
A zooarchaeological contribution to establish occupational patterns at Level J of Abric Romaní...
16
A zooarchaeological contribution to establish occupational patterns at Level J of Abric Romaní (Barcelona, Spain) Jordi Rosell a, * , Isabel Cáceres a , Ruth Blasco a , María Bennàsar a , Pilar Bravo a , Gerard Campeny a , Montserrat Esteban-Nadal a , M. Cristina Fernández-Laso a , M. Joana Gabucio a , Rosa Huguet a , Nuria Ibáñez a , Patricia Martín a , Florent Rivals b , Antonio Rodríguez-Hidalgo a , Palmira Saladié a a IPHES (Institut Català de Paleoecologia Humana i Evolució Social), Unidad asociada al CSIC, Àrea de Prehistòria, Universitat Rovira i Virgili, Plaça Imperial Tarraco 1, 43005 Tarragona, Spain b ICREA and Institut Català de Paleoecologia Humana i Evolució Social (IPHES), Universitat Rovira i Virgili, Plaça Imperial Tarraco 1, 43005 Tarragona, Spain article info Article history: Available online xxx abstract In recent decades, many research questions focus on occupational patterns in the Middle Palaeolithic. Many discussions on that topic address the temporal resolution of formation of assemblages and the character of palimpsests that can be frequently observed at archaeological sites. The Abric Romaní site (Barcelona, Spain) is a good example to analyze questions related to temporal resolution of the human occupations. This site is filled by a succession of travertine platforms of rapid formation, which clearly separate different archaeological levels. With the objective of characterizing the settlement patterns developed at this site, a combination of zooarchaeological and spatial data from Level J (sublevels Ja and Jb) is presented here. On one hand, primary and immediate anthropogenic accesses to the carcasses (mainly horses and red deer) and well-established patterns of animal processing are observed. On the other hand, these activities are always developed around hearths of inhabitable areas. From a nutritional perspective, faunal refits suggest synchronic relationships between the different activity areas and, therefore, the anthropic occupation of the total surface of the rock-shelter during the formation of Level J. However, this certain synchrony is combined with the presence of other elements, such as overlapped hearths, which show a succession of several occupations with the same settlement patterns. In this way, Level J of Abric Romaní could be understood as the succession of several occupations formed by groups that maintained the same traditions through time. From this perspective, the aim of this paper is to improve data to understand the settlement patterns and the social organization systems of the Neanderthal groups during MIS 3. Ó 2011 Elsevier Ltd and INQUA. All rights reserved. 1. Introduction The study of subsistence strategies and occupational patterns is fundamental to understand the ways of life of the human communities during the Pleistocene and their evolution through time. Subsistence strategies are the set of systems that human groups use to organize themselves socially and economically in a territory with the aim of guaranteeing the survival of their community (Díez and Rosell, 1998). On the other hand, the pattern concept can be understood as regular ways of acting or doing something. Thus, occupational patterns are the result of the rela- tionships of the members of the same group among themselves and with the environment. The developed activities in one archaeo- logical site can help to understand these occupational patterns, because they can be interconnected and performed simultaneously (Bendixen et al., 2006). In recent times, studies related to zooarchaeology are a referent to understand the subsistence strategies of the human groups in the past. Nevertheless, this type of study can encounter significant difficulties related to the sedimentation conditions during the formation of the archaeological assemblages. In this sense, events of different nature are overlapped at several sites generating palimpsests, i.e. an amalgam of remains of different origins, for * Corresponding author. E-mail addresses: [email protected] (J. Rosell), [email protected] (I. Cáceres), [email protected] (R. Blasco), [email protected] (M. Bennàsar), pbravo@ codex.cat (P. Bravo), [email protected] (G. Campeny), [email protected] (M. Esteban-Nadal), [email protected] (M.C. Fernández-Laso), [email protected] (M.J. Gabucio), [email protected] (R. Huguet), [email protected] (N. Ibáñez), [email protected] (P. Martín), fl[email protected] (F. Rivals), [email protected] (A. Rodríguez-Hidalgo), [email protected] (P. Saladié). Contents lists available at ScienceDirect Quaternary International journal homepage: www.elsevier.com/locate/quaint 1040-6182/$ e see front matter Ó 2011 Elsevier Ltd and INQUA. All rights reserved. doi:10.1016/j.quaint.2011.01.020 Quaternary International xxx (2011) 1e16 Please cite this article in press as: Rosell, J., et al., A zooarchaeological contribution to establish occupational patterns at Level J of Abric Romaní (Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.2011.01.020
Transcript of A zooarchaeological contribution to establish occupational patterns at Level J of Abric Romaní...

lable at ScienceDirect
Quaternary International xxx (2011) 1e16
Contents lists avai
Quaternary International
journal homepage: www.elsevier .com/locate/quaint
A zooarchaeological contribution to establish occupational patterns at LevelJ of Abric Romaní (Barcelona, Spain)
Jordi Rosell a,*, Isabel Cáceres a, Ruth Blasco a, María Bennàsar a, Pilar Bravo a, Gerard Campeny a,Montserrat Esteban-Nadal a, M. Cristina Fernández-Laso a, M. Joana Gabucio a, Rosa Huguet a,Nuria Ibáñez a, Patricia Martín a, Florent Rivals b, Antonio Rodríguez-Hidalgo a, Palmira Saladié a
a IPHES (Institut Català de Paleoecologia Humana i Evolució Social), Unidad asociada al CSIC, Àrea de Prehistòria, Universitat Rovira i Virgili, Plaça Imperial Tarraco 1,43005 Tarragona, Spainb ICREA and Institut Català de Paleoecologia Humana i Evolució Social (IPHES), Universitat Rovira i Virgili, Plaça Imperial Tarraco 1, 43005 Tarragona, Spain
a r t i c l e i n f o
Article history:Available online xxx
* Corresponding author.E-mail addresses: [email protected] (J. Rosell), ic
[email protected] (R. Blasco), [email protected] (P. Bravo), [email protected] (G. Cam(M. Esteban-Nadal), [email protected]@iphes.cat (M.J. Gabucio), [email protected] (N. Ibáñez), [email protected]@icrea.es (F. Rivals), antonio.ro(A. Rodríguez-Hidalgo), [email protected] (P. Sal
1040-6182/$ e see front matter � 2011 Elsevier Ltd adoi:10.1016/j.quaint.2011.01.020
Please cite this article in press as: Rosell, J., e(Barcelona, Spain), Quaternary Internationa
a b s t r a c t
In recent decades, many research questions focus on occupational patterns in the Middle Palaeolithic.Many discussions on that topic address the temporal resolution of formation of assemblages and thecharacter of palimpsests that can be frequently observed at archaeological sites. The Abric Romaní site(Barcelona, Spain) is a good example to analyze questions related to temporal resolution of the humanoccupations. This site is filled by a succession of travertine platforms of rapid formation, which clearlyseparate different archaeological levels. With the objective of characterizing the settlement patternsdeveloped at this site, a combination of zooarchaeological and spatial data from Level J (sublevels Ja and Jb)is presented here. On one hand, primary and immediate anthropogenic accesses to the carcasses (mainlyhorses and red deer) and well-established patterns of animal processing are observed. On the other hand,these activities are always developed around hearths of inhabitable areas. From a nutritional perspective,faunal refits suggest synchronic relationships between the different activity areas and, therefore, theanthropic occupation of the total surface of the rock-shelter during the formation of Level J. However, thiscertain synchrony is combined with the presence of other elements, such as overlapped hearths, whichshow a succession of several occupations with the same settlement patterns. In this way, Level J of AbricRomaní could be understood as the succession of several occupations formed by groups that maintainedthe same traditions through time. From this perspective, the aim of this paper is to improve data tounderstand the settlement patterns and the social organization systems of the Neanderthal groups duringMIS 3.
� 2011 Elsevier Ltd and INQUA. All rights reserved.
1. Introduction
The study of subsistence strategies and occupational patterns isfundamental to understand the ways of life of the humancommunities during the Pleistocene and their evolution throughtime. Subsistence strategies are the set of systems that humangroups use to organize themselves socially and economically in
[email protected] (I. Cáceres),.cat (M. Bennàsar), pbravo@peny), [email protected] (M.C. Fernández-Laso),@iphes.cat (R. Huguet),tudiants.urv.cat (P. Martín),[email protected]é).
nd INQUA. All rights reserved.
t al., A zooarchaeological conl (2011), doi:10.1016/j.quaint.
a territory with the aim of guaranteeing the survival of theircommunity (Díez and Rosell, 1998). On the other hand, the patternconcept can be understood as regular ways of acting or doingsomething. Thus, occupational patterns are the result of the rela-tionships of themembers of the same group among themselves andwith the environment. The developed activities in one archaeo-logical site can help to understand these occupational patterns,because they can be interconnected and performed simultaneously(Bendixen et al., 2006).
In recent times, studies related to zooarchaeology are a referentto understand the subsistence strategies of the human groups inthe past. Nevertheless, this type of study can encounter significantdifficulties related to the sedimentation conditions during theformation of the archaeological assemblages. In this sense, eventsof different nature are overlapped at several sites generatingpalimpsests, i.e. an amalgam of remains of different origins, for
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

J. Rosell et al. / Quaternary International xxx (2011) 1e162
example hominids and carnivores (Villa and Bartram, 1996; Villaand Soressi, 2000; Rosell and Blasco, 2008, 2009). However, themain accumulator agent can be unique in some Pleistoceneassemblages (only hominids or carnivores). The abundance ofarchaeological remains in these sets can be related to severaloccupational events and, therefore, with a formation time more orless prolonged. Archaeologically, assessing this formation time iscomplicated, although it must be taken into account to makereconstructions of prehistoric human groups’ways of life. From thisperspective, it is important to find elements in the archaeologicalrecord that allow identification of unique events (i.e. limited intime). Faunal refits can provide information on this matter. Ingeneral, refitted bones with anthropogenic breakage (e.g., impactflakes with percussion notches) are related to animal processingand to consumption, and, therefore, can establish certainsynchrony in the deposition of the faunal remains. In this way, theposition of these refitted bones on the site’s surface can be used toconnect different areas and to establish temporal relationshipsbetween them.
Unfortunately, there are few works related to these subjects inthe Middle Palaeolithic. At Saint-Césaire (France), faunal refits areused to show the absence of mixed occupations at the Cha-telperronian levels (Morin et al., 2005). In the same way, theobjective of refitting bones in Kobeh Cave (Iran) is to reduce thenumber of unidentified bones and to providemore accurate data forthe archaeozoological analyses (Marean and Kim,1998). In contrast,studies that relate food sharing with faunal refits are more commonin Upper Palaeolithic sites: Pincevent and Verberie in France (Enloeand David, 1989; Audouze and Enloe, 1997; Enloe, 2003). Subse-quently, in the palaeoindian period, the sites of Horner (Todd andFrison, 1992) and Bugas-Holding in USA (Rapson and Todd, 1987)and Palangana in Argentina (Waguespack, 2002) follow this line.
From this perspective, the Abric Romaní site (Barcelona, Spain)presents unique characteristics to look into this matter. At this site,an alternation of travertine platforms with archaeological levels isobserved. These travertine platforms are formed in a shortgeological time period. For this reason, the archaeological levels aregenerated and sealed quickly. This fact hampers the action ofsubsequent taphonomical agents on the assemblage, which mayperturb the original location of remains. In this way, several spatialunits can be identified at this site following the criteria described insome ethnoarchaeological works (Yellen, 1977; Binford, 1987):specific activity areas always related to hearths, sleeping areas andtoss zones (Carbonell et al., 1996; Vaquero and Pastó, 2001;Vallverdú et al., 2005a, 2010).
According toVaquero andPastó (2001), hearths can be consideredas the main structure that focuses on the domestic activities. Thisconception isnotonlybasedon the studiesof theAbricRomaní site. Inother more ancient sites, such as Beeches Pit in United Kingdom(Preece et al., 2006), Rosh Ein Mor and Qesem Cave in Israel (Hietalaand Stevens, 1977; Karkanas et al., 2007) and Tor Faraj in Jordan(Henry et al., 1996), a similar phenomenon is also observed.
Initially, it is easy to fall into thedynamicof treating the remainsofeach level as the result of a unique occupational event. However, thetemporal dimension and the possibility of the conserved assemblagebeing the result of different occupational events should not be dis-carded (Carbonell et al., 1996, 2007; Vallverdú et al., 2005a). Thehuman occupation time and the number ofmembers included in thegroupdetermine the archaeological record. Several shortoccupationsof small groups (diachrony) can leave the same number of remains asa permanent occupation of a large group (synchrony).
Obviously, a discussion of this fact cannot be tackled from onesingle discipline. From this perspective, the aim of this paper is toimprove data on this question from faunal remains (zooarchaeo-logical and spatial analyses) at Level J of Abric Romaní to understand
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
the settlement patterns and the social organization systems of theNeanderthal groups during MIS 3.
2. The Abric Romaní site
The town of Capellades (Barcelona, Spain) is located on the rightmargin of the Anoia River 300 m above sea level. The town is builton a travertine platform (40e60 m high), dated to Lower andMiddle Pleistocene, that drains to the river from numerous water-falls called Cinglera del Capelló. Currently, these waterfalls areinactive. Multiple cavities and karst morphologies originated fromthis process, such as Abric Agut, Abric de la Consagració, Balma delsPinyons, Costa d’en Manel, and Abric Romaní (Fig. 1).
The Abric Romaní site is filled by a succession of more than 20mhigh travertine platforms originated by water filtration from thewall and cornices (Giralt and Julià, 1996). These travertine plat-forms cover the entire surface of the rock-shelter, and they may bemore than 1 m thick. When the water is not circulating into therock-shelter, platform construction is stopped and deposits ofdetrital materials are formed. Therefore, an alternation of travertineplatforms and thin deposits of clay and silt can be observed atstratigraphic sequence of Abric Romaní.
In general, the archaeological materials are recovered in thesedetrital deposits. The archaeological record is composed of lithicindustry, faunal remains, charcoals from hearths and wood pseu-domorphs (Castro-Curel and Carbonell, 1995). To date, 16 archae-ological levels have been excavated. Level A, at the top, is the onlyone that presents Upper Palaeolithic (circa 39 ka) materials(Bischoff et al., 1988). The other levels are classified in the MiddlePalaeolithic and are dated to MIS 3.
The lithic industry of Abric Romaní is mainly elaborated in chert,quartz and limestone. All these raw materials can be located ina close radius of the site, 15 km at maximum (Gómez de Soler,2009). Small and medium flakes and debitage are the most abun-dant objects. Occasionally, these flakes are retouched to producedenticulates. On the other hand, cores are not common, and theyare always exhausted at technological level. In general, the knap-ping sequences are completed in the site, suggesting that stone toolproductionwas a frequent activity at Abric Romaní (Carbonell et al.,1996; Vaquero, 1997; Martínez and Rando, 2000; Vallverdú et al.,2010).
Faunal remains have allowed identification of 13 species.Nevertheless, red deer (Cervus elaphus) and horses (Equus ferus) arethe most numerous animals throughout the stratigraphic sequence(Table 1) (Fernández-Laso et al., 2010). At some levels, the presenceof rhino (Stephanorhinus hemitoechus), aurochs (Bos primigenius),and Proboscidea (Mammuthus?) are also documented. These taxaare related to plain and forest ecosystems located at the end ofvalleys. To a lesser extent, some animal elements have beenrecovered which are found in rocky areas today, such as thechamois (Rupicapra pyrenaica). On the other hand, remains of Ursussp., Canis lupus, Panthera leo spelaea, Panthera pardus, Lynx sp., Felissilvestris and Crocuta crocuta have also been identified. Thesecarnivore remains are frequently recovered in the travertine plat-forms, without any relation to the archaeological levels. From thisfact, the presence of non-human predators at Abric Romaní sitecould be related to sporadic dens or another type of refuge. Occa-sionally, isolated remains of these animals are recovered inarchaeological levels. Nevertheless, the absence of anthropogenicmarks (cutmarks, intentional bone breakage or burned bones)suggests natural intrusions and, perhaps, a similar origin for thecarnivores recovered in the travertine platforms.
Level J is dated byU-series between 49.3�1.6 ka and 50.4�1.6 kaand by 14C AMS to 47 ka. This level is located in the central part of thestratigraphic sequence (Bischoff et al., 1988) and was excavated
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

Table 1Macromammals recovered at the Abric Romaní site and their location in generalstratigraphy.
Levels A B C D E F G H I J K L M
Ursus sp.Canis lupusPanthera leo spelaeaPanthera pardusLynx sp.Felis silvestrisCrocuta crocutaProboscideaStepohanorhinus hemitoechusEquus ferusCervus elaphusBos primigeniusRupicapra pyrenaica
Fig. 1. Location of the Abric Romaní site in the context of Iberian Peninsula and in relation to the Capellades town (left). Position of Level J in the general stratigraphy of the site(right).
J. Rosell et al. / Quaternary International xxx (2011) 1e16 3
extensively (circa 300 m2) during the seasons 1993e1998. Palae-oclimatically, Level J corresponds to the cold and dry phase of MIS 3,witha significantdeclineof treepollenand thedominanceof a steppeenvironment (Burjachs and Julià, 1994, 1996) (Fig. 1).
Several previous travertine structures characterize the forma-tion of Level J. Among these, the presence of numerous large blocksof travertine, fallen from the rock-shelter cornice, and the traver-tine structures with steep slopes (more than 45�) near the wall arethe most important. This phenomenon formed a basin at the centerof the rock-shelter (circa 125 m2) separated from the outside areaby a line of large blocks forming a semicircle. This basin is filled bydetrital sediments and provides more stratigraphic resolution thanzones near the wall, where the slope prevents the accumulation ofsediments. During the field work, Level J was divided into twosublevels in this basin (Ja and Jb). Both sublevels were clearlyseparated by a thin travertine platform. However, this platformwasnot continuous. Its edges were attached to themargins of the basin,and the subdivision between Ja and Jb was not possible at zonesnear the wall due to absence of this platform.
A minimum of 67 hearths have been identified at Level J: 42 atsublevel Ja and 25 at sublevel Jb (Carbonell et al., 2007). With someexceptions, these combustion structures are well-preserved. Lithicremains show,more or less, the same characteristics as other levels:scarce diversity of raw materials, with a predominance of chert,abundance of flakes and debitage and the presence of somedenticulates and exhausted cores.
3. Materials and methods
The zooarchaeological study from Level J includes all therecovered faunal remains. These were studied at anatomical andtaxonomical level (Lyman, 1994; Reitz and Wing, 1999). Theunidentified bones are grouped in weight sizes, following themodified criteria established by Bunn (1986): 1) very small size orsize 1 of Bunn (<20 kg); 2) small size or size 2 (100e20 kg); 3)medium size or sizes 3A and 3B (300e100 kg); 4) large size or size 4
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
(1000e300 kg) and; 5) very large size or size 5 (>1000 kg). Toassess the completeness of the sample, NR (Number of Remains),NISP (Number of Identified Specimens), MNE (MinimumNumber ofElements), MNI (Minimum Number of Individuals) and % SkeletalSurvival Rate are calculated (Brain, 1981; Lyman, 1994).
Following the criteria of birth of modern animals, teethreplacement sequence and dental wear have been used to establishthe age of death and seasonality of the ungulates, mainly Equusferus (Guadelli, 1998; Fernandez et al., 2006), Cervus elaphus(Mariezkurrena and Altuna, 1983a, b) and Bos primigenius (modernbison data from Brugal, 1999; Johnson et al., 2005). In this regard,four categories have been identified: 1) infantile (deciduous teeth),2) juvenile (M3 in eruption), 3) adult (complete permanent denti-tion) and, 4) senile (with an accused wear of the permanentdentition).
With the aim to check the incidence of differential conservationin the skeletal profile bias, a bivariate test was carried out. This testconfronts the % Skeletal Survival Rate with the values of mineraldensityobtainedonbonesof different currentmammals. In this case,
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

J. Rosell et al. / Quaternary International xxx (2011) 1e164
the data of horse mineral densities are used for large-sized animals(Lam et al., 1998), red deer mineral densities for medium-sizedanimals (Hillson, 1992) and sheep mineral densities for small-sizedanimals (Lyman, 1985).
Alterations observed on bone surfaces were treated at bothmacroscopic and microscopic level: Olympus SZ11 stereoscopic(magnification up to 110) and ESEM (FEI QUANTA 600). Cutmarks,intentional bone breakage, burning patterns and carnivore tooth-marks are the most common damage.
The breaks on faunal remains were analyzed and classifiedaccording to Villa and Mahieu (1991). The outlines (transverse,curved/V-shaped, longitudinal), fracture angles (oblique, right,mixed) and fracture edges (smooth, jagged) were analyzed to knowthe state of the bones at the moment of breakage (green or dry).Damage caused during the bone breakage was also studied and thediagnostic elements of anthropogenic breakage are documented onfaunal remains: percussion notches, impact flakes, percussion pitsand peeling. Percussion notches are semicircular-shaped indenta-tions on fracture edges with corresponding negative flake scars(Capaldo and Blumenschine, 1994; Pickering and Egeland, 2006).Impact flakes refer to positive flakes of the percussion notches anddisplay the same basic technical attributes as stone flakes (mainlyventral face with point of detachment and bulb). Percussion pits orpercussion marks are often closely associated with and/or havingemanated from the patches of striae that result from slippage ofstone against bone during impact events (Blumenschine andSelvaggio, 1988; Pickering and Egeland, 2006). Peeling definesa roughened surface with parallel grooves or fibrous textureproduced when fresh bone is fractured and peeled apart, similar tobending a small fresh twig with two hands (White,1992; Díez et al.,1999).
Regarding anthropogenic damage, cutmarks have been clas-sified as incisions, scrapes and chopmarks (Binford, 1981; Pottsand Shipman, 1981; Shipman, 1983; Shipman and Rose, 1983;Shipman et al., 1984a). Incisions are defined as striationswith a linear outline of variable length, width, and depth, witha V-shaped section and with internal microstriation. Otherfeatures of incisions can be Hertzian cones, shoulder effects andbarbs (Shipman and Rose, 1983; Bromage and Boyde, 1984). Thescrape marks are shallow sub-parallel cutmarks caused whena stone tool is dragged transversally along the length of the bone(Noe-Nygaard, 1989). Chopmarks are the result of blows, usinga bladed cutting instrument that causes short and deep cuts,
Table 2NR, NISP, MNE and MNI by taxonomic groups of animals from sublevels Ja and Jb of Abr
Ja
NR NISP MNE MNI MNI by ages
inf. juv.
Equus ferus 351 351 88 15 3 2Cervus elaphus 497 497 202 12 3 2Bos primigenius 88 88 47 5 e 2Stephanorhinus hemitoechus 33 33 17 3 1 1Rupicapra pyrenaica 6 6 5 2 e 1Ursus sp. 1 1 1 1 e e
Lynx sp. 1 1 1 1 e e
Canis lupus 2 2 2 2 1 e
Vulpes vulpes 1 1 1 1 e e
Crocuta sp. e e e e e e
Carnivora indet. 4 4 2 e e e
Large size 2150 e 87 e e e
Medium size 2165 e 109 e e e
Small size 352 e 35 e e e
Unidentified 1087 e e e e e
Total 6738 984 597 42 8 8
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
which are generally isolated, and in which no internal micro-striations are observed. The analysis of cutmarks takes intoaccount the number of striations, location (size and portion ofbone), and distributions on the surface (isolated, clustered,crossed), orientations (oblique, longitudinal, transverse) anddelineation (straight or curved).
Burning is also identified on faunal remains from Level J. Severalresearchers have been studying the modifications produced onfaunal remains by fire and it is possible to identify the bone states atthe moment of cremation (fresh bones, dry bones, bones with flesh,without flesh, buried bones, etc.) (Shipman et al., 1984b; Buikstraand Swegle, 1989; Nicholson, 1993; Stiner, 1994; Stiner et al.,1995; Bennett, 1999; and others). The main feature of burnedbones is the change of coloration: natural color to black (carbon-ized) to white (calcined). These changes are related to the intensityof the fire and the time of exposition. Following a modification ofthe criteria described by Stiner et al. (1995) to increase the items bycategories, the burned bones from Level J can be clustered into4 color groups: 1) Grade 0 for unburned bones; 2) Grade 1 forbrown stains more or less homogeneous across the bone surface(grades 1e2 of Stiner et al., 1995); 3) Grade 2 for black stains whenthe bone was charred (Grades 3e4 of Stiner et al., 1995); and 4)Grade 3 for gray and white stains (calcined), although occasionallyveins appear with bluish tones (Grade 5 of Stiner et al., 1995).
Regarding carnivore damage, pits, punctures and scores havebeen identified (Haynes, 1980, 1983; Binford, 1981; Stiner, 1994;Blumenschine, 1995; and others). Punctures are pits that deeplypenetrate the cortical of the bones. In general, they are located nearthe edges of broken bones. Scores are surfacemarks with the along-axis more than four times the perpendicular axis. As well as thecutmarks, the number and spatial characteristics of toothmarks onbones have also been recorded. Regarding the carnivore breakage,crenulated edges are mainly observed. These present small nega-tive scars on the fracture and longitudinal breakages. For theidentification of the carnivore type that acts in the assemblages,toothmarks have been measured and compared using the data ofAndrews and Fernández-Jalvo (1997), Selvaggio and Wilder (2001)and Dominguez-Rodrigo and Piqueras (2003).
The main postdepositional modifications from the Abric Romaníassemblages are related to water and vegetation. Polished androunded edges generated by water and striae caused by roots arethe most frequent alterations. To assess the influence of theseagents, 4 categories have been established: 1) Grade 0 or
ic Romaní.
Jb
NR NISP MNE MNI MNI by ages
ad. sen. inf. juv. ad. sen.
8 2 139 139 27 4 e 1 2 15 2 96 96 37 4 1 e 3 e
3 e 15 15 9 2 e 1 1 e
1 e 27 27 4 2 1 e 1 e
1 e 2 2 2 1 e 1 e e
1 e 0 e e e e e e e
1 e 1 1 1 1 e e 1 e
1 e 0 e e e e e e e
1 e 0 e e e e e e e
e e 1 1 1 1 1 e e e
e e 0 e e e e e e e
e e 297 e 20 e e e e e
e e 617 e 32 e e e e e
e e 221 e 18 e e e e e
e e 306 e e e e e e e
22 4 1722 281 151 15 3 3 8 1
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

Table 3Young ungulates from sublevels Ja and Jb and age of death expressed in months and season (pren ¼ prenatal).
Horse Red deer Auroch
Indiv. Months Season Indiv. Months Season Indiv. Months Season
JaInfantile 3 Pren-1 Spring 3 4e8 Autumn
Pren-1 Spring 4e8 AutumnPren-1 Spring 4e8 Autumn
Juvenile 2 e e 2 16e17 Autumn 2 30e33 Winterespringe e 27e30 Autumn e e
JbInfantile 1 5e8 AutumnJuvenile 1 30e36 Autumn 1 e e
J. Rosell et al. / Quaternary International xxx (2011) 1e16 5
unmodified, 2) Grade 1 or slightly modified, 3) Grade 2 or moder-ately modified and, 4) Grade 3 or highly modified. Other post-depositional agents, such as trampling or weathering, have beenalso identified, but with insignificant effects.
To assess the temporal connections between the faunal remainsfrom Level J, some archaeostratigraphical projections have beenmade, both in horizontal and vertical axes. The aim of theseprojections is to identify clusters of faunal remains or possiblespatial units at the site (Binford, 1978a, b; Vaquero and Pastó, 2001;Vaquero et al., 2001; Vallverdú et al., 2010). Bone refits are one ofthe best elements to check the temporal and spatial connectionsbetween these areas (Fernández-Laso, 2010; Fernández-Laso et al.,2010). Two types of faunal refits are described: 1) anatomicalconnections and, 2) connection of several fragments from the samebone (Morin et al., 2005; Lyman, 2008). The second type is the onlymethod documented at Level J because of the generalized absenceof epiphyses.
For bone refits, all faunal remains recovered at sublevel Ja havebeen tested. However, particular attention has been paid to boneswith evidences of anthropogenic or green breakage. This alsoincludes small remains, mainly impact flakes.
4. Results
At Level J of Abric Romaní, 8460 large mammal remains havebeen recovered. Of these, 6738 belong to sublevel Ja and 1722 tosublevel Jb. 984 (14.6%) remains from Ja and 281 (16.3%) from Jbhave been identified anatomically (using taxonomical groups)
Table 4MNE (% Survival Rate) of ungulates grouped by sizes of weight from sublevels Ja and Jb.
Ja
Large size % Survival Medium size % Survival Small size % Surv
MNI 17 11 9Cranium 7 41.2 6 27.3 1 11.1Maxilla 14 41.2 14 63.6 3 16.7Mandible 20 59 13 59.0 4 22.2Vertebra 1 0.2 23 8.0 15 6.4Rib 2 0.4 17 5.9 10 4.3Pelvis 2 5.9 e 0.0 e 0.0Scapula 6 17.6 7 31.8 0 0.0Humerus 23 67.6 20 90.9 2 11.1Radius-ulna 19 55.9 11 50.0 3 16.7Carpus 1 0.4 2 1.3 e 0.0Metacarpus 5 14.7 19 86.4 1 5.5Femur 16 47.0 17 77.3 3 16.7Patella e 0.0 1 4.5 e 0.0Tibia 20 58.8 22 100.0 3 16.7Tarsus 2 1.0 6 4.5 e 0.0Metatarsus 6 17.6 15 68.2 e 0.0Metapodium 8 5.9 2 4.5 1 2.8Phalanx 2 0.6 9 3.5 3 1.9
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
(Table 2). The low number of identified bones is related to the highrate of bone breakage and the absence of practically all theepiphysis at both sublevels.
As other levels of Abric Romaní, red deer (Cervus elaphus) andhorses (Equus ferus) are the most numerous animals. Other ungu-lates, such as aurochs, rhino and chamois were also recovered butin lower numbers. Carnivore remains recovered at Level J are alsorare.
Within the ungulates, adult individuals are the most common inthe two assemblages. Younger and older animals have also beendocumented. From the teeth replacement sequence and dentalwear of younger animals, a seasonal dichotomy related to age ofdeath of these species has been identified: spring or early summerfor horses and aurochs and autumn for red deer (Table 3). Theskeletal profile shows a domain of cranial and limb bones (Table 4).Axial bones are almost absent in both assemblages. These valuesare clearer if the animals are grouped into following size classes(including all the unidentified remains). From % Skeletal SurvivalRate, the values of cranial elements and proximal limb bones arehigh for large- and medium-sized animals and low for axial skel-eton at both sublevels (Fig. 2). On the other hand, the small-sizedanimals present incoherent values because of the scarce number ofremains from the represented individuals.
The bias observed in skeletal profile is not caused from differ-ential preservation processes. In this regard, the results of bivariatetests of bone mineral densities are not significant (Table 5).
Cutmarked bones are rare at both sublevels and never exceed 4%of the recovered remains (Tables 6 and 7). Incisions are the most
Jb
ival Large size % Survival Medium size % Survival Small size % Survival
7 4 22 28.6 3 75.0 1 50.06 42.8 2 25.0 1 25.0
12 85.7 7 87.5 2 50.05 2.4 10 9.6 6 11.54 2.2 8 7.7 6 11.52 14.3 2 25.0 1 25.02 14.3 3 37.0 2 50.06 42.8 8 100.0 1 25.03 21.0 7 50.0 1 12.5
e 0.0 2 3.6 e 0.02 14.0 5 62.0 1 25.05 36.0 6 75.0 1 25.0
e 0.0 e 0.0 e 0.012 85.7 4 50.0 2 50.0e 0.0 1 2.0 e 0.03 21.0 5 75.0 1 25.03 10.7 1 e 0.0
e 0.0 2 2.2 4 8.3
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

Fig. 2. % Skeletal Survival Rate by anatomic portions and size classes from sublevels Ja and Jb of Abric Romaní.
J. Rosell et al. / Quaternary International xxx (2011) 1e166
common type of cutmark, although scrape marks and chopmarkswere also identified. The incisions are usually located on the boneshafts and they are either longitudinal or oblique. These cutmarksare mainly related to defleshing activities. However, the incisionssituated on some cranial remains and ribs suggest skinning andviscera removal (Fig. 3). Fat and periosteum removal is documentedfrom scrapemarks. On the other hand, chopmarks are few, and theyare associated with disarticulation and dismembering processes ortendon removal. Therefore, all the activities related to the carcassprocessing are identified.
Following the criteria established by Villa and Mahieu (1991),the faunal assemblage from Level J is dominated by oblique, curvedand smooth fractures. This fact indicates that the bones are mainlybroken green. On the other hand, numerous diagnostic elements ofanthropogenic breakage are identified (Table 8). Impact flakes andpercussion notches are the most represented. The breakage processgenerates an assemblage characterized by small bones brokenwhile green: 38% of the recovered bone remains at Ja and 26% of therecovered bones at Jb are smaller than 2 cm. It is important to pointout that all the skeletal elements were not documented, and thenumber of epiphyses remains is very few. Only 1.2% at sublevel Ja
Table 5Maximum densities of the skeletal elements and skeletal survival rate (% survival) from sthe different weight sizes. Correlation (coefficient between 1 and -1 of highmagnitude) is(data from horse) and Hillson (1992) for medium-sized animals (data from red deer) an
Density % Survival Density
Equidae Large size Deer
Sublevels Ja Jb
Mandible 60 59 86 57Vertebra 54 0.2 2.4 30Rib 55 0.4 2.2 40Pelvis 65 5.9 14.3 49Scapula 67 17.6 14.3 49Humerus 64 67.6 42.8 63Radius 84 41.2 21 68Ulna 65 14.7 0 45Femur 59 47 36 57Tibia 82 58.8 85.7 74Carpus 62 0.4 0 98Tarsus 72 1 0 64Metacarpus 84 14.7 14 72Metatarsus 91 17.6 21 74Phalanx 67 0.6 0 57Correlations Coefficient Significance Coefficientr Pearson 0.157 0.578 0.594 0.333rho Spearman 0.205 0.464 0.810 0.339
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
and 0.9% at sublevel Jb of remains are epiphyses or cancellous tissuefragments.
Burning is also present in the faunal record, and several thermo-alteration degrees are documented (Table 9). 31.4% of remains fromsublevel Ja and 18.4% of sublevel Jb shows burning evidences. Thelowest grades of burning (Grade 1) aremost common at both levels.In addition, several bones with double colorations on the same sidewere recovered.
Bone modifications generated by carnivores are low at Level J: 61remains at sublevel Ja (0.9% of the total) and only 4 remains atsublevel Jb (0.2%). Regarding carnivore damage, punctures, pits,scores and crenulated edges on the diaphyses of large- andmedium-sized animals are the main identified alterations (Table 10). On theother hand, the dimensions of the carnivore toothmarks could indi-cate that the non-humanpredators that acted on Level J assemblagesmight bemedium-sized carnivores, such aswolves (Fig. 4). However,occasional toothmarks from a larger carnivore, probably a hyaenid,have also been documented.
Postdepositional modifications are more common at Jb than Ja.The presence of root-etching, chemical corrosions and cracking ishigh at Jb. However, sublevel Ja shows good preservation and, in
ublevels Ja and Jb. Note that Pearson’’s r and Spearman’s rho have been calculated bysignificant (p< 0.05). Bone density data from Lam et al. (1998) for large-sized animalsd Lyman (1985) for small-sized animals (data from sheep).
% Survival Density % Survival
Medium size Sheep Small size
Ja Jb Ja Jb
59 87 55 22.2 508 9.6 26 6.4 11.55.9 7.7 37 4.3 11.50 25 49 0 25
31.8 37 33 0 5090.9 100 42 11.1 2540.9 50 52 16.7 259.1 37 26 0 0
77.3 75 36 16.7 25100 50 59 16.7 50
1.3 3.6 68 0 04.5 2 63 0 0
86.4 62 67 5.5 2568.2 75 68 0 253.4 2.2 55 1.9 8.3
Significance Coefficient Significance0.225 0.597 �0.026 0.927 0.9030.216 0.553 �0.112 0.690 0.974
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020
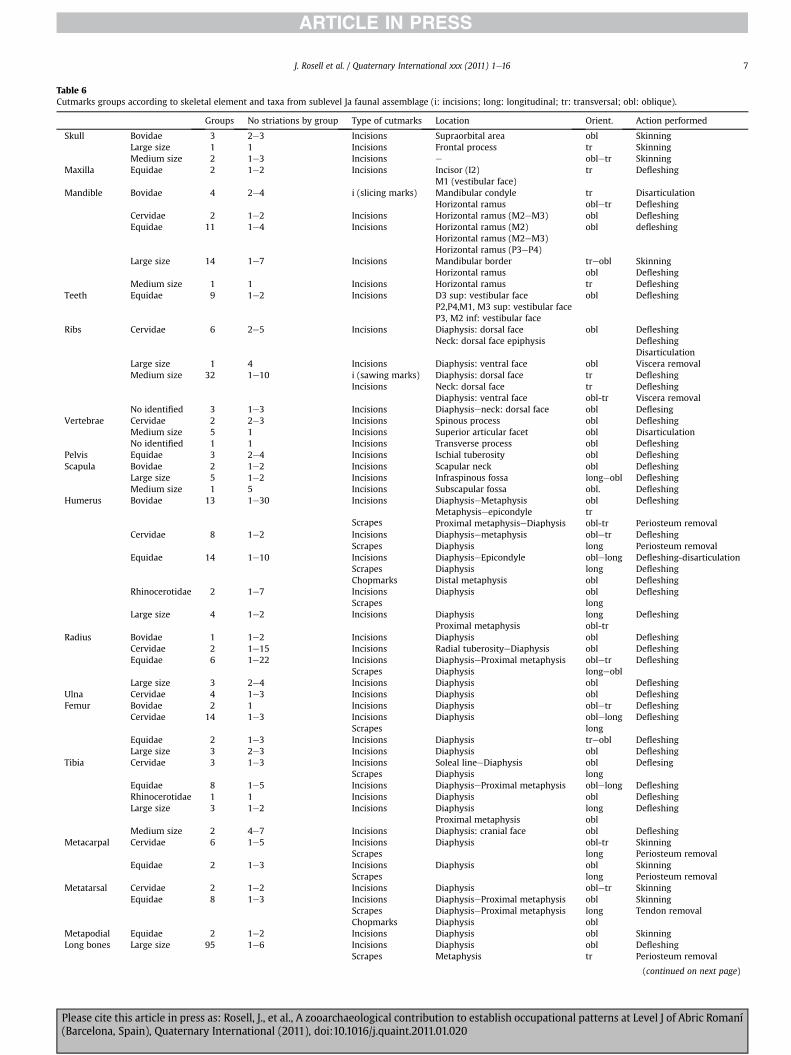
Table 6Cutmarks groups according to skeletal element and taxa from sublevel Ja faunal assemblage (i: incisions; long: longitudinal; tr: transversal; obl: oblique).
Groups No striations by group Type of cutmarks Location Orient. Action performed
Skull Bovidae 3 2e3 Incisions Supraorbital area obl SkinningLarge size 1 1 Incisions Frontal process tr SkinningMedium size 2 1e3 Incisions e obletr Skinning
Maxilla Equidae 2 1e2 Incisions Incisor (I2)M1 (vestibular face)
tr Defleshing
Mandible Bovidae 4 2e4 i (slicing marks) Mandibular condyleHorizontal ramus
trobletr
DisarticulationDefleshing
Cervidae 2 1e2 Incisions Horizontal ramus (M2eM3) obl DefleshingEquidae 11 1e4 Incisions Horizontal ramus (M2)
Horizontal ramus (M2eM3)Horizontal ramus (P3eP4)
obl defleshing
Large size 14 1e7 Incisions Mandibular borderHorizontal ramus
treoblobl
SkinningDefleshing
Medium size 1 1 Incisions Horizontal ramus tr DefleshingTeeth Equidae 9 1e2 Incisions D3 sup: vestibular face
P2,P4,M1, M3 sup: vestibular faceP3, M2 inf: vestibular face
obl Defleshing
Ribs Cervidae 6 2e5 Incisions Diaphysis: dorsal faceNeck: dorsal face epiphysis
obl DefleshingDefleshingDisarticulation
Large size 1 4 Incisions Diaphysis: ventral face obl Viscera removalMedium size 32 1e10 i (sawing marks)
IncisionsDiaphysis: dorsal faceNeck: dorsal faceDiaphysis: ventral face
trtrobl-tr
DefleshingDefleshingViscera removal
No identified 3 1e3 Incisions Diaphysiseneck: dorsal face obl DeflesingVertebrae Cervidae 2 2e3 Incisions Spinous process obl Defleshing
Medium size 5 1 Incisions Superior articular facet obl DisarticulationNo identified 1 1 Incisions Transverse process obl Defleshing
Pelvis Equidae 3 2e4 Incisions Ischial tuberosity obl DefleshingScapula Bovidae 2 1e2 Incisions Scapular neck obl Defleshing
Large size 5 1e2 Incisions Infraspinous fossa longeobl DefleshingMedium size 1 5 Incisions Subscapular fossa obl. Defleshing
Humerus Bovidae 13 1e30 Incisions
Scrapes
DiaphysiseMetaphysisMetaphysiseepicondyleProximal metaphysiseDiaphysis
obltrobl-tr
Defleshing
Periosteum removalCervidae 8 1e2 Incisions
ScrapesDiaphysisemetaphysisDiaphysis
obletrlong
DefleshingPeriosteum removal
Equidae 14 1e10 IncisionsScrapesChopmarks
DiaphysiseEpicondyleDiaphysisDistal metaphysis
oblelonglongobl
Defleshing-disarticulationDefleshingDefleshing
Rhinocerotidae 2 1e7 IncisionsScrapes
Diaphysis obllong
Defleshing
Large size 4 1e2 Incisions DiaphysisProximal metaphysis
longobl-tr
Defleshing
Radius Bovidae 1 1e2 Incisions Diaphysis obl DefleshingCervidae 2 1e15 Incisions Radial tuberosityeDiaphysis obl DefleshingEquidae 6 1e22 Incisions
ScrapesDiaphysiseProximal metaphysisDiaphysis
obletrlongeobl
Defleshing
Large size 3 2e4 Incisions Diaphysis obl DefleshingUlna Cervidae 4 1e3 Incisions Diaphysis obl DefleshingFemur Bovidae 2 1 Incisions Diaphysis obletr Defleshing
Cervidae 14 1e3 IncisionsScrapes
Diaphysis oblelonglong
Defleshing
Equidae 2 1e3 Incisions Diaphysis treobl DefleshingLarge size 3 2e3 Incisions Diaphysis obl Defleshing
Tibia Cervidae 3 1e3 IncisionsScrapes
Soleal lineeDiaphysisDiaphysis
obllong
Deflesing
Equidae 8 1e5 Incisions DiaphysiseProximal metaphysis oblelong DefleshingRhinocerotidae 1 1 Incisions Diaphysis obl DefleshingLarge size 3 1e2 Incisions Diaphysis
Proximal metaphysislongobl
Defleshing
Medium size 2 4e7 Incisions Diaphysis: cranial face obl DefleshingMetacarpal Cervidae 6 1e5 Incisions
ScrapesDiaphysis obl-tr
longSkinningPeriosteum removal
Equidae 2 1e3 IncisionsScrapes
Diaphysis obllong
SkinningPeriosteum removal
Metatarsal Cervidae 2 1e2 Incisions Diaphysis obletr SkinningEquidae 8 1e3 Incisions
ScrapesChopmarks
DiaphysiseProximal metaphysisDiaphysiseProximal metaphysisDiaphysis
obllongobl
SkinningTendon removal
Metapodial Equidae 2 1e2 Incisions Diaphysis obl SkinningLong bones Large size 95 1e6 Incisions
ScrapesDiaphysisMetaphysis
obltr
DefleshingPeriosteum removal
(continued on next page)
J. Rosell et al. / Quaternary International xxx (2011) 1e16 7
Please cite this article in press as: Rosell, J., et al., A zooarchaeological contribution to establish occupational patterns at Level J of Abric Romaní(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.2011.01.020

Table 6 (continued )
Groups No striations by group Type of cutmarks Location Orient. Action performed
Medium size 91 1e10 i (sawing marks)Incisions
DiaphysisEpiphysis
obletrtr
DefleshingDisarticulation
Small size 3 1 Incisions Diaphysis obl DefleshingNo identified 7 1e2 Incisions DiaphysiseMetaphysis obl-tr Defleshing
Flat bones Large size 36 1e9 IncisionsScrapesChopmarks
e obllongtr
Defleshing
Medium size 80 1 Incisions e obletr DefleshingSmall size 3 1 Incisions e obletr DefleshingNo identified 14 1e2 Incisions e obletr Defleshing
Unidentified. Large size 1 3 Incisions Diaphysis obl e
Medium size 8 1e>8 Incisions Diaphysis obletr e
No identified 22 1e2 Incisions e obletr e
J. Rosell et al. / Quaternary International xxx (2011) 1e168
general, movements of remains due to water abrasion are notobserved (Fig. 5). For this reason, a series of faunal refits werecarried out only with remains from Ja. In this sense, 92 refits (207remains) are registered, identifying 115 connection lines: 76 refitsof 2 remains, 12 refits of 3, 2 refits of 4, 1 refit of 5, and 1 refit of 6(Bravo, 2001). In general, these refits are locals, i.e. the connectedremains are located at a few cm distance. Nevertheless, some long-distance connections (between 4 and 8 m) can be also observed.These are located on the flat zones of the rock-shelter. The estab-lished connection lines do not seem to be affected by the layerslope.
From this perspective, two differentiated areas at sublevel Jacan be observed (Fig. 7). The first area corresponds to the outerzone of the rock-shelter. This area contains large travertine blocksand a pronounced layer slope towards the east and southeast (i.e.towards the river). Large, medium and small archaeologicalobjects have been recovered between these travertine blocks. Allthese elements are scattered, without apparent organization.Nevertheless, one group of 13 hearths have also been located(squares R-L/57-59), but only the combustion structures located inthe southeast contain archaeological remains. The second areacorresponds to the internal zone of the rock-shelter. This space is
Table 7Cutmarks groups according to skeletal element and taxa from sublevel Jb faunal assemb
Groups No striations by group Type o
Mandible Large Size 1 1 Incisio
Ribs Large Size 2 3e7 Incisio
Medium Size 2 2e3 IncisioSmall Size 1 1 Incisio
Vertebrae Medium Size 1 2 IncisioPelvis Cervidae 1 1 Incisio
Large Size 1 1 IncisioHumerus Cervidae 4 2e5 Incisio
Large Size 2 1e2 IncisioScrape
Femur Bovidae 1 2 IncisioTibia Equidae 3 3e5 Incisio
Large Size 3 1e10 IncisioMetatarsals Cervidae 4 1e2 Incisio
i (sawiMetapodials Large Size 1 1 IncisioLong bones Large Size 12 1e6 Incisio
i (sawiMedium Size 26 1e6 IncisioSmall Size 5 1e3 Incisio
Flat bones Large Size 1 2 IncisioMedium Size 8 1e3 IncisioSmall Size 1 1 Incisio
Unidentified No identified 1 1 Incisio
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
characterized by a flat surface without travertine blocks. There isa slight slope towards the southeast. Six hearth groups can beobserved: squares R-S/41-43 (Group 1), N-P/41-44 (Group 2), N-O/46-49 (Group 3), M-O/50-52 (Group 4), R-Q/51-52 (Group 5) andS-V/51-54 (Group 6). Around these groups of hearths, smallarchaeological remains are abundant, mainly charcoals, lithicflakes of debitage and faunal fragments. In general, these remainsdo not exceed 2 cm length. The large (>10 cm) and medium bones(10e2 cm) are very scarce.
The empty areas correspond to the travertine structures withsteep slopes prior to formation of Level J. The maximum concen-tration of remains is located in the center of the rock-shelter and inzones near the wall (mainly around the hearths). In the outer areas(outside the semicircle of large blocks), the faunal remains seem tobe more dispersed. On the other hand, burned remains are oftenassociated with hearths, except in the outer areas. From these data,a spatial organization is identified: remains with the highestdegrees of burning are often located in the center of the hearths,while remains with the lowest degrees are placed around thehearths. This is distinct from carnivore damage: gnawed remainsare scattered on the entire surface of the rock-shelter. Somethingsimilar is observed regarding the coprolites. Several coprolites were
lage (i: incisions; long: longitudinal; tr: transversal; obl: oblique).
f cutmarks Location Orient. Action performed
ns Horizontal ramus obl Defleshing
ns Diaphysis: ventral faceDiaphysis: dorsal face
tr Viscera removalDefleshing
ns Diaphysis: dorsal face obl Defleshingns Diaphysis: dorsal face obl Defleshingns Spinous process obl Defleshingns Ilium obl Defleshingns Acetabular fossa obl Dismemberingns DiaphysiseMetaphysis obl Defleshingns Diaphysis obl
longDefleshingPeriosteum removal
ns Diaphysis obl Defleshingns Diaphysis obl Defleshingns Diaphysis obl defleshingnsng marks)
Diaphysis obl SkinningTendon removal
ns Diaphysis obl e
nsng marks)
Diaphysis obltr
Defleshing
ns Diaphysis obletr Defleshingns Diaphysis oblelong Defleshingns Diaphysis obl Defleshingns Diaphysis oblelong Defleshingns Diaphysis obl Defleshingns e obl e
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

Fig. 3. Anthropogenic damage fromLevel J faunal assemblage of Abric Romaní: (a,b) cutmarks on internal surface of ribs belonging tomiddle-sized animals. These incisions are relatedto viscera removal; (c) long bone diaphysis belonging to middle-sized animal with oblique cutmarks related to defleshing; (d) burned bones with double coloration and cutmarks.
J. Rosell et al. / Quaternary International xxx (2011) 1e16 9
recovered at both sublevels (5 at Ja and 12 at Jb) with a disperseddistribution, although these are usually located on the hearths.
5. Discussion
From an ecological point of view, the Abric Romaní site is locatedin an ecotone formedby the contact between lowland andmountainecosystems. In this sense, the Anoia River represents a naturalcorridor between the coast plains (Penedès Depression) and theinternal plains (Òdena Basin) through the Catalanides Mountains.Abric Romaní is located at the entrance of this canyon and, therefore,it offers a high viewabove the plain. This fact could favor the controlof animal resources. Currently, horses and large bovids are migra-tory animals, and there is no evidence of change in past behavior. Inthe case of Abric Romaní, the herds of these animals could use theAnoiaValley in their migratory movements. On the other hand, reddeer is actually an ubiquitous species. For most of the year, theseanimals live solitary or in small groups of three or four females with
Table 8Number of remains with diagnostic criteria of anthropogenic bone breakage observed at
Percussionmarks
Percussionnotches
Impact flakes
Ja Jb Ja Jb Ja Jb
Equidae 7 e 46 6 3 e
Cervidae 4 e 76 8 5 e
Large bovids 1 e 33 2 4 e
Large size 24 1 49 12 113 25Medium size 6 1 21 6 103 41Small size 2 e e 1 e 3Unidentified 5 e 8 e 102 11Total NR/NISP 49 2 233 35 330 80
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
their young. However, herds for breading are formed in autumn.During this period, the animals usually do not run away if they areattacked. The behavior of these ungulates in addition to the peculiarlocation of Abric Romaní could be related to the human occupationperiodwithin the year. The seasonal use of strategic locations for thecontrol of the gregarious species during the Middle Palaeolithic isalso observed in other sites, such as the reindeer of Salzgitter Leb-enstedt, in Germany (Gaudzinski, 2000). At the Ortvale Klde site(Georgia), the same seasonal hunting strategies between Middleand Upper Palaeolithic human groups are observed (Adler et al.,2006). According to these authors, these practices are successfulonly considering the territory and knowledge of the seasonalmovements of the ungulates (Caucasian tur in this case).
The Ja and Jb faunal assemblages show similar archae-ozoological features: 1) two dominant taxa (red deer and horses),2) adult animals, 3) abundance of cranial elements (maxilla andmandibles) and limb bones (mainly stylopodials and zeugopodials),4) low number of axial elements, 5) cutmarks related to several
the sublevels Ja and Jb of Abric Romaní.
Adheringflakes
Corticalflakes
Peeling Countercoup
Ja Jb Ja Jb Ja Jb Ja Jb
1 e 1 e e e 3 e
2 1 4 e 7 e 4 11 e 2 e e e 2 e
1 e 19 5 4 1 3 e
4 1 38 10 18 1 4 e
e e e 3 3 e e e
e e 26 9 7 e e e
9 2 90 27 39 2 16 1
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

Table 9Burned bones classified by grades of colour (including double colorations) fromsublevels Ja and Jb of Abric Romaní.
Grade 1 Grade 2 Grade 3 Doublecolour
Total NR
Ja Jb Ja Jb Ja Jb Ja Jb Ja Jb
Large size 399 24 90 8 44 1 91 46 624 79Medium size 518 28 146 20 108 21 124 58 896 127Small size 157 11 37 8 33 6 26 9 253 34Unidentified 125 15 110 11 50 12 55 39 340 77Total NR 1199 78 383 47 235 40 296 152 2113 317
J. Rosell et al. / Quaternary International xxx (2011) 1e1610
performed actions (skinning, visceral removal, defleshing, disar-ticulation and periosteum and tendon removal), 6) high degree ofintentional breakage of limb bones for the marrow removal, and, 7)repetition of the same processing patterns on all the carcasses. Allthese characteristics suggest an anthropic origin of the assemblagesrelated to a primary and immediate access to the ungulates and anintensive exploitation of the carcasses (Binford, 1981; Domínguez-Rodrigo, 1999; Gaudzinski and Roebroeks, 2000; and others). Fromthis perspective, hunting was probably the most used technique toobtain these animals.
On the other hand, features of carnivore dens, such as animportant number of carnivore damage on bones, presence ofdiaphyseal cylinders, deciduous teeth (in the case of hyena) anddigested bones, are not identified in either of the two assemblages.
Table 10Carnivore damage identified at the sublevels Ja and Jb of Abric Romaní.
NR Types of damage Location
Carnivora Ja Rib 2 Score-pit e
Bovidae Ja Mandible 1 Score e
Humerus 1 Pitting DiaphysisFemur 1 Pit DiaphysisTibia 1 Score Proximal
metaphysisEquidae Ja Mandible 1 Scores e
Vertebrae 1 Pit Proximalapophysis
Radius 1 Score-pit DiaphysisMetacarpus 1 Score Proximal
MetaphysisDiaphysis
Tibia 2 Score-pit MetaphysisJb Tibia 1 Crenulated edge Diaphysis
Cervidae Ja Rib 2 Score-pit e
Humerus 2 Score-pitCrenulated edge
MetaphysiseDiaphysisDiaphysis
Radius 2 Score-pit DiaphysisFémur 2 Score-pit Diaphysise
EpiphysisTibia 2 Score-pit DiaphysisMetacarpus 2 Score DiaphysisMetatarsus 3 Score DiaphysisPhalange 1 Pit Epiphysis
Jb Metacarpus 1 Score-pit DiaphysisLarge size Ja Long bones 3 Puncture-score Diaphysis
Flat bones 3 Score-pit DiaphysisRibs 2 Score-pit Diaphysis
Mediumsize
Ja Long bones 11 Crenulated edgePits-score-puncture
MetaphysisDiaphysis
Flat bones 7 Pit DiaphysisJb Long bone 2 Pit Diaphysis
Small size Ja Long bones 1 Score-punctures DiaphysisFlat bones 1 Pit Diaphysis
Unident. Ja Long bones 1 Score DiaphysisFlat bones 1 Score DiaphysisUnidentified 3 Pit-score crenulated
edgeDiaphysis
Fig. 4. Dimensions of carnivore toothmarks on diaphysis of bones from sublevel Ja. Thenumber of toothmarked bones from sublevel Jb is more scarce and the graphicalrepresentation is not significant.
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
In contrast to ungulates, the presence of isolated carnivore remainsshould be interpreted as sporadic refuges or natural intrusions inmoments of human non-occupation of the rock-shelter.
The low number of axial elements cannot be explained from thedifferential preservation processes caused by postdepositionalagents. The results of bivariant tests between % Survival Rate andbone density are not statistically significant. For this reason,hominids and carnivores are themain agents that could cause theseabsences.
According to several naturalistic current studies (Blumenschine,1988) and in experimental works (Marean and Spencer, 1991;Marean et al., 1992), vertebrae, ribs and epiphyses of long bonesare usually disappeared during this process. This is used to explain
Fig. 5. Taphonomic modifications on faunal remains from sublevels Ja and Jb of AbricRomaní. Note that human damage and root action are the most common alterations inboth assemblages.
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

Fig. 6. Examples of anthropogenic breakage patterns on several skeletal elements from the Level J faunal assemblage: (a) on mandibles of horses; (b) on mandibles of red deer; (c)on a maxilla of horse; (d) on humeri of aurochs; (e) on humeri of reed deer. Note that the continuous repetition of the same breakage patterns generates remains with the samemorphological features.
Fig. 7. (a) Location of large travertine blocks that separate the inside of the outer zone of rock-shelter during the sublevel Ja formation and situation of the different groups ofhearths; (b) graphical representation of the connections established between several hearths areas based on faunal refits.
J. Rosell et al. / Quaternary International xxx (2011) 1e16 11
Please cite this article in press as: Rosell, J., et al., A zooarchaeological contribution to establish occupational patterns at Level J of Abric Romaní(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.2011.01.020

J. Rosell et al. / Quaternary International xxx (2011) 1e1612
the bias in the skeletal profiles in some Mousterian assemblages,such as the Iranian sites of Kobe Cave (Marean and Kim, 1998) andKujin (Marean and Cleghorn, 2003).
In this regard, sublevels Ja and Jb show the same features ofthese anthropogenic assemblages scavenged by carnivores (Roselland Blasco, 2008, 2009). However, the number of vertebrae, ribsand epiphyses recovered in the naturalistic observations, in theexperimental studies and in Kobeh Cave are higher than LevelJ (Table 11). For this reason, the bias observed in Level J could beinterpreted as a consequence of the anthropic differential transportof skeletal parts. From this perspective, skulls and limb elementsare mainly the anatomical portions transported, while ribs andvertebrae are left at the kill site. If the entire animals were carried tothe site, numerous rib and vertebrae remains should be found (andalso, more fragments of spongy tissue).
From the cost-effectiveness point of view, the human transportmodality based on skeletal elements with high nutritional value(bothmedulla andmeat) has several advantages. Once the animal isdismembered, these anatomical portions are easy to transport,while the thoracic skeleton presents more difficulties in moving.After removal of the visceral and back muscles, axial elementscould be abandoned. According to Oliver (1993), the axial bones donot have a high nutritional value for the human groups without anadvanced technology, such as boiling (Oliver, 1993). This couldexplain the almost total absence of elements belonging to axialskeletons. Similar transport decisions are observed at the MiddlePalaeolithic site of Qesem Cave, in Israel (Stiner et al., 2009).
On the other hand, the absence of epiphyses of limb bonescannot be explained from the same reasons. The oblique andlongitudinal cutmarks on diaphyses suggest that limb bones wereintroduced complete into the rock-shelter and that the epiphysesdisappeared after defleshing (Domínguez-Rodrigo, 1999). This factis not incompatible with the identified strategies of transport ofcarcasses and the absence of axial skeletons in both assemblages. Ifthese elements had followed the same processes as did limb bones,more fragments of flat bones should be recovered, mainly diaph-yses of ribs.
Another possible explanation for the absence of epiphyses couldbe carnivore activities after the human occupations. However, all theexperiments on anthropogenic assemblages scavenged by carnivoresshow a higher presence of epiphyses than those observed at Level Jof Abric Romaní (Blumenschine, 1988; Marean and Spencer, 1991;Marean et al., 1992; Marean and Kim, 1998). In this sense, anotherpossibility could be found in the anthropic processing andconsumption patterns. The fat contained in the epiphyses can beextracted by heating the bones. For this purpose, it is required tobreak the bones and to separate the epiphyses from the shafts. Fromthis, the epiphyses could be crushed and fat could be extracted andconsumed. The result of this process is the presence of bonebreakage patterns similar to those identified at both sublevels. Onthe other hand, a systematic breakage is also observed in other
Table 11Number of epiphyses, number of epiphyses recovered with carnivore toothmarksand percentage in Level J in relation to naturalistic and experimental studies onanthropogenic assemblages scavenged by carnivores.
Total number ofepiphyses
Epiphyses withtoothmarks
%
AR-Ja 78 2 2.5AR-Jb 18 0 0Firts hominid e Experiment 73 16 21.9First hominid e Natural model 3 2 66.7KobehCave 270 64 23.7First carnivore e Experiment 32 13 40.6First carnivore e Natural model 2 2 100.0
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
elements, such as mandibles and maxillae (Fig. 6). The absence ofepiphyses is also observed in other Middle Palaeolithic sites ofFrance, such as Les Pradelles and Grotte du Noisetier. In these sites,the presence of small fragments of cancellous tissue (unidentifiedfragments) is interpreted by Costamagno (personal communication)as the result of processing techniques for bone grease consumption.
In any case, the use of fire must have been important for thehuman groups of the Abric Romaní. However, the number ofburned bones is low. This fact could be explained by twophenomena: 1) animals were cooked before defleshing (doublecolorations at the same surface of the bones) (Rosell, 2001) and 2)cleaning up the inhabitable surface.
In the first case, double colorations on bones are caused by thedifferential exposure of the bone to fire. This phenomenon has beentreated by several authors with the aim of distinguishing theassemblages burned after their burial (De Graaff, 1961; Stiner et al.,1995; Bennett, 1999). In the case of Abric Romaní, double colora-tions are observed on the same surface of bones. Their features donot coincide with those described by Bennett (1999) for post-burialthermo-alterations. As well, the burned bones of sublevels Ja and Jbare not located below the hearths. According to Rosell (2001), thepresence of double colorations on the same bone surface can berelated to defleshing after cooking. The presence of meat cangenerate differential thermal alterations. During cooking, the heataffects more intensively the areas where the meat is scarce orabsent. Therefore, the result is the presence of double colorationson the same surface.
On the other hand, several authors suggest that burned bones atMiddle Palaeolithic sites could be related to their use as fuel(Costamagno et al., 2005, 2009). However, if this was carried out atLevel J, both the number of burned bones and cremation degreesshould be higher than those documented at Abric Romaní (degrees4 and 5 of Stiner et al., 1995). On the contrary, the burned bonesfrom Level J show a distribution: higher degrees are located in thecenter of the hearths and lower degrees are situated around thecombustion structures. In the sameway, this phenomenon suggestsgood preservation of the Level J record and the limited interventionof taphonomical processes, which spatially perturb the assem-blages, mainly at Ja. The presence of root-etching is due to thepresence of vegetative action at both sublevels, but these aremainly located in the outer areas of the rock-shelter. Abrasionmarks on some bone surfaces caused by water action have alsobeen observed in the same areas. However, this process does notseem to disturb the sample excessively: 1) there is no significantdisappearance of small items carried by water (<2 cm), 2) there isno preferential orientation of large bones in these areas and, 3)there are no rolled bones in these zones at any of the sublevels. Forthese reasons, the position of bones could be close to their originalsituation.
According to Vaquero and Pastó (2001), the features of this type ofdiscrete accumulations are related to domestic areas, and they can beconsidered basic spatial units in the settlement strategies at theMiddle Palaeolithic levels of Abric Romaní. The organization ofactivity areas around hearths is also observed ethnoarchaeologically,such as the !Kung in Botswana (Yellen, 1977), the Alyawara inAustralia (O’Connell, 1987) or the Efe in Zaire (Fisher and Strickland,1991), among others.
Therefore, a certain spatial organization at sublevel Ja can beobserved. Nevertheless, it is possible to think that each activity areawas independent of the others. This fact might suppose an absenceof temporal connections between them. In this way, the Ja assem-blage should be considered a succession of small occupations limitedin time and space. However, two elements suggest connectionsbetween these areas: 1) the large number of recovered ungulates(this implies a significant quantity of food) and, 2) the faunal refits
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020
MJ
Resaltado

J. Rosell et al. / Quaternary International xxx (2011) 1e16 13
(these connect activity areas between them). On the contrary, ifthese areas had been generated independently and at differenttimes, only local refits (inside of the same area) would have beenrecovered. At sublevel Ja, several long-distance refits with diagnosticevidences of intentional breakage (impact flakes and percussionnotches or fresh breakage) have been documented (Fig. 8). This factindicates a nutritional character on these movements. On the otherhand, faunal refits do not describe a random scattering on thesurface of the rock-shelter, as it would be expected if movementswere accidental or related to other anthropogenic processes, such aschildren’s games (Hammond and Hammond, 1981; Kamp, 2001;Politis, 2005; Baxter, 2006; Shea, 2006). On the contrary, faunalrefits connect groups of hearths between them. This phenomenonsuggests the use of different areas simultaneously and, therefore, theoccupation of a large area at the rock-shelter.
Several elements show that sublevel Ja is formed in differentoccupational events: 1) the seasonality of the main species (horsesand aurochs during spring and red deer during autumn); 2) thepresence of unconnected areas, such as Groups 1 and 5 (indepen-dent areas); 3) the presence of overlapped hearths (hearth forma-tion separated in time); 4) the carnivore damage related to thescavenging of remains left by hominids (the presence of non-human predators is incompatible with the presence of humangroups) and 5) the presence of isolated carnivore remains withoutanthropogenic damage and related to natural intrusions duringmoments of human inoccupation (refuges). From this perspective,occupations placed on a limited surface might be combined withoccupations located on a large surface of the rock-shelter during theformation of Level J. Nevertheless, these data should be comparedwith those obtained from stone tool refits. Lithic artefacts can besubject to recycling events, and stone removal can occur at differentmoments (including different occupational events). However,faunal refits (mainly thosewith anthropogenic breakage evidences)have a nutritional character, and therefore they should correspond
Fig. 8. Examples of bone refits (percussion notches with impact flakes) from sublevelJa of Abric Romaní.
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
the time between the processing and consumption (same occupa-tional event).
A similar phenomenon is observed in Upper Palaeolithic sites,such as Pincevent and Ververie in France (Enloe and David, 1989;Audouze and Enloe, 1997; Enloe, 2003) and in paleoindian sites,such as Horner (Todd and Frison, 1992) and Bugas-Holding in USA(Rapson and Todd, 1987) and Palangana in Argentina (Waguespack,2002). In these sites, the connected activity areas with bone refitsare interpreted as evidence of food sharing between differentfamilies. In this regard, it is possible to consider sublevel Ja in thesame way. All the connected activity areas show the samecomposition: one hearth with small faunal and lithic remainsassociated. This phenomenon suggests the presence of simulta-neous multifunctional activity areas.
As well, at other levels of the Abric Romaní site, specifically atLevel I (46 ka), Level K and Level L (52 ka), both lithic and faunal refitsare scarce and located in the same area. This shows activitiesrestricted to the provisioning strategies of lithic rawmaterials and tothe obtaining of animals. The presence of small-occupied surfacessuggests several episodes of short occupations related to huntingand a high linear mobility of Neanderthal groups in the territory(Vallverdú et al., 2005a, b; Fernández-Laso, 2010; Fernández-Lasoet al., 2010). Furthermore, the faunal remains of levels I, K and Lhave the same features observed at level J: 1) primary and imme-diate anthropic access to the carcasses; 2) two dominant taxa(horses and red deer); 3) anatomical bias of carcasses (presence ofcranial and limb bones); 4) adult individuals; 5) high degree ofintentional bone breakage; and 5) presence of cutmarked andburned bones related to the processing and consumption. However,the short occupations of these levels contrastwith the larger surfaceused at sublevel Ja. This phenomenon is also observed at level M ofthe same site (Fernández-Laso, 2010). In this sense, a diversity ofsettlement patterns and food-sharing strategies can be observed atthe Middle Palaeolithic period of Abric Romaní.
Regarding subsistence strategies, the directionality of hunting inNeanderthal contexts has always provided controversy (d’Erricoand Sánchez Goñi, 2003; Adler et al., 2006). In Upper Paleolithiccontexts, the predominance of a unique species is usually related tospecialization and the capacity of Homo sapiens to develop complexhunting techniques. In contrast, at the Middle Palaeolithic sites ofEurope, this directionality is mainly attributed to the abundance ofthe same species in the environment and to the lack of possibilitiesfor Neanderthal groups to exploit different resources. However,several Middle Palaeolithic sites have been interpreted as accu-mulations resulting from specialized hunting. Examples of this arethe aurochs and the bears from Biache-Saint-Vaast in Belgium(Auguste, 1995), the bison from Mauran in France (Farizy et al.,1994), the reindeer from Salzgitter Lebenstedt in Germany(Gaudzinski and Roebroeks, 2000), and the saiga from Chokurcha,Buran Kaya, Starocelié and Kabazi II in Crimea (Patou-Mathis,2006). This directionality contrasts with other sites where a diver-sity of obtaining strategies is observed, such as the case of level XIIat Bolomor Cave (MIS 6), in Spain (Blasco et al., 2010).
6. Conclusions
The zooarchaeological study indicates that the origin of LevelJ (sublevels Ja and Jb) is mainly anthropic. The presence of seasonaloccupations based on age at death of red deer, horses and aurochsand the presence of unconnected areas suggest several occupa-tional events. Nevertheless, a combination of occupations on largesurface and occupations on a limited surface is observed (mainly bythe presence of unconnected areas). However, all the occupationalevents are characterized by a repetition of the same patterns of
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020
MJ
Resaltado
MJ
Resaltado
MJ
Resaltado
MJ
Resaltado
MJ
Resaltado
MJ
Resaltado
MJ
Resaltado
MJ
Resaltado
MJ
Resaltado

J. Rosell et al. / Quaternary International xxx (2011) 1e1614
carcasses processing and of spatial organization (same location ofthe hearths and activity areas around them).
According to the seasonal pattern, it is possible that humangroups used the rock-shelter during the periods of the year withmost available faunal resources in the environment, specificallyspring for horses and autumn for red deer. This interest in thecapture of specific animals is combined with occasional huntingepisodes of other taxa, such as aurochs, rhinoceros, or chamois.
Well-established patterns are used in the carcass processing,which are repeated throughout the formation of both assemblages.The human groups prepared the carcasses at the obtaining sitewiththe aim of moving the selected anatomical portions (cranial andlimb bones) to the rock-shelter. This involves the development ofactivities as such skinning, visceral removal and dismembering. Theabsence of ribs and vertebrae in the assemblages can be explainedfrom the human decision to transport and not from carnivore-scavenging activities.
Once inside the rock-shelter, cranial and limb boneswere roasted,defleshed and fractured. In this sense, the bone fragments resultingfrom bone breakage suggest a double purpose: 1) to remove themarrow in excellent condition and 2) to obtain the epiphyses asa whole. This processing leads to the appearance of some very well-established bone morphotypes in which the disappearance of theepiphyses is constant (probably for fat removal).
All this systematization in the butchery process is related toa complex distribution of space. The activity areas are alwayslocated around the hearths of the internal zone of the rock-shelter. However, the faunal remains recovered in the outer zones(a combination between large bones and small bones) suggestthe existence of toss areas. On the other hand, several move-ments of faunal remains, which connect different activity areas,are observed in the internal zone. These movements havea nutritional character, and they indicate the occupation of therelatively large spaces with the use of several activity areassimultaneously.
Subsequent to the human activities, some carnivores sporadi-cally visited the cavity to scavenge faunal remains left by hominids(mainly wolves and probably, hyena). However, the intensive use ofthe animal resources by the hominids implies that the activities ofthese marauders did not excessively perturb the assemblages. Inthe same way, postdepositional processes do not disrupt theassemblages, so the remains are recovered very close to theiroriginal position.
In summary, Level J of Abric Romaní shows patterns in animalprocessing, which did not change during thewhole time of the levelformation. It is possible to think that these well-establishedpatterns were the result of traditions maintained through time.These parameters suggest an important territorial componentamong the groups that occupied the Abric Romaní site in thisperiod. From this perspective, the site would have an importantrole in the seasonal displacements of these groups. This generalmodel shows that the human groups of Abric Romaní had a highcapacity of organization of their activities, which can be attributedto the hominids of the European Middle Palaeolithic.
Acknowledgements
We want to thank the excavation team of Abric Romaní. Specialthanks to Elena Petrova for English corrections. This research wassupported by Ministerio de Educación y Ciencia SpanishGovernment Grants CGL2009-12703-C03-02 and CGL2009-7896,and by Generalitat de Catalunya Grant 2009 SGR 188.Ruth Blascoand M.Joana Gabucio are beneficiaries of an FI Grant fromGeneralitat de Catalunya and financed by European Social Found.Antonio Rodríguez-Hidalgo is beneficiary of FPI Grant assigned to
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
project from Ministerio de Educación y Ciencia Spanish Govern-ment. The field excavationwork was supported by Departament deCultura i Mitjans de Comunicació de la Generalitat de Catalunya,Diputació de Barcelona, Ajuntament de Capellades and Arts Gràfi-ques Romanyà-Valls S.A.
References
Adler, D.S., Bar-Oz, G., Belfer-Cohen, A., Bar-Yosef, O., 2006. Ahead of the game.Middle and upper Palaeolithic hunting behaviors in the Southern Caucasus.Current Anthropology 47 (1), 89e118.
Andrews, P., Fernández-Jalvo, Y., 1997. Surface modifications of the Sima de losHuesos fossil humans. Journal of Human Evolution 33 (2/3), 191e216.
Audouze, F., Enloe, J.G., 1997. High resolution archaeology at Verberie: limits andinterpretations. World Archaeology 29 (2), 195e207.
Auguste, P., 1995. Chasse et charognage au Paléolithique Moyen: l’apport du gise-ment de Biache-Saint-Vaast (Pas-de-Calais). Bulletin de la Société PréhistoriqueFrançaise 92 (2), 155e167.
Baxter, J.E., 2006. Making space for children in archaeological interpretations.Archeological Papers of the American Anthropological Association 15 (1),77e88.
Bendixen, H.J., Kroksmark, U., Magnus, E., Jakobsen, K., Alsaker, S., Nordell, K., 2006.Occupational pattern: a renewed definition of the concept. Journal of Occupa-tional Science 13 (1), 3e10.
Bennett, J.L., 1999. Thermal alteration of buried bone. Journal of ArchaeologicalScience 26, 1e8.
Binford, L.R., 1978a. Nunamiut Ethnoarchaeology. Academic Press, New York.Binford, L.R., 1978b. Dimensional analysis of behavior and site structure: learning
from an eskimo hunting stand. American Antiquity 43 (3), 330e361.Binford, L.R., 1981. Bones: Ancient Men and Modern Myths. Academic Press, New
York.Binford, L.R., 1987. Researching ambiguity: frames of reference and site structure.
In: Kent, S. (Ed.), Method and Theory for Activity Area Research. ColumbiaUniversity Press, New York, pp. 449e512.
Bischoff, J.L., Julià, R., Mora, R., 1988. Uranium series dating of the Mousterianoccupation at Abric Romaní, Spain. Nature 332, 68e70.
Blasco, R., Fernández-Peris, J., Rosell, J., 2010. Plusieurs stratégies différentes pourl’obtention de la nourriture à la fin du Pléistocène moyen: le cas du niveau XIIde la Grotte du Bolomor (Valencia, Espagne). Comptes rendues Paleovol 9 (4),171e184.
Blumenschine, R.J., Selvaggio, M., 1988. Percussion marks on bone surfaces as a newdiagnostic of hominid behavior. Nature 333, 763e765.
Blumenschine, R.J., 1988. An experimental model of the timing of hominid andcarnivore influence on archaeological bone assemblages. Journal of Archaeo-logical Science 15, 483e502.
Blumenschine, R.J., 1995. Percussion marks, tooth marks, and experimental deter-minations of the timing of hominid an carnivore access to long bones at FLKZinjanthropus, Olduvai Gorge, Tanzania. Journal of Human Evolution 29, 21e51.
Brain, C.K., 1981. The Hunters or the Hunted? An Introduction to African CaveTaphonomy. University of Chicago Press, Chicago.
Bravo, P., 2001. Estudio zooarqueológico y de remontajes del subnivel Ja del AbricRomaní (Capellades, Barcelona). Tesis de Licenciatura. Dpt. d’Història i Geo-grafia. Universitat Rovira i Virgili, Tarragona.
Bromage, T.G., Boyde, A., 1984. Microscopic criteria for the determinationof directionality cutmarks on bone. American Journal of Physical Anthropology65, 359e366.
Brugal, J.-Ph., 1999. Étude de populations de grands Bovidés européens: intéret pourla connaissance des comportements humains au Paléolithique. In: Brugal, J.-Ph.,David, F., Enloe, J.G., Jaubert, J. (Eds.), Le Bison: Gibier et Moyen de Subsistancedes Hommes du Paléolithique aux Paléoindiens des Grandes Plaines. Antibes:Actes du Colloque International, Toulouse, 1995, pp. 85e103.
Buikstra, J., Swegle, M., 1989. Bone modification due to burning: experimentalevidence. In: Bonnichsen, R., Sorg, E. (Eds.), Bone Modification. University ofMaine, Orono, pp. 247e258.
Bunn, H., 1986. Patterns of skeletal representation and hominid subsistence activ-ities at Olduvai Gorge, Tanzania, and Koobi for a, Kenya. Journal of HumanEvolution 15, 673e690.
Burjachs, F., Julià, R., 1994. Abrupt climatic changes during the last glaciation basedon pollen analysis of the Abric Romani, Catalonia, Spain. Quaternary Research42, 308e315.
Burjachs, F., Julià, R., 1996. Palaeoenvironmental evolution during the Middle-Upper Palaeolithic transition in the NE of the Iberian Peninsula. In:Carbonell, E., Vaquero, M. (Eds.), The Last Neandertals/The First AnatomicallyModern Humans. Cultural Change and Human Evolution: The Crisis at40 ka BP, pp. 377e383. Igualada, Barcelona, Spain.
Capaldo, S.D., Blumenschine, R.J., 1994. A quantitative diagnosis of notches made byhammerstones percussion and carnivore gnawing on bovid long bones.American Antiquity 59 (4), 724e748.
Carbonell, E., Cebrià, A., Allué, E., Cáceres, I., Castro, Z., Díaz, R., Esteban, M., Ollé, A.,Pastó, I., Rodríguez Álvarez, X.P., Rosell, J., Sala, R., Vallverdú, J., Vaquero, M.,Vergés, J.M., 1996. Behavioural and organizational complexity in the MiddlePalaeolithic from Abric Romaní. In: Carbonell, E., Vaquero, M. (Eds.), The Last
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

J. Rosell et al. / Quaternary International xxx (2011) 1e16 15
Neandertals/The First Anatomically Modern Humans. Cultural Change andHumanEvolution: The Crisis at 40 ka BP, pp. 385e434. Igualada, Barcelona, Spain.
Carbonell, E., Lorenzo, C., Vallverdú, J., 2007. Centralidad espacial y operativa de losneandertales. Análisis espacial diacrónico de las actividades de combustión enel Abric Romaní (Anoia, Capellades, Barcelona). In: Baquedado, E. (Ed.), ElUniverso del Neandertal I. Ed. Ibersaf, Madrid, pp. 197e219.
Castro-Curel, Z., Carbonell, E., 1995. Wood pseudomorphs from Level I at AbricRomaní (Barcelona, Spain). Journal of Field Archaeology 22, 376e384.
Costamagno, S., Théry-Parisot, I., Brugal, J.P., Guilbert, R., 2005. Taphonomicconsequences of the use of bones as fuel. Experimental data and archaeologicalapplications. In: O’Connor, T. (Ed.), Biosphere to Lithosphere, New Studies inVertebrate Taphonomy, (Proceed. 9th Conf. ICAZ, Durham, Aug. 02). OxbowBooks, Durham, pp. 51e62.
Costamagno, S., Théry-Parisot, I., Castel, J.-C., Brugal, J.P., 2009. Combustible ou non?Analyse multifactorielle et modèles explicatifs sur des ossements brûlés palé-olithiques. In: Théry-Parisot, I., Costamagno, S., Henry, A. (Eds.), Gestion descombustibles au Paléolithique et au Mésolithique: nouveaux outils, nouvellesinterprétations. Proceed. Worshop 21, XV congress UISPP, Lisboa. BAR Inter-national Series, Oxford, pp. 65e84.
De Graaff, G., 1961. Gross effects of a primitive hearth. South African ArchaeologicalBulletin 16, 25e26.
d’Errico, F., Sánchez Goñi, M.F., 2003. Neandertal extinction and the millennial scaleclimatic variability of OIS 3. Quaternary Science Reviews 22, 769e788.
Díez, J.C., Rosell, J., 1998. Estrategias de subsistencia de los homínidos de la Sierra deAtapuerca. In: Aguirre, E. (Ed.), Atapuerca y la Evolución Humana. FundaciónRamón Areces, Madrid, pp. 361e390.
Díez, J.C., Fernández-Jalvo, Y., Rosell, J., Cáceres, I., 1999. Zooarchaeology andtaphonomy of Aurora stratum (Gran Dolina, Sierra de Atapuerca, Spain. Journalof Human Evolution 37 (3/4), 623e652.
Domínguez-Rodrigo, M., Piqueras, A., 2003. The use of tooth pits to identifycarnivore taxa in tooth-marked archeofaunas and their relevance to recon-struct hominid carcass processing behaviours. Journal of ArchaeologicalScience 30, 1385e1391.
Domínguez-Rodrigo, M., 1999. Flesh availability and bone modification incarcasses consumed by lions. Palaeogeography, Palaeoclimatology, Palae-oecology 149, 373e388.
Enloe, J.G., David, F., 1989. Le remontage des os par individus: le partage du rennechez les Magdaléniens de Pincevent (La Grande Paroisse, Seine-et-Marine).Bulletin de la Société Prehistorique Française 86 (9), 275e281.
Enloe, J.G., 2003. Acquisition and processing of reindeer in the Paris Basin. In:Costamagno, S., Laroulandie, V. (Eds.), Zooarchaeological Insights into Magda-lenian Lifeways. BAR International Series, Oxford, pp. 23e31.
Farizy, D., David, F., Jaubert, J., 1994. Hommes et Bisons du Paléolithique Moyen àMauran (Haute-Garonne). XXXe Suppl., Gallia Préhistoire Paris.
Fernandez, P., Guadelli, J.-L., Phose, Ph, 2006. Applying dynamics and comparinglife tables for Pleistocene Equidae in anthropic (Bau de l’Aubesier, Combe-Grenal) and carnivore (Fouvent) contexts with modern feral horse pop-ulations (Akagera, PryorMountain). Journal of Archaeological Science 33,176e184.
Fernández-Laso, M.C., Rivals, F., Rosell, J., 2010. Intra-site changes in seasonality andtheir consequences on the faunal assemblages from Abric Romaní (MiddlePalaeolithic, Spain). Quaternaire 21 (2), 255e263.
Fernández-Laso, M.C., 2010. Remontajes de restos faunísticos y relaciones entreáreas domésticas en los niveles K, L y M del Abric Romaní (Capellades, Barce-lona, España). Ph D. Dissertation. Història i Història de l’Art. Universitat Rovira iVirgili, Tarragona.
Fisher, J.W., Strickland, H.C., 1991. Dwellings and fireplaces: keys to Efe Pygmycampsite structure. In: Gamble, C.S., Boismier, W. (Eds.), EthnoarchaeologicalApproaches to Mobile Campsites. International Monographs in Prehistory,Michigan, pp. 215e236.
Gaudzinski, S., Roebroeks, W., 2000. Adults only. reindeer hunting at the MiddlePalaeolithic site Salzgitter Lebenstedt, Northern Germany. Journal of HumanEvolution 38, 497e521.
Gaudzinski, S., 2000. On the variability of Middle Palaeolithic procurement tactics:the case of Salzgitter Lebenstedt, Northern Germany. International Journal ofOsteoarchaeology 10, 396e406.
Giralt, S., Julià, R., 1996. The sedimentary record of the Middle-Upper Palaeolithictransition in the Capellades area (NE. Spain). In: Carbonell, E., Vaquero, M.(Eds.), The Last Neandertals/The First Anatomically Modern Humans. CulturalChange and Human Evolution: The Crisis at 40 ka BP, pp. 365e376. Igualada,Barcelona, Spain.
Gómez de Soler, B., 2009. Àrees de captació, tecnologia lítica i estratègies d’apro-visionament de roques silícies en el nivell L de l’Abric Romaní (Capellades,Barcelona). Quadern de Prehistòria Catalana 17, 11e56.
Guadelli, J.L., 1998. Détermination de l’age des chevaux fossiles et établissement desclasses d’age. Paleo 10, 87e93.
Hammond, G., Hammond, N., 1981. Child’s play: a distorting factor in archaeologicaldistribution. American Antiquity 46 (3), 634e636.
Haynes, G., 1980. Evidence of carnivore gnawing on Pleistocene and recentmammalian bones. Paleobiology 6, 341e351.
Haynes, G., 1983. A guide for differentiating mammalian carnivore taxa responsiblefor gnaw damage to herbivore limb bones. Paleobiology 9, 164e172.
Henry, D.O., Hall, S.A., Hietala, H.J., Demidenko, Y.E., Usik, V.I., Rosen, A.M.,Thomas, P.A., 1996. Middle Paleolithic behavioral organization: 1993 excavationof Tor Faraj, Southern Jordan. Journal of Field Archaeology 23 (1), 31e53.
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
Hietala, H.J., Stevens, D.E., 1977. Spatial analysis: multiple procedures in patternrecognition studies. American Antiquity 42 (4), 539e559.
Hillson, S., 1992. Mammal Bones and Teeth: An Introductory Guide to Methods ofIdentification. Institute of Archaeology, University College London, London.
Johnson, W.G., Sharpe, S.E., Bullard, T.F., Lupo, K., 2005. Characterizing a firstoccurrence of bison deposits in Southeastern Nevada. Western North AmericanNaturalist 65, 24e35.
Kamp, K.A., 2001. Where have all the children gone?: the archaeology of childhood.Journal of Archaeological Method and Theory 8 (1), 1e34.
Karkanas, P., Shahack-Gross, R., Ayalon, A., Bar-Matthews, M., Barkai, R., Frumkin, A.,Gopher, A., Stiner, M.C., 2007. Evidence for habitual use of fire at the end of theLower Paleolithic: site-formation processes at Qesem Cave, Israel. Journal ofHuman Evolution 53, 197e212.
Lam, Y.M., Chen, X., Marean, C.W., Frey, C.J., 1998. Bone density and long bonerepresentation in archaeological faunas: comparing results from CT and Photondensiometry. Journal of Archaeological Science 25, 559e570.
Lyman, R.L., 1985. Bone frequencies: differential survivorship of fossil classes.Journal of Archaeological Science 12, 221e236.
Lyman, R.L., 1994. Vertebrate Taphonomy. Cambridge University Press, Cambridge.Lyman, R.L., 2008. Archaeological refitting a consideration of methods and analyt-
ical search radius. Journal of Anthropological Research 64, 229e248.Marean, C.W., Cleghorn, N., 2003. Large mammal skeletal element transport:
applying foraging theory in a complex taphonomy system. Journal ofTaphonomy 1 (1), 15e42.
Marean, C.W., Kim, S.Y., 1998. Mousterian large-mammal remains from KobehCave.Behavioral implications for Neanderthals and Early Modern Humans. CurrentAnthropology 39 (Suppl.), 79e113.
Marean, C.W., Spencer, L., 1991. Impact of carnivore ravaging on zooarchaeologicalmeasures of element abundance. American Antiquity 56, 645e658.
Marean, C.W., Spencer, L.M., Blumenschine, R.J., Capaldo, S.D., 1992. Captive hyenabone choice and destruction, the schlepp effect, and Olduvai Gorge archae-ofaunas. Journal of Archaeological Science 18, 101e121.
Mariezkurrena, K., Altuna, J., 1983a. Contribución al conocimiento del desarrollo dela dentición y el esqueleto postcraneal de Cervus elaphus. Munibe 35, 149e202.
Mariezkurrena, K., Altuna, J., 1983b. Biometría y dimorfismo sexual en el esqueletode Cervus elaphus würmiense, postwürmiense y actual del Cantabrico. Munibe35, 203e246.
Martínez, K., Rando, J.M., (2000). Organización espacial y de la producción lítica enel desarrollo de las actividades durante ocupaciones del Paleolítico medio. NivelJa del Abric Romaní(Capellades, Barcelona)Actas do 3�Congresso de Arqueolo-gia Peninsulr, vol. II, Oporto. pp215e234
Morin, E., Tsanova, T., Sirakov, N., Rendu, W., Mallye, J.-B., Lévêque, F., 2005. Bonerefits in stratified deposits: testing the chronological grain at Saint-Césaire.Journal of Archaeological Science 32, 1083e1098.
Nicholson, R.A., 1993. A morphological investigation of burnt animal bone andan evaluation of its utility in Archaeology. Journal of Archaeological Science20, 411e428.
Noe-Nygaard, N., 1989. Man-made trace fossils on bones. Journal of HumanEvolution 4 (6), 461e491.
O’Connell, J.F., 1987. Alyawara site structure and its archaeological implications.American Antiquity 52 (1), 74e108.
Oliver, J.S., 1993. Carcass processing by the Hadza: bone breakage from butchery toconsumtion. In: Hudson, J. (Ed.), From Bones to Behavior. Ethnoarchaelogicaland Experimental Contributions to the Interpretation of Faunal Remains.Occasional Paper, vol. 21. Southern Illinois University at Carbondale, Center forArchaeological Investigations, pp. 200e227.
Patou-Mathis, M., 2006. Comportements de subsistence des Néandertaliens d’Eu-rope. In: Demansin, B., Otte, M. (Eds.), Neanderthals in Europe. Proceedings ofthe International Conference held in the Gallo-Roman Museum in Tougereu(September 17e19th 2004), vol. 117, pp. 67e76. Liège, Eraul.
Pickering, T.R., Egeland, C.P., 2006. Experimental patterns of hammerstonepercussion damage on bones: implications for inferences of carcass processingby humans. Journal of Archaeological Science 33, 459e469.
Politis, G.G., 2005. Children’s activity in the production of the archaeological record ofhunter-gatherers. An ethnoarchaeological approach. In: Funari, P.P., Zarankin, A.,Stovel, E. (Eds.), Global Archaeology Theory, I. Springer, pp. 121e143.
Potts, R., Shipman, P., 1981. Cutmarks made by stone tools on bones from OlduvaiGorge, Tanzania. Nature 291, 577e580.
Preece, R.C., Gowlett, J.A., Parfitt, K., Bridgland, D.R., Lewis, S.G., 2006. Humans inthe Hoxnian: habitat, context and fire use at Beeches Pit, West Stow, Suffolk,UK. Journal of Quaternary Science 21 (5), 485e496.
Rapson, D.J., Todd, L.C., 1987. Attribute based spatial analysis: an example froma hunter-gatherer site in Northwestern Wyoming. 52nd Annual Meeting ofSociety for American Archaeology, Toronto.
Reitz, E.J., Wing, E.S., 1999. Zooarchaeology. CambridgeUniversity Press, New York.Rosell, J., Blasco, R., 2008. La presencia de carnívoros en conjuntos antrópicos del
Pleistoceno Medio: El caso del nivel TD10-Sup de Gran Dolina (Sierra de Ata-puerca, Burgos) y del nivel XII de la Cova del Bolomor (La Valldigna, Valencia).In: Díez, J.C. (Ed.), Zooarqueología Hoy. Encuentros Hispano-Argentinos. Uni-versidad de Burgos, Burgos, pp. 43e58.
Rosell, J., Blasco, R., 2009. Home sharing: carnivores in anthropogenic assemblagesof the Middle Pleistocene. Journal of Taphonomy 7 (4), 305e324.
Rosell, J., 2001. Patrons d’Aprofitament de les Biomasses Animals durant el PleistocèInferior i Mig (Sierra de Atapuerca, Burgos) i Superior (Abric Romaní, Barce-lona). Història i Geografia, Universitat Rovira i Virgili, Tarragona, 329 p.
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020

J. Rosell et al. / Quaternary International xxx (2011) 1e1616
Selvaggio, M.M., Wilder, J., 2001. Identifying the involvement of multiple carnivoretaxa with archaeological bone assemblages. Journal of Archaeological Science28, 465e470.
Shea, J.J., 2006. Child’s play: reflections on the invisibility of children in thePaleolithic record. Evolutionary Anthropology 15, 212e216.
Shipman, P., Rose, J., 1983. Early hominid hunting, butchering and carcass-pro-cessing behaviors: approaches to the fossil record. Journal of AnthropologicalArchaeology 2, 57e98.
Shipman, P., Fisher, D.C., Rose, J., 1984a. Mastodon butchery: microscopic evidenceof carcass processing and bone tool use. Paleobiology 10 (3), 358e365.
Shipman, P., Foster, G.F., Schoeninger, M., 1984b. Burnt bones and teeth: andexperimental study of colour, morphology, crystal structure and shrinkage.Journal of Archaeological Science 11, 307e325.
Shipman, P., 1983. Early hominid livestyle: hunting and gathering or foraging andscavenging? In: Clutton-Brock, J., Grigson, C. (Eds.), Animals and ArchaeologyHunters and Their Prey, vol. 1, pp. 31e49. Oxford, BAR International Series163.
Stiner, M.C., Kuhn, S.L., Weiner, S., Bar-Yosef, O., 1995. Differential burning, recrys-tallization, and fragmentation of archaeological bones. Journal of ArchaeologicalScience 22, 223e237.
Stiner, M.C., 1994. Honor Among Thieves: A Zooarchaeological Study of NeandertalEcology. Princeton University Press, Princeton.
Stiner, M.C., Barkai, R., Gopher, A., 2009. Cooperative hunting and meat sharing400-200 kya at Qesem Cave, Israel. Proceedings of the National Academy ofSciences 106 (32), 13207e13212.
Todd, L.C., Frison, G.C.,1992. Reassembly of bison skeletons from theHorner site: a studyinanatomical refitting. In:Hofman, J.L., Enloe, J.G. (Eds.), Putting thePieces Together:An Introduction to Refitting. BAR International Series, vol. 578, pp. 63e82.
Vallverdú, J., Allué, E., Bischoff, J.L., Cáceres, I., Carbonell, E., Cebrià, A., Garciá-Antón,Huguet, R., Ibáñez, N.,Martínez, K., Pastó, I., Rosell, J., Saladié, P., Vaquero, M., 2005a.
Please cite this article in press as: Rosell, J., et al., A zooarchaeological con(Barcelona, Spain), Quaternary International (2011), doi:10.1016/j.quaint.
Shorthumanoccupations in theMiddlePalaeolithic Level I of theAbricRomaní rock-shelter (Capellades, Barcelona, Spain). Journal of Human Evolution 48, 157e174.
Vallverdú, J., Allué, E., Cáceres, I., Carbonell, E., Cebrià, A., Huguet, R., Ibáñez, N.,Martínez, K., Pastó, I., Rosell, J., Saladié, P., Vaquero, M., 2005b. Modelos deocupación de corta duración en el nivel I del Abric Romaní (Capellades, Bar-celona, España). Zephyrus 57, 97e114.
Vallverdú, J., Vaquero, M., Cáceres, I., Allué, E., Rosell, J., Saladié, P., Chacón, G.,Ollé, A., Canals, A., Sala, R., Courty, M.A., Carbonell, E., 2010. Sleeping activityarea within the site structure of archaic human groups. Evidence from AbricRomaní Level N combustion activity areas. Current Anthropology 51, 137e145.
Vaquero, M., Pastó, I., 2001. The definition of spatial units in Middle Palae-olithic sites: the hearth-related assemblages. Journal of ArchaeologicalScience 28, 1209e1220.
Vaquero, M., Vallverdú, J., Rosell, J., Pastó, I., Allué, E., 2001. Neandertal behavior atthe Middle Palaeolithic site of Abric Romaní, Capellades, Spain. Journal of FieldArchaeology 28 (1e2), 93e114.
Vaquero, M., 1997. Tecnología Lítica y Comportamiento Humano: Organización delas Actividades Técnicas y Cambio Diacrónico en el Paleolítico Medio del AbricRomaní (Capellades, Barcelona). Universitat Rovira i Virgili, Tarragona.
Villa, P., Bartram, L., 1996. Flaked bone from a hyena den. Paléo 8, 143e159.Villa, P., Mahieu, E., 1991. Breakage patterns of human long bones. Journal of Human
Evolution 21, 27e48.Villa, P., Soressi, M., 2000. Stone tools in carnivore sites: the case of Bois Roche.
Journal of Anthropological Research 56 (2), 187e215.Waguespack, N.M., 2002. Caribou sharing and storage: refitting the Palangana site.
Journal of Anthropological Archaeology 21, 396e417.White, T.D., 1992. Prehistoric Cannibalism at Mancos 5MTURM-2346. Princeton
University Press, Princeton.Yellen, J.E., 1977. Archaeological Approaches to the Present: Models for Recon-
structing the Past. Academic Press, New York.
tribution to establish occupational patterns at Level J of Abric Romaní2011.01.020


















