
A politica de oferta educacional no Estado do Parana 2001 2006

A new species of Creagrutus Gunther (Teleostei:Ostariophysi: Characiformes), from the upper Rio
Parana basin, central Brazil
A. C. RIBEIRO*†, R. C. BENINE* AND C. A. FIGUEIREDO‡
*Laboratorio de Ictiologia de Ribeirao Preto, Departamento de Biologia, FFCLRP–Universidade de Sao Paulo, Av. Bandeirantes 3900, 14 040-901, Ribeirao Preto, SP,Brazil and ‡Museu de Zoologia da Universidade de Sao Paulo, Av. Nazare 481,
04 263-000, Sao Paulo, SP, Brazil
(Received 3 April 2003, Accepted 18 November 2003)
Creagrutus varii n. sp. is described based on material from the Rio Sao Marcos basin, in the
upper Rio Parana system, Goias State, Brazil. This is the first record of Creagrutus in the upper
Rio Parana. The new species is proposed to be the sister group of the remaining species of
Creagrutus. Creagrutus varii differs from congeners in the morphology of the maxilla, primor-
dial ligament, premaxillary dentition and other features. The evolution of premaxillary tooth
morphology in Piabina and Creagrutus and the biogeography of the Rio Paranaıba basin are
discussed. # 2004 The Fisheries Society of the British Isles
Key words: Creagrutus; new species; phylogeny; Rio Parana basin.
INTRODUCTION
The genus Creagrutus has been the subject of a series of recent taxonomic andphylogenetic studies. These revealed a high level of previously undetectedspecies-level diversity within Creagrutus, and provided insights into the phylo-genetic and biogeographic relationships of the components of the genus (Harold& Vari, 1994; Vari & Harold, 1998; Vari & Harold, 2001). Prior to the presentpaper, the genus consisted of 65 species, eight of these with trans-Andean, andthe remaining 57 with cis-Andean distributions (Harold & Vari, 1994; Vari &Harold, 2001; Vari & Lima, 2003).
Although Creagrutus is widespread across most of the inland waters of SouthAmerica (Harold & Vari, 1994; Vari & Harold, 2001), no Creagrutus specieshad been recorded in the upper Rio Parana system, a vast area encompassingc. 891 000 km2 (Paiva, 1982). This river system is one of the most impacted byanthropogenic activities in Brazil. Although it is also the most investigatedregion of that country in terms of biological surveys, the continued descriptionsof new species indicate that the fish fauna of the upper Rio Parana remains
†Author to whom correspondence should be addressed. Tel.: þ55 16 602 3710; fax: þ55 16 602 3666;
email: [email protected]
Journal of Fish Biology (2004) 64, 597–611
doi:10.1046/j.1095-8649.2003.00324.x,availableonlineathttp://www.blackwell-synergy.com
597# 2004TheFisheries Society of theBritish Isles

poorly known, particularly in the more remote portions of the basin above theconfluence of the Rio Paranaıba and Rio Grande in the states of Minas Geraisand Goias.
In this paper, a new species of Creagrutus is described from headwaterstreams of the Rio Sao Marcos, a tributary of the upper portion of the RioParana basin, and its placement within the phylogenetic framework for thegenus proposed by Vari & Harold (2001) is investigated. The implications ofthe discovery of this new species for the phylogeny and biogeography of thegenus Creagrutus are also discussed.
MATERIALS AND METHODS
Counts and measurements were taken from the left side of specimens with digitalcalipers and are those described by Harold & Vari (1994). All specimen lengths arestandard lengths (LS) in mm. Cleared and stained preparations follow the methods ofTaylor & Van Dyke (1985). Vertebral counts include the Weberian apparatus as fourelements. The terminal half centrum, hypural bones and associated vertebral elements,together usually designated as PU1þU1 were counted as one vertebra. All osteologicalobservations, including vertebral and branchiostegal ray counts were taken from clearedand stained specimens. Colour in life was described from a specimen photographedimmediately after capture.
The phylogenetic analyses were performed using the character matrix published byVari & Harold (2001), including the outgroup data provided in that study, with theaddition of the new species. The analysis was performed using PAUP version 3.1.1(Swofford, 1993), with all characters unordered and equally weighted.
Institutional abbreviations are as listed in Leviton et al. (1985), with the addition of:LIRP, Laboratorio de Ictiologia de Ribeirao Preto, Faculdade de Filosofia Ciencias eLetras de Ribeirao Preto, Universidade de Sao Paulo, Ribeirao Preto, Brazil; DZSJRP,Colecao de Peixes do Departamento de Zoologia e Botanica do Instituto de Biociencias,Letras e Ciencias Exatas, Universidade Estadual Paulista, Sao Jose do Rio Preto, Brazil.
Osteological terminology is that used by Weitzman (1962) with the modificationsproposed by Vari & Harold (2001). Abbreviations of bones in the figures are: a, antor-bital; aa, anguloarticular; ac, anterior ceratohyal; b, basihyal; bb, basibranchial; bb4,fourth basibranchial; cb, ceratobranchial; cb5, fifth ceratobranchial; br, branchiostegalrays; c, cleithrum; co, coracoid; d, dentary; dh, dorsal hypohyal; eb, epibranchial; ep,epioccipital; ept, ectopterygoid; es, extrascapular; f, frontal; gr, gill rakers; hb, hypobran-chial; hm, hyomandibular; ih, interhyal; i1–i6, infraorbitals 1–6; io, interopercle; le,lateral ethmoid; m, maxilla; mc, mesocoracoid; me, mesethmoid; msp, mesopterygoid;mtp, metapterygoid; n, nasal; o, opercle; os, orbitosphenoid; pa, parietal; pal, palatine;pc, posterior ceratohyal; pc1, postcleithrum 1; pc2, postcleithrum 2; pc3, postcleithrum 3;pm, premaxilla; po, preopercle; ps, parasphenoid; pt, pterotic; pts, pterosphenoid; ptt,posttemporal; q, quadrate; ra, retroarticular; rs, rhinosphenoid; s, scapula; sc, supraclei-thrum; so, supraoccipital; sop, subopercle; sp, suspensory pharyngeals; up, upperpharyngeals; v, vomer; vh, ventral hypohyal.
RESULTS
CREAGRUTUS VARII N. SP. (FIG. 1)
HOLOTYPE
LIRP 4342, 38�0 mm; Brazil: Goias, Municıpio de Catalao, Rio Sao Marcosbasin (tributary of Rio Paranaıba basin), Corrego da Prata (17�4702600 S;47�3504800 W); E.S.R. Sa & C.A. Figueiredo; 25 April 2002.
598 A. C . RIBEIRO ET AL .
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

PARATYPES
LIRP 3358, 32, 2 cleared and stained, 22�1–42�0 mm; MZUSP 81 069, 20,17�6–37�5 mm; MCP 31 989, 20, 17�5–35mm; ZUEC 6165, 8, 15�5–30�5 mm;DZSJRP 5424, 8, 16�5–32�5 mm; collected with holotype. LIRP 3625, 14,16�0–40�3 mm; collected at type locality; E.S.R. Sa & C.A. Figueiredo, 26 July2002. LIRP 3359, 7, 15�2–32�2mm; Brazil: Goias, Municıpio de Paracatu, RioSao Marcos basin (tributary of Rio Paranaıba basin), Ribeirao da Batalha atFazenda Batalha (17�2502500 S; 47�2701100 W); E. S. R Sa & C.A. Figueiredo, 27April 2002.
DIAGNOSIS
Creagrutus varii differs from congeners in having a maxilla with a relativelyslender anterior process and a posterior portion transversely compressed (v. witha thick anterior process and robust posterior portion); a maxilla without adistinct flexion in the region where it extends past the posterior limit of thepremaxilla (v. possessing a flexion); a primordial ligament that is relatively flattransversely and attaches to the posteromedial process of the ascending processof the maxilla (v. being rotund in cross-section and attaching primarily to thedistal one-half of the maxilla, but not to the distal portion of that bone); themesopterygoid and metapterygoid overlapping along their area of contact(v. being separated); a quadrate without a variably developed vertical ridge orprocess extending dorsally from the main body of the quadrate in, and posteriorto, the region where the posterodorsal process of the quadrate contacts theanteroventral process of metapterygoid (v. the presence of this structure); andthe presence of the temporal foramen (v. its absence).
Another feature useful in distinguishing C. varii from other species ofCreagrutus is the presence of two rows of teeth on the premaxilla, with twoconical teeth in the outer row.
DESCRIPTION
Morphometric and meristic data for C. varii are presented in Table I. Bodyrelatively slender in juveniles, becoming increasingly robust in larger specimens.Greatest body depth at vertical situated in region between humeral spot anddorsal-fin origin. Dorsal profile of body convex from snout tip to dorsal-finorigin, slightly concave from posterior end of dorsal-fin base to adipose fin,
FIG. 1. Creagrutus varii holotype, LIRP 4342, 38�0mm; Brazil, Goias, Municıpio de Catalao, Corrego da
Prata, 17�470260 0 S; 47�350480 0 W.
NEW CREAGRUTUS SPECIES FROM CENTRAL BRAZIL 599
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

ranging between slightly concave and straight between adipose fin and caudal-finbase. Ventral profile of head obtuse at anteroventral corner of dentary, straight togently convex from that angle to isthmus. Ventral profile of body convex (largerindividuals) to straight (juveniles) from isthmus to anal-fin origin, straight toslightly concave along anal-fin base, and gently concave from posterior end ofanal-fin base to caudal-fin base.
Head elliptical in lateral view; moderately compressed in dorsal view, butwider in interorbital region. Anterior portion of snout fleshy, with discreet
TABLE I. Morphometric and meristic character values of holotype and paratypes ofCreagrutus varii
Paratypes
Holotype Range Mean S.D.
MorphometricsStandard length (LS) (mm) 38�0 22�1–42�0 34�3 3�9Percentage of LS
Snout to anal-fin origin 57�9 51�6–60�8 59�0 1�7Snout to pelvic-fin origin 43�9 40�7–47�8 44�5 1�2Snout to pectoral-fin origin 23�9 22�4–25�9 24�0 0�9Snout to dorsal-fin origin 48�7 48�7–53�8 50�8 0�9Dorsal-fin origin to hypural joint 53�7 48�6–54�3 52�2 1�1Dorsal-fin origin to anal-fin insertion 26�6 24�1–27�9 26�3 0�8Dorsal-fin origin to pelvic-fin insertion 26�6 24�0–27�9 26�5 0�9Dorsal-fin origin to pectoral-fin insertion 33�4 24�4–35�7 33�8 2�6Caudal peduncle depth 8�7 7�8–10�5 9�3 0�6Pectoral-fin length 18�7 14�5–21�1 18�9 1�3Pelvic-fin length 13�7 12�2–15�4 14�0 0�8Dorsal-fin length 22�6 20�4–25�4 23�1 1�2Anal-fin length 18�4 16�3–20�7 18�6 0�9Head length 24�2 23�2–26�0 24�6 0�8
Percentage of head lengthPostorbital head length 38�0 37�3–44�2 40�6 1�9Snout length 26�1 21�0–36�1 26�6 2�4Bony orbital diameter 35�9 33�3–40�3 36�5 1�7Interorbital width 30�4 25�8–33�3 30�1 1�8
MeristicsMode
Lateral line scales 39 38–40 39Scale rows above lateral line 4 5–4 5Scale rows below lateral line 6 5–6 5Predorsal medium scales 11 11–16 13Branched dorsal-fin rays 8 8 8Branched anal-fin rays 16 15–17 17Branched pelvic-fin rays 6 6 6Pectoral-fin rays 10 10 10
600 A. C . RIBEIRO ET AL .
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

papillae along ventral surface of upper lips; papillae present between outer andmedial premaxillary teeth. Papillae on dorsal surface of lower lips larger thanpapillae on upper lips.
Appressed pectoral fin nearly reaching base of pelvic fin. Dorsal-fin originlocated at vertical situated slightly posterior of pelvic fin. Anal-fin originsituated at a vertical that extends through posterior portion of dorsal-fin base.Adipose fin inserted at a vertical through bases of last tree anal-fin rays.
OSTEOLOGY
Portion of cranial fontanel anterior to epiphyseal bar distinctly smaller thanportion located posteriorly. Anteriormost two thirds of frontal bones in contactalong middorsal line.
Infraorbital series consists of six well-developed elements. Third to sixthinfraorbitals contribute to posterior margin of infraorbital series. Second infra-orbital overlaps anterior portion of third infraorbital. Antorbital distinctlywider anteriorly and with distinct posteroventral angle along its longest axis.Supraorbital absent; dorsal margin of eye delimited by frontal bone (Fig. 2).Distal portion of lateral surface of sphenotic spine wide.
Upper jaw longer than, and overlapping, lower jaw (Fig. 2). Maxilla extend-ing from point located anterior to nares posteriorly to vertical slightly posteriorto ventral wing of lateral ethmoid (Fig. 2).
Premaxilla and maxilla attached by well-developed ligaments, extendingbetween posteroventral corner of premaxilla and anterior surface of maxilla,and by two distinct ligaments extending from anterodorsal tip of maxillato dorsal surface of premaxilla and from midventral portion of ascendingprocess of maxilla to dorsal surface of premaxilla (Fig. 3). Primordial ligament
soptt
ep
es
sc
pc1
o
sop
ciopo
bri1-i6
ra qaad
m
pm
a
n me
f le
rs os
pspts
pt pa
msp
FIG. 2. Lateral view of skull of Creagrutus varii, paratype, LIRP 3358, 33�0 mm; anterior to left (see text
for explanation of the abbreviations).
NEW CREAGRUTUS SPECIES FROM CENTRAL BRAZIL 601
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

bifurcates ventrally with attachments to both lateral surface of anguloarticularand lateral margins of shelf-like lateral process of the quadrate (Fig. 4).
Anterior process of mesethmoid ventrally expanded into well-developedventral process (Fig. 5).
Ectopterygoid and quadrate not in contact. Quadrate bearing shelf-likelateral process. Anteromedial portion of hyomandibula anteriorly expanded,extending over posterodorsal region of metapterygoid. Dorsoarticular surfaceof hyomandibula asymmetrical; hyomandibula with dorsolateral buttress(Fig. 4).
me n
2
pm
m
1
3
4
FIG. 3. Schematic lateral view of region of premaxilla, maxilla and associated ligaments of Creagrutus
varii, paratype, LIRP 3358, 36�4 mm, right side: 1, ligament between anterior portion of maxilla
and lateral process of mesethmoid; 2, ligaments from anterodorsal tip of maxilla to dorsal surface
of premaxilla; 3, primordial ligament; 4, ligament from midventral portion of ascending process of
maxilla to dorsal surface of premaxilla; anterior to right.
pal msp
mtp
hm
2 o
sop
iopo
1
qept
FIG. 4. Lateral view of suspensorium and opercular complex of Creagrutus varii, paratype, LIRP 3358,
46�4mm; 1, shelf-like lateral process of quadrate; 2, dorsolateral buttress of hyomandibula; anterior
to left.
602 A. C . RIBEIRO ET AL .
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

Dentigerous surface of premaxilla longitudinally elongate (Fig. 6). Premaxil-lary dentition in two series: inner row with four tricuspid teeth aligned inregular, curved series. Inner row teeth with second tooth in series slight widerthan other teeth, progressively decreasing in size posteriorly. Outer tooth rowwith five teeth, arranged in an undulatory pattern. First, tooth of outer rowslender and conical, located slightly externally to rest of its series and followedby three, bicuspid teeth of about same size, located more internally, and onemore externally positioned conical tooth (Figs 6 and 7). Dentary dentitionconsisting of three tricuspid teeth followed by four to six conical, smallerteeth that gradually decreases in size posteriorly (Fig. 8). Anguloarticularhorizontally foreshortened (Fig. 8). Dentary foramen located immediatelyanterior to tip of Meckel’s cartilage.
Mesocoracoid and scapula in contact through small intervening cartilage(Fig. 9). Scapula, coracoid, and cleithrum contacting each other anterior toscapular foramen.
Vertebrae 37–38. Precaudal vertebrae 16. Caudal vertebrae 21–22. First ptery-giophore of dorsal-fin located between the neural spines of 9th and 10thvertebrae or 8th and 9th vertebrae. Insertion of pelvic-fin at a vertical through
me
v
1le
rs
os
FIG. 5. Anterior portion of skull of Creagrutus varii, lateral view, paratype, LIRP 3358, 46�4mm; 1,
anteroventral process of mesethmoid; anterior to left.
pmB
C
BA
FIG. 6. Ventral view of premaxilla and premaxillary dentition of Creagrutus varii, paratype, LIRP 3358,
46�4mm. Letters indicate homologous system proposed by Vari and Harold (2001): A, triangular
cluster of teeth; B, primary premaxillary tooth series; C, single lateral tooth.
NEW CREAGRUTUS SPECIES FROM CENTRAL BRAZIL 603
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

5th and 6th pleural ribs. First pterygiophore of anal-fin located between haemalspines of 15th and 16th vertebrae or 14th and 15th vertebrae. Caudal skeletonwith two epurals and six separate hypurals. Hypural 6 free from the compoundvertebra. Eleven or 12 dorsal, and nine or 10 ventral procurrent rays.
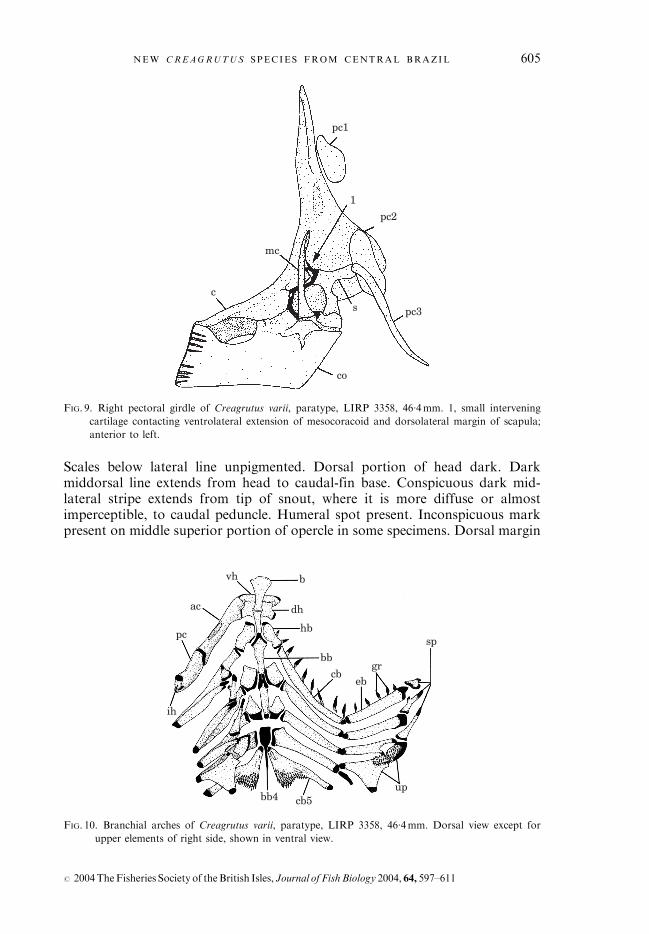
Branchiostegal rays four; three originating on anterior ceratohyal and one onposterior ceratohyal. Dorsal surface of fourth basibranchial cartilaginous, with-out bony surface plate (Fig. 10). Dentigerous portion of fifth ceratobranchialwell-developed and elongate, meeting main shaft of ossification at approxi-mately a right angle (Fig. 10). Gill rakers 14–15; nine originating from cerato-branchial and five to six from epibranchial and hypobranchial (Fig. 10).
COLOUR IN ALCOHOL
Overall ground colouration yellowish. Scales on dorsal region of body darklypigmented along posterior margins and forming reticulated colour pattern.
pm
m
FIG. 7. Lateral view of premaxilla and maxilla of Creagrutus varii, paratype, LIRP 3358, 46�4mm;
anterior to right.
d
ra
aa
FIG. 8. Lateral view of lower jaw of Creagrutus varii, paratype, LIRP 3358, 46�4mm; anterior to right.
604 A. C . RIBEIRO ET AL .
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611
Scales below lateral line unpigmented. Dorsal portion of head dark. Darkmiddorsal line extends from head to caudal-fin base. Conspicuous dark mid-lateral stripe extends from tip of snout, where it is more diffuse or almostimperceptible, to caudal peduncle. Humeral spot present. Inconspicuous markpresent on middle superior portion of opercle in some specimens. Dorsal margin
pc1
pc2
1
pc3
co
s
mc
c
FIG. 9. Right pectoral girdle of Creagrutus varii, paratype, LIRP 3358, 46�4mm. 1, small intervening
cartilage contacting ventrolateral extension of mesocoracoid and dorsolateral margin of scapula;
anterior to left.
vh b
dh
hb
bb
cbeb
gr
upcb5bb4
ih
pc
ac
sp
FIG. 10. Branchial arches of Creagrutus varii, paratype, LIRP 3358, 46�4mm. Dorsal view except for
upper elements of right side, shown in ventral view.
NEW CREAGRUTUS SPECIES FROM CENTRAL BRAZIL 605
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

of eye dark brown. Fins hyaline, with scattered dark pigmentation along someof dorsal, anal and caudal-fin rays. Scales at base of anal-fin with dark greycromatophores. Obvious spot present on base of median rays of caudal-finconstituted by an aggregation of pigments, which becomes gradually diffuseposteriorly.
COLOUR IN LIFE
Dark colouration as described under colour in alcohol. Overall groundcolouration yellowish. Dorsal portion of body darker, and becoming markedlybright silvery below lateral line. Middle superior portion of opercle with con-spicuous dark brown spot, with about the same size of humeral spot. Eyes darkred immediately dorsal to pupil.
Dorsal-fin rays yellowish and outlined with dark brown chromatophores.Paired fins yellowish to hyaline, with rays outlined and overlain by darkchromatophores. Procurrent and branched caudal-fin rays yellowish and out-lined by small black chromatophores. Anal fin yellowish along basal two thirdsof branched rays.
DISTRIBUTION
Known from only two localities in the upper Rio Paranaıba basin (Fig. 11).
ETYMOLOGY
The specific epithet, varii, is in honour of Richard Vari of the SmithsonianInstitution in recognition of his contributions to the knowledge of SouthAmerican fishes, especially on Creagrutus systematics.
MJW
49°00′ 48°30′ 48°00′ 47°30′ 47°00′ 46°30′ 46°00′ 45°30′
19°00′
18°30′
18°00′
17°30′
17°00′
16°30′
4
32
1
FIG. 11. Map of South American drainage showing collection localities (area in detail) of Creagrutus varii
and adjacent drainage. H, type locality (Brazil, Goias, Municıpio de Catalao, Corrego da Prata,
17�470260 0 S; 47�350480 0 W); *, nontype locality (Brazil, Goias, Municıpio de Paracatu, stream at
Fazenda Batalha, 17�250250 0 S; 47�270110 0 W; 1, Rio Sao Marcos; 2, Rio Paranaıba; 3, Rio Paracatu;
4, Rio Sao Francisco.
606 A. C . RIBEIRO ET AL .
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

DISCUSSION
In their phylogenetic and taxonomic study of Creagrutus and Piabina, Vari &Harold (2001) recognized nine clades, which encompassed Piabina argenteaReinhardt and 60 Creagrutus species. As noted by these authors, the degree ofresolution was relatively poor, as might be expected given the relatively low CIof c. 0�40 (Vari & Harold, 2001). The sister group relationships between Crea-grutus and P. argentea, as well as the monophyly of Creagrutus were, however,well corroborated.
To elucidate the phylogenetic placement of C. varii, it was included as aterminal in the analysis of Vari & Harold (2001). The strict consensus demon-strates that it is a member of the PiabinaþCreagrutus clade, positioned as thesister group of all other Creagrutus species. This position is a consequence of theabsence, in C. varii, of several derived characters proposed by Vari & Harold(2001) for all other Creagrutus species. Table II shows character states for 57characters presented by Vari & Harold (2001) and Fig. 12 shows the strictconsensus derived from the manipulation of their matrix.
The new species shares eight synapomorphies with the PiabinaþCreagrutusclade: 1) the presence of a well-developed ligament between the posteroventralcorner of the premaxilla and the anterior surface of the maxilla; 2) the anteriorterminus of dentary dentition is distinctly posterior to anterior terminus ofpremaxillary dentition; 3) the separation of the ectopterygoid and quadrate;4) an asymmetrical dorsal articular surface of the hyomandibula, with thehyomandibula having a dorsolateral buttress; 5) the expansion of the anteriorportion of mesethmoid ventrally into a relatively large, transversely flattened,vertical plate; 6) a hyomandibular fossa that is visible laterally as a consequenceof the reorientation of that portion of the sphenotic; 7) the distal portion oflateral surface of sphenotic spine variously elaborated posteriorly; 8) thepresence of two epurals [derived conditions of characters 10, 13, 21, 27, 35,42, 43 and 55, respectively of Vari & Harold (2001)]. Although belonging to thesame monophyletic group, C. varii differs from Piabina (the sister group ofCreagrutus) by having a fourth infraorbital that contributes to posterior margin
TABLE II. Character states of Creagrutus varii for 57 characters presented by Vari &Harold (2001) in their Creagrutus phylogeny. Characters are listed from top to bottom,
arranged in separated groups in columns
Characters 1–10 11–20 21–30 31–40 41–50 51–57
0 1 1 0 0 00 1 0 1 1 00 1 0 0 1 00 0 0 0 0 0
Character states 0 1 0 1 0 10 0 1 0 0 00 1 1 1 1 00 0 1 0 01 1 0 1 11 0 0 0 0
NEW CREAGRUTUS SPECIES FROM CENTRAL BRAZIL 607
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

of the infraorbital series (v. being approximately triangular and excluded fromthe posterior margin of the infraorbital series), a dorsal surface of fourthbasibranchial without a bony plate (v. with a bony plate), and by having anarrow ring-like process forming the anterior border of the scapular foramen(in contrast to the absence of this structure in Piabina).
The hypothesis that this new species is a member of Creagrutus is based on itspossession of the following derived characters: 1) a ventrally bifurcating pri-mordial ligament with attachments to both the lateral surface of anguloarticularand the lateral margins of the shelf-like lateral process of the quadrate; 2) adistinct ligament extending between the anterodorsal tip of the maxilla and thedorsal surface of the premaxilla; 3) a ligament extending between the midventralportion of the ascending process of the maxilla and the dorsal surface of thepremaxilla; 4) a horizontally foreshortened anguloarticular; 5) the anteriorlyexpanded anterior margin of hyomandibula extending over the posterodorsalregion of the metapterygoid; 6) the ventrolateral extension of the mesocoracoidcontacting the dorsolateral margin of the scapula through a small interveningcartilage; 7) the absence of an opening delimited by the scapula, coracoid andcleithrum anterior to the scapular foramen [derived conditions of characters 9,11, 12, 19, 26, 47 and 49, respectively, of Vari & Harold (2001)].
Some of the derived characters presented in C. varii are homoplasies sharedwith other clades under the most parsimonious hypothesis of relationships. Theclade consisting of C. cracentis Vari & Harold and C. maxillaris (Myers) shareswith C. varii: 1) a medial opening of the dentary foramen located immediatelyanterior to the tip of the Meckelian cartilage and 2) the presence of more thanseven dentary teeth [derived conditions of characters 15 and 17, respectively ofVari & Harold (2001)]. Both characters, however, are ambiguously optimized. Ahomoplastic character present in C. varii, shared with C. melasma Vari, Harold &Taphorn and C. lepidus Vari, Harold, Lasso & Machado-Allison is an antorbitalbone that is distinctly wider anteriorly and with a distinct posteroventral angle on
Piabina Creagrutus variiRemaining
Creagrutus species
3
2
1
FIG. 12. Strict consensus tree of relationships of Piabina argentea, Creagrutus varii and the remaining
Creagrutus species, based on 100 equally parsimonious trees of 151 steps in length (CI¼ 0�48,
RI¼ 0�78). Characters supporting branches are derived conditions of characters 10, 13, 21, 27, 35,
42, 43 and 55 in clade 1, characters 9, 11, 12, 19, 26, 47 and 49 in clade 2, and characters 5, 6, 22,
23, 25, 29, 30, 41 and 48 in clade 3 of Vari & Harold’s (2001) phylogeny.
608 A. C . RIBEIRO ET AL .
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

its longest axis. This feature [derived condition of character 28 of Vari & Harold(2001)], is homoplastic according to both the present analysis and that of Vari &Harold (2001). The absence of a bony plate on the dorsal surface of the fourthbasibranchial [derived condition of character 32 of Vari & Harold (2001)] is also ahomoplasy shared by C. varii and some Creagrutus species. According to Vari &Harold (2001), this ossification is present in P. argentea and in all but fourteenCreagrutus species (C. atratus Vari & Harold, C. barrigai Vari & Harold,C. bolivari Shultz, C. gephyrus Bohlke & Saul, C. gracilis Vari & Harold,C. machadoi Vari & Harold, C. magdalenae Eigenmann, C. mucipu Vari &Harold, C. paralacus Harold & Vari, C. pila Vari & Harold, C. seductus Vari &Harold, C. taphorni Vari & Harold, C. vexillapinnus Vari & Harold andC. zephyrus Vari & Harold).
Employing the ACCTRAN optimization criterion, the shape of the premax-illa and the arrangement of teeth on that bone are reversals in C. varii. Vari &Harold (2001) noted that most of the Creagrutus species and P. argentea have apremaxilla that approximates an irregular triangle when examined from ventralview (Vari & Harold, 2001). In these taxa the premaxillary teeth are large withrespect to both the size of the upper jaw and the dentigerous surface of thepremaxilla. As a consequence, the dentition covers an unusually high propor-tion of the ventral surface of the premaxilla relative to the condition present inmost characids. Vari & Harold (2001) also noted that previously proposeddescriptive schemes failed to resolve the homology of components of the pre-maxillary dentition among various genera of Characidae and Alestidae, whichtogether form the major portion of the Characidae of many previous authors.They consequently propose an alternative scheme based on the consistent toothpattern presented by P. argentea and by all but a few Creagrutus species. Thisscheme recognizes three tooth groups. A triad of teeth (element A), a series offour to seven, slightly to distinctly smaller teeth, whose arrangements rangefrom a nearly straight line, to an arch, or a sigmoid pattern (element B), and asingle, typically tricuspidate tooth (element C).
Although their system is easily recognizable in C. varii, the shape of itspremaxilla and of the arrangements of its dentition are those typical of othertetragonopterin fishes, especially Bryconamericus which has four inner row teethand five outer row teeth. In the case of the application of the DELTRANoptimization criterion, it is equally parsimonious to interpret that the toothpattern of Piabina and Creagrutus are independently acquired. In this situation,the two posteriormost teeth of element A, and the two posteriormost teeth ofelement B are considered to be elements of the inner row, and the remainingteeth as elements of the outer row. The premaxillary dentition of C. varii clearlysuggests an intermediate condition between the pattern present in most otherCreagrutus species and a Bryconamericus-like dentition, present in the putativeancestor of Piabina and Creagrutus.
Recently, Pavanelli & Britski (1999) suggested that the Rio Paranaıba basincorresponds to an area of endemism that has a fish fauna somewhat distinctfrom the remainder of the upper Rio Parana basin. Those authors proposedthat a more careful inventory of the Rio Paranaıba and of its tributaries couldreveal additional endemic species. The discovery of C. varii fits well withPavanelli & Britski’s (1999) prediction.
NEW CREAGRUTUS SPECIES FROM CENTRAL BRAZIL 609
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

Traditionally, the South American drainages have been divided into a seriesof biogeographic units or provinces, corresponding to the major hydrographicbasins (Vari, 1988; Menezes, 1996; Schaefer, 1997). According to Costa (2001),the delineation of these biogeographic regions seems to be intuitive, given thatthe criteria employed to delineate them were not explicitly stated. Costa (2001)also argued that, for at least north-eastern Brazil, various biogeographic studies(Vari, 1989; Vari et al., 1995; Costa, 1996; Schaefer, 1997) strongly suggestrelationships between this region and several adjacent drainages. Costa (2001)proposed that headwaters are typically areas of complex faunal interchangebetween adjacent regions rather than repeating components of the faunal stocksof a major unit represented by the basin as a whole.
The occurrence of C. varii in upper portions of the Rio Paranaıba basinsuggests the involvement of this area in early vicariant events prior to thediversification of the genus in the Tocantins, Araguaia and Amazon basins.None of the species of Creagrutus occurring in the Rio Tocantins and Araguaiabasins, the closest adjacent drainages, however, have resolved phylogeneticplacement within the phylogeny presented by Vari & Harold (2001). Insteadthey appear in a polytomy with the remaining species (except for C. figueiredoiVari & Harold, which is the sister group of C. cochui Gery, from the westernAmazon basin). Additional studies are needed in order to identify the biogeo-graphic history of the putative area of endemism of the Rio Paranaıba, and toresolve its relationships with both the rests of the upper Rio Parana andadjacent basins.
We thank R. Vari, F.A. Bockmann, M.R. de Carvalho and R.M.C. Castro forcomments and suggestions on the manuscript. This work was partially supported bythe State of Sao Paulo Research Foundation (FAPESP) within the BIOTA/FAPESP –The Biodiversity Virtual Institute Program (www.biota.org.br) through the ThematicProject ‘Diversidade de Peixes de riachos e cabeceiras da bacia do Alto Rio Parana noEstado de Sao Paulo, Brasil/Fish diversity of the headwater and streams of the UpperParana River system in State of Sao Paulo, Brazil’ (FAPESP grant No. 98/05 072-8 and00/1920-6) and by PRONEX ‘Conhecimento, Conservacao e Utilizacao Racional daDiversidade da Fauna de Peixes do Brasil’ (FINEP/CNPq grant No. 661 058/1997-2).
References
Costa, W. J. E. M. (1996). Phylogenetic and biogeographic analysis of the neotropicalannual fish genus Simpsonichthys (Cyprinodontiformes: Rivulidae). Journal ofComparative Biology 1, 129–140.
Costa, W. J. E. M. (2001). The neotropical annual fish genus Cynolebias (Cyprinodonti-formes: Rivulidae): phylogenetic relationships, taxonomic revision and biogeography.Ichthyological Exploration of Freshwaters 12, 333–383.
Harold, A. S. & Vari, R. P. (1994). Systematics of the trans-andean species of Creagrutus(Ostariophysi: Characiformes: Characidae). Smithsonian Contributions to Zoology551, 1–31.
Leviton, A. E., Gibbs, R. H., Jr., Heal, E. & Dawson, C. E. (1985). Standards inherpetology and ichthyology: Part I. Standard symbolic codes for institutionalresource collections in herpetology and ichthyology. Copeia 1985, 802–832.
Menezes, N. A. (1996). Methods for assessing freshwater fish diversity. In Biodiversity inBrazil: a First Approach (Bicudo, C. E. M. & Menezes, N. A., eds), pp. 289–295.Sao Paulo: CNPq.
610 A. C . RIBEIRO ET AL .
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611

Paiva, M. P. (1982). Grandes Represas do Brasil. Brasılia: Editerra.Pavanelli, C. S. & Britski, H. A. (1999). Description of a new species of Steindachnerina
(Teleostei: Characiformes: Curimatidae) from the upper Rio Parana basin, Brazil.Ichthyological Exploration of Freshwaters 10, 211–216.
Schaefer, S. A. (1997). The neotropical cascudinhos: systematics and biogeography of theOtocinclus catfishes (Siluriformes: Loricariidae). Proceedings of the Academy ofNatural Sciences of Philadelphia 148, 1–120.
Swofford, D. L. (1993). ‘‘PAUP’’: Phylogenetic Analysis Using Parsimony, Version 3.1.1.Washington, DC: Smithsonian Institution.
Taylor, W. R. & Van Dyke, G. C. (1985). Revised procedures for staining and clearingsmall fishes and other vertebrates for bone and cartilage study. Cybium 9, 107–109.
Vari, R. P. (1988). The Curimatidae, a lowland Neotropical fish family (Pisces:Characiformes); distribution, endemism, and phylogenetic biogeography. InProceedings of a Workshop on Neotropical Distribution Patterns (Vanzolini, P. E.& Heyer, W. R., eds), pp. 343–377. Rio de Janeiro: Academia Brasileira deCiencias.
Vari, R. P. (1989). Systematics of the neotropical characiform genus Curimata Bosc(Pisces: Characiformes). Smithsonian Contributions to Zoology 474, 1–63.
Vari, R. P. & Harold, A. S. (1998). The genus Creagrutus (Teleostei: Characiformes:Characidae): monophyly, relationships, and undetected diversity. In Phylogeny andClassification of Neotropical Fishes (Malabarba, L. R., Reis, R. E., Vari, R. P.,Lucena, Z. M. & Lucena, C. A. S., eds), pp. 245–260. Porto Alegre: Edipucrs.
Vari, R. P. & Harold, A. S. (2001). Phylogenetic study of the neotropical fishgenera Creagrutus Gunther and Piabina Reinhardt (Teleostei: Ostariophysi:Characiformes), with a revision of the cis-andean species. Smithsonian Contribu-tions to Zoology 613, 1–239.
Vari, R. P. & Lima, F. C. T. (2003). New species of Creagrutus (Teleostei: Characiformes:Characidae) from the Rio Uaupes basin, Brazil. Copeia 2003, 583–587.
Vari, R. P., Castro, R. M. C. & Raredon, S. J. (1995). The Neotropical fish familyChilodontidae (Teleostei: Characiformes): a phylogenetic study and a revision ofCaenotropus Gunther. Smithsonian Contributions to Zoology 577, 1–32.
Weitzman, S. H. (1962). The osteology of Brycon meeki, a generalized characid fish, withan osteological definition of the family. Stanford Ichtyological Bulletin 8, 1–77.
NEW CREAGRUTUS SPECIES FROM CENTRAL BRAZIL 611
# 2004TheFisheries Society of theBritish Isles, Journal of FishBiology 2004, 64, 597–611
Copyright © 2022 FDOKUMEN
















