
A spinose stem group brachiopod with pedicle from the Middle Cambrian Burgess Shale: Middle Cambrian...
18
Acta Zoologica (Stockholm) 87: 273– 290 ( October 2006) © 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences Abstract Holmer, L.E. and Caron, J-B. 2006. A soft-shelled spinose stem group brachiopod with pedicle from the Middle Cambrian Burgess Shale. — Acta Zoologica (Stockholm) 87: 273–290 A new genus and species of a Middle Cambrian stem group brachiopod, Acanthotretella spinosa n. gen. and n. sp., is described from the Burgess Shale Formation. Most of the 42 specimens studied came from the Greater Phyllopod bed (Walcott Quarry) and were collected from five bed assemblages, each representing a single obrution event. Specimens are probably preserved within their original habitat. In contrast to all brachiopods known from the Burgess Shale, the shells of the new stem group brachiopod are often deformed and do not show signs of brittle breakage, which suggests that the valves were originally either entirely organic in composition or, more likely, had just a minor mineral component. Acanthotretella spinosa differs from all the other described Cambrian brachiopods in that it is covered by long, slender and possibly partly mineralized spines that are posteriorly inclined at an oblique angle away from the anterior margin. The spines penetrate the shell and are mainly comparable with the thorn-like organic objects that have been inferred from early siphonotretoid brachiopods. The pedicle was slender and was composed of a central coelomic region and emerged from an apical foramen at the end of an internal pedicle tube. The finding of a pedicle attached to the macrobenthic algae Dictyophycus and other epibenthos implies that A. spinosa did not have an infaunal mode of life. The visceral region and interior characters are poorly preserved. Dr L. E. Holmer, Uppsala University, Department of Earth Sciences/ Palaeobiology, Norbyv. 22, SE-75236 Uppsala, Sweden. E-mail: [email protected] Blackwell Publishing Ltd A spinose stem group brachiopod with pedicle from the Middle Cambrian Burgess Shale Lars Erik Holmer 1 and Jean-Bernard Caron 2 1 Uppsala University, Department of Earth Sciences/Palaeobiology, Norbyv. 22, SE- 75236 Uppsala, Sweden; 2 Department of Natural History, Royal Ontario Museum, 100 Queen’s Park, Toronto, Ontario, Canada M5S 2C6 Keywords: epibenthos, filter-feeder, fossil Lagerstätten, lophophorate, non-biomineralized preservation, siphonotretoid Accepted for publication: 26 June 2006 Introduction The Burgess Shale brachiopods have not been re-evaluated since Walcott (1912, 1924) and Resser (1938). Most species are only known from their shells, which are thought to have been originally biomineralized (see e.g. Briggs et al. 1994) such as ‘Lingulella’ waptensis Walcott, Acrothyra gregaria Walcott and Diraphora bellicostata (Walcott). As for other organisms from the Burgess Shale (e.g. Scenella, trilobites, Haplophrentis) biomineralization of the shells is mainly inferred based on their three-dimensional preservation and on the presence of cracks created by compression during diagenesis. The original mineral composition of the shell, however, remains unknown. Claims that some species retained their original calcium phosphate composition (Briggs et al. 1994) have not been confirmed. The only previously known examples of exceptional preservation of non-biomineralized characters in Burgess Shale brachiopods were described from the paterinids ( Linguliformea, Paterinata, Paterinida). Micromitra burgessensis Resser is certainly the most famous example. This species is often preserved with the delicate, long setae along the margins of the biomineralized shells. The presence of setae and the fact that many speci- mens are attached to hard substrates, commonly on spicules of the sponge Pirania (e.g. Walcott 1912, 1924; Whittington 1985; Briggs et al. 1994), support the view that these speci- mens were buried alive or soon after death. The presence of setae in other species is rather dubious. The long marginal
Transcript of A spinose stem group brachiopod with pedicle from the Middle Cambrian Burgess Shale: Middle Cambrian...

Acta Zoologica
(Stockholm)
87
: 273–290 (October 2006)
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences
Abstract
Holmer, L.E. and Caron, J-B. 2006. A soft-shelled spinose stem group brachiopodwith pedicle from the Middle Cambrian Burgess Shale. —
Acta Zoologica
(Stockholm)
87
: 273–290
A new genus and species of a Middle Cambrian stem group brachiopod,
Acanthotretella spinosa
n. gen. and n. sp., is described from the Burgess ShaleFormation. Most of the 42 specimens studied came from the Greater Phyllopodbed (Walcott Quarry) and were collected from five bed assemblages, eachrepresenting a single obrution event. Specimens are probably preserved withintheir original habitat. In contrast to all brachiopods known from the BurgessShale, the shells of the new stem group brachiopod are often deformed and donot show signs of brittle breakage, which suggests that the valves were originallyeither entirely organic in composition or, more likely, had just a minor mineralcomponent.
Acanthotretella spinosa
differs from all the other described Cambrianbrachiopods in that it is covered by long, slender and possibly partly mineralizedspines that are posteriorly inclined at an oblique angle away from the anteriormargin. The spines penetrate the shell and are mainly comparable with thethorn-like organic objects that have been inferred from early siphonotretoidbrachiopods. The pedicle was slender and was composed of a central coelomicregion and emerged from an apical foramen at the end of an internal pedicletube. The finding of a pedicle attached to the macrobenthic algae
Dictyophycus
and other epibenthos implies that
A. spinosa
did not have an infaunal mode oflife. The visceral region and interior characters are poorly preserved.
Dr L. E. Holmer, Uppsala University, Department of Earth Sciences/Palaeobiology, Norbyv. 22, SE-75236 Uppsala, Sweden. E-mail: [email protected]
Blackwell Publishing Ltd
A spinose stem group brachiopod with pedicle from the Middle Cambrian Burgess Shale
Lars Erik Holmer
1
and Jean-Bernard Caron
2
1
Uppsala University, Department of Earth Sciences/Palaeobiology, Norbyv. 22, SE-75236 Uppsala, Sweden;
2
Department of Natural History, Royal Ontario Museum, 100 Queen’s Park, Toronto, Ontario, Canada M5S 2C6
Keywords:
epibenthos, filter-feeder, fossil Lagerstätten, lophophorate, non-biomineralized preservation, siphonotretoid
Accepted for publication:
26 June 2006
Introduction
The Burgess Shale brachiopods have not been re-evaluatedsince Walcott (1912, 1924) and Resser (1938). Most speciesare only known from their shells, which are thought to havebeen originally biomineralized (see e.g. Briggs
et al
. 1994)such as ‘
Lingulella
’
waptensis
Walcott,
Acrothyra gregaria
Walcott and
Diraphora bellicostata
(Walcott). As for otherorganisms from the Burgess Shale (e.g.
Scenella
, trilobites,
Haplophrentis
) biomineralization of the shells is mainlyinferred based on their three-dimensional preservation andon the presence of cracks created by compression duringdiagenesis. The original mineral composition of the shell,however, remains unknown. Claims that some species
retained their original calcium phosphate composition(Briggs
et al
. 1994) have not been confirmed. The onlypreviously known examples of exceptional preservation ofnon-biomineralized characters in Burgess Shale brachiopodswere described from the paterinids (Linguliformea, Paterinata,Paterinida).
Micromitra burgessensis
Resser is certainly themost famous example. This species is often preserved withthe delicate, long setae along the margins of the biomineralizedshells. The presence of setae and the fact that many speci-mens are attached to hard substrates, commonly on spiculesof the sponge
Pirania
(e.g. Walcott 1912, 1924; Whittington1985; Briggs
et al
. 1994), support the view that these speci-mens were buried alive or soon after death. The presence ofsetae in other species is rather dubious. The long marginal

Middle Cambrian stem group brachiopod
•
Holmer and Caron Acta Zoologica
(Stockholm)
87
: 273–290 (October 2006)
© 2006 The Authors
Journal compilation © 2006 The Royal Swedish Academy of Sciences
structures associated with a single specimen of
Nisusiaburgessensis
Walcott (Rhynchonelliformea, Kutorginata),and illustrated by Walcott in one of his monographs (Walcott1924; pl. 110 : 1) are probably not setae because one of thesestructure appears to be branching. A random associationbetween
Nisusia
and putative algae or cyanobacteria (e.g.
Marpolia
) is probably more likely in this case.Brachiopods with preserved pedicles from the Burgess
Shale were first reported by Jin
et al
. (1993; Fig. 3 : 6), whodiscovered an incomplete and poorly preserved specimen(USNM356645) with the proximal part of the pedicle,which was referred to as
Lingulella
sp. Here we describean additional 42 specimens, mostly complete of the samepedunculate form as
Acanthotretella spinosa
n. gen. and n. sp.Most of the new specimens were collected
in-situ
in five bedassemblages from a single Burgess Shale locality, the GreaterPhyllopod bed (Walcott Quarry). This species was provisionallyidentified in an earlier palaeoecological study as a non-biomineralized lingulid with pedicle,
Lingulella
sp. A (Caron2005a), but had not been described formally. The new materialdemonstrates that the combination of characters in
A. spinosa
is unlike that of any previously described brachiopod and itis referred to the stem group of the Brachiopoda.
Previous records of soft part preservation in Cambrian brachiopods
The records of exceptional preservation of non-biomineralizedparts within Cambrian Brachiopoda have increased rapidlyduring the last 15 years. The most important new findings arefrom the Lower Cambrian Chengjiang Fauna (‘Lagerstätte’)of Chengjiang County, Yunnan, South China, where a totalof six brachiopods with preservation of non-biomineralizedparts (including impressions of the pedicle, setae, lophophoreand digestive tracts) have been described to date; the faunallist comprises the following:
Lingulellotreta malongensis
Rong,‘
Lingulella
’
chengjiangensis
Jin, Hou and Wang,
Heliomedusaorienta
Sun and Hou,
Xianshanella haikouensis
Zhang andHan,
Diandongia pista
Rong, and
Longtancunella chengjiangensis
Hou, Bergström, Wang, Feng and Chen (see e.g., Jin andWang 1992; Jin
et al
. 1993; Hou
et al
. 1999; Zhang X.G.
et al
. 2003; Zhang Z.F.
et al
. 2003; Hou
et al
. 2004; Zhangand Han 2004; Zhang
et al
. 2004a; Jin 2005).
Pedicle
All previously described, well-preserved Cambrian pedicleimpressions come from the Chengjiang Fauna, where four ofthe six better known brachiopod species have been found tohave relatively long pedicles (Jin
et al
. 1993; Zhang X.G.
et al
. 2003; Zhang Z.F.
et al
. 2003; Hou
et al
. 2004; Zhangand Han 2004; Zhang
et al
. 2004a, 2004b, 2005, 2006, inpress).
Jin
et al
. (1993) also reviewed the known fossil record oflingulid-like pedicle impressions and compared them in
detail with the pedicles of modern lingulid brachiopods.Lingulids with this type of pedicle may have lived infaunallyin burrows like their modern representatives (e.g., Jin
et al
.1993), semi-infaunally (e.g., Holmer
et al
. 1997; Hou
et al
.2004; Jin 2005) or epifaunally. Direct evidence for anepifaunal life habit are known both from the two problematicbrachiopods
Longtancunella chengjiangensis
Hou, Bergström,Wang, Feng and Chen and
X. haikouensis
Zhang and Han, aswell as the botsfordiid
Diandongia pista
Rong, which all havebeen found attached to various hard epibenthic substrates(Zhang Z.F.
et al
. 2003; Hou
et al
. 2004; Zhang and Han2004; Zhang
et al
. 2006, in press). However, the detailedmorphology of the pedicles varies (see further below).
Setae
Although the presence of various soft organic setal structurescan be inferred from the study of morphological charactersof the shells in Cambrian brachiopods (e.g., Williams andHolmer 1992; Skovsted and Holmer 2003; Balthasar 2004;Williams
et al
. 2004) preservation of the setae themselves isextremely rare. Apart from the Burgess Shale taxa
Micromitra
,all other records are from the Chengjiang Fauna, whereprimarily the enigmatic
Heliomedusa orienta
Sun and Hou(Jin and Wang 1992) have extraordinarily well-preservedsetae. More recently, various types of exceptionally preservedsetae have been described from
L. malongensis
(Rong),‘
Lingulella
’
chengjiangensis
Jin, Hou and Wang,
X. haikouensis
Zhang and Han and
Longtancunella chengjiangensis
Hou,Bergström, Wang, Feng and Chen (Zhang
et al
. 2005, 2006,in press; see further below).
Lophophore and digestive system
Exceptional preservation of the lophophore and digestivetract is most unusual, and again the only Cambrian recordscome from the Chengjiang Fauna. The lingulids
L. malongensis
(Rong), ‘
Lingulella
’
chengjiangensis
Jin, Hou and Wang andthe two problematic brachiopods
Longtancunella chengjiangensis
Hou, Bergström, Wang, Feng and Chen and
X. haikouensis
Zhang and Han all have preserved imprints of a trocholophe,schizolophe to simple spiroloph as well as a U-shaped digestivesystem, much like that of Recent linguloids. In contrast
Heliomedusa orienta
Sun and Hou has a unique and enigmatictype of lophophore, which was first described in detail byZhang
et al
. (2003); but see also Jin and Wang (1992). Thedigestive system of this taxon, however, is not well preserved.
Stratigraphical setting, depositional environment and synecology
This study is based on 42 specimens from the BurgessShale Formation housed at the Royal Ontario Museum. Allthe specimens, except for three, were collected
in situ
onFossil Ridge in British Columbia (Fig. 1). Thirty-nine speci-mens came from the Greater Phyllopod Bed (GPB, Walcott

Acta Zoologica
(Stockholm)
87
: 273–290 (October 2006)
Holmer and Caron
•
Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences
Quarry Member) and a single specimen was collected froma talus above the Walcott and Raymond Quarries and probablycomes from the Emerald Lake Oncolite Member (Fletcherand Collins 1998). Specimens were recovered in fine-grainedsiliciclastic beds showing preservation of non-biomineralizedorganisms. A bed assemblage (BA) represents the sum of allfossils present within the thickness of a single depositionalevent (see details in Caron 2005a). Note: the use of ‘BA’ inthis study is not to be confused with ‘benthic assemblage’
sensu
Boucot (1975), which refers to depth-zoned groups ofcommunities. Animals were buried catastrophically at thebase of the Cathedral Escarpment (Aitken and McIlreath1984) as a result of storm-generated back-currents (Aitkenand McIlreath 1990; Gostlin and Miall 2005) or more likelyby high-density fluidized mud-flows (Allison and Brett 1995).Taphonomic evidence shows that most benthic organismswere probably buried within their habitat (Caron 2005a;Caron and Jackson in press). Each bed is interpreted as asingle obrution deposit preserving a time-averaged and censusassemblage. The benthic community was probably livingin a relatively deep-water setting and below storm-wavebase and was probably influenced in part, by variation in thebottom oxygen water (Allison and Brett 1995; Powell
et al
.2003), and by other ecological and environmental factors,
such as the repeated disturbances caused by the burial eventsthemselves (Caron 2005a).
In the GPB, the community is mostly composed of infaunaland epifaunal organisms represented by deposit or filter-feeding organisms (Caron 2005a). Arthropods are usuallydominant and the brachiopods represent only a very smallfraction of the GPB community (2.8% in average relativeabundance of individuals out of 50 900 specimens in 26BAs). The new brachiopod is very rare in the GPB both interms of its number of occurrences (five out of 26 BAs) andin terms of the relative abundance of individuals per BA (max0.4%). Quantitative community analyses have shown thatBAs associated with this new brachiopod tend to have similarcommunity composition (Caron 2005a).
Materials and Methods
Line drawings and camera lucida drawings were producedwith both the part and counterpart when available. Pictureswere usually taken by applying polarizing filters to both thecamera and the light-source to increase contrast and reducethe amount of reflected light coming from some areas of thespecimens (i.e., glare). The sediment-coated parts of somefossils were prepared using an engraving tool along with a
Fig. 1—Location and stratigraphic origin of Acanthotretella spinosa n. sp. collected by the Royal Ontario Museum from the Middle Cambrian Greater Phyllopod Bed (Burgess Shale) in relation to the Cathedral Escarpment. The top of the ‘lower ginger layer’ represents the Royal Ontario Museum reference datum. Acanthotretella spinosa n. sp. occurs in the bed assemblages represented in black.

Middle Cambrian stem group brachiopod
•
Holmer and Caron Acta Zoologica
(Stockholm)
87
: 273–290 (October 2006)
© 2006 The Authors
Journal compilation © 2006 The Royal Swedish Academy of Sciences
reamer made of tungsten carbide. The posterior parts of thepedicles could generally not be cleared of the coated sedi-ment because the fossils were too thin to separate the rockcoat properly. Scanning electron microscope (SEM) photo-graphs of uncoated specimens were taken at the Universityof Uppsala using a Zeiss Supra 35 VP microscope. We useddodge and burn tools in
© 6.0 to lightenor darken areas of SEM images to enhance features that aredifficult to see because of the important variations ofexposure within the same image. Measurements (Appendix 1)were taken from live digital images using
softwareversion 1.2. Live digital images were produced using a NikonDS-5M five megapixels digital camera mounted on a NikonSMZ-1500 stereoscopic zoom microscope.
Systematic palaeontology
‘Stem group Brachiopoda’
Although the list of potential Early Cambrian stem groupbrachiopods is gradually expanding (Holmer 2001; Holmer
et al
. 2002, 2003, 2004; Williams and Holmer 2002; Peel2003; Skovsted and Holmer 2003; Balthasar 2004; Li andXiao 2004) the exact phylogenetic relationships of the taxawithin the stem lineages have yet to be worked out in moredetail; there have been no attempts to formally establish anynamed higher taxon and this practice is followed here.
Genus
Acanthotretella
nov.
Etymology.
From
akantha
(Gr.) – thorn,
tretos
(Gr.) – perforated,and the diminutive suffix -
ellus
(L), alluding to the small shellwith spinose exterior perforated by a pedicle foramen.
Diagnosis.
Shell essentially organic in composition, probablywith very minor organo-phosphatic mineral component,ventri-biconvex, inequivalved; exterior of both valves coveredby shell-penetrating, long, slender spine-like structures oforganic and partially mineralized composition, which aregenerally posteriorly inclined at an oblique angle away fromthe anterior margin; ventral valve apsacline, with slender,long, flexible and lingulid-like pedicle with a central coelomicregion, emerging through an external tube from a circularapical foramen, continued internally as a long internalpedicle tube; visceral area of both valves short, triangular,not extending to mid-valve, possibly with single umbonalscar; other interior features poorly known.
Type and only species. Acanthotretella spinosa
n. sp.
Species
Acanthotretella spinosa
n. sp.
Synonymy. Lingulella sp. – Jin et al. (1993), Fig. 3 : 6
Holotype. ROM 57749 – complete articulated shell, with theproximal part of the pedicle preserved (Figs 2D–F, 3B).
Type and localities. Fossil ridge between Wapta Mountainand Mount Field: Emerald Oncolite Shale Member andWalcott Quarry Shale Member (Greater Phyllopod bed),Burgess Shale Formation.
Other material. A total of 42 specimens. Illustrated specimensROM 56958, 57709, 57748–57757. Unillustrated specimensROM 57185, 57758–57775.
Etymology. Spinosus (L) – thorny, alluding to the spinoseexterior.
Diagnosis. As for Genus.
Description
Preservation. In stark contrast to the usual three-dimensionalpreservation of most Burgess Shale brachiopods which arebelieved to have been originally biomineralized (e.g., Diraphora,Nisusia see Briggs et al. 1994), A. spinosa is preserved likeother non-biomineralized animals from the Burgess Shale.The fossils appear darker than the surrounding matrix inlow-angle light and some areas are more reflective in high-angle light. This may suggest that the valves were originallymostly of organic composition; however, it is more likely thatthey were poorly mineralized and just had a minor organ-ophosphatic mineral component. This cannot be positivelydemonstrated, however, because Burgess Shale organismsappear to have been mainly replaced in clay minerals duringdiagenesis with some remnants of organic carbon altered tographite (Orr et al. 1998). The rare preservation of growthlines (e.g., Figs 2A, 3A) suggests that external features of theshell are mostly missing, in particular along the anteriormargins. Marginal areas are often very poorly preserved.Growth lines are more prominent towards the posteriorpart of the shells, suggesting that the shell was thicker towardsthe pedicle insertion (e.g., Fig. 4A,E). The valve interiorscommonly have somewhat thickened areas, corresponding
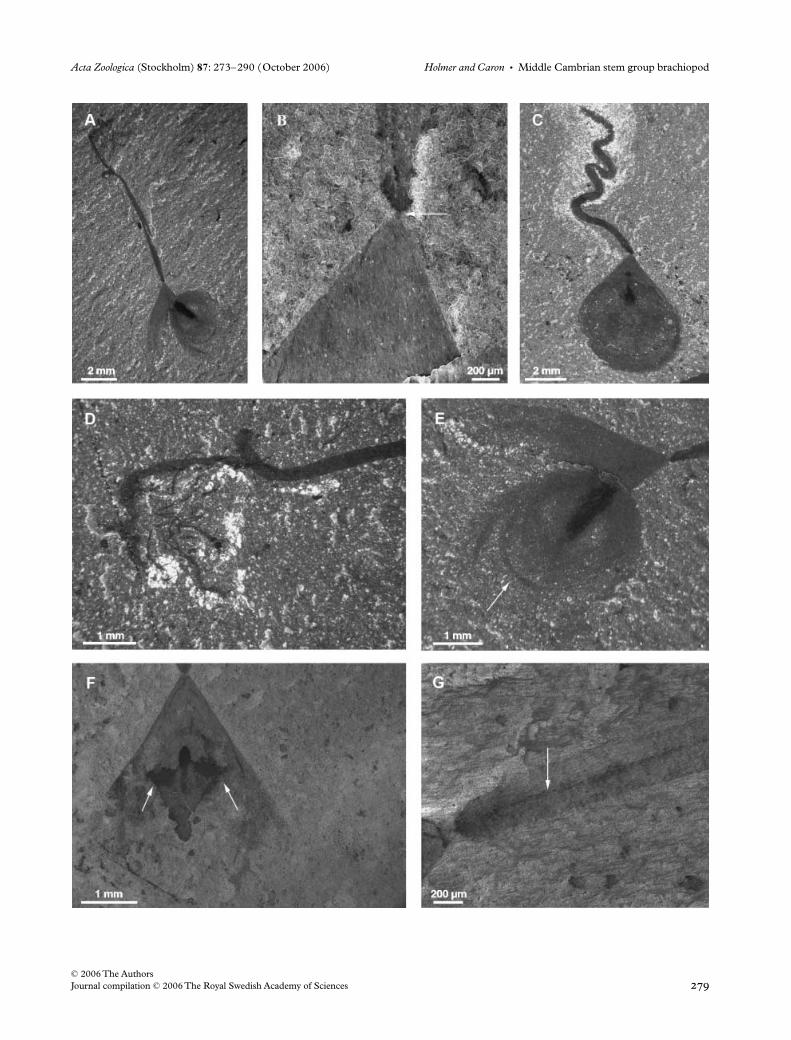
Fig. 2—Scanning electron microscope photographs of uncoated specimens of Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale); complete valves with the proximal part of the pedicle broken. —A–C. ROM 57748. —A. Overall view, arrows point to putative mantle canals. —B. Detail of spines, the arrow points to a spine in the central region of the valve. —C. Detail of the pseudointerarea region. —D–F. Holotype, ROM 57749. —D. Overall view. —E. Detail of the pseudointerarea region, the vertical arrow points to broken spines, the horizontal arrow point to a putative dorsal mesentery. —F. Detail of spines, arrows point to several posteriorly directed spines in the central region of the valve.

Acta Zoologica (Stockholm) 87: 273–290 (October 2006) Holmer and Caron • Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences

Middle Cambrian stem group brachiopod • Holmer and Caron Acta Zoologica (Stockholm) 87: 273–290 (October 2006)
© 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences
to the visceral areas, which are often filled with spherulesof framboidal pyrite, some 3 µm across (Fig. 5A,F). Thevisceral areas are usually much better preserved (with theexception of the lophophore) than the shell, which seems to berather a paradox. The better preservation of gut areas com-pared to the cuticle or carapace of many non-biomineralizedorganisms from the Burgess Shale is another example of asimilar phenomena (e.g., Conway Morris 1979; Caron 2005b)It is possible that muscles and digestive glands would havebeen stabilized and preserved first, possibly as the result ofhigher chemical reactivity (see also Butterfield 2002). Mostspecimens are typically preserved with the valves parallel tobedding planes (85%) suggesting that there was little over-turning of the specimens during burial and supporting theview that the valves were relatively flat.
Spines appear to have been brittle as suggested by anumber of broken spines along the margins of the valves(e.g., Figs 2E, 7E). Moreover, they often appear blacker thanthe rest of the shell and show little evidence of bending, sup-porting the view that they were presumably originally slightlymineralized.
The pedicle is present in most specimens (90%); it issometimes broken along its length, suggesting that it wasrelatively fragile and may have decayed quickly (Figs 5B, 7G).In contrast, the attachment zone of the pedicle to the shell,which is much narrower, must have been relatively strongbecause few pedicles are broken at this level (after the pedicle
tube, see below). Broken pedicles also suggest that theadherence of the pedicle bulb to elements of the epibenthoswas presumably strong (see below).
Pedicle. The total length of the pedicle is difficult to measurewith confidence because it does not generally appear to bepreserved in its full length or is too thin to be able to removethe rock-coating layer mechanically; however, in five pre-sumably complete specimens it varies between 20 and25 mm long, and between three and four times the total valvelength (Table 1). It invariably emerges through the ventralapical foramen through a narrow external pedicle tube, around0.1 µm wide and long (Figs 2C, 3A, 4B), which is thecontinuation of the internal pedicle tube (Figs 5A,C, 6B).The width enlarges abruptly directly above the tube, andthe remainder of the pedicle is generally of even thickness,averaging 0.4 mm in diameter, corresponding to just 8–10%of the total width of the valve (Table 1). In most specimensthere is evidence for a coelomic space, which is preservedas a thin, dark central line running throughout most of thepedicle on the SEM images and appearing more reflective inhigh-angle light (Figs 4B,G, 6A, 9D). The surface is commonlystrongly wrinkled throughout most of the length, with majorwrinkles at around 0.1–0.2 mm apart, and evidence of finerwrinkles in between, up to around 10 µm (Figs 5B,D,E, 6B).
In the majority of the specimens, the pedicle is tightlyfolded into S-shapes (Figs 4C, 5B, 6B, 7A, 8A) and in
Fig. 3—Line drawings of specimens of Acanthotretella spinosa n. sp. from Fig. 2. —A. ROM 57748. —B. ROM 57749.
Fig. 4—Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale); complete specimens with pedicle. —A, B, D, E. ROM 57750. —A. Overall view. —B. Detail of the proximal part of the pedicle (scanning electron micrograph), arrow points to the external pedicle tube. —D. Detail of the distal part of the pedicle attached to the putative algae Dictyophycus (visible as small threads). —E. Detail of anterior part showing dislocation of the valves, arrow points to growth lines. —C. Overall view, ROM 57751. —F, G. Scanning electron micrograph, ROM 57752. —F. Detail of the valves, arrows point to putative central muscle scars. —G. Detail of the pedicle, arrow points to the putative coelomic cavity.
Acta Zoologica (Stockholm) 87: 273–290 (October 2006) Holmer and Caron • Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences

Middle Cambrian stem group brachiopod • Holmer and Caron Acta Zoologica (Stockholm) 87: 273–290 (October 2006)
© 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences
Fig. 5—Scanning electron microscope photographs of an uncoated specimen of Acanthotretella spinosa n. sp. ROM 57753 from the Greater Phyllopod Bed (Burgess Shale). —A. Overall view of the posterior part of the shell and anterior part of the pedicle, arrow points to the visceral cavity (see F), the zone underlined by the black frame is enlarged in G. —B. Overall pedicle, presumably broken along its length. —C. Detail of the pseudointerarea, arrow points to the inferred pedicle tube. —D. Detail of the pedicle, the zone underlined by the black frame is enlarged in E. —E. Close up view of the pedicle from D. —F. Close up view of the visceral cavity showing framboidal pyrite from A. —G. Close up view of the posterior shell margin, from A, showing growth lines and spines.

Acta Zoologica (Stockholm) 87: 273–290 (October 2006) Holmer and Caron • Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences
some it has rotated across itself (Figs 4A,D, 6A). In a fewspecimens the distal end appears to be provided with asmall, poorly preserved terminal bulb (Figs 4D, 6A). Thereis some evidence that the terminal bulb could have beenused to adhere to living and dead elements of the epibenthos(see below).
Shell morphology and ornamentation. The dorsal and ventralvalves are invariably still conjoined, usually compressed
together with the exception of a few specimens in whichthe valves are slightly shifted (e.g., Figs 4A,E, 6A). The shellis ventri-biconvex and inequivalved, with the ventral valve onaverage 126% as long as the dorsal valve (Table 1, Fig. 10).The outline is tear-shaped with the maximum width in theanterior half of the shell. The maximum length of the shell is7.8 mm long, average 6.0 mm, and 139% as long as it is wide(Table 1). The conical ventral valve grows mixoperipherally,and has a wide and long, undivided, apsacline pseudointerarea,
Table 1 Different measurements and ratios of the brachiopod Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale)
Mean Std dev Min Max Count
LV 6.0 1.0 3.9 7.8 27Wt 4.4 0.8 2.7 5.8 27Ld 4.7 0.8 2.8 6.4 27Wi 2.3 0.5 1.2 3.2 34Li 1.3 0.3 0.8 1.8 34Lvi 2.6 0.4 1.6 3.3 34Lped 22.0 2.7 19.5 25.1 5Wped 0.4 0.1 0.3 0.5 25
Ratio Min Max Count
LV/Ld 126% 116% 141% 27LV/Wt 139% 115% 165% 27Ld/Wt 110% 93% 139% 27Wi/Wt 52% 41% 65% 27Li/LV 21% 15% 29% 27 LV Length ventral valveLvi/LV 42% 39% 50% 27 Wt Total widthLped/LV 347% 295% 437% 3 Ld Length dorsal valveLped/Wped 9% 8% 10% 9 Wi Pseudointerarea widthLi/Wi 57% 38% 93% 34 Li Pseudointerarea lengthLi/Ld 27% 17% 41% 27 Lvi Visceral length
Fig. 6—Line drawings of specimens of Acanthotretella spinosa n. sp. from Figs 4 and 5. —A. ROM 57750. —B. ROM 57753.

Middle Cambrian stem group brachiopod • Holmer and Caron Acta Zoologica (Stockholm) 87: 273–290 (October 2006)
© 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences

Acta Zoologica (Stockholm) 87: 273–290 (October 2006) Holmer and Caron • Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences
Fig. 8—Line drawings of specimens of Acanthotretella spinosa n. sp. from Fig. 7. —A. ROM 57709. —B. ROM 57754.
Fig. 9—Scanning electron microscope photographs of an uncoated specimen of Acanthotretella spinosa n. sp. ROM 57755 from the Greater Phyllopod Bed (Burgess Shale). —A. Overall view of the shell. —B. Detail of the pseudointerarea, arrow points to the inferred pedicle tube. —C. Detail of the spines. —D. Detail of the pedicle, arrow points to the putative coelomic cavity.
Fig. 7—Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale). All photographs are scanning electron microscope photographs, except A, of uncoated specimens. —A–F. Complete individual with pedicle, ROM 57709. —A. Overall view. —B. Detail of the anterior part of the pedicle. —C. Detail of the posterior part of the pedicle. —D. Detail of the posterior part of the shell. —E. Close up view of D, arrows point to broken spines. —F. Close up view of spines along the anterior margin of the shell. —G. ROM 57754, specimen with broken pedicle, arrows point to the putative lophophore.

Middle Cambrian stem group brachiopod • Holmer and Caron Acta Zoologica (Stockholm) 87: 273–290 (October 2006)
© 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences
on average 57% as long as it is wide, and occupying some-what more than half the valve width and around a quarter ofthe valve length (Table 1). The dorsal valve is flattened onlysomewhat conical, on average 110% as long as wide, with amarginal apex (Figs 4E, 6A). The dorsal pseudointerarea ispoorly preserved, but appears to have a median groove(Figs 7G, 8B).
The ontogeny is unknown and the smallest recorded shellis close to 4 mm long. The width of the ventral valve, however,grows proportionally with the length (Appendix 1). Apartfrom the spine-like structures, the exterior of both valves issmooth with faint concentric growth lines (Figs 2A, 3A,4A,E, 5A,G, 6A,B).
Spines. The exterior of both valves is covered by long (up to3 mm long), slender (between < 10 µm and 40 µm thick),shell-penetrating, spine-like structures of assumed organicand partially mineralized composition (Figs 2B,F, 3, 7F, 8A,9A,C, 11A,C,D,F,G); the spine-like structures are notpreserved as often as compared with the pedicle (present in65% of specimens). Most frequently they are preservedalong the margin of the anterior half of the valves, and theyare only occasionally preserved in the posterior half andin the central sections of the shell (Figs 2F, 3B). This is pro-bably because the anterior and central parts of many valvesare poorly preserved as a result of compaction.
Many spines are gently curved, whereas others arecompletely straight (Figs 2B,F, 3 and 11D,F). The spines
are generally posteriorly inclined at an oblique angle awayfrom the anterior margin, and some of the curved spines areorientated almost parallel to the curvature of the shell margin(Figs 2F and 3B). The density of spines varies betweenspecimens, but a 1-mm long section along the shell curvaturehas up to 10 spines.
In the few specimens where evidence for exterior growthlines can be seen, there are also traces of shorter spine-likeobjects, up to 240 µm long and 20 µm thick, which may bethe remains of a second order of spines (Fig. 5A,G).
Interior characters. The interior characters of A. spinosa areextremely poorly preserved. The ventral interior is providedwith a long internal pedicle tube (up to 45 µm in diameterand more than 0.5 mm long in a 4-mm wide valve), andextending from the apical pedicle foramen (Figs 5C, 6B).The visceral area of both valves is short, triangular, notextending to mid-valve, but only extending for on average42% of the total valve length, but on average just 27% of thedorsal valve length (Table 1). It is completely divided by apossible dorsal mesentery in some specimens (Figs 2E, 3B).Individual muscle scars cannot be distinguished with theexception of an undivided umbonal scar generally placed inclose proximity to the anterior end of the pedicle tube andthe dorsal pseudointerarea (Figs 2C,E, 3, 5A,C, 6B, 7A,G,8). A single shell has the possible imprints of a double centralmuscle (Fig. 4F).
One specimen bears faint impressions of a pair of circularareas, which may represent a lophophore, directly in front ofthe visceral cavity (Figs 7G, 8B). A second specimen showsfaint impressions of two diverging possible mantle canalsclose to the anterior margin as well as a single lateral mantlecanal (Figs 2A, 3A).
Comparative discussion and interpretation
Pedicle
The pedicle of A. spinosa differs from those of all otherknown pedunculate Cambrian brachiopods, which as notedabove, are all from the Lower Cambrian Chengjiang Fauna.The pedicle emerges through a circular apical foramen at theend of a narrow external tube. This external tube representsan outgrowth of the ventral valve and is continuous andparallel to the internal pedicle tube. The pedicle attains itsfull width less than 1 mm after the foramen. An internal pedicletube is also present in the lingulellotretid L. malongensis, butin this species the pedicle emerges from an oval foramen onthe linguloid pseudointerarea (Holmer et al. 1997; Holmerand Popov 2000) and the proximal part of the pedicle encloses
Fig. 10—Reconstruction of the shell of Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale) in lateral and dorsal views (drawing by Hannah Caine). The dorsal view is drawn so as to show internal features: the internal pedicle tube, the umbonal scar and the visceral cavity (from top to bottom).
Fig. 11—Scanning electron microscope photographs of an uncoated specimen of Acanthotretella spinosa n. sp. ROM 57756 from the Greater Phyllopod Bed (Burgess Shale). —A, B, D, F. Part. —C, E, G. Counterpart. —A. Overall view of the shell, arrow points to a spine orientated posteriorly. —B. Detail of the pseudointerarea, arrow points to putative growth lines. —C. Overall view of the shell. —D. Detail of the spines. —E. Detail of the pseudointerarea. —F. Close up view of the spines. —G. Detail of the anterior part of the shell.

Acta Zoologica (Stockholm) 87: 273–290 (October 2006) Holmer and Caron • Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences

Middle Cambrian stem group brachiopod • Holmer and Caron Acta Zoologica (Stockholm) 87: 273–290 (October 2006)
© 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences
the apex (Jin et al. 1993). Apart from A. spinosa, the only otherlinguliforms that are provided with internal and externalpedicle tubes as well as an apical foramen are the sipho-notretides and the acrotretides (e.g., Holmer and Popov2000), but unfortunately the pedicle morphology in neithergroup is known from the actual preservation of the pediclesthemselves.
The pedicle of A. spinosa also differs from that ofL. malongensis in that it is narrower and only up to 0.5 mmin diameter, as compared with 0.8–1.2 mm in diameter inthe latter. The recorded relative pedicle length/total valvelength ratios in A. spinosa, however, are within the largerange of variation recorded both from fossil and Recentlinguliform brachiopods (e.g., Jin et al. 1993), and boththe Canadian and the two Chinese species essentially haveevenly thick pedicles, whereas the pedicle of ‘Lingulella’chengjiangensis has a long tapering distal part (Jin et al. 1993).The surface wrinkles and evidence of central coelomicspaces are similarly developed in the two Chinese speciesand A. spinosa, but the terminal ends of L. malongensis and‘Lingulella’ chengjiangensis lack wrinkles, whereas in A. spinosathe poorly preserved terminal end seems to be providedwith a shorter terminal bulb. A bulb-like termination is alsopresent in L. malongensis (Zhang et al. 2005).
The pedicle morphology of the problematic brachiopodX. haikouensis (Zhang et al. 2006) is also quite similar tothat of A. spinosa, L. malongensis, as well as of ‘Lingulella’chengjiangensis; it shows evidence of a central coelomic spaceand exterior wrinkles, but the pedicle is much thickerand somewhat shorter compared to the pedicles in theother three species. Unlike A. spinosa, however, the pedicleof X. haikouensis apparently emerges between the valvesand its proximal part encloses the entire apex (Zhanget al. 2006).
The pedicle of both the problematic Longtancunellachengjiangensis as well as the botsfordiid Diandongia pistaapparently lack evidence of a central coelomic space; theexterior of the pedicle of Longtancunella chengjiangensis iswrinkled, while the pedicle exterior of D. pista is smooth(Zhang Z.F. et al. 2003; Zhang et al. in press).
The tightly folded S-shapes and rotations of the pedicle ofA. spinosa have not been commonly recorded in other LowerCambrian pedunculate linguliforms, which may form moregentle S-shaped curves (Jin et al. 1993; Zhang et al. 2005) orelse are most commonly completely straight (Zhang Z.F.et al. 2003; Zhang et al. 2006, in press).
Shell morphology. The combination of morphologicalcharacters in A. spinosa is not found in any other linguliform.The ventribiconvex shells of A. spinosa with its conicalapsacline ventral valves and dorsal and ventral pseudointer-areas are most similar to those found within the OrderSiphonotretida; however, all known siphonotretides havewell-mineralized stratiform organophosphatic shells (e.g.,Holmer and Popov 2000; Williams et al. 2004), while the
shell of A. spinosa was entirely organic in composition ormore likely had just a minor mineral component.
Spines. Apart from the pedicle, the most striking distinguishingfeatures of A. spinosa are the spines. They clearly differ fromthe normal marginal follicular mantle setae that are foundin living brachiopods in that they also cover the surface of theshell; thus, they cannot be compared directly with most otherknown exceptionally preserved marginal setae in LowerCambrian linguliforms.
In L. malongensis and ‘Lingulella’ chengjiangensis as wellas in Longtancunella chengjiangensis, the preserved marginalsetae are thin, short, delicate structures that emerge atright angles to the shell margin (Zhang et al. 2005, in press),whereas those of the Burgess paterinid Micromitra burgessensisare considerably longer (e.g., Walcott 1912, 1924; Whittington1985; Briggs et al. 1994). The only previously recorded,exceptionally preserved ‘setae’ that can be directly comparedwith those of A. spinosa have been described for Heliomedusaorienta (as re-interpreted by Holmer et al. (2003; see alsoZhang et al. 2003), and the closely related Mickwitzia (Holmeret al. 2004)); both species are provided with exceptionallypreserved setae that emerge from setal tubes all over the shell.However, there is no evidence that the spine-like structuresin A. spinosa emerged from tubular structures, and thus thereis no direct evidence to support homology with the ‘setae’ ofHeliomedusa and Mickwitzia.
The exceptionally preserved ‘setae’ of X. haikouensis appearto be more spine-like and comparable to those of A. spinosa;however, they are thicker at the base and tapering and possiblyhollow (Zhang et al. 2006). Moreover they were evidentlyrigid enough to support attached tubular epifauna (Zhangand Han 2004).
The main orientation of the spines in A. spinosa differsfrom all the previous records discussed above in that they aregenerally posteriorly inclined at an oblique angle away fromthe anterior margin; in general structure they are mostsimilar to the thorn-like organic objects that have beeninferred from early siphonotretides (Williams et al. 2004). Inspecies belonging to the siphonotretide Helmersenia, normalorganophosphatic spines are lacking and the shells areperforated by numerous canals that terminate with anexternal impression at the valve surface; the depressionmust have been caused by an organic, degraded, possiblychitinous, structure (Williams et al. 2004). The exact shapeof these chitinous structures is not known, but it is quitelikely that they were spine-like, similar to those recorded herefrom A. spinosa. Williams et al. (2004) proposed that thesestructures (as well as the regular siphonotretide spines)were secreted by specialized cells (so-called acanthoblasts)within the mantle, and this interpretation is here alsoextended to explain the formation of the spines in A. spinosa.The shell around the spines is not preserved in A. spinosa,however, and it is impossible to make a further detailedcomparison.

Acta Zoologica (Stockholm) 87: 273–290 (October 2006) Holmer and Caron • Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences
Visceral cavity and shell musculature. Only the general outlineof the visceral cavity and the possible imprint of an undividedumbonal muscle can be discerned on most specimens ofA. spinosa. The short, triangular visceral cavity is quite unlikethat of any Lower Cambrian linguloid brachiopod, whichtypically has an elongated shape, with a dorsal medianprojection (Holmer and Popov 2000). Moreover, the linguloidumbonal muscle scars are generally double, for exampleL. malongensis (Holmer et al. 1997). In shape and outline, thevisceral cavity is most similar to that of X. haikouensis andthe umbonal muscle scar in this species is undivided as inA. spinosa.
Autecology. The long and slender pedicle and the shape of theshell suggest that this species was not adapted to burrowing.Rather, the presence of the terminal bulb attached to theputative macrobenthic algae Dictyophycus and other epibenthosprovide evidence that this animal was living above the sediment–water interface (Fig. 12 and see Fig. 13 palaeoenvironmentalreconstruction). The pedicle was probably able to maintainthe shells in an upright position, possibly reaching relativelyhigh strata, in particular because the shell would have beenconsiderably lighter compared to the biomineralized brachiopodspecies that are commonly found in the Burgess Shale.Higher tiering level, however, is also reached in somebiomineralized brachiopods, albeit in a different way. For
example, Micromitra is commonly attached to spicules of thesponge Pirania, but the pedicle of this species was probablyvery short (Williams et al. 1998). In both cases, higher stratawould have been necessary, possibly to protect the lophophorefrom being clogged with mud. Such an adaptation providesan indirect indication of the original degree of firmness of thesubstrate. It is highly possible that at least some portionsof the substrate were composed of relatively soft sedimentsusceptible to easy remobilization by weak currents, reducingthe ability of many brachiopods to settle.
The finding of a cluster of six specimens of A. spinosa onan unidentified organic structure (Fig. 12A,B) supports theview that this species was presumably highly dependenton anchoring points to survive and was gregarious. Thereare some indications, however, that parts of the substratewere also covered by sheet-like colonies of the cyanobacte-rium Morania that supported a grazing community (Caronet al. in press). Such a substrate would have been firmer andpresumably more favourable for A. spinosa to colonize.Interestingly Morania is usually abundant in layers withA. spinosa.
The function of the spines in A. spinosa is more difficult tointerpret. They may represent an anti-predatory mechanismbecause the valves were not biomineralized. It is also possiblethat collectively they could be used as a support for the shellsto lay flat on the substrate.
Fig. 12—Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale). —A, B. ROM 57757. —A. Overall view showing a cluster of six specimens, of which three show pedicles, preserved on an unidentified organic structure. —B. Camera lucida drawings of A. —C, D. ROM 56958. —C. Overall view, the black frame emphasizes the position of one specimen of A. spinosa n. sp. with its pedicle looping under one antenna of a Sidneyia carcass. —D. Close up view of the black frame area from C.

Middle Cambrian stem group brachiopod • Holmer and Caron Acta Zoologica (Stockholm) 87: 273–290 (October 2006)
© 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences
Phylogenetic position and affinities
The detailed morphology and anatomy of A. spinosa is stilltoo poorly known to evaluate its phylogenetic position usingcladistic analysis. The combination of known characters inA. spinosa is unlike that of any previously described brachiopod,but many of the features related to the shell morphologyand the spines indicate that it has some affinity to the OrderSiphonotretida.
As defined by Holmer and Popov (2000), the sipho-notretides belong within the Subphylum Linguliformea,and Class Lingulata where they constitute a small, relativelyshort-lived order (comprising 15 genera), appearing near the endof the Middle Cambrian, and with most forms becomingextinct near the end of the Late Ordovician. Williams et al. (2004)noted that the siphonotretides are very different from allother lingulates, and that they possibly do not belong within thelinguliforms. If this assumption is correct, Acanthotretella maybelong within a separate stem group clade together with the
siphonotretides; this, as yet hypothetical, clade would bedefined by the acquisition of the siphonotretide acanthoblasts.
Acknowledgements
J.B.C. acknowledges the support of a Natural Sciences andEngineering Research Council of Canada postdoctoralfellowship. This is Royal Ontario Museum Burgess ShaleResearch Project number 2. We thank Parks Canada forgranting access to Royal Ontario Museum field crews (ledby Desmond Collins) to collect specimens from the BurgessShale. Grants from the Swedish Research Council (VR)supported Holmer’s work.
References
Aitken, J. D. and McIlreath, I. A. 1984. The Cathedral ReefEscarpment; A Cambrian great wall with humble origins. – Geos13: 17–19.
Fig. 13—Habitat reconstruction of Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale). The pedicle is attached to different types of epibenthos (e.g. Dictyophycus, carcasses of animals such as Sidneyia) and was probably able to raise the shell above the water–sediment interface. The biomineralized brachiopod Micromitra is commonly found in bed assemblages with A. spinosa n. sp. attached to the sponge Pirania and showing preservation of setae (drawing by Hannah Caine).

Acta Zoologica (Stockholm) 87: 273–290 (October 2006) Holmer and Caron • Middle Cambrian stem group brachiopod
© 2006 The AuthorsJournal compilation © 2006 The Royal Swedish Academy of Sciences
Aitken, J. D. and McIlreath, I. A. 1990. Comment (reply ofLudvigsen, R 1989; The Burgess shale: not in the shadow ofthe Cathedral Escarpment: Geoscience Canada, 16: 51–59). –Geoscience Canada 17: 111–116.
Allison, P. A. and Brett, C. E. 1995. In situ benthos and paleo-oxygenation in the Middle Cambrian Burgess Shale, BritishColumbia, Canada. – Geology 23: 1079–1082.
Balthasar, U. 2004. Shell structure, ontogeny and affinities of theLower Cambrian bivalved problematic fossil Mickwitzia muralensisWalcott, 1913. – Lethaia 37: 381–400.
Boucot, A. J. 1975. Evolution and Extinction Rate Controls. Elsevier,New York.
Briggs, D. E. G., Erwin, D. H. and Collier, F. J. 1994. The Fossils ofthe Burgess Shale. − Smithsonian Institution Press, Washington.
Butterfield, N. J. 2002. Leanchoilia guts and the interpretation ofthree-dimensional structures in Burgess Shale-type fossils. –Paleobiology 28: 155–171.
Caron, J. B. 2005a. Taphonomy and Community Analysis of the MiddleCambrian Greater Phyllopod Bed, Burgess Shale. PhD Thesis.University of Toronto, Toronto.
Caron, J. B. 2005b. Banffia constricta, a putative vetulicolid from theMiddle Cambrian Burgess Shale. – Transactions of the Royal Societyof Edinburgh 96: 95–111.
Caron, J. B., Jackson, D. A. in press. Taphonomy of the GreaterPhyllopod Bed Community, Burgess Shale. Palaios 21.
Caron, J. B., Scheltema, A. H., Schander, C. and Rudkin, D. 2006.A soft-bodied mollusc with radula from the Middle CambrianBurgess Shale. – Nature 442: 159–163.
Conway Morris, S. 1979. Middle Cambrian polychaetes from theBurgess Shale of British Columbia. – Philosophical Transactions ofthe Royal Society of London, Series B 285: 227–274.
Fletcher, T. P. and Collins, D. 1998. The Middle Cambrian BurgessShale and its relationship to the Stephen Formation in the southernCanadian Rocky Mountains. – Canadian Journal of Earth Sciences35: 413–436.
Gostlin, K. E. and Miall, A. D. 2005. Sedimentology of the BurgessShale: Refining the Conditions for Soft-Bodied Fossil Preservation.Geological Society of America, Annual meeting, Salt Lake City,37: 547. Geological Society of America, Boulder, Colorado.
Holmer, L. E. 2001. Phylogeny and classification: Linguliformeaand Craniiformea. In Carlson, S. J., Sandy, M. R. (Eds): BrachiopodsAncient and Modern: A Tribute to G. Arthur Cooper, pp. 11–26. ThePaleontological Society Papers, Pittsburgh, PA.
Holmer, L. E. and Popov, L. E. 2000. Class Lingulata. In Kaesler, R. L.(Ed.): Treatise on Invertebrate Paleontology. Part H. Brachiopoda(Revised), pp. 30–146. Geological Society of America and Univer-sity of Kansas Press, Lawrence, KS.
Holmer, L. E., Popov, L. E., Koneva, S. P. and Jia-Yu, R. 1997.Early Cambrian Lingulellotreta (Lingulata, Brachiopoda) fromSouth Kazakhstan (Malyi Karatau range) and South China(Eastern Yunnan). – Journal of Paleontology 71: 577–584.
Holmer, L. E., Skovsted, C. B. and Williams, A. 2002. A stem groupbrachiopod from the Lower Cambrian – support for a Micrina(halkieriid) ancestry. – Palaeontology 45: 875–882.
Holmer, L. E., Li, G. X. and Maoyan, Z. 2003. The systematicposition of the Lower Cambrian brachiopod Heliomedusa Sun andHou. In Donoghue, P. C. J. (Ed.): Palaeontological Association 47thAnnual Meeting, p. 139. The Palaeontological Association,University of Leicester, Leicester.
Holmer, L. E., Streng, M., Skovsted, C. B. and Babcock, L. E.2004. Exceptional preservation in the stem group brachiopodMickwitzia from the Early Cambrian of Nevada. – GFF 126:112.
Hou, X., Bergström, J., Wang, H., Feng, X. and Chen, A. 1999. TheChengjiang Fauna Exceptionally Well-Preserved Animals from 530Million Years Ago. Yunnan Science and Technology Press,Kunming.
Hou, X. G., Aldridge, R. J., Bergström, J., Siveter, D. J., Siveter, D.J. and Feng, X. H. 2004. The Cambrian Fossils of Chengjiang,China: the Flowering of Early Animal Life. Blackwell Publishing,Oxford.
Jin, Y. G. 2005. Life styles of early Cambrian brachiopods on mudsubstrate. In Peng, S., Zhu, M. Y., Li, G. X., Van Iten, H. (Eds):The Fourth International Symposium on the Cambrian System and theTenth International Conference of the Cambrian Stage SubdivisionWorking Group, p. 77. Acta Micropalaeontologica Sinica, Nanjing.
Jin, Y. and Wang, H. 1992. Revision of the Lower Cambrian brachiopodHeliomedusa Sun & Hou, 1987. – Lethaia 25: 35–49.
Jin, Y. G., Hou, X. G. and Wang, H. Y. 1993. Lower Cambrianpediculate lingulids from Yunnan, China. – Journal of Paleontology67: 788–798.
Li, G. and Xiao, S. 2004. Tannuolina and Micrina (Tannuolinidae)from the Lower Cambrian of Eastern Yunnan, South China, and theirscleritome reconstruction. – Journal of Paleontology 78: 900–913.
Orr, P. J., Briggs, D. E. G. and Kearns, S. L. 1998. CambrianBurgess Shale animals replicated in clay minerals. – Science 281:1173–1175.
Peel, J. S. 2003. A problematic cap-shaped metazoan from theLower Cambrian of Estonia. – GFF 125: 157–161.
Powell, W. G., Johnston, P. A. and Collom, C. J. 2003. Geochemicalevidence for oxygenated bottom waters during deposition offossiliferous strata of the Burgess Shale Formation. – Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 201: 249–268.
Resser, C. E. 1938. Fourth contribution to nomenclature of Cambrianfossils. – Smithsonian Miscellaneous Collections 97: 1–43.
Skovsted, C. B. and Holmer, L. E. 2003. The Early Cambrian(Botomian) stem group brachiopod Mickwitzia from NortheastGreenland. – Acta Palaeontologica Polonica 48: 1–20.
Walcott, C. D. 1912. Cambrian Brachiopoda. – United States GeologicalSurvey Monograph 51: − Part 1: 1–872, Part 872: 871–363.
Walcott, C. D. 1924. Cambrian and Ozarkian Brachiopoda.Cambrian geology and paleontology IV. – Smithsonian MiscellaneousCollections 67: 477–554.
Whittington, H. B. 1985. The Burgess Shale. Yale University Press,New Haven.
Williams, A. and Holmer, L. E. 1992. Ornamentation and shellstructure of acrotretoid brachiopods. – Palaeontology 35: 657–692.
Williams, A. and Holmer, L. E. 2002. Shell structure and inferredgrowth, functions and affinities of the sclerites of the problematicMicrina. – Palaeontology 45: 845–873.
Williams, A., Holmer, L. E. and Cusack, M. 2004. Chemico-structureof the organophosphatic shells of siphonotretide brachiopods. –Palaeontology 47: 1313–1337.
Williams, A., Popov, L. E., Holmer, L. E. and Cusack, M. 1998.The diversity and phylogeny of the paterinate brachiopods. –Palaeontology 41: 221–262.
Zhang, X. G., Hou, X. G. and Emig, C. C. 2003. Evidence oflophophore diversity in Early Cambrian brachiopoda. – Proceedingsof the Royal Society of London, Series B: Biological Sciences 270:S65–S68.
Zhang, Z. F. and Han, J. 2004. A new lingulid brachiopod from theChengjiang Lagerstätte. – Journal of Northwest University (NaturalScience Edition) 34: 449–453.
Zhang, Z. F., Han, J., Zhang, X. L., Liu, J. and Shu, D. G. 2003.Pediculate brachiopod Diandongia pista from the Lower Cambrianof South China. – Acta Geologica Sinica 77: 288–293.

Middle Cambrian stem group brachiopod • Holmer and Caron Acta Zoologica (Stockholm) 87: 273–290 (October 2006)
© 2006 The Authors Journal compilation © 2006 The Royal Swedish Academy of Sciences
Zhang, Z. F., Han, J., Zhang, X. G., Liu, J. and Shu, D. 2004a.Soft-tissue preservation in the Lower Cambrian linguloidbrachiopod from South China. – Acta Palaeontologica Polonica 49:259–266.
Zhang, Z. F., Shu, D., Han, J. and Liu, J. N. 2004b. New data onthe lophophore of Early Cambrian linguloids from the ChengjiangLagerstätte, Southwest China. – Carnets de Géologie Letter 2004/04 (CG2004-L04)-online.
Zhang, Z F., Shu, D., Han, J. and Liu, J. 2005. Morpho-anatomicaldifferences of the Early Cambrian Chengjiang and Recent lingulidsand their implications. – Acta Zoologica 86: 277–288.
Zhang, Z F., Shu, D., Han, J. and Liu, J. 2006. New data on therare Chengjiang (Lower Cambrian, South China) linguloidbrachiopod Xianshanella haikouensis. – Journal of Paleontology 80:203–211.
Zhang, Z. F., Shu, D., Han, J. and Liu, J. N. in press. A gregariouslingulid brachiopod Longtancunella chengjiangensis from the LowerCambrian, South China. – Lethaia.
Appendix 1. Length (LV) − width (Wt) relationship of the brachiopod Acanthotretella spinosa n. sp. from the Greater Phyllopod Bed (Burgess Shale).




















