
Aboveground biomass allocation, leaf growth, and photosynthesis patterns in tundra plant forms in...
15
Oecologia (Berl.) 24, 159-173 (1976) Oecologia 9 by Springer-Verlag 1976 Aboveground Biomass Allocation, Leaf Growth, and Photosynthesis Patterns in Tundra Plant Forms in Arctic Alaska Douglas A. Johnson* and Larry L. Tieszen Department of Biology,Augustana College, Sioux Falls, South Dakota 57102, USA Summary. Tundra plant growth forms can generally be characterized as consisting predominantly of low-growing perennial grasses and sedges, peren- nial herbaceous forbs, dwarf deciduous shrubs, and dwarf evergreen shrubs. Gross aboveground carbon allocation, leaf growth, and photosynthesis pat- tern studies were initiated to develop a quantitative understanding of the functional importance of these particular tundra growth forms. Photosynthet- ic capacities of 13 species were determined under standardized exposure conditions using a 14CO2 field system and ranged between 5 and 47 mg CO2.g dry wt-~.h -1. These results, in conjunction with detailed leaf growth determinations, support the generalization that species with an ever- green growth form have lower photosynthetic capacities than species with a perennial graminoid, forb, or deciduous shrub growth form. However, these low photosynthetic capacities in evergreen shrubs are associated with relatively extended leaf longevities. Conversely, deciduous shrub forms exhib- ited high photosynthetic capacities, but were offset by relatively short leaf longevity periods. The perennial grasses, sedges, and forbs showed patterns intermediate to these. As a result, it appears that among tundra species of different growth form, photosynthetic capacity is inversely related to leaf longevity. Introduction The composition and structure of plant communities through evolutionary time are dependent upon the action of abiotic and biotic variables on the available biological pool. In arctic Alaska these interactions are expressed as: 1) commu- nities in which perenniality and vegetative reproduction predominate (Billings and Mooney, 1968; Johnson and Tieszen, 1973), 2) a decrease in numbers * Present address and address for offprint requests: Crops Research Laboratory, Utah State University, UMC 63, Logan, Utah 84322, USA
Transcript of Aboveground biomass allocation, leaf growth, and photosynthesis patterns in tundra plant forms in...

Oecologia (Berl.) 24, 159-173 (1976) Oecologia �9 by Springer-Verlag 1976
Aboveground Biomass Allocation, Leaf Growth, and Photosynthesis Patterns in Tundra Plant Forms in Arctic Alaska
Douglas A. Johnson* and Larry L. Tieszen Department of Biology, Augustana College, Sioux Falls, South Dakota 57102, USA
Summary. Tundra plant growth forms can generally be characterized as consisting predominantly of low-growing perennial grasses and sedges, peren- nial herbaceous forbs, dwarf deciduous shrubs, and dwarf evergreen shrubs. Gross aboveground carbon allocation, leaf growth, and photosynthesis pat- tern studies were initiated to develop a quantitative understanding of the functional importance of these particular tundra growth forms. Photosynthet- ic capacities of 13 species were determined under standardized exposure conditions using a 14CO2 field system and ranged between 5 and 47 mg CO2.g dry wt-~.h -1. These results, in conjunction with detailed leaf growth determinations, support the generalization that species with an ever- green growth form have lower photosynthetic capacities than species with a perennial graminoid, forb, or deciduous shrub growth form. However, these low photosynthetic capacities in evergreen shrubs are associated with relatively extended leaf longevities. Conversely, deciduous shrub forms exhib- ited high photosynthetic capacities, but were offset by relatively short leaf longevity periods. The perennial grasses, sedges, and forbs showed patterns intermediate to these. As a result, it appears that among tundra species of different growth form, photosynthetic capacity is inversely related to leaf longevity.
Introduction
The composition and structure of plant communities through evolutionary time are dependent upon the action of abiotic and biotic variables on the available biological pool. In arctic Alaska these interactions are expressed as: 1) commu- nities in which perenniality and vegetative reproduction predominate (Billings and Mooney, 1968; Johnson and Tieszen, 1973), 2) a decrease in numbers
* Present address and address for offprint requests: Crops Research Laboratory, Utah State University, UMC 63, Logan, Utah 84322, USA

160 D.A. Johnson and L.L. Tieszen
of species from the south [e.g., 250 species at Umiat (Britton, 1957)] to the arctic coast [e.g., 120 species in the Barrow area (Dennis and Johnson, 1970)], and 3) a decrease in community stature as the arctic coast is approached.
Even though arctic tundra areas may differ in the number and kinds of species which make up their vegetation, tundra plant growth forms can generally be characterized as consisting predominantly of low-growing perennial grasses and sedges, perennial herbaceous forbs, dwarf deciduous shrubs, and dwarf evergreen shrubs. The degree to which these differences in growth form are related to functional differences is not thoroughly documented; however, work of Bliss (1962), Hedberg (1964), and Mooney and Dunn (1970b) has suggested that these growth form distributions have functional importance and that each growth form is characterized by a particular allocation strategy which presum- ably allows successful exploitation of particular habitats.
Indeed, distributions of major growth forms have recently been quantitatively related to specific environmental complexes in tundra (Wielgolaski and Webber, 1976). In addition Lewis and Callaghan (1975) and Tieszen and Wieland (1975) have described distinct growth strategies of meadow grasses, sedges, and fellfield plants. These specific growth form strategies likely involve allocation differences for carbon and nutrients (Mooney and Dunn, 1970b; Chapin et al., 1975; Mooney et al., 1975). The allocation patterns of these important plant resources to growth, storage, maintenance, and protection likely represent the means by which plants meet the demands of their environment (Mooney and Chu, 1974) and suggest differences in physiological processes and primary production.
This present study was initiated to develop a quantitative understanding of the relationships between gross aboveground carbon allocation, leaf growth, and photosynthesis patterns in different tundra growth forms.
Methods
The research was conducted at the Meade River Field Station (70~ 157~ 15 m elev.) of the Naval Arctic Research Laboratory located on the Meade River on the Alaskan North Slope and is approximately 110 km SSW of Barrow, Alaska. Climate at Meade River is somewhat more continental than the strongly marit ime dominated climate at Barrow. For example, the 1975 summer mean wind velocity and air temperature at 1 m was 11.4 km .h ~ and 5 ~ C, respectively, while total precipitation during this same period was 80 ram. A detailed geological as well as pedological description of the area can be found in Rickert and Tedrow (1967). The vegetational communit ies of the area have been described by Tieszen and Johnson (1968) and generally consist of low perennial grasses, sedges, shrubs, and herbs. Plant nomenclature follows Hultbn (1968).
For the leaf growth study, 20 plants each of Salix pulchra, Betula nana, Eriophorum vaginatum, and Ledum palustre were selected in June and followed during leaf elongation and senescence throughout the growing season. For Salix and Betula, both deciduous shrub species, leaf groups produced at a given node were followed with as many as 6 leaves produced per leaf group. For these species leaves within a leaf group were consecutively numbered from the most basal to the most distal leaf within the leaf group. Eriophorum, a tussock graminoid, consists of collections of individual tillers comprising a tussock. Individual tillers produced as many as 5 leaves during the growing season. Leaves in Eriophorum were consecutively numbered from the first green leaf to emerge and thus the oldest living leaf on the plant to the last leaf to emerge. Because of
-the evergreen growth form of Ledum, leaves of different age classes were followed throughout the growing season.

Allocation, Leaf Growth, and Photosynthesis in Tundra Plants 161
Individual leaf maps of each plant allowed the same leaf to be followed throughout the growing season. Leaf length was measured from the point of petiole attachment to the leaf blade, or in the case of Eriophorum from the base of the plant, to the most distal green portion of the outstretched leaf. Leaf area to length and leaf dry weight to area observations using similar leaf material from the same vicinity as the selected plants were collected three times during the growing season and consisted of 200 observations for each species, except for Salix which consisted of 400 observations. Regression equations were determined for each species and enabled leaf lengths to be converted to their respective leaf dry weights.
Leaf photosynthetic measurements were determined using a portable 1~CO2 field system. The field system and procedure have been described by Tieszen et al. (1974). The field system consists of a plexiglass leaf chamber which is temperature controlled by a Peltier thermoelectric stage. The procedure involves exposing an intact leaf to a t4COz air mixture, immediately cooling the leaf sample in the field following exposure, drying the exposed leaf sample, combusting this sample, and radioactivity counting using liquid scintillatio n techniques.
According to studies by Ludwig and Canvin (1971), net 14CO2 uptake in a leaf is maximum at exposure periods less than 30 s. This initial period may approximate gross CO2 influx. After 30 s, net CO2 uptake decreases as the exposure time in 14CO2 is extended and after 10 min approaches normal net CO 2 exchange rates. This decrease in 14CO2 uptake is associated with a4cQ2 evolution from the leaf. Thus, with exposure times of 1 min used in this study, COz uptake measured was likely closer to gross CO2 uptake than net photosynthesis.
Air temperatures within the leaf exposure chamber were measured using a shielded copper- constantan fine wire thermocouple. Leaf temperatures were also monitored and were within 1 ~ C of chamber air temperatures. Artificial irradiation was provided by a high intensity incandescent aircraft landing lamp (100 W) and was monitored with a Lambda Co. Model LI-190S quantum sensor. In addition, the incoming chamber air stream was bubbled through water which yielded a high relative humidity within the plexiglass leaf chamber. A period of 5 min was allowed for the leaf to attain a steady state gas exchange rate before 14CO2 exposure. Only leaves which were the longest, most recently fully expanded on the plant were used during these field experiments. Areas of the exposed leaf samples were outlined on paper with subsequent leaf area (one side) determinations made in the laboratory using a Lambda Model LI-3000 portable area meter.
Results
As an indica t ion o f a b o v e g r o u n d ca rbon ut i l izat ion by the ma jo r species present
in the Meade River C a m p area, dry weight analyses o f individual aboveg round
plant par t componen t s were ob ta ined at peak season and are presented in
Figure 1. These results for Betula nana, Salix pulchra, and Ledum palustre repre-
sent p lant par t dry weights for an average a b o v e g r o u n d individual or in the
case o f Eriophorum vaginatum for an individual tiller. A n aboveg round individual
was def ined in this study as all aboveg round biomass issued f rom a stem which
goes be lowground . A l t h o u g h a b o v e g r o u n d individuals may have been intercon-
nected be lowground , each aboveg round shoot issued f rom a stem which goes
be lowground was considered an individual . Even though non-cur ren t stems
compr i sed the largest s tanding aboveg round b iomass c o m p a r t m e n t for Betula, Salix, and Ledurn, the bulk o f current aboveg round p roduc t ion was compr ised
o f leaf p roduc t ion as was the case for Eriophorum. Values presented in Figure 1 would likely vary f r o m year to year depending on env i ronmenta l condit ions.
Lea f length measurements o f individual leaf posi t ions were taken th roughou t
the 1975 growing season for these same tundra species. These leaf lengths were
conver ted to their respective areas using regression equat ions presented in Table
1. These regression relat ionships were significant at the 0.995 level and correla-

162
1.2
D.A. Johnson and L.L. Tieszen
Bet ula nano
(Q)
1.2
Q8
O.4
&
Solix pulchra
Eriophorum Voginotum
(e)
Ledum polustre
(d)
Q2
0.1
Current Current Lost Lost 2 -6 2 -I0 > I0 Fruits Leaves Stems Year Year Year Year Year
Leaves Stems Leaves Stems Stems
Fig. l a -d , Average dry weight biomass allocation for different plant compartments of an above- ground individual for Betula nana (a), Salix pulchra (b), Eriophorum vaginatum (c), and Ledum palustre (d). Average compar tment allocations were estimated from a sample of 40 aboveground individuals for Betula, Salix, and Ledum and 50 tillers for Eriophorum. Determinations were obtained 21-26 July
Table 1. Leaf area to leaf length regression relationships for Betula nana, Eriophorum vaginatum, Ledum palustre, and Salix pulehra
Species Equation r F
Betula loge(Area) = - 0 . 3 3 8 + 1 . 8 5 2 [loge(length)] 0.94 1,946"* Eriophorum loge(Area) = - 2 . 4 6 6 + 1 . 0 6 5 [loge(length)] 0.90 1,513"* Ledum loge(Area) = - 1.856 + 1.621 [loge(length)] 0.79 521 ** Salix loge(Area) = - 1.413 + 1.811 [loge(length)] 0.94 4,042 **
** All regression relations are significant at P~0.995 and were determined using 200 observations for Betula, Eriophorum, and Ledum and 400 observations for Salix

Allocation, Leaf Growth, and Photosynthesis in Tundra Plants 163
s
�9 Salix
~3 / '~ / Eriophorum
/ x �9 �9 _~_~ , Ledum x ~ /
I = ~ m �9 x
Betula i I i x l
o 3'0 2'0 30 9 ,9 29 June July August
Fig. 2. Mean leaf dry weight seasonal progressions for Salix pulchra, Eriophorum vaginatum, Ledum palustre, and Betula nana with 220, 77, 140, and 240 leaves, respectively, followed for leaf length throughout the growing season. Dry weights were derived from these seasonal leaf length measure- ments using leaf area to length and leaf dry weight to area relationships determined three times during the growing season
tion coefficients, r, were 0.94, 0.90, 0.79, and 0.94 for Betula, Eriophorum, Ledum, and Salix, respectively. Leaf dry weights were then estimated using leaf dry weight and area relationships determined three times during the growing season for each species.
Seasonal progressions of mean leaf dry weights for each species are presented in Figure 2. More detailed individual leaf growth dynamics depicted in Figures 3 and 4 show that Betula and Salix, deciduous shrubs, produce a sudden burst of leaf material; whereas, Eriophorum, a tussock cottongrass, exhibits a sequen- tial pattern of leaf development. Ledum, an evergreen shrub species, has leaves which persist for more than one growing season. Estimates of leaf mortality for Ledum suggest that 70% of leaves initiated the same season have abscised two growing seasons after their initiation and that over 96% have abscised after four growing seasons.
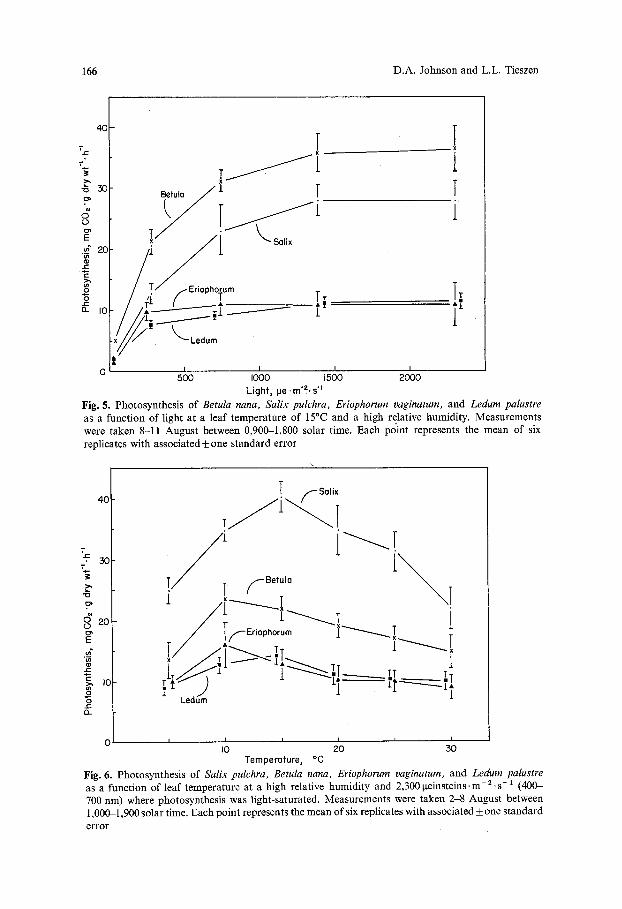
The response of CO2 uptake to light and temperature was also investigated for these four tundra species (Figs. 5 and 6). These relationships were determined the 1st week in August and even though some degree of senescence was present, these relative responses may still be representative of responses earlier in the growing season. Photosynthesis was light saturated in Salix, Betula, and Ledum at 1,380 ~teinsteins-m- 2. s- 1 or approximately 60% of maximum solar noon intensity measured directly into the solar beam at Meade River, while Eriopho- rum became light saturated at 740 ge- m- 2 S- 1 Temperature optimum for Salix and Ledum was at 15~ while optimum for Betula and Eriophorum was 10~

164 D.A. Johnson and L.L. Tieszen
12
Solix pulchra
IO (o)
8
6
4
2
f o i : n
" ' I0
3 8
6
4
2
0
. , / ~ e ~ Leaf
A ~ Position
X / x
i X 2
, i I f I s I I I I ~ 1
,./ . . . . . 0,/", , ,
20 30 I 0 20 50 9 19 29 June July August
Fig. 3a and b. Leaf dry weight seasonal progressions for different leaf positions for Salix pulchra (a) and Eriophorum vaginatum (b). For Salix and Eriophorum 220 and 77 leaves, respectively. were followed for leaf length throughout the growing season. Dry weights were derived from these seasonal leaf length measurements using leaf area to length and leaf dry weight to area relationships three times during the growing season
Photosynthetic capacities for a number of tundra species are presented in Table 2. These determinations were obtained using a standardized irradiation and temperature of 2,300 ge .m- 1. s- 1 and 15~ Data from Figure 5 as well as work of Mooney and Billings (1961) and Tieszen (1973) suggest that photosyn- thesis at this light intensity was likely saturated in all species. However, since temperature optima in Figure 6 were between 10-15~ and since 10-20~ optima have been found for other tundra species (Tieszen, 1973; Gerasimenko and Zalenskiy, 1974; Tieszen and Wieland, 1975), 15~ was not likely the tempera- ture optimum for all species examined.
Leaf photosynthetic capacity for Ledum was highest in one year old leaves and declined in capacity with increasing age. For Vaccinium highest photosyn- thetic capacities were exhibited in current leaves with characteristically lower, but constant photosynthesis through 3-year old leaves. Additionally, photosyn-

Allocation, Leaf Growth, and Photosynthesis in Tundra Plants 165
..c
@
E3
-g ,_1
Leaf Type Ledurn palustre . Current x Last Year (a) �9 2 Year Old �9 3, Year Old
x_ .x .x .x. _ _
/ i , t i/ i i i i
Leaf Position �9 I
x 2 �9 3
A 4
o 5 a 6
20 ' June
iX&
I I
. IXA
i i i i i i
Betula nana
(b)
i a ---"~"L h I e i t e i e e
30 I0 20 30 9 19 29 duly August
Fig. 4a and b. Leaf dry weight seasonal progression for different leaf types for Ledum palustre (a) and different leaf positions for Betula nana (b). For Ledum and Betula 140 and 240 leaves, respectively, were followed for leaf length throughout the growing season. Dry weights were derived from these seasonal leaf length measurements using leaf area to length and leaf dry weight to area relationships determined three times during the growing season
thetic capacity appears quite consistent within a growth form. The highest photosynthetic capacities were found in deciduous shrubs and grasses and sedges. Evergreen shrub species typically exhibited the lowest photosynthetic capacities with perennia ! forbs having intermediate CO2 uptake rates.
Estimates of leaf growth phase duration, leaf longevity, and photosynthetic capacity for typical leaves of different growth forms are presented in Table 3. Photosynthetic capacities were highest for species with shortest periods of leaf longevity. For example, Betula and Salix, both deciduous shrubs, have leaf longevities of 61 and 64 days, respectively, and exhibited CO2 uptake rates of 37 and 40mg CO2.g dry w t - l . h -1, respectively. Conversely, species with typically long leaf longevities had leaves with relatively low photosynthetic capac- ities. For example, Ledum and Vaccinium, evergreen shrubs, have leaf longevity periods of 200 and 360 days, respectively, and exhibited CO2 uptake capacities of 14 and 6mg CO2.g dry w t - l . h -1, respectively. Carex and Eriophorum, both sedges, have intermediate leaf longevities and photosynthetic capacities.
166 D.A. Johnson and L.L. Tieszen
40
c ~ o
0
20
*E
o
o_ I0
! j J ~ Betula / I
i'/i / / / k,..._ Ledum
I I I 500 I000 1500
Light, pe .m'2.s "l
l , , - x
T i
I I I
2OOO
Fig. 5. Photosynthesis of Betula nana, Salix pulchra, Eriophorum vaginatum, and Ledum palustre as a function of light at a leaf temperature of 15~ and a high relative humidity. Measurements were taken 8-11 August between 0,900-1,800 solar time. Each point represents the mean of six replicates with associated + one standard error
40
30 7"
o~ z0
IC o o
/~- Salix
l , , i ~ I .
/ I / ' ' - Eriophorum J ' ~ ! "
T / /-: ~'~-<'.T ;~!
Z a. LeduJmm i " I i
I I I I I I
I0 20 30 Temperature, ~
Fig. 6. Photosynthesis of Salix pulchra, Betula nana, Eriophorum vaginatum, and Ledum palustre as a function of leaf temperature at a high relative humidity and 2,300 i~einsteins, m -2 . s -1 (400- 700 nm) where photosynthesis was light-saturated. Measurements were taken 2-8 August between 1,000-1,900 solar time. Each point represents the mean of six replicates with associated _+ one standard error

Allocation, Leaf Growth, and Photosynthesis in Tundra Plants 167
Table 2. Photosynthetic capacities of species within four main growth forms. These capacities represent the means with associated standard errors of 12-16 replicates using most fully expanded leaves and were conducted at leaf temperatures of 15 ~ C, high humidity, and saturating light. For Arctagrostis, Petasites, and Polygonum means are of 6 replicates
Species Photosynthetic capacity (mg CO2.g dry wt -1 .h-1)
Deciduous shrubs: 1. Betula nana 37 _+ 4 2. Salix pulchra 40 _+ 3
Grasses and sedges: 1. Arctagrostis latifolia 47 _+ 7 2. Carex aquatilis 34 _+ 3 3. Carex bigelowii 37 _+ 2 4. Eriophorum vaginatum 27 _+ 2
Perennial forbs: 1. Petasites frigidus 21 +_ 2 2. Polygonum viviparum 29 +_ 3 3. Rubus chamaemorus 15_+ 1
Evergreen shrubs: 1. Cassiope tetragona
Current leaves 9 _+ 1 Noncurrent leaves 10 _+ 1
2. Dryas integrifolia Current leaves 10_+ 1
3. Ledum palustre Current leaves 10 • 1 1-year old leaves 18 +_ 1 2-year old leaves 17_+ 1 3-year old leaves 11 _+ 1
4. Vaccinium vitis-idaea Current leaves 7 + 1 1-year old leaves 5 + 1 2-year old leaves 5 + 1 3-year old leaves 5 _+ 1
D i s c u s s i o n
Pa t t e rns o f c a r b o n a l loca t ion a n d u t i l i za t ion are only poo r ly k n o w n for arc t ic species, and a review by M o o n e y (1972) documen t s very litt le c a r b o n a l loca t ion da t a even for t empe ra t e speices. In the A l a s k a n arct ic , recent s tudies have he lped e luc ida te b r o a d t e m p o r a l changes in c a r b o n up t ake and a l loca t ion (Ties- zen, 1976) as well as wi th in t i l ler a l loca t ion pa t t e rns (Allessio and Tieszen, 1975). Tieszen, for example , suggests tha t in a p r e d o m i n a n t l y g r a m i n o i d c o m m u - ni ty at Bar row, A laska , the last th i rd o f the seasonal c a r b o n up t ake is used for b e l o w g r o u n d growth , s torage, a n d / o r main tenance . Similar ly, the results o f Al less io and Tieszen (1975) are beginning to d o c u m e n t the pheno logy o f

168 D.A. Johnson and L.L. Tieszen
Table 3. Duration of different growth phases for a typical leaf in arctic tundra species as estimated from seasonal leaf dry weight progressions for each species. Previous seasons refers to growing season days during prior seasons, growth phase refers to growing season days from visible exsertion to 80% maximum leaf weight, maturation phase refers to growing season days for 80% maximum leaf weight to 20% senescence, and senescence phase refers to growing season days from 20% senescence to complete senescence. Photosynthetic capacities are the means with associated standard errors of 12 determinations using most fully expanded leaves and were conducted during peak season of leaf temperatures of 15 ~ C, high humidity, and saturating light. The photosynthetic capacities for the evergreen shrubs represent the means of four age classes of leaves. Length of growth phases for Carex aquatilis is taken from Tieszen (unpublished data). Leaf longevities for Ledum and Vaccinium represent estimates taken from leaf mortality data for an average leaf
Species Length of interval (growing season days) Photosynthetic capacity
Previous Growth Matura- Senes- Leaf (mgCO2.g seasons tion cence longevity dry wt- 1. h- 1)
Deciduous shrubs: Salix pulchra - 18 34 12 64 40 + 3 Betula nana - 17 33 11 61 37+4
Grasses and sedges: Carex aquatilis ~25 17 26 17 85 34+ 3 Eriophorum vaginatum ~ 27 29 24 15 95 27 + 2
Evergreen shrubs: Ledum palustre ~ 120 80 200 14 + 1 Vaccinium vitis-idaea ~ 280 80 360 6 + 1
a l loca t ion pa t t e rns in Dupontia fischeri. However , l i t t le compara t i ve in forma- t ion on a l loca t ion in different g rowth forms is avai lable , and this s tudy ini t iates an a t t emp t to u n d e r s t a n d the signif icance o f a l loca t ion pa t t e rns in t u n d r a plants .
The peak season a b o v e g r o u n d dry weight charac te r i za t ions presented in F igure 1 show tha t for bo th the dec iduous and evergreen shrubs between 80 -90% of cur ren t a b o v e g r o u n d b iomass was compr i s ed in the leaf c o m p a r t m e n t , while 10-15% was con ta ined in cur ren t stems. Similar ly , Eriophorum vaginatum, a t und ra sedge, had 96% of cur ren t a b o v e g r o u n d b iomass con ta ined in leaves. These values o f c a r b o n c o m m i t m e n t s agree closely wi th those o f M o o n e y and Chu (1974) for a Ca l i fo rn ia c h a p p a r a l evergreen shrub where 80 -90% of cur ren t a b o v e g r o u n d p roduc t i on was con ta ined in leaves and a p p r o x i m a t e l y 10% in
stems. Add i t i ona l ly , M o o n e y and Chu (1974) es t ima ted ca rbon c o m m i t m e n t s nea r
10% in r ep roduc t ive par t s as c o m p a r e d to a p p r o x i m a t e l y 5% in these t u n d r a shrub species a n d 4 % in Eriphorum vaginatum. These rela t ively low values o f c a r b o n inves tment in r eproduc t ive par t s in these t und ra species l ikely reflects the large degree o f vege ta t iona l r ep roduc t ion inves tment exhib i ted in t u n d r a envi ronments . Bliss (1971) suggests tha t t und ra env i ronments wi th thei r general lack o f hea t in the shor t g rowing season m a y no t be conducive to sexual r epro- duc t ion which requires cons iderab le hea t inputs for ma tu r i t y and seed set.
Lea f g rowth studies have been conduc t ed in the arct ic t u n d r a by Mat the i s et al. (1976) and Tieszen a n d Johnson (unpubl i shed data) . These studies were

Allocation, Leaf Growth, and Photosynthesis in Tundra Plants 169
concerned primarily with tundra grasses and sedges and agree quite closely with the results obtained for Eriophorum vaginatum in this present study (Fig. 3). These species have a sequential pattern of leaf development where a typical leaf initiates growth and dies back the first season and then completes exsertion and development the following season. Leaves of these species characteristically exhibited a rather long growth phase, a shorter maturation phase, and a some- what extended senescence phase (Table 3).
Contrasting this sequential pattern of leaf development was the rather synchro- nous leaf growth pattern of the deciduous shrubs (Figs. 3 and 4). These tundra species, similar to their temperate counterparts, produced a sudden burst of leaf material which had a relatively rapid expansion phase followed by an extended maturation phase, and a short senescence phase (Table 3). A similar pattern of synchronous leaf growth was found in the dwarf evergreen shrub, Ledum palustre (Fig. 4). However, even though leaves for this species appear in early July, full leaf development was not attained until near the end of the first growing season. A typical leaf of Ledum had leaf longevity extending over more than two growing seasons.
In tundra plants as well as plants in other ecosystems long-term survival is strongly hinged on the ability of these plants to fix and utilize energy. This ability is in part a function of leaf photosynthetic potential which can be viewed as an upper limit attained under optimum conditions. However, this physiolog- ical potential only accounts for a portion of the total carbon-gaining capacity of the leaf and additionally depends on the existing environmental conditions to which the leaf is exposed. Nevertheless, the potential rate at which a species can fix carbon is an important factor in considering photosynthetic gain.
Even though arctic tundra areas are often remote and relatively inaccessible, a number of field studies have examined photosynthetic capacities in tundra plants (Shvetsova and Voznesenskii, 1971; Gerasimenko and Zelenskiy, 1974; Mayo et al., 1973; Tieszen, 1973, 1976; Tieszen and Johnson, 1975). The photo- synthetic capacities presented in Table 2 agree with the results of similar species from those previous studies. For example, Shvetsova and Voznesenskii (1971) reported rates between 3 and 33rag CO2"g dry w t - l . h -1 for a comparably wide range of tundra species in the Taimyr region of the USSR.
The patterns of leaf photosynthesis in different age leaf material of two dwarf evergreen shrub species, Ledum palustre and Vaccinium vitis-idaea, are particularly interesting (Table 2). Leaf photosynthetic capacity for Ledum was highest in one year old leaves and declined in capacity with increasing age. These results agree closely with Hiramatu (1939) who found photosynthesis most active in one year old leaves of two shrubby species of evergreen holly. For Vaccinium highest photosynthetic capacities were exhibited in current leaves with characteristically lower, but constant photosynthesis exhibited through 3-year old leaves. This pattern is similar to other evergreen tundra species (Hadley and Bliss, 1964; Grace and Woolhouse, 1970) and to pine, spruce, and fir species where photosynthetic capacity reached a maximum during the first season of growth and subsequently decreased with increasing age (Freeland, 1952; Clark, 1961; E1 Aouni and Mousseau, 1974).
Ludlow and Jarvis (1971) attribute photosynthetic declines in aging evergreen

170 D.A. Johnson and L.L. Tieszen
leaf material to both increased stomatal and mesophyll resistances. They suggest that increased stomatal resistance may be due to either reduced stomatal activity and/or accumulations of wax in the ante-chamber. Increased mesophyll resist- ance in older leaf material may in part be due to winter chloroplast degradation which becomes less reversible with needle age (Perry and Baldwin, 1966; E1 Aouni and Mousseau, 1974).
Terrestrial ecosystems have been described by their growth form and struc- ture or physiognomy since Raunkier (1934). This description is based on the recognition of a variety of growth form categories into which plants are grouped regardless of taxonomic status. Bliss (1962) stated that the growth form spectrum of a flora can be used as a rough measure of the prevailing climate and the selective action for the climate on those plant forms best adapted for survival. Indeed, Mooney and Dunn (1970a) have shown qualitatively and Parsons and Moldenke (1975) quantitatively that plant form is highly determined by environ- mental parameters, regardless of genetic constraints.
Results of this study (Table 2) support the generalization that species with an evergreen growth form have lower photosynthetic capacities than species with a graminoid, forb, or deciduous growth form (Harrison et al., 1971; Mooney and Dunn, 1970b). A possible explanation of these photosynthetic capacity differences between these diverse groups of tundra plants could be differences in carbon assimilation pathways. Three basic strategies of carbon metabolism in plant photosynthesis are widely recognized and include the C3, C4, and Crassulacean Acid Metabolism (CAM) pathways. However, since Tieszen and Sigurdson (1973) and Tieszen (unpublished data) found that all arctic tundra species examined had ribulose - 1, 5-diphosphate as the principal carboxylating enzyme, indicative of C3 carbon fixation, it is likely that these varied photo- synthetic capacities are not due to carbon assimilation pathway differences.
These low CO2 uptake rates in the evergreen shrub growth form could be due to the high stomatal resistances of evergreens (e.g., Whiteman and Koller, 1964). These high resistances may be an adaptive strategy to limit water loss but also necessarily result in reductions in CO2 uptake. Additionally, Mooney and Dunn (1970a) suggested that these low photosynthetic capacities in evergreen species are related to features enabling these species to maintain leaves for more than one growing season. This retention of leaves for more than one growing season likely requires synthesis of compounds involving over- wintering protection as well as predator protection. Small (1972) indicated that modifications of evergreen leaves necessary to survive the rigors of winter likely decrease availability of CO2 through the mesophyll and hence decrease photosyn- thetic capacity.
Cates and Orians (1975) indicated that late seral species such as evergreen shrubs make a large commitment of resources to chemical defenses against herbivores. Even though these chemicals may represent considerable energy resource expenditures for the evergreen plant, these compounds likely enhance the competitive abilities of the plant by reducing loss of photosynthetic tissue to herbivores (Cates, 1975). These factors in addition to the large consumption of assimilates necessary for year-round maintenance (Penning de Vries, 1975) likely results in high growth costs for evergreen leaves, and may result in less plant resources available to invest in high capacity photosynthetic apparatus.

Allocation, Leaf Growth, and Photosynthesis in Tundra Plants 171
Whatever the cause for the low photosynthetic capacities in evergreen shrubs, these are associated with relatively extended leaf longevities (Table 3). Presum- ably, low photosynthetic capacity evergreen leaves must be retained for more than one growing season in order to attain a positive carbon contribution to the plant. Contrasting the typically low photosynthetic capacities in evergreen shrubs were the relatively high CO2 uptake rates of deciduous shrubs. Photosyn- thetic capacities in this growth form were nearly twice as high as those for evergreen shrubs; however, these high carbon fixation rates in deciduous shrubs were offset by relatively short leaf longevity periods. Apparently growth forms with high photosynthetic capacity leaves contribute positively to the carbon budget of the plant in only one season. Intermediate to these patterns were those exhibited by the grasses and sedges. This growth form had photosynthetic rates between those of evergreen and deciduous shrubs and also exhibited inter- mediate leaf longevities.
Thus, it appears from the results of this study that among tundra species of different growth form, photosynthetic capacity is inversely related to leaf longevity. As a result, even though photosynthetic capacity and leaf longevity patterns differ markedly between growth forms, net carbon gain when considered over the life span of a leaf may be quite similar for these tundra species.
Acknowledgments. This research was supported by the U.S. National Science Foundation grant number OPP75-12944 to the authors as part of the Research on Arctic Tundra Environments (RATE) program. We gratefully acknowledge the assistance of Kathy Johnson, Katie Berg, Marie Slack, and Phil Mattheis and the field support of the Naval Arctic Research Laboratory, Barrow, Alaska. The secretarial assistance of Christa Vollstedt is greatly appreciated.
References
Allessio, M.L., Tieszen, L.L.: Patterns of carbon allocation in an arctic tundra grass, Dupontia fischeri (Gramineae), at Barrow, Alaska. Amer. J. Bot. 62, 797-807 (1975)
Billings, W.D., Mooney, H.A.: The ecology of arctic and alpine plants. Biol. Rev. 43, 481 529 (1968)
Bliss, L.C. : Adaptations of arctic and alpine plants to environmental conditions. Arctic 15, 117-144 (1962)
Bliss, L.C. : Arctic and alpine plant life cycles. Ann. Rev. Ecol. System 2, 405438 (1971) Britton, M.E.: Vegetation of the arctic tundra. In: Arctic biology (H.P. Hansen, ed.), pp. 67-113.
Corvallis, Oregon: Oregon State Univ. Press 1957 Cares, R.G.: The interface between slugs and wild ginger: Some evolutionary aspects. Ecology
56, 391-400 (1975) Cates, R.G., Orians, G.H. : Successional status and the palatability of plants to generalized herbivores.
Ecology 56, 410-418 (1975) Chapin, F.S., Van Cleve, K., Tieszen, L.L. : Seasonal nutrient dynamics of tundra vegetation at
Barrow, Alaska. Arctic Alpine Res. 7, 209-226 (1975) Clark, J. : Photosynthesis and respiration in white spruce and balsam fir. N.Y. State Univ. College
of Forestry Tech. PuN. 85, 1-72 (1961) Dennis, J.G., Johnson, P.L. : Shoot and rhizome-root standing crops of tundra vegetation at Barrow,
Alaska. Arctic Alpine Res. 2, 253-266 (1970) E1 Aouni, M.H., Mousseau, M. : Relation of CO2 exchange in needles of Austrian black pine
(Pinus nigra Arn.) with age, chlorophyll content, and reassimilation. Photosynthetica 8, 78-86 (1974)
Freeland, R.O.: Effect of age of leaves upon the rate of photosynthesis in some conifers. Plant Physiol. 27, 685-690 (1952)

172 D.A. Johnson and L.L. Tieszen
Gerasimenko, T.V., Zalenskiy, O.V. : Diurnal and seasonal dynamics of photosynthesis in plants of Wrangel Island [in Russian]. International Tundra Biome Transl. 11, Univ. Alaska. Fairbanks, Alaska: Tundra Biome Center 1974
Grace, J., Woolhouse, H.W. : A physiological and mathematical study of growth and productivity of a Calluna-Sphagnum community. I. Net photosynthesis of Calluna vulgar&. L. Hull. J. appl. Ecol. 7, 363-381 (1970)
Hadley, E.B., Bliss, L.C. : Energy relationships of alpine plants on Mr. Washington, New Hampshire. Ecol. Monogr. 34, 331-357 (1964)
Harrison, A.T., Small, E., Mooney, H.A. : Drought relationships and distribution of two mediter- ranean-climate California plant communities. Ecology 52, 869-875 (1971 )
Hedberg, O. : Features of afroalpine plant ecology. Acta Phytogeographiea Suecica 49, 1-144 (1964) Hiramatu, K. : The relation of COz-assimilation activity to the age of evergreen leaves. Ecol.
Rev. 5, 25-32 (1939) Hult6n, E. : Flora of Alaska and neighboring territories. Stanford, California: Stanford Univ.
Press 1968 Johnson, P.L., Tieszen, L.L.: Vegetative research in arctic Alaska. Arctic Inst. North America
Tech. Rep. 25, 169-198 (1973) Lewis, M.C., Callaghan, T.V.: Ecological efficiency of tundra vegetation. In: Vegetation and
atmosphere (J.L. Monteith, ed.), pp. 399-433. London: Academic Press 1975 (in press) Ludlow, M.M., Jarvis, P.G.: Photosynthesis in Sitka spruce (Picea sitehensis (Bong.) Carr.). I.
General characteristics. J. appl. Ecol. 8, 925-953 (1971) Ludwig, L.J., Canvin, D.T.: An open gas-exchange system for the simultaneous measurement
of the CO 2 and laCO2 fluxes from leaves. Canad. J. Bot. 49, 1299-1313 (1971) Mattheis, P.J., Tieszen, L.L., Lewis, M.C. : Responses of Dupontia fischeri to lemming grazing
in an Alaskan arctic tundra. Ann. Bot. (in press, 1976) Mayo, J.M., Despain, D.G., Zinderen Bakker, E.M. van: CO2 assimilation by Dryas integrifolia
on Devon Island, Northwest Territories. Canad. J. Bot. 51, 581-588 (I973) Mooney, H.A. : The carbon balance of plants. Ann. Rev. Ecol. Syst. 3, 315-346 (1972) Mooney, H.A., Billings, W.D. : Comparative physiological ecology of arctic and alpine populations
of Oxyria digyna. Ecol. Monogr. 31, 1-29 (1961) Mooney, H.A., Chu, C. : Seasonal carbon allocation in Heteromeles arbutifolia, a California ever-
green shrub. Oecologia (Bed.) 14, 295-306 (1974) Mooney, H.A., Dunn, E.L. : Convergent evolution of mediterranean-climate evergreen sclerophyll
shrubs. Evolution 24, 292-303 (1970a) Mooney, H.A., Dunn, E.L. : Photosynthetic systems of mediterranean-climate shrubs and trees
of California and Chile. Amer. Naturalist 104, 44%453 (1970b) Mooney, H.A., Harrison, A.T., Morrow, P.A. : Environmental limitations of photosynthesis on
a California evergreen shrub. Oecologia (Berl.) 19, 293-301 (1975) Parsons, D.J., Moldenke, A.R.: Convergence in vegetation structure along analogous climatic
gradients in California and Chile. Ecology 56, 950-957 (1975) Penning de Vries, F.W.T. : The cost of maintenance processes in plant cells. Ann. Bot. 39, 77-92
(1975) Perry, T.O., Baldwin, G.W. : Winter breakdown of the photosynthetic apparatus of evergreen
species. Forest Sci. 12, 298-300 (1966) Raunkier, C. : The life forms of plants and statistical plant geography. London: Clarendon Press
1934 Rickert, D.A., Tedrow, J.C.F. : Pedologic investigations on some aeolian deposits of northern
Alaska. Soil Sci. 104, 250-262 (1967) Shvetsova, V.M., Voznesenskii, V.L. : Diurnal and seasonal variations in the rate of photosynthesis
in some plants of western Taimyr [in Russian]. International Tundra Biome Transl. 2, Univ. Alaska. Fairbanks, Alaska: Tundra Biome Center 1971
Small, E. : Photosynthetic rates in relation to nitrogen recycling as an adaptation to nutrient deficiency in peat bog plants. Canad. J. Bot. 50, 2227-2233 (1972)
Tieszen, L.L.: Photosynthesis and respiration in arctic tundra grasses: Field light intensity and temperature responses. Arctic Alpine Res. 5, 239-251 (1973)
Tieszen, L.L. : CO e exchange in the Alaskan arctic tundra: Seasonal changes in the rate of photosyn- thesis of four species. Photosynthetica (in press, 1976)

Allocation, Leaf Growth, and Photosynthesis in Tundra Plants 173
Tieszen, L.L., Johnson, D.A.: Seasonal pattern of photosynthesis in individual grass leaves and other plant parts in arctic Alaska with a portable 14CO2 system. Bot. Gaz. 136, 99-105 (1975)
Tieszen, L.L., Johnson, D.A., Caldwell, M.M. : A portable system for the measurement of photosyn- thesis using 14-carbon dioxide. Photosynthetica 8, 151-160 (1974)
Tieszen, L.L., Johnson, P.L.: Pigment structure of some arctic tundra communities. Ecology 49, 370-373 (1968)
Tieszen, L.L., Sigurdson, D.C:: Effect of temperature on carboxylase activity and stability in some Calvin cycle grasses from the arctic. Arctic Alpine Res. 5, 59-66 (1973)
Tieszen, L.L., Wieland, N.K. : Physiological ecology of arctic and alpine photosynthesis and respira- tion. In: Physiological adaptation to the environment (F.J. Vernberg, ed.), pp. 157-200. New York: Intext Educational Publ. 1975
Whiteman, P.C., Koller, D.: Environmental control of photosynthesis and transpiration in Pinus halepensis. Israel J. Bot. 13, 166-176 (1964)
Wielgolaski, F.E., Webber, P.J. : A comparison of circumpolar arctic and alpine tundra vegetation by classification and ordination techniques. Arctic Alpine Res. (in press, 1976)
Received March 17, 1976




















