
A phytochrome-dependent embryonic factor modulates gibberellin responses in the embryo and...
11
ORIGINAL ARTICLE Marı´a Vero´nica Arana Lucila Cecilia de Miguel Rodolfo Augusto Sa´nchez A phytochrome-dependent embryonic factor modulates gibberellin responses in the embryo and micropylar endosperm of Datura ferox seeds Received: 12 August 2005 / Accepted: 9 September 2005 / Published online: 7 October 2005 ȑ Springer-Verlag 2005 Abstract The promotion of germination by phytochrome is associated with extensive changes both in the embryo and in the micropylar region of the endosperm (ME) of Datura ferox seeds. These changes require de novo gibberellins (GAs) biosynthesis in the embryo, the site where the light stimulus is perceived. GAs stimulate embryo growth potential and move to ME, promoting the expression of genes related with weakening. We re- port here that, in addition, phytochrome stimulates the sensitivity of the seeds to gibberellic acid (GA). The phytochrome-induced signal is produced in the embryo and enhances the stimulus by GA of embryo growth potential (EGP) and the promotion of the expression of proteins thought to participate in ME weakening: endo- b-mannanase (EC 3.2.1.78), endo-b-mannosidase (EC 3.2.1.25) and expansin. Our results suggest that the cytokinins may be a component of the embryonic signal. Phytochrome also modulates DfPHOR and DfMYB transcript levels in ME. These genes show a high identity with components of GAs signaling identified in other species. Expression of DfPHOR in the ME is apparently regulated by phytochrome through the supply of GAs from the embryo to ME, whereas DfMYB expression is regulated by an embryonic factor with some of the characteristics of the one that modulates seed sensitivity to GAs. Keywords Coat-imposed dormancy Embryo growth potential Endosperm softening Germination Gibberellins Phytochrome Abbreviations EGP: Embryo growth potential FR: Far-red GA: Gibberellic acid GAs: Gibberellins ME: Micropylar region of the endosperm R: Red ZEA: Zeatin Introduction The successful adjustment of germination to a given environmental scenario requires the acquisition by the seeds of precise information about their situation and light is one of the main factors providing relevant signals (Bewley and Black 1994; Casal and Sa´ nchez 1998; Benech-Arnold et al. 2000). Gibberellins (GAs) are an important link between the light signals perceived by the seeds through the phytochromes and the processes controlling germination (Casal and Sa´ nchez 1998; Kamiya and Garcı´a-Martı´nez 1999). It has been shown that Pfr produces an increase in the content of GAs (Toyomasu et al. 1993) and the requirement of red light (R) for germination is circumvented by exogenous GA (Khan et al. 1957). The changes in GAs content are related with the control of biosynthetic genes. In lettuce seeds (Lactuca sativa L. cv Grand Rapids) the low fluence response (LFR) mode of the phytochromes control the expression of Ls3h1, a gene encoding a GA3b-hydroxylase, an enzyme that catalyzes the final biosynthetic step to produce bioactive GAs (Toyomasu et al. 1998). In Arabidopsis, Pfr induces GA4 and GA4H expression, also GA3b-hydroxylases (Yamaguchi et al. 1998). Similarly, in Datura ferox, Pfr stimulates tran- script accumulation of a GA3b-hydroxylase in a way consistent with its effect on germination (Burgin et al. 2000). Light-mediated changes in the sensitivity of seeds to GAs are scarcely known. It has been shown that R en- hances the response of germination to GA 4 in Arabid- opsis. This effect is mediated neither by phytochrome A nor by phytochrome B, since the WT and both phyto- chrome-deficient simple and double mutants exhibit similar light-mediated increments in GAs sensitivity M. V. Arana L. C. de Miguel R. A. Sa´nchez (&) IFEVA. Facultad de Agronomı´a, Universidad de Buenos Aires and CONICET, Avenida San Martı´n 4453, 1417 Buenos Aires, Argentina E-mail: [email protected] Tel.: +54-011-45248071 Fax: +54-011-45148730 Planta (2006) 223: 847–857 DOI 10.1007/s00425-005-0134-7
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of A phytochrome-dependent embryonic factor modulates gibberellin responses in the embryo and...

ORIGINAL ARTICLE
Marıa Veronica Arana Æ Lucila Cecilia de Miguel
Rodolfo Augusto Sanchez
A phytochrome-dependent embryonic factor modulates gibberellinresponses in the embryo and micropylar endosperm of Datura ferox seeds
Received: 12 August 2005 / Accepted: 9 September 2005 / Published online: 7 October 2005� Springer-Verlag 2005
Abstract The promotion of germination by phytochromeis associated with extensive changes both in the embryoand in the micropylar region of the endosperm (ME) ofDatura ferox seeds. These changes require de novogibberellins (GAs) biosynthesis in the embryo, the sitewhere the light stimulus is perceived. GAs stimulateembryo growth potential and move to ME, promotingthe expression of genes related with weakening. We re-port here that, in addition, phytochrome stimulates thesensitivity of the seeds to gibberellic acid (GA). Thephytochrome-induced signal is produced in the embryoand enhances the stimulus by GA of embryo growthpotential (EGP) and the promotion of the expression ofproteins thought to participate in ME weakening: endo-b-mannanase (EC 3.2.1.78), endo-b-mannosidase (EC3.2.1.25) and expansin. Our results suggest that thecytokinins may be a component of the embryonic signal.Phytochrome also modulates DfPHOR and DfMYBtranscript levels in ME. These genes show a high identitywith components of GAs signaling identified in otherspecies. Expression of DfPHOR in the ME is apparentlyregulated by phytochrome through the supply of GAsfrom the embryo to ME, whereas DfMYB expression isregulated by an embryonic factor with some of thecharacteristics of the one that modulates seed sensitivityto GAs.
Keywords Coat-imposed dormancy Æ Embryo growthpotential Æ Endosperm softening Æ Germination ÆGibberellins Æ Phytochrome
Abbreviations EGP: Embryo growth potential ÆFR: Far-red Æ GA: Gibberellic acid Æ
GAs: Gibberellins Æ ME: Micropylar region of theendosperm Æ R: Red Æ ZEA: Zeatin
Introduction
The successful adjustment of germination to a givenenvironmental scenario requires the acquisition by theseeds of precise information about their situation andlight is one of the main factors providing relevant signals(Bewley and Black 1994; Casal and Sanchez 1998;Benech-Arnold et al. 2000). Gibberellins (GAs) are animportant link between the light signals perceived by theseeds through the phytochromes and the processescontrolling germination (Casal and Sanchez 1998;Kamiya and Garcıa-Martınez 1999). It has been shownthat Pfr produces an increase in the content of GAs(Toyomasu et al. 1993) and the requirement of red light(R) for germination is circumvented by exogenous GA(Khan et al. 1957). The changes in GAs content arerelated with the control of biosynthetic genes. In lettuceseeds (Lactuca sativa L. cv Grand Rapids) the lowfluence response (LFR) mode of the phytochromescontrol the expression of Ls3h1, a gene encoding aGA3b-hydroxylase, an enzyme that catalyzes the finalbiosynthetic step to produce bioactive GAs (Toyomasuet al. 1998). In Arabidopsis, Pfr induces GA4 and GA4Hexpression, also GA3b-hydroxylases (Yamaguchi et al.1998). Similarly, in Datura ferox, Pfr stimulates tran-script accumulation of a GA3b-hydroxylase in a wayconsistent with its effect on germination (Burgin et al.2000).
Light-mediated changes in the sensitivity of seeds toGAs are scarcely known. It has been shown that R en-hances the response of germination to GA4 in Arabid-opsis. This effect is mediated neither by phytochrome Anor by phytochrome B, since the WT and both phyto-chrome-deficient simple and double mutants exhibitsimilar light-mediated increments in GAs sensitivity
M. V. Arana Æ L. C. de Miguel Æ R. A. Sanchez (&)IFEVA. Facultad de Agronomıa,Universidad de Buenos Aires and CONICET,Avenida San Martın 4453, 1417 Buenos Aires, ArgentinaE-mail: [email protected].: +54-011-45248071Fax: +54-011-45148730
Planta (2006) 223: 847–857DOI 10.1007/s00425-005-0134-7

(Yang et al. 1995). The photoreceptors involved andboth the site of the changes (e.g., whether they occur inembryos, endosperm, or both) and the biochemical andmolecular bases of the modulation of GAs perceptionare unknown. On the other hand, it should be noted thatin lettuce phytochrome influence on the sensitivity ofgermination to GAs is not as clear as in Arabidopsis(Toyomasu et al. 1998). Therefore, the relationship be-tween the sensitivity of the seeds to GAs and phyto-chrome requires more research.
Several genes participating in the transduction of theGAs signal have been proposed to play regulatory rolesin seeds. SPY is a negative regulator of GAs action and,in contrast with wild-type seeds; spy mutants of Ara-bidopsis are capable of germinating in the presence ofGAs biosynthesis inhibitors (Jacobsen et al. 1996). Therelation between SPY expression and changes in GAssensitivity in seeds, however, needs to be further ex-plored. In monocots, GAMYB is upregulated by GAsand has been implicated in the promotion of amylaseactivity in cereal aleurone layers (Gocal et al. 1999;Gubler et al. 2002). GAMYB also plays roles in transi-tion to flowering as well as in anther development (Gocalet al. 1999; Murray et al. 2003), indicating that this geneis involved in various regulatory processes. GAs actionin the aleurone may be comparable with its action inendosperm softening, since in both circumstances, GAsoperate by inducing the expression of several hydrolyticenzymes including those degrading cell wall polysac-charides. In Arabidopsis there are four GAMYB-relatedgenes (Kranz et al. 1998). Three of them (AtMYB33,AtMYB65 and AtMYB101) are capable of transacti-vating the barley a-amylase promoter and, in contrastwith monocots, only one (AtMYB33) is upregulated byGAs. AtMYB33 is expressed in germinating seeds, and ithas been proposed that AtMYB33 and AtMYB65 haveroles in GAs regulation of germination (Gocal et al.2001), but this idea has not been tested yet. Theexpression of PHOR1, a positive regulator of GAs sig-naling, is diurnally regulated in potato leaves and a peakof PHOR1 mRNA accumulation occurs immediatelyafter the transition to light (Amador et al. 2001). On theother hand, although DELLA genes have been proposedto participate in the control of germination, Bassel et al.(2004) did not find a good relation between DELLAgene expression and induction of germination in Ara-bidopsis, tomato and soybean. It seems likely that thesensitivity of germination to GAs may be affected byphytochrome through the modulation of the expressionof GA-signal transduction genes; however, no informa-tion is available on this subject.
Promotion of germination by phytochrome is com-mon in seeds with coat-imposed dormancy, as is the caseof D. ferox (Soriano et al. 1964; Casal and Sanchez1998), and GAs have an important role in micropylarendosperm (ME) weakening, a necessary step for ger-mination in these seeds (Sanchez and de Miguel 1997).Endosperm hardness is related to the b1-4mannan con-tent of the cell walls, and the softening mediated by
phytochrome includes an increment in the activity oftwo enzymes involved in mannan degradation (endo-b-mannanase and b-mannosidase) (Sanchez et al. 1990; deMiguel and Sanchez 1992) as well as an increment in thetranscripts of DfMan (endo-b-mannanase) and DfExpa1(expansin) (Burgin et al. 2000; Mella et al. 2004). In D.ferox, these light-induced changes in mannan-degradingactivities and ME softening involve embryo–endosperminteractions, in which the embryo, in response to R,provides the GAs required for endosperm softening(Sanchez and de Miguel 1997; Mella et al. 2000). Simi-larly, exogenous GA promotes endo-b-mannanaseactivity as well as LeMAN2 mRNA accumulation in theME of gibberellin-deficient mutant (gib-1) tomato seeds(Lycopersicon esculentum Mill.) (Groot and Karssen1987; Dahal et al. 1997; Nonogaki et al. 2000). Anotherrelevant process related to the germination in seeds withcoat-imposed dormancy is the increment of the expan-sive force of the embryo (Carpita et al. 1979b; Bewley1997). Pfr formation promotes an increment of the em-bryo growth potential (EGP) in D. ferox (de Miguel andSanchez 1992), as well as in lettuce where it is also en-hanced by GAs (Carpita et al. 1979a; Carpita et al.1979b). Potentially, then, there is more than one processthat might be influenced by any effect of the phyto-chromes on the sensitivity to GAs. Since these GAs ac-tions take place in different tissues and the possibilityexists that regulation of GAs signaling may not beidentical in all cells, it seems therefore appropriate tostudy the bases of the changes in sensitivity separately ineach tissue.
In this work we investigated the influence of phyto-chrome on GA responses in the endosperm and theembryo of D. ferox seeds by assessing their relation withthe expression of gene homologs of SPY, GAMYB andPHOR. We also evaluated the role of the embryo, as thesite of light perception, and the influence of exogenouscytokinins as a probable embryonic signal, in the mod-ulation of these GA responses.
Materials and methods
Source of seeds and general incubation conditions
Datura ferox seeds were collected from plants invadingsoybean fields in Monte, province of Buenos Aires,Argentina. After harvest, the seeds were stored in darkglass jars at room temperature until their use in theexperiments.
The seeds were sown on cotton wool saturated withdistilled water in clear plastic boxes, wrapped in blackplastic sheets for 24 h unless otherwise stated, (prein-cubation on water), decoated and incubated on Pac-lobutrazol 4 ppm ((2RS, 3RS-1-(4-clorofenil)-4,4-dimetil-2-(1H1,2,4-Triazol-1-il)pentan-3-ol, CRESTAR,SYNTGENTA), GA3 (SIGMA) with or without theaddition of cytokinins (zeatin 50 lM, SIGMA). Theseeds were then treated with a R or Far Red (FR) pulse.
848

The incubations were performed at alternating temper-atures (9 h 30�C, 15 h 20 �C).
Germination tests
The germination was measured 4 days after light treat-ments. A total of 30 seeds per sample was used in fourindependent experiments. For statistical analysis one-way analysis of variance (ANOVA) of the angulartransformation of the data was used.
EGP measurements
After light treatments, the seeds were incubated indarkness for 44 h and de-tipping was achieved byexcising the conical 0.5 mm micropylar end of the seeds.The de-tipped seeds where then transferred to water andtreated with a FR pulse in order to eliminate the Pfrformed during dissection (Figs. 1, 2a). The EGP wasmeasured after dark incubation for 24 h, as described inde Miguel and Sanchez (1992).
Treatments of seed parts and ME dissection
The experimental procedure for studying the lightinfluence in GA sensitivity in ME is described in theFig. 3a. After preincubation in darkness, the seeds weredecoated under white light, incubated on paclobutrazol
4 ppm and irradiated with a saturating R or FR lightpulse. Twenty-four hours after light treatments, the de-coated seeds were surface-sterilized with NaHClO4
(5.5 g Cl /l ) then washed with distilled water and dis-sected with a surgical blade. The seed portions, 2-mmlength (containing ME, part of lateral endosperm andradicle plus hypocotyls of the embryo), were placed inplastic boxes on cotton saturated with the appropriatehormone solutions. During the dissection the seeds wereexposed to white light, and the seed parts received againthe corresponding light treatments immediately afterthey were exposed to the hormone solutions. The incu-bation continued in darkness (16 h for enzyme activitiesmeasurements or 6 h for RNA analysis) until sampling.
Fig. 1 Phytochrome regulates the response of seed germination toGA in Datura ferox. a Experimental protocol. Seeds were imbibedfor 40 h, de-coated and incubated on Paclobutrazol (Pc) 4 ppm,100 lM GA3 with or without the addition of 50 lM zeatin (ZEA)and immediately treated with a R or FR pulse. After the lighttreatments, seeds were incubated in darkness, at alternatedtemperatures (9 h 30 C, 15 h 20 C). b Germination measured4 days after light treatments. Error bars show the SEM from fourindependent experiments. Percent of seed germination in controls(dashed lines): R H20: 84±5; R Pc GA ZEA: 61±8; FR H20: 5±3;FR Pc ZEA: 0; R Pc ZEA: 0; R Pc: 0
Fig. 2 GA promotes EGP, and R light regulates the response ofthe embryo to GA. a Experimental protocol. Seeds were imbibed indarkness for 24 h, de-coated, incubated on water or solutionscontaining Pc plus different hormones as indicated in b and c andtreated with a R or FR pulse. After 44 h of further incubation indarkness the endosperm cap was removed, the seeds weretransferred to water and treated with a FR pulse. Embryo lengthwas measured 24 h after FR treatment. b, c Embryo lengthsfrequency distribution after different light and hormone treatments.Data are the results of three independent experiments. A total of 25embryos per sample was measured in three independent experi-ments. Pc Paclobutrazol 4 ppm, GA gibberellic acid 100 lM, ZEAzeatin 50 lM, H2O incubation on water
849

In order to evaluate the influence of the embryo inGA sensitivity of ME after preincubation in darknessthe seeds were de-coated under white light, surface-sterilized with NaHClO4 and dissected, as shown inFig. 4a. Removal of the embryo part remaining insidethe dissected section was achieved by applying gentlepressure on the endosperm tip. Seed portions wereexposed to a R pulse and subsequently incubated indarkness for 20 h in cotton saturated with different GA3
concentrations with or without the addition of cytoki-nins (zeatin 50 lM). The ME (0.5 mm) was dissectedand assayed for mannan-degrading activities or RNAanalysis. Paclobutrazol 4 ppm was included in theincubation medium of all seed portions.
DfPHOR, DfSPY and DfMYB partial cDNA isolation
In order to isolate partial PHOR1, SPY and GAMYBcDNA sequences of D. ferox seeds, different sets ofprimers were designed and used for PCR amplificationof cDNA synthesized from total RNA of embryos andMEs.
DfPHOR primers (upper primer: 5¢AGCTCACCGGGATACACAAC3¢, lower primer: 5¢ACTCACAATCGCATCACTCG3¢) were designed from potato PHOR1sequence using Primer3 program (Whitehead Institutefor Biomedical Research, http://frodo.wi.mit.edu/cgi-bin/primer3/primer3) to amplify products of 180 bpexpected size for use in RT-PCR assays. DfSPY primers(upper primer: 5¢CAGGTGACTTGGATTGGATACCC3¢, lower primer: 5¢GCTTCAGGAGAGGGTGTATAAC3¢) and DfMYB primers (upper primer: 5¢ GGGTAACTGGAATGCTGTG3¢, lower primer: 5¢GCCATCTGTGCCCATTTATT3¢) were designed from conservedregions within known SPY and GAMYB genes in otherspecies.
To ensure that authentic PHOR1, SPY and MYB-related transcripts were obtained, the PCR productswere ligated into pGEM-T using the pGEM-T EasyVector System (Promega), and clones were sequenced atFundacion Leloir, University of Buenos Aires, Argen-tina. Homology searches of the databases were per-formed using the BLAST program. Alignments ofnucleotide sequences were carried out using the CLUS-TAL W program.
Enzyme activities measurement and RT-PCR reactions
Endo-b-mannanase and b-mannosidase activities weremeasured as described in Sanchez and de Miguel (1997).
RT-PCR reactions were performed using total RNAextracted from 30 endosperm caps or embryos by theTRIZOL procedure (Gibco). Twelve microliters of RNAsamples (endosperm caps) or 3 lg of RNA (embryos)were subjected to DNAse treatment (DNAseI, RNAse
Fig. 3 GA responses of ME cells is under phytochrome control. aExperimental procedure. Seeds were imbibed in darkness for 24 h,de-coated and treated with a R or FR pulse. After a further 24-hincubation on Pc in darkness the seeds were dissected and the seedportions were transferred to a incubation medium containing100 lM GA3 with or without the addition of 50 lM zeatin,irradiated again with R or FR and incubated for 6 or 16 h beforemRNA analysis or enzyme activities measurements, respectively.Pc 4 ppm was included in the medium of all seed portions. b, cEndo-b-mannanase and b-mannosidase activities in ME, as afunction of external GA3 concentration. d DfExpa1 and DfManRT-PCR band densitometry: means from at least three indepen-dent experiments, normalizing the expression in R-values as 1.Error bars indicate S.E.M. Asterisks in d show significantdifferences with the R plus GA treatment (P<0.05)
850

free, Promega) and then denatured by heating at 65�Cfor 5 min. Then the RNA was converted into cDNAwith 300 U of M-MLV Reverse Transcriptase (Pro-mega), 20 U RNAsin (Promega), 1 mM DDT, 10 mMdNTPs (Promega) and 3.3 lM oligo dT (Biodynamics).The reaction was stopped by heat inactivation at 95�Cfor 5 min. PCR reactions were performed using GoTaqenzyme (Promega) and 2 ll of cDNA as template.Annealing temperatures were: 45�C (DfMan), 57�C(actin, DfPHOR and DfSPY), 60�C (DfExpa) and 42�C(DfMYB). PCR calibration curves with different cyclenumbers were performed for each gene, in order todetermine the number of amplification events that yieldRT-PCR reactions in the lineal phase. The optimal cyclenumber range determined and then used for theRT-PCR reactions were: Actin: 20–23 cycles (embryos);23–25 cycles (MEs), DfMan: 20–23 cycles; DfExpa 25–28cycles, DfPHOR: 23–25 cycles (embryos); 23–27cycles(MEs), DfSPY: 32–36 cycles (embryos) 37–39 cycles(MEs), DfMYB: 30–32 cycles (embryos); 32–34 cycles(MEs). DfMan upper and lower primers were5¢CGCGGATCCACAATTATGGCATGGGA 3¢ and5¢CGCGGATCCATGGATAGTAGCAAAATC 3¢,respectively. The primers used for DfExpa1, and actinamplification were described in Mella et al. (2004),whereas the primers used for DfPHOR, DfSPY andDfMYB amplification were described above.
The PCR products were separated by electrophoresisin agarose gels 2% (w/v), transferred onto nylon mem-branes and probed with the corresponding [32P]dCTPlabeled cDNA fragments. The DNA probes were gen-erated by PCR amplification of the cDNA previouslyligated to pGEM-T vectors (pGEM-T Easy VectorSystem, Promega) and sequenced at Fundacion Leloir,University of Buenos Aires, Argentina. The radioactiveprobes were generated by labeling the PCR productswith the Prime A Gene Labeling System (Promega).Hybridization was carried out at 60�C in UltraHYB(Ambion) hybridization solution for 24 h, and blotswere washed at 60�C two times for 20 min 2X SSC,0.1% SDS; 0.5X SSC, 0.1% SDS and one-time0.1XSSC, 0.1%SDS. Blots were exposed to Phosphorscreen at room temperature for about 4–24 h. The ra-diolabeled signal was detected using the PhosphorIm-ager Storm 840 (Molecular Dynamics). Band intensitieswere analyzed by the Image Quant 5.2 program(Molecular Dynamics). The results are expressed as theratio of DfExpa1, DfMan, DfPHOR, DfMYB or DfSPYto actin and normalized to 1 as indicated in legends. Forstatistical analysis paired t test has been used. Data weretransformed using the arcsine of the square root. Sig-nificant results were taken at P values lower than 0.1 andmeans with differences larger than twofold.
Light sources
Red light (15 lmol m�2 seg�1) was provided by 36-WPhilips fluorescent tubes filtered through a R glass filter
(Acrılicos Anbyn, Bs. As. Argentina). FR light (40 lmolm�2 seg�1) was provided by 150-W incandescent inter-nal reflector lamp filtered through an RG9 Schott glassfilter (Mainz, Germany) and a 10-cm water filter. Thecalculated proportion of Pfr/P was 0.87 and 0.1 for theR and the FR sources, respectively.
Accession numbers
The Gene Bank accession numbers for the DfExpa1 andDfMan cDNAs are AF442773 and AF323949, respec-tively. DfPHOR: AY706991 (embryo sequence) andAY729690 (ME sequence). DfSPY: AY729691 andAY729692 embryo and ME sequence, respectively.DfMYB: AY729693 (embryo sequence) and AY729694(ME sequence).
Results
Pfr enhances the promotion of seed germinationby GA and can be substituted by zeatin
The promotion of germination by phytochrome in sev-eral species such as Arabidopsis thaliana, lettuce and D.ferox is related with an increase in active GAs content(Toyomasu et al. 1993; Sanchez and de Miguel 1997;Yamaguchi et al. 1998; Mella et al. 2000). We exploredhere the possibility that phytochrome can also modulatethe sensitivity of D. ferox seeds to GA3 (GA).
The promotion of germination by exogenous GA waslarger in R than in R followed by FR-treated seeds(Fig. 1); and the increase in sensitivity of germination toGA displayed a typical LFR of phytochrome. Zeatin(ZEA), addition to the incubation medium, enhancedthe GA effect on germination in FR-treated seeds,demonstrating that cytokinins can replace the Pfrrequirement for promotion of the response to GA.When seeds were treated with R, ZEA had no effect inthe stimulus of germination by GA, suggesting anoverlapping of the pathways for R and cytokinins actionin the stimulus of GA response. This suggestion is inagreement with the fact that the effect of R on GApromotion of germination was cancelled when ZEA wassupplied to the incubation medium. Independent of thelight treatment, no germination was observed whenseeds were incubated with ZEA plus paclobutrazol (i.e.,in the absence of either biosynthesis or external supplyof GA) (Fig. 1), suggesting that ZEA does not promotegermination directly, but affects the germination re-sponse by enhancing sensitivity to GA.
Pfr enhances the GA promotion of EGP
Red promotes EGP in D. ferox, in a FR-reversiblefashion and this effect is related to the induction of seedgermination (Fig. 2b; de Miguel and Sanchez 1992).
851
Inhibiting GAs synthesis with paclobutrazol preventedthe Pfr-mediated promotion of EGP (Fig. 2b), dem-onstrating that GAs biosynthesis is necessary for thisPfr effect. As expected, exogenous GA reversed the
effects of paclobutrazol on the promotion of EGP by R(Fig. 2b).
The effect of GA on EGP was larger in R-treatedseeds, in comparison with R followed by FR-treatedseeds, showing that the response of embryo cells to GAis under phytochrome control (Fig. 2c). The inclusion ofZEA in the incubation medium of FR-treated seedsproduced a significant increase in the response to GA,demonstrating that cytokinins enhance the response ofembryo cells to GA. On the other hand, there was noeffect of ZEA when no GA was added to the incubationmedium, and GA synthesis was blocked by pac-lobutrazol, suggesting that ZEA promotes EGP byenhancing the sensitivity of embryo cells to GA.
Phytochrome regulates the response of ME cells to GA
Since phytochrome promotion of germination inD. ferox involves changes in the seed sensitivity to GA(Fig. 1) and, on the other hand, the response of embryocells to GA is light dependent (Fig. 2), we investigatedthe possibility that the response of ME cells to GA isalso under phytochrome control. For this purpose theresponses to GA in terms of mannan-degrading enzymeactivities and DfMan and DfExpa1 transcript levels wereassessed in dissected ME that had been in contact withembryos of seeds treated either with R or FR andincubated on paclobutrazol to prevent GA biosynthesis(Fig. 3a).
The promotion by exogenous GA of endo-b-mannanase activity in the ME was strongly enhanced byR. When seeds were irradiated with R the endo-b-mannanase activity in ME was larger (1.6 times at GA100 lM) than in ME from seeds irradiated with FR(Fig. 3b). Similar results were obtained in the case ofb-mannosidase activity (Fig. 3c). Moreover, the accu-mulation of DfMan and DfExpa1 transcripts in responseto exogenous GA was also strongly promoted by R(Fig. 3d), showing that phytochrome regulates theresponses to GA linked with ME weakening. Actuallythe promotion by R of GAs influence on DfMantranscripts level is somewhat larger than that ofb-mannanase activity, suggesting that phytochromecontrol of the enzyme activity may involve more thanchanges in mRNA amounts. The addition of ZEA to the
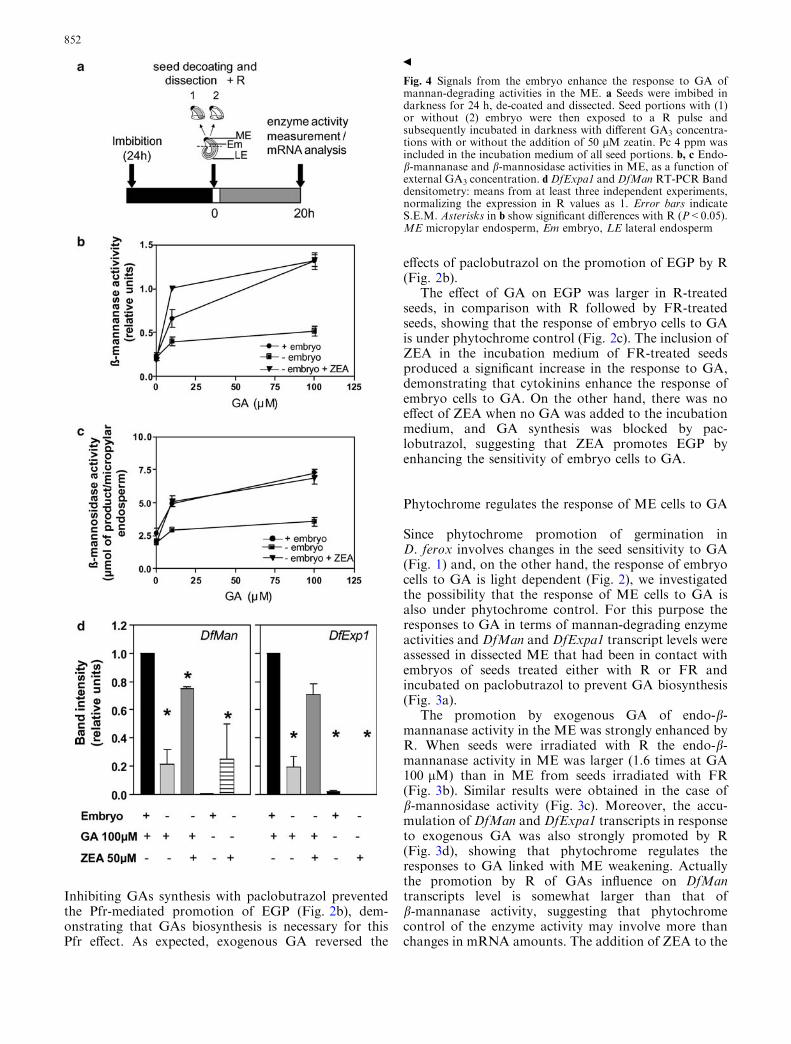
Fig. 4 Signals from the embryo enhance the response to GA ofmannan-degrading activities in the ME. a Seeds were imbibed indarkness for 24 h, de-coated and dissected. Seed portions with (1)or without (2) embryo were then exposed to a R pulse andsubsequently incubated in darkness with different GA3 concentra-tions with or without the addition of 50 lM zeatin. Pc 4 ppm wasincluded in the incubation medium of all seed portions. b, c Endo-b-mannanase and b-mannosidase activities in ME, as a function ofexternal GA3 concentration. d DfExpa1 andDfMan RT-PCR Banddensitometry: means from at least three independent experiments,normalizing the expression in R values as 1. Error bars indicateS.E.M. Asterisks in b show significant differences with R (P<0.05).ME micropylar endosperm, Em embryo, LE lateral endosperm
b
852

incubation medium significantly promoted the stimulusby GA of mannan-degrading enzyme activities and theaccumulation of transcripts of DfMan and DfExpa1 inME from seeds treated with FR (Fig. 3b–d), showingthat ZEA is capable of replacing the R requirements forenhancing responses to GA in ME.
The embryo perceives the light signal that modulatesGA responses in ME
Endosperm weakening is a result of GAs action in theME, and the most likely origin of GAs is the embryofrom where it would move to the ME promoting cellwall-degrading activities (Sanchez and de Miguel 1997).Light had no effect on the GA promotion of endo-b-mannanase activity in isolated endosperms (data notshown). Furthermore, in R treatments, values of endo-b-mannanase and b-mannosidase activities in response toGA were higher in ME incubated in contact with theembryo in comparison with isolated endosperm(Fig. 4b, c), showing that the light signal that modulatesthe changes in GA sensitivity in ME is not perceived by
the ME itself, and that the Pfr formed in the embryostimulates the production of a factor that moves to MEstimulating GA action. In portions from R-treated seedsthe embryo also stimulated the ME responses to GA forDfExpa1 and DfMan1 transcript accumulation(Fig. 4d); moreover, exogenous ZEA could replace theembryo in the promotion of GA action in ME (Fig. 4),suggesting that cytokinins may be components of theembryonic signal that stimulates GA responses in ME.
Relation between DfPHOR, DfSPY and DfMYBexpression in seeds and the changes mediatedby the phytochromes in the sensitivity to GA of MEand embryos
In order to study possible molecular bases of the changesin GA sensitivity in embryos and ME, we investigatedthe influence of phytochrome in the level of transcriptsof components of the GAs transduction pathway inD. ferox seeds. We cloned cDNA fragments with highidentity with PHOR1 (Amador et al. 2001), SPINDLY(Jacobsen et al. 1996) and AtMYB65 (Gubler et al. 1995)
Fig. 5 Phytochrome controlsDfPHOR and DfMYBtranscript levels in ME of D.ferox seeds. DfPHOR, DfSPYand DfMYB RT-PCR banddensitometry of (a) ME andembryos 6 h after lighttreatments or (b) ME of seedportions with or withoutembryo incubated on Pc plus100 lM GA3 with or withoutthe addition of 50 lM zeatin.The experimental protocol for bis described in the Fig. 4a. Errorbars indicate S.E.M. Data aremeans of at least fourindependent experiments,normalizing the expression in R(a) or in seed portions withembryo plus GA (b) as 1.Asterisks show significantdifferences with R (a) or seedportions with embryo plus GA(b) (P<0.1)
853

genes, which were called DfPHOR, DfMYB and DfSPY,respectively.
Pfr formation stimulated DfPHOR and DfMYBtranscripts accumulation in ME, and this effect wasnoted few hours after the light treatments, long beforethe changes in DfMan or DfExpa1. The effect of R wasreversed when a FR pulse immediately followed the Rpulse, displaying a typical LFR of the phytochromes. Inembryos, however, light had no effect on DfPHOR orDfMYB accumulation (Fig. 5a). On the other hand, theLFR mode of the phytochromes did not modulate theaccumulation of DfSPY transcripts, since no differencesin DfSPY transcripts level were seen in either embryosor ME after light treatments (Fig. 5a).
In view that the light signal that stimulates GA re-sponse in ME is perceived by the embryo, we exploredthe possibility that DfPHOR and/or DfMYB transcriptslevels may be related to the embryonic factor that en-hances the sensitivity to GA in ME. As shown inFig. 5b, there were no differences in DfPHOR tran-scripts accumulation between ME from seed portionscontaining the embryo in comparison with those fromseed portions without it. However, when GAs synthesiswas blocked and no exogenous GA was supplied, thelevel of DfPHOR transcripts was low. This indicates thatGA regulates DfPHOR transcripts level in ME andsuggests that phytochrome may control DfPHOR in MEindirectly by affecting the GAs contents in this tissue.
On the other hand, DfMYB transcript accumulationwas larger in ME of seed portions incubated withembryo in comparison with ME of seed portionswithout embryo, and in the last case, the inclusion ofZEA in the incubation of the seed portions replaced theembryo in the stimulus of DfMYB expression in ME.The effect of the embryo or the addition of ZEA wasindependent of the presence of GA (Fig. 5b). Theseresults show that the embryo stimulates the accumu-lation of DfMYB transcripts in ME, and suggest that itmay promote GA sensitivity in ME through changes inDfMYB expression.
Neither the contact with the embryo nor GA supplymodified the transcripts level of DfSPY in ME (Fig. 5b).These results are consistent with the hypothesis that theregulation of DfSPY expression in ME is independent ofphytochrome action.
Discussion
In seeds with coat-imposed dormancy, the induction ofgermination by phytochrome involves coordinatedchanges in the embryo and the micropylar region of theendosperm that allow the embryo to overcome thephysical constraints that limit its growth (Carpita et al.1979b; Psaras et al. 1981; Sanchez and de Miguel 1985;Sanchez and de Miguel 1992; Bewley and Black 1994).
GAs biosynthesis is required for the promotion bylight of both ME weakening (Sanchez and de Miguel1997) and EGP (Fig. 2a). While the reduction in the
mechanical resistance of the ME promoted by GA iswidely recognized as necessary for germination in sev-eral species (Groot and Karssen 1987; Bewley 1997;Chen and Bradfrod 2000) the importance of the increasein EGP is still debated (Mo and Bewley 2003). Ourobservation of the requirement of GA synthesis for Pfreffect on EPG is in agreement with the results of Carpitaet al. (1979b) with lettuce seeds, arguing for a contri-bution of the increase in EGP in the promotion of ger-mination by light. On the other hand, observations inthe mutant gib-1 of tomato (Groot and Karssen 1987)and ga-1 of Arabidopsis (Debeaujon and Koornneef2000), which are impaired in their capacity to synthesizeGAs, show that the deficiency of this hormone does notprevent embryo growth, when relieved from the oppo-sition of the ME. However, it should be noted that thegrowth rate of gib-1 embryos is lower than in wild-type(Groot and Karssen 1987), indicating that GAs stimu-late embryo growth. It is likely then that GAs maycontribute to EGP by allowing the embryo to overcomeinternal growth constraints, i.e., enhancing cell wallextensibility (Chen et al. 2001; Mella et al. 2004), andthat this may facilitate emergence through the coveringtissues.
Phytochrome promotes germination by two differentcontrol pathways: enhancing bioactive GAs synthesis(Toyomasu et al. 1993; Mella et al. 2000) and increasingthe capacity of GA sensing by the seeds (Fig. 1, Yanget al. 1995). Our data show that the changes in the re-sponse to GA of whole seeds are consistent with changesmediated by the phytochromes in the sensitivity to GAin ME and embryos (Figs. 2, 3). This provides the firstevidence that the phytochrome promotes GA responsein two relevant processes for germination: the weakeningof the ME and the stimulus of the EGP. In D. ferox, thephytochrome-dependent signal that modulates the re-sponse to GA in embryos and ME is originated in theembryo, the site where the light stimulus is perceived.This signal has a local effect, upon the embryo cells(Fig. 2) and also diffuses to ME, where promotes theresponse to GA (Fig. 4). The embryonic factor stronglypromotes in the ME the GA stimulus of mannan-degrading activities and the accumulation of DfMan andDfExpa1 transcripts (Fig. 4 and de Miguel et al. 2002).In barley, aleurone cells proximal to the embryo aremore sensitive to GA for vacuolization, induction ofa-amylase activity and b-glucanase secretion than thedistal ones (Ritchie et al. 1999). This differentialresponse suggests the production by the embryo of adiffusible factor establishing a gradient through thealeurone layer, remaining more abundant in the proxi-mal zone. Similarly, in tomato, the contact of the MEwith the embryo enhances the response to GA for thestimulus of mannanase activity (Mo and Bewley 2003).
Our data also suggest that the phytochrome-depen-dent diffusible factor that mediates the communicationbetween the embryo and ME may be a cytokinin, sinceexogenous supply of ZEA replaces the contact with theembryo in the promotion of the responses of the ME to
854

GA (Fig. 4). The suggestion that cytokinins may becomponents of the light signal promoting germination issupported by earlier work with Pinus Sylvestris, were thestimulus of germination by the phytochromes is associ-ated with a strong but transitory increase of isopentenyladenosine content (Qamaruddin and Tillberg 1989).Moreover, exogenous cytokinins supply to lettuce seedslowers the dose of light required for germination (Miller1956) and a mixture of GAs and cytokinins is capable ofreplacing the light requirements for germination incelery (Thomas et al. 1975).
An important aspect of GAs action in germination isthe way in which this hormone is perceived by the seeds.In the last decade, important efforts have been made fordetermining the way in which GAs signal is sensed thatresulted in the identification of several components ofthe GAs signaling in plants (Jacobsen et al. 1996; Gocalet al. 1999; Peng et al. 1999; Amador et al. 2001; Leeet al. 2002; Wen and Chang 2002). Our results show thatthe stimulus of germination by the phytochromes is wellcorrelated with the increment of mRNA accumulationof DfMYB and DfPHOR (Fig. 5a). These genes presenthigh homology to AtMYB65 and PHOR1, which arepositive regulators of GA signaling (Amador et al. 2001;Gocal et al. 2001). Six hours after the R pulse, there is anincrement of transcripts accumulation of DfMYB andDfPHOR in ME in comparison with FR controls. Bythat time, structural changes in ME cells are notdetectable (Mella et al. 1994), and no induction ofmannan-degrading activities by R is still measurable(Sanchez and de Miguel 1997). These results constitutethe first pointing to a correlation between alteration inthe expression of components of the GAs signalingpathway and phytochrome action in seeds, and suggestthat MYB and PHOR-related genes have a role in GA
action on weakening. Although R stimulates bothDfPHOR and DfMYB transcript accumulation in ME,the way in which it is achieved is different for each gene.DfPHOR expression is dependent on GAs, and wepropose that the accumulation of DfPHOR transcripts isstimulated by the increment of bioactive GAs in the MEin response to R. This is supported by the fact thatDfPHOR expression in ME is high whenever GA ispresent in the incubation medium independently of thecontact with the embryo or the supply of ZEA (Fig. 5b).This result is in agreement with reports in potato, whereGA stimulates PHOR1 transcripts level in roots(Amador et al. 2001). On the other hand, DfMYBexpression in ME might be promoted by the cytokininsreleased by the embryo in response to R, consideringthat the contact with the embryo or the supply of ZEApromote DfMYB transcript accumulation (Fig. 5b).This suggests that while DfPHOR expression could berelated to the action of GA in ME, changes in DfMYBmay be implicated in GA perception in this tissue.DfSPY transcript accumulation in ME is not modifiedby light, the embryo or ZEA (Fig. 5). This is consistentwith the observations in Arabidopsis, where no interac-tion between cytokinins and SPY expression has beendetected in germination (Greenboim-Wainberg et al.2005).
The phytochromes also modulate the responses toGA in embryos. However, this event is not related to thechanges in the expression of DfPHOR, DfSPY orDfMYB (Fig. 5a). There are no differences betweentranscript accumulation of these genes 6 h after R, timein which the differences in DfPHOR and DfMYBexpression in ME are visible (Fig. 5), and the same isseen 24 and 44 h after R (data not shown). Althoughposttranscriptional changes cannot be discarded, it islikely that genes different from DfPHOR, DfMYB andDfSPY are related to the changes in GA sensitivity inembryos.
The data presented in this work show that in D. feroxthe control of GA response of germination is modulatedby light perceived by the embryo, and is associated withchanges in GA sensitivity in embryos and ME. AlthoughR enhances the sensitivity to GA in the embryo and theME, the pathways of GA perception are not identical ineach case, since the action of phytochromes involveschanges in the expression of DfMYB and DfPHOR inME cells but not in embryos (Fig. 6). The study of theway in which DfMYB and DfPHOR act in ME as well asthe search of genes that control the response of embryosto GA are interesting topics in the transition fromdormancy to germination triggered by environmentalsignals.
Acknowledgments We thank Dr. Roberto Benech-Arnold for thecritical reading of the manuscript and Ms. Gabriela Auge for hermost valuable technical help. We also appreciate the hospitality ofthe Fundacion Leloir for allowing us the use of laboratory facilitiesand thank Dr Roberto Staneloni for his valuable advice and sup-port. This work was supported by the Agencia Nacional dePromocion Cientıfica y Tecnologica (grant no. 09586 to R.A.S.)
Fig. 6 Proposed model for light stimulus of germination in D.ferox
855

and the University of Buenos Aires (grant no. G073 and G6091 toR.A.S.). M.V.A. was supported by a fellowship from the Universityof Buenos Aires.
References
Amador V, Monte E, Garcıa-Martınez J, Prat S (2001) Gibberellinsignal nuclear import of PHOR1, a photoperiod-responsiveprotein with homology to Drosophila armadillo. Cell 106:343–354
Bassel G, Zielinska E, Mullen R, Bewley D (2004) Down-regula-tion of DELLA genes is not essential for germination oftomato, soybean, and Arabidopsis seeds. Plant Physiol136:2782–2789
Benech-Arnold R, Sanchez R, Forcella F, Kruk B, Ghersa C (2000)Environmental control of dormancy in weed seed banks in soil.Field Crops Res 65:105–122
Bewley D (1997) Seed germination and dormancy. Plant Cell9:1055–1066
Bewley D, Black M (1994) Seeds: Physiology of Development andGermination, Ed 2. Plenum, New York
Burgin M, Perez Flores L, Mella R, Staneloni R, Sanchez R (2000)The transcription of -mannanase and GA 3 -hydroxylase genesof Datura ferox seeds is regulated by phytochrome. In: PlantPhysiology, San Diego, p 124:S146
Carpita N, Nabors M, Ross C, Petretic N (1979a) The growthphysics and water relations of red-light-induced germination inlettuce seeds. Planta (Berl) 144:217–224
Carpita N, Ross C, Nabors M (1979b) The influence of plantgrowth regulators on the growth of the embryonic axes of redand far-red treated lettuce seeds. Planta (Berl) 145:511–516
Casal J, Sanchez R (1998) Phytochromes and seed germination.Seed Sci Res 8:317–329
Chen F, Bradfrod K (2000) Expression of an expansin is associatedwith endosperm weakening during tomato seed germination.Plant Physiol 124:1265–1274
Chen F, Dahal P, Bradford K (2001) Two tomato expansin genesshow divergent expression and localization in embryos duringseed development and germination. Plant Physiol 127:928–936
Dahal P, Nevins D, Bradford K (1997) Relationship of endo-b-mannananse activity and cell wall hydrolysis in tomato endo-sperm to germination rates. Plant Physiol 113:1243–1252
de Miguel L, Arana M, Burgin M, Sanchez R (2002) The responseof ME cells to gibberellin stimulus of ß-mannanase activity andexpansin gene expression is increased by a factor produced bythe embryo in Datura ferox seeds. The International Society forSeed Science (ISSS), Salamanca, p 26
de Miguel L, Sanchez R (1992) Phytochrome-induced germination,endosperm softening and embryo growth potential in Daturaferox seeds: sensitivity to low water potential and time to escapeto FR reversal. J Exp Bot 43:969–974
Debeaujon I, Koornneef M (2000) Gibberellin requirement forArabidopsis seed germination is determined both by testacharacteristics and embryonic abscisic acid. Plant Physiol122:415–424
Gocal G, Poole A, Gubler F, Watts R, Blundell C, King R (1999)Long-day up regulation of a GAMYB gene during Lolium tem-ulentum inflorescence formation. Plant Physiol 119:1271–1278
Gocal G, Sheldon CC, Gubler F, Moritz T, Bagnall DJ, Mac-Millan CP, Li SF, Parish RW, Dennis ES, Weigel D, King RW(2001) GAMYB-like genes, flowering, and gibberellin signalingin Arabidopsis. Plant Physiol 127:1682–1693
Greenboim-Wainberg Y, Maymon I, Borochov R, Alvarez J,Olszewski N, Ori N, Eshed Y, Weiss D (2005) Cross talk be-tween gibberellin and cytokinin: the Arabidopsis GA responseinhibitor SPYNDLY plays a positive role in cytokinin signal-ing. Plant Cell 17:92–102
Groot S, Karssen C (1987) Gibberellins regulate seed germinationin tomato by endosperm weakening: a study with gibberellin-defficient mutants. Planta (Berl) 171:525–531
Gubler F, Chandler P, White R, Llewellyn D, Jacobsen J (2002)Gibberellin signaling in barley aleurone cells. Control of SLN1and GAMYB expression. Plant Physiol 129:191–200
Gubler F, Kalla R, Roberts J, Jacobsen J (1995) Gibberellin-regulated expression of a MYB gene in barley aleurone cells:evidence for MYB transactivation of a high-pl a-amylase genepromoter. Plant Cell 7:1879–1891
Jacobsen S, Binkowski K, Olszewski N (1996) SPINDLY, a tet-ratricopeptide repeat protein involved in gibberellin signaltransduction in Arabidopsis. Proc Natl Acad Sci USA 93:9292–9296
Kamiya Y, Garcıa-Martınez J (1999) Regulation of gibberellinbiosynthesis by light. Curr Opin Plant Biol 2:398–403
Khan A, Goss J, Smith D (1957) Effect of gibberellin on germi-nation of lettuce seeds. Science 125:645–646
Kranz H, Denekamp M, Greco R, Jin H, Leyva A, Meissner RC,Petroni K, Urzainqui A, Bevan M, Martin C, Smeekens S,Tonelli C, Paz-Ares J, Weisshaar B (1998) Towards functionalcharacterisation of the members of the R2R3-MYB gene familyfrom Arabidopsis thaliana. Plant J 16:263–276
Lee S, Cheng H, King K, Wang W, He Y, Hussain A, Lo J,Harberd N, Peng J (2002) Gibberellin regulates Arabidopsisseed germination via RGL2, a GAI/RGA-like gene whoseexpression is up-regulated following imbibition. Gene Dev16:646–658
Mella R, Burgin M, Sanchez R (2004) Expansin gene expression inDatura ferox L. seeds is regulated by the low-fluence response,but not by the high irradiance response, of phytochromes. SeedSci Res 14:61–71
Mella R, Burgin M, Staneloni R, Sanchez R (2000) Expansins geneexpression is regulated by phytochrome during dormancybreakage in Datura ferox L. seeds. Plant Physiol, San Diego,USA, p 124: S152
Mella R, Maldonado S, Sanchez R (1994) Phytochrome-inducedstructural changes and protein degradation prior to radicleprotrusion in Datura ferox seeds. Can J Bot 73:1371–1378
Miller C (1956) Similarity of some kinetin and red light effects.Plant Physiol 31:318–319
Mo B, Bewley D (2003) The relationship between b-mannosidaseand endo-b-mannanase activities in tomato seeds during andfollowing germination: a comparison of seed populations andindividual seeds. J Exp Bot 54:2503–2510
Murray F, Kalla R, Jacobsen J, Gulber F (2003) A role forHvGAMYB in anther development. Plant J 33:481–491
Nonogaki H, Gee O, Bradford K (2000) A germination specificendo-b-mannanase gene is expressed in the micropylar endo-sperm cap of tomato seeds. Plant Physiol 123:1235–1245
Peng J, Richards D, Moritz T, Cano-Delgado A, Harberd N (1999)Extragenic suppressors of the Arabidopsis gaimutation alter thedose-response relationship of diverse gibberellin responses.Plant Physiol 119:1199–1207
Psaras G, Georghiou K, Mitrakos K (1981) Red-light inducedendosperm preparation for radicle protrusion of lettuce em-bryos. Bot Gaz 142:13–18
Qamaruddin M, Tillberg E (1989) Rapid effects of red light on theisopentenyladenosine content in Scots Pine seeds. Plant Physiol91:5–8
Ritchie S, McCubbin A, Ambrose G, Kao T, Gilroy S (1999) Thesensitivity of barley aleurone tissue to gibbeellin is heteroge-neous and may be spatially determined. Plant Physiol 120:361–370
Sanchez R, de Miguel L (1985) The effect of red light, ABA, andK+ on the growth of Datura ferox embryos and their relationswith photocontrol of germination. Bot Gaz 146:472–476
Sanchez R, de Miguel L (1992) Phytochrome-induced germination,endosperm softening and embryo growth potential in Daturaferox seeds: sensitivity to low water potential and time to escapeto FR reversal. J Exp Bot 43:969–974
Sanchez R, de Miguel L (1997) Phytochrome promotion of man-nan-degrading enzyme activities in the micropylar endospermof Datura ferox seeds requieres the presence of the embryo andgibberellin synthesis. Seed Sci Res 7:27–33
856

Sanchez R, Sunell L, Labavitch J, Bonner B (1990) Changes inendosperm cell walls of two Datura species before radicle pro-trusion. Plant Physiol 93:89–97
Soriano A, Sanchez R, Eilberg B (1964) Factors and processes inthe germination of Datura ferox L. Can J Bot 42:1189–1203
Thomas T, Palevich D, Biddington N, Austin R (1975) Growthregulators and the phytochrome-mediated dormancy of celeryseeds. Physiol Plantarum 35:101–106
Toyomasu T, Kawaide H, Mitsuhayi W, Inoue Y, Kamiya Y(1998) Phytochrome regulates gibberellin biosynthesis duringgermination of photoblastic lettuce seeds. Plant Physiol118:1517–1523
Toyomasu T, Tsuji H, Yamane H, Nakayama M, Yamaguchi I,Murofushi N, Takahashi N, Inoue Y (1993) Light effects on
endogenous levels of gibberellins in photoblastic lettuce seeds.J Plant Growth Regul 12:85–90
Wen C, Chang C (2002) Arabidopsis RGL1 encodes a negativeregulator of gibberellin responses. Plant Cell 14:87–100
Yamaguchi S, Smith M, Brown R, Kamiya Y, Sun T (1998) Phy-tochrome regulation and differential expression of gibberellin3b-hydroxylase genes in germinating Arabidopsis seeds. PlantCell 10:2115–2126
Yang Y, Nagatani A, Zhao Y, Kang B, Kendrick R, Kamiya Y(1995) Effects of gibberellins on seed germination of phyto-chrome-deficient mutants of Arabidopsis thaliana. Plant CellPhysiol 36:1205–1211
857




















