
A central role for gamma-glutamyl hydrolases in plant folate homeostasis
11
A central role for gamma-glutamyl hydrolases in plant folate homeostasis Tariq A. Akhtar 1,†,* , Giuseppe Orsomando 2 , Payam Mehrshahi 3 , Aurora Lara-Nu ´n ˜ ez 4 , Malcolm J. Bennett 3 , Jesse F. Gregory III 4 and Andrew D. Hanson 1 1 Horticultural Sciences Department, University of Florida, Gainesville, FL 32611, USA, 2 Dipartimento Patologia Molecolare e Terapie Innovative, Universita ` Politecnica delle Marche, Ancona, 60131, Italy, 3 Plant Sciences Division, School of Biosciences, University of Nottingham, Sutton Bonington Campus, Loughborough LE12 5RD, UK, and 4 Food Science and Human Nutrition, University of Florida, Gainesville, FL 32611, USA Received 23 May 2010; accepted 28 July 2010; published online 16 September 2010. * For correspondence (fax +734 647 0884; e-mail [email protected]). † Present address: University of Michigan, Department of Molecular, Cellular and Developmental Biology, Kraus Natural Sciences Building, 830 North University, Ann Arbor, MI 48109, USA. SUMMARY Most cellular folates carry a short poly-c-glutamate tail, and this tail is believed to affect their efficacy and stability. The tail can be removed by c-glutamyl hydrolase (GGH; EC 3.4.19.9), a vacuolar enzyme whose role in folate homeostasis remains unclear. In order to probe the function of GGH, we modulated its level of expression and subcellular location in Arabidopsis plants and tomato fruit. Three-fold overexpression of GGH in vacuoles caused extensive deglutamylation of folate polyglutamates and lowered the total folate content by approximately 40% in Arabidopsis and tomato. No such effects were seen when GGH was overexpressed to a similar extent in the cytosol. Ablation of either of the major Arabidopsis GGH genes (AtGGH1 and AtGGH2) alone did not significantly affect folate status. However, a combination of ablation of one gene plus RNA interference (RNAi)-mediated suppression of the other (which lowered total GGH activity by 99%) increased total folate content by 34%. The excess folate accumulated as polyglutamate derivatives in the vacuole. Taken together, these results suggest a model in which: (i) folates continuously enter the vacuole as polyglutamates, accumulate there, are hydrolyzed by GGH, and exit as monoglutamates; and (ii) GGH consequently has an important influence on polyglutamyl tail length and hence on folate stability and cellular folate content. Keywords: folate, polyglutamate, gamma-glutamyl hydrolase, vacuole, tomato. INTRODUCTION Tetrahydrofolate (THF) and its derivatives, collectively termed folates, are indispensable cofactors in one-carbon (C 1 ) metabolism. Folates provide C 1 units at various oxida- tion levels (methyl, methylene, or formyl) to acceptor mol- ecules during the synthesis of purines, thymidylate, pantothenate, formyl-methionyl tRNA, and methionine (Scott et al., 2000). Moreover, as methionine is required to drive the activated methyl cycle, the synthesis of methylated products such as choline, lignin, and chlorophyll depends indirectly upon folates (Hanson and Roje, 2001; Re ´ beille ´ et al., 2006). Plants and microbes synthesize folates de novo, whereas animals have a strict dietary requirement, most of which comes from plant sources. Plant folates are assembled in the mitochondrion from pterin, p-aminobenzoate (pABA), and glutamate precursors (Figure 1). In vivo, the majority of folates are conjugated to a c-linked polyglutamyl tail of up to eight residues. The tail is added, one glutamate at a time, by folylpolyglutamate synthetase (FPGS; EC 6.3.2.17). In ani- mals and plants, multiple FPGS isoforms exist, each tar- geted to a distinct subcellular location (Chen et al., 1996; Ravanel et al., 2001). The polyglutamyl tail is shortened or removed by c-glutamyl hydrolase (GGH; EC 3.4.19.9), a vacuolar endo- and/or exo-peptidase found in animals and plants (Wang et al., 1993; Orsomando et al., 2005). As part of the folate salvage pathway, GGH also hydrolyzes polygluta- mates of the folate breakdown product, pABA polygluta- mate (Orsomando et al., 2006). The relative activities of FPGS and GGH are considered to determine the extent of intracellular folate polyglutamylation. 256 ª 2010 The Authors Journal compilation ª 2010 Blackwell Publishing Ltd The Plant Journal (2010) 64, 256–266 doi: 10.1111/j.1365-313X.2010.04330.x
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of A central role for gamma-glutamyl hydrolases in plant folate homeostasis

A central role for gamma-glutamyl hydrolases in plant folatehomeostasis
Tariq A. Akhtar1,†,*, Giuseppe Orsomando2, Payam Mehrshahi3, Aurora Lara-Nunez4, Malcolm J. Bennett3, Jesse F. Gregory III4
and Andrew D. Hanson1
1Horticultural Sciences Department, University of Florida, Gainesville, FL 32611, USA,2Dipartimento Patologia Molecolare e Terapie Innovative, Universita Politecnica delle Marche, Ancona, 60131, Italy,3Plant Sciences Division, School of Biosciences, University of Nottingham, Sutton Bonington Campus, Loughborough
LE12 5RD, UK, and4Food Science and Human Nutrition, University of Florida, Gainesville, FL 32611, USA
Received 23 May 2010; accepted 28 July 2010; published online 16 September 2010.*For correspondence (fax +734 647 0884; e-mail [email protected]).†Present address: University of Michigan, Department of Molecular, Cellular and Developmental Biology, Kraus Natural Sciences Building, 830 North University,
Ann Arbor, MI 48109, USA.
SUMMARY
Most cellular folates carry a short poly-c-glutamate tail, and this tail is believed to affect their efficacy and
stability. The tail can be removed by c-glutamyl hydrolase (GGH; EC 3.4.19.9), a vacuolar enzyme whose role in
folate homeostasis remains unclear. In order to probe the function of GGH, we modulated its level of
expression and subcellular location in Arabidopsis plants and tomato fruit. Three-fold overexpression of GGH
in vacuoles caused extensive deglutamylation of folate polyglutamates and lowered the total folate content by
approximately 40% in Arabidopsis and tomato. No such effects were seen when GGH was overexpressed to a
similar extent in the cytosol. Ablation of either of the major Arabidopsis GGH genes (AtGGH1 and AtGGH2)
alone did not significantly affect folate status. However, a combination of ablation of one gene plus RNA
interference (RNAi)-mediated suppression of the other (which lowered total GGH activity by 99%) increased
total folate content by 34%. The excess folate accumulated as polyglutamate derivatives in the vacuole. Taken
together, these results suggest a model in which: (i) folates continuously enter the vacuole as polyglutamates,
accumulate there, are hydrolyzed by GGH, and exit as monoglutamates; and (ii) GGH consequently has an
important influence on polyglutamyl tail length and hence on folate stability and cellular folate content.
Keywords: folate, polyglutamate, gamma-glutamyl hydrolase, vacuole, tomato.
INTRODUCTION
Tetrahydrofolate (THF) and its derivatives, collectively
termed folates, are indispensable cofactors in one-carbon
(C1) metabolism. Folates provide C1 units at various oxida-
tion levels (methyl, methylene, or formyl) to acceptor mol-
ecules during the synthesis of purines, thymidylate,
pantothenate, formyl-methionyl tRNA, and methionine
(Scott et al., 2000). Moreover, as methionine is required to
drive the activated methyl cycle, the synthesis of methylated
products such as choline, lignin, and chlorophyll depends
indirectly upon folates (Hanson and Roje, 2001; Rebeille
et al., 2006).
Plants and microbes synthesize folates de novo, whereas
animals have a strict dietary requirement, most of which
comes from plant sources. Plant folates are assembled in the
mitochondrion from pterin, p-aminobenzoate (pABA), and
glutamate precursors (Figure 1). In vivo, the majority of
folates are conjugated to a c-linked polyglutamyl tail of up to
eight residues. The tail is added, one glutamate at a time, by
folylpolyglutamate synthetase (FPGS; EC 6.3.2.17). In ani-
mals and plants, multiple FPGS isoforms exist, each tar-
geted to a distinct subcellular location (Chen et al., 1996;
Ravanel et al., 2001). The polyglutamyl tail is shortened or
removed by c-glutamyl hydrolase (GGH; EC 3.4.19.9), a
vacuolar endo- and/or exo-peptidase found in animals and
plants (Wang et al., 1993; Orsomando et al., 2005). As part of
the folate salvage pathway, GGH also hydrolyzes polygluta-
mates of the folate breakdown product, pABA polygluta-
mate (Orsomando et al., 2006). The relative activities of
FPGS and GGH are considered to determine the extent of
intracellular folate polyglutamylation.
256 ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd
The Plant Journal (2010) 64, 256–266 doi: 10.1111/j.1365-313X.2010.04330.x

Polyglutamylation can directly affect C1 metabolism,
because folate-dependent enzymes prefer polyglutamates
to monoglutamyl forms. For example, methionine synthases
from various organisms cannot use monoglutamyl
5-methyl-THF (5-CH3-THF) (Whitfield et al., 1970; Cherest
et al., 2000; Ravanel et al., 2004). The polyglutamyl tail may
also help to ‘channel’ folates between the active sites of
folate-dependent enzymes, many of which exist as com-
plexes (Schirch and Strong, 1989). Furthermore, because
polyglutamylation favors binding to enzymes, and enzyme-
bound polyglutamates are protected from oxidative break-
down, folates tend to be stabilized by polyglutamylation
(Suh et al., 2001). The ability of folate transporters to shuttle
folates across membranes decreases as polyglutamyl tail
length increases (Shane and Stokstad, 1975). Polyglutamy-
lation also increases the anionic nature of folates and
impedes the passive diffusion of folates across biomem-
branes (Appling, 1991). Hence, polyglutamylation generally
favors the retention of cellular and organellar polyglutamates in
the compartments in which they reside (Shane, 1989).
In plants, however, almost nothing specific is known
about the in vivo significance of polyglutamylation. Nor is
the functional role of GGH clear in relation to folate
homeostasis. In bacteria, overexpression of GGH causes
extensive deconjugation of polyglutamates and decreased
folate retention, without a major effect on growth rate
(Sybesma et al., 2003; Akhtar et al., 2008). In animal sys-
tems, enhanced GGH expression decreases polyglutamate
abundance and intracellular folate levels (Li et al., 1993;
Rhee et al., 1993; Cole et al., 2001), whereas increased FPGS
activity is associated with higher intracellular folate levels
(Lowe et al., 1993; Sakamoto et al., 2008). Taken together,
these observations imply that intracellular folate levels are
positively correlated with polyglutamate tail length.
To further examine this relationship, we turned to the
recently described GGH systems of Arabidopsis and tomato
(Solanum lycopersicum) (Orsomando et al., 2005; Akhtar
et al., 2008). The Arabidopsis GGH family comprises three
genes, AtGGH1, AtGGH2, and AtGGH3, arranged in tandem
on chromosome 1. Tomato also contains three GGH genes
(LeGGH1, LeGGH2, and LeGGH3) of which the latter encodes
a catalytically inactive isoform. Modulation of GGH activity
in Arabidopsis and tomato fruit revealed that total cellular
folate levels are influenced by the extent of GGH-mediated
hydrolysis of vacuolar polyglutamates. These results thus
demonstrate a direct impact of polyglutamylation on intra-
cellular folate content.
RESULTS
Overexpression of GGH in Arabidopsis and tomato fruit
The metabolic significance of GGH was first explored in
Arabidopsis and tomato fruit by overexpressing AtGGH2
and LeGGH2, respectively. These proteins exhibit both
endo- and exo-peptidase activity towards polyglutamates.
Two versions of these GGHs were introduced: the native full-
length (FL) protein containing the predicted signal peptide or
a truncated version (TR) missing the predicted signal pep-
tide. In Arabidopsis, a hexahistidine (His6) epitope was fused
to the C-terminus of AtGGH2-FL and to the N-terminus of
AtGGH2-TR, respectively. The constitutive 35S promoter
from figwort mosaic virus was used to drive expression in
Arabidopsis (Richins et al., 1987), while in tomato the E8
promoter was utilized to restrict expression to late stages of
fruit development (Kneissl and Deikman, 1996). From
among the independent transgenic lines obtained for each
species and each construct we selected three for further
analysis; these lines showed the largest increase in total
GGH activity, which was about threefold (Figure 2a). The
lines expressing the truncated GGHs were indistinguishable
from the wild type; tomato fruit from the LeGGH2-FL lines
and siliques from the AtGGH2-FL lines were consistently
smaller, with few seeds.
Targeting of the overexpressed GGHs was checked in
Arabidopsis by assaying GGH activity in vacuoles isolated
from wild-type leaves and those expressing AtGGH2-FL or
AtGGH2-TR. The truncated protein was predicted to be
cytosolic, as is usual for vacuolar proteins without their
targeting peptide (Frigerio et al., 1998). Vacuolar GGH
activity was about fourfold higher for AtGGH2-FL transfor-
mants than for the wild type, whereas that for AtGGH2-TR
transformants was the same as the wild type (Figure 2b).
-Glu tail
COOH
CH
COOH
CH2
CH2
NH
CH
COOH
CH2
CH2
COOH
n
C
O
O
C
C
N
NHNR
5
NH
NH2
O
CH2
9
HH
HNR′
10CHN
HCH2
CH2
NH
O
pABAPterin Glu
Folate R R′
THF
5-Methyl-THF
H H
CH3 H
5-Formimino-THF CH = NH
5-Formyl-THF
10-Formyl-THF
CHO H
H CHO
5, 10-Methylene-THF
5, 10-Methenyl-THF
-CH2-
-CH-
H
Figure 1. Structures of tetrahydrofolate (THF) polyglutamates and their
C1-substituted derivatives.
Folates are composed of pterin, p-aminobenzoate (pABA) and glutamate
moieties. A c-linked polyglutamate tail of up to eight residues is attached to
the first glutamate. One-carbon (C1) units at various oxidation levels are
attached to the N5 and/or N10 position of the pterin moiety. Folates readily
undergo oxidative cleavage of the C9–N10 bond yielding pterin and pABA-
glutamate moieties. Arrows indicate the bonds cleaved by c-glutamyl
hydrolase (GGH).
Gamma-glutamyl hydrolases and folate homeostasis 257
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

Moreover, when vacuolar protein extracts from both trans-
genic lines were probed for the His6 epitope, only those from
AtGGH2-FL gave positive signals (Figure 2b). The full-length
native protein was thus sent to the vacuole and the truncated
one was not. Since the fourfold increase in activity in the
AtGGH2-FL vacuoles is similar to that in whole-leaf extracts
it may be further concluded that native AtGGH2 was sent
solely to vacuoles. We observed no difference in GGH
activity in protoplasts isolated from the lines overexpressing
the native or truncated enzymes, suggesting that the
enhanced activity was intracellular in both cases.
Folate content and polyglutamylation status in plants
overexpressing GGH
To examine the relationship between GGH and folate
homeostasis, we measured total folate levels and the degree
of polyglutamylation in both Arabidopsis and tomato lines
with enhanced GGH activity. In Arabidopsis leaves and
tomato fruit expressing vacuole-targeted, native GGH, folate
levels dropped by 39 and 46%, respectively (Figure 3a). In
contrast, overexpression of the truncated AtGGH2 or
LeGGH2 did not significantly alter folate content. The major
folate species in wild-type Arabidopsis leaf tissue was
5,10-methenyl-THF (5,10 = CH-THF), whereas 5-CH3-THF
dominated in tomato fruit; these species were 70 and 90%
polyglutamylated, respectively (Figure 3c). The degree of
polyglutamylation of these folate species was severely
impaired only in lines expressing the native versions of GGH
(Figure 3b,c). In these plants, the trend toward an increas-
ingly monoglutamylated folate pool was at the expense of
the longer chain folates (six or more glutamates).
T-DNA insertion mutants of GGH and expression of the
Arabidopsis GGH family
The consequences of GGH inactivation were investigated in
Arabidopsis (Orsomando et al., 2005). We first used real-
time PCR to probe the expression of each family member in
various tissues (Figure 4a). AtGGH1 and AtGGH2 were
expressed in all tissues examined, with AtGGH1 predomi-
nating in roots and AtGGH2 in leaves. Conversely, AtGGH3
was expressed at very low levels in all tissues, suggesting
that the corresponding isoform makes little contribution to
total cellular GGH activity and is consequently not physio-
logically relevant. Moreover, when AtGGH3 was expressed
in Escherichia coli, we failed to detect any GGH activity (data
not shown).
Based on the premise that AtGGH1 and AtGGH2 are
essentially the sole contributors to Arabidopsis GGH activity,
two independent T-DNA mutant lines were obtained for each
gene (Figure 4b). Wild type segregants and plants homozy-
gous for the T-DNA insertion were identified by PCR using
primers flanking the insertion site along with a third primer
specific for the T-DNA left border (Figure 4c). The position of
the T-DNAs within the exon regions of each corresponding
Vac
uola
r G
GH
act
ivity
(nm
ol m
in–1
mg–
1 pr
otei
n)
0%
4%
10%
50 ─
37 ─
Anti-His6
TR FL
0
10
20
30
40
50
0
2
4
6
WT 7-8
WT
AtGGH2-
TR
AtGGH2-
FL
7-10 7-5 2-4 2-12 2-9
Arabidopsis
WT0
0.1
0.2
0.3
0.4
T-4 T-6 T-11
Wild type TR-GGH2
GG
H a
ctiv
ity(n
mol
min
–1 m
g–1
prot
ein)
GG
H a
ctiv
ity(n
mol
min
–1 m
g–1
prot
ein)
Tomato
FL-GGH2
F-2 F-4 F-14
(a)
(b)
Figure 2. Overexpression of c-glutamyl hydrolase (GGH) in tomato and
Arabidopsis.
(a) Overexpression of tomato and Arabidopsis cDNAs encoding full-length
GGH2 protein (FL-GGH2) or a truncated version missing the signal peptide
(TR-GGH2). Expression in tomato was driven by the fruit-specific E8
promoter and in Arabidopsis by the figwort mosaic virus 35S promoter.
Data for three independent lines exhibiting significantly (P < 0.05) higher
GGH activity in tomato fruit (T2 generation) and Arabidopsis leaf tissue (T3
generation) are presented. The GGH activity was assayed in desalted
tissue extracts from red tomato fruit or 4-week-old Arabidopsis rosette
leaf tissue. Specific activity values are the means � SE of three indepen-
dent experiments from three to five different plants from each indepen-
dent line.
(b) Arabidopsis vacuolar GGH activity in wild-type (WT) and AtGGH2
overexpressing lines. Vacuoles were purified on a Ficoll gradient and
harvested from the 0 to 4% Ficoll interface as shown. The neutral red dye
staining indicates the presence of intact vacuoles. Note the enhanced GGH
activity only in vacuoles from AtGGH2-FL lines. Vacuolar protein extracts
(8–10 lg) from AtGGH2-TR (TR) and AtGGH2-FL (FL) lines were also
subject to immunoblot analysis using antibodies against the His6 epitope
(inset). Equal protein loading was verified by staining the immunoblot with
Ponceau red. Specific activity values are the means � SE of three
independent vacuole preparations.
258 Tariq A. Akhtar et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266
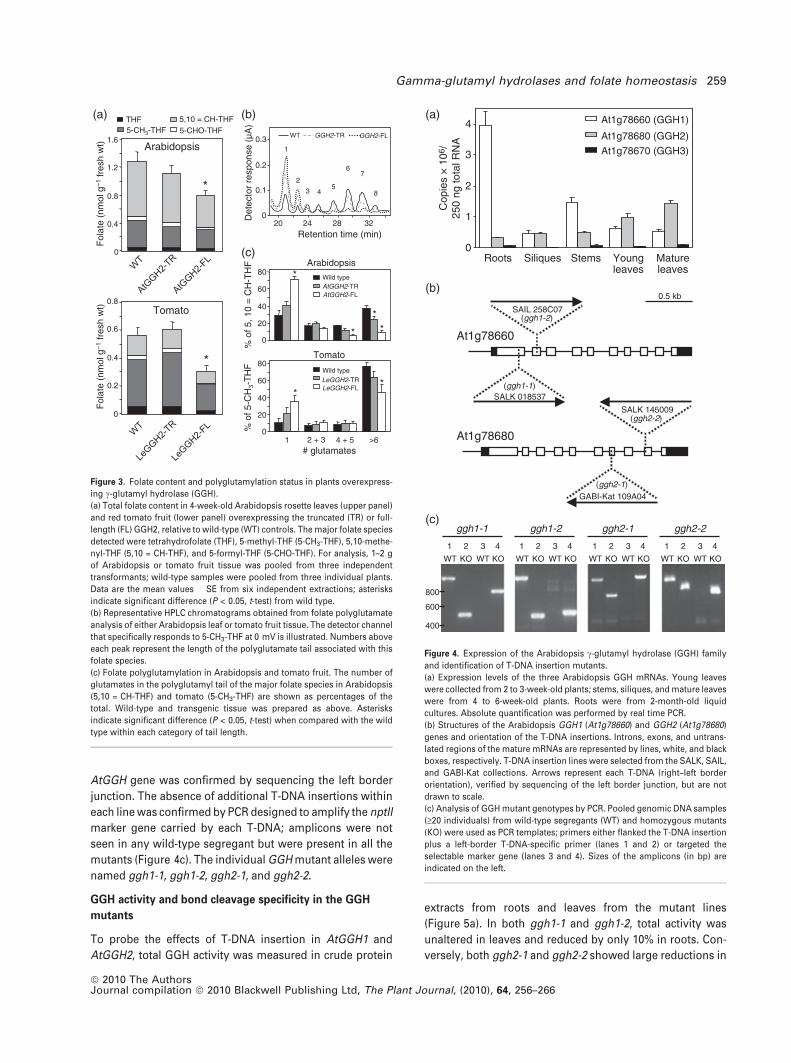
AtGGH gene was confirmed by sequencing the left border
junction. The absence of additional T-DNA insertions within
each line was confirmed by PCR designed to amplify the nptII
marker gene carried by each T-DNA; amplicons were not
seen in any wild-type segregant but were present in all the
mutants (Figure 4c). The individual GGH mutant alleles were
named ggh1-1, ggh1-2, ggh2-1, and ggh2-2.
GGH activity and bond cleavage specificity in the GGH
mutants
To probe the effects of T-DNA insertion in AtGGH1 and
AtGGH2, total GGH activity was measured in crude protein
extracts from roots and leaves from the mutant lines
(Figure 5a). In both ggh1-1 and ggh1-2, total activity was
unaltered in leaves and reduced by only 10% in roots. Con-
versely, both ggh2-1 and ggh2-2 showed large reductions in
Roots Siliques Stems Youngleaves
Matureleaves
0
1
2
3
4 At1g78660 (GGH1)
At1g78680 (GGH2)
At1g78670 (GGH3)
0.5 kb
SAIL 258C07
SALK 018537
At1g78660
SALK 145009
GABI-Kat 109A04
At1g78680
(ggh1-2)
(ggh1-1)
(ggh2-2)
(ggh2-1)
800
400
600
ggh1-1
41 2 3
WT KO WT KO
ggh2-2
1 2 3 4
WT KO WT KO
ggh2-1
1 2 3 4
WT KO WT KO
1 2 3 4
WT KO WT KO
ggh1-2
Cop
ies
× 1
06/
250
ng to
tal R
NA
(a)
(b)
(c)
Figure 4. Expression of the Arabidopsis c-glutamyl hydrolase (GGH) family
and identification of T-DNA insertion mutants.
(a) Expression levels of the three Arabidopsis GGH mRNAs. Young leaves
were collected from 2 to 3-week-old plants; stems, siliques, and mature leaves
were from 4 to 6-week-old plants. Roots were from 2-month-old liquid
cultures. Absolute quantification was performed by real time PCR.
(b) Structures of the Arabidopsis GGH1 (At1g78660) and GGH2 (At1g78680)
genes and orientation of the T-DNA insertions. Introns, exons, and untrans-
lated regions of the mature mRNAs are represented by lines, white, and black
boxes, respectively. T-DNA insertion lines were selected from the SALK, SAIL,
and GABI-Kat collections. Arrows represent each T-DNA (right–left border
orientation), verified by sequencing of the left border junction, but are not
drawn to scale.
(c) Analysis of GGH mutant genotypes by PCR. Pooled genomic DNA samples
(‡20 individuals) from wild-type segregants (WT) and homozygous mutants
(KO) were used as PCR templates; primers either flanked the T-DNA insertion
plus a left-border T-DNA-specific primer (lanes 1 and 2) or targeted the
selectable marker gene (lanes 3 and 4). Sizes of the amplicons (in bp) are
indicated on the left.
Det
ecto
r re
spon
se (
µA)
0
0.3
0.2
0.1
WT GGH2-TR GGH2-FL
1
2
3 45
67
8
20 24 28 32Retention time (min)
5-CH3-THFTHF 5,10 = CH-THF
5-CHO-THF
Fol
ate
(nm
ol g
–1 fr
esh
wt)
0
0.4
0.8
1.2
1.6Arabidopsis
*
Fol
ate
(nm
ol g
–1 fr
esh
wt)
0
0.2
0.4
0.6
0.8Tomato
*
# glutamates1 2 + 3 4 + 5 >6
LeGGH2-FL
Wild typeLeGGH2-TR
0
20
40
60
80
% o
f 5-C
H3-
TH
F
Tomato
**
% o
f 5, 1
0 =
CH
-TH
F
AtGGH2-TRAtGGH2-FL
Wild type
0
20
40
60
80Arabidopsis
*
* **
(a) (b)
(c)
Figure 3. Folate content and polyglutamylation status in plants overexpress-
ing c-glutamyl hydrolase (GGH).
(a) Total folate content in 4-week-old Arabidopsis rosette leaves (upper panel)
and red tomato fruit (lower panel) overexpressing the truncated (TR) or full-
length (FL) GGH2, relative to wild-type (WT) controls. The major folate species
detected were tetrahydrofolate (THF), 5-methyl-THF (5-CH3-THF), 5,10-methe-
nyl-THF (5,10 = CH-THF), and 5-formyl-THF (5-CHO-THF). For analysis, 1–2 g
of Arabidopsis or tomato fruit tissue was pooled from three independent
transformants; wild-type samples were pooled from three individual plants.
Data are the mean values � SE from six independent extractions; asterisks
indicate significant difference (P < 0.05, t-test) from wild type.
(b) Representative HPLC chromatograms obtained from folate polyglutamate
analysis of either Arabidopsis leaf or tomato fruit tissue. The detector channel
that specifically responds to 5-CH3-THF at 0 mV is illustrated. Numbers above
each peak represent the length of the polyglutamate tail associated with this
folate species.
(c) Folate polyglutamylation in Arabidopsis and tomato fruit. The number of
glutamates in the polyglutamyl tail of the major folate species in Arabidopsis
(5,10 = CH-THF) and tomato (5-CH3-THF) are shown as percentages of the
total. Wild-type and transgenic tissue was prepared as above. Asterisks
indicate significant difference (P < 0.05, t-test) when compared with the wild
type within each category of tail length.
Gamma-glutamyl hydrolases and folate homeostasis 259
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

GGH activity in both leaves and roots, averaging approxi-
mately 90 and 82%, respectively. Additionally, with primers
designed to measure expression of AtGGH1 or AtGGH2, no
mRNA signal in either corresponding GGH mutant plant
could be detected by semi-quantitative RT-PCR (Figure 5a).
These data show clearly that AtGGH2 is the main contributor
to total Arabidopsis GGH activity.
Because AtGGH1 and AtGGH2 exhibit distinct c-glutamate
bond cleavage patterns (Orsomando et al., 2005), their
individual contributions to total GGH activity can be mon-
itored in total protein extracts. Specifically, GGH1 cleaves
folate pentaglutamate (PteGlu5) to PteGlu2 and PteGlu3,
while GGH2 yields mainly PteGlu1 and a modest amount of
PteGlu2. On this basis, comparison of wild-type tissues
revealed that leaves contain almost exclusively GGH2 and
roots both GGH1 and GGH2 isozymes (Figure 5b). Likewise,
inspection of the cleavage patterns in mutant lines showed
that AtGGH1 and AtGGH2 mutants exhibited bond cleavage
profiles identical to recombinant AtGGH2 and AtGGH1,
respectively (Figure 5b). The reciprocal consistency of these
data further reinforces the probability that the third gene,
AtGGH3, makes little if any contribution to total GGH
activity.
RNA interference (RNAi)-mediated knockdown of residual
GGH in individual GGH knockouts
As the single GGH mutants retained substantial GGH activity
and the tandem organization of the Arabidopsis GGH genes
precludes generation of double mutants, we pursued RNAi-
mediated downregulation of AtGGH1 and AtGGH2 in the
ggh2-1 and ggh1-2 mutant backgrounds, respectively.
In ggh2-1 mutants targeted for AtGGH1 knockdown (At-
GGH1RNAi ggh2-1), three independent lines were obtained
that exhibited an average knockdown in AtGGH1 mRNA of
79%, relative to wild-type levels. Among three independent
lines targeted for AtGGH2 knockdown (AtGGH2RNAi ggh1-2),
AtGGH2 mRNA levels were reduced on average by 88%
(Figure 6a). In these lines with lowered GGH gene expres-
sion, total GGH activity in rosette leaf tissue was measured
and compared with the wild type (Figure 6b). The average
residual GGH activities in the three independent RNAi lines
targeting AtGGH1 and AtGGH2 were 0.008 and 0.02 nmol
min)1 mg)1 protein, respectively, representing 99 and 98%
knockdown in activity.
Total leaf folate content in the RNAi lines was measured
and compared with wild type and with the individual mutant
lines into which each RNAi construct was introduced.
Relative to wild-type plants, there were no significant
changes in total folate content in either ggh1-2 or ggh2-1
single mutants (Figure 6c) even though the latter had only
approximately 10% of wild-type GGH activity. However, both
AtGGH1RNAi ggh2-1 and AtGGH2RNAi ggh1-2 lines exhibited
significantly higher folate content (approximately 30%),
the increase being due mainly to 5,10 = CH-THF, the
PteGlu1 PteGlu2 PteGlu3
Extent of reaction
Rel
ativ
e pr
oduc
t con
cent
ratio
n 0 0.2 0.40
0.1
0.2
0.3Wild type root
0.10
0 0.05
0.025
0.05
0.075Wild type leaf
0 0.05 0.10
0.025
0.05
0.075GGH2 KO
0 0.20
0.1
0.2
0.3GGH1 KO
0.4
0 0.2 0.40
0.1
0.2
0.3
RecombinantGGH1
0 0.2 0.40
0.1
0.2
0.3
RecombinantGGH2
GG
H a
ctiv
ity(n
mol
min
–1 m
g–1
prot
ein)
ggh1-2 ggh2-1
KOWT
ggh2-2
WT KO
ggh1-1
KOWT KO WT
WT ggh1-1 ggh1-2 ggh2-1 ggh2-2
RootLeaf
0
1
2
10
8——
600 —500 —
* *
*
* *
*
(a)
(b)
Figure 5. c-Glutamyl hydrolase (GGH) activity and bond cleavage specificity
in the GGH mutants.
(a) Total GGH activity in wild-type (WT) and individual GGH mutant lines.
Roots were collected from 6 to 8-week-old plants grown in liquid medium and
leaf tissue was from rosette leaves from 4 to 6-week-old plants grown in
potting soil. Semiquantitative RT-PCR indicates the presence and absence of
functional mRNAs coding for GGH1 and GGH2 in WT and individual mutant
(KO) lines, respectively (inset). Data are the means and SE from three
independent experiments and asterisks indicate significant difference
(P < 0.05, t-test) when compared with the wild type.
(b) Specificity of GGH bond cleavage in wild-type and individual mutant lines.
The GGH activity was assayed in desalted tissue extracts. Progress curves for
the initial stages of hydrolysis are presented as plots of relative concentration
of each reaction product formed (folic acid monoglutamate, PteGlu1, folic acid
diglutamate, PteGlu2, or folic acid triglutamate, PteGlu3) versus the extent of
reaction (see Experimental Procedures). Progress curves from wild-type leaf
and root tissue (upper panel) are compared with those from the individual
GGH mutants (middle panel) and those from purified recombinant GGH1 and
GGH2 (lower panel). Data for the individual GGH mutants were derived from
pooled leaf and root tissue from the two GGH1 mutant lines (ggh1-1 and
ggh1-2) or from the two GGH2 mutant lines (ggh2-1 and ggh2-2). Data are
representative of results obtained in three independent experiments.
260 Tariq A. Akhtar et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

predominant folate species in rosette leaf tissue (Figure 3a).
The degree of polyglutamylation of this folate was strikingly
enhanced in both RNAi lines, where >96% of it was
polyglutamylated (Figure 6c).
Folate content and polyglutamylation status in Arabidopsis
leaf vacuoles
The increase in total folate content observed upon GGH
downregulation coupled with the decrease in total folate
resulting from GGH overexpression in its native vacuolar
location implies a significant role for the vacuole in folate
homeostasis. We therefore measured the vacuolar folate
content in Arabidopsis plants with up- or downregulated
GGH activity.
Intact vacuoles were isolated from leaf protoplasts
derived from plants overexpressing GGH (AtGGH2-FL), from
plants with 1% of normal GGH activity (AtGGH1RNAi ggh2-1),
and from wild-type controls. The total vacuolar folate
contents essentially mirrored those of whole-leaf extracts
from the same plants, i.e. vacuolar folate contents decreased
by 48% in AtGGH2-FL plants and increased by 42% in
AtGGH1RNAi ggh2-1 plants, relative to vacuoles from wild-
type plants (Figure 7a). In lines expressing cytosolic GGH
(AtGGH2-TR), no changes in vacuolar folate content or
degree of polyglutamylation were observed (not shown).
The two main folate species in all vacuoles isolated were
10-formyl-dihydrofolate (10-CHO-DHF), an oxidation
product routinely formed from 10-formyl-tetrahydrofolate
(10-CHO-THF) during organelle isolation (Goyer et al., 2005)
and 5-CH3-THF, which is the predominant folate in pea leaf
vacuoles (Orsomando et al., 2005). The extent of 5-CH3-THF
polyglutamylation was less in vacuoles from AtGGH2-FL
plants than from wild-type plants (33% versus 52%) but was
greater in vacuoles from AtGGH1RNAi ggh2-1 plants (>97%)
(Figure 7b).
DISCUSSION
The widely held notion that polyglutamylation has a critical
impact on folate homeostasis is only indirectly supported in
animal systems (Suh et al., 2001) and has not been explored
in plants at all. Additionally, the few studies in animal sys-
tems and bacteria that examined relationships between
GGH activity and folate content have yielded conflicting
results (Cole et al., 2001; Sybesma et al., 2003; Sakamoto
et al., 2008). This study accordingly aimed to assess whether
GGH contributes to the control of folate polyglutamylation
in vivo, and to clarify the relationship between the degree
of polyglutamylation and plant folate content.
Our results are consistent with a model (Figure 8) in which
folate polyglutamates are continuously imported into the
vacuole. The resulting vacuolar polyglutamate pool is
hydrolyzed by GGH, and the monoglutamates produced
are exported to the cytosol. This model parallels one
proposed for folate polyglutamate turnover in mammalian
AtGGH2RNAi ggh1-2
0
20
40
60
80
100
120
WT
AtGGH1 AtGGH2
AtGGH1RNAi ggh2-1R
elat
ive
expr
essi
on (
% W
T)
WT1–38 1–6 1–29 2–37 2–29 2–22
GG
H a
ctiv
ity (
nmol
min
–1 m
g–1 )
AtGGH1RNAi ggh2-1
AtGGH2RNAi ggh1-2
——
0.08
0
0.04
0.12
2.0
2.2
1–38 1–6 1–29 2–37 2–29 2–22
5, 10 = CH-THF
5-CHO-THF
Fol
ate
(nm
ol g
–1 fr
esh
wt)
THF
5-CH3-THF
0
0.5
1.0
1.5
2.0
2.5
**
% o
f 5, 1
0 =
CH
-TH
F
0
10
20
30
40
50
60
70
80
# glutamates
AtGGH1RNAi ggh2-1
Wild type
AtGGH2RNAi ggh1-2
1 2 + 3 4 + 5 > 6
*
*
*
**
** *
(a)
(b)
(c)
Figure 6. RNA interference (RNAi)-mediated knockdown of residual c-glut-
amyl hydrolase (GGH) in the individual GGH knockouts and its effect on folate
status.
(a) Transcript abundance of GGH1 (left) and GGH2 (right) in wild type (WT)
and three independent RNAi lines targeting GGH1 or GGH2 in the ggh2-1 or
ggh1-2 mutant backgrounds, respectively. Relative quantification of GGH
gene expression was by real-time PCR.
(b) Total GGH activity in the three RNAi lines described above. For compar-
ison, GGH activity in the wild type and in the individual GGH mutants is
presented (right panel).
(c) Folate content and polyglutamyl tail length distribution. Total folate and its
major forms [tetrahydrofolate (THF), 5-methyl-THF (5-CH3-THF), 5,10-methe-
nyl-THF (5,10 = CH-THF), 5-formyl-THF (5-CHO-THF)] in the wild type and
individual GGH mutants are compared with those in the RNAi lines (left
panel). Asterisks indicate significant difference (P < 0.05, t-test) when com-
pared with the wild type. Polyglutamyl tail length (right panel) as a percentage
of the major folate species (5,10 = CH-THF) in the same plants. Data are the
means � SE from six independent extractions from rosette leaf tissue
randomly sampled from three different plants for each genetic background.
For folate analysis of each RNAi line, leaf tissue from the three independent
lines described above was pooled. Asterisks indicate significant difference
(P < 0.05, t-test) when compared with the wild type within each category of
tail length.
Gamma-glutamyl hydrolases and folate homeostasis 261
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

lysosomes (Sirotnak and Tolner, 1999; Schneider and Ryan,
2006). Because monoglutamates are generally poor enzyme
substrates, they tend to remain unbound and are conse-
quently more susceptible to oxidative breakdown (Suh
et al., 2001; Gregory and Quinlivan, 2002). It might therefore
be expected that the release of monoglutamates from the
vacuole – and, by extension, GGH activity – is subject to strict
control. In support of this idea, Arabidopsis plants and
tomato fruit with just a threefold elevation of vacuolar GGH
activity contained approximately 40% less total folate, while
knockdown of Arabidopsis GGH activity caused a approxi-
mately 30% increase in folate content.
Our model raises several important issues. First, it invokes
a tonoplast transport system for polyglutamates. This would
be unlike all folate carriers so far described in plants, which
accept only monoglutamates (Bedhomme et al., 2005; Klaus
et al., 2005; Raichaudhuri et al., 2009). An analogous polyg-
lutamate transport system has, however, been demon-
strated in mammalian lysosomes, although its molecular
basis has not been elucidated (Barrueco and Sirotnak, 1991).
The model also envisions monoglutamate export from the
vacuole via a second transporter or perhaps via a single
transporter that exchanges mono- for polyglutamates. Nei-
ther the import nor export carrier appears to be identical
with the ATMRP1 vacuole transporter, since the latter is a
monoglutamate importer (Raichaudhuri et al., 2009).
A second issue is that overexpressing GGH in the cytosol
altered neither the degree of polyglutamylation nor the level
of folate. However, the cytosolic folate pool is relatively small
(Orsomando et al., 2005) and is probably protein bound, as in
animals. In animal cells, the concentration of folate-utilizing
enzymes is roughly equivalent to that of folates, implying a
very low proportion of free, unbound folate (Schirch and
Strong, 1989). Therefore, cytosolic GGH may simply have
been denied access to cytosolic folates, so that the rate of
deglutamylation was negligible. That little if any cytosolic
deglutamylation occurred as a result of cytosolic GGH
expression is supported by the observation that cytosolic
expression did not affect vacuolar folate polyglutamylation
or folate content. Assuming that vacuoles acquire polyglu-
tamates from the cytosol (Figure 8), a cytosolic polygluta-
mate deficit would have been mirrored in vacuoles.
Vacuole Cytosol
GGH
Folate-Glun
Folate-Glu1
Folate-Glun
2
3
Folate-Glun
Folate-Glu1
1
4
6
7
5
FPGS
Figure 8. A model for the role of c-glutamyl hydrolase (GGH) in folate
polyglutamate homeostasis.
Polyglutamates from the cytosol are continuously imported (1) into the
vacuole by a transport system. Vacuolar polyglutamates are either hydrolyzed
by GGH to the corresponding monoglutamates (2) or sequestered from GGH
attack and serve as potential storage forms of the cofactor (3). A second
transport system (4) exports monoglutamates from the vacuole to the cytosol
where they are routed to organelles (5) or polyglutamylated by folylpolyg-
lutamate synthetase (FPGS) (6). Polyglutamates represent the preferred
substrates for folate-dependent enzymes (7).
Monoglutamates
AtGGH2-FL
Wild type
AtGGH1RNAi
ggh2-1
Glu1 Glu2 Glu3 Glu4 Glu5+
Polyglutamates
Percent of 5-CH3-THF020406080100 20 40 60 80 100
0
2
4
6
8
10F
olat
e (n
mol
mg–
1 pr
otei
n)
THF 5-CH3-THF
5, 10 = CH + 10-CHO-THF
10-CHO-DHF
5-CHO-THF
*
*
(a)
(b)
Figure 7. Folate content and polyglutamylation status in Arabidopsis leaf
vacuoles.
(a) Total vacuolar folate content [tetrahydrofolate (THF), 5-methyl-THF (5-CH3-
THF), 10-formyl-dihydrofolate (10-CHO-DHF), 5-formyl-THF (5-CHO-THF),
10-formyl-THF (10-CHO-THF), 5,10-methenyl-THF (5,10 = CH-THF)] in the wild
type, c-glutamyl hydrolase (GGH2) overexpressing lines (FL-GGH2), and RNAi
lines (AtGGH1RNAi ggh2-1) with <1% of normal GGH activity.
(b) Vacuolar polyglutamyl tail length distribution. Data show the percentages
of vacuolar 5-CH3-THF that were polyglutamylated (2–6 glutamate residues) or
in the monoglutamyl form in the plants described above. Data are the
means � SE from three to six independent preparations from 4-week-old
rosette leaf tissue; asterisks indicate significant difference (P < 0.05, t-test)
compared with wild type.
262 Tariq A. Akhtar et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

A third and particularly enigmatic issue is why any
polyglutamates at all survive in the vacuole alongside
GGH, since vacuolar GGH activity in our experiments with
Arabidopsis (approximately 7 nmol min)1 mg)1 protein) is
sufficient to hydrolyze all vacuolar polyglutamates within
seconds, as was found in other studies (Orsomando et al.,
2005). Moreover, although tripling vacuolar GGH activity in
Arabidopsis lowered vacuolar polyglutamate levels, it did so
by only 19%. These observations imply that vacuolar
polyglutamates are in some way substantially protected
from attack by resident GGH. There is no evidence for plant
folate-binding proteins analogous to those in animals, for
vacuolar GGH inhibitors, or for rapid intravacuolar resyn-
thesis of polyglutamyl tails (Henderson, 1990; Scott et al.,
2000; Orsomando et al., 2005). An attractive alternative
possibility is that vacuolar folates are partially sequestered
in intravacuolar inclusions, such as those described for
anthocyanins (Pecket and Small, 1980).
Finally, the relationship established between polygluta-
mate tail length and folate content in this study is relevant to
metabolic engineering (‘biofortification’) of folate content in
plants (Bekaert et al., 2008). The strategies used to date have
all targeted the initial steps of the biosynthesis pathway, and
have not deliberately sought to manipulate polyglutamyla-
tion. In fact, however, engineering efforts in tomato and rice
resulted in enhanced folate content predominantly in mono-
glutamyl forms (Dıaz de la Garza et al., 2007; Storozhenko
et al., 2007). In theory, suppressing GGH activity could be
combined with engineering the synthesis pathway to pro-
mote vacuolar storage of polyglutamates.
EXPERIMENTAL PROCEDURES
Plant material and growth conditions
Wild-type Arabidopsis thaliana (Col-0 ecotype) seeds were fromLehle Seeds (http://www.arabidopsis.com/) and tomato (Solanumlycopersicum cv. Micro-Tom) seed were from the Ball seed com-pany (http://www.ballhort.com/). Arabidopsis and tomato plantswere grown in potting soil supplemented with Osmocote� in agrowth chamber at 22–25�C under a 12-h photoperiod (approxi-mately 80 and 200 lmol m)2 sec)1 fluence rate, respectively). Ara-bidopsis roots were obtained from liquid cultured plants grown inhalf-strength Murashige and Skoog medium containing 1% sucrose(Prabhu et al., 1996). Red and red-ripe tomato fruit were defined asbreaker plus 3 and 7 days, respectively.
Identification of Arabidopsis T-DNA mutants
Two GGH1 mutant lines, SALK 018537 and SAIL 258C07, and oneGGH2 mutant line, SALK 145009, were from the ArabidopsisBiological Resource Center (ABRC; http://abrc.osu.edu/). The GGH2mutant T-DNA insertion line GABI-Kat 109A04 was providedby Bernd Weisshaar (Max-Planck-Institute for Plant BreedingResearch, Cologne, Germany). Wild-type or homozygous mutantsegregants were identified by PCR analysis using gene-specificprimers flanking the T-DNA insertion and a T-DNA-specific primer.Genomic DNA was extracted according to Edwards et al. (1991).The insertion site was confirmed by sequencing the PCR ampliconobtained from screened mutant homozygotes. In each line, PCR
targeting the selectable marker gene (nptII for SALK lines, bar forSAIL lines, or dhps for GABI-Kat lines) on the T-DNA failed toamplify any products in wild-type segregants indicating theabsence of secondary T-DNA insertions. Homozygous mutantsand wild-type segregants were selfed and their progeny was usedfor experiments. All primers are listed in Table S1 in SupportingInformation.
Plasmids and constructs
Standard molecular methods were used to generate overexpressionand RNAi constructs, as summarized below. The pFMV, pHK1001,and pMON10086 vectors were kindly provided by Dr H. J. Klee(University of Florida, Gainesville, FL, USA); Arabidopsis andtomato cDNAs were as previously described (Orsomando et al.,2005; Akhtar et al., 2008). RNAi constructs targeting AtGGH1 andAtGGH2 were prepared as follows. From GGH1 cDNA, two PCRproducts corresponding to bases 532–863 and 532–1068 of the openreading frame (ORF) were amplified by high-fidelity PCR withprimers 1AB-5¢/1AB-3¢ and 1AC-5¢/1AC-3¢, respectively. The PCRproducts were digested with KpnI and ligated in a sense/antisenseorientation to form the hairpin construct targeting AtGGH1. Like-wise, primers 2AB-5¢/2AB-3¢ and 2AC-5¢/2AC-3¢ were used to PCR-amplify two fragments from AtGGH2 cDNA (corresponding to bases232–518 and 232–721 of the ORF, respectively), digested with KpnIand ligated to form the hairpin construct targeting AtGGH2. Eachhairpin was digested with BamHI/XbaI and ligated into the corre-sponding sites of pFMV, which places each hairpin under the con-trol of the constitutively active figwort mosaic virus (FMV) promoterand nopaline synthase (nos) terminator. To overexpress AtGGH2,the full-length AtGGH2 cDNA (At1g78680) was amplified usingprimers AtG2FLF/AtG2FLR, digested with NcoI/XhoI and ligatedinto the corresponding sites in pET28b (Novagen, http://www.emdchemicals.com/), which adds a C-terminal His6 epitope to thecorresponding protein. Using this construct as a template, a C-ter-minally tagged His6 full-length AtGGH2 sequence was amplifiedusing HISFLG2F/HISFLG2R, digested with XbaI/KpnI, and ligated tothe corresponding sites of pFMV. To overexpress a truncated ver-sion of AtGGH2 without the signal peptide, the short version of thiscDNA as described by Orsomando et al. (2005) was used as a tem-plate to amplify a N-terminally His6-tagged cDNA with FMVATG2F/FMVATG2R primers. This amplicon was digested with XbaI/KpnIand ligated to the corresponding sites of pFMV. The pFMV con-structs, containing either RNAi or overexpression cassettes, weredigested with NotI and ligated to the NotI-digested binary vectorpHK1001 and subsequently introduced into Agrobacterium tum-efaciens (ABI strain) by electroporation. To overexpress LeGGH2 intomato, full-length and truncated versions of LeGGH2 (Akhtar et al.,2008) were amplified using primers FLLEGGH2F/FLLEGGH2R orTRCLEGGH2F2/FLLEGGH2R, digested with NotI/AscI, and ligatedinto the corresponding sites of pMON10086. Each construct wasintroduced into A. tumefaciens (ABI strain) by electroporation.
Arabidopsis and tomato transformation
Transformation of wild-type or individual Arabidopsis GGH mutantswas performed by the ‘floral-dip’ method (Clough and Bent, 1998;Zhang et al., 2006). Kanamycin-resistant seedlings were recoveredfrom surface-sterilized T1 seeds plated on solid Murashige andSkoog medium containing 1% sucrose and 0.6% agar supplementedwith 50 mg L)1 kanamycin. T2 progenies segregating 3:1 (kanamy-cin-resistant:kanamycin-sensitive) were identified and resistantplants were screened for GGH activity (see below). Several inde-pendent lines with enhanced GGH activity were selfed and homo-zygous T3 plants exhibiting 100% resistance on selection medium
Gamma-glutamyl hydrolases and folate homeostasis 263
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

were used for analysis. Agrobacterium-mediated tomato transfor-mation was performed according to the method of Clough and Bent,1998. Transformants were selected and regenerated on mediumsupplemented with 100 mg L)1 kanamycin. Plantlets containingthe ProE8::LeGGH2-FL or ProE8::LeGGH2-TR construct wereidentified by PCR performed on genomic DNA prepared usingprimers located in the E8 promoter and within the coding region ofLeGGH2 (see Table S1). Independent primary transformants weretransferred to potting soil and those with T0 fruit exhibiting signif-icantly higher GGH activity were identified and selfed. Transgenic T1
plants were confirmed by PCR as above, and T1 fruit was used foranalysis.
GGH activity assay
Plant tissues were pulverized in liquid nitrogen and extracted in100 mM potassium phosphate, pH 6.0, containing 10% (v/v) glyc-erol, 10 mM b-mercaptoethanol, and 3% (w/v) polyvinylpyrrolidone(PVP-40). After centrifugation (20 000 g, 4�C, 20 min), supernatantswere desalted on PD-10 columns (Amersham Biosciences, http://www.gelifesciences.com) equilibrated in extraction buffer, withoutPVP-40. Protein was estimated by the method of Bradford (1976).The GGH assays were performed using 0.1 mM folic acid pentag-lutamate (PteGlu5) substrate as previously described (Orsomandoet al., 2005). The GGH activity was calculated as the sum of allproducts (from PteGlu1 to PteGlu4), linearly accumulating as afunction of time. The extent of hydrolysis and the relative concen-tration of each PteGlu reaction product were calculated and plottedaccording to Orsomando et al. (2005). The slope of the line corre-sponding to each product measures the relative extent of its for-mation, thus indicating the c-glutamyl bond specificity of theenzyme.
Quantitative real-time RT-PCR
Absolute quantification of individual GGH gene expression duringdevelopment was performed on the Applied Biosystems Gene-Amp 5700 PCR system (ABI, http://www.appliedbiosystems.com/).Total RNA from Arabidopsis tissues was extracted using theRNeasy Plant Mini Kit (Qiagen, http://www.qiagen.com/), andtreated with DNase (DNA-free� Kit, Ambion, http://www.ambi-on.com/) according to the manufacturer’s instructions. The RT-PCRreactions (25-ll final volumes) included the Taq-Man One-Step RT-PCR Master Mix reagents (ABI), plus 250 ng total RNA, 300 nM ofeach primer, and 150–250 nM of Taq-Man probe. Primers and Taq-Man probes were designed using the Applied Biosystems PRIMER
EXPRESS software. Standard curves for each GGH amplicon wereconstructed according to Goyer et al. (2004). Plasmid templateswere from RZPD (Deutsches Ressourcenzentrum fur Genomfors-chung GmbH, Berlin, Germany) for GGH1 (clone IDMPIZp2001J243Q, GenBank CF653065) and from ABRC for bothGGH2 (clone IDs 89D12, GenBank AY096428) and GGH3 (clone IDU12630, GenBank BT000554). In vitro transcription in the presenceof [5,6-3H]UTP was performed using the MAXIscript� kit (Ambion),according to the manufacturer’s instructions. Controls withoutreverse transcriptase were run in parallel to exclude genomic DNAcontamination. Relative quantification of GGH gene expression intransgenic Arabidopsis plants was conducted using an AppliedBiosystems 7300 Real-time PCR system (ABI). Total RNA wasextracted from 4-week-old rosette leaf tissue as above, except thatDNase treatment was performed using the RNase-Free DNase Set(Qiagen). The RNA samples were reverse transcribed using theHigh-Capacity cDNA Reverse Transcription Kit (ABI) with randomhexamers. The PCR employed ABI universal cycling conditionsusing SYBR GREEN PCR Master Mix (ABI) in a 25 ll reaction con-taining 6.25 ll of diluted cDNA (1:20) and a 300 nM concentration of
each primer. The UBI10 gene (At4g05320) served as an internalcontrol and expression values were calculated according to the2)DC¢T method (Livak and Schmittgen, 2001). SemiquantitativeRT-PCR was also performed on cDNA prepared as described aboveunder the following cycling conditions: 95�C for 10 min, 35 cyclesof 95�C for 1 min, 4�C or 58�C (depending on primer pairs for GGH1or GGH2, respectively) for 1 min, 72�C for 1 min, then 72�C for10 min. All primers are listed in Table S1.
Vacuole isolation and immunoblot analysis
Arabidopsis rosette leaf vacuoles were isolated as described byRobert et al. (2007), with the following modifications. Leaf tissuewas digested in 30-cm Petri plates containing protoplast enzymesolution without vacuum infiltration for a period of 3–4 h. Protop-lasts were recovered by passing digested tissue through a 70-lM
nylon mesh (Spectrum Laboratories, http://www.spectrumlabs.com/). The vacuolar marker enzyme, a-mannosidase, was assayedas described (Nok et al., 2000) to assess the relative enrichment andpurity of each vacuole preparation relative to the protoplasts fromwhich they were isolated. For immunoblot analysis, proteins from asingle vacuole preparation (approximately 8–10 lg) were precipi-tated using trichloroacetic acid and applied to pre-cast 2-amino-2-(hydroxymethyl)-1,3-propanediol (TRIS)-HCl gels formulated to15% acrylamide (Bio-Rad, http://www.bio-rad.com/). Followingelectrophoretic separation, proteins were transferred to polyviny-lidene difluoride membranes (Immobilon P, Millipore, http://www.millipore.com/) using a Trans-Blot SD Semi-Dry transfer system(Bio-Rad). Transfer efficiency was verified by staining the mem-branes with Ponceau S (Sigma, http://www.sigmaaldrich.com/).Blotted membranes were incubated in StartingBlock BlockingBuffer (Pierce Biotechnology, http://www.piercenet.com/) in TRIS-buffered saline (TBS; 20 mM TRIS–HCl, 150 mM NaCl, pH 7.5) con-taining 0.05% Tween 20 for 1 h, washed in TBS and incubatedovernight with anti-His5 antibody (1:1000) (Qiagen) in TBS at 4�C.Membranes were then washed in TBS containing 0.05% Tween 20(TBS-t) and incubated with horseradish peroxidase (HRP)-conju-gated anti-mouse IgG (1:2500) (Affinity Bioreagents, http://www.bioreagents.com/) for 1 h. Membranes were washed in TBS-t,rinsed in TBS, and HRP activity was detected using the ImmobilonWestern Chemiluminescent HRP Substrate (Millipore), according tothe manufacturer’s instructions. The chemiluminescent signal wascaptured with an Alpha Innotech Fluorchem SP imager (AlphaInnotech, http://www.alphainnotech.com/).
Folate analysis
Folates were extracted from leaf tissue (approximately 0.5 g) andred-ripe tomato fruit (approximately 2 g) by Polytron homogeniza-tion in 10–15 ml of 50 mM Na-HEPES, 50 mM N-cyclohexyl-2-aminoethanesulfonic acid (CHES), adjusted to pH 7.85 with HCl,containing 2% (w/v) sodium ascorbate, 1 mM CaCl2, and 10 mM
b-mercaptoethanol. Extracts were immediately incubated in a 100�Cwater bath for 10 min and then centrifuged (13,000 g, 15 min).Clarified crude extracts were split in two and treated plus or minus ratplasma conjugase to determine total folate or degree of polyglut-amylation. Folates were purified on folate-binding columns andanalyzed as previously described (Gregory and Toth, 1988). Forvacuolar folate analysis, the folate-binding column was scaled downto 1 ml and washing and elution buffers were reduced proportion-ately. Folates were quantified by HPLC with electrochemical detec-tion as previously described (Goyer et al., 2005; Orsomando et al.,2005). Detector response was calibrated with authentic THF, 5-for-myl-THF, 5-methyl-THF, 5,10-methenyl-THF, and 10-formyl-DHF,obtained from Schircks Laboratories (http://www.schircks.com/).
264 Tariq A. Akhtar et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

ACKNOWLEDGEMENTS
We thank Drs Denise Tieman and Mark Taylor for advice on tomatotransformation and construct design, Drs Valeria Naponelli andRocıo Dıaz de la Garza for help with folate analysis, Hazel Lees forArabidopsis vacuole analysis, and Drs Harry Klee and Kevin Folta forcritical reading of the manuscript. This project was supported byNational Science Foundation grant no. MCB-0443709 and by anendowment from the C. V. Griffin, Sr Foundation.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:Table S1. Synthetic oligonucleotides used in this study.Please note: As a service to our authors and readers, this journalprovides supporting information supplied by the authors. Suchmaterials are peer-reviewed and may be re-organized for onlinedelivery, but are not copy-edited or typeset. Technical supportissues arising from supporting information (other than missingfiles) should be addressed to the authors.
REFERENCES
Akhtar, T.A., McQuinn, R.P., Naponelli, V., Gregory, J.F. 3rd, Giovannoni, J.J.
and Hanson, A.D. (2008) Tomato gamma-glutamylhydrolases: expression,
characterization, and evidence for heterodimer formation. Plant Physiol.
148, 775–785.
Appling, D.R. (1991) Compartmentation of folate-mediated one-carbon
metabolism in eukaryotes. FASEB J. 5, 2645–2651.
Barrueco, J.R. and Sirotnak, F.M. (1991) Evidence for the facilitated transport
of methotrexate polyglutamates into lysosomes derived from S180 cells.
J. Biol. Chem. 266, 11732–11737.
Bedhomme, M., Hoffmann, M., McCarthy, E.A., Gambonnet, B., Moran, R.G.,
Rebeille, F. and Ravanel, S. (2005) Folate metabolism in plants. An Ara-
bidopsis homolog of the mammalian mitochondrial folate transporter
mediates folate import into chloroplasts. J. Biol. Chem. 280, 34823–34831.
Bekaert, S., Storozhenko, S., Mehrshahi, P., Bennett, M.J., Lambert, W.,
Gregory, J.F. 3rd, Schubert, K., Hugenholtz, J., Van der Straeten, D. and
Hanson, A.D. (2008) Folate biofortification in food plants. Trends Plant Sci.
13, 28–35.
Bradford, M.M. (1976) A rapid and sensitive method for the quantification of
microgram quantities of protein utilizing the principle of protein dye
binding. Anal. Biochem. 72, 248–254.
Chen, L., Qi, H., Korenberg, J., Garrow, T.A., Choi, Y.J. and Shane, B. (1996)
Purification and properties of human cytosolic folylpoly-gamma-glutamate
synthetase and organization, localization, and differential splicing of its
gene. J. Biol. Chem. 271, 13077–13087.
Cherest, H., Thomas, D. and Surdin-Kerjan, Y. (2000) Polyglutamylation of
folate coenzymes is necessary for methionine biosynthesis and mainte-
nance of intact mitochondrial genome in Saccharomyces cerevisiae.
J. Biol. Chem. 275, 14056–14063.
Clough, S.J. and Bent, A.F. (1998) Floral dip: a simplified method for Agro-
bacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16,
735–743.
Cole, P.D., Kamen, B.A., Gorlick, R., Banerjee, D., Smith, A.K., Magill, E. and
Bertino, J.R. (2001) Effects of overexpression of c-glutamyl hydrolase on
methotrexate metabolism and resistance. Cancer Res. 61, 4599–4604.
Dıaz de la Garza, R., Gregory, J.F. 3rd and Hanson, A.D. (2007) Folate bio-
fortification of tomato fruit. Proc. Natl Acad. Sci. USA, 104, 4218–4222.
Edwards, K., Johnstone, C. and Thompson, C. (1991) A simple and rapid
method for the preparation of plant genomic DNA for PCR analysis. Nucleic
Acids Res. 19, 1349.
Frigerio, L., Vitale, A., Lord, J.M., Ceriotti, A. and Roberts, L.M. (1998) Free
ricin A chain, proricin, and native toxin have different cellular fates when
expressed in tobacco protoplasts. J. Biol. Chem. 273, 14194–14199.
Goyer, A., Johnson, T.L., Olsen, L.J., Collakova, E., Shachar-Hill, Y., Rhodes,
D. and Hanson, A.D. (2004) Characterization and metabolic function of a
peroxisomal sarcosine and pipecolate oxidase from Arabidopsis. J. Biol.
Chem. 280, 26137–26142.
Goyer, A., Collakova, E., Dıaz de la Garza, R., Quinlivan, E.P., Williamson, J.,
Gregory, J.F. 3rd, Shachar-Hill, Y. and Hanson, A.D. (2005) 5-formyltetra-
hydrofolate is an inhibitory but well tolerated metabolite in Arabidopsis
leaves. J. Biol. Chem. 280, 26137–26142.
Gregory, F.F. 3rd and Quinlivan, E.P. (2002) In vivo kinetics of folate metab-
olism. Annu. Rev.Nutr. 22, 199–220.
Gregory, J.F. 3rd and Toth, J.P. (1988) Chemical synthesis of deuterated folate
monoglutamate and in vivo assessment of urinary excretion of deuterated
folates in man. Anal. Biochem. 170, 94–104.
Hanson, A.D. and Roje, S. (2001) One-carbon metabolism in higher plants.
Annu. Rev. Plant Physiol. Plant Mol. Biol. 52, 119–137.
Henderson, G.B. (1990) Folate-binding proteins. Annu. Rev. Nutr. 10, 319–335.
Klaus, S., Kunji, E.R.S., Bozzo, G.G., Noiriel, A., Dıaz de la Garza, R., Basset,
G.J.C., Ravanel, S., Rebeille, F., Gregory, J.F. 3rd and Hanson, A.D. (2005)
Higher plant plastids and cyanobacteria have folate carriers related to those
of trypanosomatids. J. Biol. Chem. 280, 38457–38463.
Kneissl, M.L. and Deikman, J. (1996) The tomato E8 gene influences ethylene
biosynthesis in fruit but not flowers. Plant Physiol. 112, 537–547.
Li, W.W., Waltham, M., Tong, W., Schweitzer, B.I. and Bertino, J.R. (1993)
Increased activity of c-glutamyl hydrolase in human sarcoma cell lines: a
novel mechanism of intrinsic resitance to methotrexate (MTX). Adv. Exp.
Med. Biol. 338, 635–638.
Livak, K.J. and Schmittgen, T.D. (2001) Analysis of relative gene expression
data using real-time quantitative PCR and the 2)DDCT method. Methods, 25,
402–408.
Lowe, K.E., Osborne, C.B., Lin, B.F., Kim, J.S., Hsu, J.C. and Shane, B. (1993)
Regulation of folate and one-carbon metabolism in mammalian cells II.
Effect of folylpoly-c-glutamate synthetase substrate specificity and level on
folate metabolism and folylpoly-c-glutamate specificity of metabolic cycles
of one-carbon metabolism. J. Biol. Chem. 268, 21665–21673.
Nok, A.J., Shuaibu, M.N., Kanbara, H. and Yanagi, T. (2000) Purification and
partial characterization of alpha-mannosidase from Trypanosoma rangeli.
Parisitol. Res. 86, 923–928.
Orsomando, G., Dıaz de la Garza, R., Green, B.J., Peng, M., Rea, P.A., Ryan,
T.J., Gregory, J.F. 3rd and Hanson, A.D. (2005) Plant gamma-glutamyl
hydrolases and folate polyglutamates: characterization, compartmenta-
tion, and co-occurrence in vacuoles. J. Biol. Chem. 280, 28877–28884.
Orsomando, G., Bozzo, G.G., Dıaz de la Garza, R., Basset, G.J., Quinlivan, E.P.,
Naponelli, V., Rebeille, F., Ravanel, S., Gregory, J.F. 3rd and Hanson, A.D.
(2006) Evidence for folate-salvage reactions in plants. Plant J. 46, 426–435.
Pecket, C.R. and Small, C.J. (1980) Occurrence, location, and development of
anthocyanoplasts. Phytochemistry, 19, 2571–2576.
Prabhu, V., Chatson, K.B., Abrams, G.D. and King, J. (1996) 13C nuclear
magnetic resonance detection of interactions of serine hydroxymethyl-
transferase with C1-tetrahydrofolate synthase and glycine decarboxylase
complex activities in Arabidopsis. Plant Physiol. 112, 207–216.
Raichaudhuri, A., Peng, M., Naponelli, V., Sanchez-Fernandez, R., Gu, H.,
Gregory, J.F. 3rd, Hanson, A.D. and Rea, P.A. (2009) Plant vacuolar ATP-
binding cassette transporters that translocate folates and antifolates
in vitro and contribute to antifolate tolerance in vivo. J. Biol. Chem. 284,
8449–8460.
Ravanel, S., Cherest, H., Jabrin, S., Grunwald, D., Surdin-Kerjan, Y., Douce, R.
and Rebeille, F. (2001) Tetrahydrofolate biosynthesis in plants: molecular
and functional characterization of dihydrofolate synthetase and three iso-
forms of folylpolyglutamate synthetase in Arabidopsis thaliana. Proc. Natl
Acad. Sci. USA, 98, 15360–15365.
Ravanel, S., Block, M.A., Rippert, P., Jabrin, S., Curien, G., Rebeille, F. and
Douce, R. (2004) Methionine metabolism in plants. Chloroplasts are
autonomous for de novo methionine synthesis and can import S-adeno-
sylmethionine from the cytosol. J. Biol. Chem. 279, 22548–22557.
Rebeille, F., Ravanel, S., Jabrin, S., Douce, R., Storozhenko, S. and Van der
Straeten, D. (2006) Folates in plants: biosynthesis, distribution, and
enhancement. Physiol. Plant. 126, 330–342.
Rhee, M.S., Wang, Y., Gopel Nair, M. and Galivan, J. (1993) Acquisition of
resistance to antifolates caused by enhanced c-glutamyl hydrolase activity.
Cancer Res. 53, 2227–2230.
Richins, R.D., Scholthof, H.B. and Shepard, R.J. (1987) Sequence of figwort
mosaic virus DNA (caulimovirus group). Nucleic Acids Res. 15, 8451–8466.
Robert, S., Zouhar, J., Carter, C. and Raikhel, N. (2007) Isolation of intact
vacuoles from Arabidopsis rosette leaf-derived protoplasts. Nat. Protoc. 2,
259–262.
Gamma-glutamyl hydrolases and folate homeostasis 265
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266

Sakamoto, E., Tsukioka, S., Oie, S. et al. (2008) Folylpolglutamate synthase
and c-glutamyl hydrolase regulate leucovorin-enhanced 5-fluorouracil
anticancer activity. Biochem. Biophys. Res. Commun. 365, 801–807.
Schirch, V. and Strong, W.B. (1989) Interaction of folylpolyglutamates with
enzymes in one-carbon metabolism. Arch. Biochem. Biophys. 269, 371–380.
Schneider, E. and Ryan, T.J. (2006) Gamma-glutamyl hydrolase and drug
resistance. Clin. Chim. Acta 374, 25–32.
Scott, J., Rebeille, F. and Fletcher, J. (2000) Folic acid and folates: the feasi-
bility for nutritional enhancement in plant foods. J. Sci. Food. Agric. 80,
795–824.
Shane, B. (1989) Folylpolyglutamate synthesis and role in the regulation of
one-carbon metabolism. Vitam. Horm. 45, 263–335.
Shane, B. and Stokstad, E.L. (1975) Transport and metabolism of folates by
bacteria. J. Biol. Chem. 250, 2243–2253.
Sirotnak, F.M. and Tolner, B. (1999) Carrier-mediated membrane transport of
folates in mammalian cells. Annu. Rev. Nutr. 19, 91–122.
Storozhenko, S., De Brouwer, V., Volckaert, M., Navarrete, O., Blancquaert,
D., Zhang, G.F., Lambert, W. and Van Der Straeten, D. (2007) Folate
fortification of rice by metabolic engineering. Nat. Biotechnol. 25, 1277–
1279.
Suh, J.R., Herbig, A.K. and Stover, P.J. (2001) New perspectives on folate
catabolism. Annu. Rev. Nutr. 21, 255–282.
Sybesma, W., Van Den Born, E., Starrenburg, M., Mierau, I., Kleerebezem, M.,
De Vos, W.M. and Hugenholtz, J. (2003) Controlled modulation of folate
polyglutamyl tail length by metabolic engineering of Lactococcus lactis.
Appl. Environ. Microbiol. 69, 7101–7107.
Wang, Y., Nimec, Z., Ryan, T.J., Dias, J.A. and Galivan, J. (1993) The proper-
ties of the secreted gamma-glutamyl hydrolases from H35 hepatoma cells.
Biochim. Biophys. Acta 1164, 227–235.
Whitfield, C.D., Steers, E.J. and Weissbach, H. (1970) Purification and prop-
erties of 5-methyltetrahydropteroyltriglutamate-homocysteine transmeth-
ylase. J. Biol. Chem. 245, 390–401.
Zhang, X., Henriques, R., Lin, S.S., Nie, Q.W. and Chua, N.H. (2006)
Agrobacterium-mediated transformation of Arabidopsis thaliana using the
floral dip method. Nat. Protoc. 1, 641–646.
266 Tariq A. Akhtar et al.
ª 2010 The AuthorsJournal compilation ª 2010 Blackwell Publishing Ltd, The Plant Journal, (2010), 64, 256–266




















