
A new method of genetic engineering - CRISPR/Cas9. (text in Russian)
23
Новый метод генной инженерии - CRISPR/Cas9 Дмитрий Джагаров Мы портняжки хоть куда – перекроим ДНК, Ген мутантный удалим и здоровым заменим Аннотация Обладающая способностью к адаптации система элементов геномной последовательности, называемая CRISPR (Clustered Regularly Interspaced Short Palindromic Repeat) и различные ассоциированные с ней белки (Cas) выполняют у бактерий и архей роль своеобразной иммунной системы помогающей им распознавать и уничтожать чужеродный генетический материал, обеспечивая, таким образом их защиту от бактериофагов, плазмид и других мобильных генетических элементов. Умение этой системы избирательно направлять белки Cas на участки ДНК с определенной последовательностью нуклеотидов позволило разработать новые сравнительно недорогие и не трудоемкие технологии для генной инженерии и функциональной геномики. A new method of genetic engineering - CRISPR/Cas9 Dmitry Dzhagarov Abstract To combat potentially deadly viral infections, prokaryotic microbes enlist small RNA-based adaptive immune systems (CRISPR- Cas systems) that protect through sequence-specific recognition and targeted destruction of viral nucleic acids. This Prokaryotic defense weapon was redirected for the purposes of biotechnology. Currently, CRISPR/Cas systems represent an effective and versatile biotechnological tool, which will have significant impact on future advancements in genome engineering. Short palindromic repeat (CRISPR) technology, an important new approach for generating RNA-guided nucleases, such as Cas9, with 1
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of A new method of genetic engineering - CRISPR/Cas9. (text in Russian)

Новый метод генной инженерии - CRISPR/Cas9 Дмитрий Джагаров
Мы портняжки хоть куда – перекроим ДНК,Ген мутантный удалим и здоровымзаменим
АннотацияОбладающая способностью к адаптации система элементов геномной последовательности, называемая CRISPR (Clustered Regularly Interspaced Short Palindromic Repeat) и различные ассоциированные с ней белки (Cas) выполняют у бактерий и архей роль своеобразной иммунной системы помогающей им распознавать и уничтожать чужеродный генетический материал, обеспечивая, таким образом их защиту от бактериофагов, плазмид и других мобильных генетических элементов.Умение этой системы избирательно направлять белки Cas на участкиДНК с определенной последовательностью нуклеотидов позволило разработать новые сравнительно недорогие и не трудоемкие технологии для генной инженерии и функциональной геномики.
A new method of genetic engineering - CRISPR/Cas9Dmitry DzhagarovAbstractTo combat potentially deadly viral infections, prokaryotic microbes enlist small RNA-based adaptive immune systems (CRISPR-Cas systems) that protect through sequence-specific recognition and targeted destruction of viral nucleic acids. This Prokaryotic defense weapon was redirected for the purposes of biotechnology. Currently, CRISPR/Cas systems represent an effective and versatile biotechnological tool, which will have significant impact on future advancements in genome engineering.Short palindromic repeat (CRISPR) technology, an important new approach for generating RNA-guided nucleases, such as Cas9, with
1

customizable specificities. Genome editing mediated by these nucleases has been used to rapidly, easily and efficiently modify endogenous genes in a wide variety of biomedically important cell types and in organisms that have traditionally been challenging to manipulate genetically.
В 1987 году в геноме кишечной палочки Escherichia coli, по соседству с геном изоформы щелочной фосфатазы, был обнаружен участок генома со странной загадочной архитектурой, состоящей из многочисленных повторов [1]. Функция повторов этого участка, названного КРАЙСПР (CRISPR) локусом, долгое время оставалась загадочной. Но в 2005 году сразу три группы сообщили, что разделяющие эти повторы промежуточные последовательности в локусе CRISPR зачастую идентичны последовательностям, найденным в геномах у бактериофагов (фагов) и плазмид. Эти данные позволили предположить, что локус CRISPR не что иное как часть ранее неизвестного иммунного механизма, базирующегося на последовательностях ДНК, который предназначен для защиты бактерий и архей от инфекции, вызванной вирусами и другими генетическими паразитами. Такая гениальная система защиты не осталась незамеченной – на протяжении 2012-2013 годов на ее основе были изобретены не только новые «инструменты» и методы редактирования генов, но также способы управления активностью генов и изучения поведения хроматина в живой клетке. Предполагается, что в ближайшем будущем CRISPRs станет одним изосновных высокоточных инструментов генной инженерии, поскольку эта технология проще в использовании и гораздо дешевле нынешних методов. Даже у таких простых организмов, какими являются бактерии и археи (одноклеточные микроорганизмы, не имеющие ядра, и мембранных органелл), есть многочисленные способы обеспечения безопасности от угрожающей им инфекции.Одним из способов обороны является локус CRISPR защищающий их от экзогенных (посторонних) нуклеиновых кислот. Система CRISPR (clustered regularly interspaced short palindromic repeats, в переводе с английского: скопление регулярно разделенных промежутками коротких симметричных повторов) является хорошо адаптирующимся наследуемым механизмом распознавания и уничтожения захватчиков. Свое название этот механизм получил благодаря особенностям строения его локуса представляющего собой массив из серии коротких повторов чередующихся с неповторяющимися промежуточными последовательностями ДНК. В каждом конкретном локусе все повторы практически одинаковы по
2

длине и последовательности и имеют длину от 24 до 48 пар нуклеотидов. Промежутки также примерно одинаковы по длине, но ихсодержимое весьма вариабельно по последовательности. Их длина от21 до 72 пар нуклеотидов. Эти промежуточные последовательности, называемые спейсерами, часто происходят из плазмид и фагов, благодаря чему система может приспособиться к тому, чтобы целенаправленно бороться с этими патогенными микроорганизмами [2]. Оказалось, что в результате процесса называемого «адаптацией», бактерии, выжившие после атаки фага, расширяют массив своего CRISPR, спейсерами, идентичными «трофейным» небольшим участкам ДНК из напавшего на них фага [3]. Таким образом, спейсерные последовательности массива CRISPR представляют собой память бактерии-хозяина о вирусных инфекциях и встречах с инородным генетическим материалом.С края к локусу CRISPR примыкает эволюционно консервативная такназываемая лидерная последовательность, состоящая из нескольких сотен пар оснований (до 550 пар), а также CRISPR ассоциированные (CAS) гены, кодирующие белки семейства Cas. Лидерная последовательность играет роль промотора (стартовой площадки для начала транскрипции) массива CRISPR. Кроме того лидерная последовательность может функционировать в качестве последовательности узнающей белки участвующие во встраивании новых спейсеров. В пользу этого свидетельствует то, что встраивание участка ДНК от нападавшего микроорганизма, а также удвоение повтора обычно просходит на границе между ней и массивом CRISPR.
1 Nakata, A., Amemura, M., & Makino, K. (1989). Unusual nucleotide arrangement with repeated sequences in the Escherichia coli K-12 chromosome. Journal of bacteriology, 171(6), 3553-3556. http://jb.asm.org/content/171/6/3553.short
3
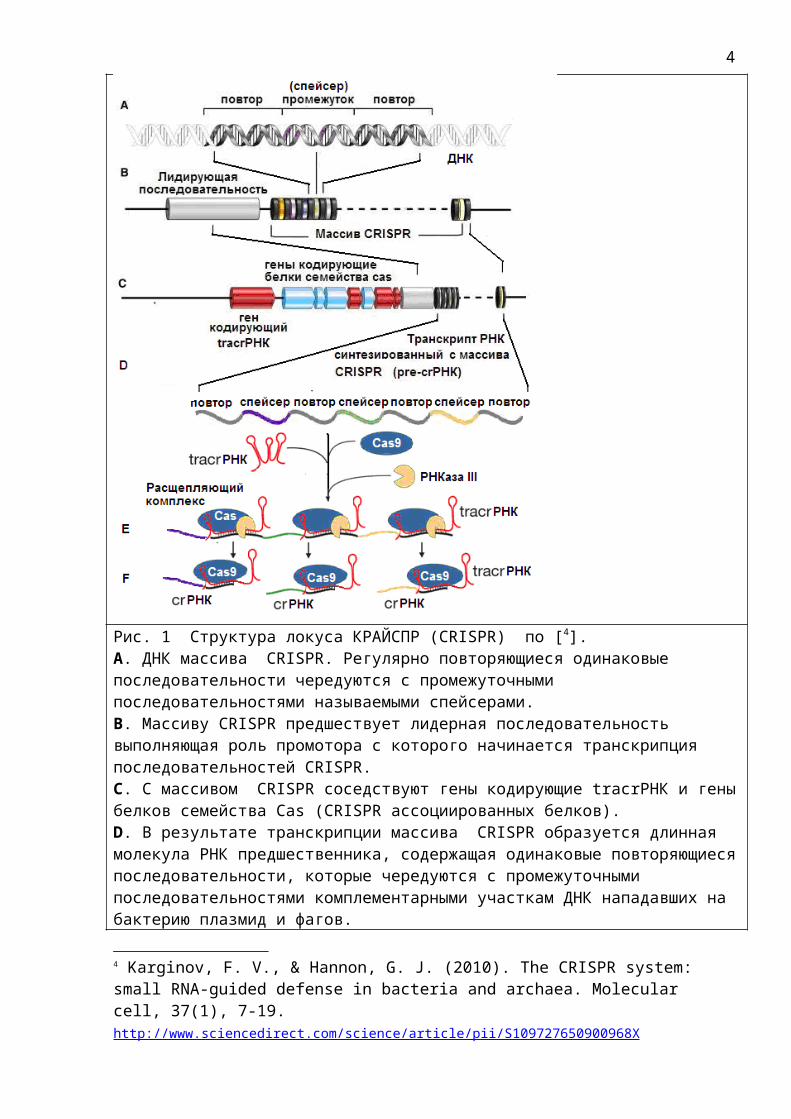
Рис. 1 Структура локуса КРАЙСПР (CRISPR) по [4]. А. ДНК массива CRISPR. Регулярно повторяющиеся одинаковые последовательности чередуются с промежуточными последовательностями называемыми спейсерами.В. Массиву CRISPR предшествует лидерная последовательность выполняющая роль промотора с которого начинается транскрипция последовательностей CRISPR.С. С массивом CRISPR соседствуют гены кодирующие tracrРНК и геныбелков семейства Cas (CRISPR ассоциированных белков). D. В результате транскрипции массива CRISPR образуется длинная молекула РНК предшественника, содержащая одинаковые повторяющиесяпоследовательности, которые чередуются с промежуточными последовательностями комплементарными участкам ДНК нападавших на бактерию плазмид и фагов.
4 Karginov, F. V., & Hannon, G. J. (2010). The CRISPR system: small RNA-guided defense in bacteria and archaea. Molecular cell, 37(1), 7-19. http://www.sciencedirect.com/science/article/pii/S109727650900968X
4
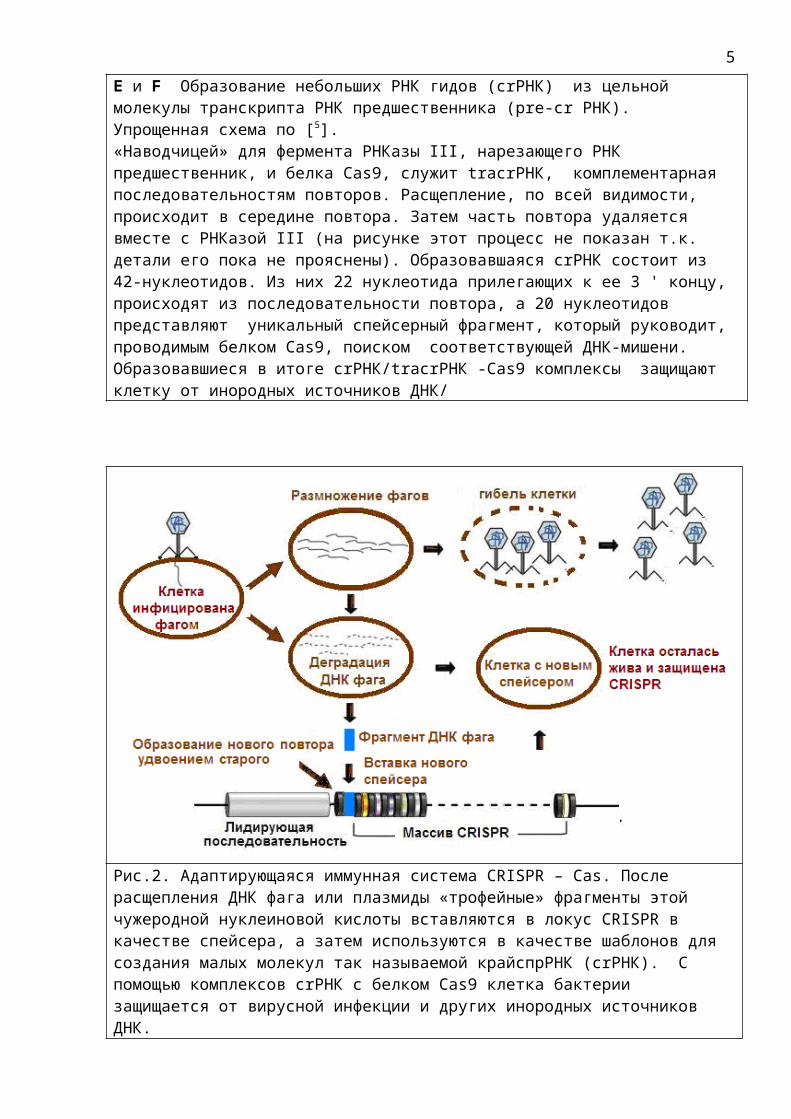
E и F Образование небольших РНК гидов (crРНК) из цельной молекулы транскрипта РНК предшественника (pre-cr РНК). Упрощенная схема по [5]. «Наводчицей» для фермента РНКазы III, нарезающего РНК предшественник, и белка Cas9, служит tracrРНК, комплементарная последовательностям повторов. Расщепление, по всей видимости, происходит в середине повтора. Затем часть повтора удаляется вместе с РНКазой III (на рисунке этот процесс не показан т.к. детали его пока не прояснены). Образовавшаяся crРНК состоит из 42-нуклеотидов. Из них 22 нуклеотида прилегающих к ее 3 ' концу, происходят из последовательности повтора, а 20 нуклеотидов представляют уникальный спейсерный фрагмент, который руководит, проводимым белком Cas9, поиском соответствующей ДНК-мишени. Образовавшиеся в итоге crРНК/tracrРНК -Cas9 комплексы защищают клетку от инородных источников ДНК/
Рис.2. Адаптирующаяся иммунная система CRISPR – Cas. После расщепления ДНК фага или плазмиды «трофейные» фрагменты этой чужеродной нуклеиновой кислоты вставляются в локус CRISPR в качестве спейсера, а затем используются в качестве шаблонов для создания малых молекул так называемой крайспрРНК (crРНК). С помощью комплексов crРНК с белком Cas9 клетка бактерии защищается от вирусной инфекции и других инородных источников ДНК.
5

После транскрипции массива CRISPR, образовавшийся цельный транскрипт РНК предшественника разрезается на отдельные фрагменты CRISPR РНК (crРНК), содержащие спейсер и часть повтора. В процессе нарезки РНК предшественника участвует небольшая РНК комплементарная повторам. Эта РНК, называемая tracrРНК служит «наводчицей» для фермента нарезающего РНК предшественник ориентируясь на повторы. Спейсерные участки crРНК затем находят комплементарные им участки цепей «вражеских» нуклеиновых кислот и таким образом направляют на них Cas белки, вызывающие их расщепление и последующую деградацию. Таким образом crРНК выполняет роль проводника – путеводителя, за что была названа «РНК гид».
Cas9 белок является нуклеазой - ферментом, специализирующимся наразрезанииДНК. Он режет двухцепочечную ДНК, используя два независимых нуклеазных домена (называемых RuvC и HNH), по одному для каждой нити ДНК. При этом домен нуклеазы HNH расщепляет цепь, которая комплементарна спейсеру, тогда как домен RuvC расщепляет противоположную цепь [6]. Интересно отметить, что белок Cas9, сам по себе, существует в неактивном состоянии, но при связывании с направляющей РНК, белок Cas9 претерпевает радикальные изменения в его трехмерной структуре, что позволяет ему взаимодействовать с ДНК-мишенью. Он состоит из двух долек: домена нуклеаз и так называемого альфа-спирального домена. На каждой из долек есть паз (канавка) для нуклеиновой кислоты. Поэтому строение Cas9 в неактивном состоянии чем то напоминает двустворчатую раковину, а после присоединения комплекса crРНК/tracrРНК, когда дольки смыкаются, и их пазы (канавки) объединяются в центральный канал (туннель) обхватывающий ДНК-мишень, строение Cas9 напоминает замок от застежки-молнии [7].
6 Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna J. A . (2012). A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science 337(6096): 816–21. DOI: 10.1126/science.1225829 http://www.sciencemag.org/content/337/6096/816.short
6
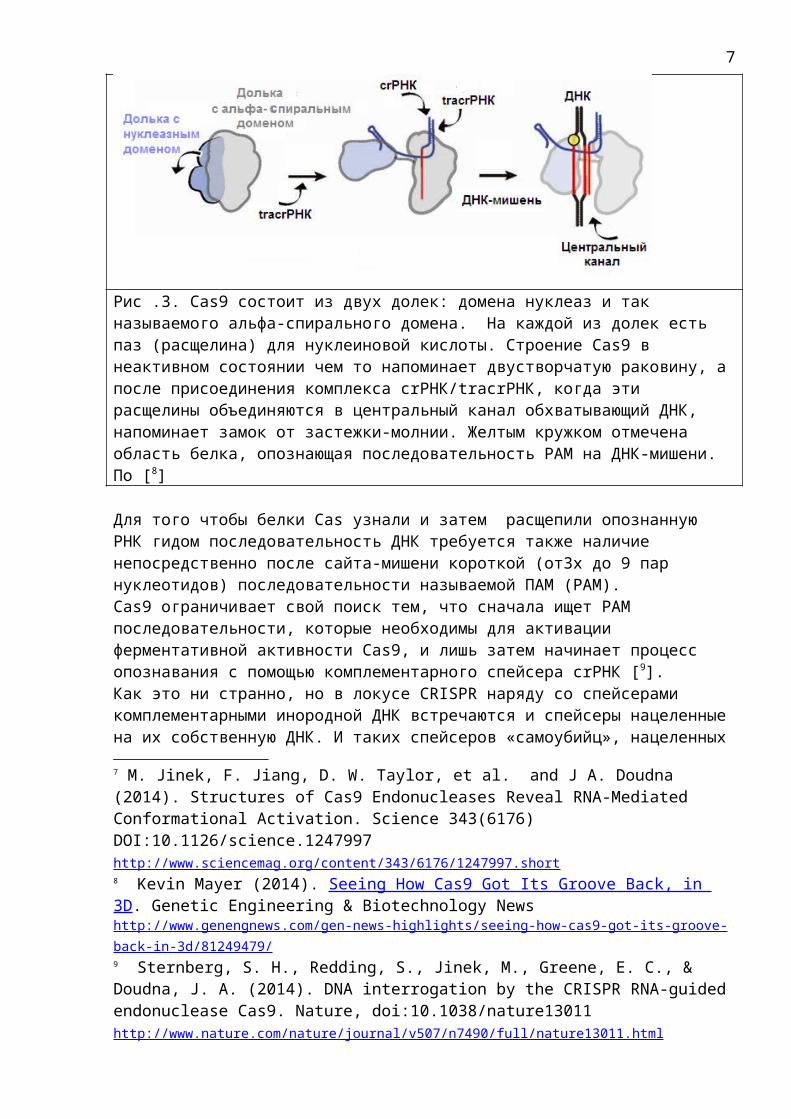
Рис .3. Cas9 состоит из двух долек: домена нуклеаз и так называемого альфа-спирального домена. На каждой из долек есть паз (расщелина) для нуклеиновой кислоты. Строение Cas9 в неактивном состоянии чем то напоминает двустворчатую раковину, апосле присоединения комплекса crРНК/tracrРНК, когда эти расщелины объединяются в центральный канал обхватывающий ДНК, напоминает замок от застежки-молнии. Желтым кружком отмечена область белка, опознающая последовательность PAM на ДНК-мишени. По [8]
Для того чтобы белки Cas узнали и затем расщепили опознанную РНК гидом последовательность ДНК требуется также наличие непосредственно после сайта-мишени короткой (от3х до 9 пар нуклеотидов) последовательности называемой ПАМ (PAM). Cas9 ограничивает свой поиск тем, что сначала ищет PAM последовательности, которые необходимы для активации ферментативной активности Cas9, и лишь затем начинает процесс опознавания с помощью комплементарного спейсера crРНК [9].Как это ни странно, но в локусе CRISPR наряду со спейсерами комплементарными инородной ДНК встречаются и спейсеры нацеленныена их собственную ДНК. И таких спейсеров «самоубийц», нацеленных7 M. Jinek, F. Jiang, D. W. Taylor, et al. and J A. Doudna (2014). Structures of Cas9 Endonucleases Reveal RNA-Mediated Conformational Activation. Science 343(6176) DOI:10.1126/science.1247997 http://www.sciencemag.org/content/343/6176/1247997.short 8 Kevin Mayer (2014). Seeing How Cas9 Got Its Groove Back, in 3D. Genetic Engineering & Biotechnology News http://www.genengnews.com/gen-news-highlights/seeing-how-cas9-got-its-groove-back-in-3d/81249479/ 9 Sternberg, S. H., Redding, S., Jinek, M., Greene, E. C., & Doudna, J. A. (2014). DNA interrogation by the CRISPR RNA-guidedendonuclease Cas9. Nature, doi:10.1038/nature13011 http://www.nature.com/nature/journal/v507/n7490/full/nature13011.html
7

на собственный организм, немало – они составляют порядка 20% . Роль их предстоит еще выяснить. Возможно, такие спейсеры могут быть необходимы бактерии, чтобы регулировать транскрипцию определенных генов, а не их расщепление [10]. Предотвратить саморазрушение таких генов, очевидно, может отсутствие рядом с ними PAM.Интересно отметить, что если заразить клетки совершенно неизвестным им фагом, то лишь три процента выживших бактерий удлиняют свою CRISPR-кассету на один спейсер, соответствующий новому фагу, то есть приспосабливаются к этой угрозе. Если же клетки заразить фагом, ДНК которого хоть и не полностью соответствует спейсеру в CRISPR кассете клетки, но все же является схожей, то адаптация в такой системе происходит на два порядка более эффективно - от 50% до 90% популяции набирает в CRISPR кассеты спейсеры из последовательности фага. То есть адаптация в таком случае идет гораздо эффективнее и выживаемостьвозрастает в десятки раз. Этот процесс, открытый российскими учеными, был назван праймингом. Связан он, по-видимому, с тем, что после опознания белком Cas9 стартовой площадки – PAM, с ДНКмишенью сначала связывается лишь небольшой участок спейсера (6-12 нуклеотидов) называемый затравкой и лишь затем остальная часть спейсера. Если на участке затравки комплементарность строго соблюдается, то на остальной части спейсера возможны отклонения, иногда достигающие 3- 5 пар нуклеотидов. Для бактерии такие отклонения только на пользу – благодаря этому она может успешно опознать фаги мутанты и, порезав их ДНК, быстро адаптироваться на новую цель, вставив новый фрагмент ДНК в виде спейсера в свой локус CRISPR. А вот ученым предстоит разработать методы позволяющие свести на нет ошибки в опознании мишени ДНК – без этого точность и надежность работы генного скальпеля будет недостаточно велика и трудно будет избежать незапланированных нежелательных мутаций при манипуляциях с геномом [11]. Возможным выходом может стать использование crРНК с усеченной спейсерной частью называемых tru-gРНК (truncated gRNAs), что по некоторым данным, позволяет на 2-3 порядка повысить точность работы молекулярной машины [12], а также использование, так называемых, спаренных никаз, о которых мы расскажем позднее.
8
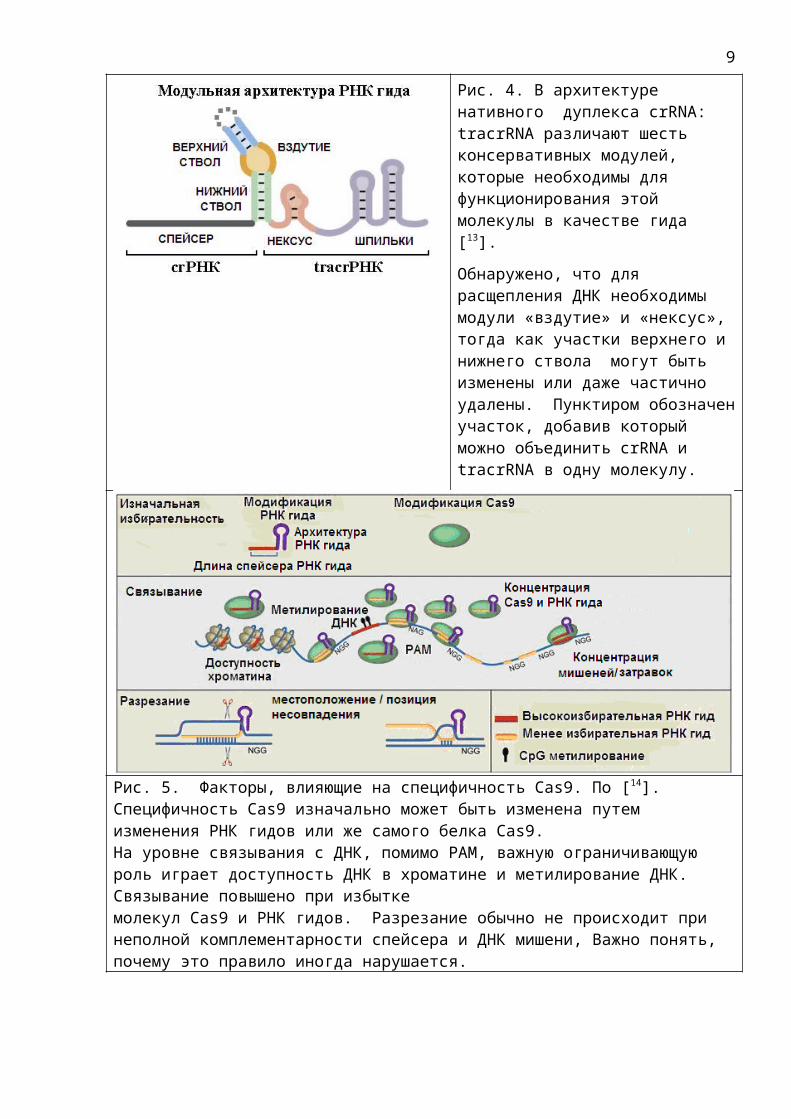
Рис. 4. В архитектуре нативного дуплекса crRNA: tracrRNA различают шесть консервативных модулей, которые необходимы для функционирования этой молекулы в качестве гида [13].Обнаружено, что для расщепления ДНК необходимы модули «вздутие» и «нексус», тогда как участки верхнего и нижнего ствола могут быть изменены или даже частично удалены. Пунктиром обозначенучасток, добавив который можно объединить crRNA и tracrRNA в одну молекулу.
Рис. 5. Факторы, влияющие на специфичность Cas9. По [14]. Специфичность Cas9 изначально может быть изменена путем изменения РНК гидов или же самого белка Cas9.На уровне связывания с ДНК, помимо РАМ, важную ограничивающую роль играет доступность ДНК в хроматине и метилирование ДНК. Связывание повышено при избыткемолекул Cas9 и РНК гидов. Разрезание обычно не происходит при неполной комплементарности спейсера и ДНК мишени, Важно понять, почему это правило иногда нарушается.
9

Интересно отметить, что некоторые фаги разработали способы успешной борьбы с этой противовирусной системой. У фагов найдены гены, способные вызвать ингибирование системы CRISPR / Cas. Существование анти-CRISPR генов предоставляет ученым новые возможности для выяснения функциональных механизмов и разработкиновых способов использования системы CRISPR / Cas [15 16].
Для чего и как используют CRISPR-Cas9 технологииИспользование адаптирующейся иммунной системы бактерий началось еще в 2007 году, когда фирме DuPont удалось создать устойчивые квирусной инфекции бактериальные штаммы для производства продуктов питания. Для этого они вакцинировали бактерию разрушенными вирусами. Однако подлинный бум начался в конце 2012года после того как удалось объединить tracrРНК и crРНК в одну цельную молекулу РНК, называемую теперь РНК гидом или sgРНК (от англ. single-guide RNA) и был изобретен вектор для клонирования этой РНК [17].Оказалось, что такая синтетическая sgРНК если ее смешать в пробирке с белком Cas9 может работать ничуть не хуже комплекса tracrРНК с crРНК и также может связавшись с белком Cas9 находить соответствующие последовательности ДНК и правильно ориентировать Cas9 чтобы создавать в них двухцепочечные разрывы.Изобретение синтетической одноцепочечной шпильки sgРНК, 15 Bondy-Denomy J, Pawluk A, Maxwell KL, Davidson AR (2013). Bacteriophage genes that inactivate the CRISPR/Cas bacterial immune system. Nature. 493(7432):429-32. doi: 10.1038/nature11723. http://www.nature.com/nature/journal/v493/n7432/abs/nature11723.html 16 Julie E. Samson, Alfonso H. Magadán, Mourad Sabri & Sylvain Moineau (2013). Revenge of the phages: defeating bacterial defences. Nature Reviews Microbiology, 11, 675–687 doi:10.1038/nrmicro3096 http://www.nature.com/nrmicro/journal/v11/n10/abs/nrmicro3096.html 11 Seung Woo Cho, Sojung Kim, Yongsub Kim, et al. (2014). Analysisof off-target effects of CRISPR/Cas-derived RNA-guided endonucleases and nickases. Genome Res. 24: 132-141 doi: 10.1101/gr.162339.113 http://genome.cshlp.org/content/24/1/132.short 12 Fu, Y., Sander, J. D., Reyon, D., Cascio, V. M., & Joung, J. K.(2014). Improving CRISPR-Cas nuclease specificity using truncated guide RNAs. Nature biotechnology. 32, 279–284 doi:10.1038/nbt.2808 http://www.nature.com/nbt/journal/v32/n3/abs/nbt.2808.html
10
заменившей двухцепочечный комплекс tracrРНК с crРНК, позволилозаменить многостадийный процесс образования этого комплекса (см.Рис. 1), легко осуществимым в лабораторных условиях, молекулярным клонированием (Рис.7). Для молекулярного клонирования фрагмент ДНК, называемый «all-in-one CRISPR-Cas9 cloning vector» (на котором записана последовательность sgРНК, а заодно и последовательность необходимая для синтеза матричной РНК белка Cas9), встраивают, к примеру, в кольцевую плазмидную ДНК. Образовавшуюся гибриднуюплазмиду вводят в бактериальную клетку. Размножаясь, бактерия многократно увеличивает и количество введенной ДНК, в точности сохраняя её структуру. В этом и состоит суть клонирования. Полученную ДНК выделяют и используют для опытов.
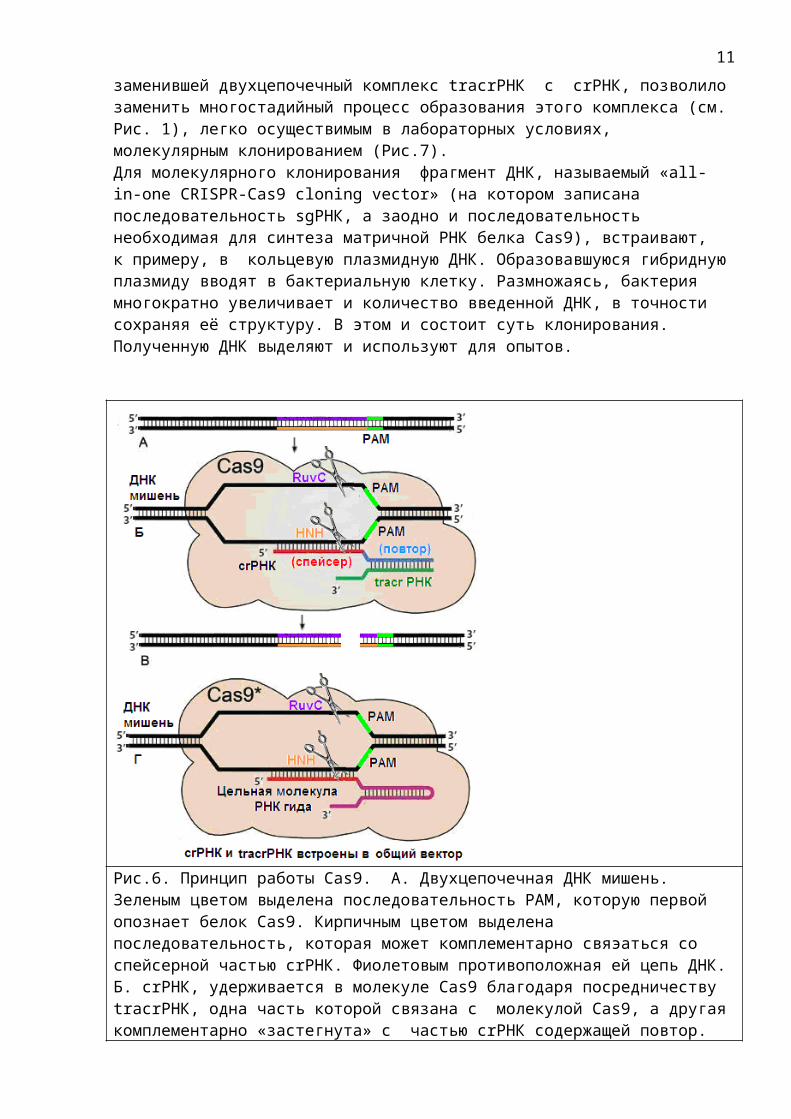
Рис.6. Принцип работы Cas9. А. Двухцепочечная ДНК мишень. Зеленым цветом выделена последовательность PAM, которую первой опознает белок Cas9. Кирпичным цветом выделена последовательность, которая может комплементарно свяэаться со спейсерной частью crРНК. Фиолетовым противоположная ей цепь ДНК.Б. crРНК, удерживается в молекуле Cas9 благодаря посредничеству tracrРНК, одна часть которой связана с молекулой Cas9, а другаякомплементарно «застегнута» с частью crРНК содержащей повтор.
11
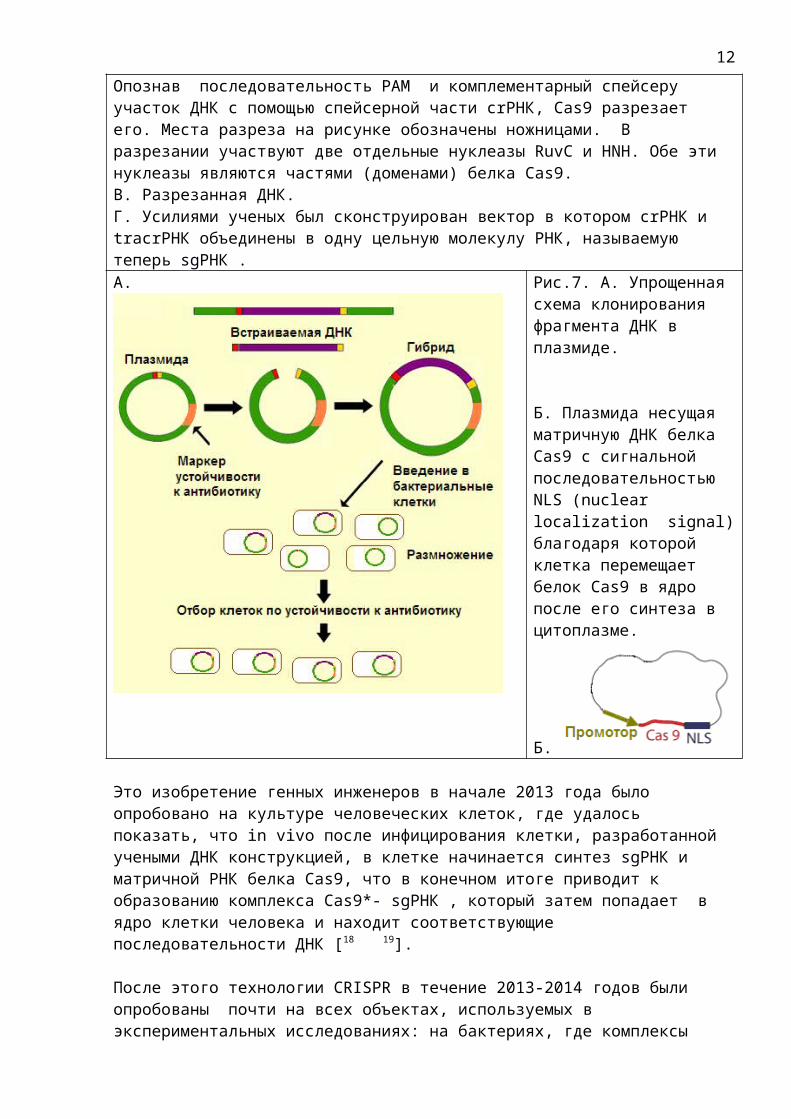
Опознав последовательность PAM и комплементарный спейсеру участок ДНК с помощью спейсерной части crРНК, Cas9 разрезает его. Места разреза на рисунке обозначены ножницами. В разрезании участвуют две отдельные нуклеазы RuvC и HNH. Обе эти нуклеазы являются частями (доменами) белка Cas9.В. Разрезанная ДНК.Г. Усилиями ученых был сконструирован вектор в котором crРНК и tracrРНК объединены в одну цельную молекулу РНК, называемую теперь sgРНК . А. Рис.7. А. Упрощенная
схема клонирования фрагмента ДНК в плазмиде.
Б. Плазмида несущая матричную ДНК белка Cas9 с сигнальной последовательностью NLS (nuclear localization signal)благодаря которой клетка перемещает белок Cas9 в ядро после его синтеза в цитоплазме.
Б.
Это изобретение генных инженеров в начале 2013 года было опробовано на культуре человеческих клеток, где удалось показать, что in vivo после инфицирования клетки, разработанной учеными ДНК конструкцией, в клетке начинается синтез sgРНК и матричной РНК белка Cas9, что в конечном итоге приводит к образованию комплекса Cas9*- sgРНК , который затем попадает в ядро клетки человека и находит соответствующие последовательности ДНК [18 19].
После этого технологии CRISPR в течение 2013-2014 годов были опробованы почти на всех объектах, используемых в экспериментальных исследованиях: на бактериях, где комплексы
12

Cas9- sgРНК были использованы в качестве высокоизбирательного «умного» антибиотика, для селективного удаления из смешанных культур отдельных видов и штаммов бактерий [20]; на дрожжах, гдеони продемонстрировали себя, как простой и мощный инструмент генной инженерии для мутагенеза и замены аллельных участков [21];на спроектированных винных мушках, которых теперь можно получитьвсего за месяц [22 23 24]; на эмбрионах рыбок данио рерио, для целенаправленной наследуемой генетической модификации in vivo; для прицельного мутагенеза на эмбрионах мыши [25 26 27] и обезьяны [28]. Оказалось, что с помощью инъекции матричной РНК Cas9 и sgRNA в клетки зиготы можно быстро, всего за одну стадию,получить свиней с модифицированным геномом и таким образом создавать модели для изучения генетических заболеваний человека и животных и тестировать на них новые методики лечения [29]. Можно этот метод генной инженерии также применять и для улучшения пород в животноводстве [30] и растений для сельского хозяйства [31]. Помимо вышеизложенного технология CRISPR предоставляет возможность вырезать из генома мутировавшую ДНК и заменить ее правильной последовательностью, что позволит лечитьмногие генетические заболевания [32], в том числе мышечную дистрофию [33], β-талассемию [34], наследственную тирозинемию, причем прямо в организме in vivo [35], что демонстрирует возможностьлечебной корректировки генетических заболеваний у взрослого человека.
Рис.8. Использование Cas9* в качестве высокоизбирательного «умного» антибиотика, для селективного удаления отдельных видов и штаммов бактерий из смешанных культур. В перспективе такой умный антибиотик можно будет использовать и для лечения человекаот устойчивых к обычным антибиотикам бактерий и вирусов [36 37].
13

Рис.9. Примеры генноймодификации с использованием CRISPR-Cas9.А. У нематод. «Коренастый» мутант (верхнее фото)Б. У эмбрионов рыбокданио (избыток вентральной ткани)В. У плодовых мушек
2 Mojica, F. J., Diez-Villasenor, C., Garcia-Martinez, J. & Soria, E. (2005) Intervening sequences of regularly spaced prokaryotic repeats derive from foreign genetic elements. J. Mol. Evol. 60,174–182 http://link.springer.com/article/10.1007/s00239-004-0046-3 3 Barrangou R, et al. & Horvath P (2007). CRISPR provides acquired resistance against viruses in rokaryotes.Science ;315:1709-1712. http://www.sciencemag.org/content/315/5819/1709.short 5 Deltcheva, E., Chylinski, K., Sharma, C. M., Gonzales, K., Chao, Y., Pirzada, Z. A., ... & Charpentier, E. (2011). CRISPR RNA maturation by trans-encoded small RNA and host factor RNase III. Nature, 471(7340), 602-607. http://www.nature.com/nature/journal/v471/n7340/abs/nature09886.html 10 David Bikard and Luciano A. Marraffini (2013) Control of gene expression by CRISPR-Cas systems. F1000Prime Rep. 2013; 5: 47. doi: 10.12703/P5-47 http://www.ncbi.nlm.nih.gov/pmc/articles/PMC3816762/ 13 Briner, A. E., Donohoue, P. D., Gomaa, A. A., Selle, K., Slorach, E. M., Nye, C. H., ... & Barrangou, R. (2014). Guide RNA Functional Modules Direct Cas9 Activity and Orthogonality. Molecular Cell. http://www.sciencedirect.com/science/article/pii/S1097276514007515 14 Wu, X., Kriz, A. J., & Sharp, P. A. Target specificity of the CRISPR-Cas9 system. Quantitative Biology, 1-12. http://link.springer.com/article/10.1007/s40484-014-0030-x 17 Martin Jinek, Krzysztof Chylinski, Ines Fonfara, et al. (2012).A programmable dual-RNA-guided DNA endonuclease in adaptive bacterial immunity. Science, 337, 816-821, doi: 10.1126/science.1225829 http://www.sciencemag.org/content/337/6096/816.short 18 Martin Jinek, Alexandra East, Aaron Cheng, Steven Lin, Enbo Ma,Jennifer Doudna
14
А
Б
В Г
ДГ

Эти исследования уже не ограничиваются только опытами на животных, программируемые РНК гидом, эндонуклеазы (RGENs) были использованы для целенаправленного редактирования генома в клетках человека [39], Предполагается, что редактирование генома в культивируемых клетках человека сможет помочь выяснениюфункциональной роли различных генов. Выключая с помощью CRISPR-Cas9 нокаута (Gecko) отдельные гены можно идентифицировать те изних, которые, например, необходимы для жизнеспособности клеток
(2013) RNA-programmed genome editing in human cells. . eLife; 2:e00471 DOI: http://dx.doi.org/10.7554/eLife.00471 http://elifesciences.org/content/2/e00471.short 19 Cho, S.W.; Kim, S.; Kim, J.M.; Kim, J.S. Targeted genome engineering in human cells with the Cas9 RNA-guided endonuclease. Nat. Biotechnol. 2013, 31, 230–232 doi:10.1038/nbt.2507 http://www.nature.com/nbt/journal/v31/n3/abs/nbt.2507.html 20 Gomaa, A. A., Klumpe, H. E., Luo, M. L., Selle, K., Barrangou, R., & Beisel, C. L. (2014). Programmable Removal of Bacterial Strains by Use of Genome-Targeting CRISPR-Cas Systems. mBio, 5(1), e00928-13. doi: 10.1128/mBio.00928-13 http://mbio.asm.org/content/5/1/e00928-13.short 21 DiCarlo JE, Norville JE, Mali P, Rios X, et al. (2013). Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucleic Acids Res 41: 4336–43. http://nar.oxfordjournals.org/content/early/2013/03/03/nar.gkt135.short 22 Gratz SJ, Wildonger J, Harrison MM, O'Connor-Giles KM (2013) .CRISPR/Cas9-mediated genome engineering and the promise of designer flies on demand. Fly (Austin), 7(4). 249–255; http://www.landesbioscience.com/journals/fly/2013FLY0024R.pdf 23 Yu Z, Ren M, Wang Z, Zhang B, et al. (2013). Highly efficient genome modifications mediated by CRISPR/Cas9 in Drosophila. Genetics 195(1): 289–91. http://www.genetics.org/content/195/1/289.short 24 Gratz, S. J., Ukken, F. P., Rubinstein, C. D. et al. & O'Connor-Giles, K. M. (2014). Highly Specific and Efficient CRISPR/Cas9-Catalyzed Homology-Directed Repair in Drosophila. Genetics, genetics-113.160713 doi: 10.1534/genetics.113.160713 http://www.genetics.org/content/196/4/961.short 25 Sung, Y. H., Kim, J. M., Kim, H. T. et al. & Kim, J. S. (2014). Highly efficient gene knockout in mice and zebrafish with RNA-guided endonucleases. Genome research, 24(1), 125-131. (Novembe, 2013), . Genome Res. . 24: 125-131
15

при раке, что необходимо знать для разработки эффективных путей лечения этого страшного заболевания [40]. Обнаружив соматические мутации, которые ведут к развитию рака, можно будетс помощью эндонуклеаз RGEN провести генную терапию убрав вредныемутации.
. Для целенаправленного разрезания ДНК генные инженеры до 2013 года обычно использовали искусственные ферменты, получаемые путем слияния ДНК-разрезающего домена нуклеазы либо с ДНК-
doi:10.1101/gr.163394.113 http://genome.cshlp.org/content/24/1/125.short 26 Jiankui Zhou, Bin Shen, Wensheng Zhang, et al. & Xingxu Huang(2014). One-step generation of different immunodeficient mice with multiple gene modifications by CRISPR/Cas9 mediated genome engineering. The International Journal of Biochemistry & Cell Biology, 46, 49–55 http://www.sciencedirect.com/science/article/pii/S1357272513003270 27 Harms, D. W., Quadros, R. M., Seruggia, D., Ohtsuka, M., Takahashi, G., Montoliu, L., & Gurumurthy, C. B. (2014). Mouse Genome Editing Using the CRISPR/Cas System. Current Protocols in Human Genetics, 15-7. http://onlinelibrary.wiley.com/doi/10.1002/0471142905.hg1507s83/abstract 28 Niu, Y., Shen, B., Cui, Y., et al. & Sha, J. (2014). Generation of Gene-Modified Cynomolgus Monkey via Cas9/RNA-Mediated Gene Targeting in One-Cell Embryos. Cell. 156(4), 836-843 http://www.sciencedirect.com/science/article/pii/S0092867414000798 29 Peng, Y., Clark, K. J., Campbell, J. M., Panetta, M. R., Guo, Y., & Ekker, S. C. (2014). Making designer mutants in model organisms. Development,141(21), 4042-4054 doi: 10.1242/dev.102186 http://dev.biologists.org/content/141/21/4042.abstract 30 Tang Hai, Fei Teng, Runfa Guo, Wei Li and Qi Zhou (2014). One-step generation of knockout pigs by zygote injection of CRISPR/Cas system. Cell Research.; doi: 10.1038/cr.2014.11 http://www.nature.com/cr/journal/v24/n3/full/cr201411a.html 31 Khaoula Belhaj, Angela Chaparro-Garcia, Sophien Kamoun and Vladimir Nekrasov (2013). Plant genome editing made easy: targeted mutagenesis in model and crop plants using the CRISPR/Cas system. Plant Methods 2013, 9:39 doi:10.1186/1746-4811-9-39 http://www.biomedcentral.com/content/pdf/1746-4811-9-39.pdf 32 Yin, H., Xue, W., Chen, S., Bogorad, R. L., Benedetti, E., Grompe, M., ... & Anderson, D. G. (2014). Genome editing with Cas9 in adult mice corrects a disease mutation and phenotype. Nature Biotechnology. doi:10.1038/nbt.2884 http://www.nature.com/nbt/journal/v32/n6/full/nbt.2884.html
16

связывающим доменом, так называемых цинковых пальцев (такую нуклеазу называют ZFN - Zinc-finger nuclease), либо с ДНК-связывающим доменом TAL эффектора (такую нуклеазу называют TALEN - Transcription activator-like effector nuclease). И те и другие требовали длительного кропотливого труда по разработке новой белковой конструкции для каждой новой ДНК мишени. По сравнению с ними система CRISPR, где в качестве нуклеазы (такую нуклеазу называют RGEN - RNA-guided endonuclease) используется один и тот же белок - Cas9, оказаласьнамного проще и дешевле, потому что для изготовления новой sgРНК требуется сменить всего 20 нуклеотидов в спейсерной части [41]. То, на что уходили месяцы и годы труда, удается сделать за неделю. За короткие сроки была синтезирована целая библиотека РНК гидов с различными последовательностями - 73,000 различных sgРНК [42].С помощью этого грандиозного набора sgРНК можно направлять Cas9на 80- 90% всех последовательностей генома человека. Для повышения эффективности и упрощения экспериментов по генной инженерии in vivo была создана линия мышей, организм которых, при определенных условиях сам производит Cas9 [43].Наряду с обычным «природным» Cas9, который осуществляет двухцепочечные разрывы ДНК, ученые используют его измененные формы, такие как: (А). Cas9 никаза - белок, у которого активна только одна из егонуклеаз, а другая инактивирована – благодаря чему Cas9 никаза вносит разрыв только на одной из двух цепей ДНК. Используя парочку таких никаз с разными sgРНК, можно значительно повысить точность генного скальпеля и кроме того вырезать большие участкиДНК [44].33 Long, C., McAnally, J. R., Shelton, J. M., Mireault, A. A., Bassel-Duby, R., & Olson, E. N. (2014). Prevention of muscular dystrophy in mice by CRISPR/Cas9–mediated editing of germline DNA. Science, 345(6201), 1184-1188. http://www.sciencemag.org/content/345/6201/1184.short 34 Xie, F., Ye, L., Chang, J. C., Beyer, A. I., Wang, J., Muench, M. O., & Kan, Y. W. (2014). Seamless gene correction of β-thalassemia mutations in patient-specific iPSCs using CRISPR/Cas9 and piggyBac. Genome research, 24(9), 1526-1533. http://genome.cshlp.org/content/24/9/1526.short 35 Yin, H., Xue, W., Chen, S., Bogorad, R. L., Benedetti, E., Grompe, M., ... & Anderson, D. G. (2014). Genome editing with Cas9 in adult mice corrects a disease mutation and phenotype. Nature biotechnology. 32, 551–553 http :// www . nature . com / nbt / journal / v 32/ n 6/ full / nbt .2884. html
17

(Б). dCas9 – мутантный белок, у которого инактивированы обе нуклеазы. Белок dCas9 может находить ДНК мишень, но не способен осуществить ее разрезание. Казалось бы, для чего может пригодиться такой инактивированный белок? Но ученые нашли для него массу способов применения. Оказалось, что его можно соединить с нуклеазой FokI. Полученный в результате такого слияния белок мономер dCas9-FokI может разрезать ДНК только в том случае, если он образует димер с. другим мономером FokI-dCas9. Благодаря этому специфичность связывания зависит от двухразных sgРНК [45], а значит значительно повышается.Помимо этого мутантный белок dCas9 можно использовать для инактивации генов на стадии транскрипции, блокируя продвижение РНК-полимеразы (способ, называемый CRISPR интерференцией или CRISPRi, от англ. CRISPR interference ) [46], а также для создания искусственных регуляторов транскрипции. Предполагается использовать систему CRISPR / dCas9 для безопасной активации экспрессии такого противовоспалительного и анти-апоптозного гена, каким является гемоксигеназа-1 (HO-1), чтобы повысить выживаемость трансплантированных стволовых клеток в в ишемизированном миокарде [47].
Как известно, транскрипция регулируется белками, называемыми факторами транскрипции. Эти белки связываются со специфическими последовательностями ДНК в промоторной области гена и либо активируют, либо наоборот блокируют ферменты, необходимые для копирования этого гена в мРНК. Так вот, если к белку dCas9 добавить домен транскрипционного фактора, который активирует илиподавляет активность генов, воздействуя на транскрипционный механизм клетки, а затем снабдить его соответствующей sgРНК, которая доставит эту конструкцию на нужный промотор гена, можно в итоге избирательно изменять активность выбранного гена по волеисследователя [48]. Сменив sgРНК такой транскрипционный фактор набазе dCas9 можно перенаправить на промотор другого гена [49 50].Разработан набор правил - «грамматика» - описывающих, как спроектировать синтетический транскрипционный фактор (STFS) и создана программа для его автоматизированного проектирования [51].перестраиваемый модуляция генных сетей. Разрабатываются дажестратегии для «перестраиваемой модуляции генных сетей. в том числе многоступенчатых каскадов генной регудяции» [52], посколькуCRISPR-Cas9 оказался надежной технологией, позволяющей проводитьсистематическое исследование клеточных последствий подавления или индукции отдельных транскриптов [53].
18
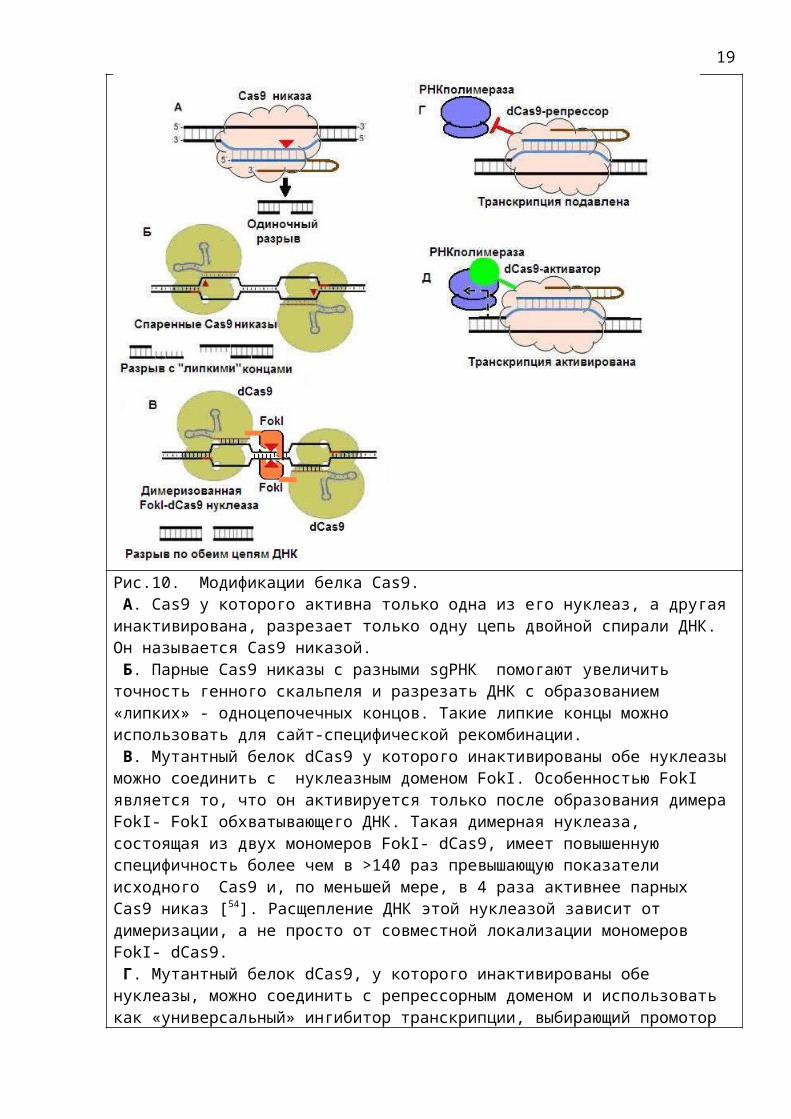
Рис.10. Модификации белка Cas9. А. Cas9 у которого активна только одна из его нуклеаз, а другаяинактивирована, разрезает только одну цепь двойной спирали ДНК. Он называется Cas9 никазой. Б. Парные Cas9 никазы с разными sgРНК помогают увеличить точность генного скальпеля и разрезать ДНК с образованием «липких» - одноцепочечных концов. Такие липкие концы можно использовать для сайт-специфической рекомбинации. В. Мутантный белок dCas9 у которого инактивированы обе нуклеазыможно соединить с нуклеазным доменом FokI. Особенностью FokI является то, что он активируется только после образования димераFokI- FokI обхватывающего ДНК. Такая димерная нуклеаза, состоящая из двух мономеров FokI- dCas9, имеет повышенную специфичность более чем в >140 раз превышающую показатели исходного Cas9 и, по меньшей мере, в 4 раза активнее парных Cas9 никаз [54]. Расщепление ДНК этой нуклеазой зависит от димеризации, а не просто от совместной локализации мономеров FokI- dCas9. Г. Мутантный белок dCas9, у которого инактивированы обе нуклеазы, можно соединить с репрессорным доменом и использовать как «универсальный» ингибитор транскрипции, выбирающий промотор
19

гена по наводке sgРНК. Д. Если белок dCas9 соединить с доменом активатором (например,VP64), то он может избирательно активировать синтез определенныхгенов. См. также рис 2 в [55 ].
Прикрепив к белку dCas9 зеленый флуоресцентный белок EGFP можно получить инструмент для визуального наблюдения за поведением выбранных исследователем последовательностей в хромосомах живых клеток млекопитающих в течение клеточного цикла, а также визуально определять длину теломер хромосомы [56].
Рис.11. Прикрепив к белку dCas9зеленый флуоресцентный белок EGFP можно получить инструмент для визуального наблюдения за поведением выбранных исследователемпоследовательностей в хромосомах живых клеток
Перспективы
. Благодаря беспрецедентной эффективности и потрясающей простотеи дешевизне в использовании, технология редактирования ДНК, основанная на прокариотической системе CRISPR-Cas9 совершила революцию в генной инженерии, За неполных 2 года было опубликовано свыше 700 статей. Редактирование с ее помощью было успешно осуществлено во множестве модельных организмов и типов клеток. Крошечная молекулярная машина, используемая бактериями для защиты от атакующих ее вирусов, позволила разработать принципиально новые методы генной терапии, с помощью которых можно будет лечить многие болезни, а также изучать пути генной регуляции.
Литература
20

21

22

23




















