
A critical analysis of the biological impacts of plasticizers on wildlife
17
doi: 10.1098/rstb.2008.0242 , 2047-2062 364 2009 Phil. Trans. R. Soc. B R. Tyler Charles Kusk, Leah Wollenberger, Eduarda M. Santos, Gregory C. Paull, Katrien J. W. Van Look and Jörg Oehlmann, Ulrike Schulte-Oehlmann, Werner Kloas, Oana Jagnytsch, Ilka Lutz, Kresten O. wildlife A critical analysis of the biological impacts of plasticizers on References http://rstb.royalsocietypublishing.org/content/364/1526/2047.full.html#ref-list-1 This article cites 127 articles, 10 of which can be accessed free Subject collections (159 articles) environmental science (658 articles) ecology Articles on similar topics can be found in the following collections Email alerting service here right-hand corner of the article or click Receive free email alerts when new articles cite this article - sign up in the box at the top http://rstb.royalsocietypublishing.org/subscriptions go to: Phil. Trans. R. Soc. B To subscribe to This journal is © 2009 The Royal Society on 15 June 2009 rstb.royalsocietypublishing.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of A critical analysis of the biological impacts of plasticizers on wildlife

doi 101098rstb20080242 2047-2062364 2009 Phil Trans R Soc B
R Tyler
CharlesKusk Leah Wollenberger Eduarda M Santos Gregory C Paull Katrien J W Van Look and Joumlrg Oehlmann Ulrike Schulte-Oehlmann Werner Kloas Oana Jagnytsch Ilka Lutz Kresten O wildlifeA critical analysis of the biological impacts of plasticizers on
Referenceshttprstbroyalsocietypublishingorgcontent36415262047fullhtmlref-list-1
This article cites 127 articles 10 of which can be accessed free
Subject collections
(159 articles)environmental science (658 articles)ecology
Articles on similar topics can be found in the following collections
Email alerting service hereright-hand corner of the article or click
Receive free email alerts when new articles cite this article - sign up in the box at the top
httprstbroyalsocietypublishingorgsubscriptions go to Phil Trans R Soc BTo subscribe to
This journal is copy 2009 The Royal Society
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Phil Trans R Soc B (2009) 364 2047ndash2062
doi101098rstb20080242
Review
A critical analysis of the biological impactsof plasticizers on wildlife
Jorg Oehlmann1 Ulrike Schulte-Oehlmann1 Werner Kloas2
Oana Jagnytsch2 Ilka Lutz2 Kresten O Kusk3 Leah Wollenberger3dagger
Eduarda M Santos4 Gregory C Paull4 Katrien J W Van Look5
and Charles R Tyler4
AuthordaggerPresent88-92 1
One conand hum
1Department of Aquatic Ecotoxicology Goethe University Frankfurt am Main Siesmayerstr 7060323 Frankfurt am Main Germany
2Department of Inland Fisheries Leibniz-Institute of Freshwater Ecology and Inland FisheriesMuggelseedamm 310 12587 Berlin Germany
3Department of Environmental Engineering Technical University of Denmark Miljoevej 1132800 Lyngby Kgs Denmark
4School of Biosciences Hatherly Laboratories University of Exeter Prince of Wales RoadExeter EX4 4PS UK
5Institute of Zoology Zoological Society of London Regentrsquos Park London NW1 4RY UK
This review provides a critical analysis of the biological effects of the most widely used plasticizersincluding dibutyl phthalate diethylhexyl phthalate dimethyl phthalate butyl benzyl phthalate andbisphenol A (BPA) on wildlife with a focus on annelids (both aquatic and terrestrial) molluscscrustaceans insects fish and amphibians Moreover the paper provides novel data on the biologicaleffects of some of these plasticizers in invertebrates fish and amphibians Phthalates and BPA havebeen shown to affect reproduction in all studied animal groups to impair development in crus-taceans and amphibians and to induce genetic aberrations Molluscs crustaceans and amphibiansappear to be especially sensitive to these compounds and biological effects are observed at envir-onmentally relevant exposures in the low ng l21 to mg l21 range In contrast most effects in fish(except for disturbance in spermatogenesis) occur at higher concentrations Most plasticizersappear to act by interfering with the functioning of various hormone systems but some phthalateshave wider pathways of disruption Effect concentrations of plasticizers in laboratory experimentscoincide with measured environmental concentrations and thus there is a very real potential foreffects of these chemicals on some wildlife populations The most striking gaps in our currentknowledge on the impacts of plasticizers on wildlife are the lack of data for long-term exposuresto environmentally relevant concentrations and their ecotoxicity when part of complex mixturesFurthermore the hazard of plasticizers has been investigated in annelids molluscs and arthropodsonly and given the sensitivity of some invertebrates effects assessments are warranted in otherinvertebrate phyla
Keywords bisphenol A dibutyl phthalate diethylhexyl phthalate dimethyl phthalatebutyl benzyl phthalate endocrine disruption
1 INTRODUCTIONThe history of phthalate-based plasticizers and bisphe-nol A (BPA) dates back to the 1920s and 1930srespectively Phthalates have been applied as polyvinylchloride (PVC) additives since 1926 but were alsoused for healthcare purposes as insect repellents andcercaricides (Ferguson et al 1946) The developmentof BPA had a very different genesis driven primarily
for correspondence (oehlmannbiouni-frankfurtde)address Federal Institute for Risk Assessment Thielallee4195 Berlin Germany
tribution of 15 to a Theme Issue lsquoPlastics the environmentan healthrsquo
204
by the search for synthetic oestrogens for clinical use(Cook et al 1933) Dodds amp Lawson (1936 1938)demonstrated the oestrogenic activity of BPA in ovari-ectomized rats but the formulation of a more effectivesynthetic oestrogen diethylstilboestrol at around thesame time rendered BPA redundant as a clinicaloestrogen (Dodds et al 1938) In the 1950s BPA wasrediscovered when a Bayer chemist Hermann Schnellreacted BPA with phosgene to produce polycarbonateplastic and its use in plastics has subsequently becomeits primary commercial application
The emollient properties of phthalates andpolymerization of BPA have driven their widespreaduse in the production of plastics (Thompson et al
7 This journal is q 2009 The Royal Society
2048 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
2009ab) Today phthalates and BPA are found inmany mass-produced products including medicaldevices food packaging perfumes cosmetics chil-drenrsquos toys flooring materials computers CDs etc(Andrady amp Neal 2009) Phthalates may constitute upto 50 per cent of the total weight of PVC plastics andtheir worldwide annual production is approximately27 million metric tonnes (Bauer amp Herrmann 1997)For BPA worldwide annual production is 25 millionmetric tonnes (Staples et al 2002) Plasticizers workby reducing the chemical affinity between moleculeswhen embedded between chains of plastic raw materials(or act as monomers in polycarbonate plastic) but asplasticizers are not especially stable in these productsthey can leach out and thus end up in the environmentPhthalates are not classified as persistent compounds(Staples et al 1997) but their occurrence in theenvironment has been reported widely possibly arguingagainst a rapid biodegradation in some environments(Fatoki amp Vernon 1990 Bauer amp Herrmann 1997Heemken et al 2001 Fromme et al 2002Beauchesne et al 2007) Bisphenol A is easily degraded(Howard 1989) but nevertheless regularly detected inaquatic ecosystems owing to its continuous releaseinto the environment
The European Union Environmental Risk Assessment(ERA) reports provide predicted environmental con-centrations (PECs) for phthalates and BPA using theEuropean Union System for the Evaluation ofSubstances (EUSES) The reliability of some of thesePECs however is disputed as monitoring data from sur-face waters have found significantly higher environmentalconcentrations of phthalates While the PEC for dibutylphthalate (DBP) is 04 mg l21 and 0231 mg kg21 (drywt) for water and sediments respectively (EU 2004)Fatoki amp Vernon (1990) report a maximum measuredconcentration of 335 mg l21 in UK rivers The ERAdraft report calculates a PEC for diethylhexyl phthalate(DEHP) of 22 mg l21 for water and 34 mg kg21 (drywt) for sediments (EU 2006) This figure is exceededin some German surface waters where measured concen-trations range between 033 and 978 mg l21 (Frommeet al 2002) and in Japanese freshwaters and sedimentswhere peak concentrations have been measured at58 mg l21 and 210 000 mg kg21 (dry wt) respectively(average concentrations in Japanese sediments rangebetween 46 and 326 mg kg21 (dry wt Naito et al2006) For di-isodecyl phthalate (DIDP) the EUSES-based calculations state PECs of 18 mg l21 in surfacewaters and 32 mg kg21 (dry wt) in sediments (EU2003a) and for di-isononyl phthalate (DINP)07 mg l21 in surface waters and 18 mg kg21 (dry wt)in sediments (EU 2003b) There is a lack of dataon measured environmental concentrations of thesecompounds to confirm or refute these calculations
Bisphenol A is present in many environmentalcompartments including the aquatic environmentand enters water systems through point dischargessuch as landfill leachates and sewage treatment planteffluents Concentrations of BPA vary widely in theaquatic environment but they may even reach concen-trations up to 21 mg l21 in freshwater systems (Crainet al 2007) Concentrations of BPA in sediments aregenerally several orders of magnitude higher than
Phil Trans R Soc B (2009)
those in the water column For example in the RiverElbe in Germany BPA was measured at 0776 mg l21
in the water compared with 343 mg kg21 (dry wt) inthe sediment (Heemken et al 2001) These findingscontrast with a PEC of 012 mg l21 for water and16 mg kg21 (dry wt) for sediments as indicated bythe ERA report for BPA (EU 2003c)
Heightening our concern about possible healthimplications of exposure to plasticizers phthalates andBPA have been shown to bioaccumulate in organismsBioconcentration factors (BCFs) for plasticizers how-ever appear to differ widely between species Some ofthese differences relate to the type and nature of theplasticizers but others relate to the differences in theexperimental designs employed diet life and speciesrsquohistory and the tissues investigated Bioconcentrationfactors reported in invertebrates are generally higherthan those in vertebrates As an example for DEHPBCFs reported are between 42 and 842 in differentfish species but 3600 for the amphipod Gammaruspseudolimnaueus 2500 in the mussel Mytilus edulis andup to 5380 in the copepod Acartia tonsa (EU 2003d)
In this review paper we analyse critically theeffects on wildlife of the most prominent plasticizerswith a focus on effects in annelids (both aquatic andterrestrial) molluscs crustaceans insects fish andamphibians The paper further presents novel datafrom the authors on the biological effects of someplasticizers on animal species The paper concludeswith an analysis on the level of harm (and theuncertainties) associated with the discharge ofplasticizers into the environment and identifies keyresearch needs to address some of the uncertainties
2 MATERIAL AND METHODSIn this paper a series of experiments are reportedupon that provide novel data on the effects of butylbenzyl phthalate (BBP) on a fish (Danio rerio) andBPA on a crustacean (A tonsa) and an amphibian(Xenopus laevis) These experiments are describedbriefly as detailed information on study designs canbe obtained from the references cited
(a) Danio rerio (zebrafish)A study was conducted to investigate the effects ofBBP on reproduction in adult D rerio Fish were main-tained in breeding colonies of six males and sixfemales according to Santos et al (2007) The chemi-cal exposure tanks were set up in duplicate and dosedwith 0 (solvent controls with methanol 001permil) 3 6and 15 mg BBP l21 and run for 20 days The totalnumber of eggs produced and the number of viableembryos at 8 h post fertilization (hpf) were determinedaccording to Santos et al (2007) At the end of theexperiment quantification of vitellogenin (VTG) inthe plasma was performed according to Fenske et al(2001) with an adjustment of the primary VTG anti-body concentrations to 1 10 000 and a primaryantibody incubation time of 30 min One gonad fromall parent males was dissected and immediately usedfor sperm quantity and motility assessments (as measuresof sperm quality) using computer-assisted sperm
Review Effects of plasticizers on wildlife J Oehlmann et al 2049
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis (CASA v 7V3B Hobson Vision Ltd UK) ona Hobson sperm tracker (Hobson Vision Ltd) accord-ing to the protocols and settings described in Santoset al (2007)
Water samples were collected from each tank on days1 4 7 14 and 20 fixed with 5 methanol (FisherUK) extracted using Sep-Pack C18 cartridges(Waters Corporation USA) and BBP concentrationsof the extracted samples were determined by HPLCanalysis (Shimadzu USA) Samples were analysedthrough separation on a C18 column (SpherisorbODS2) using an isocratic gradient of 70 per centmethanol in water and the absorbance was monitoredat 277 nm on a UV HPLC detector (AppliedBiosystems UK) Quantification was achieved bycomparison of the absorbance of the BBP peak ineach sample with the absorbance of serially dilutedBBP standards (ranging from 00001 to 1 mg ml21)
The statistical comparisons on the VTG andfecundity data were performed using one-wayANOVA followed by Tukeyrsquos multiple comparison post-test analysis (using JANDEL Sigmastat 20 statisticalsoftware) The sperm data were arcsine transformedbefore a one-way ANOVA and Tukeyrsquos multiple compari-son post-test analysis using Excel (Microsoft Excel 2002)and GraphPad Prism (v 402 San Diego CA USA)
(b) Acartia tonsa (calanoid copepod Crustacea)Full life-cycle experiments were conducted with Atonsa starting with eggs and extending for three weeksto reproductively active adults with BPA following anOECD guideline proposal (Kusk amp Wollenberger2007) with minor modifications Animals were exposedin mass cultures (800 ml of mediumtest solution with400 copepods) Nominal BPA test concentrationswere 10 50 100 300 1000 mg l21 Semi-static expo-sures were performed at 20+058C with a photoperiodof 12 L 12 D cycle using a synthetic seawater mediumwith a salinity of 18permil (Kusk amp Wollenberger 1999) Onday 14 the number of eggs produced within a 24 hperiod was determined in isolated groups of 5 females(5 replicates per concentration 10 control groups)and expressed as eggs per female Eggs produced onday 18 were transferred to a clean medium (6 replicatesper concentration each containing 30 eggs) in order toobserve the hatching rate and larval mortality of thesecond generation determined on day 21 The larvaldevelopment ratio (LDR) describing the proportionof animals completing metamorphosis from thenauplius to the copepodite morphology was recordedon day 5 The LDR test (Wollenberger et al 2005)was conducted in parallel to the exposure of the massculture under the same exposure conditions with nom-inal BPA concentrations of 30 60 125 250 5001000 mg l21 with 4 replicates per concentration eachcontaining 30 copepods and 12 control groups
(c) Xenopus laevis (African clawed frog)The impacts of BPA on the thyroid system of X laeviswere determined as described previously for tetrabro-mobisphenol A by Jagnytsch et al (2006) Brieflythe levels of thyroid receptor b (TRb) mRNA weremeasured in the brain as biomarker for the
Phil Trans R Soc B (2009)
bioavailability of thyroid hormones (THs) (Opitzet al 2006) Bisphenol A was applied at concentrationsranging between 25 and 500 mg l21 (25 25 250500 mg l21) with 2 replicates per concentration and20 tadpoles per replicate in a long-term Xenopusmetamorphosis assay (XEMA 21 d) and at BPA concen-trations of 100 250 and 500 mg l21 with and without 01and 10 nM tri-iodothyronine (T3) in short-termexposures for 1 2 and 3 days (starting with duplicatetanks each containing 30 tadpolesaquarium andsubsampling 10 individuals in each replicate everyday) The effects of exposure to BPA during larvaldevelopment on sexual differentiation were also inves-tigated using a flow-through system to the point ofcompletion of metamorphosis (see Lutz et al 2008for details of protocol) Briefly tadpoles were exposedto BPA at concentrations of 1029 1028 1027 and1026 M (228 ng l21 to 228 mg l21) and to dilutionwater without BPA (negative control) A positiveoestrogen control was also included (02 mg 17b-oestradiol l21) In this study there were 4 replicatesfor all treatments and 25 individual tadpoles wereexposed in each tank (n frac14 100 per treatment) Theendpoints investigated were survival rate time to com-pletion of metamorphosis weight sex (based on grossmorphology) and histology of testes In addition geneexpression for hepatic insulin-like growth factor-I(IGF-I) and VTG mRNA was analysed in male Xlaevis immediately at completion of metamorphosisusing semi-quantitative RTndashPCR as describedpreviously (Kloas et al 1999)
3 RESULTS AND DISCUSSION(a) Effects of phthalates
Assessing the biological effects of phthalates such asDEHP in aquatic environments presents a significantchallenge owing to its low water solubility and itstendency to form colloidal dispersions above 3 mg l21Many experiments exposing DEHP via the water atconcentrations 3 mg l21 have nevertheless clearlyestablished concentration-related biological effects(Forget-Leray et al 2005) Despite this risk assess-ments of DEHP in Japan (Naito et al 2006) andthe EU (EU 2003d) have disregarded most studiesadopting concentrations 3 mg l21 and the Japaneseassessment has set the threshold effect level (PNEC)to 3 mg DEHP l21 (Naito et al 2006) This omissionneeds addressing given the effects reported for thisphthalate in this review Working with other highlylipophilic phthalates presents similar solubility prob-lems as for DEHP Di-isononyl phthalate and DIDPhave a log Kow of 88 and a water solubility of only02 mg l21 (EU 2003ab) which makes them evenmore difficult to handle in aquatic media than DEHP
(i) Annelids and molluscsDespite their ecological importance annelids andmolluscs have received relatively little attention inassessing the effects of plasticizers compared withvertebrates or other invertebrates (especially crus-taceans and insects Weltje amp Schulte-Oehlmann2007) Early investigations on annelids and molluscs
2050 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
focused on bioaccumulation and acute toxicity ofphthalates but more recently wider biological effectshave been shown including mitotic inhibition induc-tion of chromosomal aberrations and effects on larvaldevelopment
Considering soil-dwelling organisms studies on theuptake and toxicity of the phthalates dimethyl phthalate(DMP) diethyl phthalate (DEP) DBP and DEHP (ata concentration of 5 mg kg21 for 25 days) in the wormEisenia fetida have shown that high molecular weightphthalates (DBP and DEHP) were incorporated butwith relatively low accumulation factors (0242ndash0307and 0073ndash0244 respectively) and DMP and DEPwere apparently not taken up (Xiao-yu et al 2005) Itis possible that the lack of any uptake of DMP andDEP may be as a consequence of their higher microbialdegradation rates and a faster metabolic degradation(Albro et al 1993) Neuhauser et al (1986) investigatedthe acute toxicity of phthalates and found similar LC50
values (1064ndash3335 mg kg21 for a 14 day soil exposure)for DMP in four soil-dwelling annelid species(Allolobophora tuberculata E fetida Eudrilus eugeniaePerionyx excavatus) Together these studies demonstratethat phthalates enter terrestrial worms but they have alow acute toxicity It is worth emphasizing that soils canreceive very high inputs of phthalates and the totalamount of DEHP deposited in US landfills to 1992has been estimated at 5 million metric tonnes
Comparisons of the acute toxicity of severalphthalate esters [DMP DEP DBP BBP di-n-hexyl-phthalate (DHP) and DEHP] in various aquatic organ-isms including the annelid Lumbriculus variegatus haveshown that the higher molecular weight phthalatesDHP and DEHP were not acutely toxic for the concen-tration range tested (23ndash478 mg DHP l21 102ndash691 mg DEHP l21 Call et al 2001) The toxicity oflower molecular weight phthalates was positively corre-lated with their Kow indicating non-polar narcosis asthe underlying mode of action The same study foundL variegatus was the species least sensitive to DMPthe 10 day LC50 (10 day) was 246 compared with281 and 682 mg l21 for the crustacean Hyalellaazteca and the insect Chironomus tentans respectivelyIn adult marine tubeworms Pomatoceros lamarckiiexposure to DMP was shown to decrease fertilizationsuccess at a threshold concentration of 11025 MDMP (194 mg l21) and induce a significant increasein the number of aberrations in chromosome separ-ations in oocytes at anaphase at concentrations11027 M (194 mg l21) (Dixon et al 1999 Wilsonet al 2002) DMP has been shown to affect larvaldevelopment in the polychaete worm at a thresholdconcentration 1 1025 M (194 mg l21)
The effects of phthalates have also been studied onvarious biochemical pathways such as antioxidant andperoxisomal marker enzymes in marine mussels(Mytilus galloprovincialis Orbea et al 2002 M edulisCajaraville amp Ortiz-Zarragoitia 2006) Exposure to500 mg DEHP l21 for 21 days was shown to result ina significant increase in the catalase and acyl-CoAoxidase (AOX) activity and an inhibition of the super-oxide and manganate superoxide dismutase inM galloprovincialis The peroxisomal volume densityin the digestive gland of M edulis was significantly
Phil Trans R Soc B (2009)
enhanced at 50 mg diallylphthalate (DAP) per litreafter three weeks exposure whereas the chemicalhad no significant effect on AOX activity in thisspecies (Cajaraville amp Ortiz-Zarragoitia 2006) DAPwas able to decrease the phospho-protein level (amussel VTG-like protein) but did not impair ovarianfollicles oocytes and spermatogenesis in M eduliswhen exposed to 50 mg DAP l21 for three weeks(Aarab et al 2006) In another 21 day study DAPwas also shown to increase micronuclei frequency amarker of genotoxicity and fragmented-apoptoticcells in the gills of M edulis at 50 mg DAP l21
(Barsiene et al 2006)Very few studies have considered the effects of
phthalates on annelid or mollusc populations butTagatz et al (1986) found that exposure to DBPaltered the community structure and colonizationprofile of the zoomacrobenthos under laboratory andfield conditions The densities of molluscs andechinoderms were reduced by 49 and 97 per centwhile no effects were seen in annelids arthropodschordates and other groups (at concentrations of10ndash1000 mg kg21 over eight weeks)
(ii) Crustaceans and insectsLower molecular weight phthalates that are relativelywater soluble (Howard et al 1985) have been shownto have a lower acute and chronic toxicity in Daphniamagna For example 48 h immobilization tests withD magna have shown IC50 values of 284 220 and678 mg l21 for DMP DEP and DBP respectively(Jonsson amp Baun 2003) Acute toxicity for phthalateshowever differs significantly for different species ofcrustaceans Hyalella azteca is 10ndash20-fold more sensi-tive than D magna although the higher molecularweight phthalates DHP and DEHP did not exhibittoxic effects in H azteca even at concentrations up to225 mg DHP l21 and 590 mg DEHP l21 (Call et al2001) Other effects of DBP reported in crustaceansinclude decreased locomotor activity in Gammaruspulex at an exposure concentration of 500 mg l21
(Thuren amp Woin 1991)In the Drosophila melanogaster BII cell line DEP
exhibited a weak ecdysteroid antagonistic activity(Dinan et al 2001) However the concentrationsrequired to cause an effect were several orders ofmagnitude higher than that for 20-hydroxy ecdysonethus classifying DEP as a very weak environmentalanti-ecdysteroid only
DEHP has been shown to induce sublethal effectsin Chironomus riparius and the exposure of larvaeresulted in an increased female body volume atexposure concentrations of 03 mg l21 (Kwak amp Lee2005) It also increased the expression of heat shockprotein (HSP) and haemoglobin genes in exposedlarvae of C tentans at a concentration of 05 mg l21
(Lee et al 2006a) but the same compound was notacutely toxic to this species at concentrations up to691 mg DEHP l21 (Call et al 2001) The significanceof such effects on gene expression is not known but invertebrates HSPs are involved in steroid hormonereceptor stabilization and activation (Gillesby ampZacharewski 1998) and if they act similarly in
Review Effects of plasticizers on wildlife J Oehlmann et al 2051
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
arthropods a change in the HSP70 titre might resultin a disruption of the ecdysone axis by influencingthe ecdysone receptor
(iii) FishGiven the widespread occurrence of phthalates in theaquatic environment fish are also likely to be exposedto phthalates via the water column food andor viathe sediments depending on their ecological nicheIn support of this a study in wild fish inThe Netherlands measured median concentrations ofDEHP and DBP at 17 and 10 mg kg21 (wet wt)respectively with 95th percentile values reaching upto 141 mg DEHP kg21 and 26 mg DBP kg21 (wet wt)(Peijnenburg amp Struijs 2006) Laboratory studies havefurther shown that phthalates bioconcentrate in fishFor example the BCF for DEHP was 120 in the carpand for BBP 94 in bluegill sunfish (Staples et al1997) The presence of phthalates in wild fish andtheir ability to concentrate in body tissues demonstratetheir potential to induce adverse effects on the health ofwild populations but there has been little bearing onthis expectation
Very few studies have comprehensively addressedthe biological effects of phthalates on fish and invivo studies have reported mixed findings BBP hasbeen reported to be oestrogenic and induces VTG syn-thesis in male fathead minnow (Pimephales promelas) ata concentration of 100 mg l21 via the water (Harrieset al 2000) and similarly to induce VTG in carp(Cyprinus carpio) exposed to 1ndash20 mg DEP l21 overa period of four weeks (Barse et al 2007) In salmon(Salmo salar) exposure to 1500 mg DEHP kg21 inthe food for a prolonged period (four months)during early life resulted in a small incidence of inter-sex (Norman et al 2007) These in vivo effects areconsistent with the mode of action of BBP andDEHP established in vitro where they have beenshown to bind to the oestrogen receptor (ER) andelicit a weakly oestrogenic response in fish cell cultures(Jobling et al 1995) and in the yeast oestrogen screen(YES containing the human ER) (Harris et al 1997)In female Japanese medaka (Oryzias latipes) exposureto concentrations in excess of 1 mg DEHP l21 caused amarked reduction in both VTG and in the percentageof females containing mature oocytes suggesting ananti-oestrogenic mode of action (Kim et al 2002)This anti-oestrogenic effect presumably arises throughcompetition with the endogenous oestrogens for thereceptor In vitro receptor interaction studies havealso shown that BBP and DEHP possess anti-androgenic activity (Sohoni amp Sumpter 1998) Otherin vivo effects of phthalates in fish include effects onthe metabolic pathways involved in steroid biosyn-thesis and metabolism In a study by Thibaut ampPorte (2004) DEHP and DBP altered the activity ofenzymes involved in the synthesis of endogenoussteroid hormones and their metabolism in carp effectsthat can lead to the alteration of the balance betweenendogenous oestrogens and androgens and thusalterations in their reproductive physiology Thesefindings are supported by studies in mammals inwhich phthalates have also been shown to alter the
Phil Trans R Soc B (2009)
expression and enzymatic activity of enzymes involvedin the synthesis and metabolism of sex steroids in thetestis (Liu et al 2005) In fish exposures to lowmg l21 concentrations of DEP have been shown toinduce alterations in a number of enzymes in theliver and muscle including phosphatases and trans-aminases showing a disruptive effect on generalmetabolic functions (Barse et al 2007) Other studieshave also reported that potentially the most relevantmechanism of action of phthalates is via binding tothe peroxisome proliferator-activated receptors(PPARs) and modulation of the transcriptionalevents coordinated by this molecular pathway whichincludes steroid hormone metabolism (Fan et al2004)
Exposures to phthalates have also been shown toalter behaviour in fish Exposure to 100 mg BBP l21via the water caused alterations in shoaling and feedingbehaviour in three-spined stickleback (Gasterosteusaculeatus) (Wibe et al 2002 2004) and exposure to5 mg DEP l21 caused alterations in the general behaviourof common carp (Barse et al 2007) It should be notedthat these studies have employed very high exposureconcentrations and these are unlikely to occur in thewater column in part owing to their low solubilityExceptions to this may be for fish living incloselyassociated with the sediments in heavily contaminatedenvironments Exposures of fish to lower levels ofphthalates have generally found no adverse effects Asan example exposure of Japanese medaka to 219 and193 mg g21 DINP and DIDP respectively via thediet with a daily feeding regime of 5 per cent of thebody weight failed to identify any effects on reproduc-tion (gonad somatic index egg production embryosurvival and sex ratios) growth or survival in amulti-generational study (14 days post fertilization(dpf) to 140 dpf of the F1 generation) (Patyna et al2006) Similarly exposure of medaka to concentrationsup to 5 mg DEHP l21 for 90 days post-hatch did notaffect germ cell development (Metcalfe et al 2001)
In our exposure of adult breeding zebrafish to BBPmeasured concentrations in the tank water were 3 6and 15 mg l21 over the exposure period Exposure toBBP did not affect the concentration of VTG in theplasma at any of the concentrations tested in males(VTG concentrations in the plasma were 1310+2551500+358 948+141 1300+318 1640+220 ng ml21 for the water control solvent control 36 and 15 mg l21 BBP respectively n frac14 9ndash12 malesper treatment group) Similarly there was no evidenceof any chemical effect on the number of eggs spawnedthe total number of eggs spawned per colony over theassessment period ranged between 4109 and 7578with a maximum of 727 eggs spawned by a singlecolony on a single day with a mean number of eggsspawned per female per day (including days with noeggs) of 222+14 213+14 184+08 216+17 285+14 prior to exposure and 195+16270+17 211+12 250+21 325+14 duringthe exposure for the water control solvent control 36 and 15 mg BBP l21 respectively Furthermore therewere no effects of BBP on the mean percentage viabilityof embryos at 8 hpf (figure 1) These results are inagreement with reports in the literature where VTG
time (days)ndash20 ndash10 0 10 20
cum
ulat
ive
num
ber
of v
iabl
e eg
gs (
8 hp
f)
0
100
200
300
400
500
600
700
Figure 1 Danio rerio Cumulative number of viable eggs spawned in colonies during a 20 day period prior to BBP exposure and
a 20 day period during BBP exposure Symbols represent average results for two colonies per treatment filled circle watercontrol open circle solvent control inverted filled triangle 0003 mg l21 BBP open triangle 0006 mg l21 BBP filledsquare 0015 mg l21 BBP
2052 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
induction in males or reproductive effects on femaleshas been induced only at exposure concentrations thatfar exceed the measured concentrations in our studyIn contrast however we found consistent changes inparameters of sperm quality in BBP-exposed male zeb-rafish The solvent adopted (methanol) had a negativeeffect on zebrafish sperm motility compared with thewater control after 20 days of exposure (mean curvi-linear velocity of 14 mm s21 (mean) and 25 mm s21respectively p 005 figure 2a) and BBP exposureresulted in a concentration-dependent enhancementin curvilinear velocity of the solvent-affected spermAt the highest concentration tested (15 mg BBP l21)the average curvilinear velocity of the sperm wasrestored to levels similar to those found in the watercontrol tanks (31 mm s21 figure 2a) Both thenumber of sperm tracked and the linearity of thesperm movement showed patterns similar to thosedescribed for the curvilinear velocity (figure 2bc)
The ability of BBP to reverse the adverse effect ofthe methanol on sperm number and motility is anovel finding and we hypothesize that this may haveresulted from an interaction of BBP with the PPARreceptor which leads to increased oxidative stress inthe testis as demonstrated in mammals for phthalates(reviewed in Corton amp Lapinskas 2005) In the finalstages of sperm development maturation and acti-vation are crucially dependent on the oxidative balancewithin the seminiferous tubes and in the sperm itselfExposure to methanol may have reduced the freeoxygen radicals required to promote sperm maturationand activation and this effect may have been reversedby the increase in free radicals caused by the BBPexposure The ethanol metabolite acetaldehyde hasbeen shown to block the action of PPARs in vitro(Galli et al 2001) suggesting a mechanism by whichsperm motility might be reduced in fish exposed to asolvent Exposure to BBP in the solvent carrier maycounterbalance this effect restoring the normal
Phil Trans R Soc B (2009)
motility of the sperm The increase in oxidative stresswithin the testis as a result of phthalate exposure hasbeen previously observed in mammalian systemsincluding in rats (Kasahara et al 2002) and thiseffect was shown to be mediated through activationof PPAR pathways
(iv) AmphibiansStudies on phthalates in amphibians (reviewed inKloas 2002) are less extensive than those in fish andsome invertebrates The effects of plasticizers in amp-hibians have been focused on their toxicity theirpotential to induce teratogenic effects and their endo-crine-disrupting activities most notably effects onreproduction sexual differentiation and the thyroidsystem Focusing on toxicity Larsson amp Thuren(1987) found that exposure of Rana arvalis eggs toDEHP via sediment decreased successful hatchingswith increasing concentrations and deduced a noobserved effect concentration (NOEC) of 10 mg kg21
fresh weight Studies with DBP using the frog embryoteratogenesis assay Xenopus (FETAX) have showndevelopmental malformations at 500 mg l21 andenhanced mortality rates at concentrations of 10 and15 mg l21 (Lee et al 2005)
DBP has also been shown to affect sexual differen-tiation in male Rana rugosa tadpoles delaying gonadaldevelopment (Ohtani et al 2000) and altering thespermatogenesis process in eight-month-old X laevisafter exposure during larval development (at 1ndash10 mg DBP l21 from stage 52 to completion of meta-morphosis Lee amp Veeramachaneni 2005) Studies inXenopus investigating the potential for phthalates tobind to the ER via competitive displacement exper-iments with cytosolic liver preparations have foundthat DEP has only a moderate affinity for the ER(Lutz amp Kloas 1999) Parallel in vitro studies using Xlaevis primary hepatocytes however have shown that
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Phil Trans R Soc B (2009) 364 2047ndash2062
doi101098rstb20080242
Review
A critical analysis of the biological impactsof plasticizers on wildlife
Jorg Oehlmann1 Ulrike Schulte-Oehlmann1 Werner Kloas2
Oana Jagnytsch2 Ilka Lutz2 Kresten O Kusk3 Leah Wollenberger3dagger
Eduarda M Santos4 Gregory C Paull4 Katrien J W Van Look5
and Charles R Tyler4
AuthordaggerPresent88-92 1
One conand hum
1Department of Aquatic Ecotoxicology Goethe University Frankfurt am Main Siesmayerstr 7060323 Frankfurt am Main Germany
2Department of Inland Fisheries Leibniz-Institute of Freshwater Ecology and Inland FisheriesMuggelseedamm 310 12587 Berlin Germany
3Department of Environmental Engineering Technical University of Denmark Miljoevej 1132800 Lyngby Kgs Denmark
4School of Biosciences Hatherly Laboratories University of Exeter Prince of Wales RoadExeter EX4 4PS UK
5Institute of Zoology Zoological Society of London Regentrsquos Park London NW1 4RY UK
This review provides a critical analysis of the biological effects of the most widely used plasticizersincluding dibutyl phthalate diethylhexyl phthalate dimethyl phthalate butyl benzyl phthalate andbisphenol A (BPA) on wildlife with a focus on annelids (both aquatic and terrestrial) molluscscrustaceans insects fish and amphibians Moreover the paper provides novel data on the biologicaleffects of some of these plasticizers in invertebrates fish and amphibians Phthalates and BPA havebeen shown to affect reproduction in all studied animal groups to impair development in crus-taceans and amphibians and to induce genetic aberrations Molluscs crustaceans and amphibiansappear to be especially sensitive to these compounds and biological effects are observed at envir-onmentally relevant exposures in the low ng l21 to mg l21 range In contrast most effects in fish(except for disturbance in spermatogenesis) occur at higher concentrations Most plasticizersappear to act by interfering with the functioning of various hormone systems but some phthalateshave wider pathways of disruption Effect concentrations of plasticizers in laboratory experimentscoincide with measured environmental concentrations and thus there is a very real potential foreffects of these chemicals on some wildlife populations The most striking gaps in our currentknowledge on the impacts of plasticizers on wildlife are the lack of data for long-term exposuresto environmentally relevant concentrations and their ecotoxicity when part of complex mixturesFurthermore the hazard of plasticizers has been investigated in annelids molluscs and arthropodsonly and given the sensitivity of some invertebrates effects assessments are warranted in otherinvertebrate phyla
Keywords bisphenol A dibutyl phthalate diethylhexyl phthalate dimethyl phthalatebutyl benzyl phthalate endocrine disruption
1 INTRODUCTIONThe history of phthalate-based plasticizers and bisphe-nol A (BPA) dates back to the 1920s and 1930srespectively Phthalates have been applied as polyvinylchloride (PVC) additives since 1926 but were alsoused for healthcare purposes as insect repellents andcercaricides (Ferguson et al 1946) The developmentof BPA had a very different genesis driven primarily
for correspondence (oehlmannbiouni-frankfurtde)address Federal Institute for Risk Assessment Thielallee4195 Berlin Germany
tribution of 15 to a Theme Issue lsquoPlastics the environmentan healthrsquo
204
by the search for synthetic oestrogens for clinical use(Cook et al 1933) Dodds amp Lawson (1936 1938)demonstrated the oestrogenic activity of BPA in ovari-ectomized rats but the formulation of a more effectivesynthetic oestrogen diethylstilboestrol at around thesame time rendered BPA redundant as a clinicaloestrogen (Dodds et al 1938) In the 1950s BPA wasrediscovered when a Bayer chemist Hermann Schnellreacted BPA with phosgene to produce polycarbonateplastic and its use in plastics has subsequently becomeits primary commercial application
The emollient properties of phthalates andpolymerization of BPA have driven their widespreaduse in the production of plastics (Thompson et al
7 This journal is q 2009 The Royal Society
2048 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
2009ab) Today phthalates and BPA are found inmany mass-produced products including medicaldevices food packaging perfumes cosmetics chil-drenrsquos toys flooring materials computers CDs etc(Andrady amp Neal 2009) Phthalates may constitute upto 50 per cent of the total weight of PVC plastics andtheir worldwide annual production is approximately27 million metric tonnes (Bauer amp Herrmann 1997)For BPA worldwide annual production is 25 millionmetric tonnes (Staples et al 2002) Plasticizers workby reducing the chemical affinity between moleculeswhen embedded between chains of plastic raw materials(or act as monomers in polycarbonate plastic) but asplasticizers are not especially stable in these productsthey can leach out and thus end up in the environmentPhthalates are not classified as persistent compounds(Staples et al 1997) but their occurrence in theenvironment has been reported widely possibly arguingagainst a rapid biodegradation in some environments(Fatoki amp Vernon 1990 Bauer amp Herrmann 1997Heemken et al 2001 Fromme et al 2002Beauchesne et al 2007) Bisphenol A is easily degraded(Howard 1989) but nevertheless regularly detected inaquatic ecosystems owing to its continuous releaseinto the environment
The European Union Environmental Risk Assessment(ERA) reports provide predicted environmental con-centrations (PECs) for phthalates and BPA using theEuropean Union System for the Evaluation ofSubstances (EUSES) The reliability of some of thesePECs however is disputed as monitoring data from sur-face waters have found significantly higher environmentalconcentrations of phthalates While the PEC for dibutylphthalate (DBP) is 04 mg l21 and 0231 mg kg21 (drywt) for water and sediments respectively (EU 2004)Fatoki amp Vernon (1990) report a maximum measuredconcentration of 335 mg l21 in UK rivers The ERAdraft report calculates a PEC for diethylhexyl phthalate(DEHP) of 22 mg l21 for water and 34 mg kg21 (drywt) for sediments (EU 2006) This figure is exceededin some German surface waters where measured concen-trations range between 033 and 978 mg l21 (Frommeet al 2002) and in Japanese freshwaters and sedimentswhere peak concentrations have been measured at58 mg l21 and 210 000 mg kg21 (dry wt) respectively(average concentrations in Japanese sediments rangebetween 46 and 326 mg kg21 (dry wt Naito et al2006) For di-isodecyl phthalate (DIDP) the EUSES-based calculations state PECs of 18 mg l21 in surfacewaters and 32 mg kg21 (dry wt) in sediments (EU2003a) and for di-isononyl phthalate (DINP)07 mg l21 in surface waters and 18 mg kg21 (dry wt)in sediments (EU 2003b) There is a lack of dataon measured environmental concentrations of thesecompounds to confirm or refute these calculations
Bisphenol A is present in many environmentalcompartments including the aquatic environmentand enters water systems through point dischargessuch as landfill leachates and sewage treatment planteffluents Concentrations of BPA vary widely in theaquatic environment but they may even reach concen-trations up to 21 mg l21 in freshwater systems (Crainet al 2007) Concentrations of BPA in sediments aregenerally several orders of magnitude higher than
Phil Trans R Soc B (2009)
those in the water column For example in the RiverElbe in Germany BPA was measured at 0776 mg l21
in the water compared with 343 mg kg21 (dry wt) inthe sediment (Heemken et al 2001) These findingscontrast with a PEC of 012 mg l21 for water and16 mg kg21 (dry wt) for sediments as indicated bythe ERA report for BPA (EU 2003c)
Heightening our concern about possible healthimplications of exposure to plasticizers phthalates andBPA have been shown to bioaccumulate in organismsBioconcentration factors (BCFs) for plasticizers how-ever appear to differ widely between species Some ofthese differences relate to the type and nature of theplasticizers but others relate to the differences in theexperimental designs employed diet life and speciesrsquohistory and the tissues investigated Bioconcentrationfactors reported in invertebrates are generally higherthan those in vertebrates As an example for DEHPBCFs reported are between 42 and 842 in differentfish species but 3600 for the amphipod Gammaruspseudolimnaueus 2500 in the mussel Mytilus edulis andup to 5380 in the copepod Acartia tonsa (EU 2003d)
In this review paper we analyse critically theeffects on wildlife of the most prominent plasticizerswith a focus on effects in annelids (both aquatic andterrestrial) molluscs crustaceans insects fish andamphibians The paper further presents novel datafrom the authors on the biological effects of someplasticizers on animal species The paper concludeswith an analysis on the level of harm (and theuncertainties) associated with the discharge ofplasticizers into the environment and identifies keyresearch needs to address some of the uncertainties
2 MATERIAL AND METHODSIn this paper a series of experiments are reportedupon that provide novel data on the effects of butylbenzyl phthalate (BBP) on a fish (Danio rerio) andBPA on a crustacean (A tonsa) and an amphibian(Xenopus laevis) These experiments are describedbriefly as detailed information on study designs canbe obtained from the references cited
(a) Danio rerio (zebrafish)A study was conducted to investigate the effects ofBBP on reproduction in adult D rerio Fish were main-tained in breeding colonies of six males and sixfemales according to Santos et al (2007) The chemi-cal exposure tanks were set up in duplicate and dosedwith 0 (solvent controls with methanol 001permil) 3 6and 15 mg BBP l21 and run for 20 days The totalnumber of eggs produced and the number of viableembryos at 8 h post fertilization (hpf) were determinedaccording to Santos et al (2007) At the end of theexperiment quantification of vitellogenin (VTG) inthe plasma was performed according to Fenske et al(2001) with an adjustment of the primary VTG anti-body concentrations to 1 10 000 and a primaryantibody incubation time of 30 min One gonad fromall parent males was dissected and immediately usedfor sperm quantity and motility assessments (as measuresof sperm quality) using computer-assisted sperm
Review Effects of plasticizers on wildlife J Oehlmann et al 2049
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis (CASA v 7V3B Hobson Vision Ltd UK) ona Hobson sperm tracker (Hobson Vision Ltd) accord-ing to the protocols and settings described in Santoset al (2007)
Water samples were collected from each tank on days1 4 7 14 and 20 fixed with 5 methanol (FisherUK) extracted using Sep-Pack C18 cartridges(Waters Corporation USA) and BBP concentrationsof the extracted samples were determined by HPLCanalysis (Shimadzu USA) Samples were analysedthrough separation on a C18 column (SpherisorbODS2) using an isocratic gradient of 70 per centmethanol in water and the absorbance was monitoredat 277 nm on a UV HPLC detector (AppliedBiosystems UK) Quantification was achieved bycomparison of the absorbance of the BBP peak ineach sample with the absorbance of serially dilutedBBP standards (ranging from 00001 to 1 mg ml21)
The statistical comparisons on the VTG andfecundity data were performed using one-wayANOVA followed by Tukeyrsquos multiple comparison post-test analysis (using JANDEL Sigmastat 20 statisticalsoftware) The sperm data were arcsine transformedbefore a one-way ANOVA and Tukeyrsquos multiple compari-son post-test analysis using Excel (Microsoft Excel 2002)and GraphPad Prism (v 402 San Diego CA USA)
(b) Acartia tonsa (calanoid copepod Crustacea)Full life-cycle experiments were conducted with Atonsa starting with eggs and extending for three weeksto reproductively active adults with BPA following anOECD guideline proposal (Kusk amp Wollenberger2007) with minor modifications Animals were exposedin mass cultures (800 ml of mediumtest solution with400 copepods) Nominal BPA test concentrationswere 10 50 100 300 1000 mg l21 Semi-static expo-sures were performed at 20+058C with a photoperiodof 12 L 12 D cycle using a synthetic seawater mediumwith a salinity of 18permil (Kusk amp Wollenberger 1999) Onday 14 the number of eggs produced within a 24 hperiod was determined in isolated groups of 5 females(5 replicates per concentration 10 control groups)and expressed as eggs per female Eggs produced onday 18 were transferred to a clean medium (6 replicatesper concentration each containing 30 eggs) in order toobserve the hatching rate and larval mortality of thesecond generation determined on day 21 The larvaldevelopment ratio (LDR) describing the proportionof animals completing metamorphosis from thenauplius to the copepodite morphology was recordedon day 5 The LDR test (Wollenberger et al 2005)was conducted in parallel to the exposure of the massculture under the same exposure conditions with nom-inal BPA concentrations of 30 60 125 250 5001000 mg l21 with 4 replicates per concentration eachcontaining 30 copepods and 12 control groups
(c) Xenopus laevis (African clawed frog)The impacts of BPA on the thyroid system of X laeviswere determined as described previously for tetrabro-mobisphenol A by Jagnytsch et al (2006) Brieflythe levels of thyroid receptor b (TRb) mRNA weremeasured in the brain as biomarker for the
Phil Trans R Soc B (2009)
bioavailability of thyroid hormones (THs) (Opitzet al 2006) Bisphenol A was applied at concentrationsranging between 25 and 500 mg l21 (25 25 250500 mg l21) with 2 replicates per concentration and20 tadpoles per replicate in a long-term Xenopusmetamorphosis assay (XEMA 21 d) and at BPA concen-trations of 100 250 and 500 mg l21 with and without 01and 10 nM tri-iodothyronine (T3) in short-termexposures for 1 2 and 3 days (starting with duplicatetanks each containing 30 tadpolesaquarium andsubsampling 10 individuals in each replicate everyday) The effects of exposure to BPA during larvaldevelopment on sexual differentiation were also inves-tigated using a flow-through system to the point ofcompletion of metamorphosis (see Lutz et al 2008for details of protocol) Briefly tadpoles were exposedto BPA at concentrations of 1029 1028 1027 and1026 M (228 ng l21 to 228 mg l21) and to dilutionwater without BPA (negative control) A positiveoestrogen control was also included (02 mg 17b-oestradiol l21) In this study there were 4 replicatesfor all treatments and 25 individual tadpoles wereexposed in each tank (n frac14 100 per treatment) Theendpoints investigated were survival rate time to com-pletion of metamorphosis weight sex (based on grossmorphology) and histology of testes In addition geneexpression for hepatic insulin-like growth factor-I(IGF-I) and VTG mRNA was analysed in male Xlaevis immediately at completion of metamorphosisusing semi-quantitative RTndashPCR as describedpreviously (Kloas et al 1999)
3 RESULTS AND DISCUSSION(a) Effects of phthalates
Assessing the biological effects of phthalates such asDEHP in aquatic environments presents a significantchallenge owing to its low water solubility and itstendency to form colloidal dispersions above 3 mg l21Many experiments exposing DEHP via the water atconcentrations 3 mg l21 have nevertheless clearlyestablished concentration-related biological effects(Forget-Leray et al 2005) Despite this risk assess-ments of DEHP in Japan (Naito et al 2006) andthe EU (EU 2003d) have disregarded most studiesadopting concentrations 3 mg l21 and the Japaneseassessment has set the threshold effect level (PNEC)to 3 mg DEHP l21 (Naito et al 2006) This omissionneeds addressing given the effects reported for thisphthalate in this review Working with other highlylipophilic phthalates presents similar solubility prob-lems as for DEHP Di-isononyl phthalate and DIDPhave a log Kow of 88 and a water solubility of only02 mg l21 (EU 2003ab) which makes them evenmore difficult to handle in aquatic media than DEHP
(i) Annelids and molluscsDespite their ecological importance annelids andmolluscs have received relatively little attention inassessing the effects of plasticizers compared withvertebrates or other invertebrates (especially crus-taceans and insects Weltje amp Schulte-Oehlmann2007) Early investigations on annelids and molluscs
2050 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
focused on bioaccumulation and acute toxicity ofphthalates but more recently wider biological effectshave been shown including mitotic inhibition induc-tion of chromosomal aberrations and effects on larvaldevelopment
Considering soil-dwelling organisms studies on theuptake and toxicity of the phthalates dimethyl phthalate(DMP) diethyl phthalate (DEP) DBP and DEHP (ata concentration of 5 mg kg21 for 25 days) in the wormEisenia fetida have shown that high molecular weightphthalates (DBP and DEHP) were incorporated butwith relatively low accumulation factors (0242ndash0307and 0073ndash0244 respectively) and DMP and DEPwere apparently not taken up (Xiao-yu et al 2005) Itis possible that the lack of any uptake of DMP andDEP may be as a consequence of their higher microbialdegradation rates and a faster metabolic degradation(Albro et al 1993) Neuhauser et al (1986) investigatedthe acute toxicity of phthalates and found similar LC50
values (1064ndash3335 mg kg21 for a 14 day soil exposure)for DMP in four soil-dwelling annelid species(Allolobophora tuberculata E fetida Eudrilus eugeniaePerionyx excavatus) Together these studies demonstratethat phthalates enter terrestrial worms but they have alow acute toxicity It is worth emphasizing that soils canreceive very high inputs of phthalates and the totalamount of DEHP deposited in US landfills to 1992has been estimated at 5 million metric tonnes
Comparisons of the acute toxicity of severalphthalate esters [DMP DEP DBP BBP di-n-hexyl-phthalate (DHP) and DEHP] in various aquatic organ-isms including the annelid Lumbriculus variegatus haveshown that the higher molecular weight phthalatesDHP and DEHP were not acutely toxic for the concen-tration range tested (23ndash478 mg DHP l21 102ndash691 mg DEHP l21 Call et al 2001) The toxicity oflower molecular weight phthalates was positively corre-lated with their Kow indicating non-polar narcosis asthe underlying mode of action The same study foundL variegatus was the species least sensitive to DMPthe 10 day LC50 (10 day) was 246 compared with281 and 682 mg l21 for the crustacean Hyalellaazteca and the insect Chironomus tentans respectivelyIn adult marine tubeworms Pomatoceros lamarckiiexposure to DMP was shown to decrease fertilizationsuccess at a threshold concentration of 11025 MDMP (194 mg l21) and induce a significant increasein the number of aberrations in chromosome separ-ations in oocytes at anaphase at concentrations11027 M (194 mg l21) (Dixon et al 1999 Wilsonet al 2002) DMP has been shown to affect larvaldevelopment in the polychaete worm at a thresholdconcentration 1 1025 M (194 mg l21)
The effects of phthalates have also been studied onvarious biochemical pathways such as antioxidant andperoxisomal marker enzymes in marine mussels(Mytilus galloprovincialis Orbea et al 2002 M edulisCajaraville amp Ortiz-Zarragoitia 2006) Exposure to500 mg DEHP l21 for 21 days was shown to result ina significant increase in the catalase and acyl-CoAoxidase (AOX) activity and an inhibition of the super-oxide and manganate superoxide dismutase inM galloprovincialis The peroxisomal volume densityin the digestive gland of M edulis was significantly
Phil Trans R Soc B (2009)
enhanced at 50 mg diallylphthalate (DAP) per litreafter three weeks exposure whereas the chemicalhad no significant effect on AOX activity in thisspecies (Cajaraville amp Ortiz-Zarragoitia 2006) DAPwas able to decrease the phospho-protein level (amussel VTG-like protein) but did not impair ovarianfollicles oocytes and spermatogenesis in M eduliswhen exposed to 50 mg DAP l21 for three weeks(Aarab et al 2006) In another 21 day study DAPwas also shown to increase micronuclei frequency amarker of genotoxicity and fragmented-apoptoticcells in the gills of M edulis at 50 mg DAP l21
(Barsiene et al 2006)Very few studies have considered the effects of
phthalates on annelid or mollusc populations butTagatz et al (1986) found that exposure to DBPaltered the community structure and colonizationprofile of the zoomacrobenthos under laboratory andfield conditions The densities of molluscs andechinoderms were reduced by 49 and 97 per centwhile no effects were seen in annelids arthropodschordates and other groups (at concentrations of10ndash1000 mg kg21 over eight weeks)
(ii) Crustaceans and insectsLower molecular weight phthalates that are relativelywater soluble (Howard et al 1985) have been shownto have a lower acute and chronic toxicity in Daphniamagna For example 48 h immobilization tests withD magna have shown IC50 values of 284 220 and678 mg l21 for DMP DEP and DBP respectively(Jonsson amp Baun 2003) Acute toxicity for phthalateshowever differs significantly for different species ofcrustaceans Hyalella azteca is 10ndash20-fold more sensi-tive than D magna although the higher molecularweight phthalates DHP and DEHP did not exhibittoxic effects in H azteca even at concentrations up to225 mg DHP l21 and 590 mg DEHP l21 (Call et al2001) Other effects of DBP reported in crustaceansinclude decreased locomotor activity in Gammaruspulex at an exposure concentration of 500 mg l21
(Thuren amp Woin 1991)In the Drosophila melanogaster BII cell line DEP
exhibited a weak ecdysteroid antagonistic activity(Dinan et al 2001) However the concentrationsrequired to cause an effect were several orders ofmagnitude higher than that for 20-hydroxy ecdysonethus classifying DEP as a very weak environmentalanti-ecdysteroid only
DEHP has been shown to induce sublethal effectsin Chironomus riparius and the exposure of larvaeresulted in an increased female body volume atexposure concentrations of 03 mg l21 (Kwak amp Lee2005) It also increased the expression of heat shockprotein (HSP) and haemoglobin genes in exposedlarvae of C tentans at a concentration of 05 mg l21
(Lee et al 2006a) but the same compound was notacutely toxic to this species at concentrations up to691 mg DEHP l21 (Call et al 2001) The significanceof such effects on gene expression is not known but invertebrates HSPs are involved in steroid hormonereceptor stabilization and activation (Gillesby ampZacharewski 1998) and if they act similarly in
Review Effects of plasticizers on wildlife J Oehlmann et al 2051
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
arthropods a change in the HSP70 titre might resultin a disruption of the ecdysone axis by influencingthe ecdysone receptor
(iii) FishGiven the widespread occurrence of phthalates in theaquatic environment fish are also likely to be exposedto phthalates via the water column food andor viathe sediments depending on their ecological nicheIn support of this a study in wild fish inThe Netherlands measured median concentrations ofDEHP and DBP at 17 and 10 mg kg21 (wet wt)respectively with 95th percentile values reaching upto 141 mg DEHP kg21 and 26 mg DBP kg21 (wet wt)(Peijnenburg amp Struijs 2006) Laboratory studies havefurther shown that phthalates bioconcentrate in fishFor example the BCF for DEHP was 120 in the carpand for BBP 94 in bluegill sunfish (Staples et al1997) The presence of phthalates in wild fish andtheir ability to concentrate in body tissues demonstratetheir potential to induce adverse effects on the health ofwild populations but there has been little bearing onthis expectation
Very few studies have comprehensively addressedthe biological effects of phthalates on fish and invivo studies have reported mixed findings BBP hasbeen reported to be oestrogenic and induces VTG syn-thesis in male fathead minnow (Pimephales promelas) ata concentration of 100 mg l21 via the water (Harrieset al 2000) and similarly to induce VTG in carp(Cyprinus carpio) exposed to 1ndash20 mg DEP l21 overa period of four weeks (Barse et al 2007) In salmon(Salmo salar) exposure to 1500 mg DEHP kg21 inthe food for a prolonged period (four months)during early life resulted in a small incidence of inter-sex (Norman et al 2007) These in vivo effects areconsistent with the mode of action of BBP andDEHP established in vitro where they have beenshown to bind to the oestrogen receptor (ER) andelicit a weakly oestrogenic response in fish cell cultures(Jobling et al 1995) and in the yeast oestrogen screen(YES containing the human ER) (Harris et al 1997)In female Japanese medaka (Oryzias latipes) exposureto concentrations in excess of 1 mg DEHP l21 caused amarked reduction in both VTG and in the percentageof females containing mature oocytes suggesting ananti-oestrogenic mode of action (Kim et al 2002)This anti-oestrogenic effect presumably arises throughcompetition with the endogenous oestrogens for thereceptor In vitro receptor interaction studies havealso shown that BBP and DEHP possess anti-androgenic activity (Sohoni amp Sumpter 1998) Otherin vivo effects of phthalates in fish include effects onthe metabolic pathways involved in steroid biosyn-thesis and metabolism In a study by Thibaut ampPorte (2004) DEHP and DBP altered the activity ofenzymes involved in the synthesis of endogenoussteroid hormones and their metabolism in carp effectsthat can lead to the alteration of the balance betweenendogenous oestrogens and androgens and thusalterations in their reproductive physiology Thesefindings are supported by studies in mammals inwhich phthalates have also been shown to alter the
Phil Trans R Soc B (2009)
expression and enzymatic activity of enzymes involvedin the synthesis and metabolism of sex steroids in thetestis (Liu et al 2005) In fish exposures to lowmg l21 concentrations of DEP have been shown toinduce alterations in a number of enzymes in theliver and muscle including phosphatases and trans-aminases showing a disruptive effect on generalmetabolic functions (Barse et al 2007) Other studieshave also reported that potentially the most relevantmechanism of action of phthalates is via binding tothe peroxisome proliferator-activated receptors(PPARs) and modulation of the transcriptionalevents coordinated by this molecular pathway whichincludes steroid hormone metabolism (Fan et al2004)
Exposures to phthalates have also been shown toalter behaviour in fish Exposure to 100 mg BBP l21via the water caused alterations in shoaling and feedingbehaviour in three-spined stickleback (Gasterosteusaculeatus) (Wibe et al 2002 2004) and exposure to5 mg DEP l21 caused alterations in the general behaviourof common carp (Barse et al 2007) It should be notedthat these studies have employed very high exposureconcentrations and these are unlikely to occur in thewater column in part owing to their low solubilityExceptions to this may be for fish living incloselyassociated with the sediments in heavily contaminatedenvironments Exposures of fish to lower levels ofphthalates have generally found no adverse effects Asan example exposure of Japanese medaka to 219 and193 mg g21 DINP and DIDP respectively via thediet with a daily feeding regime of 5 per cent of thebody weight failed to identify any effects on reproduc-tion (gonad somatic index egg production embryosurvival and sex ratios) growth or survival in amulti-generational study (14 days post fertilization(dpf) to 140 dpf of the F1 generation) (Patyna et al2006) Similarly exposure of medaka to concentrationsup to 5 mg DEHP l21 for 90 days post-hatch did notaffect germ cell development (Metcalfe et al 2001)
In our exposure of adult breeding zebrafish to BBPmeasured concentrations in the tank water were 3 6and 15 mg l21 over the exposure period Exposure toBBP did not affect the concentration of VTG in theplasma at any of the concentrations tested in males(VTG concentrations in the plasma were 1310+2551500+358 948+141 1300+318 1640+220 ng ml21 for the water control solvent control 36 and 15 mg l21 BBP respectively n frac14 9ndash12 malesper treatment group) Similarly there was no evidenceof any chemical effect on the number of eggs spawnedthe total number of eggs spawned per colony over theassessment period ranged between 4109 and 7578with a maximum of 727 eggs spawned by a singlecolony on a single day with a mean number of eggsspawned per female per day (including days with noeggs) of 222+14 213+14 184+08 216+17 285+14 prior to exposure and 195+16270+17 211+12 250+21 325+14 duringthe exposure for the water control solvent control 36 and 15 mg BBP l21 respectively Furthermore therewere no effects of BBP on the mean percentage viabilityof embryos at 8 hpf (figure 1) These results are inagreement with reports in the literature where VTG
time (days)ndash20 ndash10 0 10 20
cum
ulat
ive
num
ber
of v
iabl
e eg
gs (
8 hp
f)
0
100
200
300
400
500
600
700
Figure 1 Danio rerio Cumulative number of viable eggs spawned in colonies during a 20 day period prior to BBP exposure and
a 20 day period during BBP exposure Symbols represent average results for two colonies per treatment filled circle watercontrol open circle solvent control inverted filled triangle 0003 mg l21 BBP open triangle 0006 mg l21 BBP filledsquare 0015 mg l21 BBP
2052 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
induction in males or reproductive effects on femaleshas been induced only at exposure concentrations thatfar exceed the measured concentrations in our studyIn contrast however we found consistent changes inparameters of sperm quality in BBP-exposed male zeb-rafish The solvent adopted (methanol) had a negativeeffect on zebrafish sperm motility compared with thewater control after 20 days of exposure (mean curvi-linear velocity of 14 mm s21 (mean) and 25 mm s21respectively p 005 figure 2a) and BBP exposureresulted in a concentration-dependent enhancementin curvilinear velocity of the solvent-affected spermAt the highest concentration tested (15 mg BBP l21)the average curvilinear velocity of the sperm wasrestored to levels similar to those found in the watercontrol tanks (31 mm s21 figure 2a) Both thenumber of sperm tracked and the linearity of thesperm movement showed patterns similar to thosedescribed for the curvilinear velocity (figure 2bc)
The ability of BBP to reverse the adverse effect ofthe methanol on sperm number and motility is anovel finding and we hypothesize that this may haveresulted from an interaction of BBP with the PPARreceptor which leads to increased oxidative stress inthe testis as demonstrated in mammals for phthalates(reviewed in Corton amp Lapinskas 2005) In the finalstages of sperm development maturation and acti-vation are crucially dependent on the oxidative balancewithin the seminiferous tubes and in the sperm itselfExposure to methanol may have reduced the freeoxygen radicals required to promote sperm maturationand activation and this effect may have been reversedby the increase in free radicals caused by the BBPexposure The ethanol metabolite acetaldehyde hasbeen shown to block the action of PPARs in vitro(Galli et al 2001) suggesting a mechanism by whichsperm motility might be reduced in fish exposed to asolvent Exposure to BBP in the solvent carrier maycounterbalance this effect restoring the normal
Phil Trans R Soc B (2009)
motility of the sperm The increase in oxidative stresswithin the testis as a result of phthalate exposure hasbeen previously observed in mammalian systemsincluding in rats (Kasahara et al 2002) and thiseffect was shown to be mediated through activationof PPAR pathways
(iv) AmphibiansStudies on phthalates in amphibians (reviewed inKloas 2002) are less extensive than those in fish andsome invertebrates The effects of plasticizers in amp-hibians have been focused on their toxicity theirpotential to induce teratogenic effects and their endo-crine-disrupting activities most notably effects onreproduction sexual differentiation and the thyroidsystem Focusing on toxicity Larsson amp Thuren(1987) found that exposure of Rana arvalis eggs toDEHP via sediment decreased successful hatchingswith increasing concentrations and deduced a noobserved effect concentration (NOEC) of 10 mg kg21
fresh weight Studies with DBP using the frog embryoteratogenesis assay Xenopus (FETAX) have showndevelopmental malformations at 500 mg l21 andenhanced mortality rates at concentrations of 10 and15 mg l21 (Lee et al 2005)
DBP has also been shown to affect sexual differen-tiation in male Rana rugosa tadpoles delaying gonadaldevelopment (Ohtani et al 2000) and altering thespermatogenesis process in eight-month-old X laevisafter exposure during larval development (at 1ndash10 mg DBP l21 from stage 52 to completion of meta-morphosis Lee amp Veeramachaneni 2005) Studies inXenopus investigating the potential for phthalates tobind to the ER via competitive displacement exper-iments with cytosolic liver preparations have foundthat DEP has only a moderate affinity for the ER(Lutz amp Kloas 1999) Parallel in vitro studies using Xlaevis primary hepatocytes however have shown that
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

2048 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
2009ab) Today phthalates and BPA are found inmany mass-produced products including medicaldevices food packaging perfumes cosmetics chil-drenrsquos toys flooring materials computers CDs etc(Andrady amp Neal 2009) Phthalates may constitute upto 50 per cent of the total weight of PVC plastics andtheir worldwide annual production is approximately27 million metric tonnes (Bauer amp Herrmann 1997)For BPA worldwide annual production is 25 millionmetric tonnes (Staples et al 2002) Plasticizers workby reducing the chemical affinity between moleculeswhen embedded between chains of plastic raw materials(or act as monomers in polycarbonate plastic) but asplasticizers are not especially stable in these productsthey can leach out and thus end up in the environmentPhthalates are not classified as persistent compounds(Staples et al 1997) but their occurrence in theenvironment has been reported widely possibly arguingagainst a rapid biodegradation in some environments(Fatoki amp Vernon 1990 Bauer amp Herrmann 1997Heemken et al 2001 Fromme et al 2002Beauchesne et al 2007) Bisphenol A is easily degraded(Howard 1989) but nevertheless regularly detected inaquatic ecosystems owing to its continuous releaseinto the environment
The European Union Environmental Risk Assessment(ERA) reports provide predicted environmental con-centrations (PECs) for phthalates and BPA using theEuropean Union System for the Evaluation ofSubstances (EUSES) The reliability of some of thesePECs however is disputed as monitoring data from sur-face waters have found significantly higher environmentalconcentrations of phthalates While the PEC for dibutylphthalate (DBP) is 04 mg l21 and 0231 mg kg21 (drywt) for water and sediments respectively (EU 2004)Fatoki amp Vernon (1990) report a maximum measuredconcentration of 335 mg l21 in UK rivers The ERAdraft report calculates a PEC for diethylhexyl phthalate(DEHP) of 22 mg l21 for water and 34 mg kg21 (drywt) for sediments (EU 2006) This figure is exceededin some German surface waters where measured concen-trations range between 033 and 978 mg l21 (Frommeet al 2002) and in Japanese freshwaters and sedimentswhere peak concentrations have been measured at58 mg l21 and 210 000 mg kg21 (dry wt) respectively(average concentrations in Japanese sediments rangebetween 46 and 326 mg kg21 (dry wt Naito et al2006) For di-isodecyl phthalate (DIDP) the EUSES-based calculations state PECs of 18 mg l21 in surfacewaters and 32 mg kg21 (dry wt) in sediments (EU2003a) and for di-isononyl phthalate (DINP)07 mg l21 in surface waters and 18 mg kg21 (dry wt)in sediments (EU 2003b) There is a lack of dataon measured environmental concentrations of thesecompounds to confirm or refute these calculations
Bisphenol A is present in many environmentalcompartments including the aquatic environmentand enters water systems through point dischargessuch as landfill leachates and sewage treatment planteffluents Concentrations of BPA vary widely in theaquatic environment but they may even reach concen-trations up to 21 mg l21 in freshwater systems (Crainet al 2007) Concentrations of BPA in sediments aregenerally several orders of magnitude higher than
Phil Trans R Soc B (2009)
those in the water column For example in the RiverElbe in Germany BPA was measured at 0776 mg l21
in the water compared with 343 mg kg21 (dry wt) inthe sediment (Heemken et al 2001) These findingscontrast with a PEC of 012 mg l21 for water and16 mg kg21 (dry wt) for sediments as indicated bythe ERA report for BPA (EU 2003c)
Heightening our concern about possible healthimplications of exposure to plasticizers phthalates andBPA have been shown to bioaccumulate in organismsBioconcentration factors (BCFs) for plasticizers how-ever appear to differ widely between species Some ofthese differences relate to the type and nature of theplasticizers but others relate to the differences in theexperimental designs employed diet life and speciesrsquohistory and the tissues investigated Bioconcentrationfactors reported in invertebrates are generally higherthan those in vertebrates As an example for DEHPBCFs reported are between 42 and 842 in differentfish species but 3600 for the amphipod Gammaruspseudolimnaueus 2500 in the mussel Mytilus edulis andup to 5380 in the copepod Acartia tonsa (EU 2003d)
In this review paper we analyse critically theeffects on wildlife of the most prominent plasticizerswith a focus on effects in annelids (both aquatic andterrestrial) molluscs crustaceans insects fish andamphibians The paper further presents novel datafrom the authors on the biological effects of someplasticizers on animal species The paper concludeswith an analysis on the level of harm (and theuncertainties) associated with the discharge ofplasticizers into the environment and identifies keyresearch needs to address some of the uncertainties
2 MATERIAL AND METHODSIn this paper a series of experiments are reportedupon that provide novel data on the effects of butylbenzyl phthalate (BBP) on a fish (Danio rerio) andBPA on a crustacean (A tonsa) and an amphibian(Xenopus laevis) These experiments are describedbriefly as detailed information on study designs canbe obtained from the references cited
(a) Danio rerio (zebrafish)A study was conducted to investigate the effects ofBBP on reproduction in adult D rerio Fish were main-tained in breeding colonies of six males and sixfemales according to Santos et al (2007) The chemi-cal exposure tanks were set up in duplicate and dosedwith 0 (solvent controls with methanol 001permil) 3 6and 15 mg BBP l21 and run for 20 days The totalnumber of eggs produced and the number of viableembryos at 8 h post fertilization (hpf) were determinedaccording to Santos et al (2007) At the end of theexperiment quantification of vitellogenin (VTG) inthe plasma was performed according to Fenske et al(2001) with an adjustment of the primary VTG anti-body concentrations to 1 10 000 and a primaryantibody incubation time of 30 min One gonad fromall parent males was dissected and immediately usedfor sperm quantity and motility assessments (as measuresof sperm quality) using computer-assisted sperm
Review Effects of plasticizers on wildlife J Oehlmann et al 2049
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis (CASA v 7V3B Hobson Vision Ltd UK) ona Hobson sperm tracker (Hobson Vision Ltd) accord-ing to the protocols and settings described in Santoset al (2007)
Water samples were collected from each tank on days1 4 7 14 and 20 fixed with 5 methanol (FisherUK) extracted using Sep-Pack C18 cartridges(Waters Corporation USA) and BBP concentrationsof the extracted samples were determined by HPLCanalysis (Shimadzu USA) Samples were analysedthrough separation on a C18 column (SpherisorbODS2) using an isocratic gradient of 70 per centmethanol in water and the absorbance was monitoredat 277 nm on a UV HPLC detector (AppliedBiosystems UK) Quantification was achieved bycomparison of the absorbance of the BBP peak ineach sample with the absorbance of serially dilutedBBP standards (ranging from 00001 to 1 mg ml21)
The statistical comparisons on the VTG andfecundity data were performed using one-wayANOVA followed by Tukeyrsquos multiple comparison post-test analysis (using JANDEL Sigmastat 20 statisticalsoftware) The sperm data were arcsine transformedbefore a one-way ANOVA and Tukeyrsquos multiple compari-son post-test analysis using Excel (Microsoft Excel 2002)and GraphPad Prism (v 402 San Diego CA USA)
(b) Acartia tonsa (calanoid copepod Crustacea)Full life-cycle experiments were conducted with Atonsa starting with eggs and extending for three weeksto reproductively active adults with BPA following anOECD guideline proposal (Kusk amp Wollenberger2007) with minor modifications Animals were exposedin mass cultures (800 ml of mediumtest solution with400 copepods) Nominal BPA test concentrationswere 10 50 100 300 1000 mg l21 Semi-static expo-sures were performed at 20+058C with a photoperiodof 12 L 12 D cycle using a synthetic seawater mediumwith a salinity of 18permil (Kusk amp Wollenberger 1999) Onday 14 the number of eggs produced within a 24 hperiod was determined in isolated groups of 5 females(5 replicates per concentration 10 control groups)and expressed as eggs per female Eggs produced onday 18 were transferred to a clean medium (6 replicatesper concentration each containing 30 eggs) in order toobserve the hatching rate and larval mortality of thesecond generation determined on day 21 The larvaldevelopment ratio (LDR) describing the proportionof animals completing metamorphosis from thenauplius to the copepodite morphology was recordedon day 5 The LDR test (Wollenberger et al 2005)was conducted in parallel to the exposure of the massculture under the same exposure conditions with nom-inal BPA concentrations of 30 60 125 250 5001000 mg l21 with 4 replicates per concentration eachcontaining 30 copepods and 12 control groups
(c) Xenopus laevis (African clawed frog)The impacts of BPA on the thyroid system of X laeviswere determined as described previously for tetrabro-mobisphenol A by Jagnytsch et al (2006) Brieflythe levels of thyroid receptor b (TRb) mRNA weremeasured in the brain as biomarker for the
Phil Trans R Soc B (2009)
bioavailability of thyroid hormones (THs) (Opitzet al 2006) Bisphenol A was applied at concentrationsranging between 25 and 500 mg l21 (25 25 250500 mg l21) with 2 replicates per concentration and20 tadpoles per replicate in a long-term Xenopusmetamorphosis assay (XEMA 21 d) and at BPA concen-trations of 100 250 and 500 mg l21 with and without 01and 10 nM tri-iodothyronine (T3) in short-termexposures for 1 2 and 3 days (starting with duplicatetanks each containing 30 tadpolesaquarium andsubsampling 10 individuals in each replicate everyday) The effects of exposure to BPA during larvaldevelopment on sexual differentiation were also inves-tigated using a flow-through system to the point ofcompletion of metamorphosis (see Lutz et al 2008for details of protocol) Briefly tadpoles were exposedto BPA at concentrations of 1029 1028 1027 and1026 M (228 ng l21 to 228 mg l21) and to dilutionwater without BPA (negative control) A positiveoestrogen control was also included (02 mg 17b-oestradiol l21) In this study there were 4 replicatesfor all treatments and 25 individual tadpoles wereexposed in each tank (n frac14 100 per treatment) Theendpoints investigated were survival rate time to com-pletion of metamorphosis weight sex (based on grossmorphology) and histology of testes In addition geneexpression for hepatic insulin-like growth factor-I(IGF-I) and VTG mRNA was analysed in male Xlaevis immediately at completion of metamorphosisusing semi-quantitative RTndashPCR as describedpreviously (Kloas et al 1999)
3 RESULTS AND DISCUSSION(a) Effects of phthalates
Assessing the biological effects of phthalates such asDEHP in aquatic environments presents a significantchallenge owing to its low water solubility and itstendency to form colloidal dispersions above 3 mg l21Many experiments exposing DEHP via the water atconcentrations 3 mg l21 have nevertheless clearlyestablished concentration-related biological effects(Forget-Leray et al 2005) Despite this risk assess-ments of DEHP in Japan (Naito et al 2006) andthe EU (EU 2003d) have disregarded most studiesadopting concentrations 3 mg l21 and the Japaneseassessment has set the threshold effect level (PNEC)to 3 mg DEHP l21 (Naito et al 2006) This omissionneeds addressing given the effects reported for thisphthalate in this review Working with other highlylipophilic phthalates presents similar solubility prob-lems as for DEHP Di-isononyl phthalate and DIDPhave a log Kow of 88 and a water solubility of only02 mg l21 (EU 2003ab) which makes them evenmore difficult to handle in aquatic media than DEHP
(i) Annelids and molluscsDespite their ecological importance annelids andmolluscs have received relatively little attention inassessing the effects of plasticizers compared withvertebrates or other invertebrates (especially crus-taceans and insects Weltje amp Schulte-Oehlmann2007) Early investigations on annelids and molluscs
2050 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
focused on bioaccumulation and acute toxicity ofphthalates but more recently wider biological effectshave been shown including mitotic inhibition induc-tion of chromosomal aberrations and effects on larvaldevelopment
Considering soil-dwelling organisms studies on theuptake and toxicity of the phthalates dimethyl phthalate(DMP) diethyl phthalate (DEP) DBP and DEHP (ata concentration of 5 mg kg21 for 25 days) in the wormEisenia fetida have shown that high molecular weightphthalates (DBP and DEHP) were incorporated butwith relatively low accumulation factors (0242ndash0307and 0073ndash0244 respectively) and DMP and DEPwere apparently not taken up (Xiao-yu et al 2005) Itis possible that the lack of any uptake of DMP andDEP may be as a consequence of their higher microbialdegradation rates and a faster metabolic degradation(Albro et al 1993) Neuhauser et al (1986) investigatedthe acute toxicity of phthalates and found similar LC50
values (1064ndash3335 mg kg21 for a 14 day soil exposure)for DMP in four soil-dwelling annelid species(Allolobophora tuberculata E fetida Eudrilus eugeniaePerionyx excavatus) Together these studies demonstratethat phthalates enter terrestrial worms but they have alow acute toxicity It is worth emphasizing that soils canreceive very high inputs of phthalates and the totalamount of DEHP deposited in US landfills to 1992has been estimated at 5 million metric tonnes
Comparisons of the acute toxicity of severalphthalate esters [DMP DEP DBP BBP di-n-hexyl-phthalate (DHP) and DEHP] in various aquatic organ-isms including the annelid Lumbriculus variegatus haveshown that the higher molecular weight phthalatesDHP and DEHP were not acutely toxic for the concen-tration range tested (23ndash478 mg DHP l21 102ndash691 mg DEHP l21 Call et al 2001) The toxicity oflower molecular weight phthalates was positively corre-lated with their Kow indicating non-polar narcosis asthe underlying mode of action The same study foundL variegatus was the species least sensitive to DMPthe 10 day LC50 (10 day) was 246 compared with281 and 682 mg l21 for the crustacean Hyalellaazteca and the insect Chironomus tentans respectivelyIn adult marine tubeworms Pomatoceros lamarckiiexposure to DMP was shown to decrease fertilizationsuccess at a threshold concentration of 11025 MDMP (194 mg l21) and induce a significant increasein the number of aberrations in chromosome separ-ations in oocytes at anaphase at concentrations11027 M (194 mg l21) (Dixon et al 1999 Wilsonet al 2002) DMP has been shown to affect larvaldevelopment in the polychaete worm at a thresholdconcentration 1 1025 M (194 mg l21)
The effects of phthalates have also been studied onvarious biochemical pathways such as antioxidant andperoxisomal marker enzymes in marine mussels(Mytilus galloprovincialis Orbea et al 2002 M edulisCajaraville amp Ortiz-Zarragoitia 2006) Exposure to500 mg DEHP l21 for 21 days was shown to result ina significant increase in the catalase and acyl-CoAoxidase (AOX) activity and an inhibition of the super-oxide and manganate superoxide dismutase inM galloprovincialis The peroxisomal volume densityin the digestive gland of M edulis was significantly
Phil Trans R Soc B (2009)
enhanced at 50 mg diallylphthalate (DAP) per litreafter three weeks exposure whereas the chemicalhad no significant effect on AOX activity in thisspecies (Cajaraville amp Ortiz-Zarragoitia 2006) DAPwas able to decrease the phospho-protein level (amussel VTG-like protein) but did not impair ovarianfollicles oocytes and spermatogenesis in M eduliswhen exposed to 50 mg DAP l21 for three weeks(Aarab et al 2006) In another 21 day study DAPwas also shown to increase micronuclei frequency amarker of genotoxicity and fragmented-apoptoticcells in the gills of M edulis at 50 mg DAP l21
(Barsiene et al 2006)Very few studies have considered the effects of
phthalates on annelid or mollusc populations butTagatz et al (1986) found that exposure to DBPaltered the community structure and colonizationprofile of the zoomacrobenthos under laboratory andfield conditions The densities of molluscs andechinoderms were reduced by 49 and 97 per centwhile no effects were seen in annelids arthropodschordates and other groups (at concentrations of10ndash1000 mg kg21 over eight weeks)
(ii) Crustaceans and insectsLower molecular weight phthalates that are relativelywater soluble (Howard et al 1985) have been shownto have a lower acute and chronic toxicity in Daphniamagna For example 48 h immobilization tests withD magna have shown IC50 values of 284 220 and678 mg l21 for DMP DEP and DBP respectively(Jonsson amp Baun 2003) Acute toxicity for phthalateshowever differs significantly for different species ofcrustaceans Hyalella azteca is 10ndash20-fold more sensi-tive than D magna although the higher molecularweight phthalates DHP and DEHP did not exhibittoxic effects in H azteca even at concentrations up to225 mg DHP l21 and 590 mg DEHP l21 (Call et al2001) Other effects of DBP reported in crustaceansinclude decreased locomotor activity in Gammaruspulex at an exposure concentration of 500 mg l21
(Thuren amp Woin 1991)In the Drosophila melanogaster BII cell line DEP
exhibited a weak ecdysteroid antagonistic activity(Dinan et al 2001) However the concentrationsrequired to cause an effect were several orders ofmagnitude higher than that for 20-hydroxy ecdysonethus classifying DEP as a very weak environmentalanti-ecdysteroid only
DEHP has been shown to induce sublethal effectsin Chironomus riparius and the exposure of larvaeresulted in an increased female body volume atexposure concentrations of 03 mg l21 (Kwak amp Lee2005) It also increased the expression of heat shockprotein (HSP) and haemoglobin genes in exposedlarvae of C tentans at a concentration of 05 mg l21
(Lee et al 2006a) but the same compound was notacutely toxic to this species at concentrations up to691 mg DEHP l21 (Call et al 2001) The significanceof such effects on gene expression is not known but invertebrates HSPs are involved in steroid hormonereceptor stabilization and activation (Gillesby ampZacharewski 1998) and if they act similarly in
Review Effects of plasticizers on wildlife J Oehlmann et al 2051
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
arthropods a change in the HSP70 titre might resultin a disruption of the ecdysone axis by influencingthe ecdysone receptor
(iii) FishGiven the widespread occurrence of phthalates in theaquatic environment fish are also likely to be exposedto phthalates via the water column food andor viathe sediments depending on their ecological nicheIn support of this a study in wild fish inThe Netherlands measured median concentrations ofDEHP and DBP at 17 and 10 mg kg21 (wet wt)respectively with 95th percentile values reaching upto 141 mg DEHP kg21 and 26 mg DBP kg21 (wet wt)(Peijnenburg amp Struijs 2006) Laboratory studies havefurther shown that phthalates bioconcentrate in fishFor example the BCF for DEHP was 120 in the carpand for BBP 94 in bluegill sunfish (Staples et al1997) The presence of phthalates in wild fish andtheir ability to concentrate in body tissues demonstratetheir potential to induce adverse effects on the health ofwild populations but there has been little bearing onthis expectation
Very few studies have comprehensively addressedthe biological effects of phthalates on fish and invivo studies have reported mixed findings BBP hasbeen reported to be oestrogenic and induces VTG syn-thesis in male fathead minnow (Pimephales promelas) ata concentration of 100 mg l21 via the water (Harrieset al 2000) and similarly to induce VTG in carp(Cyprinus carpio) exposed to 1ndash20 mg DEP l21 overa period of four weeks (Barse et al 2007) In salmon(Salmo salar) exposure to 1500 mg DEHP kg21 inthe food for a prolonged period (four months)during early life resulted in a small incidence of inter-sex (Norman et al 2007) These in vivo effects areconsistent with the mode of action of BBP andDEHP established in vitro where they have beenshown to bind to the oestrogen receptor (ER) andelicit a weakly oestrogenic response in fish cell cultures(Jobling et al 1995) and in the yeast oestrogen screen(YES containing the human ER) (Harris et al 1997)In female Japanese medaka (Oryzias latipes) exposureto concentrations in excess of 1 mg DEHP l21 caused amarked reduction in both VTG and in the percentageof females containing mature oocytes suggesting ananti-oestrogenic mode of action (Kim et al 2002)This anti-oestrogenic effect presumably arises throughcompetition with the endogenous oestrogens for thereceptor In vitro receptor interaction studies havealso shown that BBP and DEHP possess anti-androgenic activity (Sohoni amp Sumpter 1998) Otherin vivo effects of phthalates in fish include effects onthe metabolic pathways involved in steroid biosyn-thesis and metabolism In a study by Thibaut ampPorte (2004) DEHP and DBP altered the activity ofenzymes involved in the synthesis of endogenoussteroid hormones and their metabolism in carp effectsthat can lead to the alteration of the balance betweenendogenous oestrogens and androgens and thusalterations in their reproductive physiology Thesefindings are supported by studies in mammals inwhich phthalates have also been shown to alter the
Phil Trans R Soc B (2009)
expression and enzymatic activity of enzymes involvedin the synthesis and metabolism of sex steroids in thetestis (Liu et al 2005) In fish exposures to lowmg l21 concentrations of DEP have been shown toinduce alterations in a number of enzymes in theliver and muscle including phosphatases and trans-aminases showing a disruptive effect on generalmetabolic functions (Barse et al 2007) Other studieshave also reported that potentially the most relevantmechanism of action of phthalates is via binding tothe peroxisome proliferator-activated receptors(PPARs) and modulation of the transcriptionalevents coordinated by this molecular pathway whichincludes steroid hormone metabolism (Fan et al2004)
Exposures to phthalates have also been shown toalter behaviour in fish Exposure to 100 mg BBP l21via the water caused alterations in shoaling and feedingbehaviour in three-spined stickleback (Gasterosteusaculeatus) (Wibe et al 2002 2004) and exposure to5 mg DEP l21 caused alterations in the general behaviourof common carp (Barse et al 2007) It should be notedthat these studies have employed very high exposureconcentrations and these are unlikely to occur in thewater column in part owing to their low solubilityExceptions to this may be for fish living incloselyassociated with the sediments in heavily contaminatedenvironments Exposures of fish to lower levels ofphthalates have generally found no adverse effects Asan example exposure of Japanese medaka to 219 and193 mg g21 DINP and DIDP respectively via thediet with a daily feeding regime of 5 per cent of thebody weight failed to identify any effects on reproduc-tion (gonad somatic index egg production embryosurvival and sex ratios) growth or survival in amulti-generational study (14 days post fertilization(dpf) to 140 dpf of the F1 generation) (Patyna et al2006) Similarly exposure of medaka to concentrationsup to 5 mg DEHP l21 for 90 days post-hatch did notaffect germ cell development (Metcalfe et al 2001)
In our exposure of adult breeding zebrafish to BBPmeasured concentrations in the tank water were 3 6and 15 mg l21 over the exposure period Exposure toBBP did not affect the concentration of VTG in theplasma at any of the concentrations tested in males(VTG concentrations in the plasma were 1310+2551500+358 948+141 1300+318 1640+220 ng ml21 for the water control solvent control 36 and 15 mg l21 BBP respectively n frac14 9ndash12 malesper treatment group) Similarly there was no evidenceof any chemical effect on the number of eggs spawnedthe total number of eggs spawned per colony over theassessment period ranged between 4109 and 7578with a maximum of 727 eggs spawned by a singlecolony on a single day with a mean number of eggsspawned per female per day (including days with noeggs) of 222+14 213+14 184+08 216+17 285+14 prior to exposure and 195+16270+17 211+12 250+21 325+14 duringthe exposure for the water control solvent control 36 and 15 mg BBP l21 respectively Furthermore therewere no effects of BBP on the mean percentage viabilityof embryos at 8 hpf (figure 1) These results are inagreement with reports in the literature where VTG
time (days)ndash20 ndash10 0 10 20
cum
ulat
ive
num
ber
of v
iabl
e eg
gs (
8 hp
f)
0
100
200
300
400
500
600
700
Figure 1 Danio rerio Cumulative number of viable eggs spawned in colonies during a 20 day period prior to BBP exposure and
a 20 day period during BBP exposure Symbols represent average results for two colonies per treatment filled circle watercontrol open circle solvent control inverted filled triangle 0003 mg l21 BBP open triangle 0006 mg l21 BBP filledsquare 0015 mg l21 BBP
2052 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
induction in males or reproductive effects on femaleshas been induced only at exposure concentrations thatfar exceed the measured concentrations in our studyIn contrast however we found consistent changes inparameters of sperm quality in BBP-exposed male zeb-rafish The solvent adopted (methanol) had a negativeeffect on zebrafish sperm motility compared with thewater control after 20 days of exposure (mean curvi-linear velocity of 14 mm s21 (mean) and 25 mm s21respectively p 005 figure 2a) and BBP exposureresulted in a concentration-dependent enhancementin curvilinear velocity of the solvent-affected spermAt the highest concentration tested (15 mg BBP l21)the average curvilinear velocity of the sperm wasrestored to levels similar to those found in the watercontrol tanks (31 mm s21 figure 2a) Both thenumber of sperm tracked and the linearity of thesperm movement showed patterns similar to thosedescribed for the curvilinear velocity (figure 2bc)
The ability of BBP to reverse the adverse effect ofthe methanol on sperm number and motility is anovel finding and we hypothesize that this may haveresulted from an interaction of BBP with the PPARreceptor which leads to increased oxidative stress inthe testis as demonstrated in mammals for phthalates(reviewed in Corton amp Lapinskas 2005) In the finalstages of sperm development maturation and acti-vation are crucially dependent on the oxidative balancewithin the seminiferous tubes and in the sperm itselfExposure to methanol may have reduced the freeoxygen radicals required to promote sperm maturationand activation and this effect may have been reversedby the increase in free radicals caused by the BBPexposure The ethanol metabolite acetaldehyde hasbeen shown to block the action of PPARs in vitro(Galli et al 2001) suggesting a mechanism by whichsperm motility might be reduced in fish exposed to asolvent Exposure to BBP in the solvent carrier maycounterbalance this effect restoring the normal
Phil Trans R Soc B (2009)
motility of the sperm The increase in oxidative stresswithin the testis as a result of phthalate exposure hasbeen previously observed in mammalian systemsincluding in rats (Kasahara et al 2002) and thiseffect was shown to be mediated through activationof PPAR pathways
(iv) AmphibiansStudies on phthalates in amphibians (reviewed inKloas 2002) are less extensive than those in fish andsome invertebrates The effects of plasticizers in amp-hibians have been focused on their toxicity theirpotential to induce teratogenic effects and their endo-crine-disrupting activities most notably effects onreproduction sexual differentiation and the thyroidsystem Focusing on toxicity Larsson amp Thuren(1987) found that exposure of Rana arvalis eggs toDEHP via sediment decreased successful hatchingswith increasing concentrations and deduced a noobserved effect concentration (NOEC) of 10 mg kg21
fresh weight Studies with DBP using the frog embryoteratogenesis assay Xenopus (FETAX) have showndevelopmental malformations at 500 mg l21 andenhanced mortality rates at concentrations of 10 and15 mg l21 (Lee et al 2005)
DBP has also been shown to affect sexual differen-tiation in male Rana rugosa tadpoles delaying gonadaldevelopment (Ohtani et al 2000) and altering thespermatogenesis process in eight-month-old X laevisafter exposure during larval development (at 1ndash10 mg DBP l21 from stage 52 to completion of meta-morphosis Lee amp Veeramachaneni 2005) Studies inXenopus investigating the potential for phthalates tobind to the ER via competitive displacement exper-iments with cytosolic liver preparations have foundthat DEP has only a moderate affinity for the ER(Lutz amp Kloas 1999) Parallel in vitro studies using Xlaevis primary hepatocytes however have shown that
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

Review Effects of plasticizers on wildlife J Oehlmann et al 2049
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis (CASA v 7V3B Hobson Vision Ltd UK) ona Hobson sperm tracker (Hobson Vision Ltd) accord-ing to the protocols and settings described in Santoset al (2007)
Water samples were collected from each tank on days1 4 7 14 and 20 fixed with 5 methanol (FisherUK) extracted using Sep-Pack C18 cartridges(Waters Corporation USA) and BBP concentrationsof the extracted samples were determined by HPLCanalysis (Shimadzu USA) Samples were analysedthrough separation on a C18 column (SpherisorbODS2) using an isocratic gradient of 70 per centmethanol in water and the absorbance was monitoredat 277 nm on a UV HPLC detector (AppliedBiosystems UK) Quantification was achieved bycomparison of the absorbance of the BBP peak ineach sample with the absorbance of serially dilutedBBP standards (ranging from 00001 to 1 mg ml21)
The statistical comparisons on the VTG andfecundity data were performed using one-wayANOVA followed by Tukeyrsquos multiple comparison post-test analysis (using JANDEL Sigmastat 20 statisticalsoftware) The sperm data were arcsine transformedbefore a one-way ANOVA and Tukeyrsquos multiple compari-son post-test analysis using Excel (Microsoft Excel 2002)and GraphPad Prism (v 402 San Diego CA USA)
(b) Acartia tonsa (calanoid copepod Crustacea)Full life-cycle experiments were conducted with Atonsa starting with eggs and extending for three weeksto reproductively active adults with BPA following anOECD guideline proposal (Kusk amp Wollenberger2007) with minor modifications Animals were exposedin mass cultures (800 ml of mediumtest solution with400 copepods) Nominal BPA test concentrationswere 10 50 100 300 1000 mg l21 Semi-static expo-sures were performed at 20+058C with a photoperiodof 12 L 12 D cycle using a synthetic seawater mediumwith a salinity of 18permil (Kusk amp Wollenberger 1999) Onday 14 the number of eggs produced within a 24 hperiod was determined in isolated groups of 5 females(5 replicates per concentration 10 control groups)and expressed as eggs per female Eggs produced onday 18 were transferred to a clean medium (6 replicatesper concentration each containing 30 eggs) in order toobserve the hatching rate and larval mortality of thesecond generation determined on day 21 The larvaldevelopment ratio (LDR) describing the proportionof animals completing metamorphosis from thenauplius to the copepodite morphology was recordedon day 5 The LDR test (Wollenberger et al 2005)was conducted in parallel to the exposure of the massculture under the same exposure conditions with nom-inal BPA concentrations of 30 60 125 250 5001000 mg l21 with 4 replicates per concentration eachcontaining 30 copepods and 12 control groups
(c) Xenopus laevis (African clawed frog)The impacts of BPA on the thyroid system of X laeviswere determined as described previously for tetrabro-mobisphenol A by Jagnytsch et al (2006) Brieflythe levels of thyroid receptor b (TRb) mRNA weremeasured in the brain as biomarker for the
Phil Trans R Soc B (2009)
bioavailability of thyroid hormones (THs) (Opitzet al 2006) Bisphenol A was applied at concentrationsranging between 25 and 500 mg l21 (25 25 250500 mg l21) with 2 replicates per concentration and20 tadpoles per replicate in a long-term Xenopusmetamorphosis assay (XEMA 21 d) and at BPA concen-trations of 100 250 and 500 mg l21 with and without 01and 10 nM tri-iodothyronine (T3) in short-termexposures for 1 2 and 3 days (starting with duplicatetanks each containing 30 tadpolesaquarium andsubsampling 10 individuals in each replicate everyday) The effects of exposure to BPA during larvaldevelopment on sexual differentiation were also inves-tigated using a flow-through system to the point ofcompletion of metamorphosis (see Lutz et al 2008for details of protocol) Briefly tadpoles were exposedto BPA at concentrations of 1029 1028 1027 and1026 M (228 ng l21 to 228 mg l21) and to dilutionwater without BPA (negative control) A positiveoestrogen control was also included (02 mg 17b-oestradiol l21) In this study there were 4 replicatesfor all treatments and 25 individual tadpoles wereexposed in each tank (n frac14 100 per treatment) Theendpoints investigated were survival rate time to com-pletion of metamorphosis weight sex (based on grossmorphology) and histology of testes In addition geneexpression for hepatic insulin-like growth factor-I(IGF-I) and VTG mRNA was analysed in male Xlaevis immediately at completion of metamorphosisusing semi-quantitative RTndashPCR as describedpreviously (Kloas et al 1999)
3 RESULTS AND DISCUSSION(a) Effects of phthalates
Assessing the biological effects of phthalates such asDEHP in aquatic environments presents a significantchallenge owing to its low water solubility and itstendency to form colloidal dispersions above 3 mg l21Many experiments exposing DEHP via the water atconcentrations 3 mg l21 have nevertheless clearlyestablished concentration-related biological effects(Forget-Leray et al 2005) Despite this risk assess-ments of DEHP in Japan (Naito et al 2006) andthe EU (EU 2003d) have disregarded most studiesadopting concentrations 3 mg l21 and the Japaneseassessment has set the threshold effect level (PNEC)to 3 mg DEHP l21 (Naito et al 2006) This omissionneeds addressing given the effects reported for thisphthalate in this review Working with other highlylipophilic phthalates presents similar solubility prob-lems as for DEHP Di-isononyl phthalate and DIDPhave a log Kow of 88 and a water solubility of only02 mg l21 (EU 2003ab) which makes them evenmore difficult to handle in aquatic media than DEHP
(i) Annelids and molluscsDespite their ecological importance annelids andmolluscs have received relatively little attention inassessing the effects of plasticizers compared withvertebrates or other invertebrates (especially crus-taceans and insects Weltje amp Schulte-Oehlmann2007) Early investigations on annelids and molluscs
2050 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
focused on bioaccumulation and acute toxicity ofphthalates but more recently wider biological effectshave been shown including mitotic inhibition induc-tion of chromosomal aberrations and effects on larvaldevelopment
Considering soil-dwelling organisms studies on theuptake and toxicity of the phthalates dimethyl phthalate(DMP) diethyl phthalate (DEP) DBP and DEHP (ata concentration of 5 mg kg21 for 25 days) in the wormEisenia fetida have shown that high molecular weightphthalates (DBP and DEHP) were incorporated butwith relatively low accumulation factors (0242ndash0307and 0073ndash0244 respectively) and DMP and DEPwere apparently not taken up (Xiao-yu et al 2005) Itis possible that the lack of any uptake of DMP andDEP may be as a consequence of their higher microbialdegradation rates and a faster metabolic degradation(Albro et al 1993) Neuhauser et al (1986) investigatedthe acute toxicity of phthalates and found similar LC50
values (1064ndash3335 mg kg21 for a 14 day soil exposure)for DMP in four soil-dwelling annelid species(Allolobophora tuberculata E fetida Eudrilus eugeniaePerionyx excavatus) Together these studies demonstratethat phthalates enter terrestrial worms but they have alow acute toxicity It is worth emphasizing that soils canreceive very high inputs of phthalates and the totalamount of DEHP deposited in US landfills to 1992has been estimated at 5 million metric tonnes
Comparisons of the acute toxicity of severalphthalate esters [DMP DEP DBP BBP di-n-hexyl-phthalate (DHP) and DEHP] in various aquatic organ-isms including the annelid Lumbriculus variegatus haveshown that the higher molecular weight phthalatesDHP and DEHP were not acutely toxic for the concen-tration range tested (23ndash478 mg DHP l21 102ndash691 mg DEHP l21 Call et al 2001) The toxicity oflower molecular weight phthalates was positively corre-lated with their Kow indicating non-polar narcosis asthe underlying mode of action The same study foundL variegatus was the species least sensitive to DMPthe 10 day LC50 (10 day) was 246 compared with281 and 682 mg l21 for the crustacean Hyalellaazteca and the insect Chironomus tentans respectivelyIn adult marine tubeworms Pomatoceros lamarckiiexposure to DMP was shown to decrease fertilizationsuccess at a threshold concentration of 11025 MDMP (194 mg l21) and induce a significant increasein the number of aberrations in chromosome separ-ations in oocytes at anaphase at concentrations11027 M (194 mg l21) (Dixon et al 1999 Wilsonet al 2002) DMP has been shown to affect larvaldevelopment in the polychaete worm at a thresholdconcentration 1 1025 M (194 mg l21)
The effects of phthalates have also been studied onvarious biochemical pathways such as antioxidant andperoxisomal marker enzymes in marine mussels(Mytilus galloprovincialis Orbea et al 2002 M edulisCajaraville amp Ortiz-Zarragoitia 2006) Exposure to500 mg DEHP l21 for 21 days was shown to result ina significant increase in the catalase and acyl-CoAoxidase (AOX) activity and an inhibition of the super-oxide and manganate superoxide dismutase inM galloprovincialis The peroxisomal volume densityin the digestive gland of M edulis was significantly
Phil Trans R Soc B (2009)
enhanced at 50 mg diallylphthalate (DAP) per litreafter three weeks exposure whereas the chemicalhad no significant effect on AOX activity in thisspecies (Cajaraville amp Ortiz-Zarragoitia 2006) DAPwas able to decrease the phospho-protein level (amussel VTG-like protein) but did not impair ovarianfollicles oocytes and spermatogenesis in M eduliswhen exposed to 50 mg DAP l21 for three weeks(Aarab et al 2006) In another 21 day study DAPwas also shown to increase micronuclei frequency amarker of genotoxicity and fragmented-apoptoticcells in the gills of M edulis at 50 mg DAP l21
(Barsiene et al 2006)Very few studies have considered the effects of
phthalates on annelid or mollusc populations butTagatz et al (1986) found that exposure to DBPaltered the community structure and colonizationprofile of the zoomacrobenthos under laboratory andfield conditions The densities of molluscs andechinoderms were reduced by 49 and 97 per centwhile no effects were seen in annelids arthropodschordates and other groups (at concentrations of10ndash1000 mg kg21 over eight weeks)
(ii) Crustaceans and insectsLower molecular weight phthalates that are relativelywater soluble (Howard et al 1985) have been shownto have a lower acute and chronic toxicity in Daphniamagna For example 48 h immobilization tests withD magna have shown IC50 values of 284 220 and678 mg l21 for DMP DEP and DBP respectively(Jonsson amp Baun 2003) Acute toxicity for phthalateshowever differs significantly for different species ofcrustaceans Hyalella azteca is 10ndash20-fold more sensi-tive than D magna although the higher molecularweight phthalates DHP and DEHP did not exhibittoxic effects in H azteca even at concentrations up to225 mg DHP l21 and 590 mg DEHP l21 (Call et al2001) Other effects of DBP reported in crustaceansinclude decreased locomotor activity in Gammaruspulex at an exposure concentration of 500 mg l21
(Thuren amp Woin 1991)In the Drosophila melanogaster BII cell line DEP
exhibited a weak ecdysteroid antagonistic activity(Dinan et al 2001) However the concentrationsrequired to cause an effect were several orders ofmagnitude higher than that for 20-hydroxy ecdysonethus classifying DEP as a very weak environmentalanti-ecdysteroid only
DEHP has been shown to induce sublethal effectsin Chironomus riparius and the exposure of larvaeresulted in an increased female body volume atexposure concentrations of 03 mg l21 (Kwak amp Lee2005) It also increased the expression of heat shockprotein (HSP) and haemoglobin genes in exposedlarvae of C tentans at a concentration of 05 mg l21
(Lee et al 2006a) but the same compound was notacutely toxic to this species at concentrations up to691 mg DEHP l21 (Call et al 2001) The significanceof such effects on gene expression is not known but invertebrates HSPs are involved in steroid hormonereceptor stabilization and activation (Gillesby ampZacharewski 1998) and if they act similarly in
Review Effects of plasticizers on wildlife J Oehlmann et al 2051
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
arthropods a change in the HSP70 titre might resultin a disruption of the ecdysone axis by influencingthe ecdysone receptor
(iii) FishGiven the widespread occurrence of phthalates in theaquatic environment fish are also likely to be exposedto phthalates via the water column food andor viathe sediments depending on their ecological nicheIn support of this a study in wild fish inThe Netherlands measured median concentrations ofDEHP and DBP at 17 and 10 mg kg21 (wet wt)respectively with 95th percentile values reaching upto 141 mg DEHP kg21 and 26 mg DBP kg21 (wet wt)(Peijnenburg amp Struijs 2006) Laboratory studies havefurther shown that phthalates bioconcentrate in fishFor example the BCF for DEHP was 120 in the carpand for BBP 94 in bluegill sunfish (Staples et al1997) The presence of phthalates in wild fish andtheir ability to concentrate in body tissues demonstratetheir potential to induce adverse effects on the health ofwild populations but there has been little bearing onthis expectation
Very few studies have comprehensively addressedthe biological effects of phthalates on fish and invivo studies have reported mixed findings BBP hasbeen reported to be oestrogenic and induces VTG syn-thesis in male fathead minnow (Pimephales promelas) ata concentration of 100 mg l21 via the water (Harrieset al 2000) and similarly to induce VTG in carp(Cyprinus carpio) exposed to 1ndash20 mg DEP l21 overa period of four weeks (Barse et al 2007) In salmon(Salmo salar) exposure to 1500 mg DEHP kg21 inthe food for a prolonged period (four months)during early life resulted in a small incidence of inter-sex (Norman et al 2007) These in vivo effects areconsistent with the mode of action of BBP andDEHP established in vitro where they have beenshown to bind to the oestrogen receptor (ER) andelicit a weakly oestrogenic response in fish cell cultures(Jobling et al 1995) and in the yeast oestrogen screen(YES containing the human ER) (Harris et al 1997)In female Japanese medaka (Oryzias latipes) exposureto concentrations in excess of 1 mg DEHP l21 caused amarked reduction in both VTG and in the percentageof females containing mature oocytes suggesting ananti-oestrogenic mode of action (Kim et al 2002)This anti-oestrogenic effect presumably arises throughcompetition with the endogenous oestrogens for thereceptor In vitro receptor interaction studies havealso shown that BBP and DEHP possess anti-androgenic activity (Sohoni amp Sumpter 1998) Otherin vivo effects of phthalates in fish include effects onthe metabolic pathways involved in steroid biosyn-thesis and metabolism In a study by Thibaut ampPorte (2004) DEHP and DBP altered the activity ofenzymes involved in the synthesis of endogenoussteroid hormones and their metabolism in carp effectsthat can lead to the alteration of the balance betweenendogenous oestrogens and androgens and thusalterations in their reproductive physiology Thesefindings are supported by studies in mammals inwhich phthalates have also been shown to alter the
Phil Trans R Soc B (2009)
expression and enzymatic activity of enzymes involvedin the synthesis and metabolism of sex steroids in thetestis (Liu et al 2005) In fish exposures to lowmg l21 concentrations of DEP have been shown toinduce alterations in a number of enzymes in theliver and muscle including phosphatases and trans-aminases showing a disruptive effect on generalmetabolic functions (Barse et al 2007) Other studieshave also reported that potentially the most relevantmechanism of action of phthalates is via binding tothe peroxisome proliferator-activated receptors(PPARs) and modulation of the transcriptionalevents coordinated by this molecular pathway whichincludes steroid hormone metabolism (Fan et al2004)
Exposures to phthalates have also been shown toalter behaviour in fish Exposure to 100 mg BBP l21via the water caused alterations in shoaling and feedingbehaviour in three-spined stickleback (Gasterosteusaculeatus) (Wibe et al 2002 2004) and exposure to5 mg DEP l21 caused alterations in the general behaviourof common carp (Barse et al 2007) It should be notedthat these studies have employed very high exposureconcentrations and these are unlikely to occur in thewater column in part owing to their low solubilityExceptions to this may be for fish living incloselyassociated with the sediments in heavily contaminatedenvironments Exposures of fish to lower levels ofphthalates have generally found no adverse effects Asan example exposure of Japanese medaka to 219 and193 mg g21 DINP and DIDP respectively via thediet with a daily feeding regime of 5 per cent of thebody weight failed to identify any effects on reproduc-tion (gonad somatic index egg production embryosurvival and sex ratios) growth or survival in amulti-generational study (14 days post fertilization(dpf) to 140 dpf of the F1 generation) (Patyna et al2006) Similarly exposure of medaka to concentrationsup to 5 mg DEHP l21 for 90 days post-hatch did notaffect germ cell development (Metcalfe et al 2001)
In our exposure of adult breeding zebrafish to BBPmeasured concentrations in the tank water were 3 6and 15 mg l21 over the exposure period Exposure toBBP did not affect the concentration of VTG in theplasma at any of the concentrations tested in males(VTG concentrations in the plasma were 1310+2551500+358 948+141 1300+318 1640+220 ng ml21 for the water control solvent control 36 and 15 mg l21 BBP respectively n frac14 9ndash12 malesper treatment group) Similarly there was no evidenceof any chemical effect on the number of eggs spawnedthe total number of eggs spawned per colony over theassessment period ranged between 4109 and 7578with a maximum of 727 eggs spawned by a singlecolony on a single day with a mean number of eggsspawned per female per day (including days with noeggs) of 222+14 213+14 184+08 216+17 285+14 prior to exposure and 195+16270+17 211+12 250+21 325+14 duringthe exposure for the water control solvent control 36 and 15 mg BBP l21 respectively Furthermore therewere no effects of BBP on the mean percentage viabilityof embryos at 8 hpf (figure 1) These results are inagreement with reports in the literature where VTG
time (days)ndash20 ndash10 0 10 20
cum
ulat
ive
num
ber
of v
iabl
e eg
gs (
8 hp
f)
0
100
200
300
400
500
600
700
Figure 1 Danio rerio Cumulative number of viable eggs spawned in colonies during a 20 day period prior to BBP exposure and
a 20 day period during BBP exposure Symbols represent average results for two colonies per treatment filled circle watercontrol open circle solvent control inverted filled triangle 0003 mg l21 BBP open triangle 0006 mg l21 BBP filledsquare 0015 mg l21 BBP
2052 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
induction in males or reproductive effects on femaleshas been induced only at exposure concentrations thatfar exceed the measured concentrations in our studyIn contrast however we found consistent changes inparameters of sperm quality in BBP-exposed male zeb-rafish The solvent adopted (methanol) had a negativeeffect on zebrafish sperm motility compared with thewater control after 20 days of exposure (mean curvi-linear velocity of 14 mm s21 (mean) and 25 mm s21respectively p 005 figure 2a) and BBP exposureresulted in a concentration-dependent enhancementin curvilinear velocity of the solvent-affected spermAt the highest concentration tested (15 mg BBP l21)the average curvilinear velocity of the sperm wasrestored to levels similar to those found in the watercontrol tanks (31 mm s21 figure 2a) Both thenumber of sperm tracked and the linearity of thesperm movement showed patterns similar to thosedescribed for the curvilinear velocity (figure 2bc)
The ability of BBP to reverse the adverse effect ofthe methanol on sperm number and motility is anovel finding and we hypothesize that this may haveresulted from an interaction of BBP with the PPARreceptor which leads to increased oxidative stress inthe testis as demonstrated in mammals for phthalates(reviewed in Corton amp Lapinskas 2005) In the finalstages of sperm development maturation and acti-vation are crucially dependent on the oxidative balancewithin the seminiferous tubes and in the sperm itselfExposure to methanol may have reduced the freeoxygen radicals required to promote sperm maturationand activation and this effect may have been reversedby the increase in free radicals caused by the BBPexposure The ethanol metabolite acetaldehyde hasbeen shown to block the action of PPARs in vitro(Galli et al 2001) suggesting a mechanism by whichsperm motility might be reduced in fish exposed to asolvent Exposure to BBP in the solvent carrier maycounterbalance this effect restoring the normal
Phil Trans R Soc B (2009)
motility of the sperm The increase in oxidative stresswithin the testis as a result of phthalate exposure hasbeen previously observed in mammalian systemsincluding in rats (Kasahara et al 2002) and thiseffect was shown to be mediated through activationof PPAR pathways
(iv) AmphibiansStudies on phthalates in amphibians (reviewed inKloas 2002) are less extensive than those in fish andsome invertebrates The effects of plasticizers in amp-hibians have been focused on their toxicity theirpotential to induce teratogenic effects and their endo-crine-disrupting activities most notably effects onreproduction sexual differentiation and the thyroidsystem Focusing on toxicity Larsson amp Thuren(1987) found that exposure of Rana arvalis eggs toDEHP via sediment decreased successful hatchingswith increasing concentrations and deduced a noobserved effect concentration (NOEC) of 10 mg kg21
fresh weight Studies with DBP using the frog embryoteratogenesis assay Xenopus (FETAX) have showndevelopmental malformations at 500 mg l21 andenhanced mortality rates at concentrations of 10 and15 mg l21 (Lee et al 2005)
DBP has also been shown to affect sexual differen-tiation in male Rana rugosa tadpoles delaying gonadaldevelopment (Ohtani et al 2000) and altering thespermatogenesis process in eight-month-old X laevisafter exposure during larval development (at 1ndash10 mg DBP l21 from stage 52 to completion of meta-morphosis Lee amp Veeramachaneni 2005) Studies inXenopus investigating the potential for phthalates tobind to the ER via competitive displacement exper-iments with cytosolic liver preparations have foundthat DEP has only a moderate affinity for the ER(Lutz amp Kloas 1999) Parallel in vitro studies using Xlaevis primary hepatocytes however have shown that
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

2050 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
focused on bioaccumulation and acute toxicity ofphthalates but more recently wider biological effectshave been shown including mitotic inhibition induc-tion of chromosomal aberrations and effects on larvaldevelopment
Considering soil-dwelling organisms studies on theuptake and toxicity of the phthalates dimethyl phthalate(DMP) diethyl phthalate (DEP) DBP and DEHP (ata concentration of 5 mg kg21 for 25 days) in the wormEisenia fetida have shown that high molecular weightphthalates (DBP and DEHP) were incorporated butwith relatively low accumulation factors (0242ndash0307and 0073ndash0244 respectively) and DMP and DEPwere apparently not taken up (Xiao-yu et al 2005) Itis possible that the lack of any uptake of DMP andDEP may be as a consequence of their higher microbialdegradation rates and a faster metabolic degradation(Albro et al 1993) Neuhauser et al (1986) investigatedthe acute toxicity of phthalates and found similar LC50
values (1064ndash3335 mg kg21 for a 14 day soil exposure)for DMP in four soil-dwelling annelid species(Allolobophora tuberculata E fetida Eudrilus eugeniaePerionyx excavatus) Together these studies demonstratethat phthalates enter terrestrial worms but they have alow acute toxicity It is worth emphasizing that soils canreceive very high inputs of phthalates and the totalamount of DEHP deposited in US landfills to 1992has been estimated at 5 million metric tonnes
Comparisons of the acute toxicity of severalphthalate esters [DMP DEP DBP BBP di-n-hexyl-phthalate (DHP) and DEHP] in various aquatic organ-isms including the annelid Lumbriculus variegatus haveshown that the higher molecular weight phthalatesDHP and DEHP were not acutely toxic for the concen-tration range tested (23ndash478 mg DHP l21 102ndash691 mg DEHP l21 Call et al 2001) The toxicity oflower molecular weight phthalates was positively corre-lated with their Kow indicating non-polar narcosis asthe underlying mode of action The same study foundL variegatus was the species least sensitive to DMPthe 10 day LC50 (10 day) was 246 compared with281 and 682 mg l21 for the crustacean Hyalellaazteca and the insect Chironomus tentans respectivelyIn adult marine tubeworms Pomatoceros lamarckiiexposure to DMP was shown to decrease fertilizationsuccess at a threshold concentration of 11025 MDMP (194 mg l21) and induce a significant increasein the number of aberrations in chromosome separ-ations in oocytes at anaphase at concentrations11027 M (194 mg l21) (Dixon et al 1999 Wilsonet al 2002) DMP has been shown to affect larvaldevelopment in the polychaete worm at a thresholdconcentration 1 1025 M (194 mg l21)
The effects of phthalates have also been studied onvarious biochemical pathways such as antioxidant andperoxisomal marker enzymes in marine mussels(Mytilus galloprovincialis Orbea et al 2002 M edulisCajaraville amp Ortiz-Zarragoitia 2006) Exposure to500 mg DEHP l21 for 21 days was shown to result ina significant increase in the catalase and acyl-CoAoxidase (AOX) activity and an inhibition of the super-oxide and manganate superoxide dismutase inM galloprovincialis The peroxisomal volume densityin the digestive gland of M edulis was significantly
Phil Trans R Soc B (2009)
enhanced at 50 mg diallylphthalate (DAP) per litreafter three weeks exposure whereas the chemicalhad no significant effect on AOX activity in thisspecies (Cajaraville amp Ortiz-Zarragoitia 2006) DAPwas able to decrease the phospho-protein level (amussel VTG-like protein) but did not impair ovarianfollicles oocytes and spermatogenesis in M eduliswhen exposed to 50 mg DAP l21 for three weeks(Aarab et al 2006) In another 21 day study DAPwas also shown to increase micronuclei frequency amarker of genotoxicity and fragmented-apoptoticcells in the gills of M edulis at 50 mg DAP l21
(Barsiene et al 2006)Very few studies have considered the effects of
phthalates on annelid or mollusc populations butTagatz et al (1986) found that exposure to DBPaltered the community structure and colonizationprofile of the zoomacrobenthos under laboratory andfield conditions The densities of molluscs andechinoderms were reduced by 49 and 97 per centwhile no effects were seen in annelids arthropodschordates and other groups (at concentrations of10ndash1000 mg kg21 over eight weeks)
(ii) Crustaceans and insectsLower molecular weight phthalates that are relativelywater soluble (Howard et al 1985) have been shownto have a lower acute and chronic toxicity in Daphniamagna For example 48 h immobilization tests withD magna have shown IC50 values of 284 220 and678 mg l21 for DMP DEP and DBP respectively(Jonsson amp Baun 2003) Acute toxicity for phthalateshowever differs significantly for different species ofcrustaceans Hyalella azteca is 10ndash20-fold more sensi-tive than D magna although the higher molecularweight phthalates DHP and DEHP did not exhibittoxic effects in H azteca even at concentrations up to225 mg DHP l21 and 590 mg DEHP l21 (Call et al2001) Other effects of DBP reported in crustaceansinclude decreased locomotor activity in Gammaruspulex at an exposure concentration of 500 mg l21
(Thuren amp Woin 1991)In the Drosophila melanogaster BII cell line DEP
exhibited a weak ecdysteroid antagonistic activity(Dinan et al 2001) However the concentrationsrequired to cause an effect were several orders ofmagnitude higher than that for 20-hydroxy ecdysonethus classifying DEP as a very weak environmentalanti-ecdysteroid only
DEHP has been shown to induce sublethal effectsin Chironomus riparius and the exposure of larvaeresulted in an increased female body volume atexposure concentrations of 03 mg l21 (Kwak amp Lee2005) It also increased the expression of heat shockprotein (HSP) and haemoglobin genes in exposedlarvae of C tentans at a concentration of 05 mg l21
(Lee et al 2006a) but the same compound was notacutely toxic to this species at concentrations up to691 mg DEHP l21 (Call et al 2001) The significanceof such effects on gene expression is not known but invertebrates HSPs are involved in steroid hormonereceptor stabilization and activation (Gillesby ampZacharewski 1998) and if they act similarly in
Review Effects of plasticizers on wildlife J Oehlmann et al 2051
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
arthropods a change in the HSP70 titre might resultin a disruption of the ecdysone axis by influencingthe ecdysone receptor
(iii) FishGiven the widespread occurrence of phthalates in theaquatic environment fish are also likely to be exposedto phthalates via the water column food andor viathe sediments depending on their ecological nicheIn support of this a study in wild fish inThe Netherlands measured median concentrations ofDEHP and DBP at 17 and 10 mg kg21 (wet wt)respectively with 95th percentile values reaching upto 141 mg DEHP kg21 and 26 mg DBP kg21 (wet wt)(Peijnenburg amp Struijs 2006) Laboratory studies havefurther shown that phthalates bioconcentrate in fishFor example the BCF for DEHP was 120 in the carpand for BBP 94 in bluegill sunfish (Staples et al1997) The presence of phthalates in wild fish andtheir ability to concentrate in body tissues demonstratetheir potential to induce adverse effects on the health ofwild populations but there has been little bearing onthis expectation
Very few studies have comprehensively addressedthe biological effects of phthalates on fish and invivo studies have reported mixed findings BBP hasbeen reported to be oestrogenic and induces VTG syn-thesis in male fathead minnow (Pimephales promelas) ata concentration of 100 mg l21 via the water (Harrieset al 2000) and similarly to induce VTG in carp(Cyprinus carpio) exposed to 1ndash20 mg DEP l21 overa period of four weeks (Barse et al 2007) In salmon(Salmo salar) exposure to 1500 mg DEHP kg21 inthe food for a prolonged period (four months)during early life resulted in a small incidence of inter-sex (Norman et al 2007) These in vivo effects areconsistent with the mode of action of BBP andDEHP established in vitro where they have beenshown to bind to the oestrogen receptor (ER) andelicit a weakly oestrogenic response in fish cell cultures(Jobling et al 1995) and in the yeast oestrogen screen(YES containing the human ER) (Harris et al 1997)In female Japanese medaka (Oryzias latipes) exposureto concentrations in excess of 1 mg DEHP l21 caused amarked reduction in both VTG and in the percentageof females containing mature oocytes suggesting ananti-oestrogenic mode of action (Kim et al 2002)This anti-oestrogenic effect presumably arises throughcompetition with the endogenous oestrogens for thereceptor In vitro receptor interaction studies havealso shown that BBP and DEHP possess anti-androgenic activity (Sohoni amp Sumpter 1998) Otherin vivo effects of phthalates in fish include effects onthe metabolic pathways involved in steroid biosyn-thesis and metabolism In a study by Thibaut ampPorte (2004) DEHP and DBP altered the activity ofenzymes involved in the synthesis of endogenoussteroid hormones and their metabolism in carp effectsthat can lead to the alteration of the balance betweenendogenous oestrogens and androgens and thusalterations in their reproductive physiology Thesefindings are supported by studies in mammals inwhich phthalates have also been shown to alter the
Phil Trans R Soc B (2009)
expression and enzymatic activity of enzymes involvedin the synthesis and metabolism of sex steroids in thetestis (Liu et al 2005) In fish exposures to lowmg l21 concentrations of DEP have been shown toinduce alterations in a number of enzymes in theliver and muscle including phosphatases and trans-aminases showing a disruptive effect on generalmetabolic functions (Barse et al 2007) Other studieshave also reported that potentially the most relevantmechanism of action of phthalates is via binding tothe peroxisome proliferator-activated receptors(PPARs) and modulation of the transcriptionalevents coordinated by this molecular pathway whichincludes steroid hormone metabolism (Fan et al2004)
Exposures to phthalates have also been shown toalter behaviour in fish Exposure to 100 mg BBP l21via the water caused alterations in shoaling and feedingbehaviour in three-spined stickleback (Gasterosteusaculeatus) (Wibe et al 2002 2004) and exposure to5 mg DEP l21 caused alterations in the general behaviourof common carp (Barse et al 2007) It should be notedthat these studies have employed very high exposureconcentrations and these are unlikely to occur in thewater column in part owing to their low solubilityExceptions to this may be for fish living incloselyassociated with the sediments in heavily contaminatedenvironments Exposures of fish to lower levels ofphthalates have generally found no adverse effects Asan example exposure of Japanese medaka to 219 and193 mg g21 DINP and DIDP respectively via thediet with a daily feeding regime of 5 per cent of thebody weight failed to identify any effects on reproduc-tion (gonad somatic index egg production embryosurvival and sex ratios) growth or survival in amulti-generational study (14 days post fertilization(dpf) to 140 dpf of the F1 generation) (Patyna et al2006) Similarly exposure of medaka to concentrationsup to 5 mg DEHP l21 for 90 days post-hatch did notaffect germ cell development (Metcalfe et al 2001)
In our exposure of adult breeding zebrafish to BBPmeasured concentrations in the tank water were 3 6and 15 mg l21 over the exposure period Exposure toBBP did not affect the concentration of VTG in theplasma at any of the concentrations tested in males(VTG concentrations in the plasma were 1310+2551500+358 948+141 1300+318 1640+220 ng ml21 for the water control solvent control 36 and 15 mg l21 BBP respectively n frac14 9ndash12 malesper treatment group) Similarly there was no evidenceof any chemical effect on the number of eggs spawnedthe total number of eggs spawned per colony over theassessment period ranged between 4109 and 7578with a maximum of 727 eggs spawned by a singlecolony on a single day with a mean number of eggsspawned per female per day (including days with noeggs) of 222+14 213+14 184+08 216+17 285+14 prior to exposure and 195+16270+17 211+12 250+21 325+14 duringthe exposure for the water control solvent control 36 and 15 mg BBP l21 respectively Furthermore therewere no effects of BBP on the mean percentage viabilityof embryos at 8 hpf (figure 1) These results are inagreement with reports in the literature where VTG
time (days)ndash20 ndash10 0 10 20
cum
ulat
ive
num
ber
of v
iabl
e eg
gs (
8 hp
f)
0
100
200
300
400
500
600
700
Figure 1 Danio rerio Cumulative number of viable eggs spawned in colonies during a 20 day period prior to BBP exposure and
a 20 day period during BBP exposure Symbols represent average results for two colonies per treatment filled circle watercontrol open circle solvent control inverted filled triangle 0003 mg l21 BBP open triangle 0006 mg l21 BBP filledsquare 0015 mg l21 BBP
2052 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
induction in males or reproductive effects on femaleshas been induced only at exposure concentrations thatfar exceed the measured concentrations in our studyIn contrast however we found consistent changes inparameters of sperm quality in BBP-exposed male zeb-rafish The solvent adopted (methanol) had a negativeeffect on zebrafish sperm motility compared with thewater control after 20 days of exposure (mean curvi-linear velocity of 14 mm s21 (mean) and 25 mm s21respectively p 005 figure 2a) and BBP exposureresulted in a concentration-dependent enhancementin curvilinear velocity of the solvent-affected spermAt the highest concentration tested (15 mg BBP l21)the average curvilinear velocity of the sperm wasrestored to levels similar to those found in the watercontrol tanks (31 mm s21 figure 2a) Both thenumber of sperm tracked and the linearity of thesperm movement showed patterns similar to thosedescribed for the curvilinear velocity (figure 2bc)
The ability of BBP to reverse the adverse effect ofthe methanol on sperm number and motility is anovel finding and we hypothesize that this may haveresulted from an interaction of BBP with the PPARreceptor which leads to increased oxidative stress inthe testis as demonstrated in mammals for phthalates(reviewed in Corton amp Lapinskas 2005) In the finalstages of sperm development maturation and acti-vation are crucially dependent on the oxidative balancewithin the seminiferous tubes and in the sperm itselfExposure to methanol may have reduced the freeoxygen radicals required to promote sperm maturationand activation and this effect may have been reversedby the increase in free radicals caused by the BBPexposure The ethanol metabolite acetaldehyde hasbeen shown to block the action of PPARs in vitro(Galli et al 2001) suggesting a mechanism by whichsperm motility might be reduced in fish exposed to asolvent Exposure to BBP in the solvent carrier maycounterbalance this effect restoring the normal
Phil Trans R Soc B (2009)
motility of the sperm The increase in oxidative stresswithin the testis as a result of phthalate exposure hasbeen previously observed in mammalian systemsincluding in rats (Kasahara et al 2002) and thiseffect was shown to be mediated through activationof PPAR pathways
(iv) AmphibiansStudies on phthalates in amphibians (reviewed inKloas 2002) are less extensive than those in fish andsome invertebrates The effects of plasticizers in amp-hibians have been focused on their toxicity theirpotential to induce teratogenic effects and their endo-crine-disrupting activities most notably effects onreproduction sexual differentiation and the thyroidsystem Focusing on toxicity Larsson amp Thuren(1987) found that exposure of Rana arvalis eggs toDEHP via sediment decreased successful hatchingswith increasing concentrations and deduced a noobserved effect concentration (NOEC) of 10 mg kg21
fresh weight Studies with DBP using the frog embryoteratogenesis assay Xenopus (FETAX) have showndevelopmental malformations at 500 mg l21 andenhanced mortality rates at concentrations of 10 and15 mg l21 (Lee et al 2005)
DBP has also been shown to affect sexual differen-tiation in male Rana rugosa tadpoles delaying gonadaldevelopment (Ohtani et al 2000) and altering thespermatogenesis process in eight-month-old X laevisafter exposure during larval development (at 1ndash10 mg DBP l21 from stage 52 to completion of meta-morphosis Lee amp Veeramachaneni 2005) Studies inXenopus investigating the potential for phthalates tobind to the ER via competitive displacement exper-iments with cytosolic liver preparations have foundthat DEP has only a moderate affinity for the ER(Lutz amp Kloas 1999) Parallel in vitro studies using Xlaevis primary hepatocytes however have shown that
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

Review Effects of plasticizers on wildlife J Oehlmann et al 2051
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
arthropods a change in the HSP70 titre might resultin a disruption of the ecdysone axis by influencingthe ecdysone receptor
(iii) FishGiven the widespread occurrence of phthalates in theaquatic environment fish are also likely to be exposedto phthalates via the water column food andor viathe sediments depending on their ecological nicheIn support of this a study in wild fish inThe Netherlands measured median concentrations ofDEHP and DBP at 17 and 10 mg kg21 (wet wt)respectively with 95th percentile values reaching upto 141 mg DEHP kg21 and 26 mg DBP kg21 (wet wt)(Peijnenburg amp Struijs 2006) Laboratory studies havefurther shown that phthalates bioconcentrate in fishFor example the BCF for DEHP was 120 in the carpand for BBP 94 in bluegill sunfish (Staples et al1997) The presence of phthalates in wild fish andtheir ability to concentrate in body tissues demonstratetheir potential to induce adverse effects on the health ofwild populations but there has been little bearing onthis expectation
Very few studies have comprehensively addressedthe biological effects of phthalates on fish and invivo studies have reported mixed findings BBP hasbeen reported to be oestrogenic and induces VTG syn-thesis in male fathead minnow (Pimephales promelas) ata concentration of 100 mg l21 via the water (Harrieset al 2000) and similarly to induce VTG in carp(Cyprinus carpio) exposed to 1ndash20 mg DEP l21 overa period of four weeks (Barse et al 2007) In salmon(Salmo salar) exposure to 1500 mg DEHP kg21 inthe food for a prolonged period (four months)during early life resulted in a small incidence of inter-sex (Norman et al 2007) These in vivo effects areconsistent with the mode of action of BBP andDEHP established in vitro where they have beenshown to bind to the oestrogen receptor (ER) andelicit a weakly oestrogenic response in fish cell cultures(Jobling et al 1995) and in the yeast oestrogen screen(YES containing the human ER) (Harris et al 1997)In female Japanese medaka (Oryzias latipes) exposureto concentrations in excess of 1 mg DEHP l21 caused amarked reduction in both VTG and in the percentageof females containing mature oocytes suggesting ananti-oestrogenic mode of action (Kim et al 2002)This anti-oestrogenic effect presumably arises throughcompetition with the endogenous oestrogens for thereceptor In vitro receptor interaction studies havealso shown that BBP and DEHP possess anti-androgenic activity (Sohoni amp Sumpter 1998) Otherin vivo effects of phthalates in fish include effects onthe metabolic pathways involved in steroid biosyn-thesis and metabolism In a study by Thibaut ampPorte (2004) DEHP and DBP altered the activity ofenzymes involved in the synthesis of endogenoussteroid hormones and their metabolism in carp effectsthat can lead to the alteration of the balance betweenendogenous oestrogens and androgens and thusalterations in their reproductive physiology Thesefindings are supported by studies in mammals inwhich phthalates have also been shown to alter the
Phil Trans R Soc B (2009)
expression and enzymatic activity of enzymes involvedin the synthesis and metabolism of sex steroids in thetestis (Liu et al 2005) In fish exposures to lowmg l21 concentrations of DEP have been shown toinduce alterations in a number of enzymes in theliver and muscle including phosphatases and trans-aminases showing a disruptive effect on generalmetabolic functions (Barse et al 2007) Other studieshave also reported that potentially the most relevantmechanism of action of phthalates is via binding tothe peroxisome proliferator-activated receptors(PPARs) and modulation of the transcriptionalevents coordinated by this molecular pathway whichincludes steroid hormone metabolism (Fan et al2004)
Exposures to phthalates have also been shown toalter behaviour in fish Exposure to 100 mg BBP l21via the water caused alterations in shoaling and feedingbehaviour in three-spined stickleback (Gasterosteusaculeatus) (Wibe et al 2002 2004) and exposure to5 mg DEP l21 caused alterations in the general behaviourof common carp (Barse et al 2007) It should be notedthat these studies have employed very high exposureconcentrations and these are unlikely to occur in thewater column in part owing to their low solubilityExceptions to this may be for fish living incloselyassociated with the sediments in heavily contaminatedenvironments Exposures of fish to lower levels ofphthalates have generally found no adverse effects Asan example exposure of Japanese medaka to 219 and193 mg g21 DINP and DIDP respectively via thediet with a daily feeding regime of 5 per cent of thebody weight failed to identify any effects on reproduc-tion (gonad somatic index egg production embryosurvival and sex ratios) growth or survival in amulti-generational study (14 days post fertilization(dpf) to 140 dpf of the F1 generation) (Patyna et al2006) Similarly exposure of medaka to concentrationsup to 5 mg DEHP l21 for 90 days post-hatch did notaffect germ cell development (Metcalfe et al 2001)
In our exposure of adult breeding zebrafish to BBPmeasured concentrations in the tank water were 3 6and 15 mg l21 over the exposure period Exposure toBBP did not affect the concentration of VTG in theplasma at any of the concentrations tested in males(VTG concentrations in the plasma were 1310+2551500+358 948+141 1300+318 1640+220 ng ml21 for the water control solvent control 36 and 15 mg l21 BBP respectively n frac14 9ndash12 malesper treatment group) Similarly there was no evidenceof any chemical effect on the number of eggs spawnedthe total number of eggs spawned per colony over theassessment period ranged between 4109 and 7578with a maximum of 727 eggs spawned by a singlecolony on a single day with a mean number of eggsspawned per female per day (including days with noeggs) of 222+14 213+14 184+08 216+17 285+14 prior to exposure and 195+16270+17 211+12 250+21 325+14 duringthe exposure for the water control solvent control 36 and 15 mg BBP l21 respectively Furthermore therewere no effects of BBP on the mean percentage viabilityof embryos at 8 hpf (figure 1) These results are inagreement with reports in the literature where VTG
time (days)ndash20 ndash10 0 10 20
cum
ulat
ive
num
ber
of v
iabl
e eg
gs (
8 hp
f)
0
100
200
300
400
500
600
700
Figure 1 Danio rerio Cumulative number of viable eggs spawned in colonies during a 20 day period prior to BBP exposure and
a 20 day period during BBP exposure Symbols represent average results for two colonies per treatment filled circle watercontrol open circle solvent control inverted filled triangle 0003 mg l21 BBP open triangle 0006 mg l21 BBP filledsquare 0015 mg l21 BBP
2052 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
induction in males or reproductive effects on femaleshas been induced only at exposure concentrations thatfar exceed the measured concentrations in our studyIn contrast however we found consistent changes inparameters of sperm quality in BBP-exposed male zeb-rafish The solvent adopted (methanol) had a negativeeffect on zebrafish sperm motility compared with thewater control after 20 days of exposure (mean curvi-linear velocity of 14 mm s21 (mean) and 25 mm s21respectively p 005 figure 2a) and BBP exposureresulted in a concentration-dependent enhancementin curvilinear velocity of the solvent-affected spermAt the highest concentration tested (15 mg BBP l21)the average curvilinear velocity of the sperm wasrestored to levels similar to those found in the watercontrol tanks (31 mm s21 figure 2a) Both thenumber of sperm tracked and the linearity of thesperm movement showed patterns similar to thosedescribed for the curvilinear velocity (figure 2bc)
The ability of BBP to reverse the adverse effect ofthe methanol on sperm number and motility is anovel finding and we hypothesize that this may haveresulted from an interaction of BBP with the PPARreceptor which leads to increased oxidative stress inthe testis as demonstrated in mammals for phthalates(reviewed in Corton amp Lapinskas 2005) In the finalstages of sperm development maturation and acti-vation are crucially dependent on the oxidative balancewithin the seminiferous tubes and in the sperm itselfExposure to methanol may have reduced the freeoxygen radicals required to promote sperm maturationand activation and this effect may have been reversedby the increase in free radicals caused by the BBPexposure The ethanol metabolite acetaldehyde hasbeen shown to block the action of PPARs in vitro(Galli et al 2001) suggesting a mechanism by whichsperm motility might be reduced in fish exposed to asolvent Exposure to BBP in the solvent carrier maycounterbalance this effect restoring the normal
Phil Trans R Soc B (2009)
motility of the sperm The increase in oxidative stresswithin the testis as a result of phthalate exposure hasbeen previously observed in mammalian systemsincluding in rats (Kasahara et al 2002) and thiseffect was shown to be mediated through activationof PPAR pathways
(iv) AmphibiansStudies on phthalates in amphibians (reviewed inKloas 2002) are less extensive than those in fish andsome invertebrates The effects of plasticizers in amp-hibians have been focused on their toxicity theirpotential to induce teratogenic effects and their endo-crine-disrupting activities most notably effects onreproduction sexual differentiation and the thyroidsystem Focusing on toxicity Larsson amp Thuren(1987) found that exposure of Rana arvalis eggs toDEHP via sediment decreased successful hatchingswith increasing concentrations and deduced a noobserved effect concentration (NOEC) of 10 mg kg21
fresh weight Studies with DBP using the frog embryoteratogenesis assay Xenopus (FETAX) have showndevelopmental malformations at 500 mg l21 andenhanced mortality rates at concentrations of 10 and15 mg l21 (Lee et al 2005)
DBP has also been shown to affect sexual differen-tiation in male Rana rugosa tadpoles delaying gonadaldevelopment (Ohtani et al 2000) and altering thespermatogenesis process in eight-month-old X laevisafter exposure during larval development (at 1ndash10 mg DBP l21 from stage 52 to completion of meta-morphosis Lee amp Veeramachaneni 2005) Studies inXenopus investigating the potential for phthalates tobind to the ER via competitive displacement exper-iments with cytosolic liver preparations have foundthat DEP has only a moderate affinity for the ER(Lutz amp Kloas 1999) Parallel in vitro studies using Xlaevis primary hepatocytes however have shown that
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)
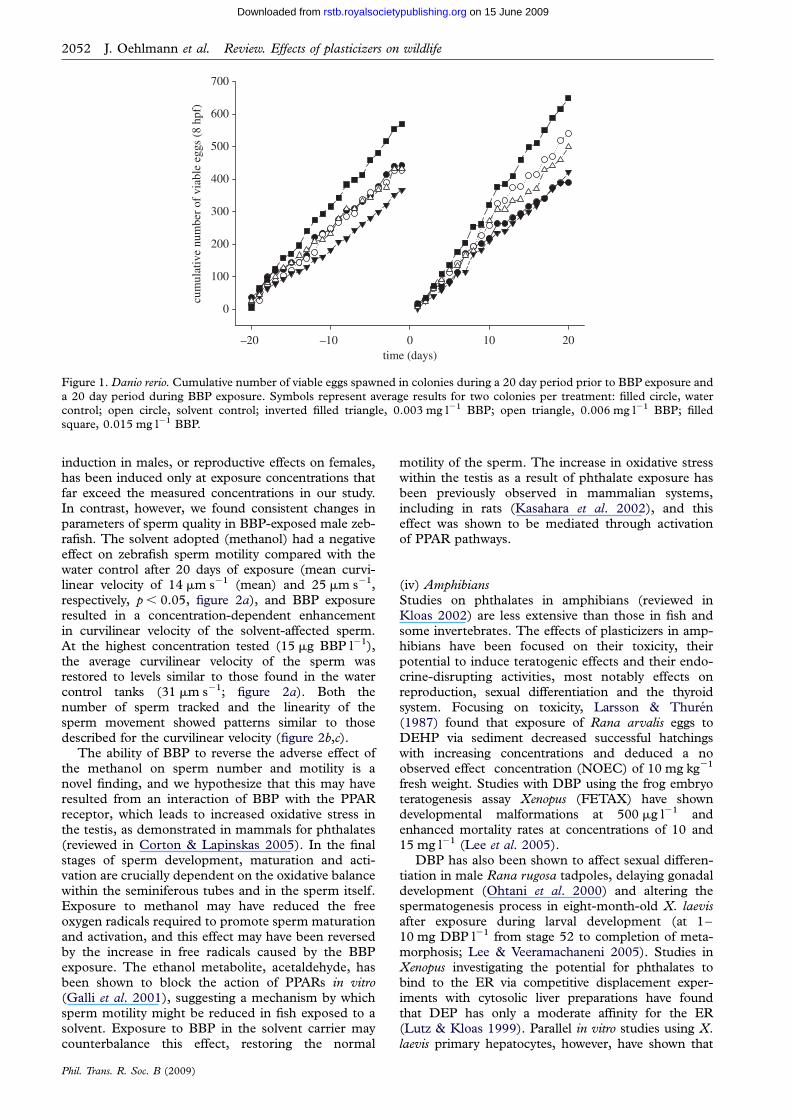
time (days)ndash20 ndash10 0 10 20
cum
ulat
ive
num
ber
of v
iabl
e eg
gs (
8 hp
f)
0
100
200
300
400
500
600
700
Figure 1 Danio rerio Cumulative number of viable eggs spawned in colonies during a 20 day period prior to BBP exposure and
a 20 day period during BBP exposure Symbols represent average results for two colonies per treatment filled circle watercontrol open circle solvent control inverted filled triangle 0003 mg l21 BBP open triangle 0006 mg l21 BBP filledsquare 0015 mg l21 BBP
2052 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
induction in males or reproductive effects on femaleshas been induced only at exposure concentrations thatfar exceed the measured concentrations in our studyIn contrast however we found consistent changes inparameters of sperm quality in BBP-exposed male zeb-rafish The solvent adopted (methanol) had a negativeeffect on zebrafish sperm motility compared with thewater control after 20 days of exposure (mean curvi-linear velocity of 14 mm s21 (mean) and 25 mm s21respectively p 005 figure 2a) and BBP exposureresulted in a concentration-dependent enhancementin curvilinear velocity of the solvent-affected spermAt the highest concentration tested (15 mg BBP l21)the average curvilinear velocity of the sperm wasrestored to levels similar to those found in the watercontrol tanks (31 mm s21 figure 2a) Both thenumber of sperm tracked and the linearity of thesperm movement showed patterns similar to thosedescribed for the curvilinear velocity (figure 2bc)
The ability of BBP to reverse the adverse effect ofthe methanol on sperm number and motility is anovel finding and we hypothesize that this may haveresulted from an interaction of BBP with the PPARreceptor which leads to increased oxidative stress inthe testis as demonstrated in mammals for phthalates(reviewed in Corton amp Lapinskas 2005) In the finalstages of sperm development maturation and acti-vation are crucially dependent on the oxidative balancewithin the seminiferous tubes and in the sperm itselfExposure to methanol may have reduced the freeoxygen radicals required to promote sperm maturationand activation and this effect may have been reversedby the increase in free radicals caused by the BBPexposure The ethanol metabolite acetaldehyde hasbeen shown to block the action of PPARs in vitro(Galli et al 2001) suggesting a mechanism by whichsperm motility might be reduced in fish exposed to asolvent Exposure to BBP in the solvent carrier maycounterbalance this effect restoring the normal
Phil Trans R Soc B (2009)
motility of the sperm The increase in oxidative stresswithin the testis as a result of phthalate exposure hasbeen previously observed in mammalian systemsincluding in rats (Kasahara et al 2002) and thiseffect was shown to be mediated through activationof PPAR pathways
(iv) AmphibiansStudies on phthalates in amphibians (reviewed inKloas 2002) are less extensive than those in fish andsome invertebrates The effects of plasticizers in amp-hibians have been focused on their toxicity theirpotential to induce teratogenic effects and their endo-crine-disrupting activities most notably effects onreproduction sexual differentiation and the thyroidsystem Focusing on toxicity Larsson amp Thuren(1987) found that exposure of Rana arvalis eggs toDEHP via sediment decreased successful hatchingswith increasing concentrations and deduced a noobserved effect concentration (NOEC) of 10 mg kg21
fresh weight Studies with DBP using the frog embryoteratogenesis assay Xenopus (FETAX) have showndevelopmental malformations at 500 mg l21 andenhanced mortality rates at concentrations of 10 and15 mg l21 (Lee et al 2005)
DBP has also been shown to affect sexual differen-tiation in male Rana rugosa tadpoles delaying gonadaldevelopment (Ohtani et al 2000) and altering thespermatogenesis process in eight-month-old X laevisafter exposure during larval development (at 1ndash10 mg DBP l21 from stage 52 to completion of meta-morphosis Lee amp Veeramachaneni 2005) Studies inXenopus investigating the potential for phthalates tobind to the ER via competitive displacement exper-iments with cytosolic liver preparations have foundthat DEP has only a moderate affinity for the ER(Lutz amp Kloas 1999) Parallel in vitro studies using Xlaevis primary hepatocytes however have shown that
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)
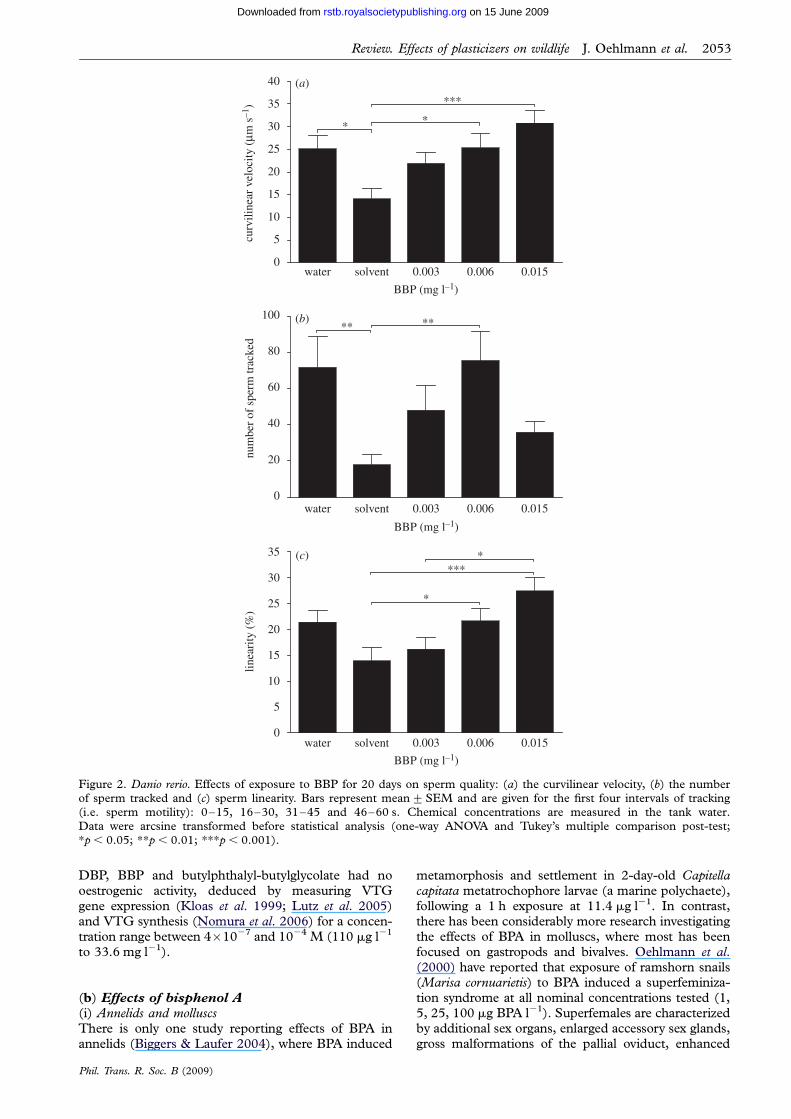
40
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
curv
iline
ar v
eloc
ity (
m s
ndash1)
100
80
60
40
20
0water solvent 0003
BBP (mg lndash1)
0006 0015
num
ber
of s
perm
trac
ked
35
30
25
20
15
10
5
0water solvent 0003
BBP (mg lndash1)
0006 0015
linea
rity
(
)
(a)
(b)
(c)
Figure 2 Danio rerio Effects of exposure to BBP for 20 days on sperm quality (a) the curvilinear velocity (b) the numberof sperm tracked and (c) sperm linearity Bars represent mean+SEM and are given for the first four intervals of tracking(ie sperm motility) 0ndash15 16ndash30 31ndash45 and 46ndash60 s Chemical concentrations are measured in the tank water
Data were arcsine transformed before statistical analysis (one-way ANOVA and Tukeyrsquos multiple comparison post-testp 005 p 001 p 0001)
Review Effects of plasticizers on wildlife J Oehlmann et al 2053
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
DBP BBP and butylphthalyl-butylglycolate had nooestrogenic activity deduced by measuring VTGgene expression (Kloas et al 1999 Lutz et al 2005)and VTG synthesis (Nomura et al 2006) for a concen-tration range between 41027 and 1024 M (110 mg l21
to 336 mg l21)
(b) Effects of bisphenol A
(i) Annelids and molluscsThere is only one study reporting effects of BPA inannelids (Biggers amp Laufer 2004) where BPA induced
Phil Trans R Soc B (2009)
metamorphosis and settlement in 2-day-old Capitellacapitata metatrochophore larvae (a marine polychaete)following a 1 h exposure at 114 mg l21 In contrastthere has been considerably more research investigatingthe effects of BPA in molluscs where most has beenfocused on gastropods and bivalves Oehlmann et al(2000) have reported that exposure of ramshorn snails(Marisa cornuarietis) to BPA induced a superfeminiza-tion syndrome at all nominal concentrations tested (15 25 100 mg BPA l21) Superfemales are characterizedby additional sex organs enlarged accessory sex glandsgross malformations of the pallial oviduct enhanced
2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

2054 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
egg production outside the main spawning season andincreased female mortality (133ndash157 in exposedsnails compared with control mortality of 38)Effects were concentration dependent (except formortality) and statistically significant at every test con-centration Limitations on this first study included thelack of analytical confirmation of the exposure regimesa lack of replication and the absence of a positivecontrol However two follow-up exposures employingthese features confirmed the original findings(Schulte-Oehlmann et al 2001 Oehlmann et al2006) Schulte-Oehlmann et al (2001) investigatedthe effects of BPA at different phases of the reproductivecycle at concentrations of 005 01 025 05 and10 mg l21 over 180 days Before and after the mainspawning season superfemales were observed withoviduct malformations and a significantly increasedegg production in all treated groups except the lowesttest concentration During the peak spawning periodno effect of BPA was seen on spawning levels probablybecause spawning activity in the controls increasedsix-fold at this time masking any effect seen forpre- and post-spawning periods Calculated NOECand EC10 values in this experiment were 79 and139 ng BPA l21 for egg production respectivelybased on measured concentrations in the tanks
Oehlmann et al (2006) demonstrated that theeffects of BPA on reproductive output and oviductmalformations in the ramshorn snail were temperaturedependent Sexually mature snails (2 replicates of 30)were exposed to 025 05 1 and 5 mg BPA l21 for fivemonths outside the main spawning season at either 20or 278C As was seen previously exposure to BPA at208C produced significantly more eggs comparedwith controls The calculated EC10 value was148 ng l21 for egg production supporting the resultsfrom Schulte-Oehlmann et al (2001) In contrast at278C none of the treatment groups produced moreeggs per female compared with controls At this temp-erature however reproductive output in controls wasconsiderably higher probably masking the stimulatoryeffects of BPA that occurred at lower temperatures Incontrast with these findings Forbes et al (2007) foundno effects of BPA on fecundity mortality and hatch-ability in M cornuarietis when exposed to 0 01 1016 160 or 640 mg BPA l21 for 12 weeks Howeverin that study snails were exposed at temperaturesof 25+18C potentially masking the effects ofBPA Furthermore the study also lacked a positivecontrol making accurate interpretations on thesedata difficult
The mud snail Potamopyrgus antipodarum has alsobeen shown to be sensitive to BPA when exposed viasediments with a stimulation in embryo productionat an EC50 of 567 mg kg21 and an EC10 of019 mg kg21 after four weeks of exposure (Duft et al2003) In another study on mud snails exposure viathe water to 1 5 25 and 100 mg BPA l21 also stimu-lated embryo production (and additionally for theethinyloestradiol-positive controls) with significanteffects at a concentration of 5 mg BPA l21 (NOEC1 mg BPA l21 Jobling et al 2004)
Bisphenol A effects have also been studied inbivalves notably Mytilus species In M edulis BPA
Phil Trans R Soc B (2009)
(at 50 mg l21 for three weeks) has been shown toincrease the phospho-protein levels induce spawningin both sexes damage oocytes and ovarian follicles(Aarab et al 2006) and induce increased micronucleifrequency in gill cells (Barsiene et al 2006) Canesiet al (2005) showed that BPA at a nominal concen-tration of 25 nM (57 mg BPA l21) in the haemolymphcaused a significant lysosomal membrane destabiliza-tion in haemocytes in M galloprovincialis Canesiet al (2007a) evaluated the lysosomal membranedestabilization in extracted haemocytes of the samespecies and reported an NOEC of 1 mM and a corre-sponding EC50 value of 345 mM (787 mg l21)Furthermore BPA induced significant changes in thephosphorylation of MAP kinases and STAT factorsresulting in a decrease in phosphorylation of thestress-activated p38 MAPK and CREB-like transcrip-tion factor (cAMP-responsive element bindingprotein) The MAP-kinase pathway is important inembryogenesis cell growth differentiation and apop-tosis and disturbances of STAT signalling favourtumour development by compromising the immunesurveillance Thus the findings of Canesi et al(2007a) may have significant implications for musselhealth Canesi et al (2007b) also reported alteredpatterns of gene expression and activity of enzymesinvolved in the redox balance following an injectionof BPA into the posterior adductor muscle (at nominaldoses of 3 15 and 60 ng BPA g21 dry wt) Inthe digestive gland an increased expression of theMytilus ER (MeER2) was observed at the lowesttest concentration 24 h after injection whereasresults for the higher test concentrations were notsignificant The positive control 17b-oestradiol hadthe same effect on both receptors (MeER1 MeER2)Metallothionein MT20 gene expression was signifi-cantly downregulated catalase GSH transferase andGSSG reductase exhibited a changed activity patternand total glutathione content was enhanced at all testconcentrations Unlike for the phthalate DAP a studyby Cajaraville amp Ortiz-Zarragoitia (2006) found noeffects of BPA on AOX activity or peroxisomalvolume density
(ii) Crustaceans and insectsBisphenol A has been shown to be acutely toxic in thehigh mg l21 range in crustaceans and aquatic insectsIn D magna nominal ICLC50 values vary widely atbetween 240 and 12 800 mg l21 (Chen et al 2002Hirano et al 2004 Park amp Choi 2007) The lowestacute value an IC50 for D magna of 240 mg l21 for24 h (Park amp Choi 2007) is remarkably low comparedwith other studies and also compared with studies foreffects on reproduction in D magna (Brennan et al2006) Differences in chemical sensitivity have beenshown to occur between strains of D magna (Bairdet al 1991) therefore Park amp Choi (2007) may havebeen working with an especially sensitive strainApplying the Comet assay to this sensitive strainDNA breakage occurred at 21 mg BPA l21 (Park ampChoi 2007)
Generally in D magna chronic BPA effects areobserved at concentrations slightly below those that
Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

Review Effects of plasticizers on wildlife J Oehlmann et al 2055
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
are acutely toxic Bisphenol A inhibits reproductionand development rates with an NOEC of1000 mg l21 (Brennan et al 2006) and affected inter-molt duration at 9000 mg l21 (Mu et al 2005) Inthe marine copepods A tonsa and Tigriopus japonicusBPA has been shown to inhibit development at 100and 01 mg l21 respectively (Andersen et al 2001Marcial et al 2003) A significant stimulation of larvaldevelopment in A tonsa was however observed at125ndash50 mg l21 (Mariager 2001) which was also seenin our own experiments at 30 and 60 mg l21 reportedin this paper In 10-day-old A tonsa there was an initialincreased egg production at 20 mg l21 BPA (Andersenet al 1999) possibly indicating an accelerated rate ofmaturation owing to a higher development rate of theearly life stages at this exposure concentration In ourexperiments with fully mature A tonsa (14 days old)egg production was reduced at a lowest observedeffect concentration (LOEC) of 300 mg BPA l21Furthermore in subsequent female offspring derivedfrom exposed parents (to all test concentrations ofBPA down to and including 50 mg l21) the hatchingrate was significantly reduced Mortality in second-gen-eration larvae was also significantly higher comparedwith controls These results indicate a maternal transferof BPA into the developing eggs andor a disruptingeffect during oogenesis In conclusion these datasetsshow that concentrations of BPA between 10 and60 mg l21 in the water stimulated development andgrowth of A tonsa while concentrations of 100 mg l21
and higher inhibited development growth and eggproduction
In studies on C riparius exposed to BPA via spikedsediments emergence of second-generation individualswas delayed (at a water phase concentration of78 ng BPA l21 and higher Watts et al 2001) Thisobservation may have resulted from effects of BPA onthe levels of HSP which has been observed inGammarus fossarum exposed to BPA (Schirling et al2006) In this study a 34 day exposure to 50 and500 mg BPA l21 accelerated maturation of oocytesreduced the number of offspring produced and resultedin reduced levels of HSP However in C tentans theexpression of HSP70 genes has been shown to bestimulated at concentrations down to 8 mg BPA l21
(the lowest concentration applied Lee et al 2006a)In the D melanogaster BII cell line BPA was shown toact as an ecdysteroid antagonist but only at a highexposure concentration of 228 mg l21 (Dinan et al2001) Studies combining BPA with other chemicalshave shown interactive effects affecting the number ofembryo abnormalities and offspring emergence (Muet al 2005 Wang et al 2005) In summary for studiesin aquatic arthropods BPA appears to interfere withtheir hormone systemsmdashpossibly by affecting thesynthesis of HSPs andor by interfering with theecdysone receptor itselfmdashand causes adverse effects atconcentrations in the low mg l21 range
(iii) FishStudies in vitro using the YES have clearly establishedthat BPA has the ability to act as an oestrogenic sub-stance in vertebrates by directly binding and activating
Phil Trans R Soc B (2009)
the ER (Harris et al 1997) In parallel studies using ahuman breast cancer cell line (MCF7) have demon-strated that BPA is able to elicit an oestrogenicresponse similar to that induced by natural oestrogens(Olea et al 1996) In these studies the concentrationsrequired to cause an effect however were severalorders of magnitude higher than that for oestradiolthus classifying BPA as a weak environmental oestrogenin vertebrates
The presence of BPA in rivers and streams at rela-tively high concentrations (up to 21 mg l21 Crainet al 2007) has led to significant research efforts inves-tigating its biological effects in fish Consistent withthese findings BPA has been shown to have feminizingeffects in vivo and to induce VTG andor zona radiataproteins (ZRPs) synthesis in a diverse range of fishspecies (carp 100 mg l21 Mandich et al 2007 fatheadminnow 160 mg l21 Brian et al 2005 Sohoni et al2001 cod 50 mg l21 Larsen et al 2006 medaka1000 mg l21 Ishibashi et al 2005 rainbow trout500 mg l21 Lindholst et al 2001) These concen-trations however have not been reported in the aquaticenvironment In vivo studies have shown that BPAaffects numerous other biological processes and inmany cases at concentrations that have been found inthe aquatic environment These effects include disrup-tions in androgen and oestrogen synthesis and metab-olism Studies in carp have reported that exposure tolow concentrations of BPA (1ndash10 mg l21) decreases theratio of oestrogen to androgen in the plasma whereasexposure to high concentrations (1000 mg l21) increasesit (Mandich et al 2007) Alterations in concentrations ofcirculating steroids have also been reported in juvenileturbot (Psetta maxima) exposed to 59 mg BPA l21
(Labadie amp Budzinski 2006) This effect is likely toresult from alterations in the levels of steroidogenicenzymes or their enzymatic activity catalysing thesynthesis andor metabolism of these hormones andordisplacement of oestradiol from sex steroid-binding glo-bulins (reviewed in Crain et al 2007) The biologicalconsequences of induced changes in the ratio betweenoestrogens and androgens are wide ranging and mayinclude masculinization or feminization of organismsandor alterations in other processes regulated by thesehormones (including growth bone morphogenesisinsulin signalling neural development cell division andapoptosis)
Studies in fish as in some invertebrates have alsoprovided evidence for differential species sensitivi-ties for exposure to BPA For example in exposuresof Atlantic cod (Gadus morhua) and turbot(Scophthalmus maximus) to 59 mg BPA l21 via thewater VTG and ZRP were more highly induced inthe cod than in turbot (Larsen et al 2006) A possiblereason for the differences in species sensitivity is in therate of metabolic transformation of BPA Supportingthis argument BPA has been shown to be metabolizedand eliminated much more rapidly in the zebrafish(D rerio) than in the rainbow trout (Oncorhynchusmykiss) (Lindholst et al 2003)
BPA has also been shown to induce alterations ingonadal development and gamete quality in fishincluding at concentrations found in the environmentThese effects include alterations in the progression of
2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

2056 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
germ cell development in fathead minnow (1 mgBPA l21 Sohoni et al 2001) alterations in the gonadalstructure in male carp (from 1 mg BPA l21) and anenhancement of atresia in oocytes in female carp(1 mg BPA l21 Mandich et al 2007) In addition inbrown trout (Salmo trutta f fario) exposures to BPAhas been shown to cause reduced sperm quality(175ndash240 mg BPA l21) delayed ovulation in females(175 mg BPA l21) and inhibition of ovulation infemales (5 mg BPA l21 Lahnsteiner et al 2005) Theeffects observed for brown trout may have population-level implications as they would lead to a delay in breed-ing with the risk that offspring would be produced atless favourable seasonal periods In carp exposure tovery high levels of BPA (1 mg l21) has also beenshown to induce intersex (Mandich et al 2007)Another reported physiological effect in turbot exposedto BPA (at 50 mg BPA l21 for three weeks) is anincreased micronuclei frequency in erythrocytesdemonstrating the capability of this compound toinduce DNA damage in fish (Bolognesi et al 2006)
Gene expression studies have been employed withgood success for identifying the molecular mechan-isms leading to the biological effects of BPA in fishStudies in Kryptolebias marmoratus (formerly Rivulusm) have shown that exposure to BPA induces changesin the expression of genes associated with oestrogensignalling including cytochrome P450 aromatase Aand B and ERa but not ERb or androgen receptor(Lee et al 2006b Seo et al 2006) Similarly BPAhas been shown to induce the expression of ERa inthe anal fin of Japanese medaka (O latipes) (Hayashiet al 2007) Studies on carp (C carpio) exposed toBPA employing gene arrays have reported alterationsin the expression of many of the same genes inducedby exposure to natural and synthetic oestrogens (oes-tradiol and ethinyloestradiol) but also some uniquegenes too (Moens et al 2006 2007) Studies onK marmoratus following exposure to BPA also demon-strated differential expression of large sets of genesand in particular genes belonging to the GeneOntologies representing catalytic activity and binding(Lee et al 2007) These studies however have gener-ally employed high BPA exposure concentrationsBPA has also been shown to modulate thesomatostatin system in the brain of teleosts resultingin deregulation of neuro-hormonal functions linkedto the reproductive system (Alo et al 2005) Takentogether these molecular mechanistic studies high-light that BPA appears to act predominantly but notexclusively through oestrogen signalling pathways inexerting its biological effects on fish
The biological effects induced in laboratoryexposures to BPA are consistent with some phenotypesobserved in wild fish living in waters receiving highlevels of effluent discharges and containing relativelyhigh concentrations of BPA Although BPA does notappear to bioconcentrate in fish and can degraderapidly in the environment (Staples et al 1998) itcan be metabolized into biologically more active com-pounds (Ishibashi et al 2005) Overall the availabledatasets support the contention that BPA may contrib-ute to adverse reproductive health effects in wild fishpopulations inhabiting contaminated sites
Phil Trans R Soc B (2009)
(iv) AmphibiansBPA has been shown to induce a wide range of adverseeffects in amphibians Teratogenic effects have beeninduced in X laevis embryos exposed from stage 6onwards (Oka et al 2003) and they include crookedvertebrae abnormal development of head and abdo-men to stage 40 (at an exposure of 2 1025 M(46 mg l21)) as well as apoptosis in the centralnervous system When exposures are started laterthan stage 10 however no adverse impacts of BPAon survival or development occur (Iwamuro et al2003 Pickford et al 2003 Levy et al 2004)Exposure of Rana temporaria to BPA in combinationwith another stressor ultraviolet-B (UVB) radiationhas been shown to induce an enhanced effect ontadpole mortality (Koponen amp Kukkonen 2002) Themechanism for this is unknown but it is not due toan effect of UVB radiation on the accumulation ordepuration kinetics of BPA (Koponen et al 2007)
BPA has been reported to affect sexual develop-ment causing sex reversal at 1027 M (228 mg l21) insemi-static exposures (Kloas et al 1999 Levy et al2004) but under flow-through conditions (at anexposure to BPA up to 500 mg l21) no such feminizingeffects were found (Pickford et al 2003) In the newdata presented here no effects of exposure to BPA(using a flow-through system and spanning concen-trations from 228 ng l21 to 228 mg l21) were seen ongross morphological sex in the controls and BPAtreatments there were between 45 and 50 per centmales and 50 and 55 per cent females and in theoestradiol (E2)-positive control (02 mg l21) therewere 19 per cent males 72 per cent females and 9per cent mixed sex However detailed histologicalanalysis of the testes (n varies between 19 in the E2treatment and 47 in 1026 M BPA treatment)mdashexam-ining the complete gonad by serial sectionsmdashidentifiedeffects for the BPA treatments At 1029 1028 1027
and 1026 M (228 ng l21 and 228 228 and228 mg l21) BPA testicular lacunae occurred at fre-quencies of 15 20 10 and 25 per cent and oogoniawith frequencies of 40 60 55 and 60 per cent demon-strating oestrogenic effects even at 1029 M BPA(228 ng l21) In addition BPA also affected growthand body weight these were significantly increased inmales exposed to concentrations of 1028 M BPA(228 mg l21) and above (p 005 MannndashWhitneyU-test) This effect on growth was supported by theinduction of hepatic IGF-I expression at 1028 MBPA (228 mg l21) and above In these treatmentsthere was a significant induction of VTG geneexpression in liver at 1026 M BPA (228 mg l21)
In competitive displacement experiments with cyto-solic liver extracts of X laevis BPA was shown to bindonly moderately to amphibian ER (Lutz amp Kloas1999) Similarly in VTG- and ER-mRNA inductionstudies with primary hepatocytes from male X laevisBPA was stimulatory at 1027 M (228 mg l21) com-pared with 1029 M (272 ng l21) for E2 (Kloas et al1999 Lutz et al 2005) In another longer term invitro study induction of VTG occurred only at aBPA concentration of 11 1025 M (25 mg l21)(compared with 41 10211 M (111 ng l21) in theE2 positive control Nomura et al 2006) Both
Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

Review Effects of plasticizers on wildlife J Oehlmann et al 2057
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
experimental approaches indicate that BPA is moder-ately to weakly oestrogenic for VTG and ER inductionBPA has also been shown to significantly reduce basalwater absorption of male pelvic patches in the Japanesetree frog Hyla japonica a known oestrogenic response(Kohno et al 2004)
BPA also affects the thyroid system in amphibiansExposure to 1025 M BPA (228 mg l21) has beenshown to inhibit spontaneous and TH-induced meta-morphic changes in X laevis and suppress TRb geneexpression both in vivo and in vitro (Iwamuro et al2003) More recently in vitro tail cultures of Xlaevis have shown to respond to BPA reducingT3-induced gene expression of TRa and TRb atexposures as low as 1027 M (228 mg l21) In contrastthe T3-induced suppression of RXRg was only moder-ately affected by exposure to BPA alone (Iwamuroet al 2006) Similar results for effects of BPA onT3-induced metamorphic changes were found byGoto et al (2006) studying Silurana tropicalis andR rugosa In the skin of Rana tagoi expression of theantimicrobial peptides preprotemporins (normallyupregulated by TH) has been shown to be reducedmarkedly following BPA treatment at 1028 M(228 mg l21) (Ohnuma et al 2006) and using trans-genic X laevis embryos bearing a THbZIP-greenfluorescent protein construct T3-induced fluorescencewas shown to be inhibited by BPA at a concentration of1025 M (228 mg l21 Fini et al 2007)
4 CONCLUSIONThe datasets reported upon here show that phthalatesand BPA can affect both development and reproduc-tion in a wide range of wildlife species Reproductiveand developmental disturbances include alterationsin the number of offspring produced andor reducedhatching success disruption of larval developmentand in insects delayed emergence Interphyla differ-ences in responses to plasticizers occur and as anexample in amphibians phthalates and BPA disruptthyroid function impacting on larval developmentbut no such effects have been reported thus far forfish even though TH is deposited into fish oocytesand subsequently promotes and regulates larval meta-morphosis Similarly in molluscs plasticizers have notbeen found to affect embryo development There areclear differences in species sensitivity to the effects ofplasticizers too Molluscs crustaceans and amphibiansappear more responsive with most effects induced inexposures via the water in the low ng l21 to mg l21 con-centration range In contrast effects in fish are gener-ally induced in the higher mg l21 and mg l21 rangewith the exception of disruption of spermatogenesisby BPA that occurs in the low mg l21 range Thesetaxon-specific effects hamper meaningful comparisonsof the relative sensitivities of different phyla toplasticizers
Given that the biological effect concentrations forplasticizers seen in the laboratory coincide withenvironmental concentrations some wildlife popu-lations are probably impacted To date howeverstudies investigating population effects have not beenreported and this constitutes a significant knowledge
Phil Trans R Soc B (2009)
gap Furthermore for invertebrate phyla most highlysensitive to the biological effects of specific plasticizerslong-term exposures should be considered as aresearch priority However only a very limitednumber of invertebrate phyla have been tested forthe effects of phthalates and BPA and representativesfrom other presently neglected taxa should be con-sidered in the future including terrestrial speciesMixture effects of plasticizers as of many otherenvironmental pollutants are also a much neglectedarea of study and given that there are likely to be addi-tive effects this warrants investigations to assessmore accurately their impacts in the environment(see discussion in Koch amp Calafat 2009 Teuten et al2009 Thompson et al 2009b) As a final noteincreasingly investigations into the mechanisms ofaction of plasticizers are finding that some (eg phtha-lates) can have multiple interaction sites in the bodyaffecting a wide range of biological processes (see dis-cussion in Meeker et al 2009 Talsness et al 2009)Thus a more thorough understanding of the modesof action of these chemicals will help in developing amore comprehensive understanding of their potentialfor harm and in the identification of the species mostvulnerable to their biological effects
This work was financially supported by the European Union(COMPRENDO project contract EVK1-CT-2002-00129)EMS and GCP were funded by the NaturalEnvironment Research Council (NERC) grant ref NEC5076611 under the Post-genomics and Proteomicsresearch programme and under the EnvironmentalGenomics Thematic (grant ref NERTS200200182)and by the Biotechnology and Biological Sciences ResearchCouncil (BBSRC grant ref 9S15001) UK to CRT
REFERENCESAarab N Lemaire-Gony S Unruh E Hansen P D
Larsen B K Andersen O K amp Narbonne J F 2006Preliminary study of responses in mussel (Mytilus edulis)exposed to bisphenol A diallyl phthalate and tetrabromo-
diphenyl ether Aquat Toxicol 78 86ndash92 (doi101016jaquatox200602021)
Albro P W Corbett J T amp Schroeder J L 1993 Themetabolism of di-(2-ethylhexyl)phthalate in the earth-
worm Lumbricus terrestris Comp Biochem Physiol C104 335ndash344 (doi1010160742-8413(93)90045-M)
Alo R Facciolo R M Madeo M Giusi G Carelli A ampCanonaco M 2005 Effects of the xenoestrogen bis-phenol A in diencephalic regions of the teleost fish Corisjulis occur preferentially via distinct somatostatin receptorsubtypes Brain Res Bull 65 267ndash273 (doi101016jbrainresbull200501006)
Andersen H R Halling-Sorensen B amp Kusk K O 1999A parameter for detecting estrogenic exposure in the
copepod Acartia tonsa Ecotoxicol Environ Saf 4456ndash61 (doi101006eesa19991800)
Andersen H R Wollenberger L Halling-Sorensen B ampKusk K O 2001 Development of copepod nauplii tocopepoditesmdasha parameter for chronic toxicity including
endocrine disruption Environ Toxicol Chem 20 2821ndash2829 (doi1018971551-5028(2001)0202821DOCNTC20CO2)
Andrady A L amp Neal M A 2009 Applications and societal
benefits of plastics Phil Trans R Soc B 364 1977ndash1984 (doi101098rstb20080304)
2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

2058 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Baird D J Barber I Bradley M Soares A M V M ampCalow P 1991 A comparative study of genotype sensi-tivity to acute toxic stress using clones of Daphniamagna Straus Ecotoxicol Environ Saf 21 257ndash265(doi1010160147-6513(91)90064-V)
Barse A V Chakrabarti T Ghosh T K Pal A K ampJadhao S B 2007 Endocrine disruption and metabolicchanges following exposure of Cyprinus carpio to diethyl
phthalate Pest Biochem Physiol 88 36ndash42 (doi101016jpestbp200608009)
Barsiene J Syvokiene J amp Bjornstad A 2006 Induction ofmicronuclei and other nuclear abnormalities in mussels
exposed to bisphenol A diallyl phthalate and tetrabromo-diphenyl ether-47 Aquat Toxicol 78 105ndash108 (doi101016jaquatox200602023)
Bauer M J amp Herrmann R 1997 Estimation of theenvironmental contamination by phthalic acid esters
leaching from household wastes Sci Total Environ 20849ndash57 (doi101016S0048-9697(97)00272-6)
Beauchesne I Barnabe S Cooper D G amp Nicell J A2007 Plasticizers and related toxic degradation productsin wastewater sludges In IWA Specialist Conf BiosolidsMoncton New Brunswick Canada 24ndash27 June
Biggers W J amp Laufer H 2004 Identification of juvenilehormone-active alkylphenols in the lobster Homarusamericanus and in marine sediments Biol Bull 20613ndash24 (doi1023071543194)
Bolognesi C Perrone E Roggieri P Pampanin D M ampSciutto A 2006 Assessment of micronuclei induction inperipheral erythrocytes of fish exposed to xenobioticsunder controlled conditions Aquat Toxicol 78 93ndash98
(doi101016jaquatox200602015)Brennan S J Brougham C A Roche J J amp Fogarty A M
2006 Multi-generational effects of four selected environ-mental oestrogens on Daphnia magna Chemosphere 6449ndash55 (doi101016jchemosphere200511046)
Brian J V et al 2005 Accurate prediction of the response offreshwater fish to a mixture of estrogenic chemicalsEnviron Health Perspect 113 721ndash728 (doi101289ehp7598)
Cajaraville M P amp Ortiz-Zarragoitia M 2006 Specificity of
the peroxisome proliferation response in mussels exposedto environmental pollutants Aquat Toxicol 78 117ndash123(doi101016jaquatox200602016)
Call D J et al 2001 An assessment of the toxicity of phtha-late esters to freshwater benthos 1 Aqueous exposures
Environ Toxicol Chem 20 1798ndash1804 (doi1018971551-5028(2001)0201798AAOTTO20CO2)
Canesi L Betti M Lorussoa L C Ciaccia C amp Gallob G2005 lsquoIn vivorsquo effects of bisphenol A in Mytilus hemocytes
modulation of kinase-mediated signalling pathways AquatToxicol 71 73ndash84 (doi101016jaquatox200410011)
Canesi L Lorussoa L C Ciaccia C Betti M Rocchi MPoiana G amp Marcomini A 2007a Immunomodulationof Mytilus hemocytes by individual estrogenic chemicals
and environmentally relevant mixtures of estrogens invitro and in vivo studies Aquat Toxicol 81 36ndash44(doi101016jaquatox200610010)
Canesi L Borghi C Ciacci C Fabbri R Vergani L ampGallo G 2007b Bisphenol A alters gene expression and
functional parameters in molluscan hepatopancreasMol Cell Endocrinol 276 36ndash44 (doi101016jmce200706002)
Chen M Y Ike M amp Fujita M 2002 Acute toxicitymutagenicity and estrogenicity of bisphenol A and
other bisphenols Environ Toxicol 17 80ndash86 (doi101002tox10035)
Cook J W Dodds E C amp Hewett C J 1933 A syntheticoestrus-exciting compound Nature 131 56ndash57 (doi101038131056b0)
Phil Trans R Soc B (2009)
Corton J C amp Lapinskas P J 2005 Peroxisome proliferator-activated receptors mediators of phthalate ester-inducedeffects in the male reproductive tract Toxicol Sci 83
4ndash17 (doi101093toxscikfi011)Crain D A Eriksen M Iguchi T Jobling S Laufer H
LeBlanc G A amp Guillette L J 2007 An ecologicalassessment of bisphenol A evidence from comparativebiology Reprod Toxicol 24 225ndash239 (doi101016j
reprotox200705008)Dinan L Bourne P Whiting P Dhadialla T S amp
Hutchinson T H 2001 Screening of environmental con-taminants for ecdysteroid agonist and antagonist activity
using the Drosophila melanogaster B-II cell in vitro assayEnviron Toxicol Chem 20 2038ndash2046 (doi101897
1551-5028(2001)0202038SOECFE20CO2)Dixon D R Wilson J T Pascoe P L amp Parry J M 1999
Anaphase aberrations in the embryos of the marine tube-
worm Pomatoceros lamarckii (Polychaeta Serpulidae) anew in vivo test assay for detecting aneugens andclastogens in the marine environment Mutagenesis 14375ndash383 (doi101093mutage144375)
Dodds E C amp Lawson W 1936 Synthetic oestrogenic
agents without the phenanthrene nucleus Nature 137996 (doi101038137996a0)
Dodds E C amp Lawson W 1938 Molecular structure inrelation to oestrogenic activity Compounds withouta phenanthrene nucleus Proc R Soc Lond B 125
222ndash232 (doi101098rspb19380023)Dodds E C Goldberg L Lawson W amp Robinson R
1938 Estrogenic activity of certain synthetic compoundsNature 141 247ndash248 (doi101038141247b0)
Duft M Schulte-Oehlmann U Weltje L Tillmann M ampOehlmann J 2003 Stimulated embryo production as aparameter of estrogenic exposure via sediments in the fresh-water mudsnail Potamopyrgus antipodarum Aquat Toxicol64 437ndash449 (doi101016S0166-445X(03)00102-4)
EU 2003a European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C9-11-branched alkyl esters C10-rich and di-rsquoisodecylrsquo phthalate (DIDP) LuxembourgOffice for Official Publications of the EuropeanCommunities
EU 2003b European Union Risk Assessment Report for 12-ben-zenedicarboxylic acid di-C8-10-branched alkyl esters C9-richand di-lsquoisononylrsquo phthalate (DINP) Luxembourg Officefor Official Publications of the European Communities
EU 2003c European Union Risk Assessment Report for 440-iso-propylidenediphenol (bisphenol-A) Luxembourg Office forOfficial Publications of the European Communities
EU 2003d European Union Risk Assessment Report DraftBis(2-ethylhexyl)phthalate (DINP) Luxembourg Office
for Official Publications of the European CommunitiesEU 2004 European Union Risk Assessment Report for dibutyl
phthalate with addendum to the environmental section 2004Luxembourg Office for Official Publications of theEuropean Communities
EU 2006 European Union Risk Assessment Report for bis(2-ethylhexyl) phthalate Luxembourg Office for OfficialPublications of the European Communities Draft ofMarch 2006
Fan L Q You L Brown-Borg H Brown S Edwards
R J amp Corton J C 2004 Regulation of phase I andphase II steroid metabolism enzymes by PPARa activatorsToxicology 204 109ndash121 (doi101016jtox200406018)
Fatoki O S amp Vernon F 1990 Phthalates esters in rivers ofthe Greater Manchester area UK Sci Total Environ 95
227ndash232 (doi1010160048-9697(90)90067-5)Fenske M van Aerle R Brack S Tyler C R amp Segner
H 2001 Development and validation of a homologouszebrafish (Danio rerio Hamilton-Buchanan) vitellogeninenzyme-linked immunosorbent assay (ELISA) and its
Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

Review Effects of plasticizers on wildlife J Oehlmann et al 2059
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
application for studies on estrogenic chemicals CompBiochem Physiol C 129 217ndash232
Ferguson M S Graham O H Bang F B amp Hairston
N G 1946 Studies on Schistosomiasis japonicaV Protection experiments against Schistosomiasis japonicaAm J Epidemiol 44 367ndash378
Fini J B Le Mevel S Turque N Palmier K Zalko DCravedi J P amp Demeneix B 2007 An in vivo multiwell-
based fluorescent screen for monitoring vertebratethyroid hormone disruption Environ Sci Technol 415908ndash5914 (doi101021es0704129)
Forbes V E Aufderheide J Warbritton R van der
Hoevend N amp Caspers N 2007 Does bisphenol Ainduce superfeminization in Marisa cornuarietis Part IItoxicity test results and requirements for statisticalpower analyses Ecotoxicol Environ Saf 66 319ndash325(doi101016jecoenv200609001)
Forget-Leray J Landriau I Minier C amp Leboulenger F2005 Impact of endocrine toxicants on survival develop-ment and reproduction of the estuarine copepodEurytemora affinis (Poppe) Ecotoxicol Environ Saf 60288ndash294 (doi101016jecoenv200406008)
Fromme H Kuchler T Otto T Pilz K Muller J ampWenzel A 2002 Occurrence of phthalates and bisphenolA and F in the environment Water Res 36 1429ndash1438(doi101016S0043-1354(01)00367-0)
Galli A Pinaire J Fischer M Dorris R amp Crabb D W
2001 The transcriptional and DNA binding activity ofperoxisome proliferator-activated receptor alpha is inhib-ited by ethanol metabolismmdasha novel mechanism for thedevelopment of ethanol-induced fatty liver J BiolChem 276 68ndash75 (doi101074jbcM008791200)
Gillesby B E amp Zacharewski T R 1998 Exoestrogensmechanisms of action and strategies for identificationand assessment Environ Toxicol Chem 17 3ndash14(doi1018971551-5028(1998)0170003EMOAAS23
CO2)Goto Y Kitamura S Kashiwagi K Oofusa K Tooi O
Yoshizato K Sato J Ohta S amp Kashiwagi A 2006Suppression of amphibian metamorphosis by bisphenolA and related chemical substances J Health Sci 52
160ndash168 (doi101248jhs52160)Harries J E Runnalls T Hill E Harris C A Maddix
S Sumpter J P amp Tyler C R 2000 Development of areproductive performance test for endocrine disruptingchemicals using pair-breeding fathead minnows
(Pimephales promelas) Environ Sci Technol 34 3003ndash3011 (doi101021es991292a)
Harris C A Henttu P Parker M G amp Sumpter J P1997 The estrogenic activity of phthalate esters in vitroEnviron Health Perspect 105 802ndash811 (doi1023073433697)
Hayashi H Nishimoto A Oshima N amp Iwamuro S2007 Expression of the estrogen receptor alpha gene inthe anal fin of Japanese medaka Oryzias latipes by
environmental concentrations of bisphenol A J ToxicolSci 32 91ndash96 (doi102131jts3291)
Heemken O P Reincke H Stachel B amp Theobald N2001 The occurrence of xenoestrogens in the Elbe riverand the North Sea Chemosphere 45 245ndash259 (doi10
1016S0045-6535(00)00570-1)Hirano M Ishibashi H Matsumura N Nagoa Y
Watanabe N Watanabe A Onikura N Kishi K ampArizono K 2004 Acute toxicity responses of two crus-taceans Americamysis bahia and Daphnia magna to endo-
crine disrupters J Health Sci 50 97ndash100 (doi101248jhs5097)
Howard P H (ed) 1989 Handbook of environmental fate andexposure data for organic chemicals vol I large productionand priority pollutants Boca Raton FL Lewis Publishers
Phil Trans R Soc B (2009)
Howard P H Banerjee S amp Robillard K H 1985Measurement of water solubilities octanol waterpartition-coefficients and vapor-pressures of commercial
phthalate-esters Environ Toxicol Chem 4 653ndash661(doi1018971552-8618(1985)4[653MOWSWP]20CO2)
Ishibashi H Watanabe N Matsumura N Hirano MNagao Y Shiratsuchi H Kohra S Yoshihara S amp
Arizono K 2005 Toxicity to early life stages and an estro-genic effect of a bisphenol A metabolite 4-methyl-24-bis(4-hydroxyphenyl)pent-1-ene on the medaka (Oryziaslatipes) Life Sci 77 2643ndash2655 (doi101016jlfs
200503025)Iwamuro S Sakakibara M Terao M Ozawa A
Kurobe C Shigeura T Kato M amp Kikuyama S2003 Teratogenic and anti-metamorphic effects ofbisphenol A on embryonic and larval Xenopus laevisGen Comp Endocrinol 133 189ndash198 (doi101016S0016-6480(03)00188-6)
Iwamuro S Yamada M Kato M amp Kikuyama S 2006Effects of bisphenol A on thyroid hormone-dependentup-regulation of thyroid hormone receptor alpha and
beta down-regulation of retinoid X receptor gamma inXenopus tail culture Life Sci 79 2165ndash2171 (doi101016jlfs200607013)
Jagnytsch O Opitz R Lutz I amp Kloas W 2006 Effects oftetrabromobisphenol A on larval development and thyr-
oid hormone regulated biomarkers of the amphibianXenopus laevis Environ Res 101 340ndash348 (doi101016jenvres200509006)
Jobling S Reynolds T White R Parker M G amp
Sumpter J P 1995 A variety of environmentallypersistent chemicals including some phthalate plastici-zers are weakly estrogenic Environ Health Perspect 103582ndash587 (doi1023073432434)
Jobling S Casey D Rodgers-Gray T Oehlmann J
Schulte-Oehlmann U Pawlowski S Braunbeck TTurner A P amp Tyler C R 2004 Comparative responsesof molluscs and fish to environmental estrogens and anestrogenic effluent Aquat Toxicol 66 207ndash222(doi101016jaquatox200401002)
Jonsson S amp Baun A 2003 Toxicity of mono- and diestersof o-phthalic esters to a crustacean a green alga and abacterium Environ Toxicol Chem 22 3037ndash3043(doi10189702-548)
Kasahara E Sato E F Miyoshi M Konaka R
Hiramoto K Sasaki J Tokuda M Nakano Y ampInoue M 2002 Role of oxidative stress in germ cell apop-tosis induced by di(2-ethylhexyl)phthalate Biochem J365 849ndash856
Kim E J Kim J W amp Lee S K 2002 Inhibition ofoocyte development in Japanese medaka (Oryziaslatipes) exposed to di-2-ethylhexyl phthalate EnvironInt 28 359ndash365 (doi101016S0160-4120(02)00058-2)
Kloas W 2002 Amphibians as model for the study of endo-crine disrupters Int Rev Cytol 216 1ndash57 (doi101016S0074-7696(02)16002-5)
Kloas W Lutz I amp Einspanier R 1999 Amphibians asmodel to study endocrine disruptors II Estrogenic
activity of environmental chemicals in vitro and in vivoSci Total Environ 225 59ndash68 (doi101016S0048-9697(99)80017-5)
Koch H M amp Calafat A M 2009 Human body burdensof chemicals used in plastic manufacture Phil Trans RSoc B 364 2063ndash2078 (doi101098rstb20080208)
Kohno S Fujime M Kamishima Y amp Iguchi T 2004Sexually dimorphic basal water absorption at the isolatedpelvic patch of Japanese tree frog Hyla japonica J ExpZool A 301 428ndash438
2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

2060 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Koponen P S amp Kukkonen J V K 2002 Effects ofbisphenol A and artificial UVB radiation on theearly development of Rana temporaria J ToxicolEnviron Health A 65 947ndash959 (doi10108000984100290071180)
Koponen P S Tuikka A amp Kukkonen J V K 2007Effects of ultraviolet-B radiation and larval growth on tox-icokinetics of waterborne bisphenol A in common frog
(Rana temporaria) larvae Chemosphere 66 1323ndash1328(doi101016jchemosphere200607018)
Kusk K O amp Wollenberger L 1999 Fully defined saltwatermedium for cultivation of and toxicity testing with marine
copepod Acartia tonsa Environ Toxicol Chem 20 2821ndash2829
Kusk K O amp Wollenberger L 2007 Towards an interna-tionally harmonized test method for reproductive anddevelopmental effects of endocrine disrupters in marine
copepods Ecotoxicology 16 183ndash195 (doi101007s10646-006-0112-2)
Kwak I S amp Lee W 2005 Endpoint for DEHP exposureassessment in Chironomus riparius Bull EnvironContam Toxicol 74 1179ndash1185 (doi101007s00128-
005-0705-0)Labadie P amp Budzinski H 2006 Alteration of steroid hor-
mone balance in juvenile turbot (Psetta maxima) exposedto nonylphenol bisphenol A tetrabromodiphenyl ether47 diallylphthalate oil and oil spiked with alkylphenols
Arch Environ Contam Toxicol 50 552ndash561 (doi101007s00244-005-1043-2)
Lahnsteiner F Berger B Kletzl M amp Weismann T 2005Effect of bisphenol A on maturation and quality of semen
and eggs in the brown trout Salmo trutta f fario AquatToxicol 75 213ndash224 (doi101016jaquatox200508004)
Larsen B K Bjornstad A Sundt R C Taban I CPampanin D M amp Andersen O K 2006 Comparison
of protein expression in plasma from nonylphenol andbisphenol A-exposed Atlantic cod (Gadus morhua) andturbot (Scophthalmus maximus) by use of SELDI-TOFAquat Toxicol 78 25ndash33 (doi101016jaquatox200602026)
Larsson P amp Thuren A 1987 Di-2-ethylhexylphthalateinhibits the hatching of frog eggs and is bioaccumulatesby tadpoles Environ Toxicol Chem 6 417ndash422(doi1018971552-8618(1987)6[417DITHOF]20CO2)
Lee S K amp Veeramachaneni D N R 2005 Subchronic
exposure to low concentrations of di-n-butyl phthalatedisrupts spermatogenesis in Xenopus laevis frogs ToxicolSci 84 394ndash407 (doi101093toxscikfi087)
Lee S K Owens G A amp Veeramachaneni D N 2005
Exposure to low concentrations of di-n-butyl phthalateduring embryogenesis reduces survivability and impairsdevelopment of Xenopus laevis frogs J Toxicol EnvironHealth A 68 763ndash772 (doi10108015287390590930243)
Lee S M Lee S B Park C H amp Choi J 2006aExpression of heat shock protein and hemoglobin genesin Chironomus tentans (Diptera chironomidae) larvaeexposed to various environmental pollutants a potentialbiomarker of freshwater monitoring Chemosphere 65
1074ndash1081 (doi101016jchemosphere200602042)Lee Y M Seo J S Kim I C Yoon Y D amp Lee J S
2006b Endocrine disrupting chemicals (bisphenol A 4-nonylphenol 4-tert-octylphenol) modulate expression oftwo distinct cytochrome P450 aromatase genes differently
in gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 345 894ndash903(doi101016jbbrc200604137)
Lee Y M Rhee J S Hwang D S Kim I C RaisuddinS amp Lee J S 2007 Mining of biomarker genes from
Phil Trans R Soc B (2009)
expressed sequence tags and differential display reversetranscriptasendashpolymerase chain reaction in the self-fertilizing fish Kryptolebias marmoratus and their
expression patterns in response to exposure to an endo-crine-disrupting alkylphenol bisphenol A Mol Cells 23287ndash303
Levy G Lutz I Kruger A amp Kloas W 2004 Bisphenol Ainduces feminization in Xenopus laevis tadpoles EnvironRes 94 102ndash111 (doi101016S0013-9351(03)00086-0)
Lindholst C Pedersen S N amp Bjerregaard P 2001Uptake metabolism and excretion of bisphenol A in therainbow trout (Oncorhynchus mykiss) Aquat Toxicol 55
75ndash84 (doi101016S0166-445X(01)00157-6)Lindholst C Wynne P M Marriott P Pedersen S N amp
Bjerregaard P 2003 Metabolism of bisphenol A in zebra-fish (Danio rerio) and rainbow trout (Oncorhynchus mykiss)in relation to estrogenic response Comp Biochem PhysiolC 135 169ndash177
Liu K J Lehmann K P Sar M Young S S amp GaidoK W 2005 Gene expression profiling following in uteroexposure to phthalate esters reveals new gene targets in
the etiology of testicular dysgenesis Biol Reprod 73
180ndash192 (doi101095biolreprod104039404)Lutz I amp Kloas W 1999 Amphibians as model to study
endocrine disruptors I Environmental pollution andestrogen receptor binding Sci Total Environ 225 49ndash57 (doi101016S0048-9697(99)80016-3)
Lutz I Blodt S amp Kloas W 2005 Regulation of estrogenreceptors in primary cultured hepatocytes of the amphi-bian Xenopus laevis as estrogenic biomarker and its appli-cation in environmental monitoring Comp BiochemPhysiol 141 384ndash392
Lutz I Kloas W Springer T A Holden L R WolfJ C Kruger H O amp Hosmer A J 2008Development standardization and refinement of pro-cedures for evaluating effects of endocrine active com-
pounds on development and sexual differentiation ofXenopus laevis Anal Bioanal Chem 390 2031ndash2048(doi101007s00216-008-1973-4)
Mandich A Bottero S Benfenati E Cevasco AErratico C Maggioni S Massari A Pedemonte F amp
Vigano L 2007 In vivo exposure of carp to gradedconcentrations of bisphenol A Gen Comp Endocrinol153 15ndash24 (doi101016jygcen200701004)
Marcial H S Hagiwara A amp Snell T W 2003 Estrogeniccompounds affect development of harpacticoid copepod
Tigriopus japonicus Environ Toxicol Chem 22 3025ndash3030(doi10189702-622)
Mariager L 2001 Effects of environmental endocrinedisrupters on a freshwater and a marine crustacean
Master thesis Aarhus University Dept Zool Instituteof Biological Sciences Aarhus Denmark
Meeker J D Sathyanarayana S amp Swan S H 2009Phthalates and other additives in plastics humanexposure and associated health outcomes Phil Trans RSoc B 364 2097ndash2113 (doi101098rstb20080268)
Metcalfe C D Metcalfe T L Kiparissis Y Koenig B GKhan C Hughes R J Croley T R March R E ampPotter T 2001 Estrogenic potency of chemicals detectedin sewage treatment plant effluents as determined by invivo assays with Japanese medaka (Oryzias latipes)Environ Toxicol Chem 20 297ndash308 (doi1018971551-5028(2001)0200297EPOCDI20CO2)
Moens L N van der Ven K Van Remortel P Del-Favero J amp De Coen W M 2006 Expression profiling
of endocrine-disrupting compounds using a customizedCyprinus carpio cDNA microarray Toxicol Sci 93298ndash310 (doi101093toxscikfl057)
Moens L N van der Ven K Van Remortel R Del-Favero J amp De Coen W M 2007 Gene expression
Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

Review Effects of plasticizers on wildlife J Oehlmann et al 2061
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
analysis of estrogenic compounds in the liver of commoncarp (Cyprinus carpio) using a custom cDNA microarrayJ Biochem Mol Toxicol 21 299ndash311 (doi101002jbt
20190)Mu X Y Rider C V Hwang G S Hoy H amp LeBlanc
G A 2005 Covert signal disruption anti-ecdysteroidalactivity of bisphenol A involves cross talk between signal-ling pathways Environ Toxicol Chem 24 146ndash152
(doi10189704-063R1)Naito W Gamo Y amp Yoshida K 2006 Screening-level risk
assessment of di(2-ethylhexyl) phthalate for aquaticorganisms using monitoring data in Japan EnvironMonit Assess 115 451ndash471 (doi101007s10661-006-7239-8)
Neuhauser E F Durkin P R Malecki M R amp AnatraM 1986 Comparative toxicity of ten organic chemicalsto four earthworm species Comp Biochem Physiol 83
197ndash200 (doi1010160742-8413(86)90036-8)Nomura Y Mitsui N Bhawal U K Sawajiri M Tooi O
Takahashi T amp Okazaki M 2006 Estrogenic activity ofphthalate esters by in vitro VTG assay using primary-cultured Xenopus hepatocytes Dent Mater J 25 533ndash537
Norman A Borjeson H David F Tienpont B ampNorrgren L 2007 Studies of uptake elimination andlate effects in Atlantic salmon (Salmo salar) dietaryexposed to di-2-ethylhexyl phthalate (DEHP) duringearly life Arch Environ Contam Toxicol 52 235ndash242
(doi101007s00244-005-5089-y)Oehlmann J Schulte-Oehlmann U Tillmann M amp
Markert B 2000 Effects of endocrine disruptors on proso-branch snails (Mollusca Gastropoda) in the laboratory Part
I bisphenol A and octylphenol as xeno-estrogensEcotoxicology 9 383ndash397 (doi101023A1008972518019)
Oehlmann J Schulte-Oehlmann U Bachmann JOetken M Lutz I Kloas W amp Ternes T A 2006Bisphenol A induces superfeminization in the ramshorn
snail Marisa cornuarietis (Gastropoda Prosobranchia) atenvironmentally relevant concentrations Environ HealthPerspect 114 127ndash133 (doi101289ehp8065)
Ohnuma A Conlon J M Kawasaki H amp Iwamuro S2006 Developmental and triiodothyronine-induced
expression of genes encoding preprotemporins in theskin of Tagorsquos brown frog Rana tagoi Gen CompEndocrinol 146 242ndash250 (doi101016jygcen200511015)
Ohtani H Miura I amp Ichikawa Y 2000 Effects of dibutyl
phthalate as an environmental endocrine disrupter ongonadal sex differentiation of genetic males of the frogRana rugosa Environ Health Perspect 108 1189ndash1193(doi1023073434832)
Oka T Adati N Shinkai T Sakuma K Nishimura T ampKurose K 2003 Bisphenol A induces apoptosis in central
neural cells during early development of Xenopus laevisBiochem Biophys Res Commun 312 877ndash882 (doi101016jbbrc200310199)
Olea N Pulgar R Perez P Oleaserrano F Rivas ANovilloFertrell A Pedraza V Soto A M ampSonnenschein C 1996 Estrogenicity of resin-basedcomposites and sealants used in dentistry EnvironHealth Perspect 104 298ndash305 (doi1023073432888)
Opitz R Lutz I Nguyen N Scanlan T amp Kloas W2006 The analysis of thyroid hormone receptor bAmRNA expression in Xenopus laevis tadpoles as a meansto detect agonism and antagonism of thyroid hormoneaction Toxicol Appl Pharmacol 212 1ndash13 (doi101016
jtaap200506014)Orbea A Ortiz-Zarragoitia M amp Cajaraville M P 2002
Interactive effects of benzo(a)pyrene and cadmium andeffects of di(2-ethylhexyl) phthalate on antioxidant andperoxisomal enzymes and peroxisomal volume density
Phil Trans R Soc B (2009)
in the digestive gland of mussel Mytilus galloprovincialisLmk Biomarkers 7 33ndash48
Park S Y amp Choi J 2007 Cytotoxicity genotoxicity and
ecotoxicity assay using human cell and environmentalspecies for the screening of the risk from pollutantexposure Environ Int 33 817ndash822 (doi101016jenvint200703014)
Patyna P J Brown R P Davi R A Letinski D J
Thomas P E Cooper K R amp Parkerton T F 2006Hazard evaluation of diisononyl phthalate and diisodecylphthalate in a Japanese medaka multigenerational assayEcotoxicol Environ Saf 65 36ndash47 (doi101016j
ecoenv200505023)Peijnenburg W amp Struijs J 2006 Occurrence of phthalate
esters in the environment of The NetherlandsEcotoxicol Environ Saf 63 204ndash215 (doi101016jecoenv200507023)
Pickford D B Hetheridge M J Caunter J E Hall A Tamp Hutchinson T H 2003 Assessing chronic toxicity ofbisphenol A to larvae of the African clawed frog(Xenopus laevis) in a flow-through exposure systemChemosphere 53 223ndash235 (doi101016S0045-6535
(03)00308-4)Santos E M Paull G C Van Look K J W Workman
V L Holt W V Van Aerle R Kille P amp Tyler C R2007 Gonadal transcriptome responses and physiologicalconsequences of exposure to oestrogen in breeding zebra-
fish (Danio rerio) Aquat Toxicol 83 134ndash142 (doi101016jaquatox200703019)
Schirling M Jungmann D Ladewig V LudwichowskiK U Nagel R Kohler H R amp Triebskorn R 2006
Bisphenol A in artificial indoor streams II Stressresponse and gonad histology in Gammarus fossarum(Amphipoda) Ecotoxicol 15 143ndash156 (doi101007s10646-005-0044-2)
Schulte-Oehlmann U Tillmann M Casey D Duft M
Markert B amp Oehlmann J 2001 OstrogenartigeWirkungen von Bisphenol A auf Vorderkiemerschnecken(Mollusca Gastropoda Prosobranchia) UWSFmdashZUmweltchem Okotox 13 319ndash333
Seo J S Lee Y M Jung S O Kim I C Yoon Y D amp
Lee J S 2006 Nonylphenol modulates expression ofandrogen receptor and estrogen receptor genes differentlyin gender types of the hermaphroditic fish Rivulus mar-moratus Biochem Biophys Res Commun 346 213ndash223(doi101016jbbrc200605123)
Sohoni P amp Sumpter J P 1998 Several environmental
oestrogens are also anti-androgens J Endocrinol 158327ndash339 (doi101677joe01580327)
Sohoni P et al 2001 Reproductive effects of long-term
exposure to bisphenol A in the fathead minnow(Pimephales promelas) Environ Sci Technol 35 2917ndash2925(doi101021es000198n)
Staples C A Peterson D R Parkerton T F amp AdamsW J 1997 The environmental fate of phthalates esters
a literature review Chemosphere 35 667ndash749Staples C A Dorn P B Klecka G M OrsquoBlock S T amp
Harris L R 1998 A review of the environmental fateeffects and exposures of bisphenol A Chemosphere 362149ndash2173 (doi101016S0045-6535(97)10133-3)
Staples C A Woodburn K Caspers N Hall A T ampKlecka G M 2002 A weight of evidence approach tothe aquatic hazard assessment of bisphenol A HumEcol Risk Assess 8 1083ndash1105 (doi1010801080-700291905837)
Tagatz M E Plaia G R amp Deans C H 1986 Toxicity ofdibutyl phthalate-contaminated sediment to laboratory-and field-colonized estuarine benthic communities BullEnviron Contam Toxicol 37 141ndash150 (doi101007BF01607741)
2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)

2062 J Oehlmann et al Review Effects of plasticizers on wildlife
on 15 June 2009rstbroyalsocietypublishingorgDownloaded from
Talsness C E Andrade A J M Kuriyama S N TaylorJ A amp vom Saal F S 2009 Components of plasticexperimental studies in animals and relevance for
human health Phil Trans R Soc B 364 2079ndash2096(doi101098rstb20080281)
Teuten E L et al 2009 Transport and release of chemicalsfrom plastics to the environment and to wildlife Phil TransR Soc B 364 2027ndash2045 (doi101098rstb20080284)
Thibaut R amp Porte C 2004 Effects of endocrine disrupterson sex steroid synthesis and metabolism pathways in fishJ Steroid Biochem Mol Biol 92 485ndash494 (doi101016jjsbmb200410008)
Thompson R C Moore C J vom Saal F S amp Swan S H2009a Plastics the environment and human health currentconsensus and future trends Phil Trans R Soc B 3642153ndash2166 (doi101098rstb20090053)
Thompson R C Swan S H Moore C J amp vom Saal
F S 2009b Our plastic age Phil Trans R Soc B 3641973ndash1976 (doi101098rstb20090054)
Thuren A amp Woin P 1991 Effects of phthalate-esters onthe locomotor-activity of the fresh water amphipodGammarus pulex Bull Environ Contam Toxicol 46
159ndash166 (doi101007BF01688270)Wang H Y Olmstead A W Li H amp LeBlanc G A 2005
The screening of chemicals for juvenoid-related endocrineactivity using the water flea Daphnia magna AquatToxicol 74 193ndash204 (doi101016jaquatox200505010)
Watts M M Pascoe D amp Carroll K 2001 Chronicexposure to 17a-ethinylestradiol and bisphenolAmdasheffects on development and reproduction in thefreshwater invertebrate Chironomus riparius (Diptera
Phil Trans R Soc B (2009)
Chironomidae) Aquat Toxicol 55 113ndash124 (doi101016S0166-445X(01)00148-5)
Weltje L amp Schulte-Oehlmann U 2007 The seven year
itchmdashprogress in research on endocrine disruption inaquatic invertebrates since 1999 Ecotoxicology 16 1ndash3(doi101007s10646-006-0116-y)
Wibe A E Billing A Rosenqvist G amp Jenssen B M2002 Butyl benzyl phthalate affects shoaling behavior
and bottom-dwelling behavior in threespine sticklebackEnviron Res 89 180ndash187 (doi101006enrs20024360)
Wibe A E Fjeld E Rosenqvist G amp Jenssen B M 2004
Postexposure effects of DDE and butylbenzylphthalate onfeeding behavior in threespine stickleback EcotoxicolEnviron Saf 57 213ndash219 (doi101016S0147-6513(03)00005-8)
Wilson J T Dixon D R amp Dixon L R J 2002
Numerical chromosomal aberrations in the early life-his-tory stages of a marine tubeworm Pomatoceros lamarckii(Polychaeta Serpulidae) Aquat Toxicol 59 163ndash175(doi101016S0166-445X(01)00249-1)
Wollenberger L Dinan L amp Breitholtz M 2005
Brominated flame retardants activities in a crustaceandevelopment test and in an ecdysteroid screening assayEnviron Toxicol Chem 24 400ndash407 (doi10189703-6291)
Xiao-yu H Bei W Shuzhen Z amp Xiao-quan S 2005
Bioavailability of phthalate congeners to earthworms(Eisenia fetida) in artificially contaminated soilsEcotoxicol Environ Saf 62 26ndash34 (doi101016jecoenv200502012)




















