SIMMONS, J. (1988). Response ofplants to interacting stresses. An evaluation of the effects of ozoneand acidic precipitation on the nutrition, growth, and physiology ofred spruce and sugar maple. Report RP2799-1. Submitted to TheElectric Power Research Institute, Palo Alto, CA.
LEBLANC, D . C . & RAYNAL, D . J. (1990). Red spruce decline onWhiteface Mountain, New York. II. Relationships betweenapical and radial growth decline. Canadian Journal of ForestResearch 20, 1415 1421.

622 5. y. L'Hirondelle, y. S. yacobson and y. P. Lassoie
LEITH, I. D . , CAPE, J.N., SHEPPARD, L . J., MURRAY, M . B.,
DEANS, J. P. & FOWLER, D . (1989). Frost hardiness and visibleinjury of spruce seedlings subjected to simulated acid mist.In: Air Pollution and Forest Decline (Ed. by J. B. Bucher and I.Bucher-Wallin), pp. 175-180. IUFRO P2.O5, Birmensdorf.
L'HIRONDELLE, S. J. (1990). Nitrogen fertilization and acidic mistaffect the growth and physiology of red spruce (Picea rubens)seedlings. Ph.D. Thesis, Cornell University, USA.
LORD, D . G . (1982). Foliar and root chemistry of declining redspruce in the Green Mountains of Vermont. M.S. Thesis,University of Pennsylvania.
MARGOLIS, H . A. , VEZINA, L . - P . & OUIMET, R. (1988). Relation oflight and nitrogen source to growth, nitrate reductase andglutamine synthetase activity of jack pine seedlings. PhysiologiaPlantarum 72, 790-795.
MARSCHNER, H . , HAUSSLING, M . & GEORGE, E . (1991). Am-
monium and nitrate uptake rates and rhizosphere pH in non-mycorrhizal roots of Norway spruce [Picea abies (L.) Karst.].Trees S, 14-21.
MoHNEN, V. A. (1988). Mountain cloud chemistry project: Wet,dry and cloud water deposition. Contract No. CR 813934-01-2.U.S. Environmental Protection Agency, Research TrianglePark, NC.
MUELLER, S . F . & WEATHERFORD, F . P. (1988). Chemical de-position to a high elevation red spruce forest. Water, Air, andSoil Pollution 38, 345-363.
NEUFELD, H . S., JERNSTEDT, J. A. & HAINES, B . L . (1985). Directfoliar effects of simulated acid rain. I. Damage, growth and gasexchange. New Phytologist 99, 389-405.
NORBY, R. J., WEERASURIYA, Y . & HANSON, P. J. (1989). Inductionof nitrate reductase activity in red spruce needles by NO^ andHNO,, vapor. Canadian Journal of Forest Research 19, 889-896.
OQUIST, G . & STRAND, M . (1986). Effects of frost hardening onphotosynthetic quantum yield, chlorophyll organization, and
energy distribution between the two photosystems in Scotspine. Canadian Journal of Botany 64, 748-753.
PERCY, K . E . , KRAUSE, C . R. & JENSEN, K . F . (1990). Effects ofozone and acidic fog on red spruce needle epicuticular waxultrastructure. Canadian Journal of Forest Research 2(i, 117-120.
PiTELKA, L. F. & RAYNAL, D . J. (1989). Forest decline and acidicdeposition. Ecology 70, 2-10.
RiNALLO, C , RADI, P . , GELLINI, R . & DI LONARDO, V. (1986).Effects of simulated acid deposition on the surface structure ofNorway spruce and silver fir needles. European Journal of ForestPathology 16, 440--^46.
ROBERTS, B . R. & CANNON, W . N . JR. (1989). Changes in xylempressure potential of red spruce seedlings treated with ozoneand simulated acid rain. Canadian Journal of Forest Research 19,1200-1203.
SMIRNOFF, N . , TODD, P. & STEWART, G . R. (1984). The
occurrence of nitrate reduction in the leaves of woody plants.Annals of Botany 54, 363-374.
SWAN, H . D . S . (1971). Relationships between nutrient supply,growth, and nutrient concentrations in the foliage of white and redspruce. Pulp and Paper Research Institute Canada, WoodlandsReport WR/134, 27 pp.
TESKEY, R. O. , GRIER, C . C . & HINCKLEY, T . M . (1984). Changein photosynthesis and water relations with age and season inAbies amabilis. Canadian Journal of Forest Research 14, 77-84.
THOMAS, R. L . , SHEARD, R. W . & MOYER, J. R. (1967). Com-
parison of conventional and automated procedures for nitrogen,phosphorus, and potassium analysis of plant material using asingle digestion. Agronomy Journal 59, 240-243.
THOMPSON, F . B . & LEYTON, L . (1971). Method for measuringthe leaf surface area of complex shoots. Nature 229, 572.
VAN DEN DRIESSCHE, R. (1971). Response of conifer seedlings tonitrate and ammonium sources of nitrogen. Plant and Soil 34,421-439.





















