
Topics in Microfinance and Behavioural Economics - CiteSeerX

J Neural Transm [P-D Sect] (1994) 8:139-148 Journal of
Neural Transmission
�9 Springer-Verlag 1994 Printed in Austria
A novel anticholinesterase THB013:
biochemical and behavioural studies
Short Communication
A. Adem, A. H. Mohammed, B. Winblad, and B. E. Henriksson
Department of Geriatric Medicine, Karolinska Institute, Huddinge University Hospital, Huddinge, Sweden
Accepted July 2, 1994
Summary. Clinical trials with tacrine (THA) have resulted in elevations of liver enzymes in Alzheimer patients that showed improvement. In an effort to mini- mize these side effects several THA analogues were synthesized. These ana- logues were compared to THA in biochemical as well as behavioural studies. In this study, the biochemical effects of THA and one of these analogs, THB 013, on plasma cholinesterase activity, cholinergic receptors as well as the ef- fect of these drugs on spatial learning in adult rats were examined. THB 013 was, at lower concentration, more efficacious in inhibiting plasma cholinest- erase as well as blocking the scopolamine induced disruption of spatial learn- ing when administered 10 min before the scopolamine injection. It is possible that THB 013 with more potent cholinergic effects than THA might be useful in the treatment of Alzheimer's disease.
Key words" Alzheimer's disease, acetylcholine release, cholinesterases, cholin- ergic, muscarinic, spatial learning, tacrine, tacrine analogues, THB 013.
Introduction
Dementia is one of the most common mental health problems in the aged population. Clinically it is characterized by memory loss associated with im- pairment in abstract thinking and judgement, disturbances in higher cortical functions, and personality change. Alzheimer's disease/senile dementia of Alzheimer type (AD/SDAT) accounts for approximately 40-60% of all cases of dementia. Biochemical changes in brains of patients with AD/SDAT in- clude reduced activity of choline acetyltransferase (CHAT) and acetylcho- linesterase (ACHE), indicating reduced activity in the cholinergic system. It is well established that pharmacological manipulations of the brain cholinergic systems affects learning and memory. Several studies have demonstrated that

140 A. Adem et al.
the muscarinic receptor antagonist scopolamine disrupts the performance of several learning tasks in rats, monkeys and humans (Bartus and Johnson, 1976; Petersen, 1977; Sutherland et al., 1982; Hagan and Morris, 1988). In studies on passive avoidance learning in mice it has been consistently shown that scopolamine t rea tment causes impairment in this type of learning. When the drug has been given during pretraining (acquisition), or post-training (consolidation) sessions, it has been found that scopolamine exerts its delete- rious effects primarily during the acquisition phase of learning (Worsham and Hamilton, 1973; Cheal, 1981). In rats scopolamine disrupts spatial learning in the radial arm maze and the Morris water maze (Stevens, 1981; Watts et al., 1981; Sutherland et al., 1982; Whishaw, 1985). Treatment with the cholinest- erase inhibitor T H A has been found to improve cognitive function in aged rats in the radial arm maze (Hodzes et al., 1990) and the Morris water maze (Mohammed, 1990). Moreover , T H A has been found to have multiple effects on the cholinergic system (Adem et al., 1990). We have therefore started to test a number of T H A analogs with the aim of obtaining drugs with more selective effects on the cholinergic system and less toxic effects than has been found with THA. We report here the neurochemical effects of one of the analogs, THB 013, and compared its effects to those of THA. Moreover , we examined whether THB 013 was more effective than T H A in blocking scopo- lamine induced disruption of spatial learning in rats.
Materials and methods
Male Sprague-Dawley rats (Alab, Sollentuna, Sweden) weighing 300-350 grams were used. They were housed under controlled laboratory environment with a 12 h on/12 h off lighting schedule (lights on at 6.00 h) in groups of four in standard laboratory cages with ad lib access to food and water.
Biochemical studies
Receptor binding assay
In displacement studies rat brain tissue homogenates were incubated with increasing concentrations of THA or THB 013 (10 11-10 4M) and 3H-quinuclidinyl benzilate (3H- QNB, 0.22 nM, 39.4 Ci/mmol, NEN) or 3H-pirenzepine (3H-PZ, 3 nM, 76Ci/mmol, NEN) or ~H-oxotremorine-M (3H-OXO-M, i nM, 87 Ci/mmol, NEN) in a final volume of i ml 50 mM Na-K phosphate buffer (pH = 7.4). After incubation for 90 minutes at 25~ the mixture was filtered through Whatman GF/B filters pretreated with 0.05% polyethylene- imine (PEI). The filters were washed three times with 3 ml of ice cold 50 mM Na-K phosphate buffer, pH = 7.4. The filters were left to dry, placed in scintillation tubes and 5 ml ready protein (Beckman) liquid was added, and the bound radioactivity counted in a Beckman scintillation counter. All assays were performed in triplicates. Atropine (10 .4 M) or pirenzepine (10 4 M) or oxotremorine (10 -4M) was used to define non-specific binding.
Measurement of cholinesterase activity
Cholinesterase (acetylcholinesterase or plasma cholinesterase) activity was measured as described before (Adem et al., 1990). Briefly, human plasma was diluted 100 times in 10 mM Na-K phosphate buffer (pH = 7.4) and aliquots (800 ul) were taken for measure- ment of cholinesterase activity using acetylthiocholine (ATC) or butyrylthiocholine

A novel anticholinesterase THB 013 141
(BTC) as substrates. Different concentrations of T H A (10-1~ -4 M) were used to inhibit the plasma cholinesterase activity. The inhibition for each concentration of inhibitor was calculated as a percent of the esterase activity in the absence of inhibitor. ICs0 values were determined f rom curves of percent inhibition versus inhibitor concentration.
Behavioural tests
Acquisition of spatial learning was evaluated in the Morris water maze. The apparatus consisted of a gray-painted circular pool, 140 cm in diameter and 70 cm deep (Mohammed et al., 1990). It was filled 40 cm high with water maintained at 24 _+ 1~ A transparent platform was placed in one location in the pool and the platform was submerged 1 cm below the water surface. The swim paths of the animal was moni tored by a v ideocamera linked to a computer. The computer calculated the latency to locate platform, distance swam and speed.
During testing the rats were gently placed in the pool with their face pointing towards the wall of the pool and allowed 60 seconds to locate the platform. If the animals failed to locate the pla t form they were placed on the platform and allowed to remain there for 30 seconds before being tested for another trial. The latency value of 65 seconds was given for an unsuccessful trial. The rats were given 4 trials daily on 4 consecutive days according to the following drug t rea tment schedules:
Table L Exper imenta l conditions of experiments I and II. Each training group consisted of 7 rats (n = 84). Saline and scopolamine (0.2 mg/kg) were injected 20 min before water maze training and T H A (2.0 mg/kg) and THB 013 (1.5 rag/ kg) 10 min before the injections of saline/scopolamine. Drugs given post-session were administered immediately after the
last trial
Experiment I Saline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Training Scopolamine . . . . . . . . . . . . . . . . . . . . . . . . . . " T H A . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . " THB 013 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . " T H A + Scopolamine . . . . . . . . . . . . . . . " T H B 013 + Scopolamine . . . . . . . . . . "
Experiment H Saline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Training . . . . . . . . . . Saline Saline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . " . . . . . . . . . . T H A Saline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . " - . . . . . . . . . THB 013 Scopolamine . . . . . . . . . . . . . . . . . . . . . . . . . . " - . . . . . . . . . Saline Scopolamine . . . . . . . . . . . . . . . . . . . . . . . . . . " - . . . . . . . . . T H A Scopolamine . . . . . . . . . . . . . . . . . . . . . . . . . . " . . . . . . . . . . T H B 013
E x p e r i m e n t I
Injections were given subcutaneously Scopolamine hydrobromide (Scop) was dissolved in saline and injected at a dose of 0.2 mg/kg 20 rain before training in the water maze. A drop of alcohol was added to T H A and T H B 013 before dissolving the substances in saline. When drug combinations were administered, T H A (2.0 mg/kg) and T H B 013 (1.5 mg/kg) were injected 10min before the injections of Scop.

142 A. Adem et al.
Experiment II
In this experiment Scop or saline was administered as in experiment I but THA or THB 013 was injected immediately after each training session. For the experiment conditions see Table 1.
Statistics
The behavioural data (latency to find platform) data was analysed by two way ANOVA, followed by Student t-test.
Results
In-vitro, T H A and its analogue THB 013 inhibit human plasma cholinesterase (Table 2). Our results indicate that THB 013 is about four times more potent than T H A in inhibiting plasma cholinesterase activity when ATC was used as a substrate.
The ability of T H A and its analogue THB 013 to displace a number of muscarinic radioligands from muscarinic receptors in rat cortex was examined (Table 2). Our results indicate that both T H A and THB 013 have similar affinities for the non-selective muscarinic antagonist 3H-QNB and the M1 selective muscarinic antagonist pirenzepine. However , T H A showed about three times higher affinity for the muscarinic agonist ~H-OXO-M binding sites.
Table 2. ICs0 (gM) values of THA and THB 013 in inhibiting human plasma cholinesterase and muscarinic ligands
ICso values (btM)
Plasmacholinesterase inhibition Muscarinic receptor binding
ATC BTC 3H-QNB 3H-PZ 3H-OXO-M
THA 0.03 • 0.001 0.04 + 0.002 6 -+ 0.2 3 _+ 0.2 0.6 +_ 0.2 THB 013 0.008 _+ 0.0001 0.01 + 0.001 3 • 0.1 4 + 0.2 2 _+ 0.2
In the behavioural studies, statistical analysis of latency data revealed a significant group effect (p < 0.05). Post hoc tests showed that animals t reated with scopolamine had longer escape latencies than saline t reated animals on Day 1 (Fig. 1). When the drugs (THA or THB 013) were given alone and compared to saline t rea tment no difference in the effects on learning were observed. However , when scopolamine was injected in T H A or THB 013 pre t rea ted animals there was a significant effect (p < 0.05) on learning only on the THB 013 pret reated rats (Fig. 1). Inspection of Fig. I shows that on days 2 and 3 pre t rea tment with THB 013 not only counteracted the deleterious effect of scopolamine but caused a dramatic improvement (p < 0.01) in the maze task. A similar pat tern of results was obtained when distance swam was analysed (data not shown).
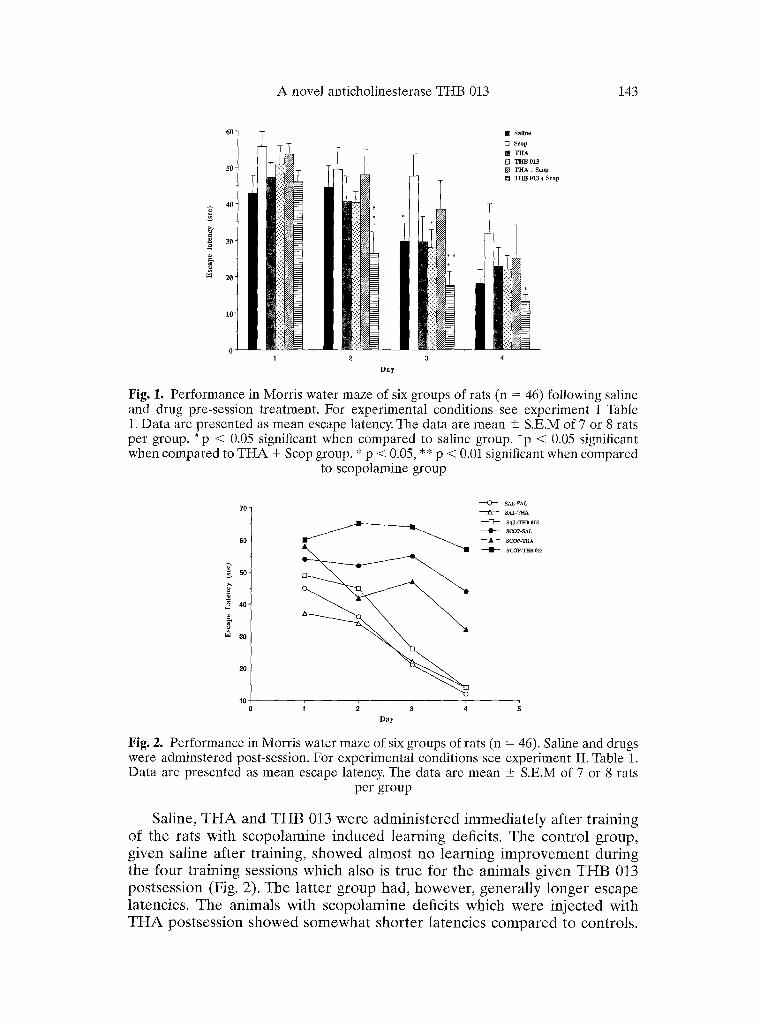
A novel anticholinesterase THB 013 143
50
i
4O
3 0 "
2oJ
10
fiii Iili
iiii
;:;: ==
2
ilii
i!ii
2
D a y
3 4
�9 SaliBe
D Stop
op Scop
Fig. 1. Performance in Morris water maze of six groups of rats (n = 46) following saline and drug pre-session treatment. For experimental conditions see experiment I Table 1. Data are presented as mean escape latency. The data are mean + S.E.M of 7 or 8 rats per group. #p < 0.05 significant when compared to saline group. *p < 0.05 significant when compared to THA + Scop group. * p < 0.05, ** p < 0.01 significant when compared
to scopolamine group
70-
60-
50-
40 -
30 -
20-
10
- - 'O- - S,kL-SAL
SAI~THA @ SAL-THB 013
SCOP-SAL
~ SCOP-THA
SCOP-TBB 013
i i ; i ; D a y
Fig, 2. Performance in Morris water maze of six groups of rats (n = 46). Saline and drugs were adminstered post-session. For experimental conditions see experiment II, Table 1. Data are presented as mean escape latency. The data are mean _+ S.E.M of 7 or 8 rats
per group
Saline, T H A and T H B 013 were adminis tered immedia te ly after training of the rats with s c o p o l a m i n e induced learning deficits. The control group, g iven saline after training, s h o w e d a lmost no learning i m p r o v e m e n t during the four training sess ions which also is true for the animals g iven T H B 013 p os t s e s s ion (Fig. 2). The latter group had, however , general ly longer escape latencies. The animals with s c o p o l a m i n e deficits which were injected with T H A pos t se s s ion s h o w e d s o m e w h a t shorter latencies c o m p a r e d to controls.

144 A. Adem et al.
However, the differences between the three scopolamine pretreated groups were not on any occasion significant (Fig. 2). All the three groups of rats which were administered saline before water maze training and then treated with saline, THA or THB 013 showed a fast, almost identical learning improve- ment during the four training sessions (Fig. 2). At day 3 and 4 there was a significant difference (p < 0.05) in the performance of the latter groups compared to that of the scopolamine pretreated ones with exception for the group injected with THA post-session.
Discussion
The cholinergic hypothesis that a defect in the cholinergic system is involved in AD/SDAT has wide acceptance in terms of a therapeutic strategy. Al- though most of these therapeutic strategies for AD/SDAT have been dis- appointing, the most promising results in clinical drug trails have been ob- tained using anticholinesterases like physostigmine. However, the use of physostigmine in AD/SDAT treatment is limited by the extremely short half- life (approximately 30 minutes) and narrow therapeutic window. The cholin- ergic hypothesis has gained momentum since the anticholinesterase THA in combination with lecithin was reported to improve the memory of Alzheimer's disease patients (Summers et al., 1986). Several clinical trials with THA have resulted in elevations of liver enzymes in those patients that showed improvement. In an effort to minimize these side effects several THA analogues were synthesized. In this study the effects of THA and one of its analogues. THB 013, on plasma cholinesterase and cholinergic receptors as well as the effects of these drugs on spatial learning was studied.
Our results indicate that THB 013 is about four times more potent than THA in inhibiting plasma cholinesterase activity when ATC was used as a substrate. Although the clinical relevance of the inhibition of plasma cho- linesterase is difficult to speculate, the inhibition of brain acetylcholinesterase may be of clinical relevance since THA or THB 013 by inhibition of brain acetylcholinesterase permits acetylcholine to diffuse from intact cholinergic synapses to cells which have been cholinergically denervated and thereby increase cholinergic activity. If one considers inhibition of cholinesterases as one mechanism of action of THA in AD/SDAT, as was suggested previously (Summers et al., 1986), it could be speculated that analogues with higher affinity for inhibiting cholinesterases would give similar effects to THA with less toxicity.
Both THA and THB 013 have similar affinities for the non-selective muscarinic antagonist 3H-QNB and the M1 selective muscarinic antagonist pirenzepine. However, THA showed about three times higher affinity for the muscarinic agonist 3H-OXO-M binding sites. Nielsen et al. (1989) reported that THA is concentrated 10-fold in the brain, probably because of its high lipid solubility. Thus, during treatment with THA high levels of the drug may be attained in the brain which may interact with cholinergic receptors. It has been suggested that the effects of THA, in AD/SDAT, may be partly medi-

A novel anticholinesterase THB 013 145
ated via reversible and competetive interaction with muscarinic cholinergic receptors (Adem et al., 1990). Thus THB 013 may have similar effects like THA on muscarinic receptors.
Tacrine inhibits the release of ACh from rat brain slices (Drukarch et al., 1987; Loiacono and Mitchelson, 1990). When the effects of increasing concen- tration (1 to 100 mM) of THA and THB 013 on 3H-ACh release from rat cortical slices were compared, a dose dependent inhibition of 3H-ACh release in the case of both THA and THB 013 were obtained (Hfikanson, personal communication). Moreover, according to HSkanson, THB 013 inhibited the 3H-ACh release at a 100 times lower concentration compared to THA. Mus- carinic receptors present in the CNS that are able to facilitate and block the release of ACh have been suggested to be of the M1 and M2 receptor subtype respectively (for review see Hfikanson, 1990). It can be speculated that the potent inhibition of 3H-ACh release by THB 013 compared to THA could be due to its potent acetylcholinesterase inhibition. Thus by inhibiting acetylcholinesterase the excess acetylcholine in the synapse blocks the M1 and/or activate the M2 muscarinic autoreceptors. However, the direct effects of THA and THB 013 on presynaptic M1 (block) and/or M2 (stimulate) receptors can not be excluded.
In AD/SDAT brain tissue, a significant increase in the release of 3H-ACh to control levels was observed in the presence of THA, AFDX 116 or atro- pine. However, in the presence of both THA and atropine or THA and pirenzepine but not THA and AFDX 116 the ACh release from Alzheimer cortical slices was inhibited, thus indicating the increased release of ACh in Alzheimer cortical slices may be mediated via muscarinic M1 receptors (Hfikanson, 1990). It can be speculated that THB 013 would give similar results to that of THA at a much lower concentration.
Scopolamine generally impairs behaviour by blocking postsynaptic mus- carinic receptors and decreases brain ACh by blocking presynaptic muscarinic receptors regulating the release of ACh (Hadfizy and Szerb, 1977). Adminis- tration of scopolamine has also, more specifically, been found to disrupt acquisition in a wider variety of learning tests. In the present study pretreat- ment of rats with scopolamine greatly impaired acquisition of the water maze task. Central cholinergic blockade by atropine sulphate has also been shown to impair place navigation in the water maze (Whishaw, 1987), and hence it was argued that the deficits in maze learning could be attributed to impaired sensorimotor functions. It is known that scopolamine also affects sensory function (For review see Hagan and Morris, 1989). It has been found that scopolamine, as in the case with natural aging, acts mainly on short term memory (Beatty and Bierley, 1985) as well as other aspects of the learning and memory process i.e. attention and motor activity. Although the direct effects of scopolamine on the processes mentioned above still are unclear, the scopo- lamine-induced state is generally considered as a relevant animal model for dementia especially Alzheimer's disease.
Our results show that the scopolamine-induced deficits in spatial learning were eliminated by THB 013 if the drug was adminstered before the training sessions in the water maze. Even compared to the control group THB 013

146 A. Adem et al.
significantly improved performance in rats with scopolamine-induced learning deficits. The facilitaing effects on the scopolamine treated animals was evident from the second day and can not be attributed to any differences in base line behaviour of the animals before the start of the spatial training. In experiment II, THA and THB 013 were administered immediately after training in the water maze and eventual improvements in performance are consequently to be related to the consolidation of the acquired information. This is in contrast to the case in experiment I where the phase of acquisition is affected by the drugs. However, the results from the experiment II indicate only a modest and non-significant improvement over time in the scopolamine animals which were iniected with THA or its analogue THB 013 after the last trial of the daily training sessions. THA post-session treated rats show, however, some- what shorter escape latencies during day 2 to 4 compared to THB 013 treated but the difference between the groups was not significant. The three groups of control rats which were treated post-session with THA, THB 013 and saline showed almost identical learning curves with very short escape latencies on the last day of water maze training. The absence of facilitating effects on the behaviour of scopolamine rats when post-session treated by THA and THB 013 might depend upon the possibility that these drugs to a very little or no extent have effect upon the memory formation of the spatial learning task. Another explanation which can not be excluded is the fact that the most intensive part of the consolidation phase is not influenced by the post-session drug treatment at all since the subcutaneous injected THA (Sherman and Messamore, 1988) and THB 013 require quite a long time until peak effects are reached.
In conclusion, our results demonstrate that under the given experimental conditions pre-session adminstration of THB 013 not only eliminates the scopolamine-induced deficits in spatial learning in adult rats but improves learning significantly in these rats compared to controls. As has been men- tioned earlier it is quite possible that scopolamine-induced deficits in spatial learning in rats could be due, in part, to changes in attention processes. An explanation of our results with emphasis upon attention will not in any way reduce the potentials of THA and its analogue THB 013 to be tested clinically in the treatment of neurodegenerative diseases such as Alzheimer's disease. We have previously suggested that the clinical efficacy of THA may be due in part to its multiple effects on the cholinergic system (Adem et al., 1990). However, clinical trails with THA have resulted in elevations of liver enzymes in Alzheimer patients that showed improvement. It is possible that THB 013, which has been found to be four times more potent in inhibiting acetylcho- linesterase compared to THA, can be useful in the treatment of Alzheimer's disease. Furthermore, it might be clinically administered in dosages which will minimize or at least reduce the negative side effects seen in the therapeutic use of THA.
Acknowledgements This study was supported by research grants from Konung Gustaf V:s och Drottning Victorias Stiftelse, Axel och Margareta Ax: son Johnsons Foundation, Stiftelse f6r Gamla

A novel anticholinesterase THB 013 147
Tjfinarinnor, Hans and Loo Osterman's Foundation, KI fonder, Stiftelsen Sigurd and Elsa Goljes Minne, and the Swedish Medical Research Council.
References
Adem A, Mohammed AK, Winblad B (1990a) Multiple effects of tetrahydro- aminoacridine on the cholinergic system: biochemical and behavioural aspects. J Neural Transm [P-D Sect] 2:113-128
Bartus RT, Johnstone HR (1976) Short term memory in the rhesus monkeys: disruption from the anti-cholinergic scopolamine. Pharmacol Biochem Behav 5:39-46
Beatty WW, Bierley RA (1985) Scopolamine degrades spatial working memory but spares spatial reference memory: dissimilarity of anticholinergic effect and restriction of distal visual cues. Pharmacol Biochem Behav 23:1-6
Cheal ML (1981) Scopolamine disrupts maintenance of attention rather than memory processes. Behav Neural Biol 33:163-187
Drukarch B, Kits KS, Van der Meer EG, Lodder JC, Stool JC (1987) 9-Amino-l,2,3,4- tetrahydroacridine (THA), an alleged drug for the treatment of Alzheimer's disease, inhibits acetylcholinesterase activity and slow outward K s current. Eur J Pharmacol 141:153-157
Had~zy R Szerb J (1977) The effect of cholinergic drugs on 3H-acetyl-choline release from slices of hippocampus, striatum and cortex. Brain Res 123:311-322
Hagan JJ, Morris REM (1988) The cholinergic hyphothesis of memory: a review of animal experiments. In: Iversen LL, Iversen SD, Snyder SH (eds) Handbook of psychopharmacology, vol 20. Plenum Press, New York
Hagan JJ, Morris REM (1989) The cholinergic hyphothesis of memory: a review of animal experiments. In: Iversen LL, Snyder SH (eds) Psychopharmacology of the aging nervous system. Plenum Press, New York, pp 237-323
Hodzes H, Ribeiro AM, Gray G, Marcheanks RM (1990) Low dose tetra- hydroaminoacridine (THA) improves cognitive function but does not affect brain acetyl-choline in rats. Pharmacol Biochem Behav 36:291-298
Hfikanson L (1990) Modulation of acetylcholine release in brain. Acta Univ Ups 72:52 Loiacono RE, Mitchelson F (1990) Effects of nicotine and facrine on acetylcholine
release from rat cerebral cortical slices. Naunyn Schmiedebergs Arch Pharmacol 342:31-35
Mohammed AK, Wahlstr6m G, Archer T, Nordberg A (1990) Learning deficits in aged rats pretreated chronically with barbital and tested late in abstinence: alleviation by tetrahydroaminoacridine. J Neural Transm [P-D Sect] 2:285-294
Nielsen JA, Mena EE, Williams IH, Nocerini MR, Liston D (1989) Correlation of brain levels of 9-amiono-l,2,3,4-tetrahydroaminoacridine (THA) with neurochemical and behavioural changes. Eur J Pharmacol 173:53-64
Petersen R (1977) Scopolamine induced learning failures in man. Psychopharmacology 52:283-289
Sherman KA, Messamore E (1988) Blood cholinesterase inhibition as a guide to the efficacy of putative therapies of Alzheimer's dementia: comparison of tacrine and physostigmine. In: Giacobini E, Becker RE (eds) Current research in Alzheimer therapy. Taylor and Francis, New York, pp 73-86
Stevens R (1981) Scopolamine impairs spatial maze performance in rats. Physiol Behav 27:385-386
Summers WK, Majorski LV, Marsh GM, Tachiki K, Kling A (1986) Oral tetrahydroaminoacridine in long term treatment of senile dementia, Alzheimer type. N Engl J Med 315:1241-1245
Sutherland RJ, Wishaw IQ, Regehi JC (1982) Cholinergic receptor blockade impairs spatial localization by use of distal cues in the rat. J Comp Physiol Psychol 96: 563- 573
Watts J, Stevens R, Robinson C (1981) Effects of scopolamine on radial maze perfor- mance in rats. Physiol Behav 26:845-851

148 A. Adem et al.: A novel anticholinesterase THB 013
Wishaw IQ (1985) Cholinergic receptor blockade in the rat impairs locale but not taxon strategies for place navigation in a swimming pool. Behav Neurosci 99: 979- 1005
Wishaw IQ, Tomie JA (1987) Cholinergic receptor blockade produce impairment in a sensorimotor subsystems for place navigation in the rat: evidence from sensory, motor, and acquisition tests in a swimming pool. Behav Neurosci 101:603-616
Worsham E, Hamilton LW (1973) Acquisition and retention of avoidance behaviours following septal lesions and scopolamine injections in rats. Physiol Psychol 1:219-266
Authors' address: A. Adem, Ph.D., Karolinska Institute, Department of Geriatric Medicine, B56, Huddinge Hospital, S-141 86 Huddinge, Sweden.
Received November 16, 1993
Copyright © 2022 FDOKUMEN




















