
A Pleistocene warming event at 1 Ma in Prydz Bay, East Antarctica: Evidence from ODP Site 1165
15
A Pleistocene warming event at 1 Ma in Prydz Bay, East Antarctica: Evidence from ODPSite 1165 G. Villa a , , C. Lupi b , M. Cobianchi b , F . Florindo c , S.F . Pekar d, e a Dipartimentodi Scienze della T erra, Università di Parma, Parco Area delle Scienze, 157A, 43100 Parma, Italy b Dipartimentodi Scienze della T erra, Università di Pavia, Via Ferrata 1, 27100 Pavia, Italy c Istituto Nazionale di Geofisica e Vulcanologia, Via Vigna Murata 605, 00143 Roma, Italy d Queens College, School of Earth and Environmental Sciences, 65-30 Kissena Boulevard, Flushing, New York 11367, USA e Lamont-Doherty Earth Observatory of Columbia University, Palisades, New York 10964, USA Received 26 April 2007; accepted 22 August 2007 Abstract Bio- and magnetostratigraphicage dataand nannofossil assemblageanalysis from ODPSite 1165 evidence an anomalous warming event of the surface waters in and around Prydz Bay during the Early Pleistocene, approximately1 Ma. This results from an increase in theabundance of nannofossils at Site 1165, that occurred at 1 Ma. Detailedhigh-resolution sampling permits a new bio-magnetostratigraphic interpretation for ODPSite 1165. A decrease in δ 18 O values at Sites 1165 and 1167 alsooccurs atthis time, supporting the presence of warming conditions in the Prydz Bay area. A return to colder surface waters, indicated by the absence orrare occurrence of nannofossils in the upper cores from Site 1165, suggests that more stable glacial conditions existed in the Prydz Baybasin, for the last 900 ka. The biogenic carbonate sequence identified at Site 1167 issimilar to the carbonate shales recovered from the Cape Roberts Project 1. Bothhave beendated at about 1 Ma, supporting the idea that a significant surface waters warming occurred during the Pleistocene. These dataand the presence of calcareous nannofossils from locations around the Antarcticcontinent also suggestthat the warming event was not limited to theanalysedbasin, but it extended around the East Antarcticcontinent. These new evidence call for a re-evaluation of the notion thatthe East Antarctic Ice-Sheet has experienced stableconditionssimilar to today since the late Neogene. © 2007 Elsevier B.V . All rights reserved. Keywords: Calcareous nannofossils; Pleistocene; Antarctica; Palaeoclimate; Prydz Bay; Ocean Drilling Program 1. Introduction The Antarctic Ice-Sheet and the surrounding South- ern Ocean are key components of global climate regime and distribution of biota, from the early Cenozoic to the present day. The Antarcticcontinentis presumed tohave been under stable polar conditions for many millions of years (Kennett and Barker, 1990; Barrett, 2003). Re- cently, a debate over stable versusmore dynamic An- tarcticclimate has arisenbased on evidence of partial collapses of the Western Antarctic Ice-Sheet (W AIS) (Scherer, 1991; Scherer et al., 1998, 2002), as well as on the temporal stabilityof the Eastern Antarctic Ice-Sheet Available online at www.sciencedirect.com Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230 – 244 www.elsevier .com/locate/palaeo Corresponding author . E-mail address: giuliana.villa@unipr .it (G. Villa). 0031-0182/$ - see front matter © 2007 Elsevier B.V . All rights reserved. doi:10.1016/j.palaeo.2007.08.017
Transcript of A Pleistocene warming event at 1 Ma in Prydz Bay, East Antarctica: Evidence from ODP Site 1165

A Pleistocene warming event at 1 Ma in Prydz Bay, East Antarctica:Evidence from ODP Site 1165
G. Villa a, , C. Lupi b, M. Cobianchi b, F. Florindo c, S.F. Pekar d,e
a Dipartimento di Scienze della Terra, Università di Parma, Parco Area delle Scienze, 157A, 43100 Parma, Italyb Dipartimento di Scienze della Terra, Università di Pavia, Via Ferrata 1, 27100 Pavia, Italyc Istituto Nazionale di Geofisica e Vulcanologia, Via Vigna Murata 605, 00143 Roma, Italy
d Queens College, School of Earth and Environmental Sciences, 65-30 Kissena Boulevard, Flushing, New York 11367, USAe Lamont-Doherty Earth Observatory of Columbia University, Palisades, New York 10964, USA
Received 26 April 2007; accepted 22 August 2007
Abstract
Bio- and magnetostratigraphic age data and nannofossil assemblage analysis from ODP Site 1165 evidence an anomalouswarming event of the surface waters in and around Prydz Bay during the Early Pleistocene, approximately 1 Ma. This results froman increase in the abundance of nannofossils at Site 1165, that occurred at 1 Ma. Detailed high-resolution sampling permits a newbio-magnetostratigraphic interpretation for ODP Site 1165. A decrease in !18O values at Sites 1165 and 1167 also occurs at thistime, supporting the presence of warming conditions in the Prydz Bay area. A return to colder surface waters, indicated by theabsence or rare occurrence of nannofossils in the upper cores from Site 1165, suggests that more stable glacial conditions existed inthe Prydz Bay basin, for the last 900 ka.
The biogenic carbonate sequence identified at Site 1167 is similar to the carbonate shales recovered from the Cape RobertsProject 1. Both have been dated at about 1 Ma, supporting the idea that a significant surface waters warming occurred during thePleistocene. These data and the presence of calcareous nannofossils from locations around the Antarctic continent also suggest thatthe warming event was not limited to the analysed basin, but it extended around the East Antarctic continent. These new evidencecall for a re-evaluation of the notion that the East Antarctic Ice-Sheet has experienced stable conditions similar to today since thelate Neogene.© 2007 Elsevier B.V. All rights reserved.
Keywords: Calcareous nannofossils; Pleistocene; Antarctica; Palaeoclimate; Prydz Bay; Ocean Drilling Program
1. Introduction
The Antarctic Ice-Sheet and the surrounding South-ern Ocean are key components of global climate regimeand distribution of biota, from the early Cenozoic to the
present day. The Antarctic continent is presumed to havebeen under stable polar conditions for many millions ofyears (Kennett and Barker, 1990; Barrett, 2003). Re-cently, a debate over stable versus more dynamic An-tarctic climate has arisen based on evidence of partialcollapses of the Western Antarctic Ice-Sheet (WAIS)(Scherer, 1991; Scherer et al., 1998, 2002), as well as onthe temporal stability of the Eastern Antarctic Ice-Sheet
Available online at www.sciencedirect.com
Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230 – 244www.elsevier.com/locate/palaeo
Corresponding author.E-mail address: [email protected] (G. Villa).
0031-0182/$ - see front matter © 2007 Elsevier B.V. All rights reserved.doi:10.1016/j.palaeo.2007.08.017

(EAIS). The stable glacial state hypothesis suggests thatthe EAIS became stable in the middle Miocene and haschanged little since (e.g. Huybrechts, 1993; Sugdenet al., 1993; Kennett and Hodell, 1995; Lear et al., 2000;Zachos et al., 2001). On the other hand, other evidence(e.g. Webb et al., 1984; Hambrey and McKelvey, 2000)suggests that the EAIS was subject to major fluctuationsup to Pliocene time implying considerable climate var-iation extending from the late Miocene through thePliocene–Pleistocene (Escutia et al., 2003).
One means to test the idea of a more dynamic climatein the high southern latitudes is the study of calcareousmicrofossils, such as foraminifers and nannofossils.However, a few studies examine the distribution andabundance of calcareous nannofossils of the Antarcticregion at latitude south of 65°S for the younger part ofthe geological record. One of the reasons is that todaythe coccolithophorid distribution is confined at latitudes
lower than the Antarctic Divergence (at about latitude65°S) (Findlay, 1998; Findlay and Giraudeau, 2000).Recent studies have reported calcareous nannofossils insediment drifts off the Antarctic Peninsula (Villa et al.,2003), and from several piston cores recovered in theWeddell Sea, Ross Sea, Maud Rise, Bellingshausen Sea(Villa et al., 2005). Though discontinuous and rare, theoccurrence of calcareous nannofossils in Pleistocenedeposits near the Antarctic margin may indicate that,differently from present-day conditions, the environ-ment was at times favourable to nannoplanktonproductivity. The environmental conditions that causedtheir paucity are generally associated with the thermalisolation of the Southern Ocean, low sea surface-watertemperatures, sea-ice extension, nutrient availability,and sea-water bottom conditions that influence nanno-fossil preservation in sediments (Villa et al., 2003,2005).
Fig. 1. Location of Site 1165 and the main oceanographic features of the area. The site was cored, on the Eastern Antarctica Rise, close to theAntarctic Divergence, off of Prydz Bay, at 64°22 S and 67°13 E. A star indicates the position of Prydz Bay with respect to the Antarctic continent(modified from Cooper and O'Brien, 2004).
231G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

Ocean Drilling Program (ODP) Leg 188 (O'Brienet al., 2001) was drilled to obtain a record of Antarcticpalaeoenvironmental changes from the Miocene toPleistocene. Site 1165 was cored on the continentalrise off Prydz Bay, East Antarctica (64°22 S, 67°13 E),which revealed a terrigenous and hemipelagic sedimentrecord of early Miocene–Quaternary age. The upper-most 50 m of the succession consist of hemipelagic andpelagic Pliocene–Pleistocene sediments (O'Brien et al.,2001) characterised by brown diatom clay with minordiatom bearing green clay that contains dispersed lone-stones (Florindo et al., 2003).
Here we provide evidence from ODP Site 1165 that atransient warm interval occurred during the Pleistocene,circa 1 Ma, based on new bio-chronostratigraphic dataand nannofossil assemblage changes.
2. Site location and analytical methods
ODP Leg 188 consists of a transect of three sites(1165, 1166 and 1167) drilled across Prydz Bay alongthe Antarctic continental margin, between 68°E and78°E (Fig. 1) (O'Brien et al., 2001; Cooper and O'Brien,2004). Sediment samples used in this study were ob-tained from ODP Site 1165B, drilled in a water depth of3537 m on the continental rise offshore from Prydz Bay(latitude of 64°22 S; longitude of 67°13 E). It recovereda mix of pelagic and hemipelagic sediments of thecentral Wild Drift, which was formed by the interactionof sediment supplied from the shelf and westward-flowing currents on the continental rise. This site pene-trated 682.02 mbsf, with a recovery of ca. 63%.
2.1. Calcareous nannofossils
The upper 50 m from Cores 1 to 5 were analysed forcalcareous nannofossils, but only the uppermost sedi-ments (from 0 to 15.74 mbsf, Cores 1 and 2) werefossiliferous. Samples were collected at 15 cm intervalsand 142 smear slides were prepared following standardtechniques (Bown and Young, 1998). Smear slides wereexamined using a polarizing light microscope (LM) ata magnification of 1250×. For each sample, all thenannofossil specimens in 400 fields of view werecounted. Total abundance (Fig. 2), preservation degree,species abundance and richness were determined (Table1). Due to the typically low abundance of calcareousnannofossils, bioevents were recorded as lowest (LOw)and highest (HO) occurrences of a species, rather thantheir first and last occurrence (FO and LO). As pointedout by Aubry (1995) and Aubry and Villa (1996), LOwand HO are stratigraphic terms without a temporal
connotation and are useful for relating biostratigraphicsuccessions affected by erosion, poor preservation andlow abundance, as in our study.
The genus Gephyrocapsa was differentiated into threesize groups, following Raffi et al. (1993). Gephyrocapsidsb4 !m were assigned to the group of the ‘smallGephyrocapsa spp.’, gephyrocapsids ranging from 4 to5.5 !m to the ‘medium-sized Gephyrocapsa spp.’ and,finally, gephyrocapsids N5.5 !m were attributed to the‘large Gephyrocapsa spp.’
3. Results
3.1. Biostratigraphy
Calcareous nannofossils are unevenly distributedthroughout the samples, as also reported by Pospichal(2003). They occur discontinuously only in Cores 1 and2 with variable total abundance (Table 1), whereasCores 3, 4 and 5 are barren. Sediments of the upper6.55 m display scattered low abundances and low-di-versity assemblages; from 6.55 mbsf to 15.74 mbsf, thepreservation is moderate and the total abundanceincreases slightly. Assemblages are dominated by Coc-colithus pelagicus and Reticulofenestra minuta.
Despite the discontinuous occurrence of calcareousnannofossils, key bioevents have been detected using
Fig. 2. Magnetostratigraphy (Florindo et al., 2003), nannofossil totalabundance plot, and oxygen isotope curve of the Pleistocene intervalof Site 1165 (Warnke et al., 2004). Nannofossil total abundance isexpressed as number of specimens/mm2. Numbers on !18O curve referto oxygen isotope stages interpretation.
232 G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

species abundance data. However, the position of thebiostratigraphic events must be considered with somecaution, because of the strong variability observed in theassemblage abundance.
Six calcareous nannofossil biohorizons are detectedin the uppermost 15.74 mbsf of the section. Some ofthese bioevents have been previously included in thestandard biostratigraphic schemes of Martini (1971),Gartner (1977), Okada and Bukry (1980), Young (1991)or discussed as biohorizons (i.e. Gartner, 1977; Thier-stein et al., 1977;Matsuoka andOkada, 1990;Wei, 1993;Raffi et al., 1993; Raffi et al., 2006).
Other biostratigraphic studies focused on Pleistocenesediments from the Southern Ocean discussed the repro-ducibility of the standard bioevents at the southern highlatitudes (i.e. Wei and Wise, 1992; Flores et al., 2000;Flores and Marino, 2002).
The main biohorizons recorded in the studied cores(Fig. 3) are reported and discussed below, from youngerto older.
– LOw of Emiliania huxleyi is observed at0.25 mbsf. The LOw of this taxon is generally re-corded at 268 ka, in the Marine Isotope Stage (MIS) 8(Thierstein et al., 1977; Ahagon et al., 1993). Thisbiohorizon represents the youngest biostratigraphicevent recorded.– HO of Pseudoemiliania lacunosa is observed at3.85 mbsf. The HO of P. lacunosa was identifiedby Thierstein et al. (1977) within the MIS 12. Allspecimens recorded of P. lacunosa are b5 !m. Thelast specimens observed are at 3.85 mbsf. Above thislevel rare nannofossils are present up to 3.25 mbsf;therefore we consider this bioevent as the HO.From 3.25 to 1.15 mbsf, the samples are barren innannofossils.– HO of Reticulofenestra asanoi is observed at7.05 mbsf. The HO of R. asanoi was recognised as asynchronous event by Sato and Takayama (1992) andWei (1993), both in low and relatively high latitudes,within the late MIS 22 (830 ka). Sato and Takayama(1992) considered only the circular to subcircularspecimens with a diameter between 6 and 8 !m.Matsuoka and Okada (1989) andWei (1993) based theHO of R. asanoi considering circular and subcircularforms with diameter N6.5 !m. At Site 1165B, R.asanoi (N6.5 !m) disappears abruptly at 7.05 mbsf.– LOw of Reticulofenestra asanoi is observed at14.39 mbsf. Wei (1993) recorded the LOw of thisspecies in the interval encompassing MIS 35–MIS29, whereas Sato and Takayama (1992) placed it inMIS 30 (1060 ka). Raffi (2002) obtained an
astronomical calibration corresponding to MIS 35/34 transition (ca. 1136 ka). The low degree of reli-ability of this biohorizon could be related to differenttaxonomic concepts (see discussion above). In thePrydz Bay core, this biohorizon is preceded by aninterval where nannofossils are very rare or absent,therefore we consider the event as a lowest occur-rence rather than a first occurrence.– LOw of Gephyrocapsa oceanica N4 !m (medium-sized Gephyrocapsa) is observed at 14.39 mbsf, andthe LOw of Gephyrocapsa caribbeanica N4 !m(medium-sized Gephyrocapsa) is observed at14.99 mbsf. The occurrence of medium-sized Ge-phyrocapsa, however, falls in the lowest fossiliferousinterval, where calcareous nannofossils are very rare.Therefore, we considered them as lowest occur-rences. Calcidiscus macintyrei was not observed.Possibly, the interval corresponding to the upperportion of its range may have been eroded and thedocumented lowest occurrence of G. caribbeanicapostdates the HO of C. macintyrei.
Between 14.99 and 15.74 mbsf, calcareous nanno-fossils are present but rare, i.e. 3–4 specimens per6 mm2, and the assemblages are exclusively representedby Coccolithus pelagicus, Dictyoccocites productellus,and Reticulofenestra minuta, which display widestratigraphic ranges encompassing the Pliocene/Pleisto-cene (P/P) boundary.
3.2. Biomagnetostratigraphy and chronostratigraphy
The magnetostratigraphic interpretation for the upperpart of Hole 1165B was obtained by Florindo et al.(2003), aided by diatom and radiolarian biostratigraphy(Fig. 4). The obtained nannofossil biostratigraphicresolution, more detailed than that provided by diatoms(Whitehead and Bohaty, 2003), permits a revision ofthe previous magneto-biochronological interpretation(Figs. 3 and 4). Pospichal (2003) presented a nanno-fossil distribution based on a low-resolution sampling,which broadly agrees with our results.
Our interpretation, based on the correlation with thegeomagnetic polarity time scale of Lourens et al. (2004),is discussed below.
– A normal polarity interval from 0 to 5.37 mbsf isreferred to the Brunhes Chron based on the LOw ofEmiliania huxleyi and the HO of Pseudoemilianialacunosa.
– A reversed polarity interval extending from 5.37 to6.97 mbsf is assigned to Subchron C1r.1r (0.780–
233G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

Table 1Range chart of calcareous nannofossils from Prydz Bay. Preservation is indicated as follow: G = good, M = moderate, P = poor. Total and relative abundances are indicated as numbers ofcoccoliths/mm2. X indicates the presence of Thoracosphaera spp.
234G.Villa
etal./
Palaeogeography,P
alaeoclimatology,P
alaeoecology260
(2008)230–244

235G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

0.990 Ma), based on the HO of Reticulofenestraasanoi at 7.05 mbsf.
– A normal polarity interval from 6.97 to 14.10 mbsfcorrelates with the Jaramillo Subchron (C1r.1n,0.990–1.070 Ma) based on the absence of the largeGephyrocapsa (Raffi et al., 1993; Wei, 1993; Floreset al., 2000; Raffi et al., 2006) and the presence ofR. asanoi. In Florindo et al. (2003) this thickinterval of normal polarity was correlated to at leasta portion of the Olduvai Subchron (C2n, 1.770–1.950 Ma).
– A reversed polarity interval from 14.10 to 14.66 mbsfis associated with the LOw of R. asanoi and theabsence of large Gephyrocapsa, and is correlated toSubchron C1r.2r (1.070–1.201 Ma).
– A normal polarity interval is recognised from 14.66to the base of the studied section and may represent atleast a portion of the Olduvai Subchron (C2n, 1.770–1.950 Ma). The presence of G. caribbeanica N4 !mat 14.99 mbsf allows correlation of the uppermostportion of this normal polarity interval with the CobbMountain Subchron (C1r.2n, 1.201–1.211 Ma). Thereverse polarity C1r.3r Subchron (1.211–1.770 Ma)is inferred to be missing. This is further supported bythe absence of Calcidiscus macintyrei. Below15.00 mbsf the species recorded are not exclusivelyof Pleistocene age.
Although the P/P boundary cannot be identified atSite 1165B using calcareous nannofossil events
Fig. 3. Correlation between magnetostratigraphy and nannofossil bioevents (Raffi et al., 2006) of the Late Pliocene–Pleistocene (left column) and thebio-magnetostratigraphy recorded at the Site 1165 (right column). Calibration of magnetostratigraphy is according to Lourens et al. (2004) and themagnetic polarity zones show black (white) interval for normal (reversed) polarity. Biohorizons detected in this study are given on the right column:wavy lines on the right side of the log indicate possible hiatuses at Site 1165. LOw and HO are respectively for Lowest Occurrence and HighestOccurrence. LCO and HCO are for Lowest and highest Consistent Occurrence.
236 G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

according to the geomagnetic polarity time scale it isplaced close to the upper limit of the Olduvai Subchron,therefore in this section it is probably missing due to thesupposed unconformity described above.
Florindo et al. (2003) suggested the occurrence ofseveral hiatuses in the two upper cores in order toexplain the thinness of the Matuyama (C1r, 0.780–2.581 Ma) and Brunhes Chrons. The only hiatus that isdiscernible biostratigraphically on the basis of our datais located below Cobb Mountain Subchron. On the basisof the simple relative thickness of the Bruhnes Chronand Jaramillo Subchron, the presence of other uncon-formities in the upper part of the succession cannot beruled out. In particular, in the Bruhnes Chron one ormore hiatuses could justify its relative thinness. Yet, ahiatus from the uppermost part of the Jaramillo to thelower part of C1r.1r Subchron, could possibly be inf-erred if we consider the recorded last occurrence of R.asanoi as synchronous to the mid-latitude event, i.e.within Subchron C1r.1r. (Maiorano and Marino, 2004;Raffi et al., 2006).
The thickness of the Jaramillo Subchron implies ahigh sedimentation rate (ca. 89 m/my on average).Sedimentation rates of the same order of magnitudehave been documented during the Pleistocene–Holo-cene interval for other Antarctic areas with comparablesettings, up to 16 m/ky, i.e. in the Wilkes Land Basin(Escutia et al., 2003). The increase of the sedimentationrate could also be related to an increase of both hemi-pelagic and pelagic components. The major input ofsiliciclastic sediments was probably shed from thecontinent through the Lambert Glacier–Amery IceShelf, one of the largest drainage systems (Hambreyand Dowdeswell, 1994), whereas the pelagic componentcould be associated to enhanced primary productivity ofboth siliceous and, subordinately, calcareous organisms.
3.3. Nannofossil occurrence
At Site 1165, calcareous nannofossils are moreabundant in the Lower Pleistocene interval, whereas intheMiddle–Upper Pleistocene their abundance decreasesdrastically (Fig. 2). In particular, the uppermost 6 m ofsediments contains mainly poorly diversified assem-blages, and the microfossil associations are dominated bydiatoms. From 6 to 15 mbsf, low diversified but frequentto abundant calcareous nannofossils co-occur withThoracosphaera spp. and less abundant diatoms andsilicoflagellates. Coccolithus pelagicus dominates theassemblage and represents 60–100% of the nannoflora(Fig. 5). This species has been considered as a cold-waterindicator and was observed elsewhere in the Pleistocene
of Southern Ocean by Villa et al. (2005). Calcidiscusleptoporus is very rare, and a negative covariance withC.pelagicus (Findlay and Flores, 2000), was not confirmedhere, this may be due to the extreme environmentalconditions. Another dominant species of the assemblagesis Reticulofenestra minuta. Also well represented aremedium-sized Gephyrocapsa such as Gephyrocapsacaribbeanica and Gephyrocapsa oceanica.
3.4. Magnetic susceptibility record
The magnetic susceptibility (") includes contribu-tions from all minerals present in sediment in proportionto their abundance. It is mostly used as a relative proxy
Fig. 4. Comparison between themagnetostratigraphic interpretation forthe Site 1165 according to Florindo et al. (2003) (left) and the re-interpretation of the same site based on calcareous nannofossil biostra-tigraphy (right). Age calibration of magnetostratigraphy is according toLourens et al. (2004). Wavy lines indicate hiatuses hypothesized in thetwo interpretations.
237G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

indicator for changes in sediment composition thatcan be linked to palaeoclimate-controlled depositionalprocesses. In this record, downcore variations of " areassociated with similar changes in other bulk magneticparameters such as the artificial remanences intensities(namely, ARM and IRM), which suggested that these
fluctuations are controlled by changes in magnetiteconcentrations (Florindo et al., 2003; Richter et al.,2003; Warnke et al., 2004).
The " record shows various large- and small-scalecycles (Fig. 6), which are expressed also in the sedimentgranulometry (Warnke et al., 2004). These changes may
Fig. 6. a) Low-field magnetic susceptibility (") and sediment granulometry for the uppermost 15 m at Site 1165B (Grützner, 2003; Warnke et al.,2004). b) Amplitude of " variations expressed as deviation between magnetic susceptibility values and its 3-point moving average.
Fig. 5. Species abundance of selected nannofossil taxa as recognised through the Site 1165. The abundances are expressed as number of specimens/mm2 in 400 fields of view plotted against the magnetostratigraphy.
238 G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244
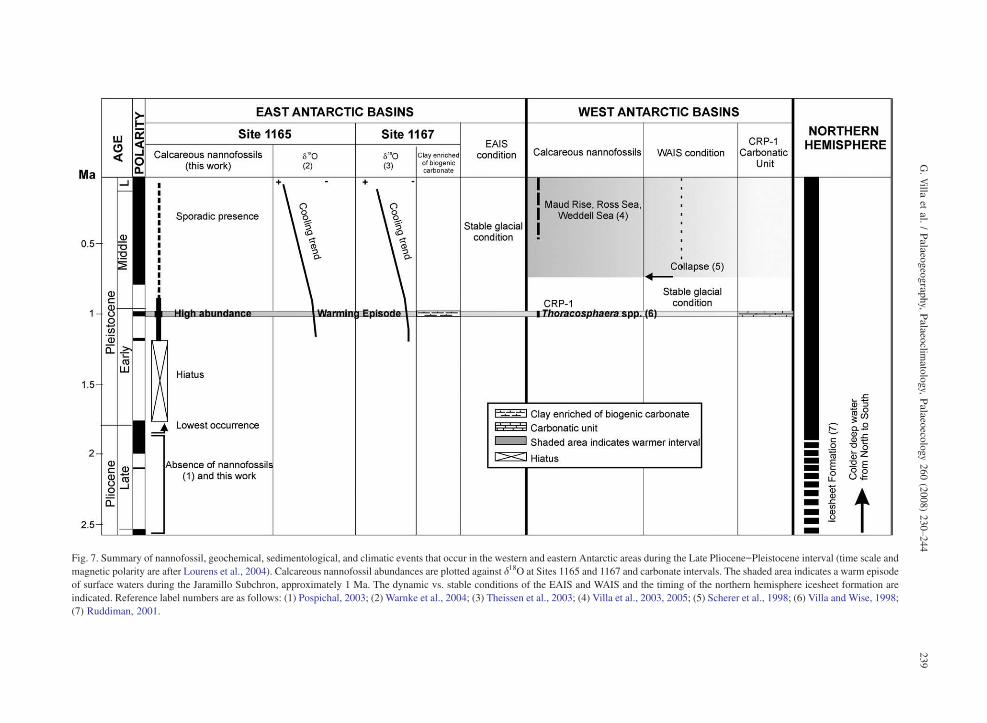
Fig. 7. Summary of nannofossil, geochemical, sedimentological, and climatic events that occur in the western and eastern Antarctic areas during the Late Pliocene–Pleistocene interval (time scale andmagnetic polarity are after Lourens et al., 2004). Calcareous nannofossil abundances are plotted against !18O at Sites 1165 and 1167 and carbonate intervals. The shaded area indicates a warm episodeof surface waters during the Jaramillo Subchron, approximately 1 Ma. The dynamic vs. stable conditions of the EAIS and WAIS and the timing of the northern hemisphere icesheet formation areindicated. Reference label numbers are as follows: (1) Pospichal, 2003; (2) Warnke et al., 2004; (3) Theissen et al., 2003; (4) Villa et al., 2003, 2005; (5) Scherer et al., 1998; (6) Villa and Wise, 1998;(7) Ruddiman, 2001.
239G.Villa
etal./
Palaeogeography,P
alaeoclimatology,P
alaeoecology260
(2008)230–244

reflect temporal variations in terrigenous input, whichare likely caused by changes in sea level and erosion ofthe continental shelf in the region of Prydz Bay. At afiner scale, from about 7.00 mbsf upward, it is worthnoting an increase in “between-sample” variability. Thismay indicate a more heterogeneous sediment composi-tion during a cooling trend, starting from 900 ka (Fig. 7),possibly associated to the presence of more floatingdebris-charged icebergs at this site.
4. Discussion
4.1. Evidence for an Early Pleistocene warming
For most of the interval studied, nannofossils aretypically low in abundance, except during some Pleisto-cene intervals, in which higher abundances wereobserved. This suggests that more favourable conditionsdeveloped leading to improvement in productivity andpreservation such as: (1) high nannofloral productivityin surface waters around Prydz Bay, possibly associatedwith warmer intervals; (2) periodic downward fluctua-tions in the calcium carbonate compensation depth(CCD), permitting carbonate preservation; (3) rapidtransport and burial downslope; or (4) minor disso-lution and overgrowth during diagenesis that can con-trol nannofloral assemblage compositions and relativeproportions.
Based on the quantitative observations, as documen-ted by the presence of Emiliania huxleyi and Pseudoe-miliania lacunosa that are dissolution-susceptible taxawith high surface/volume ratio, the assemblages aregenerally moderately well preserved. Therefore, lowspecies diversity and low total abundance are probablyrelated to the extreme environmental conditions, ratherthan dissolution. These results document that sea surfacetemperature (SST), sea-ice, and light conditions in PrydzBay became favourable for coccolithophorid productionduring interglacial intervals of the Early and MiddlePleistocene, when SST maxima were warmer than today(Villa et al., 2005). Although an enhanced preservationduring interglacial intervals would be difficult owing to arelative shallowCCD (Bonn et al., 1994), the presence ofcoccoliths at the 3537 m water depth of Site 1165 couldbe explained by a rapid burial during high primaryproductivity of siliceous and calcareous organisms(Pudsey, 2000). Also, the nannofossil assemblages donot show evidence of reworking. Species are exclusivelyof Pleistocene age and the bioevents recorded arehomotaxic with those reported in the standard biostrati-graphic schemes, therefore the observed nannofossilscan be considered in place and reliable for palaeoeco-
logical and palaeoceanographical interpretation. Thisrecord contrasts with the distribution of modern coc-colithophorids that are confined to lower and middlelatitudes (Verbeek, 1989; Honjo, 1990; Findlay andGiraudeau, 2000; Gravalosa et al., 2004) and implies thatmore favourable conditions existed for nannofossilaccumulation.
4.2. Relationship between nannofossil assemblage andpalaeoceanographic setting
During the late Early Pleistocene (Jaramillo Sub-chron), the higher abundance of nannofossils in ourcores suggests enhanced warm SST and open-marineconditions. This is supported by evidence from Site1167, which contains a clayey interval enriched ofcalcareous plankton, dated at 900–1090 ka, which isinterpreted by Theissen et al. (2003) as the response towarmer conditions and reduced ice-sheet volume. It isworth noting that in the shelly carbonate sequencerecovered in the Cape Roberts Project (CRP-1) (CapeRoberts Science Team, 1998) with an age estimated of1070 ka (Scherer et al., 2002), the unexpected presenceof Thoracosphaera spp. for such high latitudes (Villaand Wise, 1998) was related, together with otherpalaeontological evidence (Bohaty et al., 1998; Tavianiand Claps, 1998), to a warm interval in the midst of aQuaternary glacial period.
As discussed above, in the Site 1165 the highercalcareous nannofossil abundances, including the pre-sence of Thoracosphaera spp., are recorded within theJaramillo Subchron. Our biostratigraphic data supportthe hypothesis of an unconformity at the boundaryJaramillo/C1r.1r and, therefore, the top of the Jaramillomay be missing. As a result, the warmer SST intervalcould correspond to MIS 31 in the lower part of theJaramillo Subchron, as recorded at the CRP-1.
The presence of higher abundances of calcareousnannofossils also suggests that, in the Eastern Antarcticbasins, a warmer interval occurred during the Quatern-ary dominated by a glacial climate regime (Fig. 7). Infact, before this warmer interval, the absence of calcar-eous nannofossils, in our late Pliocene samples, seemsto indicate lower SST, which is in agreement with theglobal cooling trend that started at 3.2–2.5 Ma (Sikeset al., 1991; Raymo et al., 1992, Baker and Camerlen-ghi, 2002).
The sharp decrease of nannofossils from the C1r.1rSubchron (ca. the last 900 ka) upward correlates withthe trend of heavier !18O values (Warnke et al., 2004),which reflect an increase of ice volume and cooling forthis upper interval (Fig. 2), as indicated also by the
240 G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

magnetic susceptibility record. Since a minimum of 2–3 °C in surface–subsurface waters is necessary for coc-colithophorid reproduction (Burckle and Pokras, 1991;Findlay and Giraudeau, 2000), we argue that since900 ka, the SST in the Eastern Antarctic basins droppedbelow the aforementioned minimum value, and onlysporadically exceeded. Moreover, at the nearby Site1167, Theissen et al. (2003) interpreted the increasedoxygen isotope values, as the response to a cooling trendbeginning at 900 ka. The results presented in this paper,support the hypothesis of stable polar glacial conditionssince 900 ka in the Prydz Bay region, which is part ofthe EIAS.
5. Conclusions
An increase in nannofossil abundances observed incore samples obtained from ODP Site 1165 is consistentwith an anomalous warming event in the surface watersin and around Prydz Bay during the Early Pleistocene, at!1 Ma, recognizable as MIS 31 Climate proxies fromother studies around Antarctica (the oxygen isotopestratigraphy from Sites 1165 and 1167, the presence ofclay enriched of biogenic carbonate at Site 1167, and theCRP-1 shelly carbonate unit) also support a warmingevent during this time interval, suggesting that it wasextended around the Antarctic Continent. This anom-alous warming event in the Southern Ocean surfacewaters is the only known warm event recorded in thePleistocene for the Eastern Antarctic marginal basins, sofar. The warm interval during the Jaramillo Subchron,involving the EAIS and surrounding waters, indicatesthat these areas were climatically more dynamic thanpreviously thought. We conclude that this warmingevent does not support the notion that Antarctica and theSouthern Ocean have remained in a stable polarcondition since the late Neogene.
Following this warm interval, during the last900 ka, in Prydz Bay, Wilkes Land Basin (Busetti etal., 2003) and Bausan Bank (Villa et al., 2005), sea-surface conditions did not allow the growth andpropagation of coccolithophores, suggesting a stableglacial condition for the EAIS. In contrast, nannofossilassemblages reported from the Ross Sea, Weddell Sea,Maud Rise (Villa et al., 2005), the Antarctic Peninsula(Villa et al., 2003) in the Middle Pleistocene (presenceof Emiliania huxleyi), confirm that, at times, theWestern Antarctic basins experienced conditions war-mer than today, as also suggested by Scherer et al.(1998), who proposed at least a partial collapse of theWAIS from 750 ka, and specifically during MIS 11–MIS 5.
The detailed analyses and the high-resolution samplingon calcareous nannofossils permitted a refined magnetos-tratigraphy for the uppermost Pliocene–Pleistocene ofHole 1165 B. Comparisons of our data with the isotopicrecord from Sites 1165 and 1167 and with the Quaternarysequence of CRP-1 indicate the robustness of our agemodel. In particular, our obtained agemodel indicates that:
– there is no biostratigraphic evidence for a hiatus inthe Brunhes Chron although the relatively reducedthickness of the Brunhes Chron is best explained bythe presence of one or more disconformities as out-lined by Florindo et al. (2003);
– a high sedimentation rate occurred during the JaramilloSubchron; a hiatus concerning its top and the base ofC1r.1r is inferred.
– Chron C1r.3r. is absent due to a hiatus.
Acknowledgements
We thank James Pospichal, onboard nannofossilspecialist on Leg 188, for sharing his preliminary data.The Ocean Drilling Program furnished samples to G.V.
We also thank Isabella Raffi, Woody Wise, JimPospichal and two anonymous reviewers for theirconstructive review and suggestions.
This study was supported by PNRA Grant to G.V.and FAR Grant to M.C. This is a contribution to theSCAR Scientific Research Program “Antarctic ClimateEvolution” (ACE).
Appendix A. Taxonomic list
Calcareous nannofossils considered in this report (inalphabetical order by generic epithets).
Calcidiscus leptoporus (Murray and Blackman,1898) Loeblich and Tappan, 1978.
Calcidiscus macintyrei (Bukry & Bramlette, 1969)Loeblich & Tappan, 1978.
Ceratolithus telesmus Norris, 1965.Coccolithus pelagicus (Wallich, 1877) Schiller, 1930.Coccolithus pelagicus braarudii (Gaarder, 1962)
Geisen et al., 2002.Crenalithus japonicus (Nishida, 1971 emend. Nishida,
1979) de Kaenel et al., 1999.Crenalithus doronocoides (Black and Barnes) Roth,
1973.Emiliania huxleyi (Lohmann, 1902) Hay and Mohler,
in Hay et al., 1967.Gephyrocapsa aperta Kamptner, 1963.Gephyrocapsa caribbeanica Boudreaux and Hay,
1967.
241G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

Gephyrocapsa ericsonii McIntyre and Bé, 1967.Gephyrocapsa muellerae Bréhéret, 1978.Gephyrocapsa oceanica Bréhéret, 1978.Gephyrocapsa omega Bukry, 1973.Helicosphaera burkei Black, 1971.Helicosphaera carteri (Wallich, 1877) Kamptner,
1954.Pseudoemiliania lacunosa (Kamptner, 1963) Gart-
ner, 1969.Pseudoemiliania lacunosa var. lacunosa (Young,
1990) de Kaenel, 1999.Pseudoemiliania lacunosa var. ovata (Young, 1990)
de Kaenel, 1999.Pseudoemiliania pacifica (Nishida, 1979) de Kaenel,
1999.Reticulofenestra asanoi (Sato and Takayama, 1992).Reticulofenestra minuta Roth, 1970.Thoracosphaera Kamptner, 1927.Umbilicosphaera hulburtiana Gaarder, 1970.
References
Ahagon, N., Tanaka, Y., Ujiie, H., 1993. Florisphaera profunda, apossible nannoplankton indicator of late Quaternary changes insea-water turbidity at the northwestern margin of the Pacific.Marine Micropaleontology 22 (3), 255–273.
Aubry, M.-P., 1995. From chronology to stratigraphy: interpreting thelower and middle Eocene stratigraphic record in the AtlanticOcean. In: Berggren, W.A., Kent, D.V., Aubry, M.-P., Hardenbol,J. (Eds.), Geochronology, Time Scales and Global StratigraphicCorrelations: a Unified Temporal Framework for an HistoricalGeology. Special Volume, vol. 54. Society of Economic Geologistsand Mineralogists, pp. 213–274.
Aubry, M.-P., Villa, G., 1996. Calcareous nannofossil stratigraphy ofthe Lemme-Carrosio Paleogene/Neogene Global Stratotype Sec-tion and Point. Giornale di Geologia 581/1–2, 51–69 ser.3a.
Baker, P.F., Camerlenghi, A., 2002. Synthesis of Leg 178: glacialhistory of the Antarctic Peninsula from Pacific margin sediments.In: Barker, P.F., Camerlenghi, A., Acton, G.D., Ramsay, A.T.S.(Eds.), Proc. ODP, Scientific Results, vol. 178, pp. 1–40. [Online].Available from World Wide Web: http://www.odp.tamu.edu/publications/178_SR/VOLUME/SYNTH/SYNTH.PDF.
Barrett, P., 2003. Cooling a continent. Nature 421, 221–223.Bohaty, S.M., Scherer, R.P., Harwood, D.M., 1998. Quaternary
Diatom Biostratigraphy and palaeoenvironments of the CRP-1Drillcore, Ross Sea, Antarctica. Terra Antartica 5 (3), 431–453.
Bonn, W.J., Fütterer, D.K., Grobe, H., 1994. Quaternary sedimentationat the Antarctic continental margin. Terra Antartica 1 (2), 319–320.
Bown, P., Young, J.R., 1998. Techniques. In: Bown, P.R. (Ed.),Calcareous Nannofossil Biostratigraphy. British Micropalaeontol-ogy Society Series. Chapman & Hall, pp. 16–28.
Burckle, L.H., Pokras, E.M., 1991. Implications of a Pliocene stand ofNothofagus (southern beech) within 500 kilometres of the SouthPole. Antarctic Science 3 (4), 389–403.
Busetti, M., Caburlotto, A., Armand, L., Damiani, D., Giorgetti, G.,Lucchi, R.C., Quilty, P.G., Villa, G., 2003. Plio-Quaternarysedimentation on the Wilkes Land continental rise. Preliminaryresults. Deep-sea Research, Part II 50 (8–9), 1529–1562.
Cape Roberts Science Team, 1998. Initial report from CRP-1, CapeRoberts Project, Antarctica. Terra Antartica 5 (1), 1–187.
Cooper, A.K., O'Brien, P.E., 2004. Leg 188 synthesis: transitions in theglacial history of the Prydz Bay region, East Antarctica, from ODPdrilling. In: Cooper,A.K.,O'Brien, P.E., Richter, C. (Eds.), Proc.ODP,Sci. Results, vol. 188. [Online]. Available from World Wide Web:http://www-odp.tamu.edu/publications/188_SR/synth/synth.htm.
Escutia, C., Warnke, D.A., Acton, G.D., Barcena, A., Burckle, L.,Canals, M., Frazee, C.S., 2003. Sediment distribution andsedimentary processes across the Antarctic Wilkes Land marginduring the Quaternary. Deep-Sea Research. Part 2. Topical Studiesin Oceanography 50, 1481–1508.
Findlay, C.S., 1998. Living and fossil calcareous nannoplankton fromthe Australian sector of the Southern Ocean: implications forpaleoceanography. Ph.D. dissertation, University of Tasmania,(Hobbart, Australia). 148 pp. Unpublished.
Findlay, C.S., Flores, J.A., 2000. Subtropical front fluctuations south ofAustralia (45°09 S, 146°17 E) for the last 130 ka based on calcareousnannoplankton. Marine Micropaleontology 40, 403–416.
Findlay, C.S., Giraudeau, J., 2000. Extant calcareous nannoplankton inthe Australian sector of the Southern Ocean (austral summers 1994and 1995). Marine Micropaleontology 40, 417–439.
Flores, J.A., Marino, M., 2002. Pleistocene calcareous nannofossilstratigraphy for ODP Leg 177 (Atlantic sector of the SouthernOcean). Marine Micropaleontology 45, 191–224.
Flores, J.A., Gersonde, R., Sierro, F.J., Niebler, H.-S., 2000. Southernocean Pleistocene calcareous nannofossil events: calibration withisotope and geomagnetic stratigraphies. Marine Micropaleontol-ogy 40, 377–402.
Florindo, F., Bohaty, S.M., Erwin, P.S., Richter, C., Roberts, A.P.,Whalen, P.A., Whitehead, J.M., 2003. Magnetostratigraphicchronology and palaeoenvironmental history of Cenozoicsequences from ODP sites 1165 and 1166, Prydz Bay, Antarctica.Palaeogeography Palaeoclimatology Palaeoecology 198, 69–100.
Gartner, S., 1977. Clacareous nannofossil biostratigraphy and recisedzonation of the Pleistocene. Marine Micropaleontology 2, 1–25.
Gravalosa, J.M., Flores, J.A., Sierro, F.J., Gersonde, R., 2004. Livingcoccolithophores in the Amundsen and Bellinghausen Seas,Antarctic and Subantarctic Pacific sector. Journal of Nannoplank-ton Research 26 (2), 50.
Grützner, J., 2003. Data report: multisensor core logging data, coarse-fraction grain-size analyses, and biogenic silica content of upperMiocene–lower Pliocene sediments, ODP Site 1165. In: Cooper,A.K., O'Brien, P.E., Richter, C. (Eds.), Proc. ODP, Sci. Results,vol. 188. [Online]. Available fromWorld Wide Web: bhttp://www-odp.tamu.edu/publications/188_SR/007/007.htmN.
Hambrey, M.J., Dowdeswell, J.A., 1994. Flowregime of the LambertGlacier–Amery Ice Shelf System, Antarctica: structural evidencefrom satellite imagery. Annals of Glaciology 20, 401–406.
Hambrey, M.J., McKelvey, B.C., 2000. Neogene fjordal sedimentationon the western margin of the Lambert Graben, East Antarctica.Sedimentology 47, 577–607.
Honjo, S., 1990. Particle fluxes and modern sedimentation in thepolar oceans. In: SmithJr. Jr., W.O. (Ed.), Polar Oceanography:Part B. Chemistry, Biology and Geology. San Diego AcademicPress, pp. 687–739.
Huybrechts, P., 1993. Glaciological modelling of the Late CenozoicEast Antarctic ice sheet: stability or dynamism? GeografiskaAnnaler. Series A 75, 221–238.
Lear, C.H., Elderfield, H., Wilson, P.A., 2000. Cenozoic deep-seatemperatures and global ice volumes from Mg/Ca in BenthicForaminiferal Calcite. Science 287, 269–272.
242 G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

Lourens, L.J., Hilgen, F.J., Laskar, J., Schakleton, N.J., Wilson, D.,2004. The Neogene Period. In: Gradstein, F., Ogg, J., Smith, A.G.(Eds.), Geological Time Scale 2004. Cambridge University Press,Cambridge, pp. 409–440.
Kennett, J.P., Barker, P.F., 1990. Latest Cretaceous toCenozoic climate andoceanographic developments in theWeddell Sea, Antarctica; an ocean-drilling perspective. Proceedings of the Ocean Drilling Program,Weddell Sea, Antarctica, covering Leg 113 of the Cruises of theDrilling Vessel JOIDES Resolution, Valparaiso, Chile, to East Cove,Falkland Islands, sites 689–697, 25 December 1986–11 March 1987.
Kennett, J.P., Hodell, D.A., 1995. Stability or instability of Antarcticice sheets during warm climates. GSA Today 5 (1), 9–22.
Maiorano, P., Marino, M., 2004. Calcareous nannofossil bioevents andenvironmental control on temporal and spatial patterns at the early–middle Pleistocene. Marine Micropaleontology 53, 405–422.
Martini, E., 1971. Standard Tertiary and Quaternary calcareousnannoplankton zonation. In: Farinacci, A. (Ed.), Proceeding of2nd International Conference of Planktonic Microfossils Roma,vol. 2, pp. 739–785.
Matsuoka, H., Okada, H., 1989. Quantitative analysis of Quaternarynannoplankton in the subtropical northwestern Pacific Ocean.Marine Micropaleontology 14, 97–118.
Matsuoka, H., Okada, H., 1990. Time-progressive morphometricchanges of the genus Gephyrocapsa in the Quaternary sequence ofthe tropical Indian Ocean, Site 709. In: Duncan, R.A., Backman, J.,Peterson, L.C., et al. (Eds.), Proceeding ODP, Scientific Results,vol. 115. TXOceanDrilling Program, College Station, pp. 255–270.
O'Brien, P.E., Cooper, A.K., Richter, C., et al., 2001. Proc. ODP, Init.Rep., vol. 188 (online). Available from World Wide Web: http://www-odp.tamu.edu/publications/188_IR/188ir.htm.
Okada, H., Bukry, D., 1980. Supplementary modification andintroduction of code numbers to the low-latitude coccolithbiostratigraphic zonation. (Bukry, 1973; 1975). Marine Micro-paleontology 5, 321–325.
Pospichal, J.J., 2003. Calcareous nannofossils from Continental RiseSite 1165, ODP Leg 188, Prydz Bay, Antarctica. In: Copper, A.K.,O'Brien, P.E., Richter, C. (Eds.), Proceeding ODP Leg 188,Scientific Results, 188 (Online). Available from World Wide Web:http://www-odp.tamu.edu/publications/188_SR/014/014.htm.
Pudsey, C.J., 2000. Sedimentation on the continental rise west of theAntarctic Peninsula over the last three glacial cycles. MarineGeology 167, 313–338.
Raffi, I., 2002. Revision of the early–middle Pleistocene calcareousnannofossil biochronology (1.75–0.85 Ma). Marine Micropaleon-tology 45, 25–55.
Raffi, I., Backman, J., Rio, D., Shackleton, N.J., 1993. Plio-Pleistocene nannofossil biostratigraphy and calibration to oxygenisotopes stratigraphies from Deep Sea Drilling Project Site 607 andOcean Drilling Program Site 677. Paleoceanography 8, 387–408.
Raffi, I., Backman, J., Fornaciari, E., Palike, E., Rio, D., Lourens, L.,Hilgen, F., 2006. A review of calcareous nannofossil astrobio-chronology encompassing the past 25 million years. QuaternaryScience Reviews. doi:10.1016/j.quascirev.2006.07.007.
Raymo, M.E., Hodell, D., Jansen, E., 1992. Response of deepocean circulation to initiation of northern hemisphere glaciations(3–2 Ma). Paleoceanography 7, 645–672.
Richter, C., Warnke, D.A., Florindo, F., 2003. Environmentalmagnetism of late Cenozoic sediments from the East Antarcticcontinental rise (Site 1165, Prydz Bay). Eos Trans. AGU 84 (46)Fall Meet. Suppl., Abstract PP31B-0263.
Ruddiman, W.F., 2001. Earth's Climate: Past and Future. W.H.Freeman and Company, New York.
Sato, T., Takayama, T., 1992. A stratigraphically significant newspecies of the calcareous nannofossil Reticulofenestra asanoi. In:Ishizaki, K., Saito, T. (Eds.), Centenary of Japanese Micropaleon-tology. Terra Sci. Publ., Tokyo, pp. 457–460.
Scherer, R.P., 1991. Quaternary and Tertiary microfossils from beneathIce Stream B; evidence for a dynamic West Antarctic ice sheethistory. Global and Planetary Change 4, 395–412.
Scherer, R.P., Aldahan, A., Tulaczyk, S., Possnert, G., Engelhardt, H.,Kamb, B., 1998. Pleistocene collapse of the West Antarctic icesheet. Science 281, 82–85.
Scherer, R.P., Bohaty, S., Harwood, D., Roberts, A., Taviani, M., 2002.Sustained sea-ice free conditions in the Antarctic nearshore zoneduring Marine Isotope Stage 31 (1.07 MA). GSA Abs. Prog.
Sikes, E.L., Keigwin, L.D., Currey, W.B., 1991. Pliocene paleoceano-graphy: circulation and oceanography changes associated with the2.4 Ma glacial event. Paleoceanography 6, 245–257.
Sugden, D.E., Marchant, D.R., Denton, G.H., 1993. The case for astable East Antarctic ice sheet: the background. In: Sugden, D.E.,Marchant, D.R., Denton, G. (Eds.), The Case for a Stable EastAntarctica Ice Sheet. Proceedings, Geografiska Annaler. Series A:Physical Geography. Generalstabens Litografiska Anstlalt, Stock-holm, Sweden, pp. 151–154.
Taviani, M., Claps, M., 1998. Biogenic Quaternary carbonates in theCRP-1 drillhole, Victoria Land basin, Antarctica. Terra Antart 5,411–418.
Theissen, K.M., Dunbar, R.B., Cooper, A.K., Mucciarone, D.A.,Hoffmann, D., 2003. The Pleistocene evolution of the EastAntarctic Ice Sheet in the Prydz Bay region: stable isotopicevidence from ODP Site 1167. Global and Planetary Change 39,227–256.
Thierstein, H.R., Geitzenauer, K., Molfino, B., Shackleton, N.J., 1977.Global synchroneity of late Quaternary coccolith datum levels:validation by oxygen isotopes. Geology 5, 400–404.
Verbeek, J.W., 1989. Recent calcareous nannoplankton in the south-ernmost Atlantic. Polarforschung 59, 45–60.
Villa, G., Wise, S.W., 1998. Quaternary Calcareous nannofossil fromthe Antarctic region. Terra Antartica 5 (3), 479–484.
Villa, G., Persico, D., Bonci, M.C., Lucchi, R.G., Morigi, C., Rabesco,M., 2003. Biostratigraphic characterization and Quaternarymicrofossil palaeoecology in sediment drifts west of the AntarcticPenisula — implications for cyclic glacial-interglacial deposition.Paleogeography Paleoclimatology Paleoecology 198, 237–263.
Villa, G., Palandri, S., Wise, S.W., 2005. Quaternary calcareousnannofossils from Periantarctic basins: paleoecological and paleocli-matic implication. Marine Micropaleontology 56, 103–121.
Warnke, D.A., Richter, C., Florindo, F., Damuth, J.E., Balsam, W.L.,Strand, K., Ruikka, M., Juntila, J., Theissen, K., Quilty, P., 2004.Data report: HiRISC (High-Resolution Integrated StratigraphyCommittee) Pliocene–Pleistocene interval, 0–50 mbsf, at ODPLeg 188 Site 1165, Prydz Bay, Antarctica. In: Cooper, A.K.,O'Brien, P.E., Richter, C. (Eds.), Proc. ODP, Sci. Results. [Online].Available from World Wide Web: bhttp://www-odp.tamu.edu/publications/188_SR/015/015.htmN.
Webb, P.N., Harwood, D.M., McKelvey, B.C., Mercer, J.H., Stott, L.D.,1984. Cenozoic marine sedimentation and ice volume variation onthe East Antarctic craton. Geology 12 (5), 287–291.
Wei, W., 1993. Calibration of upper Pliocene–lower Pleistocenenannofossil events with oxygen isotope stratigraphy. Paleoceano-graphy 8, 85–99.
Wei, W., Wise, S.W., 1992. Oligocene–Plistocene calcareousnannofossils from Southern Ocean Sites 747, 748 and 751. Proc.Ocean Drilling Sci. Results 120, 368–373.
243G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244

Whitehead, J.M., Bohaty, S.M., 2003. Data report: Quaternary–Pliocene diatom biostratigraphy of ODP Sites 1165 and 1166,Cooperation Sea and Prydz Bay. In: Cooper, A.K., O'Brien, P.E.,Richter, C. (Eds.), Proc. ODP, Sci. Results, vol. 188. [Online].Available from World Wide Web: http://www-odp.tamu.edu/publications/188_SR/008/008.htm.
Young, J.R., 1991. A Quaternary nannofossil range chart. INANewsletter 13/1, 14–17.
Zachos, J., Pagani, M., Sloan, L., Thomas, E., Billups, K., 2001.Trends, rhythms, and aberrations in global climate 65 Ma topresent. Science 292, 686–693.
244 G. Villa et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 260 (2008) 230–244




















