
A gamut of undiscovered electrophysiological effects produced by Tityus serrulatus toxin 1 on...
9
A gamut of undiscovered electrophysiological Q1 effects produced by Tityus serrulatus toxin 1 on Na V -type isoforms Q6 Q5 Steve Peigneur a , Camilla T. Cologna b , Caroline M. Cremonez b , Bea G. Mille a , Manuela B. Pucca b , Eva Cuypers a , Eliane C. Arantes b , Jan Tytgat a, * a Toxicology and Pharmacology, University of Leuven (KU Leuven), Campus Gasthuisberg, O&N2, PO Box 922, Herestraat 49, 3000 Leuven, Belgium b Physics and Chemistry Department, Faculty of Pharmaceutical Sciences, Ribeir~ ao Preto e USP, Ribeir~ ao Preto, SP, Brazil article info Article history: Received 4 December 2014 Received in revised form 18 March 2015 Accepted 24 March 2015 Available online xxx Keywords: Voltage-gated sodium channel Scorpion toxin Tityus serrulatus Bell-shaped modulation Ion channel modulator Binding site abstract In the last decades, Ts1 has not only been the subject of many studies, it has also been considered as a very useful tool to investigate Na V channels and to explore the exact role of Na V channels in channe- lopathies. Ts1 is believed to modulate the activation process of Na V upon interaction at the neurotoxin binding site 4. Our aim was to carry out an in depth functional characterization of Ts1 on a wide array of Na v channels, in order to investigate its mechanism of action and to verify if Ts1 can indeed be considered as a prototype site 4 selective toxin, valid for all the Na v isoforms we know currently. Ts1 has been subjected to an in-depth functional investigation on 9 Na V isoforms expressed in Xenopus laevis oocytes. Ts1 does not only interfere with the activation process but also modulates the inactivation in a bell- shaped voltage-dependent matter. Furthermore, Ts1 altered the ion selectivity through insect Na V . without influencing the tetrodotoxin selectivity of the channels. Finally, Ts1 was also found to inhibit the sodium current through the cardiac Na v 1.5 isoform. On the basis of the totally unexpected plethora of Na v modulations as induced by Ts1, we demonstrate that caution is required in interpretation the in vivo experiments when using Ts1. The electrophysio- logical characterization of Ts1 indeed shows that the general accepted contours of Na V binding sites are much more obscure than believed and that interpretation of Na V pharmacology upon toxin binding is more complex than believed thus far. Q3 © 2015 Elsevier Ltd. All rights reserved. 1. Introduction Tityus serrulatus is still considered to be one of the most medi- cally important scorpions in South America with a strong impact on society due to a high envenomation rate, often with fatal outcome. In order to provide an efficient treatment against envenomation, there is a strong urge to fully understand on a molecular level the exact targets of each venom component, and moreover, to char- acterize in detail the pharmacological effects induced by binding of individual components at their target. In the last decades, Ts1 has been the subject of intense research since this peptide toxin, which is toxic to both insects and mammals, contributes significantly to the observed clinical manifestations after T. serrulatus envenom- ation. Its structure has been determined by X-ray crystallography showing a highly conserved dense core formed by three antiparallel b-strands and an a-helix interlinked by disulfide bridges. Voltage-gated sodium channels (Na V channels) are trans- membrane protein complexes constituted of an a-subunit of approximately 260 kDa which can be associated with up to four auxiliary b-subunits (b1-4) of 30e40 kDa. The pore-forming a- subunit alone is sufficient to obtain sodium current, however co- expression of b-subunits modifies expression level, kinetics and voltage dependence of channel gating (Yu and Catterall, 2003). The a-subunit is organized in four homologous domains (DI-IV). Each domain contains six putative transmembrane segments (S1eS6) connected by extracellular or intracellular loops. The S4 segments are the most conserved segments and they contain a basic residue, either lysine or arginine, in every third position. These positive charged S4 segments are believed to function as voltage sensors. They transport gating charges by moving outward upon membrane depolarization and as such initiating the voltage dependent acti- vation which results in the opening of the channel. The selectivity * Corresponding author. Tel.: þ32 16 323403; fax: þ55 16 3602 4880. Q2 E-mail address: [email protected] (J. Tytgat). Contents lists available at ScienceDirect Neuropharmacology journal homepage: www.elsevier.com/locate/neuropharm http://dx.doi.org/10.1016/j.neuropharm.2015.03.027 0028-3908/© 2015 Elsevier Ltd. All rights reserved. Neuropharmacology xxx (2015) 1e9 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 49 50 51 52 53 54 55 56 57 58 59 60 61 62 63 64 65 66 67 68 69 70 71 72 73 74 75 76 77 78 79 80 81 82 83 84 85 86 87 88 89 90 91 92 93 94 95 96 97 98 99 100 101 102 103 104 105 106 107 108 109 110 111 112 113 114 115 116 117 118 119 NP5799_proof ■ 9 April 2015 ■ 1/9 Please cite this article in press as: Peigneur, S., et al., A gamut of undiscovered electrophysiological effects produced by Tityus serrulatus toxin 1 on Na V -type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1016/j.neuropharm.2015.03.027
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A gamut of undiscovered electrophysiological effects produced by Tityus serrulatus toxin 1 on...

Q1
Q6
Q5
Q3
Q2
lable at ScienceDirect
Neuropharmacology xxx (2015) 1e9
123456789101112131415161718192021222324252627282930313233343536373839404142434445464748495051525354
55
NP5799_proof ■ 9 April 2015 ■ 1/9
Contents lists avai
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
565758596061626364656667686970717273
A gamut of undiscovered electrophysiological effects produced byTityus serrulatus toxin 1 on NaV-type isoforms
Steve Peigneur a, Camilla T. Cologna b, Caroline M. Cremonez b, Bea G. Mille a,Manuela B. Pucca b, Eva Cuypers a, Eliane C. Arantes b, Jan Tytgat a, *
a Toxicology and Pharmacology, University of Leuven (KU Leuven), Campus Gasthuisberg, O&N2, PO Box 922, Herestraat 49, 3000 Leuven, Belgiumb Physics and Chemistry Department, Faculty of Pharmaceutical Sciences, Ribeir~ao Preto e USP, Ribeir~ao Preto, SP, Brazil
7475
7677787980818283848586878889a r t i c l e i n f o
Article history:Received 4 December 2014Received in revised form18 March 2015Accepted 24 March 2015Available online xxx
Keywords:Voltage-gated sodium channelScorpion toxinTityus serrulatusBell-shaped modulationIon channel modulatorBinding site
* Corresponding author. Tel.: þ32 16 323403; fax:E-mail address: [email protected] (J.
http://dx.doi.org/10.1016/j.neuropharm.2015.03.0270028-3908/© 2015 Elsevier Ltd. All rights reserved.
9091929394
Please cite this article in press as: Peigneur,on NaV-type isoforms, Neuropharmacology
a b s t r a c t
In the last decades, Ts1 has not only been the subject of many studies, it has also been considered as avery useful tool to investigate NaV channels and to explore the exact role of NaV channels in channe-lopathies. Ts1 is believed to modulate the activation process of NaV upon interaction at the neurotoxinbinding site 4. Our aim was to carry out an in depth functional characterization of Ts1 on a wide array ofNav channels, in order to investigate its mechanism of action and to verify if Ts1 can indeed be consideredas a prototype site 4 selective toxin, valid for all the Nav isoforms we know currently. Ts1 has beensubjected to an in-depth functional investigation on 9 NaV isoforms expressed in Xenopus laevis oocytes.Ts1 does not only interfere with the activation process but also modulates the inactivation in a bell-shaped voltage-dependent matter. Furthermore, Ts1 altered the ion selectivity through insect NaV.without influencing the tetrodotoxin selectivity of the channels. Finally, Ts1 was also found to inhibit thesodium current through the cardiac Nav1.5 isoform.
On the basis of the totally unexpected plethora of Nav modulations as induced by Ts1, we demonstratethat caution is required in interpretation the in vivo experiments when using Ts1. The electrophysio-logical characterization of Ts1 indeed shows that the general accepted contours of NaV binding sites aremuch more obscure than believed and that interpretation of NaV pharmacology upon toxin binding ismore complex than believed thus far.
© 2015 Elsevier Ltd. All rights reserved.
9596
97 9899100101102103104105106107108109110111112
1. Introduction
Tityus serrulatus is still considered to be one of the most medi-cally important scorpions in South Americawith a strong impact onsociety due to a high envenomation rate, often with fatal outcome.In order to provide an efficient treatment against envenomation,there is a strong urge to fully understand on a molecular level theexact targets of each venom component, and moreover, to char-acterize in detail the pharmacological effects induced by binding ofindividual components at their target. In the last decades, Ts1 hasbeen the subject of intense research since this peptide toxin, whichis toxic to both insects and mammals, contributes significantly tothe observed clinical manifestations after T. serrulatus envenom-ation. Its structure has been determined by X-ray crystallography
þ55 16 3602 4880.Tytgat).
113114115116
S., et al., A gamut of undiscove(2015), http://dx.doi.org/10.1
showing a highly conserved dense core formed by three antiparallelb-strands and an a-helix interlinked by disulfide bridges.
Voltage-gated sodium channels (NaV channels) are trans-membrane protein complexes constituted of an a-subunit ofapproximately 260 kDa which can be associated with up to fourauxiliary b-subunits (b1-4) of 30e40 kDa. The pore-forming a-subunit alone is sufficient to obtain sodium current, however co-expression of b-subunits modifies expression level, kinetics andvoltage dependence of channel gating (Yu and Catterall, 2003). Thea-subunit is organized in four homologous domains (DI-IV). Eachdomain contains six putative transmembrane segments (S1eS6)connected by extracellular or intracellular loops. The S4 segmentsare the most conserved segments and they contain a basic residue,either lysine or arginine, in every third position. These positivecharged S4 segments are believed to function as voltage sensors.They transport gating charges by moving outward upon membranedepolarization and as such initiating the voltage dependent acti-vation which results in the opening of the channel. The selectivity
117118119
red electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027

S. Peigneur et al. / Neuropharmacology xxx (2015) 1e92
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475767778798081828384858687888990919293949596979899
100101102103104105106107108109110111112113114115116117118119120121122123124125126127128129130
NP5799_proof ■ 9 April 2015 ■ 2/9
filter and pore are formed by the transmembrane segments S5 andS6 together with the re-entrant segments that are part of the loopwhich connects the S5 and S6 of each domain. Folding of the do-mains in a clockwise orientation, in which domain I and IV are inclose proximity of each other, leads to the formation of the outervestibule and the selectivity filter (Catterall, 2000; Chanda andBezanilla, 2002). The short intracellular linker that connects thedomains III and IV contains a highly conserved sequence of threehydrophobic residues (isoleucine, phenylalanine and methionine)or IFM motif. Sodium channel inactivation is mediated by this hy-drophobic motif since it serves as an inactivation gate crucial forcausing fast inactivation by binding to a receptor. This inactivationgate receptor is located near or within the intracellular mouth ofthe sodium channel pore. It has been shown that several residues inthe intracellular loop that connects IIIS4eS5 and in the loop con-necting IVS4eS5 are contributing to the inactivation gate receptor(Dong, 2007; Yu and Catterall, 2003).
Neurotoxins acting on NaV channels can aim at six different sitesin the channels, distinguished not only by matters of localization ofthe toxin binding place but also by the results of the toxin's action.Here we describe a brief overview of what is known of the NaVbinding sites relevant for this work. There are several excellentreviews available describing the NaV channel binding sites and theirinteraction with toxins in detail (Catterall, 2012; Stevens et al.,2012). The amino acid residues that form the neurotoxin receptorsite 1 are primarily located in the pore loop which is formed by themembrane dipping part of the connecting loop between S5 and S6of each domain (Catterall et al., 2007). This site is occupied by thewater soluble heterocyclic guanidine tetrodotoxin (TTX) which isisolated from the tissue of at least 40 different species of puffer fishbut it can also be found in mollusks, crabs, octopus and frogs(Catterall et al., 2007; Hwang et al., 1991). TTX exerts its strongtoxicity and high fatality, also towards humans, by binding withinthe outer vestibule of the channel, physically occluding the ionpathway. Based on their sensitivity to TTX have the mammalianisoforms been divided into TTX-sensitive (NaV1.1eNaV1.4, NaV1.6and NaV1.7) or TTX-insensitive (NaV1.5, Nav1.8, NaV1.9) (Narahashi,2008). Besides TTX, it has also been shown that m-conotoxins,neurotoxin peptides isolated from Cone snail species, bind to amicro site within the neurotoxin receptor site 1 (Stephan et al.,1994).
The neurotoxin receptor 3 is mainly localized at the extracellularloop connecting the segments S3 and S4 from domain IV. It isbelieved that other parts of the channel such as the extracellularloops between the S5 and S6 of domain I and IV also contributesignificantly to channel recognition and binding of site 3 toxins(Bosmans and Tytgat, 2007). The voltage sensors of each domainwill normally move outward under influence of the electric fieldwhen the membrane is depolarized (Hille, 2001). Upon binding atsite 3 these toxins trap the voltage sensing segment S4 of domain IVin its inward position. As such they prevent the normal outwardmovement of these voltage sensors and herewith the conforma-tional changes necessary for fast inactivation (Catterall et al., 2007).
Summarized, toxin binding at neurotoxin receptor 3 affects thecoupling of activation and inactivation, resulting in a slowing downor inhibition of the fast inactivation. Several studies have demon-strated the functionally relevant structural differences in insect andmammalian receptor site 3 regions (Gordon et al., 1996).
Toxin binding at site 4 causes a shift in the voltage dependenceof activation towards more hyperpolarized membrane potentialsand reduces the peak sodium current amplitude (Cestele et al.,2006; Vijverberg et al., 1984). The shift in voltage dependence ofNaV channel activating causes channels to open at, or close to, theresting potential. This increase in open channel probability leads torepetitive firing and consequently increases the influx of Ca2þ into
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscovon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
the nerve terminals resulting in an increased frequency of minia-ture excitatory junctional potentials (King et al., 2008). The alter-ations in channel gating are believed to be a direct result of toxinbinding at site 4 leading to a trapping of the voltage sensor in itsoutward, activated position (Cestele et al., 2001, 2006). The receptorsite 4 has been primarily defined to specific residues in the extra-cellular loops connecting the S1eS2 and S3eS4 segments ofdomain II (Catterall et al., 2007). However, using the scorpion b-toxin Tz1 (Tityus zulianus) it was shown that three residues in thepore loop of domain III are determinant for the specificity of b-toxinfor different sodium channel isoforms (Leipold et al., 2006).
In this work we investigated the subtype selectivity of Ts1,believed to be a classical b-scorpion toxin, and it was found that thistoxin exerts surprisingly different pharmacological activities upondifferent isoforms. The obtained electrophysiological profile of Ts1confirms that the generally accepted contours of the NaV bindingsites are more obscure than believed and that a revision of existingnomenclature of the binding sites is needed.
2. Material and methods
2.1. Ts1 isolation
Ts1 was obtained from crude venom using the improved method described byCerni et al. (2014). T. serrulatus venom was fractionated on a CM-cellulose-52 col-umn connected to a Fast Protein Liquid Chromatography (FPLC) system. Pure Ts1wasobtained from fraction XIII eluted from CM-cellulose-52. A second purification stepwas performed on a C18 column (buffer A: H2Oþ 0.1% TFA; buffer B: 80% ACNþ 0.1%TFA). To ascertain ourselves that no impurities were present, sample purity wasverified by mass spectrometry and protein sequencing. Mass spectrometry analysisfor molecular mass determination was performed by an electrospray triple-quadrupole mass spectrometer. Amino acid sequence determination of nativetoxin was performed by Edman degradation using a Protein Sequencer PPSQ-33A(Shimadzu Corp.). No impurities were found.
2.2. Expression of voltage-gated ion channels in Xenopus laevis oocytes
For the expression of NaV channels (rNaV1.2, rNaV1.3, rNaV1.4, hNaV1.5, mNaV1.6,rNaV1.7, rNaV1.8, the insect channels DmNaV1, the auxiliary subunits rb1, hb1 andTipE) in Xenopus oocytes, the linearized plasmids were transcribed using the T7 orSP6 mMESSAGE-mMACHINE transcription kit (Ambion®, Carlsbad, California, USA).The harvesting of stage V-VI oocytes from anaesthetized female X. laevis frog waspreviously described (Liman et al., 1992). Oocytes were injected with 50 nL of cRNAat a concentration of 1 ng/nL using a micro-injector (Drummond Scientific®, Broo-mall, Pennsylvania, USA). The oocytes were incubated in a solution containing (inmM): NaCl, 96; KCl, 2; CaCl2, 1.8; MgCl2, 2 and HEPES, 5 (pH 7.4), supplemented with50 mg/L gentamycin sulfate.
2.3. Electrophysiological recordings
Two-electrode voltage-clamp recordings were performed at room temperature(18e22 �C) using a Geneclamp 500 amplifier (Molecular Devices®, Downingtown,Pennsylvania, USA) controlled by a pClamp data acquisition system (Axon In-struments®, Union City, California, USA). Whole cell currents from oocytes wererecorded 1e4 days after injection. Bath solution compositionwas (in mM): NaCl, 96;KCl, 2; CaCl2, 1.8; MgCl2, 2 and HEPES, 5 (pH 7.4). Voltage and current electrodeswere filled with 3 M KCl. Resistances of both electrodes were kept between 0.8 and1.5 MU. The elicited currents were filtered at 1 kHz and sampled at 20 kHz using afour-pole low-pass Bessel filter. Leak subtraction was performed using a -P/4 pro-tocol. In order to avoid overestimation of a potential toxin-induced shift in thecurrentevoltage relationships of inadequate voltage control when measuring largesodium currents in oocytes, only data obtained from cells exhibiting currents withpeak amplitude below 2 mA were considered for analysis. For the electrophysio-logical analysis of toxins a number of protocols were applied from a holding po-tential of�90mVwith a start-to-start interval of 0.2 Hz. Sodium current traces wereevoked by 100 ms depolarizations to Vmax (the voltage corresponding to maximalsodium current in control conditions). The currentevoltage relationships weredetermined by 50-ms step depolarizations between �90 and 70 mV, using 5-mVincrements. The sodium conductance (gNa) was calculated from the currents ac-cording to Ohm's law: gNa¼ INa/(V� Vrev), where INa represents the Naþ current peakamplitude at a given test potential V, and Vrev is the reversal potential. The values ofgNa were plotted as a function of voltage and fitted using the Boltzmann equation:gNa/gmax ¼ [1 þ (exp(Vg � V)/k)]�1, where gmax represents maximal gNa, Vg is thevoltage corresponding to half-maximal conductance and k is the slope factor. Toxin-induced effects on the steady-state inactivation were investigated using a standardtwo-step protocol. In this protocol, 100-ms conditioning 5-mV step prepulsesranging from �90 to 70 mV were followed by a 50-ms test pulse to �30 or �10 mV.
ered electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027

S. Peigneur et al. / Neuropharmacology xxx (2015) 1e9 3
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475767778798081828384858687888990919293949596979899
100101102103104105
NP5799_proof ■ 9 April 2015 ■ 3/9
Data were normalized to the maximal Naþ current amplitude, plotted against pre-pulse potential and fitted using the Boltzmann equation: INa/Imax ¼ [(1 � C)/(1 þ exp((V � Vh)/k))] þ C, where Imax is the maximal INa, Vh is the voltage corre-sponding to half-maximal inactivation, V is the test voltage, k is the slope factor, andC is a constant representing a non-inactivating persistent fraction (close to zero incontrol). To assess the concentrationeresponse relationships, data were fitted withthe Hill equation: y ¼ 100/[1 þ (EC50/[toxin])h], where y is the amplitude of thetoxin-induced effect, EC50 is the toxin concentration at half maximal efficacy [toxin],is the toxin concentration and h is the Hill coefficient. The time constants (t) of theNaV channel fast inactivation were measured directly from the decay phase of therecorded Naþ current using a single exponential fit. All data are presented asmean ± standard error (S.E.M) of at least 5 independent experiments (n� 5). All datawere tested for normality using a D'Agustino Pearson omnibus normality test. Alldata were tested for variance using Bonferroni test or Dunn's test. Data following aGaussian distribution were analyzed for significance using one-way ANOVA. Non-parametric data were analyzed for significance using the KruskaleWallis test. Dif-ferences were considered significant if the probability that their difference stemmedfrom chance was 55% (p < 0.05). All data was analyzed using pClamp Clampfit 10.0(Molecular Devices®, Downingtown, Pennsylvania, USA) and Origin 7.5 software(Originlab®, Northampton, Massachusetts, USA).
3. Results
Ts1 was investigated for its activity on 9 voltage-gated sodiumchannel isoforms, 8 mammalian subtypes (NaV1.1eNaV1.8), 1 insectNaV channel (DmNaV1) and 1 bacterial NaV channel (NaChBac).While Ts1 did not show any affinity for NaV1.1, NaV1.7, NaV1.8 orNaChBac channels (Supplemental Fig. 1), it was found that Ts1 ex-erts a striking differing pharmacological profile inwhich a selectivemodulation of NaV channel gating and kinetics occurred, dependingon the sensitive isoform it interacts with.
3.1. Ts1 acts as a classical b-toxin, modulating the activationprocess
At a testing concentration of 100 nM, Ts1 influenced the kineticsof activation for NaV1.2eNaV1.4 and NaV1.6 (Figs. 1 and 2). As can beseen in Table 1, Ts1 significantly shifted the midpoint of activationof these channels towards more negative potentials, causingchannels already to open at resting potentials compared to controlsituation. The strongest shift in V1/2 of activation was observed forNaV1.6 and NaV1.3 channels with a leftward 25 mV and 12 mV shift,respectively. For NaV1.4 and NaV1.6 channels, a strong reduction in
Fig. 1. Effect of Ts1 on mammalian NaV1.2 and NaV1.3 isoforms. Current traces were evokedincrements. Traces of a representative experiment are shown in control (left panel) and insteady-state inactivation (squares) curves in control (closed symbols) and in the presence o
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscoveon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
the sodium peak current of 50% and 20%, respectively, was observed(Fig. 2). Furthermore, for both channels, a significant leftward shiftof the steady-state inactivation curve was seen after application of100 nM Ts1. In contrast, no alterations of kinetics of inactivationwere observed for NaV1.2 and NaV1.3 channels.
3.2. Ts1 acts as an inhibitor of NaV1.5 channels
Ts1 inhibited the sodium peak current trough NaV1.5 channelwithout changes in the properties of gating (Fig. 3A and C). Nochanges in the activation or steady-state inactivation curves wereinduced by application of 100 nM toxin (Fig. 3B). To assess theconcentration dependence of the Ts1-induced inhibitory effects, adoseeresponse curve was constructed, in which the percentage ofcurrent inhibition was plotted as a function of the concentration(Fig. 3D). EC50 values yielded 55 ± 13 nM. To determine the bindingsite of Ts1, we investigated whether Ts1 binds to neurotoxin re-ceptor site 1. TTX, a well-known pore blocker of NaV's, was used forcompetitive binding experiments. Application of TTX at its half-maximal inhibitory concentration (IC50) resulted in a blockage ofhalf of the expressed NaV1.5 channels, as a 50% decrease of thesodium current peak amplitude was observed. Subsequent andadditional application of Ts1 at its EC50 did result in an additionalreduction of the peak amplitude (n ¼ 6) (Fig. 3E). Since it has beenestablished that voltage-clamp errors can occur in two-electrodeexperiments with heterologously expressed channels in oocytes,we verified whether or not IC50 concentration of TTX alter thevoltage-dependence of activation. The constructed activation andsteady-state inactivation curves showed no alteration of gatingkinetics of the channels when TTX was applied at the corre-sponding IC50 concentration (p > 0.05, n � 4, data not shown).
3.3. Ts1 acts as a bell-shaped voltage-dependent modulator ofinsect NaV channels
Using a test protocol, in which depolarizing pulses to 0 mV (i.e.Vmax, the test potential corresponding to maximal INa) were used,addition of 100 nM Ts1 resulted in (i) a little but significant decreaseof the peak sodium current (INa) by 13 ± 4% (n ¼ 11) (Fig. 4A).
by 100 ms depolarizations stepping from a holding potential of �90 to 10 mV in 5 mV100 nM Ts1 situation (middle panel). Right panel shows the activation (circles) andf 100 nM Ts1 (open symbols).
106107108109110111112113114115116117118119120121122123124125126127128129130
red electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027
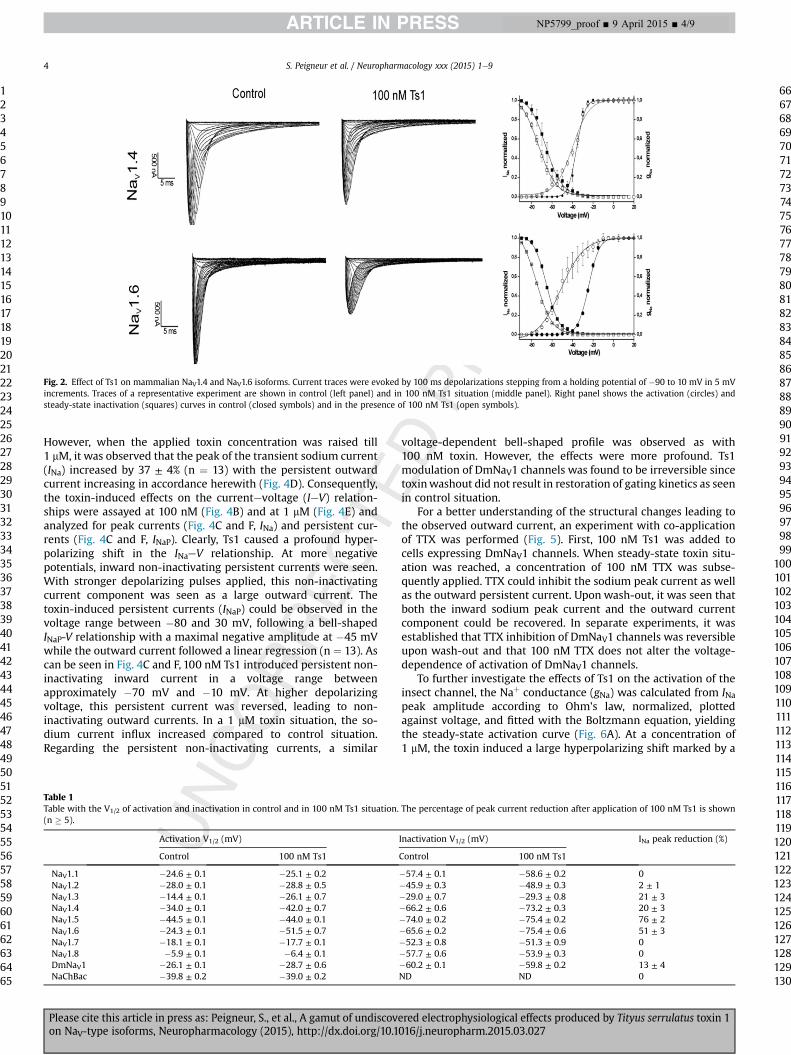
Fig. 2. Effect of Ts1 on mammalian NaV1.4 and NaV1.6 isoforms. Current traces were evoked by 100 ms depolarizations stepping from a holding potential of �90 to 10 mV in 5 mVincrements. Traces of a representative experiment are shown in control (left panel) and in 100 nM Ts1 situation (middle panel). Right panel shows the activation (circles) andsteady-state inactivation (squares) curves in control (closed symbols) and in the presence of 100 nM Ts1 (open symbols).
S. Peigneur et al. / Neuropharmacology xxx (2015) 1e94
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475767778798081828384858687888990919293949596979899
100101102103104105106107108109110111112113
NP5799_proof ■ 9 April 2015 ■ 4/9
However, when the applied toxin concentration was raised till1 mM, it was observed that the peak of the transient sodium current(INa) increased by 37 ± 4% (n ¼ 13) with the persistent outwardcurrent increasing in accordance herewith (Fig. 4D). Consequently,the toxin-induced effects on the currentevoltage (IeV) relation-ships were assayed at 100 nM (Fig. 4B) and at 1 mM (Fig. 4E) andanalyzed for peak currents (Fig. 4C and F, INa) and persistent cur-rents (Fig. 4C and F, INaP). Clearly, Ts1 caused a profound hyper-polarizing shift in the INaeV relationship. At more negativepotentials, inward non-inactivating persistent currents were seen.With stronger depolarizing pulses applied, this non-inactivatingcurrent component was seen as a large outward current. Thetoxin-induced persistent currents (INaP) could be observed in thevoltage range between �80 and 30 mV, following a bell-shapedINaP-V relationship with a maximal negative amplitude at �45 mVwhile the outward current followed a linear regression (n ¼ 13). Ascan be seen in Fig. 4C and F, 100 nM Ts1 introduced persistent non-inactivating inward current in a voltage range betweenapproximately �70 mV and �10 mV. At higher depolarizingvoltage, this persistent current was reversed, leading to non-inactivating outward currents. In a 1 mM toxin situation, the so-dium current influx increased compared to control situation.Regarding the persistent non-inactivating currents, a similar
Table 1Table with the V1/2 of activation and inactivation in control and in 100 nM Ts1 situation.(n � 5).
Activation V1/2 (mV)
Control 100 nM Ts1
NaV1.1 �24.6 ± 0.1 �25.1 ± 0.2NaV1.2 �28.0 ± 0.1 �28.8 ± 0.5NaV1.3 �14.4 ± 0.1 �26.1 ± 0.7NaV1.4 �34.0 ± 0.1 �42.0 ± 0.7NaV1.5 �44.5 ± 0.1 �44.0 ± 0.1NaV1.6 �24.3 ± 0.1 �51.5 ± 0.7NaV1.7 �18.1 ± 0.1 �17.7 ± 0.1NaV1.8 �5.9 ± 0.1 �6.4 ± 0.1DmNaV1 �26.1 ± 0.1 �28.7 ± 0.6NaChBac �39.8 ± 0.2 �39.0 ± 0.2
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscovon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
voltage-dependent bell-shaped profile was observed as with100 nM toxin. However, the effects were more profound. Ts1modulation of DmNaV1 channels was found to be irreversible sincetoxinwashout did not result in restoration of gating kinetics as seenin control situation.
For a better understanding of the structural changes leading tothe observed outward current, an experiment with co-applicationof TTX was performed (Fig. 5). First, 100 nM Ts1 was added tocells expressing DmNaV1 channels. When steady-state toxin situ-ation was reached, a concentration of 100 nM TTX was subse-quently applied. TTX could inhibit the sodium peak current as wellas the outward persistent current. Upon wash-out, it was seen thatboth the inward sodium peak current and the outward currentcomponent could be recovered. In separate experiments, it wasestablished that TTX inhibition of DmNaV1 channels was reversibleupon wash-out and that 100 nM TTX does not alter the voltage-dependence of activation of DmNaV1 channels.
To further investigate the effects of Ts1 on the activation of theinsect channel, the Naþ conductance (gNa) was calculated from INapeak amplitude according to Ohm's law, normalized, plottedagainst voltage, and fitted with the Boltzmann equation, yieldingthe steady-state activation curve (Fig. 6A). At a concentration of1 mM, the toxin induced a large hyperpolarizing shift marked by a
The percentage of peak current reduction after application of 100 nM Ts1 is shown
Inactivation V1/2 (mV) INa peak reduction (%)
Control 100 nM Ts1
�57.4 ± 0.1 �58.6 ± 0.2 0�45.9 ± 0.3 �48.9 ± 0.3 2 ± 1�29.0 ± 0.7 �29.3 ± 0.8 21 ± 3�66.2 ± 0.6 �73.2 ± 0.3 20 ± 3�74.0 ± 0.2 �75.4 ± 0.2 76 ± 2�65.6 ± 0.2 �75.4 ± 0.6 51 ± 3�52.3 ± 0.8 �51.3 ± 0.9 0�57.7 ± 0.6 �53.9 ± 0.3 0�60.2 ± 0.1 �59.8 ± 0.2 13 ± 4ND ND 0
114115116117118119120121122123124125126127128129130
ered electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027
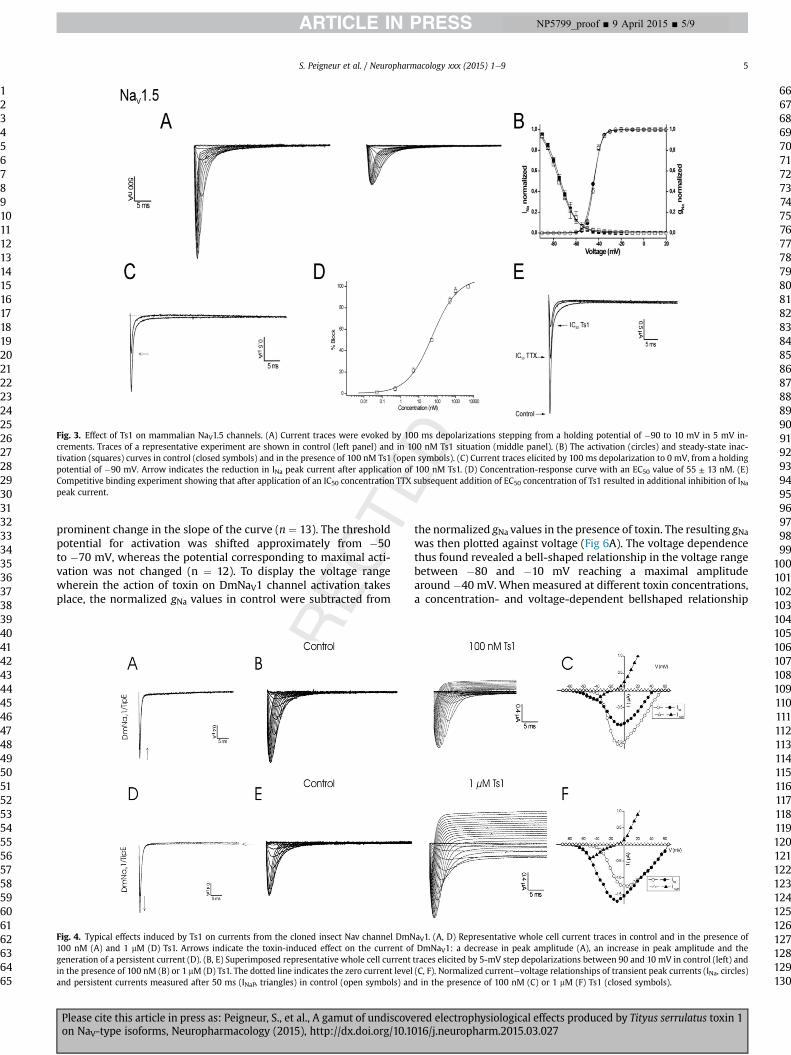
Fig. 3. Effect of Ts1 on mammalian NaV1.5 channels. (A) Current traces were evoked by 100 ms depolarizations stepping from a holding potential of �90 to 10 mV in 5 mV in-crements. Traces of a representative experiment are shown in control (left panel) and in 100 nM Ts1 situation (middle panel). (B) The activation (circles) and steady-state inac-tivation (squares) curves in control (closed symbols) and in the presence of 100 nM Ts1 (open symbols). (C) Current traces elicited by 100 ms depolarization to 0 mV, from a holdingpotential of �90 mV. Arrow indicates the reduction in INa peak current after application of 100 nM Ts1. (D) Concentration-response curve with an EC50 value of 55 ± 13 nM. (E)Competitive binding experiment showing that after application of an IC50 concentration TTX subsequent addition of EC50 concentration of Ts1 resulted in additional inhibition of INapeak current.
S. Peigneur et al. / Neuropharmacology xxx (2015) 1e9 5
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475767778798081828384858687888990919293949596979899
100101102103
NP5799_proof ■ 9 April 2015 ■ 5/9
prominent change in the slope of the curve (n ¼ 13). The thresholdpotential for activation was shifted approximately from �50to �70 mV, whereas the potential corresponding to maximal acti-vation was not changed (n ¼ 12). To display the voltage rangewherein the action of toxin on DmNaV1 channel activation takesplace, the normalized gNa values in control were subtracted from
Fig. 4. Typical effects induced by Ts1 on currents from the cloned insect Nav channel DmN100 nM (A) and 1 mM (D) Ts1. Arrows indicate the toxin-induced effect on the current ofgeneration of a persistent current (D). (B, E) Superimposed representative whole cell currentin the presence of 100 nM (B) or 1 mM (D) Ts1. The dotted line indicates the zero current leveland persistent currents measured after 50 ms (INaP, triangles) in control (open symbols) an
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscoveon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
the normalized gNa values in the presence of toxin. The resulting gNawas then plotted against voltage (Fig 6A). The voltage dependencethus found revealed a bell-shaped relationship in the voltage rangebetween �80 and �10 mV reaching a maximal amplitudearound �40 mV. When measured at different toxin concentrations,a concentration- and voltage-dependent bellshaped relationship
aV1. (A, D) Representative whole cell current traces in control and in the presence ofDmNaV1: a decrease in peak amplitude (A), an increase in peak amplitude and the
traces elicited by 5-mV step depolarizations between 90 and 10 mV in control (left) and(C, F). Normalized currentevoltage relationships of transient peak currents (INa, circles)d in the presence of 100 nM (C) or 1 mM (F) Ts1 (closed symbols).
104105106107108109110111112113114115116117118119120121122123124125126127128129130
red electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027
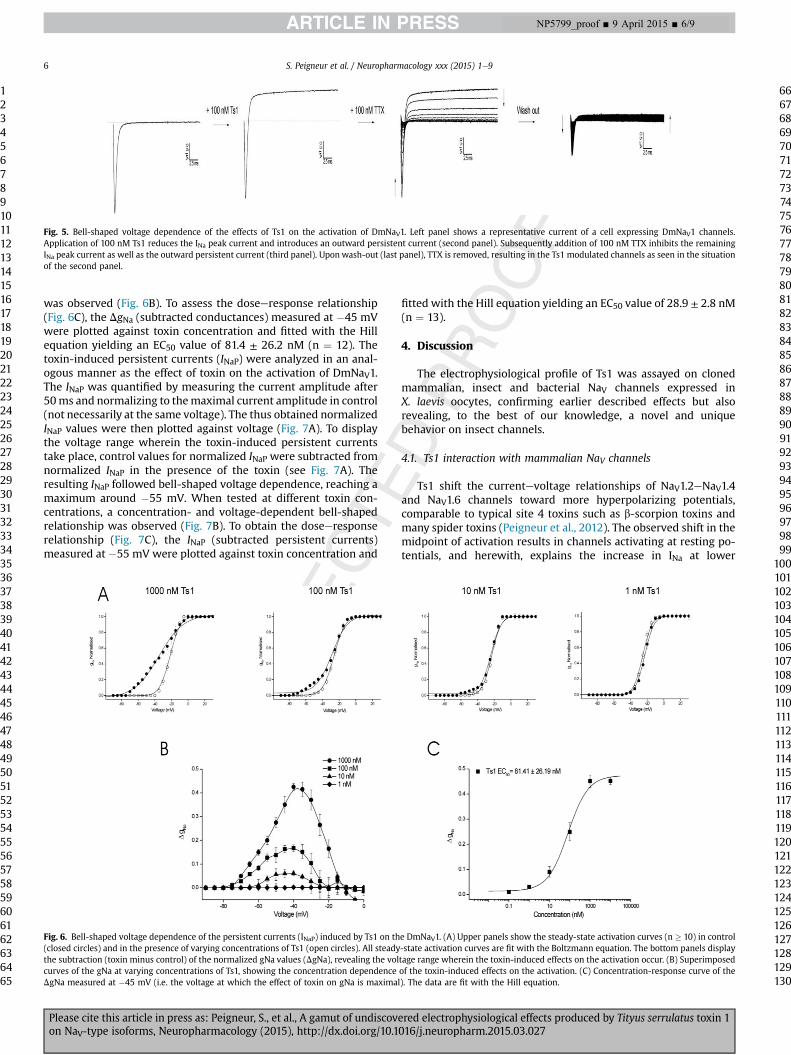
Fig. 5. Bell-shaped voltage dependence of the effects of Ts1 on the activation of DmNaV1. Left panel shows a representative current of a cell expressing DmNaV1 channels.Application of 100 nM Ts1 reduces the INa peak current and introduces an outward persistent current (second panel). Subsequently addition of 100 nM TTX inhibits the remainingINa peak current as well as the outward persistent current (third panel). Uponwash-out (last panel), TTX is removed, resulting in the Ts1 modulated channels as seen in the situationof the second panel.
S. Peigneur et al. / Neuropharmacology xxx (2015) 1e96
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475767778798081828384858687888990919293949596979899
NP5799_proof ■ 9 April 2015 ■ 6/9
was observed (Fig. 6B). To assess the doseeresponse relationship(Fig. 6C), the DgNa (subtracted conductances) measured at �45 mVwere plotted against toxin concentration and fitted with the Hillequation yielding an EC50 value of 81.4 ± 26.2 nM (n ¼ 12). Thetoxin-induced persistent currents (INaP) were analyzed in an anal-ogous manner as the effect of toxin on the activation of DmNaV1.The INaP was quantified by measuring the current amplitude after50ms and normalizing to themaximal current amplitude in control(not necessarily at the same voltage). The thus obtained normalizedINaP values were then plotted against voltage (Fig. 7A). To displaythe voltage range wherein the toxin-induced persistent currentstake place, control values for normalized INaP were subtracted fromnormalized INaP in the presence of the toxin (see Fig. 7A). Theresulting INaP followed bell-shaped voltage dependence, reaching amaximum around �55 mV. When tested at different toxin con-centrations, a concentration- and voltage-dependent bell-shapedrelationship was observed (Fig. 7B). To obtain the doseeresponserelationship (Fig. 7C), the INaP (subtracted persistent currents)measured at �55 mV were plotted against toxin concentration and
Fig. 6. Bell-shaped voltage dependence of the persistent currents (INaP) induced by Ts1 on th(closed circles) and in the presence of varying concentrations of Ts1 (open circles). All steadythe subtraction (toxin minus control) of the normalized gNa values (DgNa), revealing the volcurves of the gNa at varying concentrations of Ts1, showing the concentration dependenceDgNa measured at �45 mV (i.e. the voltage at which the effect of toxin on gNa is maximal
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscovon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
fitted with the Hill equation yielding an EC50 value of 28.9 ± 2.8 nM(n ¼ 13).
4. Discussion
The electrophysiological profile of Ts1 was assayed on clonedmammalian, insect and bacterial NaV channels expressed inX. laevis oocytes, confirming earlier described effects but alsorevealing, to the best of our knowledge, a novel and uniquebehavior on insect channels.
4.1. Ts1 interaction with mammalian NaV channels
Ts1 shift the currentevoltage relationships of NaV1.2eNaV1.4and NaV1.6 channels toward more hyperpolarizing potentials,comparable to typical site 4 toxins such as b-scorpion toxins andmany spider toxins (Peigneur et al., 2012). The observed shift in themidpoint of activation results in channels activating at resting po-tentials, and herewith, explains the increase in INa at lower
e DmNaV1. (A) Upper panels show the steady-state activation curves (n � 10) in control-state activation curves are fit with the Boltzmann equation. The bottom panels displaytage range wherein the toxin-induced effects on the activation occur. (B) Superimposedof the toxin-induced effects on the activation. (C) Concentration-response curve of the). The data are fit with the Hill equation.
100101102103104105106107108109110111112113114115116117118119120121122123124125126127128129130
ered electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027
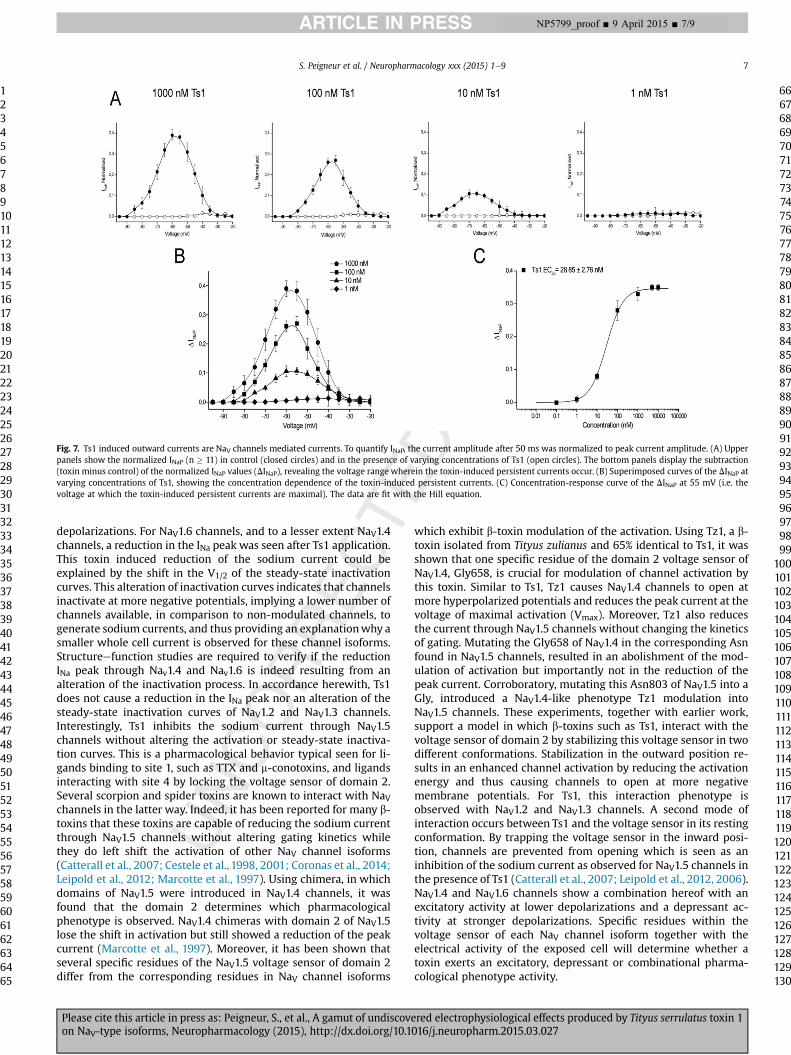
Fig. 7. Ts1 induced outward currents are NaV channels mediated currents. To quantify INaP, the current amplitude after 50 ms was normalized to peak current amplitude. (A) Upperpanels show the normalized INaP (n � 11) in control (closed circles) and in the presence of varying concentrations of Ts1 (open circles). The bottom panels display the subtraction(toxin minus control) of the normalized INaP values (DINaP), revealing the voltage range wherein the toxin-induced persistent currents occur. (B) Superimposed curves of the DINaP atvarying concentrations of Ts1, showing the concentration dependence of the toxin-induced persistent currents. (C) Concentration-response curve of the DINaP at 55 mV (i.e. thevoltage at which the toxin-induced persistent currents are maximal). The data are fit with the Hill equation.
S. Peigneur et al. / Neuropharmacology xxx (2015) 1e9 7
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475767778798081828384858687888990919293949596979899
100101102103104105106107108109110111112113114115116117118119120121122123124125126127128129130
NP5799_proof ■ 9 April 2015 ■ 7/9
depolarizations. For NaV1.6 channels, and to a lesser extent NaV1.4channels, a reduction in the INa peak was seen after Ts1 application.This toxin induced reduction of the sodium current could beexplained by the shift in the V1/2 of the steady-state inactivationcurves. This alteration of inactivation curves indicates that channelsinactivate at more negative potentials, implying a lower number ofchannels available, in comparison to non-modulated channels, togenerate sodium currents, and thus providing an explanationwhy asmaller whole cell current is observed for these channel isoforms.Structureefunction studies are required to verify if the reductionINa peak through Nav1.4 and Nav1.6 is indeed resulting from analteration of the inactivation process. In accordance herewith, Ts1does not cause a reduction in the INa peak nor an alteration of thesteady-state inactivation curves of NaV1.2 and NaV1.3 channels.Interestingly, Ts1 inhibits the sodium current through NaV1.5channels without altering the activation or steady-state inactiva-tion curves. This is a pharmacological behavior typical seen for li-gands binding to site 1, such as TTX and m-conotoxins, and ligandsinteracting with site 4 by locking the voltage sensor of domain 2.Several scorpion and spider toxins are known to interact with NaVchannels in the latter way. Indeed, it has been reported for many b-toxins that these toxins are capable of reducing the sodium currentthrough NaV1.5 channels without altering gating kinetics whilethey do left shift the activation of other NaV channel isoforms(Catterall et al., 2007; Cestele et al., 1998, 2001; Coronas et al., 2014;Leipold et al., 2012; Marcotte et al., 1997). Using chimera, in whichdomains of NaV1.5 were introduced in NaV1.4 channels, it wasfound that the domain 2 determines which pharmacologicalphenotype is observed. NaV1.4 chimeras with domain 2 of NaV1.5lose the shift in activation but still showed a reduction of the peakcurrent (Marcotte et al., 1997). Moreover, it has been shown thatseveral specific residues of the NaV1.5 voltage sensor of domain 2differ from the corresponding residues in NaV channel isoforms
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscoveon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
which exhibit b-toxin modulation of the activation. Using Tz1, a b-toxin isolated from Tityus zulianus and 65% identical to Ts1, it wasshown that one specific residue of the domain 2 voltage sensor ofNaV1.4, Gly658, is crucial for modulation of channel activation bythis toxin. Similar to Ts1, Tz1 causes NaV1.4 channels to open atmore hyperpolarized potentials and reduces the peak current at thevoltage of maximal activation (Vmax). Moreover, Tz1 also reducesthe current through NaV1.5 channels without changing the kineticsof gating. Mutating the Gly658 of NaV1.4 in the corresponding Asnfound in NaV1.5 channels, resulted in an abolishment of the mod-ulation of activation but importantly not in the reduction of thepeak current. Corroboratory, mutating this Asn803 of NaV1.5 into aGly, introduced a NaV1.4-like phenotype Tz1 modulation intoNaV1.5 channels. These experiments, together with earlier work,support a model in which b-toxins such as Ts1, interact with thevoltage sensor of domain 2 by stabilizing this voltage sensor in twodifferent conformations. Stabilization in the outward position re-sults in an enhanced channel activation by reducing the activationenergy and thus causing channels to open at more negativemembrane potentials. For Ts1, this interaction phenotype isobserved with NaV1.2 and NaV1.3 channels. A second mode ofinteraction occurs between Ts1 and the voltage sensor in its restingconformation. By trapping the voltage sensor in the inward posi-tion, channels are prevented from opening which is seen as aninhibition of the sodium current as observed for NaV1.5 channels inthe presence of Ts1 (Catterall et al., 2007; Leipold et al., 2012, 2006).NaV1.4 and NaV1.6 channels show a combination hereof with anexcitatory activity at lower depolarizations and a depressant ac-tivity at stronger depolarizations. Specific residues within thevoltage sensor of each NaV channel isoform together with theelectrical activity of the exposed cell will determine whether atoxin exerts an excitatory, depressant or combinational pharma-cological phenotype activity.
red electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027

S. Peigneur et al. / Neuropharmacology xxx (2015) 1e98
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475767778798081828384858687888990919293949596979899
100101102103104105106107108109110111112113114115116117118119120121122123124125126127128129130
NP5799_proof ■ 9 April 2015 ■ 8/9
4.2. Ts1 interaction with insect channels
Ts1 cause a 4-fold effect on the currents mediated by the insectDmNaV1 channel: (i) a decrease of peak sodium current at lowerconcentrations, (ii) an increase of INa peak, (iii) generation of a non-inactivating persistent current and (iv) a change in ion selectivity.Ts1 shifts the currentevoltage relationships of DmNaV1 channelstoward more hyperpolarizing potentials, comparable to what isseen on mammalian NaV channels. This shift in activation couldexplain the increase in peak current amplitude at low de-polarizations. Importantly, the foremost prominent difference inpharmacological activity compared to mammalian NaV channels isthe persistent current generated by Ts1 modulation of insectchannels. In fact, the increase in peak current amplitude and theappearance of persistent currents macroscopically resemble theeffects of typical site 3 toxins, such as a-scorpion toxins. Uponbinding at their binding site, which is mainly located at domain 4,site 3 toxins inhibit or slow down the fast inactivation of NaVchannels. However, the observed actions of Ts1 differ thoroughlyfrom the actions of typical site 3 toxins. The Ts1-induced persistentcurrents were only observed in the voltage range between�80 and0 mV, following bell-shaped voltage dependence with maximalamplitude around �45 mV. The interpretation of this unexpectedobservation is difficult because of the multitude effect of the toxin(the gamut of effects, cf. title). Outward persistent currents mightobscure inward persistent currents (if distinct) or perhaps thereversal potential of the persistent currents is close to zero becauseit is a mixed cation current. Besides the apparent slowing of theinactivation that could result in the persistent currents, Ts1 alsoinflicts a hyperpolarizing shift in the voltage dependence of acti-vation, causing the channels to open at lower depolarizationscompared with control conditions. Furthermore, at depolarizationsmore positive than�10mV, the persistent currents display a strongoutward component. This outward current could be inhibited byTTX, suggesting that this current is NaV channel mediated and thusthat a change in the ion selectivity of the sodium channel occurswithout altering the TTX-sensitivity.
A careful survey of the literature shows that several b-toxins, andmore specific insect selective b-toxins, induce similar effects as Ts1in regard to the persistent currents, however, up till now these ef-fects were not identified as bell-shaped voltage-dependent modu-lation of NaV channels. Using cockroach isolated axons, it was foundthatAaHIT fromAndroctonusaustralisHectorandBotIT2 fromButhusoccitanus Americana, besides shifting the activation towards hyper-polarizing potentials, also slow down the activation kinetics of NaVchannels, especially at more negative potentials (Borchani et al.,1996; De Lima et al., 1986). For IsomTx1, from Isometrus vittatus, itwas shown that the inactivation was incomplete and a maintainedcurrent persisted after toxin application, mainly at negative voltagepulses (Coronas et al., 2003). Although several studies report thedelay of inactivation at negative potentials, up till now and to thebest of our knowledge, the bell-shaped voltage-dependent modu-lation of insect NaV channels by b-scorpion toxins has not yet beeninvestigated in detail on heterologously expressed insect NaVchannels. However, for a certain family of spider toxins this peculiarpharmacological behavior has been described (Billen et al., 2010).The b/d-agatoxins are a group of 37 amino acid residues long pep-tides, isolated from the funnel web spider Agelena orientalis. Theyexert their insect toxicity by leftward shifting the midpoint of acti-vation and by inducing persistent non-inactivating sodium currentas is seen for Ts1. In contrast to Ts1, no outward component of thecurrent was seen at stronger depolarization. Furthermore, b/d-aga-toxins are insect selective since they did not interact withmammalian NaV channels, even not at higher concentrations. Uponobservation of the b/d-agatoxin modulation of NaV channels, the
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscovon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
authors argued that the classical concept of receptor sites 3 and 4 ofNaV channel binding toxins that exhibit the a-effect and b-effectbecomes rather “blurred” (Billen et al., 2010). Indeed, there is anincreasing awareness of cross-competitive binding of site 3- and 4-specific ligands (Bosmans et al., 2008; Liu et al., 2011).
It is interesting to note that Ts1 shares high sequence identitywith Ts2, a classical a-scorpion toxin, isolated from the samevenom (Cologna et al., 2012). Ts2 was found to be a potent site 3toxin, slowing down the inactivation of mammalian NaV channels.Indeed, it is well known that a- and b-scorpion toxins share a highdegree of identity. Moreover, the differences between a- and b-toxins sequences are not bigger than those foundwithin each group(Cologna et al., 2012). Furthermore, it has been proposed thatseveral b-scorpion toxins, such as AahIT4 from Androctonus aus-tralis Hector and Lqhb1 from Leiurus quinquestriatus hebraeus,represent a group of ancestral toxins from which further diversifi-cation permitted emergence of toxins with the structural featuresof b-toxins but displaying a-like activity without modulating theactivation. Also three toxins from Centuroides sculpturatus Ewing(CsEv1e3) and Ts2 are believed to belong to this group (Colognaet al., 2012; Possani et al., 1999). Taking in to account the highsequence homology between Ts1 and Ts2, together with theknowledge that a few very specific residues are determinative for apeptide to recognize with high affinity either site 3 or site 4, it canbe speculated that the bell-shaped voltage-dependent modulationof NaV channel inactivation by Ts1, and by extension other b-scor-pion toxins and the b/d-agatoxins, results from an unspecific orweak interaction with the voltage sensor of domain 4. This inter-actionwould be in agreement of the voltage sensor trapping modelwhich is applicable to many NaV channel modulating toxins(Cestele et al., 2001). Correspondingly, this would explain why theslowing down of inactivation is only seen in a bell-shaped fashionat more negative potentials. If Ts1 would bind to the voltage sensorof domain 4 with low affinity, than stronger depolarizations wouldprovide sufficient energy to remove Ts1 from its binding site,allowing restoration of the normal inactivation process. This hy-pothesis is supported with previous work showing that Ts1 indeedis capable of binding to domain 4 voltage sensors when this sensoris transferred into a KV2.1 channel (Bosmans et al., 2008). Moreover,earlier studies using voltage clamp fluorometry demonstrated thatTs1 impairs the movement of all 4 NaV channel voltage sensors(Campos et al., 2007).
In the last decade, an enormous number of studies have beenconducted in an effort to map the localization of the neurotoxinsites 3 and 4 and to provide a better understanding of the mech-anism of interaction of a-and b-scorpion toxins with these sites(Catterall, 2014; Ma et al., 2013; Song et al., 2011; Zhang et al.,2012). Consequently, there is a great knowledge of which regionor specific residues of the NaV channel contribute to each bindingsite. There is little to no argument regarding the understanding thata-scorpion toxins exert their toxicity by interactionwith site 3, as b-toxins do with site 4. However, we do strongly question the clas-sification of a-scorpion as toxins that exclusively modulate theinactivation process by binding to site 3 or the general accepteddefinition of b-scorpion toxins as toxins interfering with the acti-vation process through solely interacting with site 4. Accumulationof data available in the literature, together with the results shownhere for Ts1, clearly does show that several site 4 ligands, such as b-scorpion toxins and b/d-agatoxins do alter the inactivation processof NaV channels. Whether or not this modulation of the inactivationoccurs through toxin binding at site 4 or rather by interacting withanother binding site, such as site 3, remains the subject of furtherstructureefunction studies combining site directedmutagenesis onthe NaV channel with specific pointmutations within the Ts1 aminoacid sequence.
ered electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027

S. Peigneur et al. / Neuropharmacology xxx (2015) 1e9 9
1234567891011121314151617181920212223242526272829303132333435363738394041424344454647484950515253545556575859606162636465
66676869707172737475
NP5799_proof ■ 9 April 2015 ■ 9/9
5. Conclusion
In this work we have reported for the first time the full elec-trophysiological profile of Ts1 on NaV channels. In contrast to whatwas generally assumed from this classical b-scorpion toxin, Ts1 wasfound to exert a plethora of pharmacological effects on differentNaV channel isoforms. The observation that Ts1 modulates theinactivation of insect NaV channels in a bell-shaped voltage-dependent matter urges to a revision of the traditional classifica-tion of NaV channel toxins solely based on their site of interaction.
Q4
7677787980818283848586878889909192939495969798
Acknowledgments
The authors thank John N. Wood (University College London,London, UK) for sharing rNaV1.8; A. L. Goldin (University of Cali-fornia, Irvine, CA, USA) for sharing rNaV1.2, rNaV1.3, and mNaV1.6;G. Mandel (State University of New York, Stony Brook, NY, USA) forsharing rNaV1.4; R. G. Kallen (Roche Institute of Molecular Biology,Nutley, NJ, USA) for sharing hNaV1.5; S. H. Heinemann (Friedrich-Schiller-Universit€at Jena, Jena, Germany) for sharing the ratb1subunit; S. C. Cannon (University of Texas Southwestern MedicalCenter, Dallas, TX, USA) for sharing the hb1 subunit and Martin S.Williamson (Rothhamsted Research, Harpenden, UK) for providingthe Para and tipE clone. The NaV1.7 clone was kindly provided byRoche (Basel, Switzerland). We are grateful to Dr. Maria Elena deLima and Dr. Paulo Beir~ao for helpful suggestions. The authors aregrateful to Dr. Alexander Vassilevski for constructive discussions.This work was supported by F.W.O. Vlaanderen Grants G.0433.12,G.A071.10N, and G.0257.08; Inter-University Attraction Poles Pro-gramGrant IUAP 7/10 fromBelgian State, Belgian Science Policy; KULeuven Grant OT/12/081 and CNPq (Conselho Nacional de Desen-volvimento Científico e Tecnol�ogico) Grant 402508/2012-2.
99100101102103
Appendix A. Supplementary data
Supplementary data related to this article can be found at http://dx.doi.org/10.1016/j.neuropharm.2015.03.027.
104105106107108109110111112113114115116117118119120121122123124125126127
References
Billen, B., Vassilevski, A., Nikolsky, A., Debaveye, S., Tytgat, J., Grishin, E., 2010.Unique bell-shaped voltage-dependent modulation of Naþ channel gating bynovel insect-selective toxins from the spider Agelena orientalis. J. Biol. Chem.285, 18545e18554.
Borchani, L., Mansuelle, P., Stankiewicz, M., Grolleau, F., Cestele, S., Karoui, H.,Lapied, B., Rochat, H., Pelhate, M., El Ayeb, M., 1996. A new scorpion venomtoxin paralytic to insects that affects Naþ channel activation. Purification,structure, antigenicity and mode of action. Eur. J. Biochem. 241, 525e532.
Bosmans, F., Martin-Eauclaire, M.F., Swartz, K.J., 2008. Deconstructing voltagesensor function and pharmacology in sodium channels. Nature 456, 202e208.
Bosmans, F., Tytgat, J., 2007. Sea anemone venom as a source of insecticidal peptidesacting on voltage-gated Naþ channels. Toxicon 49, 550e560.
Campos, F.V., Chanda, B., Beirao, P.S., Bezanilla, F., 2007. beta-Scorpion toxin mod-ifies gating transitions in all four voltage sensors of the sodium channel. J. Gen.Physiol. 130, 257e268.
Catterall, W.A., 2000. From ionic currents to molecular mechanisms: the structureand function of voltage-gated sodium channels. Neuron 26, 13e25.
Catterall, W.A., 2012. Voltage-gated sodium channels at 60: structure, function andpathophysiology. J. Physiol. 590, 2577e2589.
Catterall, W.A., 2014. Structure and function of voltage-gated sodium channels atatomic resolution. Exp. Physiol. 99, 35e51.
Catterall, W.A., Cestele, S., Yarov-Yarovoy, V., Yu, F.H., Konoki, K., Scheuer, T., 2007.Voltage-gated ion channels and gating modifier toxins. Toxicon 49, 124e141.
Cerni, F.A., Pucca, M.B., Peigneur, S., Cremonez, C.M., Bordon, K.C., Tytgat, J.,Arantes, E.C., 2014. Electrophysiological characterization of Ts6 and Ts7, K(þ)channel toxins isolated through an improved Tityus serrulatus venom purifi-cation procedure. Toxins (Basel) 6, 892e913.
Please cite this article in press as: Peigneur, S., et al., A gamut of undiscoveon NaV-type isoforms, Neuropharmacology (2015), http://dx.doi.org/10.1
Cestele, S., Qu, Y., Rogers, J.C., Rochat, H., Scheuer, T., Catterall, W.A., 1998. Voltagesensor-trapping: enhanced activation of sodium channels by beta-scorpiontoxin bound to the S3-S4 loop in domain II. Neuron 21, 919e931.
Cestele, S., Scheuer, T., Mantegazza, M., Rochat, H., Catterall, W.A., 2001. Neutrali-zation of gating charges in domain II of the sodium channel alpha subunitenhances voltage-sensor trapping by a beta-scorpion toxin. J. Gen. Physiol. 118,291e302.
Cestele, S., Yarov-Yarovoy, V., Qu, Y., Sampieri, F., Scheuer, T., Catterall, W.A., 2006.Structure and function of the voltage sensor of sodium channels probed by abeta-scorpion toxin. J. Biol. Chem. 281, 21332e21344.
Chanda, B., Bezanilla, F., 2002. Tracking voltage-dependent conformational changesin skeletal muscle sodium channel during activation. J. Gen. Physiol. 120,629e645.
Cologna, C.T., Peigneur, S., Rustiguel, J.K., Nonato, M.C., Tytgat, J., Arantes, E.C., 2012.Investigation of the relationship between the structure and function of Ts2, aneurotoxin from Tityus serrulatus venom. FEBS J. 279, 1495e1504.
Coronas, F.I., Diego-Garcia, E., Restano-Cassulini, R., de Roodt, A.R., Possani, L.D.,2014. Biochemical and physiological characterization of a new Na-channelspecific peptide from the venom of the Argentinean scorpion Tityus triv-ittatus. Peptides.
Coronas, F.V., Stankiewicz, M., Batista, C.V., Giraud, S., Alam, J.M., Possani, L.D.,Mebs, D., Pelhate, M., 2003. Primary structure and electrophysiological char-acterization of two almost identical isoforms of toxin from Isometrus vittatus(family: Buthidae) scorpion venom. Toxicon 41, 989e997.
De Lima, M.E., Martin, M.F., Diniz, C.R., Rochat, H., 1986. Tityus serrulatus toxin VIIbears pharmacological properties of both beta-toxin and insect toxin fromscorpion venoms. Biochem. Biophys. Res. Commun. 139, 296e302.
Dong, K., 2007. Insect sodium channels and insecticide resistance. Invert. Neurosci.7, 17e30.
Gordon, D., Martin-Eauclaire, M.F., Cestele, S., Kopeyan, C., Carlier, E., Khalifa, R.B.,Pelhate, M., Rochat, H., 1996. Scorpion toxins affecting sodium current inacti-vation bind to distinct homologous receptor sites on rat brain and insect so-dium channels. J. Biol. Chem. 271, 8034e8045.
Hille, B., 2001. Ion Channels of Excitable Membranes. Sinauer Associates, Inc, Sun-derland, Massachusetts.
Hwang, D.F., Tai, K.P., Chueh, C.H., Lin, L.C., Jeng, S.S., 1991. Tetrodotoxin and de-rivatives in several species of the gastropod Naticidae. Toxicon 29, 1019e1024.
King, G.F., Escoubas, P., Nicholson, G.M., 2008. Peptide toxins that selectively targetinsect Na(V) and Ca(V) channels. Channels (Austin) 2, 100e116.
Leipold, E., Borges, A., Heinemann, S.H., 2012. Scorpion beta-toxin interference withNaV channel voltage sensor gives rise to excitatory and depressant modes.J. Gen. Physiol. 139, 305e319.
Leipold, E., Hansel, A., Borges, A., Heinemann, S.H., 2006. Subtype specificity ofscorpion beta-toxin Tz1 interaction with voltage-gated sodium channels isdetermined by the pore loop of domain 3. Mol. Pharmacol. 70, 340e347.
Liman, E.R., Tytgat, J., Hess, P., 1992. Subunit stoichiometry of a mammalian Kþchannel determined by construction of multimeric cDNAs. Neuron 9, 861e871.
Liu, Z.R., Ye, P., Ji, Y.H., 2011. Exploring the obscure profiles of pharmacologicalbinding sites on voltage-gated sodium channels by BmK neurotoxins. ProteinCell 2, 437e444.
Ma, Z., Kong, J., Gordon, D., Gurevitz, M., Kallen, R.G., 2013. Direct evidence thatscorpion alpha-toxins (site-3) modulate sodium channel inactivation by hin-drance of voltage-sensor movements. PLoS One 8, e77758.
Marcotte, P., Chen, L.Q., Kallen, R.G., Chahine, M., 1997. Effects of Tityus serrulatusscorpion toxin gamma on voltage-gated Naþ channels. Circ. Res. 80, 363e369.
Narahashi, T., 2008. Tetrodotoxin: a brief history. Proc. Jpn. Acad. Ser. B Phys. Biol.Sci. 84, 147e154.
Peigneur, S., Sevcik, C., Tytgat, J., Castillo, C., D'Suze, G., 2012. Subtype specificityinteraction of bactridines with mammalian, insect and bacterial sodium chan-nels under voltage clamp conditions. FEBS J. 279, 4025e4038.
Possani, L.D., Becerril, B., Delepierre, M., Tytgat, J., 1999. Scorpion toxins specific forNaþ-channels. Eur. J. Biochem. 264, 287e300.
Song, W., Du, Y., Liu, Z., Luo, N., Turkov, M., Gordon, D., Gurevitz, M., Goldin, A.L.,Dong, K., 2011. Substitutions in the domain III voltage-sensing module enhancethe sensitivity of an insect sodium channel to a scorpion beta-toxin. J. Biol.Chem. 286, 15781e15788.
Stephan, M.M., Potts, J.F., Agnew, W.S., 1994. The microI skeletal muscle sodiumchannel: mutation E403Q eliminates sensitivity to tetrodotoxin but not to mu-conotoxins GIIIA and GIIIB. J. Membr. Biol. 137, 1e8.
Stevens, M., Peigneur, S., Dyubankova, N., Lescrinier, E., Herdewijn, P., Tytgat, J.,2012. Design of bioactive peptides from naturally occurring mu-conotoxinstructures. J. Biol. Chem. 287, 31382e31392.
Vijverberg, H.P., Pauron, D., Lazdunski, M., 1984. The effect of Tityus serrulatusscorpion toxin gamma on Na channels in neuroblastoma cells. Pflugers Arch.401, 297e303.
Yu, F.H., Catterall, W.A., 2003. Overview of the voltage-gated sodium channel family.Genome Biol. 4, 207.
Zhang, J.Z., Yarov-Yarovoy, V., Scheuer, T., Karbat, I., Cohen, L., Gordon, D.,Gurevitz, M., Catterall, W.A., 2012. Mapping the interaction site for a beta-scorpion toxin in the pore module of domain III of voltage-gated Na(þ) chan-nels. J. Biol. Chem. 287, 30719e30728.
128129130
red electrophysiological effects produced by Tityus serrulatus toxin 1016/j.neuropharm.2015.03.027












