
A new species of Oreobates (Anura: Strabomantidae) from the Andes of northern Argentina
10
Accepted by M. Vences: 16 Sept. 2008; published: 17 Oct. 2008 41 ZOOTAXA ISSN 1175-5326 (print edition) ISSN 1175-5334 (online edition) Copyright © 2008 · Magnolia Press Zootaxa 1908: 41–50 (2008) www.mapress.com/ zootaxa/ A new species of Oreobates (Anura: Strabomantidae) from the Andes of northern Argentina MARCOS VAIRA 1,3 & LILIANA FERRARI 2 1 Conicet-CIBA, Universidad Nacional de Jujuy, Gorriti 237, 4600. Jujuy, Argentina. E-mail: [email protected] 2 Instituto de Herpetología, Fundación Miguel Lillo, Miguel Lillo 251, 4000. Tucumán, Argentina. E-mail: [email protected] 3 Corresponding author Abstract Oreobates barituensis sp. nov. is described from the ecoregion of southern Andean Yungas forests of north-western Argentina at elevations of 1100 to 1300 m above see level. It shares morphological characters with the sympatric and syntopic O. discoidalis, as well as with O. ibischi and O. cruralis from Bolivia. The recognition of the new species is in particular supported by considerable differences in the advertisement call among these species. We also made some con- siderations on probable conspecificity of northern Argentina and Bolivian populations of O. discoidalis and O. cf. crura- lis. Key words: Anura, Strabomantidae, Oreobates, systematics, bioacustics, Andes, Yungas, Argentina Resumen Se describe una nueva especie, Oreobates barituensis sp. nov., de la Ecorregión de las Yungas Andinas del Sur en el noroeste de Argentina a elevaciones entre los 1000 a 1300 m. La nueva especie comparte caracteres morfológicos tanto con la especie simpátrica y sintópica O. discoidalis, así como con O. ibischi y O. cruralis de Bolivia. El reconocimiento de la nueva especie se confirma especialmente por las considerables diferencias en el canto de anuncio entre las especies. Asimismo, realizamos algunas consideraciones sobre la probable co-especificidad de poblaciones de O. discoidalis y O. cf. cruralis del norte argentino y de Bolivia. Palabras claves: Anura, Strabomantidae, Oreobates, Sistemática, Bioacústica, Andes, Yungas, Argentina Introduction Recent works reviewed the highly diverse Neotropical genus Eleutherodactylus and proposed substantial changes of its systematics (Heinicke et al. 2007; Hedges et al. 2008). The authors, based on extensive DNA sequence analysis and morphological evidence, proposed a new taxon, Terrarana. It now comprises four fam- ilies formerly included in the species-rich family Brachycephalidae of Frost et al. (2006). This new arrange- ment resolved the relationships of many South American species by including them in the family Strabomantidae, consisting of 16 genera and 530 species, most of them occurring in the northwestern Andes. Now, several former members of the genera Craugastor , Eleutherodactylus, Ischnocnema, and Phrynopus are placed in the monophyletic genus Oreobates. The genus, as reviewed and redefined by Padial et al. (2008a) and Hedges et al. (2008), include fifteen species known from Colombia, Ecuador, Peru, Brazil, Bolivia and
-
Upload
universidadnacionaldejujuyconicet -
Category
Documents
-
view
5 -
download
0
Transcript of A new species of Oreobates (Anura: Strabomantidae) from the Andes of northern Argentina

Accepted by M. Vences: 16 Sept. 2008; published: 17 Oct. 2008 41
ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2008 · Magnolia Press
Zootaxa 1908: 41–50 (2008) www.mapress.com/zootaxa/
A new species of Oreobates (Anura: Strabomantidae) from the Andes of northern Argentina
MARCOS VAIRA1,3 & LILIANA FERRARI2
1Conicet-CIBA, Universidad Nacional de Jujuy, Gorriti 237, 4600. Jujuy, Argentina. E-mail: [email protected] Instituto de Herpetología, Fundación Miguel Lillo, Miguel Lillo 251, 4000. Tucumán, Argentina. E-mail: [email protected] author
Abstract
Oreobates barituensis sp. nov. is described from the ecoregion of southern Andean Yungas forests of north-westernArgentina at elevations of 1100 to 1300 m above see level. It shares morphological characters with the sympatric andsyntopic O. discoidalis, as well as with O. ibischi and O. cruralis from Bolivia. The recognition of the new species is inparticular supported by considerable differences in the advertisement call among these species. We also made some con-siderations on probable conspecificity of northern Argentina and Bolivian populations of O. discoidalis and O. cf. crura-lis.
Key words: Anura, Strabomantidae, Oreobates, systematics, bioacustics, Andes, Yungas, Argentina
Resumen
Se describe una nueva especie, Oreobates barituensis sp. nov., de la Ecorregión de las Yungas Andinas del Sur en elnoroeste de Argentina a elevaciones entre los 1000 a 1300 m. La nueva especie comparte caracteres morfológicos tantocon la especie simpátrica y sintópica O. discoidalis, así como con O. ibischi y O. cruralis de Bolivia. El reconocimientode la nueva especie se confirma especialmente por las considerables diferencias en el canto de anuncio entre las especies.Asimismo, realizamos algunas consideraciones sobre la probable co-especificidad de poblaciones de O. discoidalis y O.cf. cruralis del norte argentino y de Bolivia.
Palabras claves: Anura, Strabomantidae, Oreobates, Sistemática, Bioacústica, Andes, Yungas, Argentina
Introduction
Recent works reviewed the highly diverse Neotropical genus Eleutherodactylus and proposed substantialchanges of its systematics (Heinicke et al. 2007; Hedges et al. 2008). The authors, based on extensive DNAsequence analysis and morphological evidence, proposed a new taxon, Terrarana. It now comprises four fam-ilies formerly included in the species-rich family Brachycephalidae of Frost et al. (2006). This new arrange-ment resolved the relationships of many South American species by including them in the familyStrabomantidae, consisting of 16 genera and 530 species, most of them occurring in the northwestern Andes.Now, several former members of the genera Craugastor, Eleutherodactylus, Ischnocnema, and Phrynopus areplaced in the monophyletic genus Oreobates. The genus, as reviewed and redefined by Padial et al. (2008a)and Hedges et al. (2008), include fifteen species known from Colombia, Ecuador, Peru, Brazil, Bolivia and

VAIRA & FERRARI42 · Zootaxa 1908 © 2008 Magnolia Press
Argentina: O. choristolemma (Harvey & Sheehy), O. cruralis (Boulenger), O. discoidalis (Peracca), O. gran-ulosus (Boulenger), O. heterodactylus (Miranda-Ribeiro), O. ibischi (Reichle, Lötters & De la Riva), O. lehri(Padial, Chaparro & De la Riva), O. madidi (Padial, Gonzáles & De la Riva), O. pereger (Lynch), O. quixensisJiménez de la Espada, O. sanctaecrucis (Harvey & Keck), O. sanderi (Padial, Reichle & De la Riva), O. sax-atilis (Duellman), O. simmonsi (Lynch), and O. zongoensis (Reichle & Köhler). Prior to these thorough revi-sions, an increasing rate of field work conducted during the recent decade, mainly in Peru and Bolivia,provided new distinctive specimens and recordings of advertisement calls that led to the description of severalnew species (Reichle et al. 2001; Harvey & Sheehy 2005; Padial et al. 2005, 2007, 2008a).
Oreobates discoidalis is the only member of the genus known from Argentina, within its Argentineanrange being restricted to the northern provinces of Tucumán, Salta and Jujuy (Lavilla et al. 2000). Fieldworkin 2001 in a montane forest (Ecoregion of Southern Andean Yungas) of Salta yielded the discovery of speci-mens of Oreobates and recordings of advertisement calls which differ significantly from O. discoidalis and allother species in the genus. More recently, additional specimens were obtained at two adjacent localities of theArgentinean Yungas. We concluded that these specimens represent an unknown species, which is describedherein.
Material and methods
Morphometric measurements of preserved specimens were taken with digital calipers under a dissectingmicroscope to the nearest 0.1 mm. All measurements were taken by one of us (MV) to avoid inter-observerdifferences and rounded to only one decimal to avoid pseudo precision (Hayek et al. 2001). Terminology anddescription of morphological characters follows Hedges et al. (2008). We used morphological characters thatwere proposed as being of diagnostic value by those authors, but also follow a standardized format of a list ofcharacters included in Padial et al. (2008a) and Hedges et al. (2008), deemed to be important although notnecessarily diagnostic. A frog dissection with subsequent staining and clearing was accomplished to deter-mine the condition of the trigeminal nerve in relation to the adductor musculature (following Lynch 1986),and to describe diagnostic skeletal characters. Morphometrics measurements follow Padial et al. (2008a).Colour was registered in life and in alcohol. For comparisons with congeneric species we took data from liter-ature sources (Lynch 1975; Harvey & Keck 1995; Reichle et al. 2001; Harvey & Sheehy 2005; Padial et al.2005a,b, 2007, 2008a).
Advertisement calls of two individuals (five calls each) were recorded using a SONY TCM-20DV taperecorder and a Sennheiser ME-66 microphone by MV on 25 October 2001 at the type locality at midnight. Airtemperatures were 17.5 and 17.2 °C, respectively. Calls were digitized and analyzed by LF using a PC withCool Edit Pro software (Syntrillium Software Co.). Sampling frequency was 44,100 Hz at 16 bit resolution.Frequency information was obtained through Fast Fourier Transformation (width 256 points) at Hammingwindow function. The terminology used for the description of the calls follows Heyer et al. (1990). Quantita-tive parameters were expressed as mean ± one standard deviation. Advertisement call characteristics werecompared with published call descriptions of O. discoidalis (Padial et al. 2008b), O. cruralis (Padial et al.2008b), O. heterodactylus (Padial & De la Riva 2005), O. ibischi (Reichle et al. 2001, Padial et al. 2008b), O.madidi (Padial et al. 2005; Padial et al. 2008b), O. quixensis (Heyer & Gascon 1995), and O. sanctaecrucis(Köhler 2000). Given that published descriptions of the call of O. discoidalis are from a Bolivian populationgeographically distant from the type locality of the species in the province of Tucumán, northern Argentina(see Padial et al. 2008a), and that we heard but did not record O. discoidalis from the same locality as theunknown species, we included in call comparisons unpublished data on the calls of an Argentinean populationof O. discoidalis recorded by us at Abra de Cañas, Jujuy (380 km airline distance from the type locality of O.discoidalis, and 130 km airline distance from the three localities where we recorded the unknown species). We

Zootaxa 1908 © 2008 Magnolia Press · 43NEW OREOBATES FROM ARGENTINA-VAIRA & FERRARI
also included in the comparisons call characteristics from two geographically closer Bolivian populations ofOreobates from the Bellavista Mountains and La Hoyada, considered as O. cf. cruralis by Köhler (2000),Reichle et al. (2001), and Padial et al. (2008b). Voucher specimens indicated in this work are deposited at theherpetological collections of the Instituto de Herpetología, Fundación Miguel Lillo, Tucumán (FML).
Results
Oreobates barituensis sp. nov.Figs 1–2
Holotype. FML 21079 (field number MV 268), adult male, calling when captured, collected at Baritú, Depar-tamento Santa Victoria, Provincia Salta, Argentina (22º 29’ 58,53’’S 64º; 45’ 40,83’’W, 1100 m above sealevel), on 25 October 2001 by M. Vaira.
Paratypes. FML 21077-21078 (field numbers MV 255-256), adult males, and FML 21080 (field numberMV 269), adult female, same data as the holotype. MV 238-240 (to be catalogued at FML), adult males, samelocality as the holotype, collected on 2 December 2000 by M. Vaira. MV 410-413, adult males, collected atArazay, Departamento Santa Victoria, Provincia Salta, Argentina (22º 19’ 0’’S 64º 42’ 59,9’’W, 1300 m abovesea level), on 21 November 2007 by M. Vaira. MV 414-421, adult males, collected at Lipeo, DepartamentoSanta Victoria, Provincia Salta, Argentina (22º 25’ 52.56’’S 64º 44’ 7.17’’W, 1300 m above sea level), on 23November 2007 by M. Vaira.
Etymology. The species is named after the village of “Baritú” where the holotype specimen was col-lected.
FIGURE 1. Holotype of Oreobates barituensis sp. nov. in life (FML 21079).
VAIRA & FERRARI44 · Zootaxa 1908 © 2008 Magnolia Press
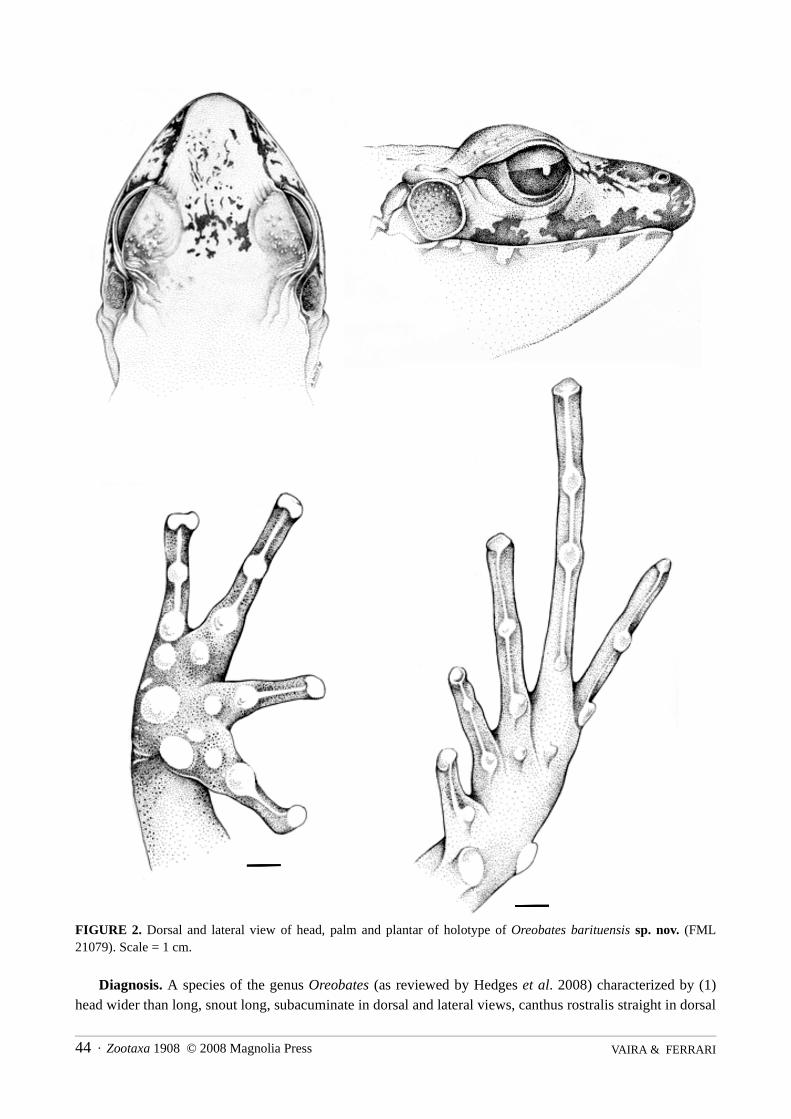
FIGURE 2. Dorsal and lateral view of head, palm and plantar of holotype of Oreobates barituensis sp. nov. (FML21079). Scale = 1 cm.
Diagnosis. A species of the genus Oreobates (as reviewed by Hedges et al. 2008) characterized by (1)head wider than long, snout long, subacuminate in dorsal and lateral views, canthus rostralis straight in dorsal

Zootaxa 1908 © 2008 Magnolia Press · 45NEW OREOBATES FROM ARGENTINA-VAIRA & FERRARI
view, rounded in profile; (2) tympanic membrane and annulus distinct, tympanum length half of the eyelength; supratympanic fold present; (3) cranial crests absent, upper eyelid with small tubercles; (4) dentiger-ous processes of vomers ovoid, prominent, posteromedial to choanae; (5) condition “S” of adductor muscle(sensu Lynch 1986); (6) males with vocal slits and a single faint nonspinous nuptial pad on thumb; (7) termi-nal discs of fingers I and II rounded not enlarged, terminal discs of fingers III and IV truncated and enlarged,circumferential grooves incomplete, ungual flap poorly defined, terminal phalanges knob-shaped, lateralfringes and keels on fingers absent, ulnar tubercles absent, prominent subarticular tubercles, enlarged and con-ical in profile, supernumerary tubercles conical, smaller than subarticular tubercles; (8) first finger slightlylonger than second; (9) toes long, lacking lateral fringes, webbing absent; fifth toe slightly shorter than third;(10) no tubercles on heel and tarsus, prominent subarticular tubercles, enlarged and conical in profile, innermetatarsal tubercle ovoid; (11) skin of dorsum scattered with soft tubercles, scapular and sacral glandspresent; (12) skin of venter smooth, discoidal fold present; (13) dorsal coloration of body and head in preser-vative brownish-gray with black marks; limbs brownish and striped with darker bands; venter whitish cream,throat finely mottled in gray.
Oreobates barituensis differs from other Oreobates species as follows (characters of the other species inparentheses): from O. choristolemma by the absence of occipital or mid-dorsal folds (occipital W-shaped foldand/or x-shaped mid-dorsal fold), lack of axillary gland (small axillary gland), and no indented ungual flap onfinger discs (notched ungual flap); from O. cruralis by head wider than long (longer than wide), and a singlefaint nuptial pad on thumb (no nuptial pads); from O. discoidalis by having head wider than long (longer thanwide), tympanum size about half of eye length (longer than half of eye length), and a single faint nuptial padon thumb (no nuptial pads); from O. granulosus by the skin of dorsum scattered with soft tubercles (low kera-tinized granules and warts); from O. heterodactylus by tympanum about half of eye length (two-thirds of theeye length), dentigerous process posteromedial to choane (medial to choane), and no ulnar tubercles (two tofour ulnar tubercles); from O. ibischi by the distinct supratympanic fold (weak tympanic fold), tympanumlength about half of eye length (more than two-thirds of eye length); and terminal discs of fingers III and IVenlarged and truncated (markedly enlarged and ovate); from O. lehri by skin of dorsum exhibiting low tuber-cles not keratinized (enlarged keratinized granules), supratympanic fold distinct and long (weak and short),fingers I and II shorter than finger IV (fingers I, II and IV of approximately the same length), and by its footlength about 50% of SVL (foot length = 60% of SVL); from O. madidi by exhibiting soft scattered tubercleson dorsum (heavily warty), and head wider than long (longer than wide); from O. pereger by the presence ofdiscoidal fold (no discoidal fold), palmar tubercle oval (bifid), and enlarged discs on fingers (no enlargeddiscs on fingers); from O. quixensis by a smaller size (SVL of adults 35–63 mm), and ventral surfaces smooth(heavily mottled); from O. sanctaecrucis by its predominantly bronze brown dorsal colouration (blackishbrown with red to scarlet spots in axiles); from O. sanderi by having no occipital and/or mid-dorsal fold (W-shaped occipital fold and/or x-shaped mid-dorsal fold), and the lack of supernumerary tubercles (conspicuoussupernumerary tubercles); from O. saxatilis by a smaller size (SVL of adults 44.5–50 mm) and the lack of anulnar tubercle (a single, round ulnar tubercle); from O. simmonsi by its dorsum and extremities with soft scat-tered tubercles (dorsum and extremities homogeneously granular), and a smooth venter (granular); and fromO. zongoensis by dorsal skin with soft scattered tubercles (strongly granular), dorsum bronze brown and ven-ter whitish cream (dorsal and ventral surfaces dark brown), and a golden iris in life (orange).
Description of the holotype. Adult male (Figs 1–2 ); snout-vent length 27.9 mm; head slightly wider thanbody, head length about 30% of SVL, wider than long (head width/head length 1.1); snout subacuminate indorsal view, and rounded in profile, tip of snout overlaps border of lower lip in lateral view, nostrils slightlyprotuberant, directed laterally; canthus rostralis straight in dorsal view, rounded in profile, loreal regionslightly concave and moderately sloping; upper eyelid with small tubercles, eyes in lateral position, no cranialcrests; supratympanic fold distinct, tympanic membrane and tympanic annulus distinct, tympanum nearlyround, its length half of eye length, a large postrictal gland. Choanae rounded partially concealed by palatal

VAIRA & FERRARI46 · Zootaxa 1908 © 2008 Magnolia Press
shelf when roof of mouth is viewed from below; dentigerous process of vomers ovate, posteriormedial to cho-anae, about the size of a choanae, tongue subcircular, vocal slits present, posterolateral to tongue. Skin of dor-sum scattered with soft tubercles, almost of the same size, with two larger glands on shoulders and two largerglands in sacral region surrounded by a kidney-shaped dark blotch; ventral surfaces smooth, dorsolateral foldscomposed by small warts, interrupted anteriorly; discoidal fold distinct.
No ulnar tubercles; palmar tubercle moderately large, outer oval larger than inner rounded tubercle; subar-ticular tubercles single, conical, and prominent; supernumerary tubercles conical, smaller than subarticulartubercles; a single faint nonspinous nuptial pad on thumb; disc of fingers I and II rounded not enlarged, disc offingers III and IV truncated and enlarged; fingers lacking lateral fringes and keels; relative length of fingersIII>IV>I>II.
Heel and tarsus lacking tubercles or folds; inner metatarsal tubercle ovoid, longer than wide; toes lackinglateral fringes and keels, webbing absent; fifth toe slightly shorter than third; terminal discs of toes I, II and Vrounded not enlarged, discs of toes III and IV truncated and moderately enlarged; relative length of toesIV>III>V>II>I.
FIGURE 3. Distribution of Oreobates barituensis sp. nov. in the montane forests of Northwestern Argentina. The squarerepresents the type locality and stars denote the other localities of collection.
Coloration: In life, dorsum bronze brown, marbled with dark brown, black and cream dots. Irregularinterocular dark stripe; interscapular region with yellowish oval blotch. Iris golden, marbled with black; blacksupratympanic stripe; lips with diffuse brown and white perpendicular bars. A black irregular stripe departsfrom the tip of the snout and continues to the midpart of the flanks. Dorsal surfaces of forelimbs and hind-limbs with diffuse blackish and brown bars. Venter uniformly whitish, finely mottled with light brown. Throatmottled dark brown. In preservative, dorsal coloration of body and head brownish-gray with black marks;limbs brownish, stripped with darker bands; venter whitish cream, throat finely mottled in gray.
Mean measurements (in mm) and proportions of holotype: SVL 26.2; head length 8.7; head width 8.8;eye length 2.9; eye to nostril distance 4.3; internarial distance 2.2; eye-eye distance 4.8; tympanic membraneheight 1.5; tympanic membrane length 1.4; arm length 6.0; thigh length 13.4; tibia length 12.6; foot length13.7. Tibia length/SVL 0.5; foot length/SVL 0.5; head length/SVL 0.3; head width/SVL 0.3; head width/headlength 1.1; eye to nostril/eye length 1.7; tympanic membrane length/tympanic membrane height 0.9.

Zootaxa 1908 © 2008 Magnolia Press · 47NEW OREOBATES FROM ARGENTINA-VAIRA & FERRARI
Variation: Range measurements (in mm) and proportions of male and female paratypes; female paratypein bold: SVL 21.2–29.9; 34.1; head length 7.6–10.5; 10.3; head width 7.7–10.1; 12.5; eye length 2.0–3.0; 3.1;eye to nostril distance 3.0–5.4; 4.8; internarial distance 1.9–2.4; 2.5; eye-eye distance 3.8–5.8; 6.2; tympanicmembrane height 1.1–2.1; 2.0; tympanic membrane length 0.9–2.1; 2.0; arm length 4.7–7.0; 7.5; thigh length10.5–15.8; 18.1; tibia length 10.5–14.5; 18.3; foot length 9.8–15.6; 16.6. Tibia length/SVL 0.4–0.5; 0.5; footlength/SVL 0.4–0.6; 0.5; head length/SVL 0.3–0.4; 0.5; head width/SVL 0.3–0.4; 0.3; head width/head length0.9–1.2; 1.3; eye to nostril/eye length 1.5–2.0; 1.5; tympanic membrane length/tympanic membrane height0.8–1.3; 0.9.
Most noticeable morphological differences compared to the holotype are as follows:a – Skin of dorsum less warty (MV 255, 269, 415, 417, 418, 419) to almost smooth (MV 256, 416, 420,
421).b – Supratympanic fold less noticeable (MV 255, 416).c – Upper eyelid with few small tubercles (MV, 420, 421) or smooth (MV 416).d – Discoidal fold less noticeable (MV 415, 417, 419, 421) Distribution and natural history. Oreobates barituensis is know from three localities (Arazay, Lipeo
and Baritú, Fig. 3), all small villages bordering the Parque Nacional Baritú, in a subtropical humid montaneforest of north-western Argentina (ecoregion of Southern Andean Yungas). The area consists of a mosaic ofagriculture, secondary shrub, and both primary and secondary forest. The individuals were collected whilecalling at dusk and night. Males were calling from the bare ground or from small rocks. The single gravidfemale was collected at Baritú together with calling males. The species occurred in syntopy with Oreobatesdiscoidalis.
Advertisement call. The call of Oreobates barituensis (Fig. 4) consists of a short trill formed by 5 to 6short notes (mean 5.3 ± 0.5), with a call duration of 168 to 211 milliseconds (mean 185.3 ± 18.9). The domi-nant frequency peak is between 2916 and 3449 Hz (mean 3226 ± 121.26). The note duration ranged from 2 to6 milliseconds (mean 4.3 ± 1.0) with a note repetition rate within calls ranging from 29.5 to 32.05 notes persecond (mean 30.6 ± 1.01).
Comparisons with published calls descriptions: The call of O. barituensis is shorter than the call of O.cruralis (179–620 ms), O. discoidalis (379–632 ms), and O. madidi (926–1075 ms); it is similar in duration tothe call of O. heterodactylus (126–245 ms), O. ibischi (89–264 ms), and O. quixensis (120–170 ms) andlonger compared to that of O. sanctaecrucis (91– 97 ms). It consists of fewer notes per call than the call of O.cruralis (13–27 notes), O. discoidalis (8–11 notes), and O. madidi (28–32 notes); it is similar to O. heterodac-tylus (5–9 notes) and O. ibischi (4–9 notes); but longer than O. quixensis (1–5 notes) and O. sanctaecrucis (1note per call). The note repetition rate is similar to O. madidi (29.3–30.3 notes/sec); smaller than O. cruralis(37.5–72.6 notes/sec) and larger than O. discoidalis (16.8–23.7 notes/sec) and O. sanctaecrucis (mean 10.8notes/sec). The dominant frequency is higher compared to calls of O. ibischi (2000–2981 Hz), O. discoidalis(2114–2472 Hz), O. madidi (2412–2584 Hz), O. quixensis (720–1620 Hz) and O. sanctaecrucis ( 1400–2200Hz), but slightly lower when compared to O. cruralis (2900–3800 Hz) and O. heterodactylus (2876–4134Hz).
Comparison with the advertisement call of O. discoidalis from Abra de Cañas, Argentina (Fig. 5): The 10call recordings of two individuals of this population consist of a long melodious call formed by 10 to 13 notes,with a call duration of 550 to 720 milliseconds (mean 634 ± 57.13). The note duration ranged from 24 to 40milliseconds (mean 31.6 ± 3.3) with a note repetition rate within calls ranging from 18 to 18.7 notes per sec-ond (mean 18.2 ± 0.27). The dominant frequency peak is between 1902 and 2164 Hz (mean 2068 ± 63.5). Incomparison, the call of O. barituensis is very distinctive with a notably shorter call length and fewer notes percall. The note duration is shorter, but with a longer repetition rate. The dominant frequency is considerablyhigher.
Comparison with the advertisement calls of O. cf. cruralis from Bolivia: The calls of O. cf. cruralis from

VAIRA & FERRARI48 · Zootaxa 1908 © 2008 Magnolia Press
the Bellavista Mountains reported by Reichle et al. (2001) and Padial et al. (2008b) are notably similar to thecalls emitted by O. barituensis (see spectogram of Fig. 2B in Padial et al. 2008b). Despite some disparity inthe number of notes per call (4 to 13 notes in O. cf. cruralis) the call length (107–392 ms), note repetition rate(33.65–37.4 ms), and dominant frequency (2880–3400 Hz) are very similar. In comparing the calls of O. cf.cruralis from la Hoyada with O. barituensis, we found noticeable differences in call length (326–338 ms),number of notes per call (9–10 notes), and dominant frequency (2680–2831 Hz).
FIGURE 4. Oscillogram and sound spectrogram of advertisement call of holotype of Oreobates barituensis sp. nov.(FML 21079, SVL 27.9 mm), recorded on 25 October 2001 at type locality. Air temperature during recording 17.5 °C.
FIGURE 5. Oscillogram and sound spectrogram of advertisement call of Oreobates discoidalis from Abra de Cañas,northern Argentina (MCN 361, SVL 27.7 mm), recorded on 08 November 1996. Air temperature during recording 20.2°C.
Discussion
Based on the data provided above, we conclude that the new species described here belongs to the genus Ore-obates as proposed by Hedges et al. (2008). Although O. barituensis is morphologically similar to the sympa-tric (and syntopic) O. discoidalis, as well as to O. ibischi and O. cruralis from Bolivia, the recognition of thenew species is particularly supported by considerable differences in the advertisement call among these spe-

Zootaxa 1908 © 2008 Magnolia Press · 49NEW OREOBATES FROM ARGENTINA-VAIRA & FERRARI
cies (Reichle et al. 2001; Padial et al. 2005; Padial et al. 2008b). Thus, with O. barituensis a second species ofOreobates is added to the amphibian fauna of the forests of northernmost Argentina. Previous studies showedthat morphological characters could be of little help to distinguish species belonging to the group formerlyconsidered eleutherodactylines, given their high phenotypic variability within populations and the morpholog-ical conservatism among species (Crawford & Smith 2005; Heinicke et al. 2007). Validity of usual morpho-logical characters, even the most reliable, was particularly accomplished by Hedges et al. (2008) to the non-Pristimantis species of the Andes of South America. However, temporal and spectral parameters of the adver-tisement call were recognized as useful characters to discriminate species in the genus Oreobates (Padial et al.2008a,b). In the absence of consistent morphological characters to distinguish species, bioacoustics haveproved to be a useful tool to diagnose the species formerly considered belonging to the Andean Eleutherodac-tylus discoidalis group. Padial et al. (2008b) utilized quantitative and qualitative acoustic characters to detectconsistent interspecific differences in this group of frogs. They found that temporal parameters such as num-ber of pulses and call duration, and also the spectral parameter of dominant frequency were useful diagnosticcharacters. Interspecific differences in call parameters were related to geographic distribution, showing latitu-dinal variation in number of pulses and pulse rate along the Andes (Padial et al. 2008b). Our analyses of theadvertisement call characteristics of specimens from Baritú were consistent with the methods used by thoseauthors to support the diagnosis of the recognized species and by others used for the description of new ones(e.g. Reichle et al. 2001).
Based on the acoustic evidence provided here, we are also able to consider some issues related to the iden-tification of Bolivian populations of O. discoidalis and O. cf. cruralis recorded by Köhler (2000), Reichle etal. (2001), and Padial et al. (2008b). As we stated above, the type locality of O. discoidalis is actually geo-graphically quite distant from the location where individuals of the Bolivian population were recorded byPadial et al. (2008b). There was no guarantee that this Bolivian population was correctly identified as belong-ing to the nominal O. discoidalis (J. Köhler, pers. comm.). Even with subtle variations, general similaritiesbetween the calls of O. discoidalis from northern Argentina provided by us with those described by Padial etal. (2008b) from Bolivia, indicate that in spite of the allopatric nature of these populations both would pertainto the nominal species. However, the absence of recordings from the type locality of O. discoidalis in northernArgentina may impede further definitions. Otherwise, overall similarities in the acoustic traits of O. barituen-sis calls with those of Bolivian O. cf. cruralis from Bellavista Mountains strongly suggests that both popula-tions might be conspecific and hence, the newly described O. barituensis would constitute a species with amuch wider distribution. Further recordings of the advertisement calls of closer populations from Bolivia andArgentina would be of great help to achieve a better understanding of this complex group.
Acknowledgments
This work is part of a project supported by a grant from CONICET to MV. It is also part of projects funded bya PIP-CONICET to R. F. Laurent, and by the Consejo de Investigaciones de la Universidad Nacional de Saltato M. Fabrezi. Permits for access and field work in Parque Nacional Baritú were provided by Delegación Téc-nica de Parques Nacionales Regional Noroeste whose cooperation is greatly appreciated. We are especiallygrateful to A. DuPuy for the drawings. To W. Maciel, U. Colina, M. Entrocassi, and F. Dobrotinich for thehelp provided during fieldwork to MV. Corrections and criticisms of J. Köhler greatly improved an earlier ver-sion of this manuscript drawing our attention to important issues related to the comparisons of the calls ofOreobates discoidalis and of O. cf. cruralis populations. We also thank D. Koscinski for corrections and sug-gestions.References

VAIRA & FERRARI50 · Zootaxa 1908 © 2008 Magnolia Press
Crawford, A.J. & Smith, O.N. (2005) Cenozoic biogeography and evolution in direct-developing frogs of Central Amer-ica (Leptodactylidae: Eleutherodactylus) as inferred from a phylogenetic analysis of nuclear and mitochondrialgenes. Molecular Phylogenetics and Evolution, 35, 536–555.
Frost, D.R., Grant, T., Faivovich, J., Bain, R.H., Haas, A., Haddad, C.F.B., de Sá, R.O., Channing, A., Wilkinson, M.,Donnellan, S.C., Raxworthy, C.J., Campbell, J.A., Blotto, B.L., Moler, P., Drewes, R.C., Nussbaum, R.A., Lynch,J.D., Green, D.M. & Wheeler, W.C. (2006) The amphibian tree of life. Bulletin of the American Museum of NaturalHistory, 297, 1–370.
Hayek, L.C., Heyer, R.W. & Gascon, C. (2001) Frog morphometrics: a cautionary tale. Alytes, 18,153–177.Harvey, M.B. & Keck, M.B. (1995) A new species of Ischnocnema (Anura: Leptodactylidae) from high elevations in the
Andes of central Bolivia. Herpetologica, 51, 56–66.Harvey, M.B. & Sheehy, C.M. (2005) A new species of Ischnocnema (Anura: Leptodactylidae) from La Paz, Bolivia.
Herpetologica, 61, 268–275.Hedges, S.B., Duellman, W.E. & Heinicke, M.P. (2008) New World direct-developing frogs (Anura: Terrarana): Molecu-
lar phylogeny, classification, biogeography, and conservation. Zootaxa, 1737, 1–182.Heinicke, M.P., Duellman, W.E. & Hedges, S.B. (2007) Major Caribbean and Central American frog faunas originated
by ancient oceanic dispersal. Proceedings of National Academy of Sciences (USA), 104,10092–10097.Heyer W.R. & Gascon, C. (1995) Collection notes and call characteristics for Ischnocnema quixensis and Phyzelaphryne
miriamae (Amphibia: Anura; Leptodactylidae). Journal of Herpetology, 29; 304–307.Heyer, W.R., Rand, A.S., Cruz, C.A. Peixoto, O.L. & Nelson, C.O. (1990) Frogs of Boracéia. Arquivos de Zoología Sao
Paulo; 31, 237–410.Köhler, J. (2000) Amphibian diversity in Bolivia: a study with special reference to montane forest regions. Bonner Zool-
ogische Monographien, 48, 1–243.Lavilla, E.O., Vaira, M., Ponssa, M.L. & Ferrari, L. (2000) Batracofauna de las Yungas Andinas de Argentina: Una sínte-
sis. Cuadernos de Herpetología, 14, 5–26.Lynch, J.D. (1975) A review of the Andean leptodactylid frog genus Phrynopus. Occasional Papers of the Museum of
Natural History, University of Kansas, 35, 1–51.Lynch, J.D. (1986) The definition of the Middle American clade of Eleutherodactylus based on jaw musculature
(Amphibia: Leptodactylidae). Herpetologica, 42, 248–258. Padial, J.M. & De la Riva, I. (2005) Rediscovery, redescription, and advertisement call of Eleutherodactylus heterodacty-
lus (Miranda Ribeiro, 1937) (Anura: Leptodactylidae), and notes on other Eleutherodactylus. Journal of Herpetol-ogy, 39, 372–379.
Padial, J.M., Gonzáles, L. & De la Riva, I. (2005) A new species of the Eleutherodactylus discoidalis group (Anura: Lep-todactylidae) from Andean humid montane forests of Bolivia. Herpetologica, 61, 318–325.
Padial, J.M., Chaparro, J.C. & De la Riva, I. (2007) A new species of the Eleutherodactylus discoidalis Group (Anura:Brachycephalidae) from cloud forests of Peru. Herpetologica, 63,114–122.
Padial, J.M., Chaparro, J.C. & De la Riva, I. (2008a) Systematics of Oreobates and the Eleutherodactylus discoidalisspecies group (Anura) based on two mtDNA genes and external morphology. Zoological Journal of the LinneanSociety, 152, 737–773.
Padial, J.M., Köhler, J., Muñoz, A. & De la Riva, I. (2008b) Assessing the taxonomic status of tropical frogs through bio-acoustics: geographical variation in the advertisement calls in the Eleutherodactylus discoidalis species group(Anura). Zoological Journal of the Linnean Society, 152, 353–365.
Reichle, S., Lötters, S. & De la Riva, I. (2001) A new species of the discoidalis group of Eleutherodactylus (Anura, Lep-todactylidae) from inner-Andean dry valleys of Bolivia. Journal of Herpetology, 35, 21–26.




















