
A survey of mangiferin and hydroxycinnamic acid ester accumulation in coffee (Coffea) leaves:...
19
A survey of mangiferin and hydroxycinnamic acid ester accumulation in coffee (Coffea) leaves: biological implications and uses Claudine Campa 1, *, Laurence Mondolot 2 , Arsene Rakotondravao 3 , Luc P. R. Bidel 4 , Annick Gargadennec 2 , Emmanuel Couturon 5 , Philippe La Fisca 2 , Jean-Jacques Rakotomalala 3 , Christian Jay-Allemand 6 and Aaron P. Davis 7 1 IRD, UMR DIADE (IRD/UM2), BP 64501, 34394 Montpellier, France, 2 Faculte ´ de Pharmacie, UMR 5175 CEFE-CNRS, BP 14491, 34093 Montpellier, France, 3 Laboratoire de Biochimie, FOFIFA, Antananarivo, Madagascar, 4 INRA, UMR DIADE (IRD/UM2), BP 64501, 34394 Montpellier, France, 5 IRD, UMR DIADE (IRD/UM2), Po ˆle de Protection des Plantes, 97410 Saint Pierre, La Re ´union, France, 6 UM2, UMR DIADE (IRD/UM2), BP 64501, 34394 Montpellier, France and 7 Royal Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK * For correspondence. E-mail [email protected] Received: 26 January 2012 Returned for revision: 5 March 2012 Accepted: 12 April 2012 Published electronically: 13 June 2012 † Background and Aims The phenolic composition of Coffea leaves has barely been studied, and therefore this study conducts the first detailed survey, focusing on mangiferin and hydroxycinnamic acid esters (HCEs). † Methods Using HPLC, including a new technique allowing quantification of feruloylquinic acid together with mangiferin, and histochemical methods, mangiferin content and tissue localization were compared in leaves and fruits of C. pseudozanguebariae, C. arabica and C. canephora. The HCE and mangiferin content of leaves was evaluated for 23 species native to Africa or Madagascar. Using various statistical methods, data were assessed in relation to distribution, ecology, phylogeny and use. † Key Results Seven of the 23 species accumulated mangiferin in their leaves. Mangiferin leaf-accumulating species also contain mangiferin in the fruits, but only in the outer (sporophytic) parts. In both leaves and fruit, mangiferin accumulation decreases with ageing. A relationship between mangiferin accumulation and UV levels is posited, owing to localization with photosynthetic tissues, and systematic distribution in high alti- tude clades and species with high altitude representatives. Analyses of mangiferin and HCE content showed that there are significant differences between species, and that samples can be grouped into species, with few excep- tions. These data also provide independent support for various Coffea lineages, as proposed by molecular phylo- genetic analyses. Sampling of the hybrids C. arabica and C. heterocalyx cf. indicates that mangiferin and HCE accumulation may be under independent parental influence. † Conclusions This survey of the phenolic composition in Coffea leaves shows that mangiferin and HCE accu- mulation corresponds to lineage recognition and species delimitation, respectively. Knowledge of the spectrum of phenolic accumulation within species and populations could be of considerable significance for adaptation to specific environments. The potential health benefits of coffee-leaf tea, and beverages and masticatory products made from the fleshy parts of Coffea fruits, are supported by our phenolic quantification. Key words: Arabica coffee, C. arabica, C. canephora, chlorogenic acids, Crop Wild Relatives (CWRs), coffee- leaf tea, hybridization, hydroxycinnamic acids, mangiferin, phenolic compounds, phylogeny, robusta coffee. INTRODUCTION Coffee (Coffea) is the second most traded commodity after oil, accounting for exports worth an estimated US$15 . 4 billion in 2009/10, when some 93 . 4 million bags were shipped [International Coffee Organization (ICO), 2011], and has an estimated annual retail value exceeding US$70 billion (Vega et al., 2003; Lewin et al., 2004). In 2010, total coffee sector employment was estimated at about 26 million people in 52 producing countries [International Coffee Organization (ICO), 2011]. Coffea arabica L. (Arabica coffee) and C. canephora Pierre ex A. Froehner (robusta coffee) are the two species used in the production of coffee, providing approx. 70 and 30 % of commercial production, respectively [International Coffee Organization (ICO), 2011]. Due to their economic importance, the biochemical composition of Arabica and robusta coffee beans (i.e. seeds; see Materials and Methods) has been extensively studied, because of their implication in taste and cup quality (Belay, 2011). Less atten- tion has been paid to other Coffea species, of which there are 124 occurring naturally in the Old Word (Davis et al., 2006, 2011; Davis, 2011). Several biochemical studies have focused on wild species, particularly those from Africa and Madagascar, although these works have generally focused on qualitative and quantitative assessments of sugar, lipid, caf- feine and esters of hydroxycinnamic acid (HCEs) in green (unroasted) coffee beans (Clifford et al., 1989; Anthony et al., 1993; Campa et al., 2004, 2005; Dussert et al., 2008). In contrast to the considerable amount of research on green beans, there are relatively few studies concerned with the me- tabolite content of the other parts of the coffee plant, such as the leaves (Mondolot et al., 2006; Clifford et al., 2008), the # The Author 2012. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved. For Permissions, please email: [email protected] Annals of Botany 110: 595–613, 2012 doi:10.1093/aob/mcs119, available online at www.aob.oxfordjournals.org by guest on June 3, 2016 http://aob.oxfordjournals.org/ Downloaded from
Transcript of A survey of mangiferin and hydroxycinnamic acid ester accumulation in coffee (Coffea) leaves:...

A survey of mangiferin and hydroxycinnamic acid ester accumulation in coffee(Coffea) leaves: biological implications and uses
Claudine Campa1,*, Laurence Mondolot2, Arsene Rakotondravao3, Luc P. R. Bidel4, Annick Gargadennec2,Emmanuel Couturon5, Philippe La Fisca2, Jean-Jacques Rakotomalala3, Christian Jay-Allemand6
and Aaron P. Davis7
1IRD, UMR DIADE (IRD/UM2), BP 64501, 34394 Montpellier, France, 2Faculte de Pharmacie, UMR 5175 CEFE-CNRS,BP 14491, 34093 Montpellier, France, 3Laboratoire de Biochimie, FOFIFA, Antananarivo, Madagascar, 4INRA, UMR DIADE
(IRD/UM2), BP 64501, 34394 Montpellier, France, 5IRD, UMR DIADE (IRD/UM2), Pole de Protection des Plantes,97410 Saint Pierre, La Reunion, France, 6UM2, UMR DIADE (IRD/UM2), BP 64501, 34394 Montpellier, France and 7Royal
Botanic Gardens, Kew, Richmond, Surrey TW9 3AB, UK* For correspondence. E-mail [email protected]
Received: 26 January 2012 Returned for revision: 5 March 2012 Accepted: 12 April 2012 Published electronically: 13 June 2012
† Background and Aims The phenolic composition of Coffea leaves has barely been studied, and therefore thisstudy conducts the first detailed survey, focusing on mangiferin and hydroxycinnamic acid esters (HCEs).† Methods Using HPLC, including a new technique allowing quantification of feruloylquinic acid together withmangiferin, and histochemical methods, mangiferin content and tissue localization were compared in leaves andfruits of C. pseudozanguebariae, C. arabica and C. canephora. The HCE and mangiferin content of leaves wasevaluated for 23 species native to Africa or Madagascar. Using various statistical methods, data were assessed inrelation to distribution, ecology, phylogeny and use.† Key Results Seven of the 23 species accumulated mangiferin in their leaves. Mangiferin leaf-accumulatingspecies also contain mangiferin in the fruits, but only in the outer (sporophytic) parts. In both leaves andfruit, mangiferin accumulation decreases with ageing. A relationship between mangiferin accumulation andUV levels is posited, owing to localization with photosynthetic tissues, and systematic distribution in high alti-tude clades and species with high altitude representatives. Analyses of mangiferin and HCE content showed thatthere are significant differences between species, and that samples can be grouped into species, with few excep-tions. These data also provide independent support for various Coffea lineages, as proposed by molecular phylo-genetic analyses. Sampling of the hybrids C. arabica and C. heterocalyx cf. indicates that mangiferin and HCEaccumulation may be under independent parental influence.† Conclusions This survey of the phenolic composition in Coffea leaves shows that mangiferin and HCE accu-mulation corresponds to lineage recognition and species delimitation, respectively. Knowledge of the spectrumof phenolic accumulation within species and populations could be of considerable significance for adaptation tospecific environments. The potential health benefits of coffee-leaf tea, and beverages and masticatory productsmade from the fleshy parts of Coffea fruits, are supported by our phenolic quantification.
Key words: Arabica coffee, C. arabica, C. canephora, chlorogenic acids, Crop Wild Relatives (CWRs), coffee-leaf tea, hybridization, hydroxycinnamic acids, mangiferin, phenolic compounds, phylogeny, robusta coffee.
INTRODUCTION
Coffee (Coffea) is the second most traded commodity after oil,accounting for exports worth an estimated US$15.4 billion in2009/10, when some 93.4 million bags were shipped[International Coffee Organization (ICO), 2011], and has anestimated annual retail value exceeding US$70 billion (Vegaet al., 2003; Lewin et al., 2004). In 2010, total coffee sectoremployment was estimated at about 26 million people in 52producing countries [International Coffee Organization(ICO), 2011]. Coffea arabica L. (Arabica coffee) andC. canephora Pierre ex A. Froehner (robusta coffee) are thetwo species used in the production of coffee, providingapprox. 70 and 30 % of commercial production, respectively[International Coffee Organization (ICO), 2011]. Due totheir economic importance, the biochemical composition of
Arabica and robusta coffee beans (i.e. seeds; see Materialsand Methods) has been extensively studied, because of theirimplication in taste and cup quality (Belay, 2011). Less atten-tion has been paid to other Coffea species, of which there are124 occurring naturally in the Old Word (Davis et al., 2006,2011; Davis, 2011). Several biochemical studies havefocused on wild species, particularly those from Africa andMadagascar, although these works have generally focused onqualitative and quantitative assessments of sugar, lipid, caf-feine and esters of hydroxycinnamic acid (HCEs) in green(unroasted) coffee beans (Clifford et al., 1989; Anthonyet al., 1993; Campa et al., 2004, 2005; Dussert et al., 2008).In contrast to the considerable amount of research on greenbeans, there are relatively few studies concerned with the me-tabolite content of the other parts of the coffee plant, such asthe leaves (Mondolot et al., 2006; Clifford et al., 2008), the
# The Author 2012. Published by Oxford University Press on behalf of the Annals of Botany Company. All rights reserved.
For Permissions, please email: [email protected]
Annals of Botany 110: 595–613, 2012
doi:10.1093/aob/mcs119, available online at www.aob.oxfordjournals.org
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

outer fleshy layers of the fruit (Lopes and Monaco, 1979;Lopes et al., 1984; Lopes and Shepherd, 1991), and vascular-ized organs (Mahesh et al., 2007). Moreover, many of thesestudies are still focused on the crop species, and most exclu-sively on HCE content, when analysing phenolic compounds(Lepelley et al., 2007). Other than Arabica and robustacoffee, only one wild species, C. pseudozanguebariaeBridson, has been studied for its leaf phenolic content. Highquantities of esters of feruloylquinic acid (FQA) were firstdescribed in fruit, green beans and leaves of this species(Bertrand et al., 2003), and then a C-glucosylxanthone, mangi-ferin, was isolated from the leaves (Talamond et al., 2008).This compound, which was initially isolated from theleaves, bark and peel of mango [Mangifera indicaL. (Anacardiaceae)] (Baretto et al., 2008), is well known forits numerous pharmacological properties such as anti-inflammatory, antidiabetic, antihyperlipidaemic and neuropro-tective activities (Garrido et al., 2004; Muruganandan et al.,2005; Campos-Esparza et al., 2009). In plants, mangiferin pro-vides antioxidant and antimicrobial protection upon bioticstress (Franklin et al., 2009). More generally, phenolics areinvolved in the response to biotic and abiotic stresses(Santiago et al., 2000; Read et al., 2009), mainly due totheir antioxidant properties (Moglia et al., 2008; Shahidiet al., 2010; Gallego-Giraldo et al., 2011). In this way, theymay play a role in adaptation to environmental change(Boudet, 2007) and in coevolution with pests and diseases(Orians, 2000; Eyles et al., 2010). Phenolics could be criticalto understanding plant–animal interactions, including thoseinvolved in the control of significantly destructive pests.Phenolics also have the potential to elucidate evolutionaryhistory when scored as characters in phylogenetic analyses,as undertaken using other secondary metabolites (Wink,2003; Dussert et al., 2008).
Although hydroxycinnamic acids ( p-coumaric, caffeic,ferulic and sinapic acids) exist as a free form in cells, theyusually occur in conjugated forms. The most common formsare glycosylated derivatives (Herrmann, 1989; Macheixet al., 1990) and esters of triterpenes (Bolzani et al., 1991)or quinic acid, shikimic acid or tartric acid (Ribereau-Gayon,1968; Schuster and Herrmann, 1985; Manach et al., 2004),named HCEs. The esters formed between hydroxycinnamicacids and quinic acid are grouped under the generic name ofchlorogenic acids, one of the most widespread in the plantkingdom being 5-O-caffeoylquinic acid (5-CQA), also calledchlorogenic acid.
The aim of this study was to undertake a survey of the accu-mulation of phenolic compounds in the leaves of Coffeaspecies, focusing on mangiferin and HCEs, in order to: (1)better understand the physical and temporal location of mangi-ferin within Coffea leaves, and make a comparison with accu-mulation in other plant tissues; (2) test for correlations betweenthe accumulation of mangiferin and HCEs in leaf tissue; (3)assess the geographical and systematic (phylogenetic) distribu-tion of leaf-accumulated mangiferin and HCEs within Coffealeaf tissue; and (4) briefly investigate the relationshipsbetween hybridization and phenolic accumulation (mangiferinand HCEs) in Coffea leaves. The first step in achieving theseobjectives was to develop a new high-performance liquid chro-matography (HPLC) procedure to quantify mangiferin together
with FQA. Identification of chemical structures was performedby liquid chromatography–mass spectrometry (LC-MS) ana-lyses on a leaf extract of C. pseudozanguebariae, the firstcoffee species elucidated as mangiferin accumulating(Talamond et al., 2008). Mangiferin and HCE content wasthen evaluated in the leaves of 23 Coffea species (24 taxa), in-cluding two hybrids (C. arabica ‘Laurina’ and C. heterocalyxStoff. cf.), originating from different localities in Africa andMadagascar (Table 1), the main centres of Coffea species di-versity (Davis et al., 2006, 2011; Davis, 2011). The fruits ofthree species were tested for the temporal and physical accu-mulation of mangiferin.
MATERIALS AND METHODS
Taxon sampling and plant material
The level of accumulation of HCEs and mangiferin was con-comitantly studied. Experiments were performed on theleaves of 23 Coffea species (Table 1), representing 40African genotypes and nine Madagascan genotypes (one geno-type per species). Genotypes represent individuals taken aswhole plants, seedlings or cuttings from wild populations orcultivation, and gathered under a common accession prefixwith different numbers. The biosynthesis of secondary meta-bolites is largely dependent on environmental conditions,and so we used trees grown under near-identical culture condi-tions for the African and Madagascan genotypes. In thismanner, a valid estimation of the genetic effect on thespecies diversity of phenolic compounds was possible. Forthe African species, leaves were collected from two or threegenotypes for each species grown in tropical greenhouses(natural daylight, 25 8C night and 28 8C day temperature,78–82 % relative humidity) at the IRD research centre inMontpellier (France), to replicate the same environment asfound in Madagascar and Reunion (see below). For theMadagascan species, leaves were harvested from one treeper species grown at the FOFIFA coffee research stationof Kianjavato (Madagascar), where trees are grown in theopen ground. Only fully expanded leaves, from the secondnode below the apex of the growing shoot, were takenfor the study of phenolic diversity. To study mangiferincontent during leaf development, four species were used:C. arabica, C. canephora, C. eugenioides S. Moore andC. pseudozanguebariae. Leaves were sampled at three devel-opmental stages: stage 1, young leaves from the apex; stage2, leaves from the first node below the apex; and stage 3,leaves from the second node below the apex. Immediatelyafter harvesting, leaves were weighed and then frozen inliquid nitrogen before being lyophilized for extraction. Fruitsfrom C. arabica were harvested at Reunion Island (IRD collec-tion). All the C. arabica samples used in this study were repre-sented by the cultivar ‘Laurina’ (Bourbon Pointu). Althoughthis entity is a cultivated mutant, the sequence profile of thespecies, based on the same markers as Maurin et al. (2007),is the same as wild origin material (A. Davis, unpubl. data).Coffea pseudozanguebariae and C. canephora fruits wereobtained from the CNRA station (Divo, Ivory Coast). Fruitswere sampled at three developmental stages: stage 1 (length,0.7 cm), fruit green with a partially formed seed (immature);
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea596
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
stage 2 (length ,1.2 cm), fruit yellowish green, pericarp(exocarp, mesocarp, endocarp) easily detached from the seed(semi-mature); and stage 3 (length .1.2 cm), fruit reddishyellow, pericarp easily detached from the seed (mature).Except for stage 1, the pericarp was isolated from the seedand each part was pooled (up to five fruits per species, perstage), weighed, and frozen in liquid nitrogen until extraction.Coffee beans are the seeds of the coffee fruit, the bulk ofwhich is made up of endosperm. The fruit skin (exocarp),the fleshy part of the fruit (mesocarp), and the parchment,which is the crustaceous pyrene coat (endocarp), are allremoved during processing.
Phenolic extraction
Extraction was performed using sonication (20 min, roomtemperature, 24 kHz, R.E.U.S-GEX 180, Contes, France) in6 mL of MeOH/H2O (80:20, v/v) of 25 mg of plant material(lyophilized leaves or frozen fruits ground under liquid nitro-gen with a mortar and pestle). After centrifugation (5 min,5000 rpm), the methanol extract was collected and filtered(Millipore, 0.25 mm porosity) before analysis. Each extractionwas realized in triplicate for leaves and fruit samples. Eachsample was characterized by its mean content of mangiferinand HCEs, expressed as a percentage of fresh or dry weight.
TABLE 1. Origin of the species studied and provenance of the genotypes analysed
Species Species rangeClade designation (and alliance, if
applicable)Genotype: native origin/germplasmcollection (accession number/code)
From AfricaC. anthonyi Stoff. & F. Anthony Cameroon, Congo EC-Afr Cameroon/IRD Montpellier (OD
71, OD 72)C. arabica L. ‘Laurina’ [BourbonPointu]
Ethiopian, SE South Sudan Hybrid origin: EC-Afr(C. eugenioides) × LG (C. canephora)
Ethiopia/IRD La Reunion (Ar 27,Ar 34, Ar 39)
C. canephora Pierre ex A. Froehner Upper Guinea Region and LowerGuinea/Congolian Region
LG/C [canephora alliance] Cameroon (BD)/IRD Montpellier(BD 62, BD 66)
C. canephora Pierre ex A. Froehner Upper Guinea Region and LowerGuinea/Congolian Region
LG/C [canephora alliance] Congo (BB)/IRD Montpellier (BB62, BB 66, BB 67)
C. congensis A. Froehner Lower Guinea/Congolian Region LG/C [canephora alliance] Cameroon/IRD Montpellier (CC61, CC 66, CC70)
C. eugenioides S. Moore Burundi, Rwanda, Zaire, southernSouth Sudan, Kenya, Tanzania,Uganda
EC-Afr Tanzania/IRD Montpellier (DA 75,DA 77)
C. heterocalyx Stoff. cf. SW Cameroon Hybrid origin: × EC-Afr(C. eugenioides) × C. liberica
Central Africa/IRD Montpellier(JC 8, JC 62, JC 69)
C. humilis A. Chev. Sierra Leone, Liberia, SW Ivory Coast UG Ivory Coast/IRD Montpellier (G52, G 68, G 72)
C. liberica var. dewevrei (De Wild.& T.Durand) Lebrun
Lower Guinea/Congolian Region LG/C [liberica alliance] Central African Republic/IRDMontpellier (EB 52, EB 63)
C. liberica var. liberica Bull. exHiern
Upper Guinea Region and LowerGuinea/Congolian Region
LG/C [liberica alliance] Ivory Coast/IRD Montpellier (EA51, EA 61, EA 63)
C. mannii (Hook.f) A.P. Davis Upper Guinea Region and LowerGuinea/Congolian Region
‘African Psilanthus’ Ivory Coast/IRD Montpellier(PSI-man)
C. pseudozanguebariae Bridson Kenya, Tanzania, Mozambique EA [2] Tanzania/IRD Montpellier [viaDivo, Cote d’Ivoire] (H 58, H 63)
C. racemosa Lour. Mozambique, Mozambique ChannelIs., KwaZulu-Natal, Zimbabwe
EA [3] Mozambique/IRD Montpellier (IB58, IB 58, IB 62)
C. salvatrix Swynn. & Phillipson Malawi, Mozambique, Tanzania,Zimbabwe
EA [2] Mozambique/IRD Montpellier (LB57, LB 63)
C. sessiliflora Bridson Kenya, Tanzania, EA [3] Mozambique/IRD Montpellier (PA58, PA 67, PB 58)
C. stenophylla G. Don Guinea, Ivory Coast, Sierra Leone UG Ivory Coast/IRD Montpellier (FB53, FB 54, FB 61)
From MadagascarC. leroyi A.P. Davis Eastern Madagascar IO [Madagascan Coffea] Kianjavato (A.315)C. andrambovatensis J.-F. Leroy Eastern Madagascar IO [Madagascan Coffea] Kianjavato (A.227)C. ankaranensis A.P. Davis &Rakotonas.
NE Madagascar IO [Madagascan Coffea] Kianjavato (A 808)
C. augagneuri Dubard Northern Madagascar IO [Madagascan Coffea] Kianjavato (A.519)C. millotii J.-F. Leroy Eastern Madagascar IO [Madagascan Coffea] Kianjavato (A.219)C. perrieri Drake ex Jum. &H. Perrier
Western and southern Madagascar IO [Madagascan Coffea] Kianjavato (A.305)
C. resinosa (Hook.f.) Radlk. Eastern Madagascar IO [Madagascan Coffea] Kianjavato (A.915)C. tsirananae J.-F. Leroy Northern Madagascar IO [Madagascan Coffea] Kianjavato (A.515)C. vohemarensis A.P. Davis &Rakotonas
Northern Madagascar IO [Madagascan Coffea] Kianjavato (A.977)
Species range after Davis et al. (2006); clade designation, terminology and alliance after Maurin et al. (2007) and Davis et al. (2011). Clade terminologyabbreviations: EA, East African clade (numbers in square brackets denote the three major clades within the EA clade); EC-Afr, East Central African clade;IO, Indian Ocean clade; LG/C, Lower Guinea/Congolian clade; UG, Upper Guinea clade. Region designation (i.e. Upper Guinea Region and Lower Guinea/Congolian Region) after White (1979, 1983).
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 597
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Electrospray-mass spectrometry analysis of the samples
Electrospray-mass spectrometry (ESI-MS) was performedon a binary HPLC system (Waters 1525 m, Waters,Manchester, UK) coupled with a Waters Micromass ZQESCi multimode ionization mass spectrometer (MicromassLtd, Manchester, UK). Separation was performed on anXTerra MS C18 column (3.5 mm particle size, 2.1 mm ×100 mm) kept at 40 8C in a thermostatic oven (Gecko 2000,Cluzeau Info Labo, France) with a binary mobile phase gradi-ent delivered at a total flow rate of 210 mL min21. The mobilephase was composed of permuted water (solvent A) and aceto-nitrile (solvent B), both phases acidified by 0.1 % (v/v) formicacid in order to prevent the ionization of phenolic acids. Thegradient profile evolved linearly from 5 % B to 40 % B in40 min, increased exponentially to 100 % B from 40 to50 min. After 5 min of isocratic elution at 100 % B, themobile phase composition returned to 5 % B with 5 minre-equilibration.
The source and capillary were heated at 80 and 450 8C, re-spectively, and the capillary voltage was set to 3.0 kV.Nitrogen was used as the desolvatation gas (400 L h21) andcone gas (50 L h21). Spectra were acquired and accumulatedover 60 s in the full scan mode over the m/z 50–1200 range,in both the negative and positive modes at cone voltages of–25, –60, +25 and 60 V, successively. Absorbance spectraover the range of 210–800 nm were acquired using a Waters996 Photodiode Array Detector. Absorbance and massspectra were handled using MassLynx 3.5 software(Micromass Ltd). The sensitivity of the mass spectrometerwas optimized using the chlorogenic acid standard.5-O-caffeoylquinic acid, mangiferin and 5-metoxyflavonewere provided by Sigma-Aldrich (Buchs, Switzerland).
Clifford et al. (2008) showed that Coffea leaves had a pro-portionately greater content of cis-isomers relative totrans-isomers, compared with coffee beans, suggesting thatUV-irradiation in vivo may also cause geometric isomeriza-tion. In our study, we noticed the presence of cis-isomers, es-pecially in leaves of cultivated species grown outdoors duringdaylight hours, and returned to greenhouses after dark. In thesecases cis-isomers never exceeded 10 % of the CGA content,and we decided to ignore these compounds for our studybecause: (1) most of the samples used in this study (allexcept those from Madagascar) were cultivated in greenhouseswith very little or no UVb, and correspondingly very lowquantities of cis-isomers; and (2) the Madagascan samples,which are cultivated outdoors, have CGAs in such low quan-tities that the cis-isomers are not quantifiable.
Phenolic quantification of samples
Quantification was carried out on 10 mL of extract using aHPLC system (Shimadzu LC 20, Japan) equipped with aphotodiode array detector and consisting of a Eclipse XDBC18 (3.5 mm) column (100 mm × 4.6 mm, Agilent). Theelution system (0.6 mL min21) involved two filtered (0.2 mmpore size filter), sonicated and degassed solvents, namely sol-vents A (water/acetic acid, 98:2, v/v) and B (H2O/MeOH/acetic acid, 5:90:5 v/v/v). The linear gradient was: 0 min,18 % solvent B; 0–5 min, 25 %; 5–8 min, 36 %; 8–10 min,
isocratic; 10–13 min, 58 %; 13–16 min, 62 %; 16–21, 18 %;isocratic, 18 % till 25 min.
The calibration curve was plotted using three replicatepoints of standard solutions of mangiferin and 5-CQA fromSigma-Aldrich (St Louis, USA), and 3,5-dicaffeoylquinicacid (3,5-diCQA), a gift from Professor Andary {extractedfrom sunflower [Helianthus annuus L. (Asteraceae)],Laboratory of Natural Substances, Faculty of Pharmacy,Montpellier} at 10, 25, 50, 75 and 100 mg mL – 1.Identification was performed by comparing spectra and reten-tion times at 280, 320 and 360 nm. Quantification of mangi-ferin, 3-, 4- and 5-CQA, FQAs and 3,4-, 3,5- and4,5-diCQA was undertaken at 320 nm by comparison withmangiferin, 5-CQA and 3,5-diCQA standards. The techniquefor FQA and mangiferin quantification is new, and is reportedhere for the first time. Except for seeds in which content wasexpressed as a percentage of fresh weight (g 100 g21 f. wt),compound content was expressed as percentage of dryweight (g 100 g21 d. wt).
Histochemical analysis
Small pieces of freshly collected Coffea leaves or fruits wereembedded in 3 % agarose (type II EEO, Panreac) beforecutting for histochemical examination. Cross-sections(40 mm) were obtained using a Leica VT 1000S vibratingblade microtome (frequency 7, speed 2). For mangiferin histo-localization, cross-sections were mounted in distilled waterwithout any reagent. Transverse sections of specimens wereviewed under a light microscope (Nikon Optiphot) with UVlight (filter UV-1A: 365 nm excitation filter). In these condi-tions, mangiferin presents a strong yellow autofluorescence.Photographs were taken with a digital Nikon Coolpix 4500camera.
Statistical analyses
All results were analysed using the Statistica softwarepackage (7.1 version, USA). The primary variables are:3-CQA, 3-caffeoylquinic acid; 4-CQA, 4-caffeoylquinicacid; 5-CQA, 5-caffeoylquinic acid; 3,4-diCQA,3,4-dicaffeoylquinic acid; 3,5-diCQA, 3,5-dicaffeoylquinicacid; 4,5-diCQA, 4,5-dicaffeoylquinic acid; FQAs, sum of3-, 4- and 5-feruloylquinic acid; and mangiferin. Secondaryvariables were: CQAs, sum of the isomers of CQA; andHCEs, sum of the isomers of CQA and FQA. Statistical ana-lysis was used to examine between-species variation, whichwas tested using one-way analysis of variance (ANOVA)with fixed effect. When the F-test was significant, aNewman and Keuls test was carried out to compare means.Standard deviations were computed from the residual meansquare, the latter being the best estimate (unbiased and accur-ate) of the residual variance. Linear regression was performedto highlight relationships between some HCE isomers.Hierarchical clustering analysis (with weighted average group-ing method and Euclidian distance as parameters) and princi-pal component analysis (PCA) were applied to data in order toidentify clusters and similarities between samples and species.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea598
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Clade and geographical terminology
The terminology for area-based clades largely followsMaurin et al. (2007): Upper Guinea (UG) clade, LowerGuinea/Congolian (LG/C) clade, East-Central African(EC-Afr) clade, East African (EA) clade; and Anthony et al.(2010) and Davis et al. (2011): Africa/Indian Ocean (A/IO)clade. The humid West and Central African forests are con-tained within the Guineo-Congolian Regional Centre ofEndemism (White, 1983). Within this major region there arethree sub-centres of endemism for humid forest species: (1)Upper Guinea; (2) Lower Guinea; and (3) Congolian (White,1979). For practical purposes, the sub-centres (2) and (3) areoften merged as the Lower Guinea/Congolian region, andthis convention has been followed here. There are three excep-tions to the area–clade relationship outlined by Maurin et al.(2007). Coffea canephora and C. liberica, (members of theLG/C clade) also occur in the Upper Guinea region; andC. anthonyi Stoff. & F.Anthony (a member of the EC-Afrclade) occurs in the Lower Guinea/Congolian region. TheEast-Central Africa area is comparable with the LakeVictoria Regional Mosaic [including most of Uganda, thewhole of eastern Rwanda and Burundi, and small parts ofZaire, Kenya and Tanzania (White, 1983)], a predominantlyhigh altitude region formed by the uplift of the Great RiftValley. Coincidently, the vegetation of this area is ‘ameeting place of five distinct floras’ (White, 1983), includingthe Guineo-Congolian flora, and this may help to explain therelationship (EC-Afr clade; Maurin et al., 2007; Davis et al.,2011) of C. anthonyi with the high altitude East-CentralAfrica species, C. eugenioides and C. kivuensis Lebrun.
RESULTS
Identification of mangiferin and HCEs
The HPLC quantitative method described herein allowedthe separation of chlorogenic acids (CQAs, diCQAs andFQAs) and mangiferin in a methanol extract ofC. pseudozanguebariae leaves (Fig. 1). Based on their reten-tion time and UV absorbance spectra, the four different peakfamilies were determined. Except for FQAs, all the
compounds detected can be quantified, although mangiferinand an FQA isomer (peak 4 and 5) presented quite similar re-tention times (9.95 and 10.17 min, respectively). It was diffi-cult to state the nature of FQA isomers in the absence ofstandards. We have therefore chosen to express FQA contentas FQAs, cumulating 4- and 5-FQA content when both werepresented (3-FQA was generally undetectable), without speci-fying the nature and the proportion of each isomer.Isomangiferin (peak 4′) was detected at 12.58 min. Becauseit was often present in trace amounts, this compound wasnot taken into account.
An analysis by ESI-MS was conducted to identify with cer-tainty the compounds accumulated in the leaves but also infruits (Table 2). An elution profile identical to that obtainedby the HPLC quantitative method allowed the observation ofnine major peaks by separation on an XTerra MS C18column. Peaks 1, 2 and 3 shared typical chlorogenic acid(5-CQA) UV absorbance spectra with a shoulder at 240 nmand maxima at 324 nm. These peaks have the same m/z 353[M-H]2 and 355 [M + H]+ parent ions, and shared m/z 191,179, 173 [M-H]2 fragment ions corresponding to [quinicacid-H]2, [caffeic acid-H]2 and [quinic acid-H-H2O]2, re-spectively. Peak 2 was assigned to 5-CQA as it co-elutedwith chlorogenic acid standard and exhibited the same frag-mentation pattern (Clifford et al., 2003). The very low m/z173 relative intensity of peak 1 compared with peak 3 indi-cated that peak 1 can be assigned to 3-CQA and peak 3 to4-CQA. Peak 4 showed the absorbance spectra of the mangi-ferin standard, with peaks at 241, 258, 318 and 366 nm anda shoulder at 275 nm within the acidified water/acetonitrileeluent. It exhibited the pseudomolecular parent ion m/z 423[M + H]+ and 421 [M-H]– at a retention time of16.26 min. In our system, the mangiferin standard gave thesame fragmentation pattern as that obtained using the API4000 LC-MS/MS system (Suryawanshi et al., 2007). In nega-tive mode, the m/z 421 [M-H]2 parent ion yielded to fragmentions at m/z 403, 331, 301, 273 and 259. The fragment ion at m/z 301 corresponded to glycoside lost [mangiferin-H-120]2 andat m/z 273 to a supplementary [-CH2O] lost.
Peaks 5 and 6 (as the latter was not detected in leaf samples,it is not shown in Fig. 1) exhibited typical UV spectra of
200
Abs
orba
nce
(mA
U)
100
0
0
1
3
2
4
54'
7 89
5 10
Retention time (min)
1500
1000
Abs
orba
nce
(mA
U)
Abs
orba
nce
(mA
U)
500
0200 300250
318366 324
296
350Wavelength (nm)
400 200 2500
50100150200250300
300 350Wavelength (nm)
400
15 20 25
A B
FI G. 1. HPLC profile of a C. pseudozanguebariae leaf extract. Absorption profile obtained at 320 nm using the technical conditions for quantitative analysis.Peak 1 ¼ 3-caffeoylquinic acid (3-CQA); peak 2 ¼ 5-caffeoylquinic acid (5-CQA); peak 3 ¼ 4-caffeoylquinic acid (4-CQA); peak 4 ¼ mangiferin; peak 4’ ¼isomangiferin; peak 5 ¼ 5-feruloylquinic acid (5-FQA); peak 6 ¼ 4-feruloylquinic acid (4-FQA); peak 7 ¼ 3,4-dicaffeoylquinic acid (3,4 diCQA); peak 8 ¼3,5-dicaffeoylquinic acid (3,5-diCQA); and peak 9 ¼ 4,5-dicaffeoylquinic acid (4,5-diCQA). The absorption spectrum between 200 and 400 nm is indicated
for mangiferin (A) and FQA (B).
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 599
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

hydroxycinnamic acids, with a shoulder at 296 nm and amaximum at 324 nm, characteristic of quinic acid derivatives.Both peaks showed the m/z 367 [M-H]2 and 369 [M + H]+
parent ions, and shared the m/z 191 [M-H]2 fragment ion, cor-responding to [quinic acid-H]2. The more abundant fragmentsof 3-FQA, 5-FQA and 4-FQA are 193, 191 and 173, respect-ively. From comparison with a C. canephora grain extract,we deduced that peaks 5 and 6 can be assigned to 5-FQAand 4-FQA, respectively (Clifford et al., 2006; Matsui et al.,2007; Alonso-Salces et al., 2009; Jaiswal et al., 2010b).Peaks 7, 8 and 9 shared typical 5-CQA UV absorbancespectra. Having the same m/z 515 [M-H]2 and 517 [M +H]+ parent ions, and a m/z 353 [M-H]2 fragment ion assignedto CQAs, they corresponded to diCQAs. Peak 7 and 9 formedfour major fragments at m/z 191 [quinic acid-H]-, 173 [quinicacid-H-H2O]-, 161 [caffeic acid-H-H2O]- and 135 [caffeicacid-H-CO2]-. Peak 7 and peak 9, exhibiting the highest 173[M-H]2 and 179 [M-H]2 fragment, respectively, can beassigned to 3,4-diCQA and 4,5-diCQA. Peak 8 appeared tobe 3,5-diCQA.
Jaiswal et al. (2010a) and Kuhnert et al. (2011) reported thepresence of numerous minor chlorogenic acids inC. canephora beans, based on sinapic or methoxycinnamicacids. We also observed some of these secondary metabolitesin C. canephora and C. pseudozanguebariae (their retentiontimes are close to those of FQAs), although their concentra-tions were very low and content comparison between specieswas difficult.
Mangiferin localization in leaf and fruit tissues
Histochemical observations for the localization of mangi-ferin in leaves and fruits were performed on three species:C. pseudozanguebariae, C. arabica and C. canephora.
Leaves. Based on mangiferin autofluorescence properties,histochemical observations revealed a preferential localizationof mangiferin in palisade and spongy (mesophyll) parenchymaof C. pseudozanguebariae leaves (Fig. 2A1). In comparison,the same transverse sections of leaves indicated that mangi-ferin is absent in C. canephora (Fig. 2A2), but present at alow concentration in C. arabica (Fig. 2A3).
Fruits. The intense yellow autofluorescence observed in thecells of the exocarp (outer fruit wall or skin) and the externallayers of the mesocarp (soft, pulpy layer of the fruits) of younggreen fruits of C. pseudozanguebariae indicate a high contentof mangiferin (Fig. 2B1). Mangiferin was not detected in thegreen fruit of C. canephora (Fig. 2B2). In C. arabica, mangi-ferin was weakly accumulated and preferentially localized incells from the exocarp and external mesocarp (Fig. 2B3), asin C. pseudozanguebariae. Mangiferin was entirely absentfrom the seeds and endocarp (pyrene layer, or ‘parchment’)of the three species examined (Fig. 2B4).
Quantitative evaluation of mangiferin during leaf and fruit ageing
Mangiferin content was evaluated in leaves and fruits at dif-ferent development stages. The study was undertaken on thesame three species used in the histochemical observations(see above) plus C. eugenioides, one of the parent species ofC. arabica, the unique amphidiploid species originating fromhybridization between C. eugenioides and C. canephora(Lashermes et al., 1999; Maurin et al., 2007).
Leaves. In C. pseudozanguebariae, C. eugenioides andC. arabica, mangiferin was present at each developmentstage, but was not detected in C. canephora (Table 3). Whenpresent, mangiferin content decreased with leaf ageing[about 30 % less mangiferin content between young leaf(from the apex) and mature leaf (from node 2)]. Irrespectiveof the developmental stage, the highest mangiferin contentwas observed in leaves of C. eugenioides.
Fruits. Mangiferin content was evaluated for the same speciesas above, except C. eugenioides, in fruits at different growthstages. As previously indicated by histochemical observations,quantitative analysis confirmed that mangiferin was actuallypresent at all stages of development in the fruit ofC. pseudozanguebariae and C. arabica (Table 4). In entirefruits from stage 1, mangiferin content was 5-fold lower inC. arabica than in C. pseudozanguebariae, where theaverage content of mangiferin is approx. 1.15 % of the freshfruit weight. Mangiferin was not detected in the fruit ofC. canephora. Even at the later stages of fruit maturity, a sep-arate analysis revealed that mangiferin continued only to accu-mulate in the endocarp and outer layers of the mesocarp, and
TABLE 2. Electrospray ionization mass–spectrometry characterization of isomers
Peak RT (min) [M-H]2 (m/z) ESI-MS fragments (m/z) UV lmax (nm) Abbreviation Identification
1 6.05 353 191 (100), 179 (82), 173 (1), 135 (10) 240, 300sh, 324 3-CQA 3-Caffeoylquinic acid2* 10.03 353 191 (100), 179 (51) 240, 300sh, 324 5-CQA 5-Caffeoylquinic acid3 11.20 353 191 (20), 179 (98), 173 (100), 135 (12) 240, 300sh, 324 4-CQA 4-Caffeoylquinic acid4* 16.26 421 403, 331 (93), 301, 271 258, 318, 366 Mangif Mangiferin5 17.90 367 193, 191, 149 296sh, 324 5-FQA 5-Feruloylquinic acid6 29.29 367 193, 173 296sh, 324 4-FQA 4-Feruloylquinic acid7 24.33 515 353, 191, 173, 161, 135 240, 300sh, 326 3,4-DiCQA 3,4-Dicaffeoylquinic acid8 28.91 515 353, 191, 179, 161, 135 240, 300sh, 326 3,5-diCQA 3,5-Dicaffeoylquinic acid9 31.63 515 353, 179 240, 300sh, 326 4,5-DiCQA 4,5-Dicaffeoylquinic acid
Peak assignments of the fresh pericarp extracts and green bean extracts using LC-ESI-MS. Characterization of phenolic compounds and mangiferin by UVabsorbance spectrum and electrospray ionization-mass spectrometry detection (LC-DAD/ESI-MS). RT, retention time.
* Identified with standard compound.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea600
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

not the pyrene coat (endocarp) or the seed (mostly endo-sperm). As with leaves, mangiferin content decreased withageing (maturity); stage 2 and 3 had .3-fold less mangiferincompared with stage 1.
Mangiferin and HCE content in leaves of wild coffee species
Mangiferin and HCE contents were analysed by liquid chro-matography in mature leaves (taken from the second nodebelow the apex of the shoot) of 49 genotypes belonging to23 species originating from different regions of Africa andMadagascar (Table 1). The nine selected compounds werereadily identified in the chromatograms (Table 5).
Mangiferin content. Seven of the 15 African species ac-cumulated mangiferin in their leaves (Table 5), i.e.C. anthonyi, C. arabica, C. eugenioides, C. heterocalyx cf.,C. pseudozanguebariae, C. sessiliflora and C. salvatrixSwynn. & Philipson. Mangiferin content among these sevenspecies was present in a range of 0.03–16.36 % d. wt.Coffea salvatrix and C. anthonyi accumulated the greatestquantities (16.36 and 12.06 % d. wt, respectively). None ofthe nine Madagascan species examined contained mangiferin.
FI G. 2. Histochemical localization of mangiferin in leaves (A) and green fruit (B) of different Coffea species. Cross-sections were observed under UV light,×400, except B4 (×40). (A) Leaves. Coffea pseudozanguebariae leaf blade (A1), showing a very high concentration of mangiferin preferentially localized in thecells forming the palisade and spongy parenchyma (mesophyll) (yellow arrows). Coffea canephora leaf blade (A2), no mangiferin accumulated (i.e. no specificyellow autofluorescence), allowing observation of chlorophyll (red autofluorescence), principally in palisade and spongy parenchyma. Blue fluorescences arespecific for cuticle constituents and of caffeoylquinic acids (in epidermal cells). Coffea arabica ‘Laurina’ leaf blade (A3), showing that the presence of mangiferin(yellow autofluorescence) has attenuated the red autofluorescence of the chlorophyll. (B) Fruits. Coffea pseudozanguebariae (B1), yellow autofluorescence oftransverse sections (T.S.) of immature fruits (stage 1), showing that mangiferin is extremely concentrated in the exocarp and mesocarp cells. Coffea canephora(B2), no mangiferin accumulated. Coffea arabica ‘Laurina’ (B3 and B4) mangiferin accumulation appears to be vacuolar and restricted to exocarp and externalmesocarp cells (arrows) in young fruits. There is no mangiferin detected in the endocarp, integument (seed coat) and seed. Abbreviations: c, cuticle; chl, chloro-
phyll; e, epidermis; en, endocarp; ex, exocarp; in, integument; m, mesocarp; pp, palisade; s, seed; sp, spongy parenchyma (mesophyll).
TABLE 3. Mangiferin content (% d. wt) in leaves at threedevelopment stages of C. pseudozanguebariae, C. canephora,
C. arabica L. cv. ‘Laurina’ and C. eugenioides
Species Stage 1 Stage 2 Stage 3
C. pseudozanguebariae 8.60+1.15 8.03+1.34 5.62+0.87C. eugenioides 11.39+1.03 7.61+0.73 6.43+0.53C. arabica ‘Laurina’ 1.11+0.26 0.92+0.32 0.89+0.22C. canephora 0 0 0
Stage 1 corresponds to leaves from the apex; and stages 2 and 3 to leavesfrom the first and second node from the apex, respectively. Values areexpressed as a percentage of dry weight (% d. wt) and corresponded to themean between three trees evaluated in triplicate.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 601
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
HCE content. Among HCEs, the three isomers of CQA werethe only compounds accumulated in the leaves of all speciesexamined, FQAs and diCQAs being exclusive to individualspecies (Table 5). The HCE composition of C. canephoraleaves was in agreement with previous results concerning thebiosynthesis pathway of CQAs in C. canephora seedlings(Mahesh et al., 2007). That is, in mature leaves ofC. canephora from Cameroon, the total CQAs representsabout 2 % d. wt and .80 % of the content in HCEs. Theaverage dry weight of the total HCE content varied from0.85 % (C. sessiliflora) to 3.96 % (C. arabica) in the leavesof African species, and from 0.64 % [C. resinosa (Hook.f.)Radlk.] to 2.02 % (C. leroyi A.P.Davis) for the Madagascanspecies tested. CQAs were the major HCEs accumulated inthe leaves of all the species examined. These two compounds(3- and 5-CQA) accounted for .80 % of the HCEs, exceptfor two species: C. stenophylla G.Don, an African speciesfrom the Upper Guinean forests, and C. andrambovatensisJ.-F. Leroy, a species from Eastern Madagascar. Both specieswere characterized by ,62 % of HCEs in the form ofCQAs, and a FQAs content higher (0.74 and 0.36 % d. wt, re-spectively) than that of 5-CQA (0.48 and 0.33 % d. wt). As innumerous other plants, 5-CQA appeared as the most abundantCQA isomer in Coffea species, except in C. augagneuriDubard, a Madagascan species in which 4-CQA is preferen-tially accumulated. FQAs were not detected in .60 % of thegenotypes, including C. pseudozanguebariae. Only fivespecies presented FQAs content .0.1 % d. wt: C. arabica(0.12 %), C. humilis (0.18 %) and C. stenophylla (0.74 %)for African species, and C. leroyi (0.31 %) andC. andrambovatensis (0.36 %) for Madagascan species.DiCQAs accumulated weakly, but were detected in 90 % ofthe species. When evaluated, total content varied from 0.001to 0.370 % d. wt in C. anthonyi and C. canephora leaves,respectively, and diCQAs were not detected in the leavesof C. stenophylla (from Africa) and C. millotii (fromMadagascar). For the 20 species containing diCQAs, totalcontent ranged from 0.61 % (C. tsirananae, a Madagascanspecies) to 13.12 % (C. canephora, an African species) ofHCEs. For all the species, except C. augagneuri, 3,4-diCQAappeared as the minor isomer.
Statistical analyses
For each of the phenolic compounds analysed, interspecificvariability was considerable, as estimated by the interspecific
maximum/minimum ratio, which varied from 4 for 5-CQA(from 3.2 % for C. arabica, to 0.64 % for C. humilis), and620 for mangiferin (from 16.36 % for C. salvatrix to 0.03 %for C. sessiliflora). There was a significant effect of species,as tested by ANOVA. A Newmann and Keuls test applied tothe means of the African species indicated that the 5-CQAcontent of C. arabica was significantly greater than that ofall other species (Table 5). In the same way, the FQAcontent of C. stenophylla and the 3,4-diCQA content ofC. canephora were significantly higher compared with theother species. Differences were highly significant with mangi-ferin, so that values can be considered as species specific, i.e.C. anthonyi and C. salvatrix (Table 5). No particular rela-tionship could be established between the content in the dif-ferent compounds studied, except for 3-CQA and 4-CQAwhich showed a weak linear relationship (Fig. 3), althoughfive African species did not present this relationship:C. arabica, C. heterocalyx cf., C. racemosa, C. salvatrix andC. stenophylla.
When species were grouped according to their positionwithin recent phylogenetic analyses of Coffea (Maurin et al.,2007; Anthony et al., 2010; Davis et al., 2011; see Table 1)and analysed statistically, the following groups were found tobe significantly different for mangiferin accumulation(Table 6): (1) C. anthonyi, C. eugenioides (EC-Afr clade);and (2) C. pseudozanguebariae, C. salvatrix (EA[2] clade).For HCE distribution, the following groups were supported:(1) C. anthonyi, C. eugenioides (EC-Afr clade), on the basisof 5-CQA and total CQAs; and (2) C. humilis andC. stenophylla (UG clade), for FQAs (Table 6). A clusteringanalysis of all African species was performed using all HCEdata (CQA isomers, FQAs and diCQA isomers) as variables.
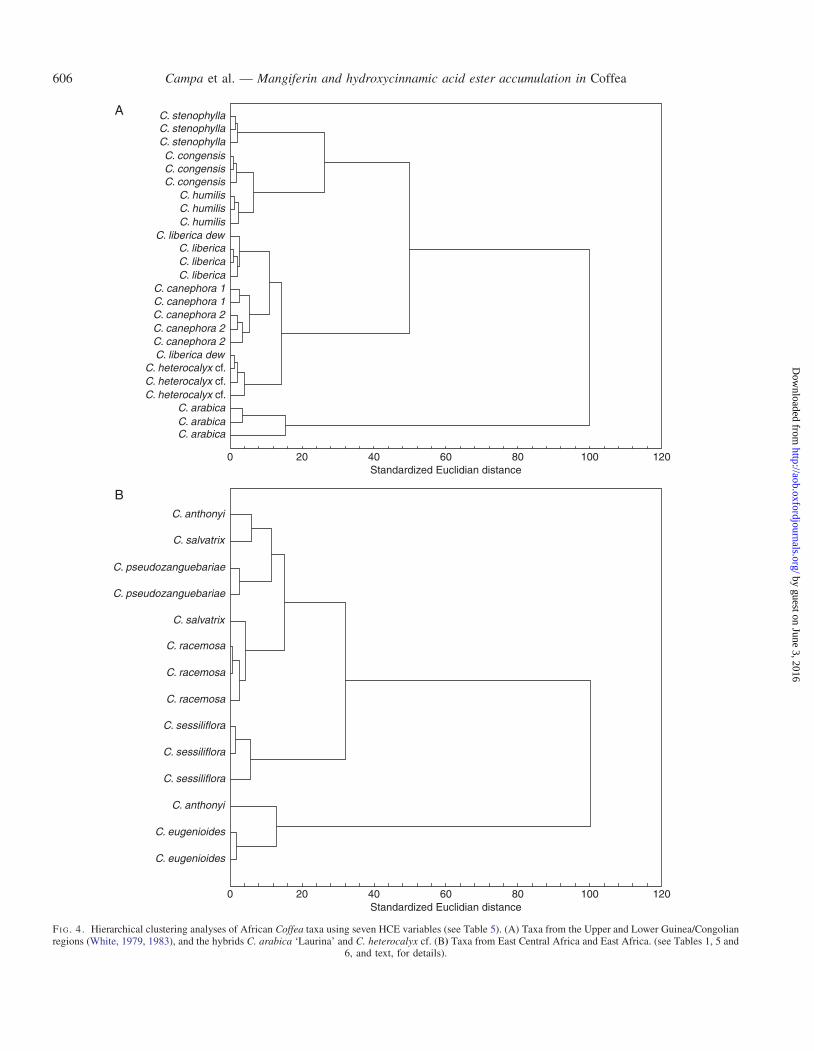
Analyses using hierarchical clustering analysis, e.g. on allsamples (49 genotypes), all African (40 genotypes) (neitheranalyses shown here), and those undertaken below, revealedweak correspondence with phylogenetic topologies based onparsimony and Bayesian analysis of molecular data (Maurinet al., 2007; Anthony et al., 2010; Davis et al., 2011). Atthe species level, however, clustering was remarkably good.In the separate analyses presented here, which are based onthe major lineages of Coffea (Maurin et al., 2007; Anthonyet al., 2010; Davis et al., 2011), the clustering ability ofHCEs at the species and population level is clearly demon-strated. In Fig. 4A, which incorporates taxa from the Upperand Lower Guinea/Congolian regions (White, 1979, 1983)and two hybrids, only one sample of C. liberica var. dewevrei
TABLE 4. Mangiferin content (% f. wt) during fruit development of C. pseudozanguebariae, C. arabica L. cv. ‘Laurina’ andC. canephora
Stage 1 (ST1) Stage 2 (ST5) Stage 3 (ST6)Species Entire fruit Pericarp Seed Pericarp Seed
C. pseudozanguebariae 1.13+0.37 0.43+0.15 0 0.61+0.23 0C. arabica ‘Laurina’ 0.23+0.07 0.07+0.02 0 0.05+0.02 0C. canephora 0 0 0 0 0
Stage 1 corresponds to the first stage of development of the perisperm; stage 2 to the completion of the development of the endosperm; and stage 3 topericarp maturation; or ST1, ST5 and ST6 stages according to De Castro and Marraccini (2006), respectively. Content is expressed as a percentage of freshweight (% f. wt). Evaluations have been made in triplicate on five entire fruits for the first stage, and on fruit fragmented between seeds and pericarp forstages 2 and 3.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea602
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
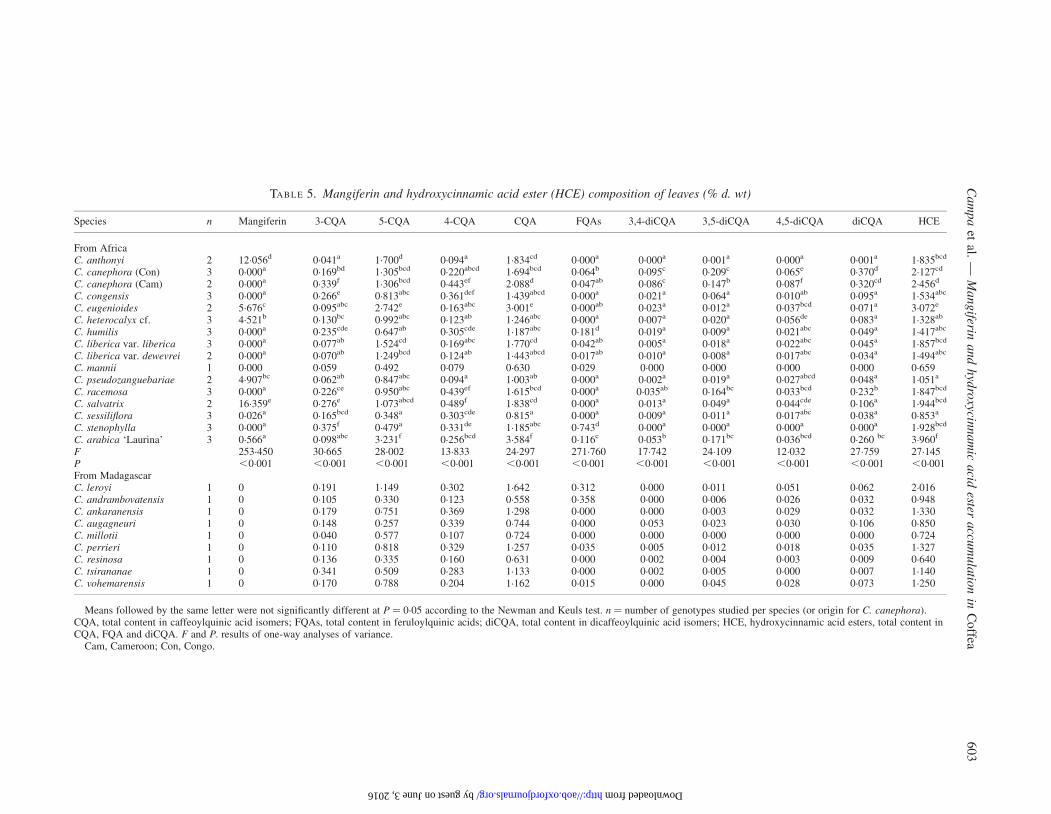
TABLE 5. Mangiferin and hydroxycinnamic acid ester (HCE) composition of leaves (% d. wt)
Species n Mangiferin 3-CQA 5-CQA 4-CQA CQA FQAs 3,4-diCQA 3,5-diCQA 4,5-diCQA diCQA HCE
From AfricaC. anthonyi 2 12.056d 0.041a 1.700d 0.094a 1.834cd 0.000a 0.000a 0.001a 0.000a 0.001a 1.835bcd
C. canephora (Con) 3 0.000a 0.169bd 1.305bcd 0.220abcd 1.694bcd 0.064b 0.095c 0.209c 0.065e 0.370d 2.127cd
C. canephora (Cam) 2 0.000a 0.339f 1.306bcd 0.443ef 2.088d 0.047ab 0.086c 0.147b 0.087f 0.320cd 2.456d
C. congensis 3 0.000a 0.266e 0.813abc 0.361def 1.439abcd 0.000a 0.021a 0.064a 0.010ab 0.095a 1.534abc
C. eugenioides 2 5.676c 0.095abc 2.742e 0.163abc 3.001e 0.000ab 0.023a 0.012a 0.037bcd 0.071a 3.072e
C. heterocalyx cf. 3 4.521b 0.130bc 0.992abc 0.123ab 1.246abc 0.000a 0.007a 0.020a 0.056de 0.083a 1.328ab
C. humilis 3 0.000a 0.235cde 0.647ab 0.305cde 1.187abc 0.181d 0.019a 0.009a 0.021abc 0.049a 1.417abc
C. liberica var. liberica 3 0.000a 0.077ab 1.524cd 0.169abc 1.770cd 0.042ab 0.005a 0.018a 0.022abc 0.045a 1.857bcd
C. liberica var. dewevrei 2 0.000a 0.070ab 1.249bcd 0.124ab 1.443abcd 0.017ab 0.010a 0.008a 0.017abc 0.034a 1.494abc
C. mannii 1 0.000 0.059 0.492 0.079 0.630 0.029 0.000 0.000 0.000 0.000 0.659C. pseudozanguebariae 2 4.907bc 0.062ab 0.847abc 0.094a 1.003ab 0.000a 0.002a 0.019a 0.027abcd 0.048a 1.051a
C. racemosa 3 0.000a 0.226ce 0.950abc 0.439ef 1.615bcd 0.000a 0.035ab 0.164bc 0.033bcd 0.232b 1.847bcd
C. salvatrix 2 16.359e 0.276e 1.073abcd 0.489f 1.838cd 0.000a 0.013a 0.049a 0.044cde 0.106a 1.944bcd
C. sessiliflora 3 0.026a 0.165bcd 0.348a 0.303cde 0.815a 0.000a 0.009a 0.011a 0.017abc 0.038a 0.853a
C. stenophylla 3 0.000a 0.375f 0.479a 0.331de 1.185abc 0.743d 0.000a 0.000a 0.000a 0.000a 1.928bcd
C. arabica ‘Laurina’ 3 0.566a 0.098abc 3.231f 0.256bcd 3.584f 0.116c 0.053b 0.171bc 0.036bcd 0.260 bc 3.960f
F 253.450 30.665 28.002 13.833 24.297 271.760 17.742 24.109 12.032 27.759 27.145P ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001 ,0.001From MadagascarC. leroyi 1 0 0.191 1.149 0.302 1.642 0.312 0.000 0.011 0.051 0.062 2.016C. andrambovatensis 1 0 0.105 0.330 0.123 0.558 0.358 0.000 0.006 0.026 0.032 0.948C. ankaranensis 1 0 0.179 0.751 0.369 1.298 0.000 0.000 0.003 0.029 0.032 1.330C. augagneuri 1 0 0.148 0.257 0.339 0.744 0.000 0.053 0.023 0.030 0.106 0.850C. millotii 1 0 0.040 0.577 0.107 0.724 0.000 0.000 0.000 0.000 0.000 0.724C. perrieri 1 0 0.110 0.818 0.329 1.257 0.035 0.005 0.012 0.018 0.035 1.327C. resinosa 1 0 0.136 0.335 0.160 0.631 0.000 0.002 0.004 0.003 0.009 0.640C. tsirananae 1 0 0.341 0.509 0.283 1.133 0.000 0.002 0.005 0.000 0.007 1.140C. vohemarensis 1 0 0.170 0.788 0.204 1.162 0.015 0.000 0.045 0.028 0.073 1.250
Means followed by the same letter were not significantly different at P ¼ 0.05 according to the Newman and Keuls test. n ¼ number of genotypes studied per species (or origin for C. canephora).CQA, total content in caffeoylquinic acid isomers; FQAs, total content in feruloylquinic acids; diCQA, total content in dicaffeoylquinic acid isomers; HCE, hydroxycinnamic acid esters, total content inCQA, FQA and diCQA. F and P. results of one-way analyses of variance.
Cam, Cameroon; Con, Congo.
Ca
mp
aet
al.—
Ma
ng
iferina
nd
hyd
roxycin
na
mic
acid
esteraccu
mu
latio
nin
Co
ffea6
03
by guest on June 3, 2016 http://aob.oxfordjournals.org/ Downloaded from

fails to cluster; the samples of C. canephora cluster accordingto their geographical origin (Congo and Cameroon). InFig. 4B, using taxa from East Central Africa and EastAfrica, only two species do not cluster (C. anthonyi andC. salvatrix). In other analyses (not shown), e.g. all Africansamples, all samples of C. liberica (var. liberica and var.dewevrei) cluster according to their varietal designations.
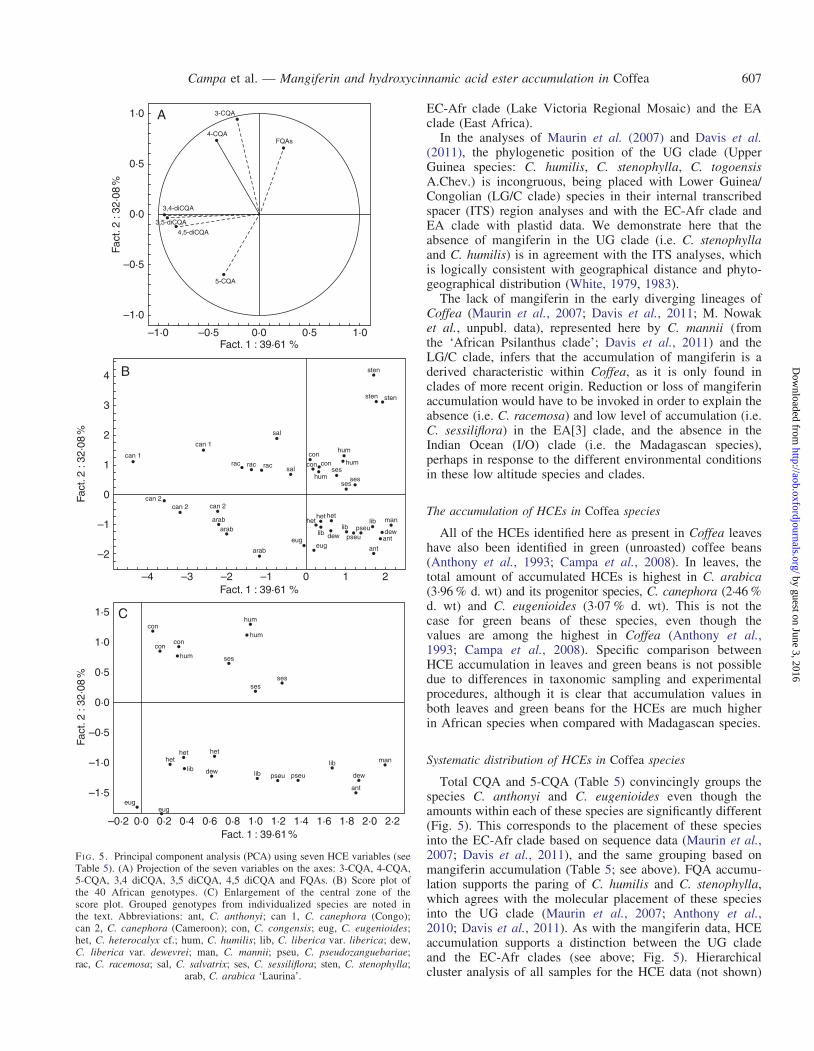
A PCA undertaken on all African species using the sevenHCE primary variables (Fig. 5) showed that two principalcomponents have an eigenvalue of .1, and accounted for39 and 32 % of the total variance, respectively. The scoreplot obtained for principal components 1 and 2 individualizedeight species: C. anthonyi, C. arabica, C. canephora,C. eugenioides, C. racemosa, C. salvatrix, C. sessiliflora andC. stenophylla. Coffea canephora and C. racemosa genotypeswere characterized by a high 3-CQA content, C. stenophyllaby a high FQAs content, and C. eugenioides and C. anthonyiby high 5-CQA contents. The remaining species in the PCAanalysis overlap with respect to their CQA, FQA and HCEprofiles to a greater (C. heterocalyx cf. and C. liberica) orlesser (C. congensis and C. humilis) extent. In a PCA analysisincluding the Madagascan species (not shown) it was clear thatthey exhibited the lowest diversity for HCEs, and this wasmanifest by their distribution in the centre of the PCA plot.
DISCUSSION
The accumulation of mangiferin in Coffea species
Mangiferin, a C-glucosylxanthone, has been recently describedin leaves of the wild Coffea species C. pseudozanguebariae(Talamond et al., 2008). The present work confirms thisresult by a double analysis involving a new HPLC technique
and LC-MS. In contrast to the study of Bertrand et al.(2003) dealing with FQA content of leaves and fruits inC. pseudozanguebariae, only traces of FQAs were recordedin leaves in which mangiferin was highly concentrated. Ouranalysis of fruit content in this species showed that mangiferinwas present in the exocarp and mesocarp, and yet was un-detectable in the endocarp (parchment) or the coffee bean(i.e. seed). The same result was obtained with one of theother leaf mangiferin-accumulating species, C. arabica(Table 4). In C. canephora, the absence of mangiferin inleaves corresponds to the absence of this compound in thefruit. Our data clearly demonstrate that mangiferin is differen-tially accumulated in separate organs of the coffee plant. Inspecies that accumulate mangiferin, this compound is foundin the leaves and outer layers of the fruit (exocarp and outermesocarp) but not in the seed (or endocarp; see above),which is largely composed of endosperm. The differential ac-cumulation within the fruit could be linked to origin, the endo-sperm being formed by the fertilization of maternal tissue,whereas the outer layers of the fruit are composed of vegeta-tive (sporophytic) tissue of the inferior fruit (i.e. the recep-tacle). Another possibility is that mangiferin production isonly associated with photosynthetic tissue, the receptacle andyoung fruit being green and photosynthetic; the internal partsof fruit (e.g. the ovary) not. Presumably, in mangiferin-accumulating species, the embryonic cotyledons contain nomangiferin and accumulation starts at a later leaf stage.
As most of the earlier biochemical analyses of Coffeahave been undertaken on seeds, the lack of accumulation ofmangiferin in this organ may explain why the compound hasonly been reported in a single species of Coffea, i.e.C. pseudozanguebariae, when the leaves were being studied(Talamond et al., 2008). In the analysis of leaf mangiferin
C. sessiliflora
C. racemosa
C. salvatrix
C. stenophylla
C. arabica
C. heterocalyx
C. mannii
4-CQA = 0·09246 + 0·95351(3-CQA)
r = 0·75155
0·000·0
0·1
0·2
0·3
0·4
0·5
0·6
0·05 0·10 0·15 0·20
3-CQA content (% d. wt)
4-C
QA
con
tent
(%
d. w
t)
0·25 0·30 0·35 0·40 0·45
FI G. 3. Linear regression between 4-CQA and 3-CQA contents in leaves. Analysis included 40 genotypes from the 15 African taxa.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea604
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

content carried out here on 23 Coffea species (24 Coffea taxa;Table 5), seven African taxa were found to accumu-late mangiferin: C. anthonyi, C. arabica, C. eugenioides,C. heterocalyx cf., C. pseudozanguebariae, C. salvatrix andC. sessiliflora. Coffea salvatrix and C. anthonyi accumulatedthe greatest quantities (16.36 and 12.06 % d. wt, respectively),which represents higher levels of mangiferin than have beendetected in the leaves of mangoes, i.e. approx. 10 % d. wt(Baretto et al., 2008).
Systematic distribution of mangiferin in Coffea species
Comparison of leaf mangiferin content among 23 Coffeaspecies (24 taxa), represented by 49 genotypes, shows thatthe level of mangiferin accumulation in leaves at the samegrowth stage is species specific in three species, C. anthonyi,C. eugenioides and C. salvatrix. These data agree with themorphological delimitation (Bridson, 1988; Stoffelen et al.,2009) and molecular characterization of these species(Maurin et al., 2007; Anthony et al., 2010; Davis et al.,2011), in that they represent distinct species. The presence/absence of mangiferin in leaves is also in good agreementwith molecular phylogenetic studies of Coffea, as detailedbelow. The clade designation of each of the 24 taxa tested isshown in Table 1 (see also ‘Clade and geographical termin-ology’, above); a statistical analysis of the accumulation ofmangiferin for each clade is shown in Table 6. Among the fivenon-hybrid African species, C. anthonyi and C. eugenioidesbelong to the EC-Afr clade, and C. pseudozanguebariae,C. salvatrix and C. sessiliflora belong to the EA clade. Coffeapseudozanguebariae, C. salvatrix and C. sessiliflora are furtherdivided into the two distinct EA clades reported by Maurinet al. (2007): EA[2] (C. pseudozanguebariae and C. salvatrix)and EA[3] (C. sessiliflora), EA[2] being a predominantly mid-to high altitude East African clade, and EA[3] a predominantlylow altitude East African clade (Davis et al., 2006; Maurinet al., 2007). As all the above clades have a strong geographicalelement, based on species/sample distribution, the mangiferindata are also aligned with the provenance of these Africancoffee species, at least in the species that we have sampled.The phylogenetic position of C. anthonyi, a species from theLower Guinea/Congolian Region (Stoffelen et al., 2009) butlocated in the EC-Afr clade (Davis et al., 2011), violates anarea–clade relationship, but, significantly, its phylogenetic pos-ition is supported by the mangiferin accumulation. Additionally,it is reported that East-Central Africa (comparable with theLake Victoria Regional Mosaic) is a meeting place of five dis-tinct floras (White, 1983), including the Guineo-Congolianflora (White, 1983). It should also be said that the exact prov-enance details for C. anthonyi are not yet fully known, andfurther work on the precise geographical distribution of thisspecies is required.
Our sampling of major clades across the Coffea genus is notcomplete: material from Asian and Australasian clade, and theMascarene clade (Davis et al., 2011) is lacking and, in add-ition, sampling within some clades is limited, except for theEC-Afr clade and the UG clade. Despite these sampling lim-itations, the accumulation of mangiferin shows a consistentpattern of phylogenetic correlation, being restricted to the
TA
BL
E6.
Mangif
erin
and
hyd
roxy
cinnam
icaci
des
ter
(HC
E)
com
posi
tion
of
leave
s(%
d.
wt)
Cla
de
nM
angif
erin
3-C
QA
5-C
QA
4-C
QA
CQ
AF
QA
3,4
-diC
QA
3,5
-diC
QA
4,5
-diC
QA
diC
QA
HC
E
UG
60
a0. 3
05
b0
. 563
a0. 3
18
ab
1. 1
86
ab
0. 4
62
b0. 0
09
a0. 0
05
a0
. 010
a0. 0
24
a1
. 672
ab
c
LG
/C8
0a
0. 1
46
ab
1. 1
89
a0. 2
30
ab
1. 5
64
a0
. 020
a0. 0
12
a0. 0
32
a0
. 016
a0. 0
61
a1
. 645
ab
c
UG
_L
G/C
50
a0. 2
37
ab
1. 3
05
a0. 3
09
ab
1. 8
52
ac
0. 0
57
a0. 0
92
c0. 1
84
b0
. 074
c0. 3
50
c2
. 259
bc
EC
-Afr
48. 8
66
b0. 0
68
a2
. 221
b0. 1
29
ab
2. 4
18
c0
a0. 0
11
a0. 0
06
a0
. 018
a0. 0
36
a2
. 453
c
EA
[2]
410. 6
33
b0. 1
69
ab
0. 9
60
a0. 2
91
ab
1. 4
21
ab
0a
0. 0
07
a0. 0
34
a0
. 021
a0. 0
77
a1
. 497
ab
EA
[3]
60. 0
13
a0. 1
95
ab
0. 6
49
a0. 3
71
b1. 2
15
ab
0a
0. 0
22
a0. 0
87
a0
. 036
ab
0. 1
35
a1
. 350
ab
IO9
0a
0. 1
58
ab
0. 6
13
a0. 2
46
ab
1. 0
16
ab
0. 0
80
a0. 0
07
a0. 0
12
a0
. 021
a0. 0
39
a1
. 136
a
Non-d
esig
nat
edsp
ecie
sC
.m
annii
10
a0. 0
59
a0
. 492
a0. 0
79
a0. 6
30
b0
. 029
a0
a0
a0
a0
a0
. 659
a
Hybri
ds
C.
ara
bic
acv
.‘L
auri
na’
30. 5
66
a0. 0
98
a3
. 231
c0. 2
56
ab
3. 5
84
d0
. 116
a0. 0
53
b0. 1
71
b0
. 036
ab
0. 2
60
b3
. 960
d
C.
het
eroca
lyx
cf.
34. 5
21
a0. 1
30
ab
0. 0
992
a0. 1
23
ab
1. 2
46
ab
0. 0
00
a0. 0
07
a0. 0
20
a0
. 056
bc
0. 0
83
a1
. 328
ab
F15. 9
32
3. 5
52
22. 9
89
2. 4
81
15. 4
49
6. 7
46
17. 4
51
12. 8
98
8. 9
76
18. 2
99
14. 2
30
P,
0. 0
01
0. 0
03
,0
. 001
0. 0
24
,0. 0
01
,0
. 001
,0. 0
01
,0. 0
01
,0
. 001
,0. 0
01
,0
. 001
Mea
ns
foll
ow
edby
the
sam
ele
tter
wer
enot
signifi
cantl
ydif
fere
nt
atP¼
0. 0
5ac
cord
ing
toth
eN
ewm
anan
dK
euls
test
.n¼
num
ber
of
gen
oty
pes
studie
dfo
rea
chbio
geo
gra
phic
ori
gin
acco
rdin
gto
Mau
rin
etal.
(2007)
and
Dav
iset
al.
(2011).
Fan
dP
,re
sult
sof
one-
way
anal
yse
sof
var
iance
.C
lade
term
inolo
gy
abbre
via
tions:
EA
,E
ast
Afr
ican
clad
e(n
um
ber
insq
uar
ebra
cket
sden
ote
the
thre
em
ajor
clad
esw
ithin
the
EA
clad
e);
EC
-Afr
,E
ast
Cen
tral
Afr
ican
clad
e;IO
,In
dia
nO
cean
clad
e;L
G/C
,L
ow
erG
uin
eae/
Congoli
ancl
ade;
UG
,U
pper
Guin
eacl
ade.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 605
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
C. stenophyllaC. stenophyllaC. stenophyllaC. congensisC. congensisC. congensis
C. humilisC. humilisC. humilis
C. liberica dewC. libericaC. libericaC. liberica
C. canephora 1C. canephora 1C. canephora 2C. canephora 2C. canephora 2C. liberica dew
C. heterocalyx cf.C. heterocalyx cf.C. heterocalyx cf.
C. arabicaC. arabicaC. arabica
C. anthonyi
C. salvatrix
C. pseudozanguebariae
C. pseudozanguebariae
C. salvatrix
C. racemosa
C. racemosa
C. racemosa
C. sessiliflora
C. sessiliflora
C. sessiliflora
C. anthonyi
C. eugenioides
C. eugenioides
0 20 40 60Standardized Euclidian distance
Standardized Euclidian distance
80 100 120
0
A
B
20 40 60 80 100 120
FI G. 4. Hierarchical clustering analyses of African Coffea taxa using seven HCE variables (see Table 5). (A) Taxa from the Upper and Lower Guinea/Congolianregions (White, 1979, 1983), and the hybrids C. arabica ‘Laurina’ and C. heterocalyx cf. (B) Taxa from East Central Africa and East Africa. (see Tables 1, 5 and
6, and text, for details).
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea606
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from
EC-Afr clade (Lake Victoria Regional Mosaic) and the EAclade (East Africa).
In the analyses of Maurin et al. (2007) and Davis et al.(2011), the phylogenetic position of the UG clade (UpperGuinea species: C. humilis, C. stenophylla, C. togoensisA.Chev.) is incongruous, being placed with Lower Guinea/Congolian (LG/C clade) species in their internal transcribedspacer (ITS) region analyses and with the EC-Afr clade andEA clade with plastid data. We demonstrate here that theabsence of mangiferin in the UG clade (i.e. C. stenophyllaand C. humilis) is in agreement with the ITS analyses, whichis logically consistent with geographical distance and phyto-geographical distribution (White, 1979, 1983).
The lack of mangiferin in the early diverging lineages ofCoffea (Maurin et al., 2007; Davis et al., 2011; M. Nowaket al., unpubl. data), represented here by C. mannii (fromthe ‘African Psilanthus clade’; Davis et al., 2011) and theLG/C clade, infers that the accumulation of mangiferin is aderived characteristic within Coffea, as it is only found inclades of more recent origin. Reduction or loss of mangiferinaccumulation would have to be invoked in order to explain theabsence (i.e. C. racemosa) and low level of accumulation (i.e.C. sessiliflora) in the EA[3] clade, and the absence in theIndian Ocean (I/O) clade (i.e. the Madagascan species),perhaps in response to the different environmental conditionsin these low altitude species and clades.
The accumulation of HCEs in Coffea species
All of the HCEs identified here as present in Coffea leaveshave also been identified in green (unroasted) coffee beans(Anthony et al., 1993; Campa et al., 2008). In leaves, thetotal amount of accumulated HCEs is highest in C. arabica(3.96 % d. wt) and its progenitor species, C. canephora (2.46 %d. wt) and C. eugenioides (3.07 % d. wt). This is not thecase for green beans of these species, even though thevalues are among the highest in Coffea (Anthony et al.,1993; Campa et al., 2008). Specific comparison betweenHCE accumulation in leaves and green beans is not possibledue to differences in taxonomic sampling and experimentalprocedures, although it is clear that accumulation values inboth leaves and green beans for the HCEs are much higherin African species when compared with Madagascan species.
Systematic distribution of HCEs in Coffea species
Total CQA and 5-CQA (Table 5) convincingly groups thespecies C. anthonyi and C. eugenioides even though theamounts within each of these species are significantly different(Fig. 5). This corresponds to the placement of these speciesinto the EC-Afr clade based on sequence data (Maurin et al.,2007; Davis et al., 2011), and the same grouping based onmangiferin accumulation (Table 5; see above). FQA accumu-lation supports the paring of C. humilis and C. stenophylla,which agrees with the molecular placement of these speciesinto the UG clade (Maurin et al., 2007; Anthony et al.,2010; Davis et al., 2011). As with the mangiferin data, HCEaccumulation supports a distinction between the UG cladeand the EC-Afr clades (see above; Fig. 5). Hierarchicalcluster analysis of all samples for the HCE data (not shown)
3-CQA
4-CQAFQAs
3,4-diCQA
3,5-diCQA
4,5-diCQA
5-CQA
–0·5
–0·5
–1·0
4
3
2
1
0
–1
–2
–4 –3 –2 –1 0
eugeug
arab
arabarab
can 2can 2
can 1
can 1
rac rac
sal
sal
con
hum
con
hum
humses
sesses
sten
sten sten
manlib
pseupseulib
hethet
lib
het
dewdewant
ant
conrac
can 2
1 2
–1·0
0·0
Fact
. 2 :
32·0
8 %
0·0
0·5
0·5
1·0
1·0
con
con
hum
conhum
ses
hum
sesses
hethet het
dew lib pseu pseu
lib
dew
man
ant
eug
–0·2
–1·5
–1·0
–0·5
0·0
Fact
. 2 :
32·0
8 %
Fact
. 2 :
32·0
8 %
0·5
1·0
1·5
0·0 0·2 0·4 0·6 0·8 1·0Fact. 1 : 39·61 %
Fact. 1 : 39·61 %
Fact. 1 : 39·61 %
1·2 1·4 1·6 1·8 2·0 2·2eug
lib
A
B
C
FI G. 5. Principal component analysis (PCA) using seven HCE variables (seeTable 5). (A) Projection of the seven variables on the axes: 3-CQA, 4-CQA,5-CQA, 3,4 diCQA, 3,5 diCQA, 4,5 diCQA and FQAs. (B) Score plot ofthe 40 African genotypes. (C) Enlargement of the central zone of thescore plot. Grouped genotypes from individualized species are noted inthe text. Abbreviations: ant, C. anthonyi; can 1, C. canephora (Congo);can 2, C. canephora (Cameroon); con, C. congensis; eug, C. eugenioides;het, C. heterocalyx cf.; hum, C. humilis; lib, C. liberica var. liberica; dew,C. liberica var. dewevrei; man, C. mannii; pseu, C. pseudozanguebariae;rac, C. racemosa; sal, C. salvatrix; ses, C. sessiliflora; sten, C. stenophylla;
arab, C. arabica ‘Laurina’.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 607
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

shows the Upper Guinea species clustering with the LowerGuinea species.
Low values for most HCEs in the representatives ofMadagascan species, observed in PCA analyses (not shown)as points clustered around the crossing point of the axes,support separation of these taxa from African Coffea. The sep-aration of Madagascan from African Coffea species on thebasis of low levels of HCE accumulation in green beans ofCoffea has been reported by Anthony et al. (1993) andCampa et al. (2005). Phenolic data for both leaves and greenbeans thus support the assumption based on molecularresults (Anthony et al., 2010; Hamon et al., 2011) that theMadagascan species represent a monophyletic lineage withinCoffea. In addition, the presence of other families of phenolicshas been identified in Madagascan species (Rakotomalala,1992). Further analysis is underway to determine the natureof these compounds and include them in future analysis of bio-chemical diversity (A. Rakotondravao, unpubl. data). The gen-erally low interspecific phenolic diversity of the Madagascanspecies, compared with the phenolic diversity across thegenus, is comparable with low DNA sequence diversityobtained in molecular phylogenetic studies (Maurin et al.,2007; Anthony et al., 2010). It has been suggested that theMadagascan species, excluding the baracoffea alliance(Davis and Rakotnasolo, 2008; M. Nowak, unpubl. data), rep-resent a recent radiation within Coffea (Maurin et al., 2007;Anthony et al., 2010).
Like mangiferin, HCEs have the ability to cluster speciesbased on multiple samples (Table 5; Figs 4 and 5). Forexample, in the PCA analysis (Fig. 5), eight species are indi-vidualized based on seven HCE variables: C. anthonyi,C. arabica, C. canephora, C. eugenioides, C. racemosa,C. salvatrix, C. sessiliflora and C. stenophylla. These resultsare comparable with the discriminatory ability of fatty acidand sterol composition of seeds to group Coffea accessionsinto species groups (Dussert et al., 2008). The leaf HCE dataalso separate geographical samples of C. canephora: samplesfrom the Congo share a more similar HCE profile withC. arabica, compared with representatives of the samespecies from Cameroon. This is consistent with geographicalproximity, and possibly genetic similarity, given thatC. canephora is implemented in the hybrid origin ofC. arabica (Lashermes et al., 1999; Maurin et al., 2007).
The effects of hybridization on mangiferin and HCE accumulation
The sampling of two well-documented Coffea hybrids,C. arabica and C. heterocalyx cf., provides an opportunityfor a preliminary study of the effects of hybridization on man-giferin and HCE accumulation. Coffea arabica is an allotetra-ploid, formed via the hybridization of C. eugenioides, a highaltitude species from the EC-Afr clade, and C. canephora, alow to mid-altitude species from the LG/C clade. Coffeaarabica is placed in the EC-Afr clade (i.e. withC. eugenioides) with ITS data (Lashermes et al., 1997;Maurin et al., 2007) and the LG/C clade (withC. canephora) with plastid data (Cros et al., 1998; Maurinet al., 2007). Coffea heterocalyx cf. (¼ C. sp. X ofLashermes et al., 1997; Cros et al., 1998; Table 1) has beenidentified (Cros et al., 1998; Maurin et al., 2007) as a hybrid
between C. eugenioides (EC-Afr clade; ITS data) andC. liberica (LG/C clade; plastid data). Close examination ofplastid sequence data for C. liberica var. dewevrei andC. heterocalyx cf. shows that the sequences of four plastidregions are almost identical, and thus this variety has beenimplemented as the closest parental taxon within C. liberica(Maurin et al., 2007). Direct examination of ITS and plastidsequence data produced by Maurin et al. (2007) forC. arabica and C. heterocalyx cf. shows that sequencedivergence from their progenitors is minimal, e.g. 2 or 3 bpdifferences for 3222 bp of plastid data, and 2 or 3 bp differencefor 831 bp for ITS. Based on these data and restrictionfragment length polymorphism (RFLP) results (Lashermeset al., 1999), it has been assumed that C. arabica andC. heterocalyx cf. are of recent origin, although no dates oforigin have been proposed. There are considerable differencesbetween these taxa, however, as C. arabica is an allotetraploidspecies and occurs over a relatively large area in south-westEthiopia and south-west South Sudan, and possibly northernKenya (Davis et al., 2006), whereas C. heterocalyx cf. is ofunknown origin and is so far only known from cultivation.The use of ITS and plastid sequence data represents a coarsemeans of investigating hybridization, and in most cases pro-vides only a preliminary level of understanding of hybridorigin, age and history. Nevertheless, our results for thesetwo taxa provide a useful starting point for investigating theeffects of hybridization on the phenolic accumulation inCoffea.
For C. heterocalyx cf., our results show that this taxon accu-mulates a similar [although significantly different (at P ¼0.05)] quantity of mangiferin (average 4.2 % d. wt) toC. eugenioides (average 5.7 % d. wt); C. liberica does not ac-cumulate mangiferin. For the HCE data (Table 5), there are nostatistical differences compared with C. liberica var. dewevrei(Table 5; Fig. 5); conversely, C. eugenioides shows profounddifferences in 5-CQA, total CQAs and total HCEs (Fig. 5).This pattern does not precisely follow any of the reported scen-arios for relative chemical concentrations of parental chemi-cals in first-generation (F1) hybrids, reported either asabsolute concentration or relative amounts (Orians, 2000),that is: (1) similar to that in one of the two parental taxa; (2)intermediate between those of the two parental taxa; (3) over-expressed or present in higher concentrations than in eitherparent; (4) underexpressed or present in lower concentrationsthan either parent; (5) deficient, in that hybrids lack chemicalsthat both parents produce; or (6) novel, in that hybrids maycontain chemicals lacking in both parental taxa. A better fitwould be inheritance of different chemical profiles fromeach parent: mangiferin from C. eugenioides and HCEcontent from C. liberica var. dewevrei, or it could be inter-preted as a mixture of scenarios: (1) for HCE accumulationand (4) for mangiferin content. Because plastids (and plastidsequences) are uniparentally inherited in flowering plants,our results, in combination with the available sequence data(Lashermes et al., 1997; Cros et al., 1998; Maurin et al.,2007), could be used to argue that in C. heterocalyx cf., man-giferin and HCE accumulation is linked to independent paren-tal influence, in agreement with studies on the accumulation ofvarious secondary compounds in hybrids of other plant groups(Orians, 2000). In Coffea, the indication that HCE and
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea608
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

mangiferin accumulation are derived from independentbranches of the same metabolite pathway, together with theabsence of competition for substrate availability, would cer-tainly not violate this assumption (see below: ‘Correlationbetween HCE and mangiferin accumulation’). It has beendemonstrated that maternal influence can be important in de-termining hybrid chemistry (Harney and Grant, 1963; Belzerand Ownbey, 1971; Spring and Schilling, 1990; Buschmannand Spring, 1995). For example, Buschmann and Spring(1995) found that some sesquiterpene lactones were absentin Helianthus debilis Nutt. × H. annuus L. hybrids butpresent in H. annuus × H. debilis crosses, with each hybridmore similar to the maternal taxon.
The situation observed here for C. arabica is more compli-cated. Mangiferin accumulation in C. arabica (average 0.57 %d. wt) is approx. 10-fold lower than in C. eugenioides (average5.67 % d. wt). HCE accumulation of C. arabica shares similarvalues with both C. canephora and C. eugenioides (Table 5),and in these three species HCE amounts are much higherthan in all the other Coffea species tested (Table 5). In thePCA analysis of the HCE data (Fig. 5), C. arabica fallsbetween C. canephora and C. eugenioides, but clusterscloser to the former, and is strongly influenced by 5-CQAcontent. The HCE profile of C. arabica, together with the pres-ence of mangiferin, is consistent with the assumption thatC. canephora and C. eugenioides are the progenitor speciesfor this hybrid taxon. In a scenario following the model ofhybrid influence in C. heterocalyx cf., we would assume thatin C. arabica the ability for mangiferin accumulation wasinherited from C. eugenioides, and its HCE chemistry fromC. canephora, and this is weakly supported by mangiferinand HCE accumulation, respectively (Table 5; Fig. 5) Thismode of inheritance is, however, far less convincing than forC. heterocalyx cf., given the intermediary of the HCE quan-tities and the 10-fold difference in mangiferin accumulation.An alternative scenario would be that the data represent chem-ical concentrations intermediate between those of the two par-ental taxa (Orians et al., 2000). In addition, it is reported thatF2 and later generation hybrids are both qualitatively and quan-titatively more variable than F1 hybrids (Orians, 2000), andthis provides a third possibility, i.e. that early generations ofC. arabica expressed HCE and mangiferin concentrationssimilar to the parental taxa, as in C. heterocalyx cf., andthen divergence occurred through differential selectionduring subsequent generations. This model would not be con-comitant with the sequence data (Maurin et al., 2007;A. Davis, unpubl. data), which shows negligible ITS andplastid sequence divergence between each hybrid and theirparental taxa (see above), but this is not an issue given thatmost plastid and ITS markers lack the fidelity to discriminatethe relatively short divergence times between tens, hundreds oreven thousands of generations. Even though both C. arabicaand C. heterocalyx cf. each possess only one parental ITStype (Lashermes et al., 1997; Cros et al., 1998; Maurinet al., 2007), the time periods for homogenization to eitherof the parental ITS types can vary considerably withinclosely related groups, especially where allopolyploids arepresent (Pillon et al., 2007). Moreover, ITS homogenizationcan be a rapid process (Ganley and Kobayashi, 2007), andmay occur in ,100 generations in natural hybrids (Franzke
and Mummenho, 1999), and Song et al. (1995) have demon-strated that extensive genome changes can occur during eachof just five generations. Based on the information, and if weaccept the third scenario for C. arabica as proposed above,the phenolic data presented here would support an olderhybrid origin, with a greater number of post-hybridization gen-erations, for C. arabica, compared with C. heterocalyx cf. Thiswould agree with the relatively wide natural distribution ofC. arabica (Davis et al., 2006), and the fact that this hybridtaxon functions like a species. In comparison, C. heterocalyxcf. is only known from cultivation. The fact that C. arabicais an allotetraploid might have some bearing on its chemicalinheritance, but so far the only conclusion made in thisrespect is that novel chemicals are more common in polyploidthan diploid hybrids (Orians, 2000). This effect was notrevealed by our experiments. It should also be said that weexamined a cultivated accession of C. arabica ‘Laurina’(Bourbon Pointu), a genetic mutant of cultivated stock, andthat the effects of mutation could have some influence onHCE and mangiferin accumulation. The possible influence ofgene silencing (Van Moerkercke et al., 2009) on the modifica-tion of metabolite production is unknown, and warrants inves-tigation, particularly in controlled crosses of Coffea taxa.
Correlation between HCE and mangiferin accumulation
We found no statistical correlation between mangiferin andHCE accumulation in Coffea, other than independent supportfor phylogenetic groupings of species (see above).Mangiferin biosynthesis occurs via the benzoic acid (BA) bio-synthetic route that leads to the formation of benzophenonederivatives (Abd El-Mawla and Beerhues, 2002; Liu et al.,2003; Long et al., 2009). This route is a branching occurringat the cinnamic acid step of the general phenylpropanoidpathway common to most of the phenolic compounds, includ-ing HCEs. The lack of correlation between metabolite contentshas been previously observed in Petunia hybrida (Hook.)P.L.Vilm. (Solanaceae). In this example, the independence ofthe metabolite pathways and the absence of competition forsubstrate availability have been shown to be caused by the si-lencing of a gene encoding 3-ketoacyl-CoA thiolase(PhKAT1), which is involved in the benzenoid biosyntheticpathway and the production of BA (Van Moerkercke et al.,2009).
The function of mangiferin in Coffea species
Mangiferin is a phenolic compound and, as such, one mightassume that it is involved in a response to stress. The periph-eral localization of mangiferin in Coffea plant organs, its asso-ciation with photosynthetic tissue and its accumulation at earlystages of fruit formation strengthen a hypothesis of a protec-tive action against UV-radiation in Coffea, as suggestedfor Aphloia theiformis (Vahl) Benn. (Aphloiaceae), aMadagascan shrub that abundantly accumulates mangiferinin leaves when plants develop under high light conditions(Danthu et al., 2010). Of the seven Coffea species (includingthe hybrid-derived species C. arabica, but excludingC. heterocalyx cf., which is of unknown natural origin) accu-mulating mangiferin, five are high altitude species or species
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 609
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

that have high altitude populations, where high UV conditionsmight be encountered [altitude data from Davis et al. (2006),with new altitude records incorporated]: C. anthonyi (350–900 m; mangiferin 12.06 % d. wt), C. arabica (950–2000 m;0.56 % d. wt), C. eugenioides (1000–2200 m; 5.68 % d. wt),C. pseudozanguebariae (0–800 m; 4.91 % d. wt) andC. salvatrix (400–1650; 16.36 % d. wt). Coffea sessiliflora(10–600 m; 0.03 % d. wt) would be the only exception, al-though this is a predominantly lowland species and only accu-mulates very small amounts of mangiferin. The influence ofaltitude on mangiferin accumulation has been studied in thegenus Swertia L. (Gentianaceae): S. mussotii Franch. showeda weaker mangiferin content when plants were artificially cul-tivated at lower altitude (Yang et al., 2005); whereasS. franchetiana Harry Sm. showed no altitudinal trend butinstead a significant latitudinal and longitudinal correlation(Yang et al., 2004). Measurement of ecological and geograph-ical variables, including direct measurement of light levels onleaves, in conjunction with further biochemical analysis,would be necessary to test for mangiferin accumulation corre-lations in Coffea. The distribution of mangiferin-accumulatingspecies within specific clades of Coffea may indicate that theonset of the xanthone pathway occurred independently inEast-Central Africa and East Africa (EC-Afr clade and EAclade), and perhaps in predominantly high altitude clades,after the formation of the early diverging lineages in Westand Central (Guinea/Congolian) Africa (LG/C clade and‘African Psilanthus clade’) as identified by sequencingstudies (Maurin et al., 2007; Davis et al., 2011; M. Nowak,unpubl data). Further sampling of African Coffea specieswould be required to test these ideas; investigation of high alti-tude Madagascan species, such as C. buxifolia A.Chev.(1250–2000 m), should also be undertaken. The precise distri-bution and influence of mangiferin in leaves of the commercialspecies, i.e. the predominantly high altitude (950–2000 m)C. arabica (0.56 % d. wt) and lower altitude [50–600(–1500) m] C. canephora (0 % d. wt) would be a priority, par-ticularly given recent interest in climate change mitigationusing improved varieties of these two species (Camargo,2010).
The presence of mangiferin has also been cited for its anti-oxidant and antimicrobial properties and for its contribution toplant defence against biotic stress (Franklin et al., 2009). Abetter understanding of the biosynthetic pathway of xanthonesand their glycosylated derivatives is essential to determine thelevel of involvement of mangiferin in stress response. In par-ticular, it would be necessary to isolate the genes controllingthis poorly understood pathway to study their expressionlevel in individuals, populations and species occurring in con-trasting environmental niches.
Phenolic accumulation in Coffea leaves and fruits andimplications for human use
The leaves and soft outer fruit layers of C. arabica andC. canephora are used to produce ‘tea’ (leaves) and coffee-like(outer fruit layers) beverages, particularly in Africa and Asia(Wellman, 1961; Wrigley, 1988). This use is largely unprac-tised in Europe and America, although some specialistcoffee and tea outlets sell limited amounts of coffee-leaf
‘tea’. In Africa, a hot beverage made from the outer parts ofthe fruits [‘skins’ or ‘husks’, i.e. the fruit skin (exocarp) andsoft, fleshy part of the fruit (mesocarp)] is referred to asQixr or Kishr, and in Yemen a hot drink called Qishr ismade from coffee husks and spices. In an early report,Sloane (1694) started that: ‘ . . . the Arabians themselves insummer heats use these husks [C. arabica] roasted after themanner of Coffee-berries, esteeming that drink more cooling,it being sourish to the taste.’ Coffee husks are also widelyused in Africa as a masticatory (Wellman, 1961; Wrigley,1988; A. Davis, pers. observ.). The food and health valuesof non-seed Coffea products is not well known. Early literature(Anonymous, 1876) suggests that coffee-leaf ‘tea’ has healthbenefits. Our findings, which show high levels of HCEs inthe leaves of both C. arabica and C. canephora, wouldsupport the assumption of a beneficial influence on humanhealth, given that research demonstrates high antioxidant po-tential for chlorogenic acids (Johnston et al., 2003; Stalmachet al., 2006; Parras et al., 2007), which can prevent oxidativedamage, cerebrovascular diseases, brain performance andmental health, ageing and cancer (Zang et al., 2003;Niggeweg et al., 2004; Jin et al., 2005; Mishima et al.,2005; Rajavelu et al., 2011), and have potential for chemother-apy (Belkaid et al., 2006; Cos et al., 2008). Like green tea andgreen coffee beans (Rajavelu et al., 2011), the main phenolicconstituent of coffee leaves is 5-CQA, although in certainCoffea species this is considerably exceeded by mangiferin.As said in the Introduction, mangiferin is recognized for its nu-merous pharmacological properties such as anti-inflammatory,antidiabetic, antihyperlipidaemic and neuroprotective activ-ities (Garrido et al., 2004; Muruganandan et al., 2005;Campos-Esparza et al., 2009). Information on the influenceof consuming products made from the ‘husk’ of Coffeaspecies is more elusive. It does seem compelling, however,that in C. arabica, where we demonstrate the accumulationof mangiferin in the husk and not the seed (endosperm), theformer is sometimes used in favour of the latter in the produc-tion of a beverage, given that mangiferin has potential benefi-cial properties. We report higher levels of mangiferin in the‘husk’ of C. eugenioides and C. pseudozanguebariae, com-pared with C. arabica, but have not been able to trace use ofthese species in beverage production or as a masticatory.Further research on the use and properties of coffee-leaf‘tea’ and coffee-husk coffee would be worthwhile.
CONCLUSIONS
We have demonstrated mangiferin accumulation in sevenAfrican Coffea taxa: C. anthonyi, C. arabica, C. eugenioides,C. heterocalyx cf., C. pseudozanguebariae, C. salvatrix andC. sessiliflora. Previously the presence of mangiferin had onlybeen detected in a single species, i.e. C. pseudozanguebariae(Talamond et al., 2008). Histochemical methods show that inCoffea mangiferin is localized in leaves and outer layers ofthe fruit, in the same regions as the photosynthetic tissue.These data, in conjunction with published work on othermangiferin-accumulating species, and the systematic distribu-tion of mangiferin in high altitude clades and species ofCoffea, suggest an association with altitude.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea610
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Statistical analysis of mangiferin and HCE accumulationidentified significant difference between species, and also pro-vided the means to place samples into: (1) clades establishedon the basis of molecular phylogenetic analyses (Maurinet al., 2007; Anthony et al., 2010; Davis et al., 2011;Hamon et al., 2011); (2) species-specific groups (with excep-tions) agreeing with molecular and morphological delimita-tion; and (3) population groups (for C. canephora). It isclear that leaf phenolic data have considerable phylogeneticutility in Coffea, at a broad range of taxonomic levels.Combining phenolic data with larger, independent data sets,such as DNA sequence data sets, is likely to be especiallyrewarding. Knowledge of the spectrum of phenolic accumula-tion in leaves and fruits of coffee species and populationsmight be of significance for understanding insect herbivory,and environmental adaptation, including climate changemitigation.
Sampling of two known hybrid taxa, C. arabica andC. heterocalyx cf., provided an opportunity to make a prelim-inary assessment of the effects of hybridization on the phenol-ic composition of Coffea leaves. We suggest that mangiferinand HCE accumulation are controlled by independent parentalinfluence, an assumption that is not a variance with the indica-tion that these two groups of compounds are derived from in-dependently regulated metabolite pathways. Further researchwith controlled crossings and their influence on phenolic accu-mulation in leaves, and seeds (beans), of Coffea could be ofsignificant value for coffee breeding and crop development, in-cluding breeding strategies for climate change mitigation(Camargo, 2010; Jaramillo et al., 2011).
The potential health benefits of coffee-leaf ‘tea’, and bev-erages and masticatory products made from the fleshy partsof Coffea fruit, are supported by our findings on the basis ofhigh accumulation of phenolic compounds in coffee leaves, al-though the accumulated influence of these compounds in thehuman body would require further research.
ACKNOWLEDGEMENTS
We would like to thank the National Center of AppliedResearch and Rural Development (Foibe Fikarohana ampihar-ina amin’ny Fampandrosoana ny eny Ambanivohitra:FOFIFA), and the Ueshima Coffee Corporation (UCC)Japan, for allowing us to access the collections at KianjavatoCoffee Research Station (FOFIFA), and Dokou Akaffoufrom the CNRA station of DIVO (Ivory Coast), for use of ma-terial. The authors also wish gratefully to acknowledge:Y. Baissac (UM2) for LC-MS support; T. Roscoe (CNRS)for critical reading and commenting on an earlier version ofthis contribution; C. Andary (Faculty of Pharmacy) for hisgenerous gift of a 3,5-diCQA sample; P. Hamon andM. Noirot (IRD) for support and advice; and two anonymousreviewers for their constructive comments. This work was par-tially funded by a grant from the Agence UniversitaireFrancophone (AUF).
LITERATURE CITED
Abd El-Mawla AMA, Beerhues L. 2002. Benzoic acid biosynthesis in cellcultures of Hypericum androsaemum. Planta 214: 727–733.
Alonso-Salces RM, Guillou C, Berrueta LA. 2009. Liquid chromatographycoupled with ultraviolet absorbance detection, electrospray ionization,collision-induced dissociation and tandem mass spectrometry on a triplequadrupole for the on-line characterization of polyphenols and methyl-xanthines in green coffee beans. Rapid Communications in MassSpectrometry 23: 363–383.
Anonymous. 1876. Coffee-leaf tea. British Medical Journal 2(830): 691.
Anthony F, Diniz LEC, Combes M-C, Lashermes P. 2010. Adaptive radi-ation in Coffea subgenus Coffea L. (Rubiaceae) in Africa andMadagascar. Plant Systematics and Evolution 285: 51–64.
Anthony F, Noirot M, Clifford MN. 1993. Biochemical diversity in the genusCoffea L.: chlorogenic acids, caffeine, and mozambioside contents.Genetic Resources and Crop Evolution 40: 61–70.
Barreto JC, Trevisan MTS, Hull WE, et al. 2008. Characterization andquantitation of polyphenolic compounds in bark, kernel, leaves, andpeel of mango (Mangifera indica L.). Journal of Agricultural and FoodChemistry 56: 5599–5610.
Belay A. 2011. Some biochemical compounds in coffee beans and methodsdeveloped for their analysis. International Journal of the PhysicalSciences 6: 6373–6378.
Belkaid A, Currie J-C, Desgagnes J, Annabi B. 2006. The chemopreventiveproperties of chlorogenic acid reveal a potential new role for the micro-somal glucose-6-phosphate translocase in brain tumor progression.Cancer Cell International 6: 7.
Belzer NF, Ownbey M. 1971. Chromatographic comparisons of Tragopogonspecies and hybrids. American Journal of Botany 58: 791–802.
Bertrand C, Noirot M, Doulbeau S, de Kochko A, Hamon S, Campa C.2003. Chlorogenic acid content swap during fruit maturation in Coffeapseudozanguebariae. Qualitative comparison with leaves. Plant Science165: 1355–1361.
Bolzani V da S, Trevisan LMV, Young CM. 1991. Caffeic acid esters andtriterpenes of Alibertia macrophylla. Phytochemistry 30: 2089–2091.
Boudet AM. 2007. Evolution and current status of research in phenolic com-pounds. Phytochemistry 68: 2722–2735.
Bridson DM. 1988. Coffea. In: Polhill RM. ed. Flora of Tropical East Africa,Rubiaceae,2. Rotterdam/Brookfield:Balkema, 415–474.
Buschmann H, Spring O. 1995. Sesquiterpene lactones as a result of interspe-cific hybridization in Helianthus species. Phytochemistry 39: 367–371.
Camargo MBP. 2010. The impact of climatic variability and climate changeon Arabic coffee crop in Brazil. Bragantia 69: 239–247.
Campa C, Ballester JF, Doulbeau S, Dussert S, Hamon S, Noirot M. 2004.Trigonelline and sucrose diversity in wild Coffea species. Food Chemistry88: 39–43.
Campa C, Doulbeau S, Dussert S, Hamon S, Noirot M. 2005. Qualitativerelationship between caffeine and chlorogenic acid contents among wildCoffea species. Food Chemistry 93: 135–139.
Campa C, Rakotomalala J-JR, de Kochko A, Hamon S. 2008. Chlorogenicacids: diversity in green beans of wild coffee species. In: HemantaranjanA. ed. Advances in plant physiology, Vol. 10. Jodhpur: ScientificPublishers, 421–437.
Campos-Esparza MR, Sanchez-Gomez MV, Matute C. 2009. Molecularmechanisms of neuroprotection by two natural antioxidant polyphenols.Cell Calcium 45: 358–368.
Clifford MN, Williams T, Bridson DM. 1989. Chlorogenic acids and caffeineas possible taxonomic markers in Coffea and Psilanthus. Phytochemistry28: 829–838.
Clifford MN, Johnston KL, Knight S, Kuhnert N. 2003. Hierarchicalscheme for LC-MSn identification of chlorogenic acids. Journal ofAgricultural and Food Chemistry 51: 2900–2911.
Clifford MN, Knight S, Surucu B, Kuhnert N. 2006. Characterizationby LC-MSn of four new classes of chlorogenic acids in greencoffee beans: dimethoxycinnamoylquinic acids, diferuloylquinic acids,caffeoyl-dimethoxycinnamoylquinic acids, and feruloyl-dimethoxycinna-moylquinic acids. Journal of Agricultural and Food Chemistry 54:1957–1969.
Clifford MN, Kirkpatrick J, Kuhnert N, Roorendaal H, Salgado PR. 2008.LC-MSn analysis of the cis isomers of chlorogenic acids. Food Chemistry106: 379–385.
Cos P, Maes L, Vlietinck A, Pieters L. 2008. Plant-derived leading com-pounds for chemotherapy of human immunodefiency virus (HIV) infec-tion – an update (1998–2007). Planta Medica 74: 1323–1337.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 611
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Cros J, Combes MC, Trouslot P, Anthony F, Hamon S, Charrier A,Lashermes P. 1998. Phylogenetic analysis of chloroplast DNA variationin Coffea L. Molecular Phylogenetics and Evolution 9: 109–117.
Danthu P, Lubrano C, Flavet L, et al. 2010. Biological factors influencingproduction of xanthones in Aphloia theiformis. Chemistry andBiodiversity 7: 140–150.
Davis AP. 2011. Psilanthus mannii, the type species of Psilanthus, transferredto Coffea. Nordic Journal of Botany 29: 471–472.
Davis AP, Rakotonasolo F. 2008. A taxonomic revision of the baracoffea al-liance: nine remarkable Coffea species from western Madagascar.Botanical Journal of the Linnean Society 158: 355–390.
Davis AP, Govaerts R, Bridson DM, Stoffelen P. 2006. An annotated taxo-nomic conspectus of the genus Coffea (Rubiaceae). Botanical Journal ofthe Linnean Society: 152: 465–512.
Davis AP, Tosh J, Ruch N, Fay MF. 2011. Growing coffee: Psilanthus(Rubiaceae) subsumed on the basis of plastid and nuclear DNAsequences; implications for the size, morphology, distribution and evolu-tionary history of Coffea. Botanical Journal of the Linnean Society 167:357–377.
De Castro RD, Marraccini P. 2006. Cytology, biochemistry and molecularchanges during coffee fruit development. Brazilian Journal of PlantPhysiolgy 18: 175–199.
Dussert S, Laffargue A, de Kochko A, Joet T. 2008. Effectiveness of thefatty acid and sterol composition of seeds for the chemotaxonomy ofCoffea subgenus Coffea. Phytochemistry 69: 2950–2960.
Eyles A, Bonello P, Ganley R, Mohammed C. 2010. Induced resistance topests and pathogens in trees. New Phytologist 185: 893–908.
Franklin G, Conceicao LFR, Kombrink E, Dias ACP. 2009. Xanthone bio-synthesis in Hypericum perforatum cells provides antioxidant and anti-microbial protection upon biotic stress. Phytochemistry 70: 60–68.
Franzke A, Mummenho K. 1999. Recent hybrid speciation in Cardamine(Brassicaceae) – conversion of nuclear ribosomal ITS sequences instatu nascendi. Theoretical Applied Genetics 98: 831–834.
Gallego-Giraldo L, Jikumaru Y, Kamiya Y, Tang Y, Dixon RA. 2011.Selective lignin down regulation leads to constitutive defence responseexpression in alfalfa (Medicago sativa L.). New Phytologist 190:627–639.
Ganley ARD, Kobayashi T. 2007. Highly efficient concerted evolution in theribosomal DNA repeats: total rDNA repeat variation revealed by whole-genome shotgun sequence data. Genome Research 17: 184–191.
Garrido G, Gonzalez D, Lemusa Y, et al. 2004. In vivo and in vitro anti-inflammatory activity of Mangifera indica L. extract (VIMANGw).Pharmacological Research 50: 143–149.
Hamon P, Duroy P-O, Dubreuil-Tranchant C, et al. 2011. Two novelTy1-copia retrotransposons isolated from coffee trees can effectivelyreveal evolutionary relationships in the Coffea genus (Rubiaceae).Molecular Genetics and Genomics 285: 447–460.
Harney PM, Grant WF. 1963. Biochemical anomaly in flower extracts ofinterspecific hybrids between Lotus species. Science 142: 1061.
Herrmann K. 1989. Occurrence and content of hydroxycinnamic and hydro-xybenzoic acid compounds in foods. Critical Reviews in Food Scienceand Nutrition 28: 315–347.
International Coffee Organization (ICO). 2011. World Coffee Trade. [http://www.ico.org/trade_e.asp?section=About_Coffee]. Accessed 3 January2012.
Jaiswal R, Patras MA, Eravuchira PJ, Kuhnert N. 2010a. Profile and char-acterization of the chlorogenic acids in green robusta coffee beans byLC-MSn: identification of seven new classes of compounds. Journal ofAgricultural and Food Chemistry 58: 8722–8737.
Jaiswal R, Sovdat T, Vivan F, Kuhnert N. 2010b. Profiling and characteriza-tion by LC-MSn of the chlorogenic acids and hydroxycinnamoylshikimateesters in Mate (Ilex paraguariensis). Journal of Agricultural and FoodChemistry 58: 5471–5484.
Jaramillo J, Muchugu E, Vega FE, Davis AP, Borgemeister C,Chabi-Olaye A. 2011. Some like it hot: the influence and implicationsof climate change on coffee berry borer (Hypothenemus hampei) andcoffee production in East Africa. PLoS One 6: e24528. http://dx.doi.org/10.1371/journal.pone.0024528.
Jin UH, Lee JY, Kang SK, Kim JK, Park WH, Kim JG, Moon SK, KimCH. 2005. A phenolic compound, 5-caffeoylquinic acid (chlorogenicacid), is a new type and strong matrix metalloproteinase-9 inhibitor: iso-lation and identification from methanol extract of Euonymus alatus. LifeSciences 77: 2760–2769.
Johnston KL, Clifford MN, Morgan LM. 2003. Coffee acutely modifiesgastrointestinal hormone secretion and glucose tolerance in humans: gly-cemic effects of chlorogenic acid and caffeine. American Journal ofClinical Nutrition 78: 728–733.
Kuhnert N, Jaiswal R, Eravuchira PJ, El-Abassy RM, von der Kammer B,Materny A. 2011. Scope and limitations of principal component analysisof high resolution LC-TOF-MS data: the analysis of the chlorogenic acidfraction in green coffee beans as a case study. Analytical Methods 3:144–155.
Lashermes P, Combes M-C, Trouslot P, Charrier A. 1997. Phylogeneticrelationships of coffee-tree species (Coffea L.) as inferred from ITSsequences of nuclear ribosomal DNA. Theoretical and AppliedGenetics 94: 947–955.
Lashermes P, Combes MC, Robert J, Trouslot P, D’Hont A, Anthony F,Charrier A. 1999. Molecular characterisation and origin of the Coffeaarabica L. genome. Molecular and General Genetics 261: 259–266.
Lepelley M, Cheminade G, Tremillon N, Simkin A, Caillet V, McCarthy J.2007. Chlorogenic acid synthesis in coffee: an analysis of CGA contentand real-time RT-PCR expression of HCT, HQT, C3H1, andCCoAOMT1 genes during grain development in C. canephora. PlantScience 172: 978–996.
Lewin B, Giovannucci D, Varangis P. 2004. Coffee markets: new paradigmsin global supply and demand. International Bank for Reconstruction andDevelopment, Agriculture and Rural Development Discussion Paper 3.
Liu B, Falkenstein-Paul H, Schmidt W, Beerhues L. 2003. Benzophenonesynthase and chalcone synthase from Hypericum androsaemum cell cul-tures: cDNA cloning, functional expression, and site-directed mutagen-esis of two polyketide synthases. The Plant Journal 34: 847–855.
Long MC, Nagegowda DA, Kaminaga Y, et al. 2009. Involvement of snap-dragon benzaldehyde dehydrogenase in benzoic acid biosynthesis. ThePlant Journal 59: 256–265.
Lopes CR, Monaco LC. 1979. Chemotaxonomic studies of some species ofthe genus Coffea. Journal of Plantation Crops 7: 6–14.
Lopes CR, Shepherd GP. 1991. Phylogenetic studies of some species of thegenus Coffea – 1 – numerical analysis of flavonoid compounds.Brazilian Journal of Genetics 14: 425–435.
Lopes CR, Musche R, Hec M. 1984. Identificacao de pigmentos flavonoides eacidos fenolicos nos cultivares Bourbon e Caturra de Coffea arabica.Brazilian Journal of Genetics 7: 657–669.
Macheix J-J, Fleuriet A, Billot J. 1990. Changes and metabolism of phenoliccompounds in fruits. In: Macheix J-J, Fleuriet A, Billot J. eds. Fruit phe-nolics. Boca Raton, FL: CRC Press, 149–237.
Mahesh V, Million-Rousseau R, Ullmann P, et al. 2007. Functional charac-terization of two p-coumaroyl ester 3’-hydroxylase genes from coffeetree: evidence of a candidate for chlorogenic acid biosynthesis. PlantMolecular Biology 64: 145–159.
Manach C, Scalbert A, Morand C, Remesy C, Jimenez L. 2004.Polyphenols: food source and bioavailability. American Journal ofClinical Nutrition 79: 727–747.
Matsui Y, Nakamura S, Kondou N, Takasu Y, Ochiai R, Masukawa Y.2007. Liquid chromatography-electrospray ionization-tandem mass spec-trometry for simultaneous analysis of chlorogenic acids and their metabo-lites in human plasma. Journal of Chromatography B 858: 96–105.
Maurin O, Davis AP, Chester M, Mvungi EF, Jaufeerally-Fakim Y, FayMF. 2007. Towards a phylogeny for Coffea (Rubiaceae): identifyingwell-supported lineages based on nuclear and plastid DNA sequences.Annals of Botany 100: 1565–1583.
Mishima S, Inoh Y, Narita Y, et al. 2005. Identification of caffeoylquinicacid derivatives from Brazilian propolis as constituents involved in induc-tion of granulocytic differentiation of HL-60 cells. Bioorganic andMedicinal Chemistry 13: 5814–5818.
Moglia A, Lanteri S, Comino C, Acquadro A, de Vos R, Beekwilder J.2008. Stress-induced biosynthesis of dicaffeoylquinic acids in globe arti-choke. Journal of Agricultural and Food Chemistry 56: 8641–8647.
Mondolot L, La Fisca P, Buatois B, Talansier E, de Kochko A, Campa C.2006. Caffeoylquinic acid content and histolocalization in Coffea cane-phora developing leaves. Annals of Botany 98: 33–40.
Muruganandan S, Srinivasan K, Gupta S, Gupta PK, Laj J. 2005. Effect ofmangiferin on hyperglycemia and atherogenicity in streptozotocin diabet-ic rats. Journal of Ethnopharmacology 97: 497–501.
Niggeweg R, Michael AJ, Martin C. 2004. Engineering plants with increasedlevels of the antioxidant chlorogenic acids. Nature Biotechnology 22:746–754.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea612
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from

Orians CM. 2000. The effects of hybridization in plants on secondary chem-istry: implications for the ecology and evolution of plant–herbivore inter-actions. American Journal of Botany 87: 1749–1756.
Parras P, Martınez-Tome M, Jimenez AM, Murci MA. 2007. Antioxidantcapacity of coffees of several origins brewed following three differentprocedures. Food Chemistry 102: 582–592.
Pillon Y, Fay MF, Hedren M, et al. 2007. Evolution and temporal diversifi-cation of Western European polyploid species complexes in Dactylorhiza(Orchidaceae). Taxon 56: 1185–1208.
Rajavelu A, Tulyasheva Z, Jaiswal R, Jeltsch A, Kuhnert N. 2011. The in-hibition of the mammalian DNA methyltransferase 3a (Dnmt3a) bydietary black tea and coffee polyphenols. BMC Biochemistry 12: 16.http://dx.doi.org/10.1186/1471-2091-12-16.
Rakotomalala JJR. 1992. Diversite biochimique des cafeiers: analyse desacides hydroxycinnamiques, bases puriques et diterpenes glycosidiques.Particularites des cafeiers sauvages de la region malgache(Mascarocoffea Chev.). PhD thesis, University of Montpellier 2, France.
Read J, Sanson GD, Caldwell E, et al. 2009. Correlations between leaf tough-ness and phenolics among species in contrasting environments ofAustralia and New Caledonia. Annals of Botany 103: 757–767.
Ribereau-Gayon P. 1968. Les composes phenoliques des vegetaux. Paris: Dunod.Santiago LJM, Louro RP, De Oliveira DE. 2000. Compartmentation of
phenolic compounds and phenylalanine ammonia-lyase in leaves ofPhyllanthus tenellus Roxb. and their induction by copper sulphate.Annals of Botany 86: 1023–1032.
Schuster B, Herrmann K. 1985. Hydroxybenzoic and hydroxycinnamic acidderivatives in soft fruits. Phytochemistry 24: 2761–2764.
Shahidi F, Chandrasekara A. 2010. Hydroxycinnamates and their in vitroand in vivo antioxidant activities. Phytochemistry Reviews 9: 147–170.
Sloane H. 1694. An account of a prodigiously large feather of the bird cuntur,brought from Chili, and supposed to be a kind of vultur; and of thecoffee-shrub. Philosophical Transactions 18: 61–64.
Song K, Lu P, Tang K, Osborn TC. 1995. Rapid genome change in syntheticpolyploids of Brassicaceae and its implications for polyploid evolution.Proceedings of the National Academy of Sciences, USA 92: 7719–7723.
Spring O, Schilling EE. 1989. Chemosystematic investigation of the annualspecies of Helianthus (Asteraceae). Biochemical Systematics and Ecology17: 519–528.
Stalmach A, Mullen W, NagaiI C, Crozier A. 2006. On-line HPLC analysisof the antioxidant activity of phenolic compounds in brewed, paper-filtered coffee. Brazilian Journal of Plant Physiology 18: 253–262.
Stoffelen P, Noirot M, Couturon E, Bontems S, De Block P, Anthony F.2009. Coffea anthonyi, a new self-compatible Central African coffeespecies, closely related to an ancestor of Coffea arabica. Taxon 58:133–140.
Suryawanshi S, Asthana RK, Gupta RC. 2007. Simultaneous estimation ofmangiferin and four secoiridoid glycosides in rat plasma using liquidchromatography tandem mass spectrometry and its application to pharma-cokinetic study of herbal preparation. Journal of Chromatography, B 858:211–219.
Talamond P, Mondolot L, Gargadennec A, et al. 2008. First report on man-giferine (C-glucosyl-xanthone) isolated from leaves of a wild coffeeplant, Coffea pseudozanguebariae (Rubiaceae). Acta Botanica Gallica155: 513–519.
Van Moerkercke A, Schauvinhold I, Pichersky E, Haring MA,Schuurink RC. 2009. A plant thiolase involved in benzoic acid biosyn-thesis and volatile benzenoid production. The Plant Journal 60:292–302.
Vega FE, Rosenquist E, Collins W. 2003. Global project needed to tacklecoffee crisis. Nature 435: 343.
Wellman FL. 1961. Coffee, botany, cultivation and utilization. London:Leonard Hill Books.
White F. 1979. The Guineo-Congolian Region and its relationships to otherphytochoria. Bulletin du Jardin Botanique National de Belgique 49:11–55.
White F. 1983. The vegetation of Africa. A descriptive memoir to accompanythe Unesco/AETFAT/UNSO vegetation map of Africa. Paris: Unesco.
Wink M. 2003. Evolution of secondary metabolites from an ecological andmolecular phylogenetic perspective. Phytochemistry 64: 3–19.
Wrigley G. 1988. Coffee – tropical agriculture series. Harlow, UK: LongmanScientific & Technical.
Yang H, Duan Y, Hu F, Liu J. 2004. Lack of altitudinal trends in phytochem-ical constituents of Swertia franchetiana (Gentianaceae). BiochemicalSystematics and Ecology 32: 861–866.
Yang H, Ding C, Duan Y, Liu J. 2005. Variation of active constituents of animportant Tibet folk medicine Swertia mussotii Franch. (Gentianaceae)between artificially cultivated and naturally distributed. Journal ofEthnopharmacology 98: 31–35.
Zang L-Y, Cosma G, Gardner H, Castranoca V, Vallyathan V. 2003. Effectof chlorogenic acid on hydroxyl radical. Molecular and CellularBiochemistry 247: 205–210.
Campa et al. — Mangiferin and hydroxycinnamic acid ester accumulation in Coffea 613
by guest on June 3, 2016http://aob.oxfordjournals.org/
Dow
nloaded from




















