
Accelerated arterial stiffening and gene expression profile of the aorta in patients with coronary...
11
Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited. Accelerated arterial stiffening and gene expression profile of the aorta in patients with coronary artery disease Ce ´ line Fassot a , Marie Briet b , Philippe Rostagno c , Pascal Barbry c , Claudine Perret a , Dominique Laude a , Pierre Boutouyrie a , Erwan Bozec a , Patrick Bruneval e , Christian Latremouille d and Ste ´ phane Laurent a Background Hypertension and chronic renal failure (CRF) are considered models of accelerated arterial stiffening. Arterial stiffness increases further when CRF is associated with hypertension. We hypothesized that, in patients with mild CRF, aortic gene expression profile would include genes involved in arterial calcifications and enlargement. Method We analysed human aorta with the ‘GeneChip Microarray’ technology, in patients with or without CRF, scheduled for a coronary artery bypass graft. Results Nine of 25 patients had high-quality RNA and were included in the study. Among the 101 transcripts differentially expressed between CRF patients and controls, 97 transcripts were overexpressed in CRF patients. Two genes had the highest overexpression in CRF patients: lumican (LUM), involved in the regulation of collagen fibrillogenesis; and ornithine decarboxylase (ODC1), involved in polyamine biosynthesis, smooth muscle cell growth and proliferation. Immunohistochemical staining revealed an increased amount of LUM and ODC1 in the vascular smooth muscle cells (VSMCs) of CRF compared to non-CRF aortic sections. Eight genes were implicated in the regulation of the cytoskeleton (including capping protein muscle Z-line 1 a and moesin) and cell migration, and five genes were implicated in extracellular matrix function and apoptosis. A trend towards an upregulation of candidate genes involved in arterial calcifications was observed in CRF patients, but did not reach statistical significance. Carotid-femoral pulse wave velocity was not correlated with gene expression level. Conclusion In conclusion, these results show that patients at an early stage of CRF have a specific gene expression profile of aortic tissue and suggest that genes implicated in collagen fibrillogenesis, and VSMCs migration and proliferation, particularly LUM and ODC1, may play a role. J Hypertens 26:747–757 Q 2008 Wolters Kluwer Health | Lippincott Williams & Wilkins. Journal of Hypertension 2008, 26:747–757 Keywords: aorta, arterial remodelling, arterial stiffness, kidney disease, microarray Abbreviations: ALPL, alkaline phosphatase; APOD, apolipoprotein D; BGLAP, osteocalcin CAD, coronary artery disease; CAP1, adenylate cyclase-associated protein 1; CAPZA1, capping protein muscle Z-line 1 alpha; CCA, common carotid artery; COL I, collagen I; COL II, collagen II; CRF, chronic renal failure; CV, cardiovascular; ESRD, end-stage renal disease; ET1, endothelin 1; GFR, glomerular filtration rate; HCT, hypercholesterolemia; LEP, leptin; MDRD, Modification of Diet in Renal Disease; MGP, matrix gla protein; MI, myocardial infarction; MSN, moesin; ODC1, ornithine decarboxylase; OPG, osteoprotegerin; PWV, pulse wave velocity; SERPIN 2, serine proteinase inhibitor; SMC, smooth muscle cell; SSP1, osteopontin; VSMC, vascular smooth muscle cell a Faculty of Medicine, University Paris-Descartes, Department of Pharmacology, Georges Pompidou European Hospital, and Institut National de la Sante ´ et de la Recherche Me ´ dicale U652, b Department of Physiology, Georges Pompidou European Hospital, Paris, c Laboratory of Physiological Genomics, Sofia-Antipolis, d Department of Cardiac Surgery, Georges Pompidou European Hospital and e Department of Pathology, Georges Pompidou European Hospital, and Institut National de la Sante ´ et de la Recherche Me ´ dicale U652, Paris, France Correspondence to Ste ´ phane Laurent, MD, PhD, Department of Pharmacology and Institut National de la Sante ´ et de la Recherche Me ´ dicale U652, Ho ˆ pital Europeen Georges Pompidou, 20 rue Leblanc, 75015 Paris, France Tel: +33 1 56 09 39 91; fax: +33 1 56 09 39 92; e-mail: [email protected] Received 25 August 2007 Revised 11 November 2007 Accepted 23 November 2007 Introduction Arterial stiffness and wave reflections are now well accepted as the most important determinants of increasing systolic and pulse pressures in ageing societies [1]. Hyper- tension and chronic renal failure (CRF) are considered models of accelerated arterial stiffening [1–3]. We recently showed that, compared to uncomplicated hypertension, arterial stiffness further increased when mild-to-moderate CRF was associated with hypertension [4]. The molecular mechanisms of arterial stiffening have been well described in hypertension [2,5] and end- stage renal disease (ESRD) [3,6], but little in mild-to-moderate CRF. CRF patients have an increased risk of cardiovascular morbidity and mortality [7], correlated to the decrease of glomerular filtration rate (GFR) [8] that impairs cardiac and arterial functions. Pathogenic mechanisms of arterial disease in CRF include endothelial dysfunction [9] lead- ing to vasoconstriction and arterial growth, oxidant stress [10], dysfunction of nitric oxide metabolism [11], increase in inflammatory factors [12,13], and vascular calcifications [6,14]. Vascular calcifications in ESRD patients have strong predictive value for cardiovascular and all-cause mortality [15]. A likely mechanism is the association with an increased stiffness of large, capacitive, elastic-type Original article 747 0263-6352 ß 2008 Wolters Kluwer Health | Lippincott Williams & Wilkins
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Accelerated arterial stiffening and gene expression profile of the aorta in patients with coronary...

C
Original article 747
Accelerated arterial stiffening
and gene expression profile ofthe aorta in patients with coronary artery diseaseCeline Fassota, Marie Brietb, Philippe Rostagnoc, Pascal Barbryc,Claudine Perreta, Dominique Laudea, Pierre Boutouyriea, Erwan Bozeca,Patrick Brunevale, Christian Latremouilled and Stephane LaurentaBackground Hypertension and chronic renal failure
(CRF) are considered models of accelerated arterial
stiffening. Arterial stiffness increases further when CRF is
associated with hypertension. We hypothesized that, in
patients with mild CRF, aortic gene expression profile would
include genes involved in arterial calcifications and
enlargement.
Method We analysed human aorta with the ‘GeneChip
Microarray’ technology, in patients with or without CRF,
scheduled for a coronary artery bypass graft.
Results Nine of 25 patients had high-quality RNA and were
included in the study. Among the 101 transcripts
differentially expressed between CRF patients and controls,
97 transcripts were overexpressed in CRF patients. Two
genes had the highest overexpression in CRF patients:
lumican (LUM), involved in the regulation of collagen
fibrillogenesis; and ornithine decarboxylase (ODC1),
involved in polyamine biosynthesis, smooth muscle cell
growth and proliferation. Immunohistochemical staining
revealed an increased amount of LUM and ODC1 in the
vascular smooth muscle cells (VSMCs) of CRF compared to
non-CRF aortic sections. Eight genes were implicated in the
regulation of the cytoskeleton (including capping protein
muscle Z-line 1 a and moesin) and cell migration, and five
genes were implicated in extracellular matrix function and
apoptosis. A trend towards an upregulation of candidate
genes involved in arterial calcifications was observed in
CRF patients, but did not reach statistical significance.
Carotid-femoral pulse wave velocity was not correlated with
gene expression level.
opyright © Lippincott Williams & Wilkins. Unauth
0263-6352 � 2008 Wolters Kluwer Health | Lippincott Williams & Wilkins
Conclusion In conclusion, these results show that patients
at an early stage of CRF have a specific gene expression
profile of aortic tissue and suggest that genes implicated in
collagen fibrillogenesis, and VSMCs migration and
proliferation, particularly LUM and ODC1, may play a role.
J Hypertens 26:747–757 Q 2008 Wolters Kluwer Health |
Lippincott Williams & Wilkins.
Journal of Hypertension 2008, 26:747–757
Keywords: aorta, arterial remodelling, arterial stiffness, kidney disease,microarray
Abbreviations: ALPL, alkaline phosphatase; APOD, apolipoprotein D;BGLAP, osteocalcin CAD, coronary artery disease; CAP1, adenylatecyclase-associated protein 1; CAPZA1, capping protein muscle Z-line 1alpha; CCA, common carotid artery; COL I, collagen I; COL II, collagen II;CRF, chronic renal failure; CV, cardiovascular; ESRD, end-stage renaldisease; ET1, endothelin 1; GFR, glomerular filtration rate; HCT,hypercholesterolemia; LEP, leptin; MDRD, Modification of Diet in RenalDisease; MGP, matrix gla protein; MI, myocardial infarction; MSN, moesin;ODC1, ornithine decarboxylase; OPG, osteoprotegerin; PWV, pulse wavevelocity; SERPIN 2, serine proteinase inhibitor; SMC, smooth muscle cell;SSP1, osteopontin; VSMC, vascular smooth muscle cell
aFaculty of Medicine, University Paris-Descartes, Department of Pharmacology,Georges Pompidou European Hospital, and Institut National de la Sante et de laRecherche Medicale U652, bDepartment of Physiology, Georges PompidouEuropean Hospital, Paris, cLaboratory of Physiological Genomics, Sofia-Antipolis,dDepartment of Cardiac Surgery, Georges Pompidou European Hospital andeDepartment of Pathology, Georges Pompidou European Hospital, and InstitutNational de la Sante et de la Recherche Medicale U652, Paris, France
Correspondence to Stephane Laurent, MD, PhD, Department of Pharmacologyand Institut National de la Sante et de la Recherche Medicale U652, HopitalEuropeen Georges Pompidou, 20 rue Leblanc, 75015 Paris, FranceTel: +33 1 56 09 39 91; fax: +33 1 56 09 39 92;e-mail: [email protected]
Received 25 August 2007 Revised 11 November 2007Accepted 23 November 2007
IntroductionArterial stiffness and wave reflections are now well
accepted as the most important determinants of increasing
systolic and pulse pressures in ageing societies [1]. Hyper-
tension and chronic renal failure (CRF) are considered
models of accelerated arterial stiffening [1–3]. We recently
showed that, compared to uncomplicated hypertension,
arterial stiffness further increased when mild-to-moderate
CRF was associated with hypertension [4]. The molecular
mechanisms of arterial stiffening have been well described
in hypertension [2,5] and end- stage renal disease (ESRD)
[3,6], but little in mild-to-moderate CRF.
CRF patients have an increased risk of cardiovascular
morbidity and mortality [7], correlated to the decrease of
glomerular filtration rate (GFR) [8] that impairs cardiac
and arterial functions. Pathogenic mechanisms of arterial
disease in CRF include endothelial dysfunction [9] lead-
ing to vasoconstriction and arterial growth, oxidant stress
[10], dysfunction of nitric oxide metabolism [11], increase
in inflammatory factors [12,13], and vascular calcifications
[6,14]. Vascular calcifications in ESRD patients have
strong predictive value for cardiovascular and all-cause
mortality [15]. A likely mechanism is the association with
an increased stiffness of large, capacitive, elastic-type
orized reproduction of this article is prohibited.

C
748 Journal of Hypertension 2008, Vol 26 No 4
arteries such as the aorta and the common carotid artery
[15]. Vascular calcifications occur more often and at an
earlier age in patients with ESRD than in normal controls.
We previously showed that arterial enlargement and
increased arterial stiffness occur in parallel with the
decline in renal function in patients with mild-to-mod-
erate CRF [4].
We hypothesized that, in patients with mildly altered
renal function, the gene expression profile of the aorta
would include genes involved in arterial calcifications and
remodelling. We used punch biopsies of patients sched-
uled for coronary artery bypass surgery and a microarray
technology [16] to characterize the gene expression pro-
file of aortic tissue. We compared patients with mild-to-
moderate CRF to patients with normal renal function,
and studied the relationship between aortic stiffness,
measured through carotid-femoral pulse wave velocity
(PWV), and underexpressed or overexpressed genes.
Materials and methodsPatientsTwenty-five patients, with and without CRF, scheduled
for coronary artery bypass surgery, were recruited at the
Department of Cardiac Surgery of Hopital Europeen
Georges Pompidou. The study was conducted according
to the guidelines of the ethics committee of the institu-
tion. We included patients with coronary artery disease
(CAD) because they were carrying an ischemic cardio-
myopathy without an associated disease that was able to
compromise the interpretation of the results. Demo-
graphic data with details of cardiovascular risk factors
and blood samples were collected on the day when aortic
stiffness was measured, that is the day before the bypass
surgery. Hypertension, diabetes and hypercholesterole-
mia (HCT) were indicated by a previous diagnosis or the
use of antihypertensive, oral hypoglycaemic or choles-
terol-lowering agents, respectively. Mild renal insuffi-
ciency was due to hypertension and/or diabetes, and
not other disease.
Glomerular filtration rate measurementsThe Modification of Diet in Renal Disease (MDRD)
simplified equation was used to estimate GFR [17]:
Glomerular filtration rateðGFRÞ½ml=min per 1:73m2�
¼ 186� ½serum Crðmg=dlÞ��1:154 � ½age��0:203
� ½0:742 if female� � ½1:210 if black�
Pulse wave velocity measurementsAortic stiffness was estimated by carotido-femoral PWV,
measured along the descending thoraco-abdominal
aorta by the foot-to-foot velocity method, as previously
published and validated [18]. Briefly, waveforms were
obtained transcutaneously over the right common
opyright © Lippincott Williams & Wilkins. Unautho
carotid artery (CCA) and femoral artery, and the time
delay (t) was measured between the feet of the two
waveforms.
We adjusted PWV on age, HCT and previous myocardial
infarction (MI) using a general linear model to determine
the multivariate b coefficients in a stepwise manner and
to calculate the predicted values on the basis of the
model. In a multivariate analysis of the 25 patients,
age, HCT and MI were independently correlated to
PWV (P< 0.05 each), and explained, in stepwise
regression analysis, 28, 16 and 31% of the variance (R2
increment) of PWV, respectively.
Aortic biopsiesThe punch biopsies (one to three samples per patient)
taken at the time of the coronary bypass surgery
were immediately conserved in RNAlater (Qiagen, Cour-
taboeuf, France) for RNA analysis or frozen in liquid
nitrogen for western blot and stored at �808C until
processing, or fixed in formaldehyde for immunohisto-
chemistry. In four patients, a pilot histological study has
shown a normal ascending aorta, without atherosclerosis
and overt calcifications. The thickness of the intima was
normal. The media was normal, with a regular organiz-
ation of elastin lamellae and smooth muscle cells (SMCs).
The adventitia was normal. These findings strongly
suggest that the RNA, extracted from the punch biopsy,
is representative of a normal SMC population, rather than
an atherosclerotic cell population, even in CRF patients.
Aortic tissue was taken in a young organ donor during
transplantation, to serve as reference. Indeed, the extent
of atherosclerosis was limited in this subject.
RNA extraction and amplificationAfter extraction of total RNA from human aortic samples
by TRIzol reagent (Invitrogen, Cergy Pontoise, France),
quality and concentration were checked by the Agilent
Technology. RNA with a good quality was amplified
by the Amino Allyl MessageAmp aRNA kit (Ambion,
Huntingdon, UK), according to the manufacturer’s
instructions.
Microarray analysisThe oligonucleotide microarrays were supplied by the
laboratory of Physiological Genomics of Sophia-Antipo-
lis. These arrays contained 27 648 oligonucleotides
probes (length � 51 bp) specific to 25 279 human tran-
scripts. The list of the probes is available online (http://
www.microarray.fr:8080/mediante/index). Antisens RNA
was coupled with Cy3 and Cy5 postlabelling reactive
dyes (Amersham, Little Chalfont, UK). After a postpro-
cessing step, microarrays were hybridized with labelled-
RNA as previously described [19]. Slides were scanned
with a ScanArray 5000 (GSI Lumonics, Munich,
Germany) and images were quantified with the GenePix
Pro 5.0 program (Axon Instruments, Chatillon, France).
rized reproduction of this article is prohibited.

C
Aortic gene expression profile Fassot et al. 749
Fig. 1
27648 oligonucleotides probes
17 897 oligonucleotides probes
1716 probes
101 probesDifferentially expressed between CRF+ and CRF--
Gene expresion level > 500
Present in all samples (n = 9)
Wilcoxon testP < 0.05
97 probesOverexpressed in CRF+
≥ 1.5-fold
Scheme for analysis of profiling data. CRF, chronic renal failure.
On each slide of microarray, we hybridized the aRNA of
a patient and the reference (donor). This technique
allowed us to normalize values of patients to reference
values and then compare patients between them. More-
over, because incorporations of Cy3 and Cy5 dyes can
differ significantly, data were further normalized using a
dye-swap method [20]. Statistical analysis was performed
using Microsoft Excel 2003 functions as described in
Fig. 1.
Protein analysisImmunohistochemistry
The proteins encoded by differentially expressed genes
were studied by immunohistochemistry on aortic biop-
sies of CRF and non-CRF patients. Paraffin-embedded
sections were treated with xylen, fixed for 10 min in cold
acetone and kept wet in Tris-buffered saline (TBS)
until processing. All antigens were detected with very
well characterized monoclonal [antidynein (Santa-Cruz
Biotechnology, Santa-Cruz, USA), moesin (MSN; Lab
Vision Corporation, Fremont, USA)] and polyclonal
[anti-CAPZA1 (Chemicon, Chandlers Ford, UK),
ornithine decarboxylase (ODC1; Santa-Cruz Biotechnol-
ogy, Santa-Cruz, USA), osteoprotegerin (OPG), osteocal-
cin and lumican (LUM; R&D Systems, Lille, France)]
antibodies. Dilutions used were done according to the
manufacturer’s instructions. Antigen presence was
revealed by the indirect immunoperoxidase method.
Western blot
A western blot analysis of proteins of interest was per-
formed following standard techniques. Proteins were
extracted from arterial biopsies of CRF and non-CRF
opyright © Lippincott Williams & Wilkins. Unauth
patients. Total protein content was determined by
the Bradford technique [21]. Equal amounts (50 mg) of
the denatured proteins were loaded per lane, separated
on a Ready Gel 10% Tris–HCl polyacrylamide gel
(Bio-Rad, France) and transferred to a nitrocellulose
membrane. Membranes were incubated with mono-
clonal antibodies directed against MSN or polyclonal
antibodies against OPG. Subsequent analysis used an
antimouse, an antigoat or an antichicken IgG peroxidase
complex, as a second antibody (dilutions 1 : 3000–
1 : 5000), and chemiluminescence emitted from luminol
oxidized by peroxidase as a detection method (ECL
System, Amersham).
ResultsPatientsOn the basis of their GFR, the 25 patients were split into
equal tertiles: lowest GFR (37–68 ml/min per 1.73 m2),
medium GFR (69–91 ml/min per 1.73 m2) and highest
values (greater than 92 ml/min per 1.73 m2) (Table 1).
The patients of the lowest tertile were considered as
CRF population and the others as non-CRF population.
These two groups were not significantly different, as far
as sex ratio, brachial SBP, DBP, MBP, PP and PWV were
concerned (Table 1). In the CRF population, time from
the beginning of the disease was 10� 4 years.
According to the quantity and quality of the extracted
RNA, nine patients, including four non-CRF patients
and five CRF patients, were analysed on oligonucleotides
microarray. The clinical characteristics of the total popu-
lation, the nine patients used for microarray analysis and
the five patients used for protein analysis, are presented
in Table 1.
Analysis of genes involved in arterial calcificationWe first tested the hypothesis that genes encoding
proteins implicated in vascular calcifications would be
differentially expressed between aortic tissues of patients
with CRF compared to patients without CRF. Although
the expression level of genes encoding proteins involved
in vascular calcifications [matrix Gla protein: MGP; osteo-
pontin: SSP1; osteoprotegerin: TNFRSF11B; osteocal-
cin: BGLAP; and osteo/chondrocytic transcription factors
(Sox9, Runx2 and Msx2); alkaline phosphatase: ALPL;
leptin: LEP; collagen I: COL I; collagen II: COL II]
was 1.2–2.2-fold higher than in non-CRF patients, no
significant difference was observed between groups
(Table 2).
Differential gene expression of the complete data setThe complete data set was filtered to remove transcripts
with low expression level (less than 500) and not present
in all of the nine samples, yielding 1716 genes (Fig. 1).
Statistically significant changes in gene expression were
identified using a Wilcoxon test with a P-value less than
0.05. There were 101 genes differentially expressed
orized reproduction of this article is prohibited.
C
750 Journal of Hypertension 2008, Vol 26 No 4
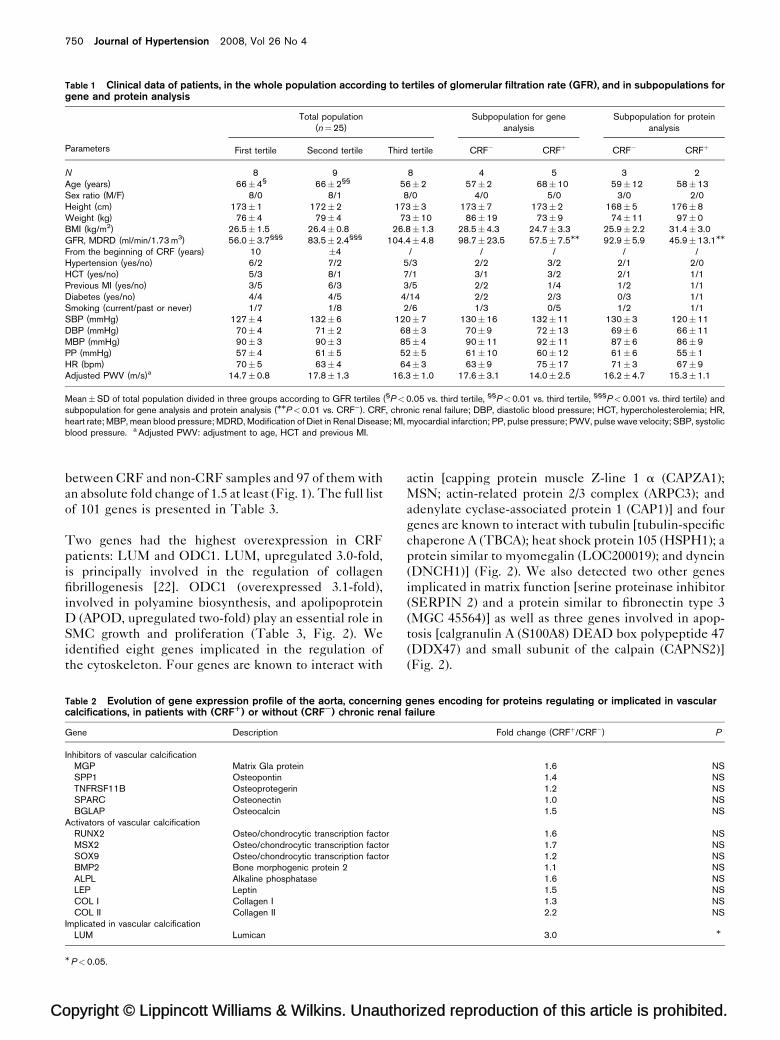
Table 1 Clinical data of patients, in the whole population according to tertiles of glomerular filtration rate (GFR), and in subpopulations forgene and protein analysis
Parameters
Total population(n¼25)
Subpopulation for geneanalysis
Subpopulation for proteinanalysis
First tertile Second tertile Third tertile CRF� CRFþ CRF� CRFþ
N 8 9 8 4 5 3 2Age (years) 66�4§ 66�2§§ 56�2 57�2 68�10 59�12 58�13Sex ratio (M/F) 8/0 8/1 8/0 4/0 5/0 3/0 2/0Height (cm) 173�1 172�2 173�3 173�7 173�2 168�5 176�8Weight (kg) 76�4 79�4 73�10 86�19 73�9 74�11 97�0BMI (kg/m2) 26.5�1.5 26.4�0.8 26.8�1.3 28.5�4.3 24.7�3.3 25.9�2.2 31.4�3.0GFR, MDRD (ml/min/1.73 m3) 56.0�3.7§§§ 83.5�2.4§§§ 104.4�4.8 98.7�23.5 57.5�7.5MM 92.9�5.9 45.9�13.1MM
From the beginning of CRF (years) 10 �4 / / / / /Hypertension (yes/no) 6/2 7/2 5/3 2/2 3/2 2/1 2/0HCT (yes/no) 5/3 8/1 7/1 3/1 3/2 2/1 1/1Previous MI (yes/no) 3/5 6/3 3/5 2/2 1/4 1/2 1/1Diabetes (yes/no) 4/4 4/5 4/14 2/2 2/3 0/3 1/1Smoking (current/past or never) 1/7 1/8 2/6 1/3 0/5 1/2 1/1SBP (mmHg) 127�4 132�6 120�7 130�16 132�11 130�3 120�11DBP (mmHg) 70�4 71�2 68�3 70�9 72�13 69�6 66�11MBP (mmHg) 90�3 90�3 85�4 90�11 92�11 87�6 86�9PP (mmHg) 57�4 61�5 52�5 61�10 60�12 61�6 55�1HR (bpm) 70�5 63�4 64�3 63�9 75�17 71�3 67�9Adjusted PWV (m/s)a 14.7�0.8 17.8�1.3 16.3�1.0 17.6�3.1 14.0�2.5 16.2�4.7 15.3�1.1
Mean�SD of total population divided in three groups according to GFR tertiles (§P<0.05 vs. third tertile, §§P<0.01 vs. third tertile, §§§P<0.001 vs. third tertile) andsubpopulation for gene analysis and protein analysis (MMP<0.01 vs. CRF�). CRF, chronic renal failure; DBP, diastolic blood pressure; HCT, hypercholesterolemia; HR,heart rate; MBP, mean blood pressure; MDRD, Modification of Diet in Renal Disease; MI, myocardial infarction; PP, pulse pressure; PWV, pulse wave velocity; SBP, systolicblood pressure. a Adjusted PWV: adjustment to age, HCT and previous MI.
between CRF and non-CRF samples and 97 of them with
an absolute fold change of 1.5 at least (Fig. 1). The full list
of 101 genes is presented in Table 3.
Two genes had the highest overexpression in CRF
patients: LUM and ODC1. LUM, upregulated 3.0-fold,
is principally involved in the regulation of collagen
fibrillogenesis [22]. ODC1 (overexpressed 3.1-fold),
involved in polyamine biosynthesis, and apolipoprotein
D (APOD, upregulated two-fold) play an essential role in
SMC growth and proliferation (Table 3, Fig. 2). We
identified eight genes implicated in the regulation of
the cytoskeleton. Four genes are known to interact with
opyright © Lippincott Williams & Wilkins. Unautho
Table 2 Evolution of gene expression profile of the aorta, concerningcalcifications, in patients with (CRFR) or without (CRFS) chronic renal
Gene Description
Inhibitors of vascular calcificationMGP Matrix Gla proteinSPP1 OsteopontinTNFRSF11B OsteoprotegerinSPARC OsteonectinBGLAP Osteocalcin
Activators of vascular calcificationRUNX2 Osteo/chondrocytic transcription factorMSX2 Osteo/chondrocytic transcription factorSOX9 Osteo/chondrocytic transcription factorBMP2 Bone morphogenic protein 2ALPL Alkaline phosphataseLEP LeptinCOL I Collagen ICOL II Collagen II
Implicated in vascular calcificationLUM Lumican
MP<0.05.
actin [capping protein muscle Z-line 1 a (CAPZA1);
MSN; actin-related protein 2/3 complex (ARPC3); and
adenylate cyclase-associated protein 1 (CAP1)] and four
genes are known to interact with tubulin [tubulin-specific
chaperone A (TBCA); heat shock protein 105 (HSPH1); a
protein similar to myomegalin (LOC200019); and dynein
(DNCH1)] (Fig. 2). We also detected two other genes
implicated in matrix function [serine proteinase inhibitor
(SERPIN 2) and a protein similar to fibronectin type 3
(MGC 45564)] as well as three genes involved in apop-
tosis [calgranulin A (S100A8) DEAD box polypeptide 47
(DDX47) and small subunit of the calpain (CAPNS2)]
(Fig. 2).
rized reproduction of this article is prohibited.
genes encoding for proteins regulating or implicated in vascularfailure
Fold change (CRFþ/CRF�) P
1.6 NS1.4 NS1.2 NS1.0 NS1.5 NS
1.6 NS1.7 NS1.2 NS1.1 NS1.6 NS1.5 NS1.3 NS2.2 NS
3.0 M

Copyright © Lippincott Williams & Wilkins. Unauthorized reproduction of this article is prohibited.
Aortic gene expression profile Fassot et al. 751
Table 3 One hundred and one transcripts differentially expressed between aorta of chronic renal failure (CRF) and non-CRF patients
Gene Description Fold change Category
STK3 Serine/threonine kinase 3 1.6 Signalling/communicationMRGX4 MAS-related gene MRGX4 1.6 Signalling/communicationIFNA2 Interferon a 2 1.5 Cell defenseTRIP15 Thyroid receptor interacting protein 15 1.8 Signalling/communicationPCDHB16 Protocadherin b16 1.6 Signalling/communicationAIM1 Absent in melanoma 1 1.6 Signalling/communicationBACH Brain acyl-CoA hydrolase 1.6 MetabolismLOC378344 Similar to riken cDNA 6332401019 gene 1.7 UnclassifiedLOC200019 Similar to myomegalin 1.5 Cell structure/motilityMGC45564 Similar to fibronectin type 3 1.6 Cell structure/motilityMGC31963 Kidney predominant protein NCU-G1 1.8 UnclassifiedFLJ21128 Hypothetical protein FLJ21128 1.7 UnclassifiedNUDT11 Nucleoside diphosphate linked moiety X m 11 2.2 Gene/protein expressionPRDM10 PR domain containing 10 1.9 Gene/protein expressionMANBA Mannosidase ba lysosomal 1.8 MetabolismBCL9 B-cell CLL/lymphoma 9 1.8 Signalling/communicationPIGA Phosphatidylinositol glycan, class A 1.9 Signalling/communicationRFWD1 TNF receptor associated factor 7 1.5 Cell divisionDNCH1 Dynein cytoplasmic heavy polypeptide 1 1.8 Cell structure/motilityC20orf11 Chromosome 20 open reading frame 11 2.1 Gene/protein expressionFLJ32752 Hypothetical protein FLJ32752 2.0 UnclassifiedGGA1 Golgi ass. g adaptin ear cont. ARF bind. prot. 1 1.5 Gene/protein expressionSENP7 Sentrin-specific protease 7 1.6 Gene/protein expressionARPC3 Actin-related protein 2/3 complex 1.7 Cell structure/motilityTPK1 Thiamin pyrophosphokinase 1 1.6 Cell structure/motilityLOC158471 Hypothetical protein BC019095 1.8 MetabolismFLJ36664 Hypothetical protein FLJ36664 1.9 UnclassifiedLOC150035 cDNA FLJ11616 fis 1.9 UnclassifiedZNF304 Zinc finger protein 304 1.6 Gene/protein expressionLALP1 Lysosomal apyrase-like protein 1 1.6 Cell divisionKIAA1107 Unknow protein mRNA, partial cds 1.5 UnclassifiedMSN Moesin 1.8 Cell structure/motilityZNF225 Zinc finger protein 225 1.7 Gene/protein expressionFLJ14816 Hypothetical protein FLJ14816 1.6 UnclassifiedH1F0 Histone family 1.8 Cell divisionTRIP3 Thyroid hormone receptor interactor 3 2.1 Signalling/communicationIRF1 Interferon regulatory factor 1 1.9 Gene/protein expressionTLOC1 Translocation protein 1 1.7 Signalling/communicationMGC16169 Hypothetical protein MGC16169 2.1 UnclassifiedMGC26610 Hypothetical protein MGC26610 1.7 UnclassifiedKSP37 Ksp 37 protein 1.5 Cell defenseSERPIN2 Serine proteinase inhibitor, clade E, member 2 1.7 Signalling/communicationKIAA1677 cDNA DKFZp434M0516 1.8 UnclassifiedCLECSF1 C-type lectin, superfamily member 1 (cart-der) 1.9 UnclassifiedDKFZp451J0118 DKFZp451J0118 protein 1.7 MetabolismFLJ21816 Hypothetical protein FLJ21216 1.9 Signalling/communicationPME-1 Protein phosphatase methytransferase 1 1.7 Gene/Protein expressionINSRR Insulin receptor-related protein 1.6 Cell structure/motilityC20orf155 Chromosome 20 open reading frame 155 1.6 Gene/protein expressionCAP1 Adenylate cyclase associated protein 1 1.5 Cell structure/motilityEBAF Endometrial bleeding associated factor 1.3 Signalling/communicationZNF18 Zinc finger protein 18 1.4 Gene/protein expressionDDX47 DEAD box polypeptide 47 2.1 Cell divisionFUT4 Fucosyltransferase 4 1.9 MetabolismPAIP2 Polybinding protein interacting protein 2 2.1 Signalling/communicationCOX7C Cytochrome c oxidase subunit viic 2.3 MetabolismCAPZA1 Capping protein muscle Z-line a 1 2.1 Cell structure/motilityAPOD Apolipoprotein D 2.0 MetabolismTUWD12 Acetylgalactosaminyltransferase 2.0 MetabolismKR19 Zinc finger protein 19 1.8 Gene/protein expressionHMGN2 High-mobility group nucleosomal-binding domain 2 2.6 Cell divisionUBN1 Ubinuclein 1 1.7 Signalling/communicationTBCA Tubulin-specific chaperone a 2.1 Cell structure/motilityTEAD2 TEA domain family member 2 1.5 Gene/protein expressionSTK38L Serine/threonine kinase 38-like 1.5 Gene/protein expressionTM4SF1 Transmembrane 4 superfamily member 1 1.7 Signalling/communicationFLJ25157 Hypothetical protein FLJ25157 1.8 Signalling/communicationTAF10 Taf 10 RNA polymerase II 1.9 UnclassifiedC15orf15 Chromosome 15 open reading frame 15 2.2 Gene/protein expressionSTC2 Stanniocalcin 2 1.9 Gene/protein expressionALS2CR17 Amyotrophic lateral sclerosis 2 chromosome region 17 2.5 Cell defenseODC1 Ornithine decarboxylase 1 3.1 MetabolismLUM Lumican 3.0 Cell structure/motilityHM13 Histocompatibility 13 2.4 Cell structure/motilityPOLS Polymerase sigma 2.1 Cell defense

C
752 Journal of Hypertension 2008, Vol 26 No 4
Table 3 (continued )
Gene Description Fold change Category
C14orf92 Chromosome 14 open reading frame 92 2.0 Gene/protein expressionZNF211 Zinc finger protein 211 1.7 Gene/protein expressionZNF217 Zinc finger protein 217 1.9 Gene/protein expressionSSX2 Synovial sarcoma X2 1.9 Gene/protein expressionSFRS7 Splicing factor 1.7 Gene/protein expressionFLJ10634 Hypothetical protein FLJ10634 1.9 UnclassifiedPIP3AP Myotubularin-related protein 12 2.0 Signalling/communicationCAPNS2 Calpain small subunit 2 2.1 Gene/protein expressionLOC340371 Hypothetical protein LOC340371 2.2 UnclassifiedRPS6KC1 Ribosomal protein S6 kinase 1.6 Gene/protein expressionMAPK10 MAP kinase 10 1.9 Signalling/communicationC18orf1 Chromosome 18 open reading frame 1 1.7 Gene/protein expressionARL7 ADP-ribosylation factor-like 7 1.8 Gene/protein expressionCCL11 Eotaxin 1 1.6 Signalling/communicationMGC20446 Hypothetical protein MGC20447 1.7 UnclassifiedNYX Nyctalopin 1.8 Signalling/communicationS100A8 Calgranulin A 2.1 Cell divisionRALBP1 Rala-binding protein 1 1.4 Signalling/communicationHSPH1 Heat shock protein 105 1.4 Cell structure/motilityTBX19 T-box 19 1.9 Gene/Protein expressionTJP4 Tight junction protein 4 1.7 Signalling/communicationCSNK2A2 Casein kinase 2 2.0 Signalling/communicationNEURL Neuralized homolog like 1.8 Cell divisionIL13RA1 Interleukin 13 receptor a1 2.0 Signalling/communicationUBE2J2 Ubiquitin-conjugated enzyme E2, J2 1.7 Gene/protein expressionHCLS1 Hematopoietic cell-specific Lyn substrate 1 1.8 Cell defense
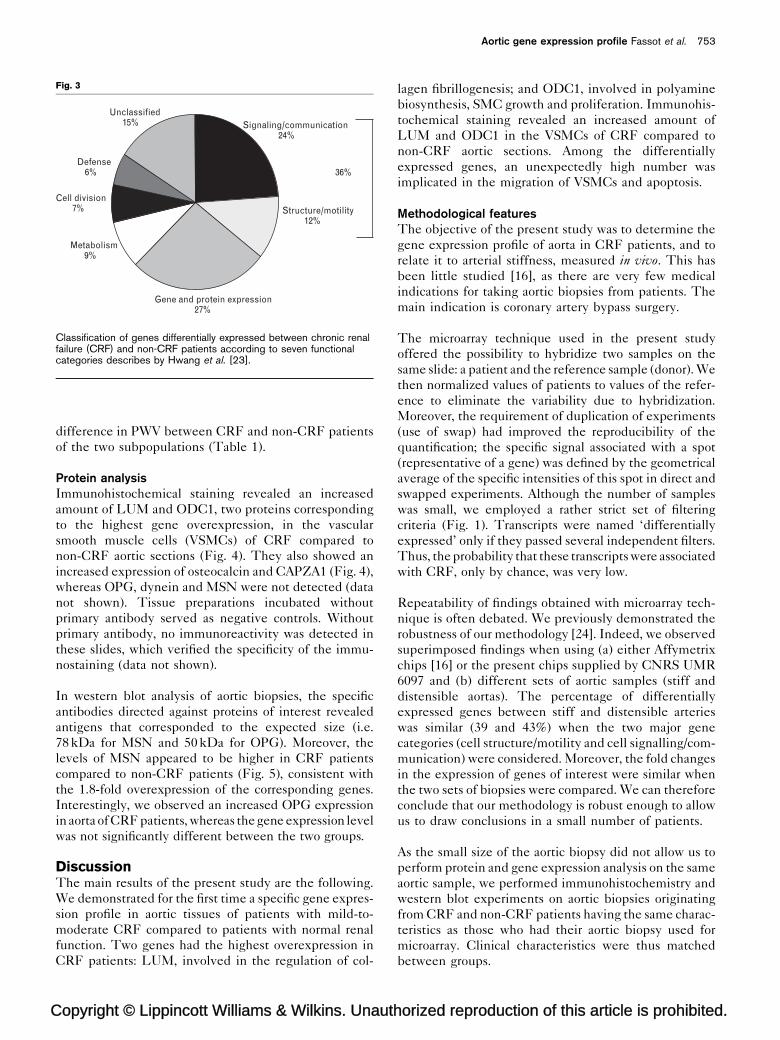
Functional classification of differentially expressedgenesWe classified the 101 transcripts differentially expressed
between groups, according to the nine functional
categories described by Hwang et al. [23] (Fig. 3). As a
group, transcripts encoding proteins potentially involved
in the mechanical regulation of vascular function (‘sig-
opyright © Lippincott Williams & Wilkins. Unautho
Fig. 2
Level of expression of genes selected for their implication in matrix, migrati
nalling/communication’ and ‘structure/motility’) consti-
tuted 36% of the annotated transcripts.
Association with arterial stiffeningAccording to the CRF status and aortic stiffness, there
was no interaction between the genes differentially
expressed. Moreover, we did not find any significant
rized reproduction of this article is prohibited.
on of smooth muscle cells (SMCs), cytoskeleton and apoptosis.
C
Aortic gene expression profile Fassot et al. 753
Fig. 3
Gene and protein expression27%
Signaling/communication24%
Structure/motility12%
Metabolism9%
Cell division7%
Defense6%
Unclassified15%
36%
Classification of genes differentially expressed between chronic renalfailure (CRF) and non-CRF patients according to seven functionalcategories describes by Hwang et al. [23].
difference in PWV between CRF and non-CRF patients
of the two subpopulations (Table 1).
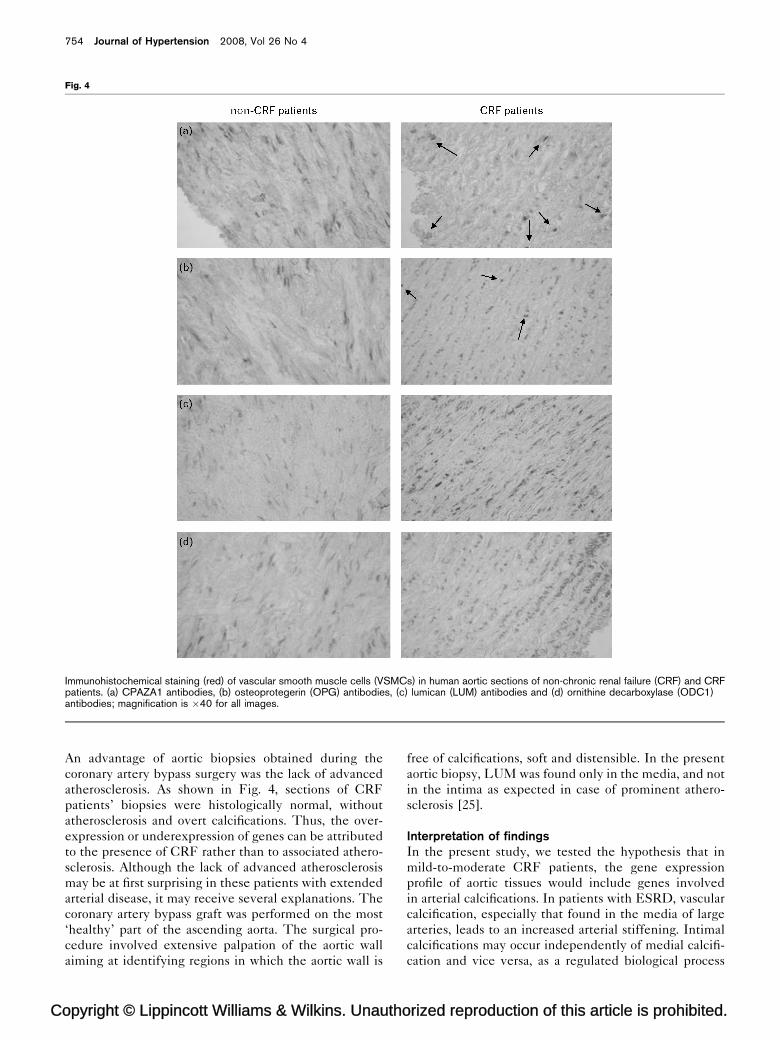
Protein analysisImmunohistochemical staining revealed an increased
amount of LUM and ODC1, two proteins corresponding
to the highest gene overexpression, in the vascular
smooth muscle cells (VSMCs) of CRF compared to
non-CRF aortic sections (Fig. 4). They also showed an
increased expression of osteocalcin and CAPZA1 (Fig. 4),
whereas OPG, dynein and MSN were not detected (data
not shown). Tissue preparations incubated without
primary antibody served as negative controls. Without
primary antibody, no immunoreactivity was detected in
these slides, which verified the specificity of the immu-
nostaining (data not shown).
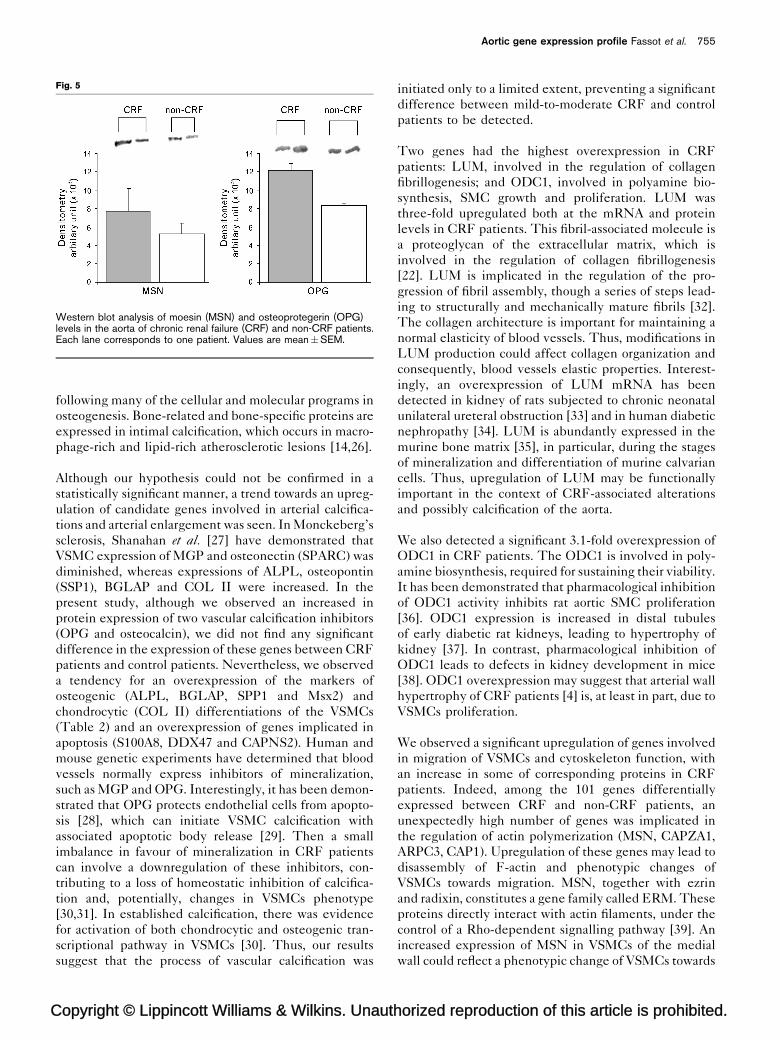
In western blot analysis of aortic biopsies, the specific
antibodies directed against proteins of interest revealed
antigens that corresponded to the expected size (i.e.
78 kDa for MSN and 50 kDa for OPG). Moreover, the
levels of MSN appeared to be higher in CRF patients
compared to non-CRF patients (Fig. 5), consistent with
the 1.8-fold overexpression of the corresponding genes.
Interestingly, we observed an increased OPG expression
in aorta of CRF patients, whereas the gene expression level
was not significantly different between the two groups.
DiscussionThe main results of the present study are the following.
We demonstrated for the first time a specific gene expres-
sion profile in aortic tissues of patients with mild-to-
moderate CRF compared to patients with normal renal
function. Two genes had the highest overexpression in
CRF patients: LUM, involved in the regulation of col-
opyright © Lippincott Williams & Wilkins. Unauth
lagen fibrillogenesis; and ODC1, involved in polyamine
biosynthesis, SMC growth and proliferation. Immunohis-
tochemical staining revealed an increased amount of
LUM and ODC1 in the VSMCs of CRF compared to
non-CRF aortic sections. Among the differentially
expressed genes, an unexpectedly high number was
implicated in the migration of VSMCs and apoptosis.
Methodological featuresThe objective of the present study was to determine the
gene expression profile of aorta in CRF patients, and to
relate it to arterial stiffness, measured in vivo. This has
been little studied [16], as there are very few medical
indications for taking aortic biopsies from patients. The
main indication is coronary artery bypass surgery.
The microarray technique used in the present study
offered the possibility to hybridize two samples on the
same slide: a patient and the reference sample (donor). We
then normalized values of patients to values of the refer-
ence to eliminate the variability due to hybridization.
Moreover, the requirement of duplication of experiments
(use of swap) had improved the reproducibility of the
quantification; the specific signal associated with a spot
(representative of a gene) was defined by the geometrical
average of the specific intensities of this spot in direct and
swapped experiments. Although the number of samples
was small, we employed a rather strict set of filtering
criteria (Fig. 1). Transcripts were named ‘differentially
expressed’ only if they passed several independent filters.
Thus, the probability that these transcripts were associated
with CRF, only by chance, was very low.
Repeatability of findings obtained with microarray tech-
nique is often debated. We previously demonstrated the
robustness of our methodology [24]. Indeed, we observed
superimposed findings when using (a) either Affymetrix
chips [16] or the present chips supplied by CNRS UMR
6097 and (b) different sets of aortic samples (stiff and
distensible aortas). The percentage of differentially
expressed genes between stiff and distensible arteries
was similar (39 and 43%) when the two major gene
categories (cell structure/motility and cell signalling/com-
munication) were considered. Moreover, the fold changes
in the expression of genes of interest were similar when
the two sets of biopsies were compared. We can therefore
conclude that our methodology is robust enough to allow
us to draw conclusions in a small number of patients.
As the small size of the aortic biopsy did not allow us to
perform protein and gene expression analysis on the same
aortic sample, we performed immunohistochemistry and
western blot experiments on aortic biopsies originating
from CRF and non-CRF patients having the same charac-
teristics as those who had their aortic biopsy used for
microarray. Clinical characteristics were thus matched
between groups.
orized reproduction of this article is prohibited.
C
754 Journal of Hypertension 2008, Vol 26 No 4
Fig. 4
Immunohistochemical staining (red) of vascular smooth muscle cells (VSMCs) in human aortic sections of non-chronic renal failure (CRF) and CRFpatients. (a) CPAZA1 antibodies, (b) osteoprotegerin (OPG) antibodies, (c) lumican (LUM) antibodies and (d) ornithine decarboxylase (ODC1)antibodies; magnification is �40 for all images.
An advantage of aortic biopsies obtained during the
coronary artery bypass surgery was the lack of advanced
atherosclerosis. As shown in Fig. 4, sections of CRF
patients’ biopsies were histologically normal, without
atherosclerosis and overt calcifications. Thus, the over-
expression or underexpression of genes can be attributed
to the presence of CRF rather than to associated athero-
sclerosis. Although the lack of advanced atherosclerosis
may be at first surprising in these patients with extended
arterial disease, it may receive several explanations. The
coronary artery bypass graft was performed on the most
‘healthy’ part of the ascending aorta. The surgical pro-
cedure involved extensive palpation of the aortic wall
aiming at identifying regions in which the aortic wall is
opyright © Lippincott Williams & Wilkins. Unautho
free of calcifications, soft and distensible. In the present
aortic biopsy, LUM was found only in the media, and not
in the intima as expected in case of prominent athero-
sclerosis [25].
Interpretation of findingsIn the present study, we tested the hypothesis that in
mild-to-moderate CRF patients, the gene expression
profile of aortic tissues would include genes involved
in arterial calcifications. In patients with ESRD, vascular
calcification, especially that found in the media of large
arteries, leads to an increased arterial stiffening. Intimal
calcifications may occur independently of medial calcifi-
cation and vice versa, as a regulated biological process
rized reproduction of this article is prohibited.
C
Aortic gene expression profile Fassot et al. 755
Fig. 5
Western blot analysis of moesin (MSN) and osteoprotegerin (OPG)levels in the aorta of chronic renal failure (CRF) and non-CRF patients.Each lane corresponds to one patient. Values are mean�SEM.
following many of the cellular and molecular programs in
osteogenesis. Bone-related and bone-specific proteins are
expressed in intimal calcification, which occurs in macro-
phage-rich and lipid-rich atherosclerotic lesions [14,26].
Although our hypothesis could not be confirmed in a
statistically significant manner, a trend towards an upreg-
ulation of candidate genes involved in arterial calcifica-
tions and arterial enlargement was seen. In Monckeberg’s
sclerosis, Shanahan et al. [27] have demonstrated that
VSMC expression of MGP and osteonectin (SPARC) was
diminished, whereas expressions of ALPL, osteopontin
(SSP1), BGLAP and COL II were increased. In the
present study, although we observed an increased in
protein expression of two vascular calcification inhibitors
(OPG and osteocalcin), we did not find any significant
difference in the expression of these genes between CRF
patients and control patients. Nevertheless, we observed
a tendency for an overexpression of the markers of
osteogenic (ALPL, BGLAP, SPP1 and Msx2) and
chondrocytic (COL II) differentiations of the VSMCs
(Table 2) and an overexpression of genes implicated in
apoptosis (S100A8, DDX47 and CAPNS2). Human and
mouse genetic experiments have determined that blood
vessels normally express inhibitors of mineralization,
such as MGP and OPG. Interestingly, it has been demon-
strated that OPG protects endothelial cells from apopto-
sis [28], which can initiate VSMC calcification with
associated apoptotic body release [29]. Then a small
imbalance in favour of mineralization in CRF patients
can involve a downregulation of these inhibitors, con-
tributing to a loss of homeostatic inhibition of calcifica-
tion and, potentially, changes in VSMCs phenotype
[30,31]. In established calcification, there was evidence
for activation of both chondrocytic and osteogenic tran-
scriptional pathway in VSMCs [30]. Thus, our results
suggest that the process of vascular calcification was
opyright © Lippincott Williams & Wilkins. Unauth
initiated only to a limited extent, preventing a significant
difference between mild-to-moderate CRF and control
patients to be detected.
Two genes had the highest overexpression in CRF
patients: LUM, involved in the regulation of collagen
fibrillogenesis; and ODC1, involved in polyamine bio-
synthesis, SMC growth and proliferation. LUM was
three-fold upregulated both at the mRNA and protein
levels in CRF patients. This fibril-associated molecule is
a proteoglycan of the extracellular matrix, which is
involved in the regulation of collagen fibrillogenesis
[22]. LUM is implicated in the regulation of the pro-
gression of fibril assembly, though a series of steps lead-
ing to structurally and mechanically mature fibrils [32].
The collagen architecture is important for maintaining a
normal elasticity of blood vessels. Thus, modifications in
LUM production could affect collagen organization and
consequently, blood vessels elastic properties. Interest-
ingly, an overexpression of LUM mRNA has been
detected in kidney of rats subjected to chronic neonatal
unilateral ureteral obstruction [33] and in human diabetic
nephropathy [34]. LUM is abundantly expressed in the
murine bone matrix [35], in particular, during the stages
of mineralization and differentiation of murine calvarian
cells. Thus, upregulation of LUM may be functionally
important in the context of CRF-associated alterations
and possibly calcification of the aorta.
We also detected a significant 3.1-fold overexpression of
ODC1 in CRF patients. The ODC1 is involved in poly-
amine biosynthesis, required for sustaining their viability.
It has been demonstrated that pharmacological inhibition
of ODC1 activity inhibits rat aortic SMC proliferation
[36]. ODC1 expression is increased in distal tubules
of early diabetic rat kidneys, leading to hypertrophy of
kidney [37]. In contrast, pharmacological inhibition of
ODC1 leads to defects in kidney development in mice
[38]. ODC1 overexpression may suggest that arterial wall
hypertrophy of CRF patients [4] is, at least in part, due to
VSMCs proliferation.
We observed a significant upregulation of genes involved
in migration of VSMCs and cytoskeleton function, with
an increase in some of corresponding proteins in CRF
patients. Indeed, among the 101 genes differentially
expressed between CRF and non-CRF patients, an
unexpectedly high number of genes was implicated in
the regulation of actin polymerization (MSN, CAPZA1,
ARPC3, CAP1). Upregulation of these genes may lead to
disassembly of F-actin and phenotypic changes of
VSMCs towards migration. MSN, together with ezrin
and radixin, constitutes a gene family called ERM. These
proteins directly interact with actin filaments, under the
control of a Rho-dependent signalling pathway [39]. An
increased expression of MSN in VSMCs of the medial
wall could reflect a phenotypic change of VSMCs towards
orized reproduction of this article is prohibited.

C
756 Journal of Hypertension 2008, Vol 26 No 4
migration, due to actin-filament (F-actin) disassembly
[40]. This is consistent with the fact that APOD is also
overexpressed (2.0-fold) in CRF patients. This major
component of HDL is associated with protease activity
and is required for breaking the cell–matrix attachments
in motile cells. An overproduction of APOD by VSMCs
may positively influence their migration [41]. As the actin
cytoskeleton is essential for maintaining the structural
and functional integrity of cells, our results suggest
that phenotypic changes towards migration and spatial
reorganization of aortic VSMCs occur at the early stage of
CRF. Indeed, in nephrectomized rats, VSMCs of the
aorta showed alteration of cell shape, orientation and
spatial arrangement of matrix, independently of blood
pressure [42]. That three genes involved in apoptosis
(S100A8, DDX47 and CAPNS2) were overexpressed in
CRF patients suggests an associated apoptosis. Both
VSMCs migration and apoptosis may contribute to the
arterial enlargement observed in CRF patients.
At the early stage of CRF, phenotypic changes of VSMCs
may favour the spatial reorganization of the arterial wall
towards lumen enlargement, in parallel with the initiation
of vascular calcification. Lumen enlargement, by loading
the stiff components of the arterial wall, in conjunction
with early vascular calcification could stiffen the arterial
wall.
In addition, the disorganization and abnormal distri-
bution of F-actin may play a role in the regulation of
vascular tone. In postischemic acute renal failure, the
disorganization of the actin cytoskeleton in VSMCs of
the renal vasculature may play a contributory role in the
increased vascular tone, impaired autoregulation of renal
blood flow and aberrant vascular reactivity [43].
Study limitationsSome study limitations should be discussed. First, the
present study included only a small number of patients
and aortic samples. This is due to the difficulty of
obtaining both aortic biopsies and noninvasive arterial
measurements before the bypass surgery, then collecting
enough aortic tissue despite the limited size of the
aortic biopsy, and finally extracting enough high-quality
RNA.
Second, by contrast to previous findings [16], aortic
stiffness was not increased in CRF compared to non-
CRF patients in the present study. This is very likely due
to the additional damage of the arterial wall in these
patients at high cardiovascular risk and scheduled for a
bypass surgery. Indeed, in patients with mild renal dys-
function and less numerous cardiovascular risk factors
[16], aortic stiffness was significantly elevated, although
mildly, by comparison with CRF hypertensives. The
patients included in the present study had a higher cf-
PWV than those of our former study [16], despite less
opyright © Lippincott Williams & Wilkins. Unautho
advanced renal dysfunction. Multivariate analyses of the
determinants of aortic stiffness [16,44] showed that GFR
had only a weak independent influence on aortic stiff-
ness. It is thus likely that CRF significantly influences
aortic stiffness only during the final stage of the disease,
whereas associated cardiovascular risk factors, leading to
atherosclerosis and coronary heart disease, are earlier
important determinants of aortic stiffness [1]. Finally,
because the entire group has very likely elevated aortic
stiffness as a result of their atherosclerosis, data of Table 1
suggest that there was widespread aortic disease in all
groups.
Third, antihypertensive agents have been implicated
in the control of vascular calcification and remodelling
[45]. Particularly, calcium channel blockers have been
reported to slow the progression of coronary calcification
in hypertensive patients [46]. In a novel model of isolated
systolic hypertension caused by arterial calcifications,
Dao et al. [47] and Essalihi et al. [48] have demonstrated
that a treatment with an ET-1 antagonist or an angio-
tensin II blocker prevents arterial calcifications and may
cause regression of vascular calcification when they are
established. It is unlikely that antihypertensive drugs
had masked any significant increase in expression of
genes involved in vascular calcification, as only two
and three patients among CRF patients and controls,
respectively, had antihypertensive medications. Never-
theless, it should be kept in mind that lipid-lowering
therapy and antihypertensive drugs, which can delay
the progression or lead to regression of atherosclerotic
disease, can have an impact on the genetic translation of
the different factors involved in these biochemical
processes.
In conclusion, these results show that patients at an early
stage of CRF have a specific gene expression profile of
aortic tissue and suggest that genes implicated in collagen
fibrillogenesis and VSMCs migration and proliferation
may play a role, particularly LUM and ODC1.
AcknowledgementThe present study was supported by grants from the
‘Fondation de France’, INSERM (Institut National de la
Sante et de la Recherche Medicale) and ANR-05-PCOD-
004-0 (Agence Nationale de la Recherche).
There are no conflicts of interest.
References1 Laurent S, Cockcroft J, Van Bortel L, Boutouyrie P, Giannattasio C, Hayoz
D, et al. Expert consensus document on arterial stiffness: methodologicalissues and clinical applications. Eur Heart J 2006; 27:2588–2605.
2 Laurent S, Boutouyrie P, Lacolley P. Structural and genetic bases of arterialstiffness. Hypertension 2005; 45:1050–1055.
3 Blacher J, Guerin AP, Pannier B, Marchais SJ, London GM. Arterialcalcifications, arterial stiffness, and cardiovascular risk in end-stage renaldisease. Hypertension 2001; 38:938–942.
rized reproduction of this article is prohibited.

C
Aortic gene expression profile Fassot et al. 757
4 Briet M, Bozec E, Laurent S, Fassot C, London G, Jacquot C, et al. Arterialstiffness and enlargement in mild to moderate chronic kidney disease.Kidney Int 2006; 69:350–357.
5 Zieman SJ, Melenovsky V, Kass DA. Mechanisms, pathophysiology, andtherapy of arterial stiffness. Arterioscler Thromb Vasc Biol 2005; 25:932–943.
6 Demer LL, Tintut Y, Parhami F. Novel mechanisms in accelerated vascularcalcification in renal disease patients. Curr Opin Nephrol Hypertens 2002;11:437–443.
7 London GM, Guerin AP, Marchais SJ, Metivier F, Pannier B, Adda H.Arterial media calcification in end-stage renal disease: impact on all-causeand cardiovascular mortality. Nephrol Dial Transplant 2003; 18:1731–1740.
8 Go AS, Chertow GM, Fan D, McCulloch CE, Hsu CY. Chronic kidneydisease and the risks of death, cardiovascular events, and hospitalization.N Engl J Med 2004; 351:1296–1305.
9 Morris ST, McMurray JJ, Rodger RS, Jardine AG. Impaired endothelium-dependent vasodilatation in uraemia. Nephrol Dial Transplant 2000;15:1194–1200.
10 Modlinger PS, Wilcox CS, Aslam S. Nitric oxide, oxidative stress, andprogression of chronic renal failure. Semin Nephrol 2004; 24:354–365.
11 Klahr S. The role of nitric oxide in hypertension and renal diseaseprogression. Nephrol Dial Transplant 2001; 16 (Suppl 1):60–62.
12 Muntner P, Hamm LL, Kusek JW, Chen J, Whelton PK, He J, et al. Theprevalence of nontraditional risk factors for coronary heart disease inpatients with chronic kidney disease. Ann Intern Med 2004; 140:9–17.
13 Shlipak MG, Fried LF, Crump C, Bleyer AJ, Manolio TA, Tracy RP, et al.Elevations of inflammatory and procoagulant biomarkers in elderly personswith renal insufficiency. Circulation 2003; 107:87–92.
14 Shanahan CM, Cary NR, Metcalfe JC, Weissberg PL. High expression ofgenes for calcification-regulating proteins in human atheroscleroticplaques. J Clin Invest 1994; 93:2393–2402.
15 Guerin AP, London GM, Marchais SJ, Metivier F. Arterial stiffening andvascular calcifications in end-stage renal disease. Nephrol Dial Transplant2000; 15:1014–1021.
16 Durier S, Fassot C, Laurent S, Boutouyrie P, Couetil JP, Fine E, et al.Physiological genomics of human arteries: quantitative relationshipbetween gene expression and arterial stiffness. Circulation 2003;108:1845–1851.
17 Levey AS, Bosch JP, Lewis JB, Greene T, Rogers N, Roth D. A moreaccurate method to estimate glomerular filtration rate from serumcreatinine: a new prediction equation. Modification of Diet in Renal DiseaseStudy Group. Ann Intern Med 1999; 130:461–470.
18 Boutouyrie P, Tropeano AI, Asmar R, Gautier I, Benetos A, Lacolley P,et al. Aortic stiffness is an independent predictor of primary coronary eventsin hypertensive patients: a longitudinal study. Hypertension 2002; 39:10–15.
19 Moreilhon C, Gras D, Hologne C, Bajolet O, Cottrez F, Magnone V, et al.Live Staphylococcus aureus and bacterial soluble factors induce differenttranscriptional responses in human airway cells. Physiol Genomics 2005;20:244–255.
20 Kerr MK, Churchill GA. Experimental design for gene expressionmicroarrays. Biostatistics 2001; 2:183–201.
21 Bradford MM. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein–dyebinding. Anal Biochem 1976; 72:248–254.
22 Chakravarti S, Magnuson T, Lass JH, Jepsen KJ, LaMantia C, Carroll H.Lumican regulates collagen fibril assembly: skin fragility and corneal opacityin the absence of lumican. J Cell Biol 1998; 141:1277–1286.
23 Hwang DM, Dempsey AA, Wang RX, Rezvani M, Barrans JD, Dai KS, et al.A genome-based resource for molecular cardiovascular medicine: toward acompendium of cardiovascular genes. Circulation 1997; 96:4146–4203.
24 Laurent S, Fassot C, Lacolley P, Boutouyrie P. Molecular determinants ofarterial stiffness. Artery Res 2007; 1:26–31.
25 Onda M, Ishiwata T, Kawahara K, Wang R, Naito Z, Sugisaki Y. Expressionof lumican in thickened intima and smooth muscle cells in human coronaryatherosclerosis. Exp Mol Pathol 2002; 72:142–149.
26 Bostrom K, Watson KE, Horn S, Wortham C, Herman IM, Demer LL. Bonemorphogenetic protein expression in human atherosclerotic lesions. J ClinInvest 1993; 91:1800–1809.
27 Shanahan CM, Cary NR, Salisbury JR, Proudfoot D, Weissberg PL,Edmonds ME. Medial localization of mineralization-regulating proteins inassociation with Monckeberg’s sclerosis: evidence for smooth musclecell-mediated vascular calcification. Circulation 1999; 100:2168–2176.
28 Malyankar UM, Scatena M, Suchland KL, Yun TJ, Clark EA, Giachelli CM.Osteoprotegerin is an alpha vbeta 3-induced, NF-kappa B-dependentsurvival factor for endothelial cells. J Biol Chem 2000; 275:20959–20962.
opyright © Lippincott Williams & Wilkins. Unauth
29 Proudfoot D, Skepper JN, Hegyi L, Bennett MR, Shanahan CM, WeissbergPL. Apoptosis regulates human vascular calcification in vitro: evidence forinitiation of vascular calcification by apoptotic bodies. Circ Res 2000;87:1055–1062.
30 Tyson KL, Reynolds JL, McNair R, Zhang Q, Weissberg PL, Shanahan CM.Osteo/chondrocytic transcription factors and their target genes exhibitdistinct patterns of expression in human arterial calcification. ArteriosclerThromb Vasc Biol 2003; 23:489–494.
31 Luo G, Ducy P, McKee MD, Pinero GJ, Loyer E, Behringer RR, et al.Spontaneous calcification of arteries and cartilage in mice lacking matrixGLA protein. Nature 1997; 386:78–81.
32 Qin H, Ishiwata T, Asano G. Effects of the extracellular matrix on lumicanexpression in rat aortic smooth muscle cells in vitro. J Pathol 2001;195:604–608.
33 Silverstein DM, Travis BR, Thornhill BA, Schurr JS, Kolls JK, Leung JC, et al.Altered expression of immune modulator and structural genes in neonatalunilateral ureteral obstruction. Kidney Int 2003; 64:25–35.
34 Schaefer L, Raslik I, Grone HJ, Schonherr E, Macakova K, Ugorcakova J,et al. Small proteoglycans in human diabetic nephropathy: discrepancybetween glomerular expression and protein accumulation of decorin,biglycan, lumican, and fibromodulin. FASEB J 2001; 15:559–561.
35 Raouf A, Ganss B, McMahon C, Vary C, Roughley PJ, Seth A, et al. Lumicanis a major proteoglycan component of the bone matrix. Matrix Biol 2002;21:361–367.
36 Ignarro LJ, Buga GM, Wei LH, Bauer PM, Wu G, del Soldato P, et al. Role ofthe arginine–nitric oxide pathway in the regulation of vascular smoothmuscle cell proliferation. Proc Natl Acad Sci U S A 2001; 98:4202–4208.
37 Deng A, Munger KA, Valdivielso JM, Satriano J, Lortie M, Blantz RC, et al.Increased expression of ornithine decarboxylase in distal tubules of earlydiabetic rat kidneys: are polyamines paracrine hypertrophic factors?Diabetes 2003; 52:1235–1239.
38 Loikkanen I, Lin Y, Railo A, Pajunen A, Vainio S. Polyamines are involved inmurine kidney development controlling expression of c-ret, E-cadherin, andPax2/8 genes. Differentiation 2005; 73:303–312.
39 Amieva MR, Furthmayr H. Subcellular localization of moesin in dynamicfilopodia, retraction fibers, and other structures involved in substrateexploration, attachment, and cell–cell contacts. Exp Cell Res 1995;219:180–196.
40 Blindt R, Zeiffer U, Krott N, Filzmaier K, Voss M, Hanrath P, et al.Upregulation of the cytoskeletal-associated protein moesin in the neointimaof coronary arteries after balloon angioplasty: a new marker of smoothmuscle cell migration? Cardiovasc Res 2002; 54:630–639.
41 Leung WC, Lawrie A, Demaries S, Massaeli H, Burry A, Yablonsky S, et al.Apolipoprotein D and platelet-derived growth factor-BB synergismmediates vascular smooth muscle cell migration. Circ Res 2004; 95:179–186.
42 Amann K, Neususs R, Ritz E, Irzyniec T, Wiest G, Mall G. Changes ofvascular architecture independent of blood pressure in experimentaluremia. Am J Hypertens 1995; 8:409–417.
43 Kwon O, Phillips CL, Molitoris BA. Ischemia induces alterations in actinfilaments in renal vascular smooth muscle cells. Am J Physiol Renal Physiol2002; 282:F1012–F1019.
44 Hermans MM, Henry R, Dekker JM, Kooman JP, Kostense PJ, Nijpels G,et al. Estimated glomerular filtration rate and urinary albumin excretion areindependently associated with greater arterial stiffness: the hoorn study.J Am Soc Nephrol 2007; 18:1942–1952.
45 Giachelli CM. Vascular calcification mechanisms. J Am Soc Nephrol 2004;15:2959–2964.
46 Motro M, Shemesh J. Calcium channel blocker nifedipine slows downprogression of coronary calcification in hypertensive patients comparedwith diuretics. Hypertension 2001; 37:1410–1413.
47 Dao HH, Essalihi R, Graillon JF, Lariviere R, De Champlain J, Moreau P.Pharmacological prevention and regression of arterial remodeling in a ratmodel of isolated systolic hypertension. J Hypertens 2002; 20:1597–1606.
48 Essalihi R, Dao HH, Yamaguchi N, Moreau P. A new model of isolatedsystolic hypertension induced by chronic warfarin and vitamin K1 treatment.Am J Hypertens 2003; 16:103–110.
orized reproduction of this article is prohibited.




















