
A new protocol for the collection and cataloguing of reference material for the study of fossil...
19
A new protocol for the collection and cataloguing of reference material for the study of fossil Cyperaceae fruits: The Modern Carpological Collection Edoardo Martinetto a, ⁎, Daniela Bouvet b , Elena Vassio a , Paola Magni c , Pedro Jiménez-Mejías d,e a Department of Earth Sciences, University of Turin, Via Valperga Caluso 35, 10125 Torino, Italy b Department of Life Sciences and Systems Biology, University of Turin, Viale Mattioli 25, 10125 Torino, Italy c Centre for Forensic Science, University of Western Australia, 35 Stirling Hwy, Crawley 6009, Australia d Botany Area, Department of Molecular Biology and Biochemical Engineering, Pablo de Olavide University, Ctra de Utrera, Km 1s.n., 41013 Seville, Spain e Department of Biodiversity and Conservation, Real Jardín Botánico de Madrid, Plaza de Murillo 2, 28014 Madrid, Spain abstract article info Article history: Received 30 May 2013 Received in revised form 22 November 2013 Accepted 25 November 2013 Available online 7 December 2013 Keywords: monocotyledons carpology taxonomy protocol Northern Italy taphonomy Fossil fruits of the species-rich family Cyperaceae (Poales) are frequently found in Cenozoic sediments of Europe and their taxonomical analysis is particularly difficult and poorly practised, because each fossil has to be com- pared with several tens of modern samples in order to pinpoint its nearest living relatives. This is only possible by using collections specifically prepared for archaeo- or palaeobotanical studies. We illustrate here the current status and organisation of one of these collections, developed in the frame of a more general “Modern Carpological Collection” (MCC) of the Turin University. Recent efforts in sampling, identifying and characterising cyperaceous fruits in Northern Italy permitted the gathering of 411 samples, covering 80% of the ca. 200 species spontaneously growing in this territory. Due to the still dynamic taxonomy, the large number of species and to the broad intraspecific variability of fruit morphology, we kept a very detailed trace of the sample's origin, by indicating whether and how they were related to the herbarium material. Modern samples from soils and sedi- ments have also been collected, being affected by taphonomic processes that make modern fruits more similar and readily comparable to palaeobotanical samples. In order to test the usefulness of the collection for the iden- tification of fossils, we selected some fossil samples of Cyperaceae fruits extracted from Pleistocene and Holocene sediments and we compared them with all the MCC samples containing similar material. The result of this trial demonstrates the advantage in using specimen-rich collections, such as the MCC, for the identification of fossils, against using only the literature and/or specimen-poor reference collections. We suggest that the procedures adopted for the MCC could become a standard protocol in collecting and cataloguing reference material for the study of fossil Cyperaceae fruits. Besides the possible future development of this relatively small and local collec- tion, an even more promising perspective is seen in the networking activity amongst carpological collections, pointing to the compilation of a large worldwide list of comparative material available for archaeo- and palaeobotanical studies. Such an easy access to an abundant reference material would certainly favour the prog- ress of fruit-morphological studies; moreover, the integration with molecular analyses would presumably yield a better definition of the phylogenetic history of the Cyperaceae within a time context. © 2013 Elsevier B.V. All rights reserved. 1. Introduction The family Cyperaceae (Poales) is the third species-richest family amongst the monocots (Joppa et al., 2011), comprising approximately 5600 species, 40% of which belong to the genus Carex L., the most diverse angiosperm genus of the temperate zones (Escudero et al., 2012). The fossil record of Cyperaceae is very rich in the Cenozoic of Europe (see Kats et al., 1965; Van der Burgh, 1978; Mai and Walther, 1988; Velichkievich and Zastawniak, 2006; Smith et al., 2009; Mai, 2010; Jiménez-Mejías and Martinetto, 2013 and references therein). However, the taxonomic knowledge of these fossils is rather poorly de- veloped in contrast to other families, usually much less species-rich (e.g.: Najadaceae, Potamogetonaceae: Velichkievich and Zastawniak, 2006), because the taxonomy of fossil remains is frequently done super- ficially (e.g.: Cavallo and Martinetto, 2001). When working on Cenozoic fruits and seeds, the lack of adequate modern reference material is one of the major problems for the precise definition of the taxonomical position of ancient remains; this is particularly true for such a highly diverse and globally distrib- uted family, in which carpological features are considered not as important as other characters (see below). Archaeobotanists and palaeobotanists are usually compelled to spend a long time in several Review of Palaeobotany and Palynology 201 (2014) 56–74 ⁎ Corresponding author. E-mail address: [email protected] (E. Martinetto). 0034-6667/$ – see front matter © 2013 Elsevier B.V. All rights reserved. http://dx.doi.org/10.1016/j.revpalbo.2013.11.003 Contents lists available at ScienceDirect Review of Palaeobotany and Palynology journal homepage: www.elsevier.com/locate/revpalbo
Transcript of A new protocol for the collection and cataloguing of reference material for the study of fossil...

Review of Palaeobotany and Palynology 201 (2014) 56–74
Contents lists available at ScienceDirect
Review of Palaeobotany and Palynology
j ourna l homepage: www.e lsev ie r .com/ locate / revpa lbo
A new protocol for the collection and cataloguing of reference materialfor the study of fossil Cyperaceae fruits: The ModernCarpological Collection
Edoardo Martinetto a,⁎, Daniela Bouvet b, Elena Vassio a, Paola Magni c, Pedro Jiménez-Mejías d,e
a Department of Earth Sciences, University of Turin, Via Valperga Caluso 35, 10125 Torino, Italyb Department of Life Sciences and Systems Biology, University of Turin, Viale Mattioli 25, 10125 Torino, Italyc Centre for Forensic Science, University of Western Australia, 35 Stirling Hwy, Crawley 6009, Australiad Botany Area, Department of Molecular Biology and Biochemical Engineering, Pablo de Olavide University, Ctra de Utrera, Km 1s.n., 41013 Seville, Spaine Department of Biodiversity and Conservation, Real Jardín Botánico de Madrid, Plaza de Murillo 2, 28014 Madrid, Spain
⁎ Corresponding author.E-mail address: [email protected] (E. Marti
0034-6667/$ – see front matter © 2013 Elsevier B.V. All rihttp://dx.doi.org/10.1016/j.revpalbo.2013.11.003
a b s t r a c t
a r t i c l e i n f oArticle history:Received 30 May 2013Received in revised form 22 November 2013Accepted 25 November 2013Available online 7 December 2013
Keywords:monocotyledonscarpologytaxonomyprotocolNorthern Italytaphonomy
Fossil fruits of the species-rich family Cyperaceae (Poales) are frequently found in Cenozoic sediments of Europeand their taxonomical analysis is particularly difficult and poorly practised, because each fossil has to be com-pared with several tens of modern samples in order to pinpoint its nearest living relatives. This is only possibleby using collections specifically prepared for archaeo- or palaeobotanical studies. We illustrate here the currentstatus and organisation of one of these collections, developed in the frame of a more general “ModernCarpological Collection” (MCC) of the Turin University. Recent efforts in sampling, identifying and characterisingcyperaceous fruits in Northern Italy permitted the gathering of 411 samples, covering 80% of the ca. 200 speciesspontaneously growing in this territory. Due to the still dynamic taxonomy, the large number of species and tothe broad intraspecific variability of fruit morphology, we kept a very detailed trace of the sample's origin, byindicating whether and how they were related to the herbarium material. Modern samples from soils and sedi-ments have also been collected, being affected by taphonomic processes that make modern fruits more similarand readily comparable to palaeobotanical samples. In order to test the usefulness of the collection for the iden-tification of fossils, we selected some fossil samples of Cyperaceae fruits extracted from Pleistocene andHolocenesediments and we compared them with all the MCC samples containing similar material. The result of this trialdemonstrates the advantage in using specimen-rich collections, such as the MCC, for the identification of fossils,against using only the literature and/or specimen-poor reference collections. We suggest that the proceduresadopted for the MCC could become a standard protocol in collecting and cataloguing reference material for thestudy of fossil Cyperaceae fruits. Besides the possible future development of this relatively small and local collec-tion, an even more promising perspective is seen in the networking activity amongst carpological collections,pointing to the compilation of a large worldwide list of comparative material available for archaeo- andpalaeobotanical studies. Such an easy access to an abundant reference material would certainly favour the prog-ress of fruit-morphological studies;moreover, the integrationwithmolecular analyses would presumably yield abetter definition of the phylogenetic history of the Cyperaceae within a time context.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
The family Cyperaceae (Poales) is the third species-richest familyamongst the monocots (Joppa et al., 2011), comprising approximately5600 species, 40% of which belong to the genus Carex L., the mostdiverse angiosperm genus of the temperate zones (Escudero et al.,2012). The fossil record of Cyperaceae is very rich in the Cenozoic ofEurope (see Kats et al., 1965; Van der Burgh, 1978; Mai and Walther,1988; Velichkievich and Zastawniak, 2006; Smith et al., 2009; Mai,
netto).
ghts reserved.
2010; Jiménez-Mejías and Martinetto, 2013 and references therein).However, the taxonomic knowledge of these fossils is rather poorly de-veloped in contrast to other families, usually much less species-rich(e.g.: Najadaceae, Potamogetonaceae: Velichkievich and Zastawniak,2006), because the taxonomyof fossil remains is frequently done super-ficially (e.g.: Cavallo and Martinetto, 2001).
When working on Cenozoic fruits and seeds, the lack of adequatemodern reference material is one of the major problems for theprecise definition of the taxonomical position of ancient remains;this is particularly true for such a highly diverse and globally distrib-uted family, in which carpological features are considered not asimportant as other characters (see below). Archaeobotanists andpalaeobotanists are usually compelled to spend a long time in several

Fig. 1. Morphological comparison (step 2 of the identification process, see Material andmethods) between four MCC fruit samples stored into resealable polyethylene bags and,at the junction of the modern preparates, a fossil fruit sample to be identified: CCN5610,Boca, NW Italy, Pleistocene. A. Carex elata subsp. elata, San Carlo Canavese, NW Italy,MCC2071; B. Carex atrata subsp. aterrima, Locana, NW Italy, MCC1946; C. Carex canescens,Locana, NW Italy, MCC2128; D. Carex nigra, Locana, NW Italy, MCC2065. Scale bar: 5 mm.
57E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
herbaria and the success in picking up a significant fruit material isusually low (Nesbitt, 1990; Nesbitt et al., 2003), because botanicalspecimens are rarely collected at full ripening time (when the fruitstend to drop down). Of course, many extensive fruit and seed collec-tions have been established for several purposes (e.g.: IBPGR, 1982;Linington, 1997; Nesbitt et al., 2003; Smith et al., 2003; Bacchettaet al., 2004) but, to our knowledge, none of them allows a quicksimultaneous analysis of large quantities of Cyperaceae fruits, asoften required during archaeo- and palaeobotanical analyses. Asmall reference collection of Cyperaceae fruits was also started,during the 1980s, at the University of Turin, in the frame of a generalcollection of Spermatophyte fruits and seeds referred to as “ModernCarpological Collection” (MCC, first cited by Jiménez-Mejías andMartinetto, 2013). This was established in parallel to thepalaeocarpological collection CENOFITA (label CCN) developed inthe same institution (Martinetto and Vassio, 2010). At first glance,this small collection (MCC) would seem to present a local solution(very specific to one laboratory) for the morphological analysis ofCyperaceae fruits. Conversely, we suggest that a deeper insight inthe history and structure of the MCC would point out some intrigu-ing solutions that, if applied worldwide, would considerably im-prove the results of fruit-morphological studies, particularly thosecarried out on fossils. One of the interesting aspects of the MCC isrepresented by the accurate collection protocol, which will be de-scribed below and suggested for application to analogous collectionsdevoted to the study of Cenozoic fossil fruits.
Thus, the aims of the present paper are:
1) to exploit the experience gained in structuring theMCC collection inorder to suggest a new approach for collecting and cataloguingmodern carpological material (not necessarily cyperaceous). Thiswill be based on the clear ranking of samples according to differentcollection procedures (see below type α–ζ samples), most ofwhich require a strict connection between the carpological samplesand the herbarium material. The reasons for such an apparentlyover-accurate cataloguing system are several, i.e. to assure a reliableand constantly updatable identification of the reference material,due to the still dynamic taxonomy, to permit the interpretation offruit-morphological diversity (often high in the samples) as linkedto individual, population-scale or intraspecific variation, toinvestigate the transformation of fruits in the transition from life todeath assemblages, and finally to prepare an adequate collectionfor parallel, molecular and fruit-morphological, taxonomic studies;
2) to illustratewhich carpological characters useful for the taxonomyofthe Cyperaceae have been so far highlighted in the literature or havebeen detected by uswhen analysing thematerial of theMCC, aswellas other carpological collections;
3) to test the potential of the MCC material in identifying fossil speci-mens and to point out its strengths and weaknesses. The paperwill also discuss the practical utility of different storage devices fora more straightforward comparison of fossil and modern materials;and
4) to stimulate the collection and publication of accurately cataloguedand specimen-rich cyperaceous fruit samples, in order to assureadequate reference material for the future characterisation ofCenozoic materials, in particular Neogene ones (not treated inthis paper).
2. Material and methods
2.1. Material collection
The reference plants of the MCC were collected mainly in Italy andonly marginally across Europe (Belgium, Bulgaria, Finland, Franceincluding Corsica, United Kingdom, Greece, Poland, and Portugal).Additionally, several samples from other geographical regions were
incorporated into the collection thanks to accessions coming fromother institutions (Appendix 1). Voucher specimens collected in con-nection to a MCC fruit sample were identified by using specialised bib-liography (Fournier, 1961; Hess et al., 1967–72;Walters, 1980; Pignatti,1982; Egorova, 1999; Aeschimann et al., 2004; Aeschimann and Burdet,2005; Jermy et al., 2007; Luceño et al., 2008) and preferentially depos-ited into the herbarium TO, but a few samples were also deposited inMRSN, AO and UPOS (abbreviations according Thiers, 2013; see Appen-dix 1). Furthermore, the MCC also received a few fruit samples fromROV, UPOS and the private herbaria of C. Argenti, R. Řepka andA. Soldano. The contribution of several persons with a good knowledgeof the local floras (see Acknowledgements) has been fundamental forincreasing the taxonomic coverage of the MCC.
2.2. Storage of MCC material
The fresh MCC fruit samples were enclosed in a porous paper en-velope and left to dry in a warm environment (20–30 °C) for at leasttwo weeks. Subsequently, each sample was enclosed in a singletransparent box or resealable polyethylene bag. Actually, over time,several storage media have been employed for the MCC fruit andseed samples: glass tubes, paper envelopes, plastic boxes, slides formicropalaeontological preparations, and some specimens from sev-eral samples were glued on thick paper. The suitability of thesedifferent storage media for the study of fossil materials will bediscussed below. We will not deal with the strategies for long-termpreservation, since this has been the object of several papers(IBPGR, 1982; Nesbitt, 1990; Linington, 1997; Nesbitt et al., 2003;Bacchetta et al., 2004 and references therein).
In the period between 1985 and 2000 carpological samples wereinserted only into plastic boxes, without carefully controlling themoisture degree. Fungal infestation, affecting a few of these samples,

58 E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
has been attributed to incomplete dehydration before the closure of theboxes. Conversely, animal infestation was only detected in freshmaterial and probably disappeared soon after the closure of the sampleinto the box, because of oxygen depletion. Recently, all the sampleshave been moved from boxes to resealable polyethylene bags. Eachsample has been split into two parts in order to facilitate both long-term preservation and analysis of materials: a protected subsampleand a consultable subsample (Fig. 1). Detailed information about eachsample has been collected in a database (Appendix 1), which is keptin two departments of the Turin University (Earth Sciences and LifeSciences/Systems Biology), and will be available on the website http://www.dst.unito.it, soon after the issue of the present paper.
It should be noted that theMCC purposes do not require keeping thematerial viable as in germoplasm banks (IBPGR, 1982; Linington, 1997;Bacchetta et al., 2004). In some cases, a moderate decay of a part of thesample was specifically favoured in order to obtain a material similar towaterlogged fossil samples, the most common preservation type in theEuropean Cenozoic. This effect can be obtained by adding watercollected in natural environments into a plastic box or resealable bagwhere diaspores are left to macerate for a period ranging from a fewmonths to several years (see Carex nigra (L.) Reichard in Plate I, 5).This process can be simulated with quicker chemical treatments(Thomlison, 1984; Nesbitt et al., 2003; Birks, 2013), e.g. by means ofsodium hypochlorite. Another useful preparation of the consultablesubsamples concerned the cleaning of achenes by hand-rubbing andneedle preparation, the last one being very important to preserve the
Plate I.1-6. Examples of utricle (1–5, lower rows) and achene variation inCarex nigra. Figs. 1a, 3aUsseglio, NW Italy,MCC1899. The extraction of achenes by hand-rubbing (60 s) did not work wpeculiar feature of this individual. 2. Achenes from a soil sample (ε-MCC) collected under a denproduced thenaturally cleaned achenes of theupper row, sometimesdid not affect the long, nonAvila, Spain, MCC2063; notice the smaller length/width ratio in comparison to 3a–3 h. 5. Speciexpose the achenes, whose appearance became similar to fossil ones (compare 5c and 6d), Pianthe stipeless base with callus in 5d (arrow). 6. Fossil achenes assigned to C. nigra from a Pleisto
style for all its length (see Jiménez-Mejías and Martinetto, 2013). Thepreservation of delicate preparations (e.g.: cross-sections) from com-pression was obtained by inserting a small and rigid plastic box intothe resealable bag.
2.3. Carpological characters in Cyperaceae
Mesoscopic carpological characters, cited as taxonomically useful inCyperaceae (Walters, 1953; Nilsson and Hjelmquist, 1967; Berggren,1969; Egorova, 1999; Browning and Gordon-Gray, 2000; Lye, 2000;Pignotti, 2003; Cappers et al., 2006; Bojňanský and Fargašová, 2007;Hroudová et al., 2007; Ferreira and Eggers, 2008; Hefler and Longhi-Wagner, 2008; Luceño et al., 2008; Smith et al., 2009; Jiménez-Mejíasand Martinetto, 2013), have been exhaustively compiled. Also the vari-ation recorded in the plates of Ercole et al. (2012) has been considered.We re-examined representative fruit and utricle (see detail in Fig. 2)samples of theMCC collection to check the consistency of these char-acters amongst different species. In those cases where a completevoucher was lacking, the re-examination was performed to revisethe identification.
The foregoing analyses were carried out by means of a Wild M3Bstereomicroscope with ×20 and ×40 magnification. Plates' pictureshave been obtained by using a Leica M205 C optical microscope inte-grated with Leica Microsystems' Leica DFC290 HD digital camera andLeica Application Suite software.
and 3d showneedle-prepared achenes inside theutricles. 1.α-MCCsample fromLac Falin,ith this sample, so that needle preparation was employed; the contorted style (arrow) is ase population of C. nigra, Lac Falin, Usseglio, NW Italy, MCC1964. The partial decay, which-jointed, attenuated style (arrow in2a). 3. Pania di Corfino, Apennines,N Italy,MCC2197. 4.mens macerated in water for 20 years in an airtight box, subsequently rubbed by hands todella Mussa, NW Italy, MCC0252. Notice the narrowly conical style base (arrow in 5b) andcene sediment sample, Boca, NW Italy, CCN5610; Scale bars = 1 mm.

Fig. 2. Example of a plant of Carex pendula in the ripening season, from which the voucherand the fruit sample (α-MCC) have been collected (Ventimiglia, NW Italy: MCC2175). Inthe right bottom corner a detail of a fresh utricle of this plant, opened by means of needlepreparation, is shown. Thus the trigonous fruit (achene), with a distinct basal stipe(“carpophore”) and the complete style (broken) at the apex, has been exposed; scale bar:1 mm.
59E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
2.4. Other carpological collections
In order to assess the present status of carpological collections, inparticular those dedicated to palaeobotanical studies, we collecteddata available in the internet (see Discussion). Only some of thesecollections were personally consulted by the authors, in other caseswe collected, directly from the respective curators, information about
Table 1List of selected fossil samples (Plates I, II and IV) from the CENOFITA collection (Martinetto andVthe MCC (Plates I, II and III).
Species No. of specimenswithin the sample
Fossil locality
Bolboschoenus maritimus “sensu lato” 6 Bucine quarry (Central ItCarex caryophyllea 1 Garosso di Rivalta (NW ICarex cf. elata 40 Buronzo (NW Italy)Carex leporina 2 Butteri quarry (Central ICarex nigra 20 Boca (NW Italy)Carex cf. nigra 2 Piànico Sèllere (NE Italy)Carex pallescens 36 Casapinta-Masserano (NCarex pendula 2 Valeriano Creek (NE ItalyCarex pseudocyperus 44 (ach.), 86
(utricles)Colombari (NW Italy)
Carex remota 3 Garosso di Rivalta (NW ICarex cf. riparia 1 Butteri quarry (Central ICarex umbrosa 4 Garosso di Rivalta (NW ICarex vesicaria 95 Colombari (NW Italy)Eleocharis palustris vel E. mamillata 10 Garosso di Rivalta (NW ISchoenoplectus lacustris 428 Colombari (NW Italy)
the storage conditions and consultability of the carpological material,which will be discussed below.
2.5. Fossil samples and method of identification
The fossils selected for the test of taxonomic usefulness of theMCC –
limited to achenes or utricles of Cyperaceae – are represented byPleistocene and Holocene materials of the CENOFITA collection(Martinetto and Vassio, 2010; Vassio, 2012), sampled in Northern-Central Italy and previously studied by Vassio (2012). Hence, 13 fossilsamples (Table 1) were singled out in order to compare them tothe MCC samples (and a few MRSN samples) and to evaluate theirusefulness for the identification of fossils. Each fossil sample was re-analysed by evaluating the absence/presence of relevant characters(see Results and Table 2), according to a 3-step procedure:
− step 1: observation of a fossil sample at the stereomicroscope andconsultation of the iconography (Nilsson and Hjelmquist, 1967;Berggren, 1969; Cappers et al., 2006; Ercole et al., 2012; Birks,2013; etc., see above), thus creating a list of morphologically similarspecies (up to 20 species);
− step 2: rapid comparative analysis of fossil samples with consultableMCC samples (consultable subsamples prepared in resealableplastic bags), selected according to the list of morphologicallysimilar species (Fig. 1). This process ends with the selection ofthe modern species (normally a few samples) showing the bestfit to fossils, according to the most stable and diagnostic characters.Due to the possible high intraspecific variability (see Plate I, 1–5),the comparison is also carried out between the fossil(s) and allthe MCC samples of each similar modern species (up to 40 MCCsamples); and
− step 3: fine-scale comparison of fossil fruits with those of the MCCsamples showing the best fit, extracted from the resealable bags.
Finally, the taxonomy was proposed on the basis of the detection inthe fossils of the same characters (or character variation) seen in one ora few modern species.
3. Results
3.1. Storage media and easiness of comparison between fossil andmodern materials
In several collections mentioned by us (directly and indirectlyconsulted: see Table 2), the storage media used were: glass tubes,paper envelopes, plastic boxes and slides for micropalaeontological
assio, 2010; Vassio, 2012), which have been compared to themodern referencematerial of
Sedimentsample label
Age CENOFITA collectionnumber
aly) BCN1 B5V Late Pleistocene CCN6076taly) RV1 B3O Holocene CCN6419
BU20 Early Pleistocene CCN0695taly) BUT1 B6X Late Pleistocene CCN5942
BOQ2 C5P Pleistocene CCN5610BVC3 B4A Middle Pleistocene CCN5811
W Italy) CP1 B5W Late Pleistocene CCN6346) SCN1 C0G Holocene CCN6284
CLB1 B8L Late Pleistocene–Holocene CCN6370 (achenes),CCN6372 (utricles)
taly) RV1 B3O Holocene CCN6438taly) BUT1 B6X Late Pleistocene CCN5926taly) RV1 B3O Holocene CCN6443
CLB1 B8L Late Pleistocene–Holocene CCN6374taly) RV1 B3O Holocene CCN6455
CLB1 B8L Late Pleistocene–Holocene CCN6375

Table 2Main characters and states (if applicable) observed for themorphological characterization of the achenes and utricles of Cyperaceae samples. The very distinct characters for Cladium arenot included: stone-fruit with longitudinal sutures, rough and lustreless surface, surrounded by the coriaceous-fleshy pericarp. Compiled fromWalters (1953), Nilsson and Hjelmquist(1967), Berggren (1969), Egorova (1999), Browning and Gordon-Gray (2000), Pignotti (2003), Hroudová et al. (2007), Jermy et al., 2007, Ferreira and Eggers (2008), Hefler andLonghi-Wagner (2008), Luceño et al. (2008), Smith et al. (2009) and Jiménez-Mejías andMartinetto (2013), including also additional characters from the variation observed in the platesof Ercole et al. (2012).
Achene characters States
Outline (see Berggren, 1969) Elliptic, ovate, obovate, circular, pyriform, etc.Length (without style) [provide range of measures]Maximumwidth [provide range of measures]Length–width ratio (L:W) [provide range of values]Outline of faces Elliptic, ovate, obovate, circular, different from face to faceEdges Sharp, rounded (Plate III, 9), thickened and raised (Plate III, 4), winged only at the base (Plate III, 10), with a
shallow depression at both sidesEdges colour Same colour as the faces (Plate III, 6), differently coloured (Plate III, 4), with a central white streak (Plate III, 8a)Arches connecting the edges apically Apparent (Carex curvula), moderate, scarce, nullFlexa in outline (see Jiménez-Mejías and Martinetto, 2013) Only one flexum. two flexaCross-section shape Linear, lenticular, biconvex or elliptic, triangular. terete/quadrangular, flat–convex, circular, etc.Number of sides 2, 3, 4Adaxial–abaxial diversification None, poor, strong, very strongSides curvature Concave, straight, convexLongitudinal rib entity Apparent, moderate, scarceLongitudinal rib number [provide value]Transversal rib (or wrinkles) entity Apparent, moderate, scarce, continuous, interruptedTransversal rib (or wrinkles) continuity Continuous, interruptedTransversal rib (or wrinkles) edges Flattened, rounded, sharpApical angle (see Jiménez-Mejías and Martinetto, 2013) [measured on each face and expressed as mean value or range]Basal angle (see Jiménez-Mejías and Martinetto, 2013) [measured on each face and expressed as mean value or range]Epidermal cell size Visible at 20×, visible at 40×Epidermal cells shape Square, circular, rectangular, polygonalOuter layer consistency (outer anticlinal walls) Mucilaginous, ephemeral thin-membranaceous, chartaceous, vitreous, coriaceousAnticlinal wall protrusion Apparent, moderate, scarceSilica body size Visible at 20×, visible at 40×, visible at 40× after removal of the outer cell wall, not visible at 40× even after
removal of the outer cell wallPericarp total thickness [provide mean value]Pericarp layers relative thickness (e.g. Bolboschoenus) [provide ratio or mean value]Utricle remains at the base of manipulated achenes (in Carex; seeJiménez-Mejías and Martinetto, 2013)
None, a few, abundant
Callus at the base Absent, thin (Plate II, 2), absent or thin but never thick, thick, thick or thin but never absentOrientation of achene to spikelet rachis Side-oriented (e.g. Cyperus), edge-oriented (e.g. Pycreus)Style branch number Entire, bifid, trifid, 4-fidStyle fracture Tending to break at the base, tending to break at (e.g.) 1/5 achene length, deciduous and leaving a notchStyle lignification Only lignified at the base (less than 1/4 achene length), much lignified in the basal half and poorly lignified in
the distal one, much lignified for most its lengthStyle base (“beak”) relative width 1/8 to ½ achene width approx.Style base (“beak”) length Mostly 0.1 to 1.0 mmStyle shape Inconspicuous, shortly cylindrical, shortly conical, cylindrical; long cylindrical, long subcylindrical and
attenuated towards the apex, long subcylindrical and conspicuously widened towards the base; truncate,acute, double-ridged towards the apex
Style jointed (articulated) to the base Present or notStyle ornamentation Smooth, fimbriate (Fimbristylis)Stilopodium shape Short conical, long conical (Eleocharis austriaca), flat-triangular (E. ovata), conspicuously pyramidal (E.
multicaulis)Ratio stilopodium width/achene width [provide mean value]Ratio stilopodium length/achene length [provide mean value]Stilopodium ornamentation Smooth, scabrid (Rhynchospora)Style base ringed Present (Plate III, 5) or notSmall discus on the style Present (Carex umbrosa: Plate III, 4) or notMedian depression Present in some achenes or notLateral invaginations Present or notBase relative size ¼ to ¾ achene width approx.Base slightly winged Yes or notStipitate base (Plate III, 6) Straight or curvedLength of stipitate base 1/10 to ¼ achene length approx.Sub-stipitate base (Plate III, 2) Always present, null in most specimens, always lackingArticulated stipe at the base Yes (e.g. Carex pendula: Fig. 1) or notBase extended into a thin scariose and stipitate end Yes (e.g. Carex montana) or notPerianth bristles (Plate III, 13) Present (e.g. Eleocharis, Scirpus) or notPerianth bristle length ¼ to 6/4 achene length approx.Number of perianth bristles [provide value]Perianth bristles ornamentation Smooth, scabrid (Plate III, 13), spathulate (e.g. Fuirena), feather-like (e.g. Schoenoplectus litoralis)Direction of hooks on perianth bristles Downward (Plate III, 13), patent, upwardPerianth bristles ornamentation on sides Outer and inner sides, only outer side, only inner side
Main utricle characters States
Incomplete, not closed (Kobresia) Present or notPresence of rachilla (Carex curvula, C. oedipostyla) Present or notOutline (see Berggren, 1969) Elliptic, ovate, obovate, circular, etc.
60 E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74

Table 2 (continued)
Achene characters States
Length [provide range of measures]Width [provide range of measures]Length–width ratio (L:W) [provide range of values]Cross-section shape Flat, circular, trigonousProportion of utricle length occupied by the achene [provide range of values]Utricle enveloping achene Loosely, tightlyLustre Dull, lustrousColour [see Berggren (1969) for standard colour classification, and consider that colour usually changes in fossils]Consistency Weak and thin-chartaceous, thick-chartaceous, coriaceous, thick and spongyEpidermal cell size Visible at 20×, visible at 40×Ornamentation Smooth, hairs, bristles, acute papillae, rounded papillae, low papillae; all the utricle or only towards the topBeak thickness Distinctly thickened (Carex aquatilis), or notNerve number [provide range of values]Nerve thickness b1/20 utricle width or N1/20 utricle width [if possible provide actual measures]Style persistence Style elongated and persistent after ripening, or notBeak shape Inconspicuous, shortly cylindrical, shortly conical; long conical, long subcylindrical, cylindrical; curved, straight
(Carex flava gr.)Beak apex (Jermy et al., 2007) Split, bifid (teeth straight, curved or divergent: Plate III, 8b), notched, truncated, obliquely truncate; with
scabrules, or not; distinctly whitish hyaline-margined, or notLongitudinal incision on dorsal side of beak symmetrical, asymmetrical (Carex vulpina)Spines or scabrules on beak Always present, present in most specimens (Plate II, 6a, 6b), null in most specimens, always lackingAdaxial–abaxial diversification None, poor, strong, very strongApical angle [provide range of measures]Marginal wing Always present, present in most specimens, null in most specimens, always lacking, only at topSub-stipitate base (Plate III, 8b) Always present, null in most specimens, always lackingStipitate base (Plate II, 6a) Always present, null in most specimens, always lacking
Main utricle characters
61E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
preparations. The trial of all of these storage media for MCC samplesallows us to discuss their usefulness degree when comparing fossiland modern materials.
Modern reference samples preserved in glass tubes, paper envelopesand most types of plastic boxes need to be extracted and placed closeto the fossil for an accurate morphological comparison. As for theCyperaceae fruits, two peculiarities must be highlighted: 1) there arevery small morphological differences between fruits of several species,that often compel the analyst to match a fossil fruit sample with up to10–30 extant species; 2) the morphological variability of fruits fromdifferent populations within the same species may be very large (seeBerggren, 1969; Egorova, 1999; Ercole et al., 2012), which implies thatseveral modern fruit samples (see Plate I, 1–5) of an individual specieshave to be placed side by side with the fossil specimens (Plate I, 6) forthe detection of the most similar ones.
Both of the foregoing operations imply a considerable risk ofsample mixing. In our experience, the use of transparent resealablepolyethylene bags facilitates the simultaneous observation of severalfruit samples either belonging to different species (Fig. 1) or to dif-ferent populations of a single species (Plate I, 1–5). Moreover, thisstorage method avoids the extraction of fruits (even for observationunder the stereomicroscope) at least in the first two steps of theidentification process employed by us (see above), right whenthere is a major risk of sample mixing. It also makes easier the peri-odical control of the samples' conservation status (animal and fungalparasites).
3.2. Sample collection and cataloguing: the MCC protocol
The final result of the preparation of several hundreds of modernfruit samples, and the constant need to improve their adequacy in theidentification of fossils, is represented by the assessment of the protocolwhich is described here. In theMCC each fruit sample is designated by acatalogue number (e.g.: MCC0001), which is also marked on its relatedherbarium sheet, with the addition of the letter “h” (i.e.: MCC0001/h).The quantity of the material collected for each sample was as abundantas possible (with the exception of rare plants), in order to permitdestructive preparations and various analyses, and to keep also asidesome catalogued material for scientific exchanges. The main peculiarityof the MCC protocol consists in keeping a very detailed trace of the
sample's origin, also indicatingwhether and howeach of them is relatedto the herbarium material. The reasons for this need of accuracy havebeen stated above. We defined six different types of samples on thebasis of their origin and the existence of complete voucher materialfor reference (Table 2). The samples from living plants linked to avoucher specimen (Fig. 2), presumably belonging to a single geneticspecimen (genet), are considered “type α” samples (in short, α-MCC).Given that, the voucher and the MCC sample would indeed representthe production of a single genome, with an obvious advantage for paral-lel molecular and fruit-morphological studies.
In case of less fruit-productive plants (e.g.: Carex pauciflora Lightf.)the collection was made by gathering a few neighbouring specimensand/or fruiting stems. These samples are considered “type β” samples(β-MCC). In the light of the current phylogeographic knowledge, thistype ofMCC samplesmay include fruits of plantswith significantly differ-ent genomes, as demonstrated to happen in contact (genome-merging)areas of different lineages of several Carex species (Schönswetter et al.,2008; Escudero et al., 2010; Jiménez-Mejías et al., 2012b): and this is tobe considered for the interpretation of fruit-morphological variationwithin the sample.
In case of absence of a voucher specimen straight-linked to the fruitsample, we define two additional sample types: “type γ” (γ-MCC), fruitspecimens of taxa documented by herbarium material collected in thesurroundings of the γ-MCC sampling site (not further than 10 kmapart), and “type δ” (δ-MCC), when only the fruits of a given individualor monospecific population have been collected (Table 3), being thewhole plant identified in the nature or in cultivation.
The “type ε” samples (ε-MCC) comprise fruit specimens which havebeen collected from the soil, the litter or the sediment closely linked to aliving plant individual or standing population, from which a vouchershould be collected.
The “type ζ” samples (ζ-MCC) are recovered from mixed plant de-bris transported far-away from the mother plant (not detectable),often in sediments or soils. The few ζ-MCC samples so far cataloguedoriginate from modern concentrations of fruits and seeds (Vassio,2012; Vassio and Martinetto, 2012; Bertolotto et al., 2012) collectedalong brooks, rivers and lakeshores. Their identification only reliesupon the fruit-morphological characterisation, but the circumscriptionof the local flora facilitates assignment to a definite species, detectedin the surroundings of the collection place.

62 E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
The usefulness of type ε and type ζ samples (Plate II, 3) in the studyof fossils (Plate II, 4 and 5) is due to the fact that they have already beenaffected by decay processes (Smith et al., 2009; Vassio, 2012; Vassio andMartinetto, 2012), which also occur in the first steps of the fossilisation.Therefore, type ε–ζ sample fruits (Plates I, 3 and II, 3) are readily compa-rable to analogous fossil specimens (Plates I, 6 and II, 5), even as for thefine details of the cell structure. Conversely, the appearance of fruitsfrom type α–δ samples (Plate II, 1 and 2) is always rather differentwith respect to the homologous fossils (Plate II, 4 and 5) because it isconditioned by the presence of decay-prone elements. It should benoted that the utricle is rarely preserved at the fossil state (Godwin,1956; Nilsson and Hjelmqvist, 1967; Dickson, 1970) and, when present(Plate II, 4), it is so brittle to be frequently destroyed by the preparationmethods which require sediment dissolution or removal.
The use of a short code (α-MCC, β-MCC, etc.) for each fruit sampletype is very useful for a quick and precise labelling of new carpologicalsamples collected in the field.
3.3. Present status of the MCC
Presently, theMCC contains 2039 carpological samples representing1473 taxa, belonging to 668 genera and 164 families. The collection isrich in Cyperaceae, Rosaceae, Hypericaceae, Ranunculaceae andLamiaceae species, because they are commonly found in those late Ce-nozoic carpological assemblages for whose identification the MCC wasbuilt up.
Most of the material has been collected from living plants, eitherspontaneous or in cultivation, and a minor quantity of accessions tothe MCC have been obtained through collaboration with other institu-tions yielding carpological collections, namely the CNR-IDPA researchinstitute of Bergamo (Consiglio Nazionale delle Ricerche - Istituto perla Dinamica dei Processi Ambientali) and the Natural Science Museumof Turin (Museo Regionale di Scienze Naturali). All of these accessionsare invariably δ samples. Regarding the family Cyperaceae, the MCCpresently includes 411 catalogued samples (plus ca. 100 not yetcatalogued), represented by 154 type α samples, 97 type β, 35 type γ,114 type δ, 5 type ε and 6 type ζ. The implementation of the MCC inthe last four years permitted cataloguing 80% of the Cyperaceae speciesoccurring in the North Italian flora (ca. 200 species after Conti et al.,2005, 2007), most of which are represented by type α, β or γ samples(associated to a voucher), since only 10 species are exclusively repre-sented by type δ samples (not associated to a voucher). For the remain-ing 20% of the north Italian species, the situation is such that either thematerial has been collected, but not yet catalogued, or it has been im-possible to collect the material because a few species are rare andhave a very localised distribution (e. g., Carex dioica L., Carex stenophyllaWahlenb., Carex supina Wahlenb., and Cyperus michelianus (L.) Delile).
Table 3Distinction of six types of fruit and seed samples (α–ζ) of the Modern Carpological Collectioncarpological collections.
Sampletype
Origin of carpological material Herbarium material
α Single living individual Individual documented bysimultaneously collected vo
β Group of living plants from a single populationassigned to a same taxon
Population documented bysimultaneously collected vo
γ Group of living plants assigned to a same taxon Population documented bynot simultaneously collecte
δ Living plants, in undetermined number No voucher
ε Soil, litter or roots of a standing population, referableto the standing plants through the voucher
Population documented by
ζ Fruits concentrations in brooks, rivers, lakes or soils,not referable a standing plant
No voucher
The refinement of the collection protocol also permitted gathering alarger number of specimen-rich samples, so that 165 samples havebeen classified as available for scientific exchanges (Appendix 1), with-out causing a significant loss to the bulk of the collection.
The account of the collection sites points out that most of the MCCsamples (80%) were collected in Italy, and the 18% in the rest ofEurope. A very subordinate part (2%) originated fromother sites outsideEurope (Canada and Japan). In the European context, Italy is the mostspecies-rich country regarding Carex (Jiménez-Mejías et al., 2012b),and also for the rest of the Cyperaceae genera, as up to 18 out of the 20native European genera are present in the Italian territory (Jiménez-Mejías and Luceño, 2011).
3.4. Taxonomic value of carpological characters in Cyperaceae andevaluation in the fossil record
Characters detected as taxonomically useful in our bibliographicalrevision are shown in Table 2. Indeed, the joint use of the works ofKats et al. (1965), Nilsson and Hjelmquist (1967), Berggren (1969),Cappers et al. (2006), Velichkievich and Zastawniak (2006), Bojňanskýand Fargašová (2007), Ercole et al. (2012) and Birks (2013) alreadyallows assessing the affinity of an unidentified cyperaceous fruit samplewith a restricted number of taxa from Europe, either modern or fossil.
The integration and revision of the mesoscopic fruit and/or utriclecharacters reported in such relevant literature revealed in some casesa really clear-cut characteristic morphology (e.g.: Carex leporina L.,Plate III, 1; Carex umbrosa Host, Plate III, 4; Carex caryophyllea LaTourr.,Plate III, 5; Carex pseudocyperus L., Plate III, 8; and Schoenoplectuslacustris (L.) Palla, Plate III, 12). However, in most cases, the characterdifference between similar species is so faint, and characters are sovariable within each species, that further analyses on fruit sampleswould be required to ensure a definitive taxonomical assignment onlyon a carpological basis. The work of Jiménez-Mejías and Martinetto(2013) on European members of Carex sect. Phacocystis Dumortillet il-lustrates those needs to be covered: 1) to observe fine morphologicaldetails, not so apparent in the above-cited papers, such as the cell struc-ture or the style fracture; and 2) to include a larger number of speci-mens simultaneously, in order to quantify the variability of charactersat the scale of the individual, the population and the taxon (e.g.: the in-constant presence of spines on the utricle beak of Carex rufina Drejer:Plate II, 6a and b).
3.5. Intraspecific variation of achenes and utricles
Interestingly, the observation of rich MCC samples has revealedbroader variability as that so far described in the literature for severaltaxa, which undoubtedly can affect the taxonomic interpretation of
(MCC) of the Turin University. This classification is proposed also for application to other
Identification Genets
one or moreuchers
Carried out on voucher specimens Single
one or moreuchers
Carried out on voucher specimens Undetermined
one or more vouchersd
Carried out on voucher specimens, confirmedon the basis of carpological features
Undetermined
Field-observation of the plant, confirmed onthe basis of carpological features
Undetermined
one or more vouchers Carried out on voucher specimens, confirmedon the basis of carpological features
Undetermined
Carpological features, facilitated by thecircumscription of the local flora
Undetermined

Plate II. 1–3. Carex elata ssp. elata, aspect of fruit samples in different ripening and decay conditions: 1. unripe achene inside the utricle,MCC2040, typeα sample (α-MCC); 2. ripe achene,MCC2202 (α-MCC); 3a,b. views of the two sides of a decayed achene in a modern fluvial deposit, MCC2191 (ζ-MCC); 4, 5. fossil Carex cf. elata, Buronzo, NW Italy, Early Pleistocene,CCN0695; 4. utricle still held by the original sediment; note the impression of the achene outline inside. 5. Achene with apparent similarities to the modern specimes 3b; 6. Carex rufina,MCC2040 (α-MCC): (a) utricle without prickles; (b) utricle with prickles at the upper right margin. Scale bar = 1 mm.
63E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
fossil samples (see Discussion). The availability of different samples of asame taxon allows assessing which characters are species-specific, ratherthan limited to a single individual, population or local type. Ercole et al.(2012) and Jiménez-Mejías and Martinetto (2013) already showed intheir plates a good number of fruit specimens from a single sample, andthemorphologywithin a sample ranged fromvery constant (e.g.: achenesofCyperus eragrostis Lam. in Ercole et al., 2012) to extremely variable (e.g.:achenes of Carex liparocarposGaudin or utricles of Carex limosa L. in Ercoleet al., 2012; see also Plate II, 1–5 as for Carex nigra).
The preparation of several MCC samples from a single speciesallows a quick estimation of the intraspecific fruit and utricle variation(Pl. I, 1–5). Carex nigra, whose high genetic diversity is well-known(Jiménez-Mejías et al., 2012a,b), constituted a good example to illus-trate the intraspecific variability of achene and utricle morphology.Remarkably, due to its abundance and wide distribution, it is also thespecies with the highest number of catalogued MCC samples (20)from several parts of Europe. Plate I represents the variability detectedin a selection of four achenes and four utricles of five modern samples(Plate I, 1–5) from three distinct geographic areas (Spain, WesternAlps, and Apennines). This modern material is used for a morphologicalcomparison with Pleistocene achenes (catalogue number CCN5610)from the NW Italian locality Boca (Plate I, 6; Table 1), recognised assimilar to some populations of C. nigra. Utricle morphology is knownto be quite variable, as can be inferred from the contrast of the descrip-tions of different local floras (e.g.: Great Britain: Jermy et al., 2007;Iberian Peninsula: Luceño et al., 2008; Russia: Egorova, 1999). Regard-ing achenes, the analysis of the α-MCC samples demonstrated that a
definite (and preservable in fossils) character, such as the contortedstyle of sample MCC1899 (Plate I, 1b and 1d), can be present just in asingle individual; in fact this was detected neither in other specimensfrom the same site (Plate I, 2a–2d) nor in those from neighbouringsites (e.g.: Plate I, 5a–5c). Jiménez-Mejías and Martinetto (2013)already noted a considerable variation in outline and size amongst thedifferent C. nigra samples from different sites. They reported the not-jointed style (Plate I, 2a and 2b), that tends to break into an acute shortlyconical remain (Plate I, 5b), and the conspicuous, mostly thick, callusbearing utricle remains (Plate I, 5d) as the best diagnostic features.Variation of the selected modern utricles and achenes from the MCCsamples is in agreement with these observations (Plate I, 1–5). Whencompared, the fossil achenes are in themean slightly smaller thanmod-ern ones (post-burial alteration), more or less heavily deformed andwith sticking sand grains (white spots). The style is poorly preserved,excepted one specimen (Plate I, 6a)which bears amore conspicuous re-main of a progressively attenuated, non-jointed style. The style basevaries from a small papilla (Plate I, 6c, see 4b as a modern analogue)to shortly conical and somewhat acute (Plate I, 6d), to robust and broad-ly conical (Plate I, 6f), as also present in modern specimens (Plate I, 3aand 3c). The considerable variation of the style base dimensionsobserved in modern individuals (Plate I, 4b and 4c) is apparent in thefossils too (Plate I, 6f and 6g). On the contrary, the callus is preservedat the base ofmost of the fossil achenes, being rather thick in three spec-imens (Plate I, 6a, 6e and 6g), whereas other specimens bear a smallercallus (Plate I, 6b and 6c). The utricle remains are mostly not preserveddue to the taphonomic process, although scanty traces are still

64 E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
noticeable in two specimens (Plate I, 6d and 6e). The outline of the fossilachenes is considerably diverse, as it can be observed within singlemodern populations (Plate I, 2a–2d, 5a–5c).
After these observations, it can be concluded that variation indimensions and outline of the fossil achenes, together with the non-jointed style preserved in some specimens, the style base shape ofmost specimens, and the presence of a thick callus, permit assigningthe fossil achenes to the C. nigra-type. Additionally, by considering therange of variability of the fossil population, the geographic position,and the Pleistocene age, we definitely assign the fossils (Plate I,6a–6h) to Carex nigra. The broad variation of outline, style base andbasal callus in different specimens does not mean that they belong todifferent species, because an analogous variation has been observed inachenes from single modern individuals (Plate I, 4a–4d, 5a–5c).
3.6. Identification test of selected fossil samples
As stated above, one of the aims of the present work is representedby the preparation of adequate modern reference material to be usedfor the morphological characterisation of Neogene and Quaternaryfossils, exemplified by the Buronzo locality specimens shown in PlateII, 4 and 5 (CCN0695, Table 1). The identification of pre-Quaternarymaterials is complicated by the demonstrated occurrence of extincttaxa before 1 Ma (see Jiménez-Mejías and Martinetto, 2013, and refer-ences therein). In the present paper we tested the usefulness of theMCC as reference material for the identification of 13 fossil samples(see Table 1 for localities and ages), dating back to not more than1 Ma, and therefore presumably belonging to still living species. Foreach of the analysed samples we describe in detail the methodologicalsteps of fossil identification by comparison to themorphological charac-ters and character variation of the MCC samples.
Fossil sample 1 (Plate IV, 1)The two biconvex achenes available (catalogue number CCN5942)
are characterised by elliptic outline, with style base (“beak”) long ¼ ofthe achene length, jointed to a poorly lignified style. The overview ofthe literature indicated that biconvex achenes with jointed style(Jiménez-Mejías and Martinetto, 2013) are characteristic of subgen.Vignea. Furthermore, a style base (“beak”) long ¼ of the achene lengthand an elliptic outline as broad as in the fossils have been detected onlyin achenes of Carex leporina L., for example those figured by Capperset al. (2006), Bojňanský and Fargašová (2007) and Ercole et al. (2012).Cappers et al.'s achene differs for having the maximum width ¼ fromthe base, whereas it is at 3/4 in our fossils. In sample MCC2198 ofC. leporina we detected nearly identical achenes (Plate II, 1), with maxi-mum width at 3/4 from the base, mixed with achenes with maximumwidth at ¼ from the base, in the same α-MCC sample. This is a clearindication that the position of achene's maximum width is variablewithin a single individual in C. leporina.
In Europe (north) there is only another species closely related toCarex leporina (sect. Ovales Kunth): Carex macloviana d'Urv., whichdiffers from the fossils, based on the data of Berggren (1969), for a def-initely shorter style base. The similar achenes of Carex bohemica Schreb.,judging from the variation observed in sample MCC0219, are alwayssmaller and narrower. However, fossil achenes as broad and largeas our specimens were assigned to C. bohemica by Velichkievich and
Plate III.Modern fruits and utricles. 1. Carex leporina, achene, MCC2198; 2. Carex remota, achenCarex umbrosa, achene, MCC1866; 5. Carex caryophyllea, achene from a sediment sample (ζ-MMCC2180; 8. Carex pseudocyperus: (a) achene and (b) utricle, MCC0294; 9. Carex vesicariaMCC1996; 12. Schoenoplectus lacustris, dorsal view, MCC1104; 13. Eleocharis palustris, MCC197
Plate IV. Fossil achenes and utricles from the CENOFITA collection (code CCN), see Table 1 for lospecimens from the same sample, CCN5811; 4. Carex umbrosa, CCN6443; 5. Carex caryophylleachene, CCN6346; 8. Carex pseudocyperus: (a) achene, CCN6370 and (b) utricle, CCN6372; 9.Bolboschoenus maritimus “sensu lato”, CCN6076; 12. Schoenoplectus lacustris, dorsal view, CCN6Scale bars = 1 mm. (see on page 11)
Zastawniak (2006: pl. 55, figs. 8–15), so that a verification of achenevariation in a larger number of modern samples would be desirable.
Several American and East-Asian species of sect. Ovales, which maywell have morphological affinities with our fossils (Hoshino and Masaki,2011; eMonocot Team, 2013), have not been considered for comparison.
Diagnosis: Carex leporina.
Fossil sample 2 (Plate IV, 2)Also the achenes of this sample show the characteristic features of
subgen. Vignea (see above), but here the style junction is very close tothe base. Specimens with nearly identical shape and dimensions asthe three biconvex achenes of this sample (CCN6438) have been figuredby Cappers et al. (2006: Carex remota L.) and Ercole et al. (2012: Carexpraecox Schreb., C. remota). The analysis of all thematerials of these spe-cies in the MCC and in MRSN suggests that the fossils are identical toachenes of C. remota (Plate III, 2), whereas they differ from the very sim-ilar achenes of one sample of C. praecox (MRSN-SPER-2006) for anarrower style base. Berggren (1969) and Cappers et al. (2006) figuredachenes of C. praecox, which differmore consistently from our fossils fora distinctly stipitate base. All theMCC samples of the other species withsimilar achenes have been checked without evidence of a completemorphological agreement with the fossils.
Diagnosis: Carex remota.
Fossil sample 3 (Plate IV, 3a, 3b)It is represented by two biconvex and narrowly elliptic achenes
(CCN5811), whose characters, in particular the rough surface, broadand flat style base and the broad truncated base with a narrow basalangle (20–30°), were only detected in the achenes of Carex mucronataAll. figured by Ercole et al. (2012) (see also Plate III, 3a). However, thedimensions are considerably smaller and the style base is conical andattenuated instead of broad and flat with an abrupt termination. In afirst time the overview of all the MCC samples with biconvex achenesshowed only another species with moderately similar achenes, Carexpulicaris L., which was excluded for the definitely narrower style baseand smoother achene surface. The examination of the recent accessionsinto the MCC permitted detection of nearly identical achenes (Plate III,3b), also as for cell pattern, in a sample of Carex nigra from theApennines (MCC2197, see Plate I, 6). It should be pointed out that theC. nigra achenes depicted in the literature (Nilsson and Hjelmquist,1967; Berggren, 1969; Cappers et al., 2006; Bojňanský and Fargašová,2007; Ercole et al., 2012) are always definitely broader, so that onlythe availability of sample MCC2197 permitted assigning the fossils toC. nigra, a species for which a broad phenotypic plasticity is well-known (Jiménez-Mejías and Martinetto, 2013).
Diagnosis: Carex cf. nigra; the open nomenclature is cautiously usedbecause of the unusual outline of these fruits. A more detailed analysis(e.g. by means of the SEM) could provide additional diagnosticelements.
Fossil sample 4 (Plate IV, 4)The affinity of these fossils (CCN6443) with Carex umbrosa (Plate III,
4) was detected by comparison to the plates of Ercole et al. (2012),whereas achenes with this characteristic morphology are not treatedby Nilsson and Hjelmquist (1967), Berggren (1969) and Cappers et al.(2006). Fresh achenes of C. umbrosa show a typical disc on the style,
e, MCC2178; 3a. Carex mucronata, achene, MCC2195. 3b. Carex nigra, achene, MCC2197; 4.CC),-MCC2196; 6. Carex pendula, achene (ζ-MCC), MCC2176; 7. Carex pallescens, achene,, achene, MCC0316; 10. Carex riparia, achene, MCC0300; 11. Bolboschoenus maritimus,7. Scale bars = 1 mm. (see on page 10)
cality data. 1. Carex leporina, CCN5942; 2. Carex remota, CCN6438; 3a–b. Carex cf. nigra: twoa, CCN6419; 6. Carex pendula, achene with traces of utricle, CCN6284; 7. Carex pallescens,Carex vesicaria, CCN6374; 10. Carex cf. riparia, immature or sterile achene, CCN5926; 11.375; 13. Eleocharis palustris vel mamillata, CCN6455. Material revised after Vassio (2012).
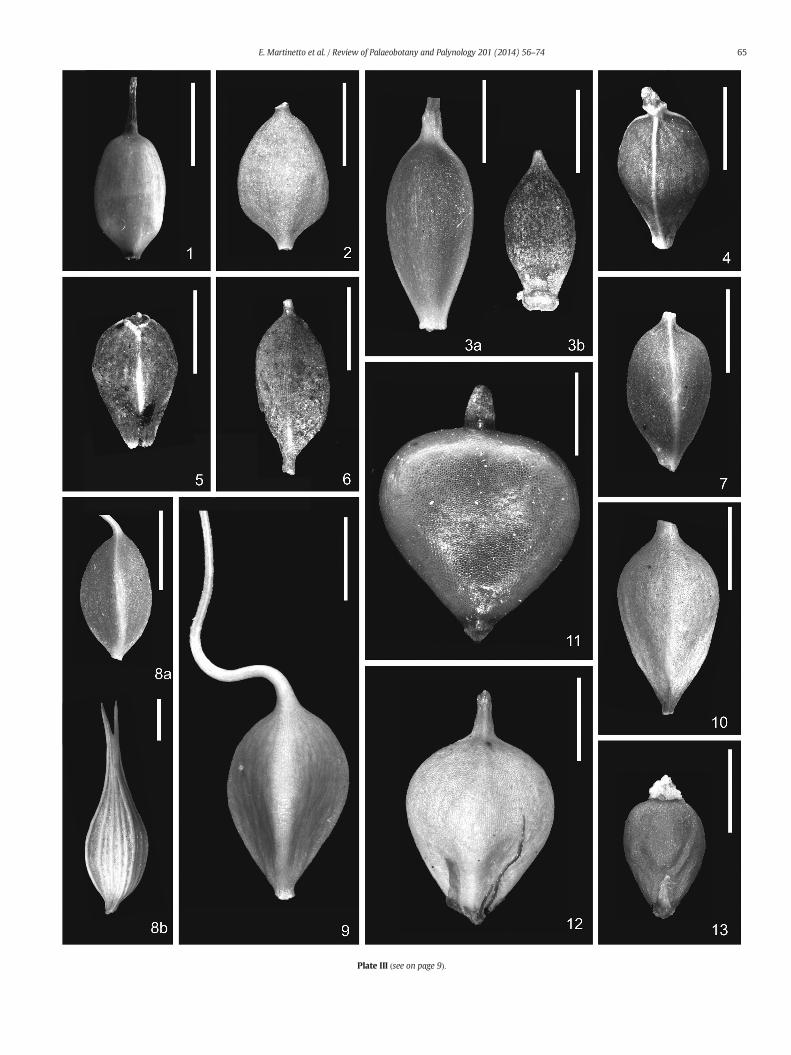
Plate III (see on page 9).
65E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74

Plate IV (see on page 9).
66 E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74

67E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
but this was not preserved in the fossils, and we verified in a ζ-MCCsample of this species (MCC2201) that the loss of style already occursduring early decay into the sediment. These remaining diagnostic fea-tures were considered: dimensions, shape, acute stipeless base, facespoorly folded, raised and clear-coloured edges, and a notch at the baseof the missing style (a definite clue for the original presence of a broaddecay-prone style base). After an overview of all the MCC sampleswith trigonous achenes of similar shape, we consider in particular thislast character as typical for sect. Mitratae Kük. (also characterised byraised and clear-coloured edges), Hallerianae (Arch. & Graebn.) Rouyand Digitatae (Fr.) H. Christ. Apart C. umbrosa, other two European spe-cies of sect.Mitratae (Carex caryophyllea, Carex depressa) differ from thefossils for a circular ridge at the achene's top (see below as for its trans-formation in the fossil state). Members of sect. Hallerianae and Digitataehave been excluded for the lack of raised and clear-coloured edges andthe presence of a stipitate base.
Diagnosis: Carex umbrosa.
Fossil sample 5 (Plate IV, 5)For this trigonous achene (CCN6419)with a flat platform at the apex
we found no match in the literature, however, identical achenes wereobtained by macerating with sodium hypochlorite the fruits of Carexcaryophyllea (MCC2033). So, the typical apical ridge that surroundsthe style base in the achenes of this species (Plate III, 5) is possibly notpreserved in the fossils. Thus, the identification has to be based on thebroad platform, which is left at the achene's top. Indeed, we suspectedthat this was the aspect of partly decomposed achenes of this species,because analogous fruits were found in a modern sediment sample(ζ-MCC; Plate IV, 5) collected in a place where C. caryophyllea is com-mon (Vassio, 2012). We also checked in the MCC all the other speciesof sect. Mitratae, Hallerianae and Digitatae (Appendix 2), but onlyCarex depressa Link, had a similar apical ridge and its smaller sizecould not leave such a large platform as observed in the fossil. ThetwoMCC samples of C. depressa also show the achene's faces abundantlyfolded, unlike the fossil.
Diagnosis: Carex caryophyllea.
Fossil sample 6 (Plate IV, 6)It is represented by two achenes, enriched by the presence of utricle
remains (CCN6284), which seem to tightly envelope the achene,and have a longitudinal vein in the middle of each of the three faces.With the additional character of a stipitate base of the achene, thissample could easily be characterised as belonging to sections Digitatae(Fr.) H. Christ. or Clandestinae G. Don. At this point a problem arises:the very small dimensions of the fossil achenes would exclude Carexdigitata L., Carex humilis Leyss., Carex ornithopoda ssp. ornithopodaWilld., and Carex pediformis C.A. Meyer (lacking into the MCC, butwhose achenes' size is described as larger by Bojňanský and Fargašová,2007). Amongst the European taxa checked, it could only matchC. ornithopoda ssp. ornithopodioides (Hausm.) Nyman. Yet, the thin andcylindrical style base of the fossil is absolutely not compatible with thethick and conical one of C. ornithopoda ssp. ornithopodioides. The studyof ε-MCC samples provided the solution of the problem: the soil sampleMCC2176 of Carex pendula Huds, with utricles originally broad and in-flated (see the α-MCC in Fig. 2), displays the utricle “tightly enveloping”the achene due to post-burial alteration. The control of all the othercharacters (dimensions, proportions, stipe, sharp edges, apparent cellpattern, etc.) confirmed that sample CCN6284 was identical to theC. pendula achenes (Plate III, 6). Many other species have very similarachenes (e.g.: Carex atrata L., Carex magellanica Lam. ssp. irrigua(Wahlenb.) Hiitonen, and Carex parviflora Host, but a careful analysisof their MCC samples did not provide evidence of a complete morpho-logical agreement with the fossils.
Diagnosis: Carex pendula.
ssil sample 7 (Plate IV, 7)
The over 30 achenes of this sample (CCN6346) lack highly diagnosticcharacters, and the general morphology resembles several species:Carex distans L., Carex hostiana DC., Carex mairei Coss. et Germ.,C. pallescens L., Carex pendula, Carex punctata Gaudin (see Plate III, 7)and, to a lesser extent, Carex binervis Sm., Carex flacca L., Carex laevigataSm., and Carex sylvatica Huds (only MCC2000). The mean achenedimensions of sample CCN6346, the width and length of style base, theconvex faces, the cell size and relief, the obtuse apical angle and thick-ness/curvature of the light-coloured edges,match only those of themod-ern achene populations of Carex pallescens (bestfit toMCC2180). In caseslike the present one, when several tens of achenes are available from asingle fossil sample (Table 1), its comparison to rich samples of allthose modern species, which bear similar achenes, is decisive. In thisway, even poorly apparent characters, detected after the examinationof a large amount of the sample, can be identified as constant and diag-nostic for a particular species.
Diagnosis: Carex pallescens.
Fossil sample 8 (Plate IV, 8)The trigonous nutlets of this sample (CCN6370), with a narrowly
elliptic (lenticular) outline have two features which are shared by a fewmodern European species: maximum width in the middle or slightlybelow, very thin non-jointed and attenuate style. The consultation ofthe literature permitted us to find achenes with the same characters insix species: Carex capillaris L., Carex microcarpa Bertol., Carex pallescens,Carex pendula, Carex pseudocyperus, and Carex strigosa Huds. Thedetailed comparison of the MCC material excludes five of this speciesfor the following main characters: presence of stipe (C. pendula,C. microcarpa, and C. strigosa), style base too short (C. capillaris,C. extensa Gooden., C. microcarpa, C. pallescens, and C. pendula). Therefore,the achenesmatch a singlemodern European species, and, when also theutricle is considered, the determination is even easier because onlyC. pseudocyperus (Plate III, 8b; CCN 6372) has a robust, many-nervedutricle.
Diagnosis: Carex pseudocyperus.
Fossil sample 9 (Plate IV, 9)The sample (CCN6374) is represented by 95 achenes whose shape
would seem typical for achenes of Carex hirta L., based on Nilsson andHjelmquist (1967), Berggren (1969), Cappers et al. (2006) andBojňanský and Fargašová (2007). Only Ercole et al. (2012) figuredsimilar, narrowly obovate achenes, with robust style base, for anotherspecies: Carex vesicaria L. In our opinion the determination of themate-rialwasnot possible on thebasis of the literature alone, sowe accuratelyanalysed the MCC samples of achenes of C. hirta, C. vesicaria and othersimilar species (Carex riparia Curtis, Carex. rostrata Stokes). The resultwas thatwe found the bestfit of the fossilswith specimens of C. vesicaria(Plate III, 9). As contrasting characters in C. hirta versus C. vesicaria wepropose the cylindrical vs. trigonous stipitate achene base and theroundly trigonous vs. circular section of the style base. When the styleis preserved completely the distinction is very easy (Nilsson andHjelmquist, 1967), since in C. hirta it is abruptly constricted (and oftenbroken) at ca. 1/2 of the achene length, in C. vesicaria it is progressivelyattenuated and often longer than the achene (Plate III, 9). These charac-ters were stable in the four MCC samples of C. hirta and in the two sam-ples of C. vesicaria (Appendix 2).
Diagnosis: Carex vesicaria.
Fossil sample 10 (Plate IV, 10)The characteristic morphology of this fossil (CCN5926), withwinged
edges at the base, did not match any figure available on atlases (Nilssonand Hjelmquist, 1967; Berggren, 1969; Cappers et al., 2006; Bojňanskýand Fargašová, 2007). After long-lasting examination of all sampleswith trigonous achenes in the MCC, nearly identical achenes werefound as unripe specimens amongst a modern sample of Carex riparia(Plate III, 10). The large cylindrical style base is an additional character
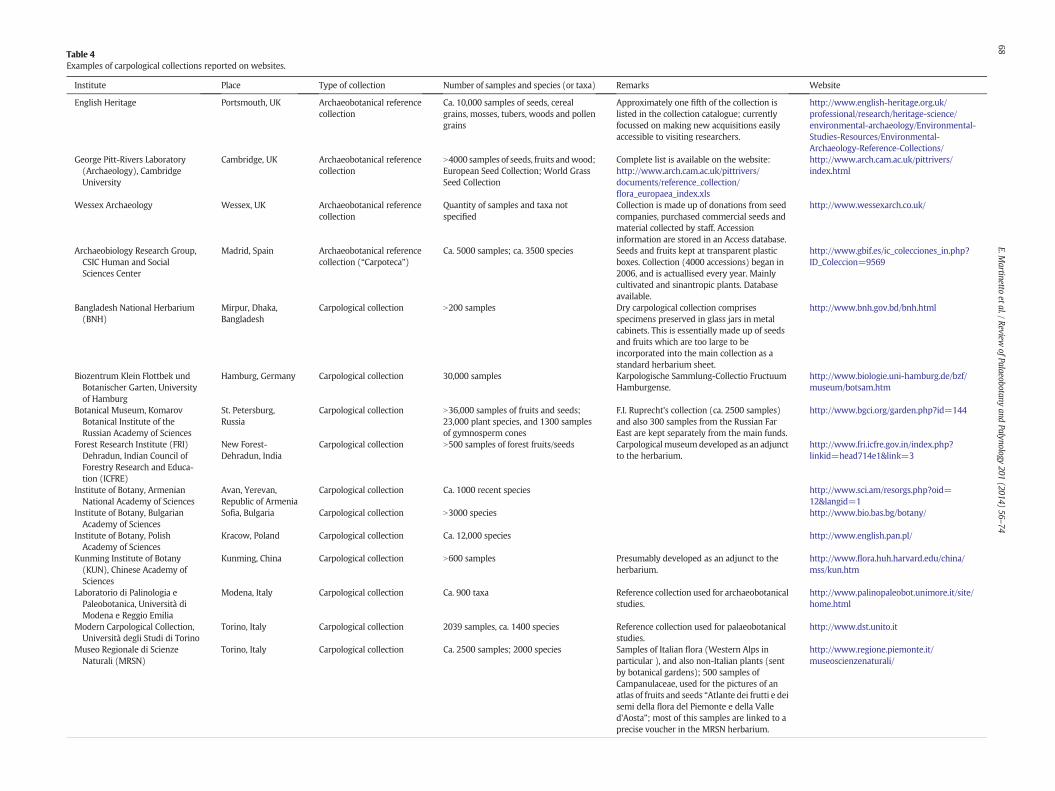
Table 4Examples of carpological collections reported on websites.
Institute Place Type of collection Number of samples and species (or taxa) Remarks Website
English Heritage Portsmouth, UK Archaeobotanical referencecollection
Ca. 10,000 samples of seeds, cerealgrains, mosses, tubers, woods and pollengrains
Approximately one fifth of the collection islisted in the collection catalogue; currentlyfocussed on making new acquisitions easilyaccessible to visiting researchers.
http://www.english-heritage.org.uk/professional/research/heritage-science/environmental-archaeology/Environmental-Studies-Resources/Environmental-Archaeology-Reference-Collections/
George Pitt-Rivers Laboratory(Archaeology), CambridgeUniversity
Cambridge, UK Archaeobotanical referencecollection
N4000 samples of seeds, fruits andwood;European Seed Collection; World GrassSeed Collection
Complete list is available on the website:http://www.arch.cam.ac.uk/pittrivers/documents/reference_collection/flora_europaea_index.xls
http://www.arch.cam.ac.uk/pittrivers/index.html
Wessex Archaeology Wessex, UK Archaeobotanical referencecollection
Quantity of samples and taxa notspecified
Collection is made up of donations from seedcompanies, purchased commercial seeds andmaterial collected by staff. Accessioninformation are stored in an Access database.
http://www.wessexarch.co.uk/
Archaeobiology Research Group,CSIC Human and SocialSciences Center
Madrid, Spain Archaeobotanical referencecollection (“Carpoteca”)
Ca. 5000 samples; ca. 3500 species Seeds and fruits kept at transparent plasticboxes. Collection (4000 accessions) began in2006, and is actuallised every year. Mainlycultivated and sinantropic plants. Databaseavailable.
http://www.gbif.es/ic_colecciones_in.php?ID_Coleccion=9569
Bangladesh National Herbarium(BNH)
Mirpur, Dhaka,Bangladesh
Carpological collection N200 samples Dry carpological collection comprisesspecimens preserved in glass jars in metalcabinets. This is essentially made up of seedsand fruits which are too large to beincorporated into the main collection as astandard herbarium sheet.
http://www.bnh.gov.bd/bnh.html
Biozentrum Klein Flottbek undBotanischer Garten, Universityof Hamburg
Hamburg, Germany Carpological collection 30,000 samples Karpologische Sammlung-Collectio FructuumHamburgense.
http://www.biologie.uni-hamburg.de/bzf/museum/botsam.htm
Botanical Museum, KomarovBotanical Institute of theRussian Academy of Sciences
St. Petersburg,Russia
Carpological collection N36,000 samples of fruits and seeds;23,000 plant species, and 1300 samplesof gymnosperm cones
F.I. Ruprecht's collection (ca. 2500 samples)and also 300 samples from the Russian FarEast are kept separately from the main funds.
http://www.bgci.org/garden.php?id=144
Forest Research Institute (FRI)Dehradun, Indian Council ofForestry Research and Educa-tion (ICFRE)
New Forest-Dehradun, India
Carpological collection N500 samples of forest fruits/seeds Carpological museumdeveloped as an adjunctto the herbarium.
http://www.fri.icfre.gov.in/index.php?linkid=head714e1&link=3
Institute of Botany, ArmenianNational Academy of Sciences
Avan, Yerevan,Republic of Armenia
Carpological collection Ca. 1000 recent species http://www.sci.am/resorgs.php?oid=12&langid=1
Institute of Botany, BulgarianAcademy of Sciences
Sofia, Bulgaria Carpological collection N3000 species http://www.bio.bas.bg/botany/
Institute of Botany, PolishAcademy of Sciences
Kracow, Poland Carpological collection Ca. 12,000 species http://www.english.pan.pl/
Kunming Institute of Botany(KUN), Chinese Academy ofSciences
Kunming, China Carpological collection N600 samples Presumably developed as an adjunct to theherbarium.
http://www.flora.huh.harvard.edu/china/mss/kun.htm
Laboratorio di Palinologia ePaleobotanica, Università diModena e Reggio Emilia
Modena, Italy Carpological collection Ca. 900 taxa Reference collection used for archaeobotanicalstudies.
http://www.palinopaleobot.unimore.it/site/home.html
Modern Carpological Collection,Università degli Studi di Torino
Torino, Italy Carpological collection 2039 samples, ca. 1400 species Reference collection used for palaeobotanicalstudies.
http://www.dst.unito.it
Museo Regionale di ScienzeNaturali (MRSN)
Torino, Italy Carpological collection Ca. 2500 samples; 2000 species Samples of Italian flora (Western Alps inparticular ), and also non-Italian plants (sentby botanical gardens); 500 samples ofCampanulaceae, used for the pictures of anatlas of fruits and seeds “Atlante dei frutti e deisemi della flora del Piemonte e della Valled'Aosta”; most of this samples are linked to aprecise voucher in the MRSN herbarium.
http://www.regione.piemonte.it/museoscienzenaturali/
68E.M
artinettoetal./Review
ofPalaeobotanyand
Palynology201
(2014)56
–74
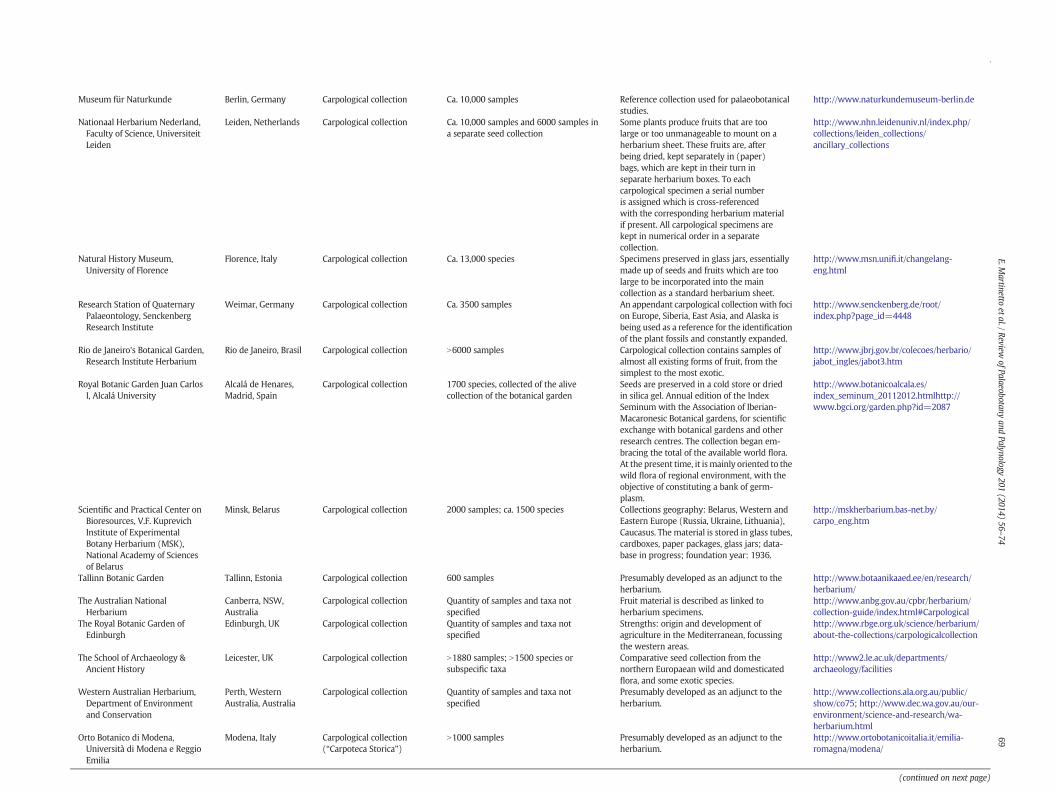
Institute
Place Type of collection Number of samples andspecies (or taxa)
Remarks Website
Museum für Naturkunde Berlin, Germany Carpological collection Ca. 10,000 samples Reference collection used for palaeobotanicalstudies.
http://www.naturkundemuseum-berlin.de
Nationaal Herbarium Nederland,Faculty of Science, UniversiteitLeiden
Leiden, Netherlands Carpological collection Ca. 10,000 samples and 6000 samples ina separate seed collection
Some plants produce fruits that are toolarge or too unmanageable to mount on aherbarium sheet. These fruits are, afterbeing dried, kept separately in (paper)bags, which are kept in their turn inseparate herbarium boxes. To eachcarpological specimen a serial numberis assigned which is cross-referencedwith the corresponding herbarium materialif present. All carpological specimens arekept in numerical order in a separatecollection.
http://www.nhn.leidenuniv.nl/index.php/collections/leiden_collections/ancillary_collections
Natural History Museum,University of Florence
Florence, Italy Carpological collection Ca. 13,000 species Specimens preserved in glass jars, essentiallymade up of seeds and fruits which are toolarge to be incorporated into the maincollection as a standard herbarium sheet.
http://www.msn.unifi.it/changelang-eng.html
Research Station of QuaternaryPalaeontology, SenckenbergResearch Institute
Weimar, Germany Carpological collection Ca. 3500 samples An appendant carpological collectionwith focion Europe, Siberia, East Asia, and Alaska isbeing used as a reference for the identificationof the plant fossils and constantly expanded.
http://www.senckenberg.de/root/index.php?page_id=4448
Rio de Janeiro's Botanical Garden,Research Institute Herbarium
Rio de Janeiro, Brasil Carpological collection N6000 samples Carpological collection contains samples ofalmost all existing forms of fruit, from thesimplest to the most exotic.
http://www.jbrj.gov.br/colecoes/herbario/jabot_ingles/jabot3.htm
Royal Botanic Garden Juan CarlosI, Alcalá University
Alcalá de Henares,Madrid, Spain
Carpological collection 1700 species, collected of the alivecollection of the botanical garden
Seeds are preserved in a cold store or driedin silica gel. Annual edition of the IndexSeminum with the Association of Iberian-Macaronesic Botanical gardens, for scientificexchange with botanical gardens and otherresearch centres. The collection began em-bracing the total of the available world flora.At the present time, it ismainly oriented to thewild flora of regional environment, with theobjective of constituting a bank of germ-plasm.
http://www.botanicoalcala.es/index_seminum_20112012.htmlhttp://www.bgci.org/garden.php?id=2087
Scientific and Practical Center onBioresources, V.F. KuprevichInstitute of ExperimentalBotany Herbarium (MSK),National Academy of Sciencesof Belarus
Minsk, Belarus Carpological collection 2000 samples; ca. 1500 species Collections geography: Belarus, Western andEastern Europe (Russia, Ukraine, Lithuania),Caucasus. The material is stored in glass tubes,cardboxes, paper packages, glass jars; data-base in progress; foundation year: 1936.
http://mskherbarium.bas-net.by/carpo_eng.htm
Tallinn Botanic Garden Tallinn, Estonia Carpological collection 600 samples Presumably developed as an adjunct to theherbarium.
http://www.botaanikaaed.ee/en/research/herbarium/
The Australian NationalHerbarium
Canberra, NSW,Australia
Carpological collection Quantity of samples and taxa notspecified
Fruit material is described as linked toherbarium specimens.
http://www.anbg.gov.au/cpbr/herbarium/collection-guide/index.html#Carpological
The Royal Botanic Garden ofEdinburgh
Edinburgh, UK Carpological collection Quantity of samples and taxa notspecified
Strengths: origin and development ofagriculture in the Mediterranean, focussingthe western areas.
http://www.rbge.org.uk/science/herbarium/about-the-collections/carpologicalcollection
The School of Archaeology &Ancient History
Leicester, UK Carpological collection N1880 samples; N1500 species orsubspecific taxa
Comparative seed collection from thenorthern Europaean wild and domesticatedflora, and some exotic species.
http://www2.le.ac.uk/departments/archaeology/facilities
Western Australian Herbarium,Department of Environmentand Conservation
Perth, WesternAustralia, Australia
Carpological collection Quantity of samples and taxa notspecified
Presumably developed as an adjunct to theherbarium.
http://www.collections.ala.org.au/public/show/co75; http://www.dec.wa.gov.au/our-environment/science-and-research/wa-herbarium.html
Orto Botanico di Modena,Università di Modena e ReggioEmilia
Modena, Italy Carpological collection(“Carpoteca Storica”)
N1000 samples Presumably developed as an adjunct to theherbarium.
http://www.ortobotanicoitalia.it/emilia-romagna/modena/
(continued on next page)
69E.M
artinettoetal./Review
ofPalaeobotanyand
Palynology201
(2014)56
–74
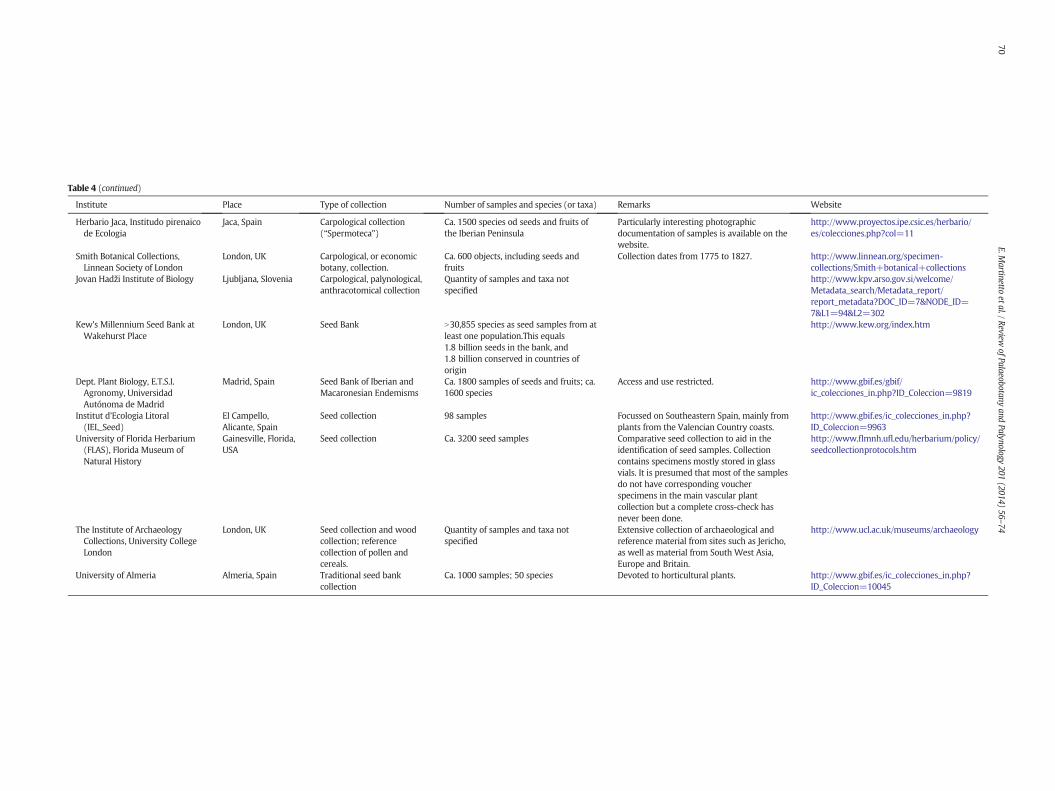
Table 4 (continued)
Institute Place Type of collection Number of samples and species (or taxa) Remarks Website
Herbario Jaca, Institudo pirenaicode Ecologia
Jaca, Spain Carpological collection(“Spermoteca”)
Ca. 1500 species od seeds and fruits ofthe Iberian Peninsula
Particularly interesting photographicdocumentation of samples is available on thewebsite.
http://www.proyectos.ipe.csic.es/herbario/es/colecciones.php?col=11
Smith Botanical Collections,Linnean Society of London
London, UK Carpological, or economicbotany, collection.
Ca. 600 objects, including seeds andfruits
Collection dates from 1775 to 1827. http://www.linnean.org/specimen-collections/Smith+botanical+collections
Jovan Hadži Institute of Biology Ljubljana, Slovenia Carpological, palynological,anthracotomical collection
Quantity of samples and taxa notspecified
http://www.kpv.arso.gov.si/welcome/Metadata_search/Metadata_report/report_metadata?DOC_ID=7&NODE_ID=7&L1=94&L2=302
Kew's Millennium Seed Bank atWakehurst Place
London, UK Seed Bank N30,855 species as seed samples from atleast one population.This equals1.8 billion seeds in the bank, and1.8 billion conserved in countries oforigin
http://www.kew.org/index.htm
Dept. Plant Biology, E.T.S.I.Agronomy, UniversidadAutónoma de Madrid
Madrid, Spain Seed Bank of Iberian andMacaronesian Endemisms
Ca. 1800 samples of seeds and fruits; ca.1600 species
Access and use restricted. http://www.gbif.es/gbif/ic_colecciones_in.php?ID_Coleccion=9819
Institut d'Ecologia Litoral(IEL_Seed)
El Campello,Alicante, Spain
Seed collection 98 samples Focussed on Southeastern Spain, mainly fromplants from the Valencian Country coasts.
http://www.gbif.es/ic_colecciones_in.php?ID_Coleccion=9963
University of Florida Herbarium(FLAS), Florida Museum ofNatural History
Gainesville, Florida,USA
Seed collection Ca. 3200 seed samples Comparative seed collection to aid in theidentification of seed samples. Collectioncontains specimens mostly stored in glassvials. It is presumed that most of the samplesdo not have corresponding voucherspecimens in the main vascular plantcollection but a complete cross-check hasnever been done.
http://www.flmnh.ufl.edu/herbarium/policy/seedcollectionprotocols.htm
The Institute of ArchaeologyCollections, University CollegeLondon
London, UK Seed collection and woodcollection; referencecollection of pollen andcereals.
Quantity of samples and taxa notspecified
Extensive collection of archaeological andreference material from sites such as Jericho,as well as material from South West Asia,Europe and Britain.
http://www.ucl.ac.uk/museums/archaeology
University of Almeria Almeria, Spain Traditional seed bankcollection
Ca. 1000 samples; 50 species Devoted to horticultural plants. http://www.gbif.es/ic_colecciones_in.php?ID_Coleccion=10045
70E.M
artinettoetal./Review
ofPalaeobotanyand
Palynology201
(2014)56
–74

71E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
pointing to this species. All the samples of other species with similarachenes have been checked without evidence of this morphology.
Diagnosis: Carex cf. riparia. The open nomenclature is justified by theinterpretation of the achene as a sterile one;wewould prefer to confirmthe report of this species on the basis of normally developed achenes.
Fossil sample 11 (Plate IV, 11)These achenes (CCN6076) are characterised by obovate shape,
stipitate base, cell pattern larger than in the majority of Carex achenes.The style base is also very characteristic: robust and conical. Based onthese characters, the identification of these fossils as Bolboschoenusmaritimus (L.) Palla would have been possible also without comparisonto the MCC material. The appearance of the epidermis of modern fruits(Plate III, 11) is completely different from the fossils: shining, smooth,light brown. However, it has been demonstrated for several water-logged fossil fruits that a shining light surface turned into dull anddarker; unfortunately, no ε-MCC or ζ-MCC is available to confirm ifsuch transformation already occurs at the stage of fruit burial into thesoil.
Diagnosis: Bolboschoenus maritimus sensu lato.
Fossil sample 12 (Plate IV, 12)This sample (CCN6375) is represented by abundant achenes (Table 1)
assignable to the genus Schoenoplectus subgenus Schoenoplectus on thebasis of the following character combination: obovate outline, acutebase and largely obtuse apex, flat on one face and convex on the other;wall very thick; epidermal cells small-sized and sub-rectangular. Thepresence of a longitudinal, blunt carina on the convex side of themajorityof the fossil fruits is highly diagnostic for Schoenoplectus lacustris (Plate III,12), but a few ones lack the carina andwould agreewith the descriptions(Berggren, 1969; Pignotti, 2003) of Schoenoplectus tabernaemontani(C.C. Gmel.) Palla and Schoenoplectus triqueter (L.) Palla. Here, the roleof theMCCmaterial has been determinant in understanding fruit varia-tion in our fossil sample: in each α-MCC of S. lacustris one could singleout a few fruits with a S. tabernaemontani-like and S. triqueter-likeshape. We suggest that the detection of a characteristic S. lacustris-morphology in ca. 80% of our fossil sample provides a reliable assign-ment at species level.
Diagnosis: All the fossil achenes would represent Schoenoplectuslacustris, and, given the broad intraspecific variation observed in MCCsamples, there is no clear evidence for the occurrence of other speciesin this rich assemblage.
Fossil sample 13 (Plate IV, 13)The sample (CCN6455) is made up by nine achenes (Table 1), and
one of them has preserved a part of stylopodium, which points to thegenus Eleocharis. The lack of perianth bristles can be attributed todamage due to taphonomical processes. The dimensions of thesebiconvex fruits, the outline, the epidermal cell pattern, and the widthof the stylopodium attachment admit only three European speciesfor comparison: Eleocharis austriaca Hayek, Eleocharis mamillata(H. Lindb.) H. Lindb and Eleocharis palustris (L.) Roem. & Schult.(Plate III, 13). Eleocharis ovata (Roth) Roem. & Schult. is also similar,but smaller, and with broader stylopodium base; Eleocharis uniglumis(Link) Schult. has larger achenes and coarser cell pattern. The modernachenes of E. mamillata, E. austriaca and E. palustris are easily sepa-rated based on stylopodium shape (Berggren, 1969). Yet, this struc-ture, in the single fossil achene where it was preserved (Plate IV,13), is distinctly incomplete. We obtained a similar condition bymacerating achenes of E. austriaca (MCC1976) and E. palustris(MCC0506, MCC1977) in sodium hypochlorite (10 h), then removingthe softened stylopodium by needle preparation (Plate III, 13). There-fore, our interpretation is that the stylopodium of the fossil wasdestroyed for most of its length.
The modern sample with the best fit to the fossils is MCC1977(Eleocharis palustris). The sample of Eleocharis austriaca differs for the
broader achenes and narrower stylopodium attachment. In addition,the fruits of Eleocharis mamillata illustrated by Berggren (1969) haveachenes definitely broader than the fossils. Unfortunately this speciesis still lacking in the MCC, and it is possible that a direct comparison ofthe fossil to the modern material will provide clues for a definitedetermination.
Diagnosis: Eleocharis palustris vel E. mamillata.
4. Discussion
4.1. Role of the MCC in the identification experiment of fossil materials
Amongst the thirteen fossil samples selected for the identificationexperiment, four samples (Carex leporina, Carex remota, Carex umbrosa,and Bolboschoenus maritimus s.l.) match the morphology reported forextant species in the literature (see above) and would have been cor-rectly identified without the MCC material. Other four samples (Carexcaryophyllea, Carex pseudocyperus, Carex cf. riparia, and Schoenoplectuslacustris) would have left, in our opinion, considerable perplexities asfor a precise specific assignment, without consulting theMCC referencesamples (but obviously also analogous material available in other col-lections). On the other hand, the correct interpretation of the remainingfive fossil samples (Carex cf. nigra, Carex pallescens, Carex pendula, Carexvesicaria, and Eleocharis palustris vel mamillata) by comparison to theliterature would have been either problematic or hardly possible, andthe comparative study of the MCC material has been decisive in thesecases. It is true that for two of these five species we were unable toprovide a definite determination, but it should be considered that, in thefirst case, theMCCmaterial avoided amisinterpretation (as C. mucronata,by comparison to Ercole et al., 2012); in the second case (uncertaintyE. palustris/mamillata), the lack of MCC material of E. mamillata possiblyhad a detrimental effect.
4.2. Applications and relevance of the MCC
The MCC has been used for palaeobotanical investigations (Cavalloand Martinetto, 2001; Martinetto, 2009; Martinetto et al., 2012;Vassio, 2012), including taphonomical studies (Martinetto and Vassio,2010; Bertolotto et al., 2012; Vassio and Martinetto, 2012) and for thepreparation of an atlas of Cyperaceae fruits (Ercole et al., 2012). In gen-eral, for modern and late Quaternary achene samples, for which thepresent flora of the territory fixes definite constraints in searching theliving relatives, our observations confirmed the views of previous au-thors (Nilsson and Hjelmquist, 1967; Berggren, 1969; Latałowa, 1999;Mai, 2010) who concluded that the morphological characterisation ofCyperaceae achenes usually permits assignment to a precise species.This is promising for the use of Cyperaceae fruit collections in re-search fields beyond archaeo-/palaeobotany, especially in ecological(e.g.: Figuerola et al., 2010) and forensic sciences (see below).
At the present state the MCC is clearly a collection of regional value,even if rich enough for comprehensive studies pointing at the character-isation of fruits at sectional and species level within Cyperaceae. Themain strength that we see in theMCC, regardless of its sample richness,is represented by its structure, expansibility and fitness for the identifi-cation of Cyperaceae fruit samples, in particular the fossil ones. The newMCC protocol could solve the usual and variable difficulties in findingadequate reference material in herbaria and conventional carpologicalcollections (Nesbitt, 1990): scarce or unripe material, strict limits tosampling of vouchers, origin of the sample not clear, determinationnot reliable and up-to date, fruits not prepared for analysis, storagemedia hampering the simultaneous observation of several samples,etc. Of course, our protocol can be improved and implemented withother useful techniques (e.g.: Nesbitt et al., 2003; Birks, 2013).
A very elementary solution applied to the MCC, i.e. the mounting ofconsultable samples in transparent polyethylene bags, constitutes anenormous advantage for the comparison of fossil samples to themodern

72 E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
ones and can promote the taxonomic interpretation of a large amount ofvaguely assigned fossil samples, which are waiting to be included in thesystematic framework for phylogenetic purposes.
At this moment, most published studies on the diversificationof Cyperaceae are based on indirect estimates (mutation rates: Hippet al., 2010; Escudero et al., 2010; secondary calibrations: Escuderoet al., 2009; geological events: Dragon and Barrington, 2009), beingjust the study of Escudero et al. (2012) calibrated using the age of a fos-sil. Perhaps, the MCC is not rich enough yet to perform the taxonomicassessment at species level in large and problematic sections, butprobably suitable for assignation at supraspecific level in mostCyperaceae groups (e.g.: Carex sect. Phacocystis, Jiménez Mejías andMartinetto, 2013). In any case, the consultation of the wide networkof institutional herbaria and carpological collections (Table 4) willalways constitute an essential support for the thorough success ofsuch studies. The lack of a large worldwide reference collection (ornetwork of collections) of comparative material is indeed one ofthe reasons why the identification of the extensive cyperaceous fos-sil fruit record is often disregarded from rigorous taxonomic treat-ment, and rarely included in phylogenetic studies (Smith et al.,2009; Jimenez-Mejias and Martinetto, 2013). The progress of fruit-morphological studies and their integration with molecular datawill presumably yield a better definition of the phylogeographichistory of those taxa with a good fossil record, as well as a better un-derstanding of the phylogenies, with subsequent feedback to thepalaeobotanical studies (Smith et al., 2010).
The broad array of modern reference samples already available inthe MCC allows assessing the extent of the intraspecific variability inmodern samples, evaluating variation in fine-scale characters, and,more important, to pinpoint the constant characters which are moresignificant for taxonomic/phylogenetic studies. However, also thedefinition of fruit or utricle variability in each modern species is an im-portant taxonomic tool, and one of themain guides that palaeobotanistsmay use for species delimitation in rich fossil fruit samples(Velichkievich and Zastawniak, 2006). Following this line, several“extinct” species (morphospecies) have been characterised on thebasis of finemorphological differenceswith respect to the nearest livingrelatives (see Cyperus glomeratoides, Schoenoplectus lacustroides inVelichkievich and Zastawniak, 2003): in these cases characteristic mor-phological details have been interpreted as time-dependant andlinked to the taxon's evolution. Such results can be very useful in bio-stratigraphic/biochronologic studies, which can have an importantgeological application, as the approximate dating of fossil-bearingdeposits.
In forensic investigations the availability of a carpological collectionis equally important as a pollen collection. Pollen grains are quite wellstudied and such botanical evidence has long been considered a usefultool when used for determining where a person or an object mayhave been (Montali et al., 2006). Seeds and fruits or their parts areoften collected at crime scenes, especially when cadavers are locatedon the ground or buried (Lombardi, 1999). A correct identification ofseed and fruit evidence could directly associate environmentswith peo-ple, whether they are suspects or victims (Miller Coyle, 2004). In partic-ular, both in the literature (e.g.: Janyszek et al., 2008) and in our ownobservations (see Plate I), it has been pointed out that local populationsof Cyperaceae fruits may show peculiar morphological traits. Therefore,the provenance of fruit material found at crime scenes would requirethe comparative analysis of several conspecific samples from variouslocalities, which is partly feasible by means of the MCC right now, andwill possibly be even easier in the perspective of a future development.The other carpological collections with reference material presentlyavailable in Italy (Table 4) do not seem to fulfil all the needs for forensicinvestigations (e.g.: the rapid and simultaneous analysis of preparedsamples for many species, and several different localities). When con-sidering seeds and fruits in a forensic context the collaboration withspecialised researchers, actively working on carpological collections
such as the MCC, is of paramount importance. This type of botanicalcollection will not only be useful for current crime scene investigations,but also in numerous cold cases involving botanical evidence (Hall andByrd, 2012; Magni et al., 2012).
4.3. Future
The availability of MCC material already provided a valuable helpin the difficult study of fossil fruits of Cyperaceae (Bertolotto et al.,2012; Martinetto et al., 2012; Vassio, 2012; Jiménez-Mejías andMartinetto, 2013). The abundance of specimens in each modernsample and the preparation of multiple samples of the same speciesprovided uswith new tools in pinpointing the close living relatives offossil samples. Future developments of the collection will beaddressed towards finding materials of the European taxa whichare now lacking in the MCC, possibly focussing on type α and β sam-ples, in order to build up a large collection of European cyperaceousfruits, hopefully in synergy with other European institutions dealingwith carpological collections (Table 4). Likewise, new accessions ofthose species which are currently represented only by typeγ or δ sam-ples (Appendix 1) will be encouraged. We also recognised a definitepractical utility in the direct comparison of fossils and ε-MCC/ζ-MCCsamples (see above for Carex caryophyllea and Carex pendula), whichare regrettably scarce in theMCC and should be increased formore spe-cies in the future.
The experience gained in NW Italy during the past few years hasshown that spontaneous contributions by researchers which are ac-tive in the field (see Acknowledgements) can permit, in a short timespan, assembling a large regional collection of fruits, focussed on theCyperaceae in our case. With this paper we also want to stimulate re-search centres operating in other continents to apply the MCC (orsimilar) protocol, thus moving the first steps towards the construc-tion of a network of collections which would facilitate the study ofCyperaceae fruits all over the world. The exchange and physical as-semblage of materials worldwide are not a compulsory action inthis process (and often not allowed by the laws against alien speciesinvasions/disease diffusion), because the gathering of high-qualityimages (e.g.: Hoshino and Masaki, 2011; Myrbo et al., 2011; Ercoleet al., 2012) of MCC-like samples (i.e.: abundant material, wellassessed provenance) would represent an acceptable compromiseagainst the physical assemblage of a worldwide collection. Adequatewebsites to host this material already exist, as eMonocot Cyperaceae(eMonocot Team, 2013), and we believe that the simple use of thepresent facilities would be sufficient to build up, in a short time, aworldwide list of taxa whose carpological materials and/or imageswould be available to the advantage of archaeo- and palaeobotanicalstudies. The possible result of this network could be a global “virtual”collection, formed bymerging the contributions from several large orsmall institutions.
Supplementary data to this article can be found online at http://dx.doi.org/10.1016/j.revpalbo.2013.11.003.
Acknowledgements
We are deeply indebted to Laura Guglielmone (herbarium TO) forthe curation of voucher specimens and to Rosa Camoletto andAnnalaura Pistarino (herbarium MRSN) for the long-lasting collabo-ration in locating fruit samples. We also thank the VertebratePalaeontology group of the Turin University, for the access to micro-photographic equipment, and Takuji Honshino and Tomomi Masakifor sending useful information on the Japanese literature andcollections. Furthermore, we wish to thank Carlo Argenti, FabrizioBertolino, Maurizio Bovio, Oreste Cavallo, Aldo Chiariglione, MicheleDa Pozzo, Giannantonio Domina, Enrico Ercole, Silvio Frattini, CaroleGee, Daniela Gigante, Claudio Gioda, Cesare Lasen, Modesto Luceño,Enrique Maguilla, Santiago Martín-Bravo, Chiara Minuzzo, Arata

73E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
Momohara, Franco Pedrotti, Renata Perego, Maria Pingen, LauraPoggio, Filippo Prosser, Cesare Ravazzi, Radomír Řepka, AdrianoSoldano and Nestor Viñals for their contribution to theMCC. A partic-ular acknowledgement is addressed to Loredana Guglielmetto forthe curation and implementation of the collection in the 1990s andto Adriano Soldano for the help provided in the identification or revi-sion of problematic voucher specimens. We are grateful to NECLIME(http://www.neclime.de/) for supporting us in collecting informa-tion about reference carpological collections.
References
Aeschimann, D., Burdet, H.M., 2005. Flore de la Suisse et des Territoires Limitrophes: leNouveau Binz. Haupt, Bern.
Aeschimann, D., Lauber, K., Moser, D.M., Theurillat, J.-P., 2004. Flora Alpina. Zanichelli,Bologna.
Bacchetta, G., Demurtas, A., Pontecorvo, C., 2004. The Biodiversity Conservation Centre ofCagliari (Italy). Scr. Bot. Belg. 29, 79–82.
Berggren, G., 1969. Atlas of Seeds and Small Fruits of Northwest European Plant Specieswith Morphological Descriptions. Part 2 Cyperaceae.Swedish Natural Science Re-search Council, Lund.
Bertolotto, G., Martinetto, E., Vassio, E., 2012. Applicazioni paleobotaniche dello studio diresti carpologici in suoli e depositi fluviali attuali del Piemonte, con particolareriferimento alle Cyperaceae. Inf. Bot. Ital. 44 (2), 72–74.
Birks, H.H., 2013. Plant macrofossil introduction. In: Elias, S., Mock, C. (Eds.), Encyclopediaof Quaternary Science. Elsevier, pp. 593–612.
Bojňanský, V., Fargašová, A., 2007. Atlas of seeds and fruits of Central and East-Europeanflora. The Carpathian Mountains Region.Springer, Dordrecht.
Browning, J., Gordon-Gray, K.D., 2000. Patterns of fruit morphology in Bolboschoenus(Cyperaceae) and their global distribution. S. Afr. J. Bot. 66, 63–71.
Cappers, R.T.J., Bekker, R.M., Jans, J.E.A., 2006. Digital Seed Atlas of the Netherlands.Barkhuis.
Cavallo, P., Martinetto, E., 2001. Flore carpologiche del Pliocene di Castelletto Cervo(Biella). Boll. Museo Regionale Sci. Nat. Torino 18 (2), 277–343.
Conti, F., Abbate, G., Alessandrini, A., Blasi, C. (Eds.), 2005. An annotated checklist of theItalian vascular flora. Palombi, Rome.
Conti, F., Alessandrini, A., Bacchetta, G., Banfi, E., Barberis, G., Bartolucci, F., Bernardo, L.,Bonacquisti, S., Bouvet, D., Bovio, M., Brusa, G., Del Guacchio, E., Foggi, B., Frattini, S.,Galasso, G., Gallo, L., Gangale, C., Gottschlich, G., Grünanger, P., Gubellini, L., Iiriti, G.,Lucarini, D., Marchetti, D., Moraldo, B., Peruzzi, L., Poldini, L., Prosser, F., Raffaelli, M.,Santangelo, A., Scassellati, E., Scortegagna, S., Selvi, F., Soldano, A., Tinti, D., Ubaldi,D., Uzunov, D., Vidali, M., 2007. Integrazioni alla checklist della flora vascolareitaliana. Nat. Vicentina 10, 5–74.
Dickson, C.A., 1970. The study of plant macrofossils in British Quaternary deposits.In: Walker, D., West, R.G. (Eds.), Studies in the Vegetational History of theBritish Isles. Essays in Honour of Harry Godwin. Cambridge University Press,pp. 233–254.
Dragon, J.A., Barrington, D.S., 2009. Systematics of the Carex aquatilis and C. lenticularis lin-eages: geographically and ecologically divergent sister clades of Carex sectionPhacocystis (Cyperaceae). Am. J. Bot. 96, 1896–1906.
Egorova, T.V., 1999. The Sedges (Carex L.) of Russia and Adjacent States. Missouri Botan-ical Garden Press, Saint-Louis.
eMonocot Team, 2013. eMonocot Cyperaceae. (http://www.cyperaceae.e-monocot.org[accessed 8 November 2013]).
Ercole, E., Pistarino, A.L., Martinetto, E., Soldano, A., Siniscalco, C., 2012. Atlante fotograficodei frutti e dei semi della flora del Piemonte e della Valle d'Aosta: Cyperaceae. Boll.Museo Regionale Sci. Nat. Torino 29 (1–2), 5–34 and 133–284.
Escudero, M., Valcárcel, V., Vargas, P., Luceño, M., 2009. Significance of ecological vicari-ance and long-distance dispersal in the diversification of Carex sect. Spirostachyae(Cyperaceae). Am. J. Bot. 96, 2100–2114.
Escudero, M., Vargas, P., Arens, P., Ouborg, N.J., Luceño, M., 2010. The east–west–northcolonization history of the Mediterranean and Europe by the coastal plant Carexextensa (Cyperaceae). Mol. Ecol. 19, 352–370.
Escudero, M., Hipp, A., Waterway, M., Valente, L., 2012. Diversification rates and chromo-some evolution in the most diverse angiosperm genus of the temperate zone (Carex,Cyperaceae). Mol. Phylogenet. Evol. 63, 650–655.
Ferreira, P.M.A., Eggers, L., 2008. Espécies de Cyperaceae do Centro de Pesquisa eConservação da Natureza Pró-Mata, município de São Francisco de Paula, RS, Brasil.Acta Bot. Bras. 22, 173–185.
Figuerola, J., Charalambidou, I., Santamaria, L., Green, A.J., 2010. Internal dispersal of seedsby waterfowl: effects of seed size on gut passage time and germination patterns.Naturwissenschaften 97, 555–565.
Fournier, P., 1961. Les Quatres Flores De France. P. Lechevalier, Paris.Godwin, H., 1956. Collection and identification of plant remains. In: Godwin, H. (Ed.), The
History of the British Flora: a Factual Basis for Phytogeography. University Press,Cambridge, pp. 6–53.
Hall, D., Byrd, J., 2012. Forensic Botany: A Practical Guide. Wiley-BlackwellPublishing.
Hefler, S.M., Longhi-Wagner, H.M., 2008. Análise da morfologia do fruto em espécies deCyperus L. subg. Cyperus–Cyperaceae. Acta Bot. Brasiliensia 22 (3), 637–651.
Hess, H.E., Landolt, E., Hirzel, R., 1967–72. Flora der Schweiz., vols. 1-3. Basel & Stuttgart.
Hipp, A.L., Rothrock, P.E., Whitkuss, R., Weber, J.A., 2010. Chromosomes tell half of the his-tory: the correlation between karyotype rearrangements and genetic diversity insedges, a group with holocentric chromosomes. Mol. Ecol. 19, 3124–3138.
Hoshino, T., Masaki, T., 2011. Illustrated Sedges of Japan. Heibonsha Ltd. Publishers,Tokyo.
Hroudová, Z., Kákravský, P., Ducháček, M., Marhold, K., 2007. Taxonomy, distribution andecology of Bolboschoenus in Europe. Ann. Bot. Fenn. 44, 81–102.
I.B.P.G.R. (International Board for Plant Genetic Resources), 1982. Design of Seed StorageFacilities for Genetic Conservation. Revised 1985 and 1990. International Board forPlant Genetic Resources, Rome.
Janyszek, M., Jagodziński, A.M., Janyszek, S., Wrońska-Pilarek, D., 2008. Morphological vari-ability of Carex spicata Huds. utricles among plant communities. Flora 203, 386–395.
Jermy, A.C., Simpson, D.A., Foley, M.J.Y., Porter, M.S., 2007. Sedges of the British Isles, BSBIHandbook no. 13rd ed. Botanical Society of the British Isles, London.
Jiménez-Mejías, P., Luceño, M., 2011. Cyperaceae. Euro + Med Plantbase—the informationresource for Euro-Mediterranean plant diversity (http://ww2.bgbm.org/EuroPlusMed/[accessed 31 July 2012]).
Jiménez-Mejías, P., Martinetto, E., 2013. Towards an accurate taxonomic interpretation ofCarex fossil fruits (Cyperaceae): a case study in section Phacocystis in the WesternPalearctic. Am. J. Bot. 100, 1580–1603.
Jiménez-Mejías, P., Luceño,M., Martín-Bravo, S., 2012a. The importance of the Italian pen-insula in the phylogeography of Carex (Cyperaceae). Inf. Bot. Ital. 44 (2), 53–57.
Jiménez-Mejías, P., Luceño, M., Lye, K.A., Brochmann, C., Gussarova, G., 2012b. Geneticallydiverse but with surprisingly little geographical structure: the complex history of thewidespread herb Carex nigra (Cyperaceae). J. Biogeogr. 39 (12), 2279–2291.
Joppa, L.N., Roberts, D.L., Pimm, S.L., 2011. How many species of flowering plants arethere? Proc. R. Soc. B 278, 554–559.
Kats, N.Y., Kats, S.V., Kipiani, M.G., 1965. Atlas and Key to Identification of FossilFruits and Seeds Occurring in the Quaternary Deposits of the USSR. Nauka, Moscow(in Russian).
Latałowa, M., 1999. Palaeoecological reconstruction of the environmental conditions andeconomy in early medieval Wolin—against a background of the Holocene history ofthe landscape. Acta Palaeobotanica 39 (2), 183–271.
Linington, S.H., 1997. The millennium seed bank project. Botanic Gardens ConservationNews 2 (9), 34–35.
Lombardi, G., 1999. The contribution of forensic geology and other trace evidence analy-ses to the investigation of the killing of Italian Prime Minister Aldo Moro. J. ForensicSci. 44 (3), 634–642.
Luceño, M., Castroviejo, S., Jiménez-Mejías, P., 2008. Cyperaceae. In: Castroviejo, S.,Luceño, M., Galán, A., Jiménez-Mejías, P., Cabezas, F., Medina, L. (Eds.), Flora IbericaXVIII. CSIC, Madrid, pp. 3–250.
Lye, K.A., 2000. Achene structure and function of structure in Cyperaceae. In: Wilson, K.L.,Morrison, D.A. (Eds.), Monocots: Systematics and Evolution. CSIRO, Melbourne,pp. 615–628.
Magni, P.A., Harvey, M.L., Saravo, L., Dadour, I.R., 2012. Entomological evidence: lessons tobe learnt from a cold case review. Forensic Sci. Int. 223, e31–e34.
Mai, D.H., 2010. Karpologische Untersuchungen in einem Interglazial von Neumark-Nord(Geiseltal). Palaeontogr. B 282, 99–187.
Mai, D.H.,Walther, H., 1988. Die pliozänen Floren von ThüringenDeutsche DemokratischeRepublik. Quartärpaläontologie 7, 55–297.
Martinetto, E., 2009. Palaeoenvironmental significance of plant macrofossils fromthe Piànico Formation, Middle Pleistocene of Lombardy, North Italy. Quat. Int.204, 20–30.
Martinetto, E., Vassio, E., 2010. Reconstructing “Plant Community Scenarios” by means ofpalaeocarpological data from the CENOFITA database, with an example from the Ca'Viettone site (Pliocene, Northern Italy). Quat. Int. 225, 25–36.
Martinetto, E., Monegato, G., Vassio, E., 2012. An Early Pleistocene plant assemblage withEast European affinity in the Venetian-Friulian Basin (NE Italy). Alp. Mediterr. Quat.25 (2), 91–104.
Miller Coyle, H., 2004. Forensic Botany: Principles and Applications to Criminal Casework.CRC Press.
Montali, E., Mercuri, A.M., Trevisan Grandi, G., Accorsi, C.A., 2006. Towards a “crime pollencalendar”—pollen analysis on corpses throughout one year. Forensic Sci. Int. 163 (3),211–223.
Myrbo, A., Morrison, A., McEwan, R., 2011. Tool for Microscopic Identification (TMI).(http://www.tmi.laccore.umn.edu. [accessed on 19th October 2013]).
Nesbitt, M., 1990. Seeds reference collections and archaeobotany. Circaea 8 (1), 21–39.Nesbitt, M., Colledge, S., Murray, M.A., 2003. Organisation and management of seed refer-
ence collections. Environ. Archaeol. 8, 77–84.Nilsson, O., Hjelmquist, H., 1967. Studies on the nutlet structure of South Scandinavian
species of Carex. Bot. Notiser 120, 461–485.Pignatti, S., 1982. Flora d'Italia 1–3. Edagricole, Bologna.Pignotti, L., 2003. Scirpus L. and related genera (Cyperaceae) in Italy. Webbia 58 (2),
281–400.Schönswetter, P., Elven, R., Brochmann, C., 2008. Trans-Atlantic dispersal and large-scale
lack of genetic structure in the circumpolar, arctic-alpine sedge Carex bigelowii s.l.(Cyperaceae). Am. J. Bot. 95, 1006–1014.
Smith, R.D., Dickie, J.B., Linington, S.H., Pritchard, H.W., Probert, R.J., 2003. Seed Conserva-tion: Turning Science Into Practice. Royal Botanic Gardens, Kew.
Smith, S.Y., Collinson, M.E., Simpson, D.A., Rudall, P.J., Marone, F., Stampanoni, M., 2009.Elucidating the affinities and habits of ancient, widespread Cyperaceae: Volkeriamesselensis gen. et sp. nov., a fossil mapanoid sedge from the Eocene of Europe.Am. J. Bot. 96, 1506–1518.
Smith, S.Y., Collinson, M.E., Rudall, P.J., Simpson, D.A., 2010. The Cretaceous and Paleogenefossil record of Poales: review and current research. In: Seberg, O., Petersen, G.,Barfod, A., Davis, J.I. (Eds.), Diversity. Phylogeny, and Evolution in Monocotyledons:

74 E. Martinetto et al. / Review of Palaeobotany and Palynology 201 (2014) 56–74
Proceedings of the Fourth International Conference on the Comparative Biology ofthe Monocotyledons & the Fifth International Symposium on Grass Systematics andEvolution. Aarhus University Press, Aarhus, pp. 333–356.
Thiers, B., 2013. Index Herbariorum: a Global Directory of Public Herbaria and AssociatedStaff. NewYork Botanical Garden's Virtual Herbarium (http://www.sweetgum.nybg.org/ih/ [accessed 21 July 2013]).
Thomlison, P., 1984. Tested, rapid techniques for clearing botanical specimensfor the study of waterlogged archaeological plant remains. Circaea 2 (2),97–101.
Van der Burgh, J., 1978. The Pliocene flora of Fortuna-Garsdorf I fruits and seeds of Angio-sperms. Rev. Palaeobot. Palynol. 26, 173–211.
Vassio, E., 2012. Palaeovegetation reconstructions and palaeoclimatic interpretations ofQuaternary carpological assemblages with an actuopalaeobotanical approach. Ph.D.Thesis University of Turin, Turin.
Vassio, E., Martinetto, E., 2012. Biases in the frequency of fruits and seeds in modern flu-vial sediments in NW Italy: the key to interpreting analogous fossil assemblages.Palaios 27 (11), 779–797.
Velichkevich, F.Y., Zastawniak, E., 2003. The Pliocene flora of Kholmech, south-easternBelarus and its correlation with other Pliocene floras of Europe. Acta Palaeobotanica43 (2), 137–259.
Velichkevich, F.Y., Zastawniak, E., 2006. Atlas of the Pleistocene Vascular Plant Macrofossilsof Central and Eastern Europe. Part I—Pteridophytes andMonocotyledons.W. Szafer In-stitute of Botany, Polish Academy of Sciences, Kraków.
Walters, S.M., 1953. Eleocharis mamillata Lindb. fil. and allied species. Ber. Schweiz. Bot.Ges. 63, 271–286.
Walters, S.M., 1980. Cyperaceae. In: Tutin, T.G., Heywood, V.H., Burges, N.A., Moore, D.M.,Valentine, D.H., Walters, S.M., Webb, D.A. (Eds.), Flora Europaea, vol. 5. UniversityPress, Cambridge.




















