
2-Cyano-lup-1-en-3-oxo-20-oic acid, a cyano derivative of betulinic acid, activates peroxisome...
10
Carcinogenesis vol.28 no.11 pp.2337–2346, 2007 doi:10.1093/carcin/bgm189 Advance Access publication August 27, 2007 2-Cyano-lup-1-en-3-oxo-20-oic acid, a cyano derivative of betulinic acid, activates peroxisome proliferator-activated receptor g in colon and pancreatic cancer cells Sudhakar Chintharlapalli 1,y , Sabitha Papineni 1,y , Shengxi Liu 1 , Indira Jutooru 2 , Gayathri Chadalapaka 2 , Sung-dae Cho 1 , Rajesh S.Murthy 3 , Youngjae You 3 and Stephen Safe 1,2, 1 Institute of Biosciences and Technology, Texas A&M University Health Science Center,Houston, TX 77030-3303, USA, 2 Department of Veterinary Physiology and Pharmacology, Texas A&M University, College Station,TX 77843-4466, USA and 3 Department of Chemistry, South Dakota State University, Brookings, SD 57007, USA To whom correspondence should be addressed. Tel: þ979 845 5988; Fax: þ979 862 4929; Email: [email protected] Betulinic acid (BA) is a phytochemical triterpenoid acid from bark extracts and is cytotoxic to cancer cells and tumors. We modified the A-ring of BA to give a 2-cyano-1-en-3-one moiety and the effects of the 2-cyano-lup-1-en-3-oxo-20-oic acid (CN-BA), 2-cyano derivative of BA, and its methyl ester (CN-BA-Me) were investigated in colon and pancreatic cancer cells. Both CN-BA and CN-BA-Me were highly cytotoxic to Panc-28 pancreatic and SW480 colon cancer cells. CN-BA and CN-BA-Me also induced differentiation in 3T3-L1 adipocytes, which exhibited a character- istic fat droplet accumulation induced by peroxisome proliferator- activated receptor g (PPARg) agonists. Based on these results, we investigated the activities of CN-BA and CN-BA-Me as PPARg agonists using several receptor-mediated responses including ac- tivation of transfected PPARg-responsive constructs, induction of p21 in Panc-28 cells and induction of caveolin-1 and Kru ¨ ppel-like factor 4 in colon cancer cells. The results clearly demonstrated that both CN-BA and CN-BA-Me activated PPARg-dependent re- sponses in colon (caveolin-1) and pancreatic (p21) cancer cells, whereas induction of KLF4 by these compounds in colon cancer cells was PPARg independent and also dependent on cell context. The PPARg agonist activities of CN-BA and CN-BA-Me were struc- ture-, response/gene- and cell context-dependent suggesting that these compounds are a novel class of selective PPARg modulators with potential for clinical treatment of colon and pancreatic cancer. Introduction Lup-20(29)-ene-3b,28-diol (betulin) is a triterpene natural product found in extracts of many bushes and trees, and betulin can constitute up to 30% of the dry weight of bark from birch trees (1,2). Betulin has been used in folk medicine for treating skin diseases; however, betu- linic acid (BA), which is both a natural product and chemical oxida- tion product of betulin, induces a broad range of pharmacological activities. BA and several derivatives exhibit anticancer activity, in- hibit human immuno virus and other viruses through multiple path- ways, are effective anti-bacterial and anti-malarial drugs and exhibit anti-inflammatory activity (1,2). The antitumorigenic activities of BA have been extensively investigated. Studies show that this compound inhibits tumor growth through multiple pathways and these responses are also cancer cell/tumor dependent (1–16). Pisha et al. (3) reported that BA selectively inhibited melanoma cancer cell and tumor growth and in in vivo studies this was accompanied by minimal toxic side effects at repeated doses of up to 500 mg/kg. Subsequent studies showed that BA was cytotoxic to many other cancer cell lines and this was associated with different activities (4–16). For example, BA induces apoptosis through decreased mitochondrial membrane poten- tial, activation of mitogen-activated protein kinase and modulation of nuclear factor jB (4,5,14). Structural modifications of BA and other lupane-derived triterpe- noids differentially affect their pharmacologic activities (17–23). For example, modification of the C-20 exocyclic position of BA did not affect the cytotoxicity of these derivatives to a panel of prostate and colon cancer and melanoma cell lines (17). In contrast, A-ring mod- ifications of BA containing a 1-ene-3-oxo moiety substituted at C-2 with electron-withdrawing groups were highly cytotoxic (19). These results were similar to ursane and oleanane triterpenoid acids where analogs containing electron-withdrawing substituents at C-2 within a 1-ene-3-one functionality were also highly cytotoxic to cancer cells compared with the parent acids (24–28). Typical among these synthetic derivatives were 2-cyano-3,12-dioxo-18b-oleana-1,19- diene-28-oic acid (CDDO; synthesized from oleanolic acid) and 2-cyano-3,12-dioxo-18b-olean-1,12-diene-30-oic acid (b-CDODA; synthesized from glycyrrhetinic acid, a major constituent of licorice extracts). The high cytotoxicity of b-CDODA, CDDO and related com- pounds was due, in part, to their peroxisome proliferator-activated receptor c (PPARc) agonist activity and ligands for this receptor are being developed as new anticancer drugs (29,30). Although there are multiple structural differences between BA and oleanolic acid (the synthetic precursor of CDDO and CDODA) (Figure 1), we hypothesized that introduction of a 2-cyano group into the lupane skeleton of BA would generate a new class of PPARc agonists. Previous studies showed that 2-cyano-lup-1-en-3-oxo-20- oic acid (CN-BA), the 2-cyano derivative of 20(29)-dihydro BA, was highly cytotoxic to cancer cells, and in this study, we compared the effects of BA, CN-BA and the corresponding methyl ester (CN- BA-Me) (Figure 1) in Panc-28 pancreatic and colon cancer cell lines. Results of growth inhibition studies showed that both CN-BA and CN-BA-Me were more cytotoxic than BA in pancreatic and colon cancer cells. CN-BA and CN-BA-Me but not BA induced PPARc- dependent transactivation; however, the receptor-dependent induction of p21, caveolin1 and Kru ¨ppel-like factor 4 expression was cell con- text and gene dependent. These results demonstrate for the first time that CN-BA and CN-BA-Me are PPARc agonists and their enhanced cytotoxicity compared with BA is due, at least in part, to activation of PPARc. Moreover, the structure- and cell context-dependent activities of CN-BA and CN-BA-Me as PPARc agonists suggest that these compounds are selective PPARc modulators. Materials and methods Cell lines and reagents SW480, HT-29 and HCT-15 human colon cancer cells were kindly provided by Dr Stan Hamilton (M.D.Anderson Cancer Center, Houston, TX). Panc-28 human pancreatic cancer cells and 3T3-L1 pre-adipocytes were obtained from American Type Culture Collection (Manassas, VA). SW480, HT-29 and Panc- 28 cells were maintained in Dulbecco’s modified/Ham’s F-12 (Sigma–Aldrich, St Louis, MO) with phenol red supplemented with 0.22% sodium bicarbonate, 0.011% sodium pyruvate, 5% fetal bovine serum and 10 ml/l 100 antibiotic anti-mycotic solution (Sigma). HCT-15 cells were maintained in RPMI 1640 (Sigma) supplemented with 0.22% sodium bicarbonate, 0.011% sodium pyru- vate, 0.45% glucose, 0.24% N-2-hydroxyethylpiperazine-N#-2-ethanesulfonic acid, 10% fetal bovine serum and 10 ml/l of 100 antibiotic anti-mycotic solution (Sigma). Cells were maintained at 37°C in the presence of 5% CO 2 . Reporter lysis buffer and luciferase reagent for luciferase studies were supplied Abbreviations: BA, betulinic acid; C-DIM, methylene-substituted diindolyl- methanes; CDDO, 2-cyano-3,12-dioxo-18b-oleana-1,19-diene-28-oic acid; b- CDODA, 2-cyano-3,12-dioxo-18b-olean-1,12-diene-30-oic acid; CN-BA, 2- cyano-lup-1-en-3-oxo-20-oic acid; CN-BA-Me, methyl ester of CN-BA; DMEM, Dulbecco’s modified Eagle’s medium; DMSO, dimethyl sulfoxide; FBS, fetal bovine serum; b-GAL, b-Galactosidase; PPARc, peroxisome pro- liferator-activated receptor c; TBST, Tris-buffered saline containing Tween-20. y Both authors contributed equally to this work. Ó The Author 2007. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] 2337 by guest on December 12, 2013 http://carcin.oxfordjournals.org/ Downloaded from
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of 2-Cyano-lup-1-en-3-oxo-20-oic acid, a cyano derivative of betulinic acid, activates peroxisome...

Carcinogenesis vol.28 no.11 pp.2337–2346, 2007doi:10.1093/carcin/bgm189Advance Access publication August 27, 2007
2-Cyano-lup-1-en-3-oxo-20-oic acid, a cyano derivative of betulinic acid, activatesperoxisome proliferator-activated receptor g in colon and pancreatic cancer cells
Sudhakar Chintharlapalli1,y, Sabitha Papineni1,y,Shengxi Liu1, Indira Jutooru2, Gayathri Chadalapaka2,Sung-dae Cho1, Rajesh S.Murthy3, Youngjae You3 andStephen Safe1,2,�
1Institute of Biosciences and Technology, Texas A&M University HealthScience Center,Houston, TX 77030-3303, USA, 2Department of VeterinaryPhysiology and Pharmacology, Texas A&M University, College Station, TX77843-4466, USA and 3Department of Chemistry, South Dakota StateUniversity, Brookings, SD 57007, USA
�To whom correspondence should be addressed. Tel: þ979 845 5988;Fax: þ979 862 4929;Email: [email protected]
Betulinic acid (BA) is a phytochemical triterpenoid acid frombark extracts and is cytotoxic to cancer cells and tumors. Wemodified the A-ring of BA to give a 2-cyano-1-en-3-one moietyand the effects of the 2-cyano-lup-1-en-3-oxo-20-oic acid (CN-BA),2-cyano derivative of BA, and its methyl ester (CN-BA-Me) wereinvestigated in colon and pancreatic cancer cells. Both CN-BA andCN-BA-Me were highly cytotoxic to Panc-28 pancreatic andSW480 colon cancer cells. CN-BA and CN-BA-Me also induceddifferentiation in 3T3-L1 adipocytes, which exhibited a character-istic fat droplet accumulation induced by peroxisome proliferator-activated receptor g (PPARg) agonists. Based on these results, weinvestigated the activities of CN-BA and CN-BA-Me as PPARgagonists using several receptor-mediated responses including ac-tivation of transfected PPARg-responsive constructs, induction ofp21 in Panc-28 cells and induction of caveolin-1 and Kruppel-likefactor 4 in colon cancer cells. The results clearly demonstratedthat both CN-BA and CN-BA-Me activated PPARg-dependent re-sponses in colon (caveolin-1) and pancreatic (p21) cancer cells,whereas induction of KLF4 by these compounds in colon cancercells was PPARg independent and also dependent on cell context.The PPARg agonist activities of CN-BA andCN-BA-Mewere struc-ture-, response/gene- and cell context-dependent suggesting thatthese compounds are a novel class of selective PPARg modulatorswith potential for clinical treatment of colon and pancreatic cancer.
Introduction
Lup-20(29)-ene-3b,28-diol (betulin) is a triterpene natural productfound in extracts of many bushes and trees, and betulin can constituteup to 30% of the dry weight of bark from birch trees (1,2). Betulin hasbeen used in folk medicine for treating skin diseases; however, betu-linic acid (BA), which is both a natural product and chemical oxida-tion product of betulin, induces a broad range of pharmacologicalactivities. BA and several derivatives exhibit anticancer activity, in-hibit human immuno virus and other viruses through multiple path-ways, are effective anti-bacterial and anti-malarial drugs and exhibitanti-inflammatory activity (1,2). The antitumorigenic activities of BAhave been extensively investigated. Studies show that this compoundinhibits tumor growth through multiple pathways and these responsesare also cancer cell/tumor dependent (1–16). Pisha et al. (3) reported
that BA selectively inhibited melanoma cancer cell and tumor growthand in in vivo studies this was accompanied by minimal toxic sideeffects at repeated doses of up to 500 mg/kg. Subsequent studiesshowed that BA was cytotoxic to many other cancer cell lines andthis was associated with different activities (4–16). For example, BAinduces apoptosis through decreased mitochondrial membrane poten-tial, activation of mitogen-activated protein kinase and modulation ofnuclear factor jB (4,5,14).
Structural modifications of BA and other lupane-derived triterpe-noids differentially affect their pharmacologic activities (17–23). Forexample, modification of the C-20 exocyclic position of BA did notaffect the cytotoxicity of these derivatives to a panel of prostate andcolon cancer and melanoma cell lines (17). In contrast, A-ring mod-ifications of BA containing a 1-ene-3-oxo moiety substituted at C-2with electron-withdrawing groups were highly cytotoxic (19). Theseresults were similar to ursane and oleanane triterpenoid acids whereanalogs containing electron-withdrawing substituents at C-2 withina 1-ene-3-one functionality were also highly cytotoxic to cancercells compared with the parent acids (24–28). Typical among thesesynthetic derivatives were 2-cyano-3,12-dioxo-18b-oleana-1,19-diene-28-oic acid (CDDO; synthesized from oleanolic acid) and2-cyano-3,12-dioxo-18b-olean-1,12-diene-30-oic acid (b-CDODA;synthesized from glycyrrhetinic acid, a major constituent of licoriceextracts). The high cytotoxicity of b-CDODA, CDDO and related com-pounds was due, in part, to their peroxisome proliferator-activatedreceptor c (PPARc) agonist activity and ligands for this receptor arebeing developed as new anticancer drugs (29,30).
Although there are multiple structural differences between BAand oleanolic acid (the synthetic precursor of CDDO and CDODA)(Figure 1), we hypothesized that introduction of a 2-cyano group intothe lupane skeleton of BA would generate a new class of PPARcagonists. Previous studies showed that 2-cyano-lup-1-en-3-oxo-20-oic acid (CN-BA), the 2-cyano derivative of 20(29)-dihydro BA,was highly cytotoxic to cancer cells, and in this study, we comparedthe effects of BA, CN-BA and the corresponding methyl ester (CN-BA-Me) (Figure 1) in Panc-28 pancreatic and colon cancer cell lines.Results of growth inhibition studies showed that both CN-BA andCN-BA-Me were more cytotoxic than BA in pancreatic and coloncancer cells. CN-BA and CN-BA-Me but not BA induced PPARc-dependent transactivation; however, the receptor-dependent inductionof p21, caveolin1 and Kruppel-like factor 4 expression was cell con-text and gene dependent. These results demonstrate for the first timethat CN-BA and CN-BA-Me are PPARc agonists and their enhancedcytotoxicity compared with BA is due, at least in part, to activation ofPPARc. Moreover, the structure- and cell context-dependent activitiesof CN-BA and CN-BA-Me as PPARc agonists suggest that thesecompounds are selective PPARc modulators.
Materials and methods
Cell lines and reagents
SW480, HT-29 and HCT-15 human colon cancer cells were kindly provided byDr Stan Hamilton (M.D.Anderson Cancer Center, Houston, TX). Panc-28human pancreatic cancer cells and 3T3-L1 pre-adipocytes were obtained fromAmerican Type Culture Collection (Manassas, VA). SW480, HT-29 and Panc-28 cells were maintained in Dulbecco’s modified/Ham’s F-12 (Sigma–Aldrich,St Louis, MO) with phenol red supplemented with 0.22% sodium bicarbonate,0.011% sodium pyruvate, 5% fetal bovine serum and 10 ml/l 100� antibioticanti-mycotic solution (Sigma). HCT-15 cells were maintained in RPMI 1640(Sigma) supplemented with 0.22% sodium bicarbonate, 0.011% sodium pyru-vate, 0.45% glucose, 0.24% N-2-hydroxyethylpiperazine-N#-2-ethanesulfonicacid, 10% fetal bovine serum and 10 ml/l of 100� antibiotic anti-mycoticsolution (Sigma). Cells were maintained at 37�C in the presence of 5% CO2.Reporter lysis buffer and luciferase reagent for luciferase studies were supplied
Abbreviations: BA, betulinic acid; C-DIM, methylene-substituted diindolyl-methanes; CDDO, 2-cyano-3,12-dioxo-18b-oleana-1,19-diene-28-oic acid; b-CDODA, 2-cyano-3,12-dioxo-18b-olean-1,12-diene-30-oic acid; CN-BA, 2-cyano-lup-1-en-3-oxo-20-oic acid; CN-BA-Me, methyl ester of CN-BA;DMEM, Dulbecco’s modified Eagle’s medium; DMSO, dimethyl sulfoxide;FBS, fetal bovine serum; b-GAL, b-Galactosidase; PPARc, peroxisome pro-liferator-activated receptor c; TBST, Tris-buffered saline containing Tween-20.
yBoth authors contributed equally to this work.
� The Author 2007. Published by Oxford University Press. All rights reserved. For Permissions, please email: [email protected] 2337
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from

by Promega (Madison, WI). b-Galactosidase (b-Gal) reagent was obtainedfrom Tropix (Bedford, MA), and LipofectAMINE reagent was purchased fromInvitrogen (Carlsbad, CA). The PPARc antagonist N-(4#-aminopyridyl)-2-chloro-5-nitrobenzamide (T007) (31) was synthesized in this laboratory, andits identity and purity (.98%) was confirmed by gas chromatography–massspectrometry.
Synthesis of CN-BA and CN-BA-Me
CN-BA and CN-BA-Me were prepared from betulin (Sigma–Aldrich) based onthe previous methods (19). The synthesis from a key intermediate, methyl lup-2-eno[2,3-d]isoxazol-28-oate, is briefly described and only definite peaks inproton NMR are recorded. Methyl Lup-2-eno[2,3-d]isoxazol-28-oate. To a so-lution of methyl lupan-2-hydroxymethylene-3-oxo-28-oate (350 mg, 0.70mmol) in ethanol (20 ml) and water (1 ml), hydroxylamine hydrochloride(488 mg, 7.02 mmol) was added. The reaction mixture was refluxed for 1 h,cooled to room temperature and concentrated under vacuum. Water was addedto the reaction mixture and extracted with ethyl acetate (2�). The organic layerwas then washed with brine (2�), separated, and the crude product was puri-fied by a flash silica gel column using a solvent system of hexanes:ethyl acetate(95:5) to yield methyl lup-2-eno[2,3-d]isoxazol-28-oate as a white cream-colored solid. 1H NMR (400 MHz, CDCl3): d 8.00 (1H, s), 3.68 (3H, s),1.31, 1.21, 0.99, 0.98, 0.83 (each 3H, s), 0.89 (3H, d, J 5 6.8 Hz), 0.78 (3H,d, J 5 6.8 Hz). 13C NMR (100 MHz, CDCl3): d 177.6, 173.8, 151.1, 109.7,57.8, 54.3, 52.0, 49.7, 49.6, 45.0, 43.4, 41.5, 39.7, 38.9, 38.1, 36.6, 35.6, 34.2,32.8, 30.6, 30.5, 30.4, 29.5, 27.7, 23.8, 23.6, 22.2, 22.0, 19.6, 16.8, 16.5, 15.5,15.4. Methyl 2-Cyano-lup-3-hydroxy-2-en-28-oate. To a solution of lup-2-eno[2,3-d]isoxazol-28-oate (250 mg, 0.50 mmol) in ether (30 ml) and metha-nol (15 ml) in an ice bath, 30% sodium methoxide in methanol (3071 mg, 56.87mmol) was added drop wise. The reaction mixture was then stirred at roomtemperature for 2 h. After dilution with ether, the reaction mixture was washedwith 5% hydrochloric acid (2�). The organic layer was separated and workedup by standard methods to yield crude methyl 2-cyano-lup-3-hydroxy-2-en-28-oate (240 mg, 96%) as a white solid, which was used for the next step withoutfurther purification. 1H NMR (400 MHz, CDCl3): d 3.90 (1H, m), 3.67 (3H, s),1.27, 1.17, 0.98, 0.95, 0.83 (each 3H, s), 0.89 (3H, d, J5 6.8 Hz), 0.79 (3H, d,J 5 6.8 Hz). CN-BA-Me. A mixture of 2-cyano-lup-3-hydroxy-2-en-28-oate(230 mg, 0.46 mmol) and 2,3-dichloro-5,6-dicyano-1,4-benzoguinone (117mg, 0.52 mmol) in benzene (30 ml) was refluxed for 3 h. The reaction mixturewas cooled in ice and filtered to remove reduced 2,3-dichloro-5,6-dicyano-1,4-benzoguinone. The filtrate was then concentrated under vacuum. The crudeproduct was purified by a flash silica gel column using a solvent system ofbenzene:acetone (98:2) to yield CN-BA-Me (170 mg, 74%) as a pale brownsolid. 1H NMR (400 MHz, CDCl3): d 7.83 (1H, s), 3.66 (3H, s), 1.25, 1.18,1.12, 1.00, 0.95 (each 3H, s), 0.87 (3H, d, J5 6.7 Hz), 0.75 (3H, d, J5 6.6 Hz).13C NMR (100 MHz, CDCl3): d 199.0, 177.4, 171.6, 115.8, 114.6, 57.6, 53.3,52.0, 49.4, 45.7, 44.8, 44.4, 43.6, 42.6, 41.5, 38.8, 37.9, 34.2, 32.6, 30.4, 30.2,28.5, 27.3, 23.7, 23.5, 22.1, 22.0, 19.6, 19.1, 17.2, 15.4, 15.2. ESI-HRMS Calcdfor (C32H47NO3 þ H): 494.3634. Found: 494.3685. Anal. (C32H47NO3) C, H.CN-BA. A mixture of CN-BA-Me (120 mg, 0.25 mmol) and lithiumiodide
(720 mg) in dimethylformamide (2.4 ml) was refluxed for 2 h. The reactionmixture was cooled to room temperature and 5% hydrochloric acid was added.The reaction mixture was extracted with ethyl acetate (2�). The organic layerwas then washed with water (2�) followed by washings with brine (2�). Theorganic layer was separated and worked up by standard methods. The crudeproduct was purified by a flash silica gel column using a solvent system ofhexanes:ethyl acetate (80:20) to yield CN-BA (93 mg, 80%) as a light yellowsolid. 1H NMR (400 MHz, CDCl3): d 10.29 (1H, broad s), 7.83 (1H, s), 1.27,1.14, 1.04, 0.99, 0.99 (each 3H, s), 0.91 (3H, d, J 5 6.8 Hz), 0.79 (3H, d, J 56.7 Hz). 13C NMR (100 MHz, CDCl3): d 199.0, 182.1, 171.5, 115.8, 114.8,57.5, 53.3, 49.3, 45.8, 44.8, 44.4, 43.7, 42.7, 41.5, 39.0, 38.1, 34.3, 32.7, 32.7,30.5, 30.3, 28.6, 27.4, 23.8, 23.5, 22.2, 22.1, 19.6, 19.2, 17.3, 15.4, 15.3. ESI-HRMS Calcd for (C31H45NO3 þ H): 480.3478. Found: 480.3540. Anal.(C31H45NO3) C, H. CN-BA and CN-BA-Me were .97% pure by spectroscopicanalysis.
Cell proliferation assay
This assay is performed in 12-well tissue culture plates at the concentration of2 � 104 cells per well, using Dulbecco’s modified Eagle’s medium (DMEM)/Ham’s F-12 media containing 2.5% charcoal-stripped fetal bovine serum(FBS). The cells were counted on the initial day using Z1 cell counter (Beck-man Coulter, Fullerton, CA) and then the cells were treated either with vehicle[dimethyl sulfoxide (DMSO)] or the indicated triterpenoid compounds, eachsample in triplicate. Every 48 h, fresh medium was added along with theindicated compounds. The count of the cells was taken after 2, 4 and 6 days.The results are expressed as means ± standard errors for each set of triplicate.
Mammalian two-hybrid assay
The GAL4 reporter construct contains 5� GAL4 response elements (p GAL4),were kindly provided by Dr Marty Mayo (University of North California,Chapel Hill, NC). The GAL4-coactivator fusion plasmids pM-SRC1,pMSRC2, pMSRC3, pM-DRIP205, pM-CARM-1 and PPARc-VP16 (Vp-PPARc) containing the DEF region of the PPARc (amino acids 183–505) fusedto the pVP16 expression vector were kindly provided by Dr Shigeaki Kato(University of Tokyo, Tokyo, Japan). SW480 colon cancer cells were plated in12-well tissue culture plates at 1 � 105 cells per well in DMEM/Ham’s F-12medium supplemented with 2.5% charcoal-stripped FBS. After allowing cellsto adhere overnight, transient transfections were carried out with GAL4-Luc(0.4 lg), b-GAL (0.04 lg), VP-PPARc (0.04 lg), pM-SRC1 (0.04 lg), pM-PGC-1 (0.04 lg), pM-SMRT (0.04 lg), pM-TRAP220 (0.04 lg), pM-DRIP205(0.04 lg) and pMCARM1 (0.04 lg) using LipofectAMINE2000 (Invitrogen)following the manufacturer’s guidelines. After 6 h of transfection, cells weretreated in triplicate either with vehicle (DMSO) or the indicated compoundsuspended in complete medium for 20–24 h. One hundred microliters per wellof 1� Reporter Lysis Buffer (Promega) was used to lyse the cells and 30 ll ofthis lysate was used to perform the luciferase and b-GAL assays using Lumi-count (Perkin-Elmer Life and Analytical Sciences, Boston, MA). The lucifer-ase activities obtained were normalized to the b-gal activity.
Transfections
Cells were seeded on to the 12-well plates and 0.4 lg of GAL4-Luc, 0.04 lg ofb-GAL, 0.04 lg of GAL4DBD-PPARc, 0.4 lg of p21-luc(FL) containing�2325 to þ8 insert, 0.4 lg of p21-luc (�124) containing �124 to þ8 insertand 0.4 lg of p21-LUC (�60) containing �60 to þ8 insert were transfectedusing LipofectAMINE reagent (Invitrogen) following the manufacturer’s pro-tocol. Cells were treated either with vehicle or respective compounds sus-pended in complete medium after 6 h of transfection. Cell lysate is extractedafter 20–22 h by adding 100 ll of 1� reporter lysis buffer per well and 30 llof this extract is used to quantitate the luciferase activity using Lumicount(Perkin-Elmer Life and Analytical Sciences). Each experiment is done intriplicate and the results are normalized to the b-GAL activity.
Western blot analysis
SW480, HT-29, HCT-15 and Panc-28 (3 � 105) colon cancer cells were seededin 6-well tissue culture plates in DMEM/Ham’s F-12 medium containing 2.5%charcoal-stripped FBS. Protein is extracted from the cells treated either withvehicle or indicated compounds suspended for 24 h except for caveolin-1protein, which was done for 72 h. Samples were extracted in high salt buffer[50 mmol/l N-2-hydroxyethylpiperazine-N#-2-ethanesulfonic acid, 500 mmol/lNaCl, 1.5 mmol/l MgCl2, 1 mmol/l ethyleneglycol-bis(aminoethylether)-tetraacetic acid, 10% glycerol and 1% Triton X-100 (pH 7.5) and 5 ll/mlprotease inhibitor cocktail (Sigma–Aldrich)]. Samples were incubated at100�C for 2 min, separated on either 10 or 12% Sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels and then transferred to polyvinylidenedifluoride membrane (Bio-Rad, Hercules, CA). The polyvinylidene difluoridemembrane was blocked in 5% TBST-Blotto (10 mM Tris–HCl, 150 mM NaCl,pH 8.0, 0.05% Triton X-100 and 5% non-fat dry milk) for about 30 min and
Fig. 1. Structures of the triterpenoid acid BA, CN-BA/CN-BA-Me and b-CDODA-Me. The b-CDODA compounds were derived from glycyrrhetinicacid using a synthetic procedure analogous as outlined in this paper forpreparation of CN-BA and CN-BA-Me from BA.
S.Chintharlapalli et al.
2338
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from

was then incubated in fresh 5% TBST-Blotto with 1:1000 for caveolin-1 (SantaCruz Biotechnology, Santa Cruz, CA), 1:1000 for p21(BD Pharmingen,Franklin Lakes, NJ) and 1:10 000 for b-actin (Sigma) primary antibody over-night with gentle shaking at 4�C. After washing with Tris-buffered salinecontaining Tween-20 (TBST) for 10 min, the membrane was incubated withrespective secondary antibody (1:5000) (Santa Cruz Biotechnology) in 5%TBST-Blotto for 3 h. The membrane is then washed with TBST for 10 min,incubated with chemiluminiscence reagent from Perkin-Elmer for 1 min andthen exposed to Kodak X-OMAT AR autoradiography film (Eastman Kodak,Rochester, NY).
Differentiation and Oil red O staining
3T3-L1 pre-adipocytes were cultured on poly-lysine-coated coverslips withDMEM and 10% FBS at 5% CO2 in 24-well plates. At 2 days after confluence,cells were incubated with fresh media supplemented with 3-isobutyl-1-methylxanthine (0.5 mM), dexamethasone (1 lM), insulin (1.7 lM), and in-dicated compounds (0.25 lM). After 48 h, cells were changed to fresh mediaand treated with DMSO or indicated compounds for 5 days. Cells without anytreatment for the entire 7 days were used as control. The cells were then fixedwith 10% formalin. Fixed cells were washed with 60% isopropanol and stainedwith filtered 60% Oil red O in deionized water. After staining, cells werewashed with water and photographed.
Semi-quantitative real-time polymerase chain reaction
SW480 and HT29 colon cancer cells were treated either with vehicle (DMSO)or indicated compounds and after 24 h total RNA was extracted using RNeasyMini kit (Qiagen, Valencia, CA). RNA concentration was measured by UV260:280 nm absorption ratio, and 2 lg RNA was used to synthesize cDNAusing Reverse Transcription System (Promega). Polymerase chain reaction con-ditions were as follows: initial denaturation at 94�C (1 min) followed by 28cycles of denaturation for 30 s at 94�C, annealing for 60 s at 55�C and extensionat 72�C for 60 s and a final extension step at 72�C for 5 min. The mRNA levelswere normalized using GAPDH as an internal housekeeping gene. Primersobtained from IDT (Coralville, IA) and used for amplification are as follows:KLF4 (sense 5#-CTA TGG CAG GGA GTC CGC TCC-3#; anti-sense 5#-ATGACC GAC GGG CTG CCG TAC-3#) and GAPDH (sense 5#-ACG GAT TTGGTC GTATTG GGC G-3#; anti-sense 5#-CTC CTG GAA GAT GGT GAT GG-3#). Polymerase chain reaction products were electrophoresed on 1% agarosegels containing ethidium bromide and visualized under UV transillumination.
Results
Figure 2 illustrates the effects of BA, CN-BA and CN-BA-Me ongrowth of SW480 and Panc-28 cells. All three compounds inhibitgrowth of both cell lines and IC50 values ranging from 1 to 5, 1 to2.5 and 1 to 2.5 lM (Panc-28) and 1 to 5, 1.0 and 1 to 2.5 lM (SW480)were observed for BA, CN-BA and CN-BA-Me, respectively. CN-BAwas the most cytotoxic compound in both cell lines and this confirmsresults of a previous report showing that 2-cyano derivatives of BAenhanced cytotoxicity (19). One of the hallmarks of PPARc agonistsis their induction of differentiation in 3T3-L1 adipocytes, which ischaracterized by accumulation of fat droplets, which can be detectedby Oil red O staining. Results in Figure 1D show that both CN-BA andCN-BA-Me induce Oil red O staining in this assay, whereas BA doesnot induce this response (data not shown). These results suggest thatthese 2-cyano derivatives of BA exhibit activity associated withPPARc agonists.
The PPARc agonist activity of BA and related compounds wasdetermined in SW480 cells transfected with PPARc-GAL4/pGAL4and a PPRE3-luc construct (Figure 3A). The results show that 2.5–10lM CN-BA and CN-BA-Me induced transactivation, whereas BA wasinactive in this assay. The PPARc agonist activities were also deter-mined in SW480 cells using the same constructs but treated withCN-BA and CN-BA-Me alone or in combination with the PPARcantagonist T007, and in all cases, the induced activities were inhibitedby T007 (Figure 3B). A similar approach was used in Panc-28 cellstransfected with PPARc-GAL4/pGAL4 and PPRE3-luc (Figure 3C),and CN-BA induced luciferase activity that was inhibited in cells co-treated with CN-BA plus T007. Not surprisingly, BA was inactive inthese assays; however, results obtained for CN-BA-Me were highlyinconsistent in Panc-28 cells compared with the colon cancer cell line(Figure 3A and B). CN-BA-Me exhibited minimal induction in cells
transfected with PPARc-GAL4/pGAL4 and no induction was ob-served in Panc-28 cells transfected with PPRE3-luc (data not shown).These results were observed in replicate experiments suggesting thatthere were structure-dependent differences (CN-BA versus CN-BA-Me)for activation of the PPARc-GAL4/pGAL4 or a PPRE3-luc constructsin Panc-28 (but not SW480) cells.
PPARc agonists are structurally diverse and induce tissue-specificreceptor-dependent responses that are typical of selective PPARcmodulators (29,30). Similar results have been observed for agonists thatbind and activate other nuclear receptors and this structure-dependenteffect is due, in part, to tissue-specific expression of coactivators andother nuclear proteins that exhibit ligand structure-dependent inter-actions with receptors. Results in Figure 3D summarize the effects ofCN-BA and CN-BA-Me on induction of luciferase activity in SW480cells transfected VP-PPARc and GAL4 coactivator and GAL4-SMRT(a co-repressor) expression plasmids. We used b-CDODA-Me, atriterpenoid methyl ester derivative, which also contains a 2-cyano-1-en-3-one function (Figure 1) and activates PPARc in colon cancercells (28), as a comparative reference compound for the mammaliantwo-hybrid assay. The results show that CN-BA, CN-BA-Me andb-CDODA-Me significantly induced luciferase activity in SW480cells transfected with VP-PPARc and GAL4-PGC-1 and GAL4-SRC-1, but not GAL4-AIB1, GAL4-TIFII, GAL4-TRAP220 andGAL4-SMRT. In contrast, only CN-BA-Me also activated GAL4-CARM1 indicating differences between CN-BA and CN-BA-Me inthe mammalian two-hybrid assay, suggesting that even among thesetwo acid–ester analogs, some tissue-specific selective PPARc modu-lator activity might be expected. The data are consistent with thedifferences observed for CN-BA and CN-BA-Me in activation oftransfected constructs in Panc-28 cells (Figure 3C).
Previous studies in this laboratory have shown that PPARc agonistsinduce p21 and p27 and decrease cyclin D1 expression in Panc-28cells, and only the former response is receptor dependent (32). Resultsin Figure 4C show that both CN-BA and CN-BA-Me induce p21protein expression in Panc-28 cells, and this is also accompanied byinduction of p27 and down-regulation of cyclin D1 (data not shown)as reported previously for a series of PPARc-active methylene-substituted diindolylmethanes (C-DIM) analogs in this cell line(32). Co-treatment of Panc-28 cells with 5 lM CN-BA and CN-BA-Me plus the 10 lM T007 significantly inhibited induction of p21,confirming that induction of p21 was PPARc dependent (Figure4A). In contrast, induction of p21 by BA was not inhibited after co-treatment with T007 and this was consistent with results of trans-activation studies showing that BA does not activate PPARc inPanc-28 or SW480 (Figure 3). Figure 4B shows that BA, CN-BAand CN-BA-Me induce transactivation in Panc-28 cells transfectedwith p21-luc(Fl), which contains the �2325 to þ8 region of the p21promoter. In cells co-treated with BA and related compounds plus thePPARc antagonist T007, the induction of luciferase activity by CN-BA and CN-BA-Me was inhibited, whereas BA-induced activity wasunaffected by T007. The results complement the immunoblot dataconfirming that induction of p21 by CN-BA/CN-BA-Me was PPARcdependent, whereas induction of p21 by BA was PPARc independent.We further investigated induction of luciferase activity in Panc-28cells transfected with constructs containing �2325 to þ8 [p21-Luc(Fl)], �124 to þ8 [p21-Luc (�124)], �101 to þ8 [p21-Luc (�101)]and �60 to þ8 [p21-Luc (�60)] p21 promoter inserts. The latter threeconstructs contain the six proximal GC-rich sites (1–6) and the resultsof the transfection studies suggest that these GC-rich sites are neces-sary for CN-BA- and CN-BA-Me-induced transactivation. Deletionanalysis of the p21 promoter indicated that loss of inducibility [i.e.p21-luc(60)] was associated with loss of GC-rich sites 3 and 4,whereas CN-BA significantly induced activity but only at the 7.5lM concentration, suggesting sites 3 and 4 were also important forthis compound but induction could also be observed using constructscontaining only GC-rich sites 5 and 6. Previous studies show thatPPARc-dependent activation of p21 by other PPARc agonists is alsodependent on GC-rich sites 3 and 4 and involves PPARc/Sp-dependentactivation of p21. The ligand-dependent recruitment of PPARc to
2-Cyano betulinic acid analogs activate PPARg
2339
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from

the p21 promoter by CN-BA and CN-BA-Me was further investigatedin a ChIP assay in Panc-28 cells treated with the BA, CN-BA andCN-BA-Me for 1 or 2 h. The results (Figure 4D) show that bothCN-BA and CN-BA-Me recruited PPARc to the proximal GC-richregion of the p21 promoter and this was also accompanied by en-hanced binding of Sp1. In contrast, BA did not induce PPARc inter-actions with the p21 promoter in the ChIP assay. This is consistentwith receptor-independent activation of p21 by BA and the mecha-
nism of this response is currently being investigated. As a control forthis experiment, transcription factor TFIIB bound to the proximalregion of the GAPDH gene promoter but not to exon 1 of the CNAP1gene.
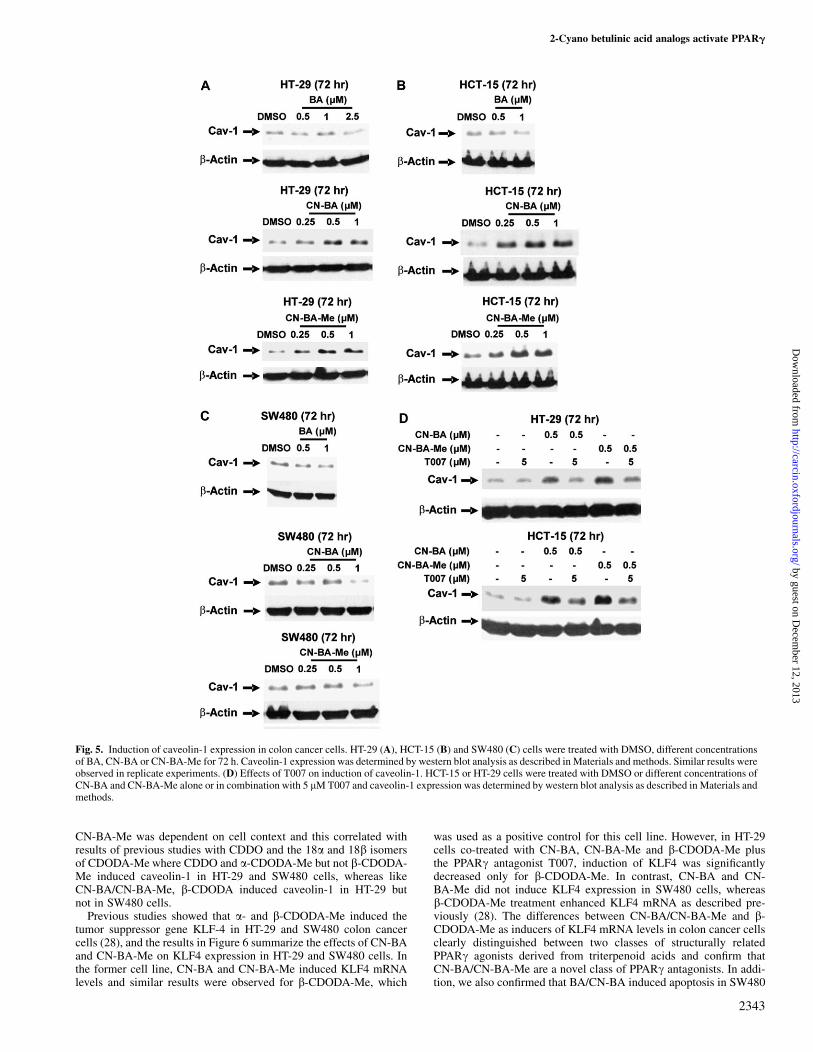
PPARc agonists such as CDDO, b-CDODA and related esters andPPARc-active C-DIMs also induce receptor-dependent expressionof caveolin-1 in colon cancer cells (28,33–35). Figure 5A showsthat CN-BA and CN-BA-Me but not BA induce caveolin-1 in HT-29
Fig. 2. Cell proliferation and adipocyte differentiation assays. Panc-28 and SW480 cells were treated with different concentrations of BA (A), CN-BA (B) or CN-BA-Me (C) for 6 days and the number of cells were counted after treatment for 2, 4 or 6 days as described in Materials and methods. Results are expressed asmeans ± standard errors for three separate determinations for each treatment group. (D) Effects of CN-BA and CN-BA-Me on differentiation of 3T3-L1adipocytes. 3T3-L1 adipocytes were treated with 0.25 lM CN-BA, CN-BA-Me or DMSO. Induction of fat droplets by Oil red O staining was determined asdescribed in Materials and methods. Induction of intense staining for fat droplets was observed in replicate (3) experiments.
S.Chintharlapalli et al.
2340
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from

cells and similar results were observed in HCT-15 cells (Figure 5B).In contrast, BA, CN-BA and CN-BA-Me did not induce caveolin-1expression in SW480 cells, and the latter two compounds decreasedexpression of this protein (Figure 5C). Co-treatment of HT-29 and
HCT-15 cells with CN-BA/CN-BA-Me plus the PPARc antago-nist T007 resulted in inhibition of the induced caveolin-1 response,confirming that induction was PPARc dependent (Figure 5D).Thus, receptor-dependent activation of caveolin-1 by CN-BA and
Fig. 3. Activation of PPARc in SW480 and Panc-28 cells by BA, CN-BA and CN-BA-Me. SW480 cells (A and B) were transfected with PPARc-GAL4/pGAL4 orPPRE3-luc treated with DMSO (control) or different concentrations of the compounds, and luciferase activity determined as described in Materials and methods.(C) Activation of PPARc in Panc-28 cells. Cells were transfected with PPARc-GAL4/pGAL4 or PPRE3-luc treated with DMSO or different concentrations of CN-BA and CN-BA-Me alone or in combination with 10 lM T007, and luciferase activity determined as described in Materials and methods. Results in (A–C) areexpressed as means ± standard errors for three replicate determinations for each treatment group, and significant (P, 0.05) induction by the BA derivatives (�) andinhibition after co-treatment with T007 (��) are indicated. For studies in Panc-28 cells, we only observed induction of luciferase activity using CN-BA and not withCN-BA-Me or BA over several sets of experiments. (D) Mammalian two-hybrid assay in SW480 cells transfected with VP-PPARc and GAL4-coactivatorchimeras. SW480 cells were transfected with VP-PPARc, coactivator-GAL4/pGAL4, treated with different concentrations of CN-BA or CN-BA-Me and 5 lMb-CDODA-Me, and luciferase activity was determined as described in Materials and methods. Results are expressed as means ± standard errors for three replicatedeterminations for each treatment group, and significant (P , 0.05) induction is indicated by an asterisk.
2-Cyano betulinic acid analogs activate PPARg
2341
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from
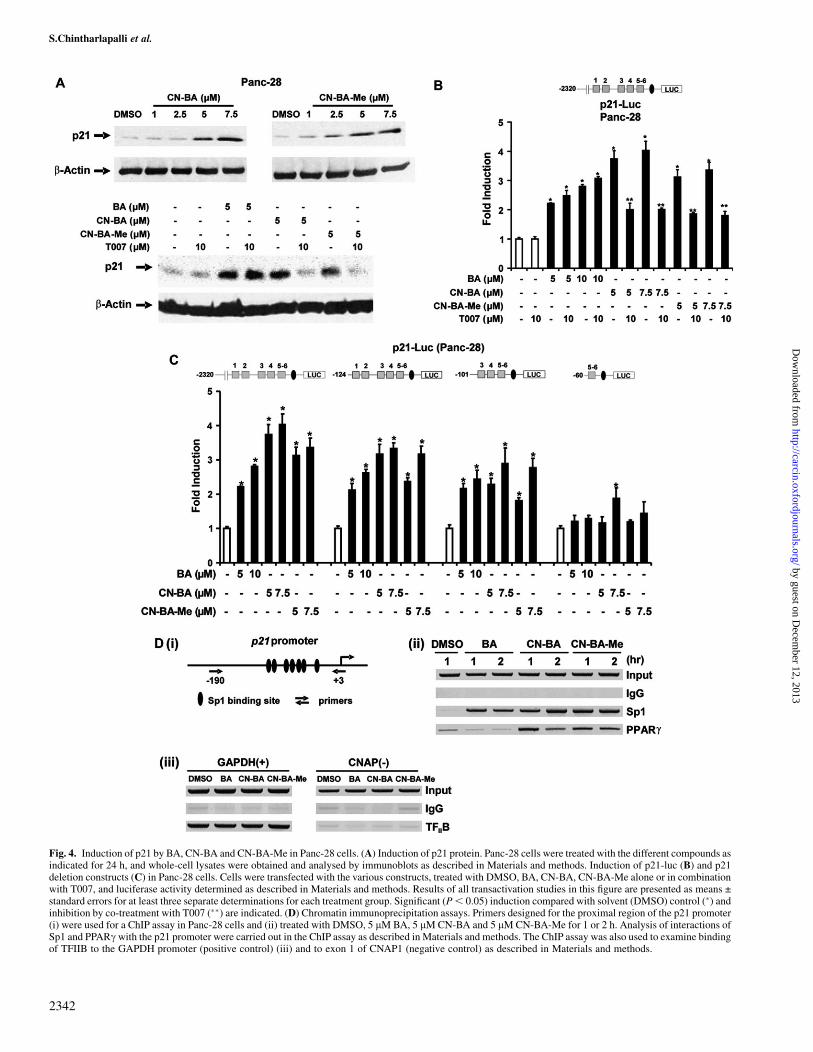
Fig. 4. Induction of p21 by BA, CN-BA and CN-BA-Me in Panc-28 cells. (A) Induction of p21 protein. Panc-28 cells were treated with the different compounds asindicated for 24 h, and whole-cell lysates were obtained and analysed by immunoblots as described in Materials and methods. Induction of p21-luc (B) and p21deletion constructs (C) in Panc-28 cells. Cells were transfected with the various constructs, treated with DMSO, BA, CN-BA, CN-BA-Me alone or in combinationwith T007, and luciferase activity determined as described in Materials and methods. Results of all transactivation studies in this figure are presented as means ±standard errors for at least three separate determinations for each treatment group. Significant (P, 0.05) induction compared with solvent (DMSO) control (�) andinhibition by co-treatment with T007 (��) are indicated. (D) Chromatin immunoprecipitation assays. Primers designed for the proximal region of the p21 promoter(i) were used for a ChIP assay in Panc-28 cells and (ii) treated with DMSO, 5 lM BA, 5 lM CN-BA and 5 lM CN-BA-Me for 1 or 2 h. Analysis of interactions ofSp1 and PPARc with the p21 promoter were carried out in the ChIP assay as described in Materials and methods. The ChIP assay was also used to examine bindingof TFIIB to the GAPDH promoter (positive control) (iii) and to exon 1 of CNAP1 (negative control) as described in Materials and methods.
S.Chintharlapalli et al.
2342
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from
CN-BA-Me was dependent on cell context and this correlated withresults of previous studies with CDDO and the 18a and 18b isomersof CDODA-Me where CDDO and a-CDODA-Me but not b-CDODA-Me induced caveolin-1 in HT-29 and SW480 cells, whereas likeCN-BA/CN-BA-Me, b-CDODA induced caveolin-1 in HT-29 butnot in SW480 cells.
Previous studies showed that a- and b-CDODA-Me induced thetumor suppressor gene KLF-4 in HT-29 and SW480 colon cancercells (28), and the results in Figure 6 summarize the effects of CN-BAand CN-BA-Me on KLF4 expression in HT-29 and SW480 cells. Inthe former cell line, CN-BA and CN-BA-Me induced KLF4 mRNAlevels and similar results were observed for b-CDODA-Me, which
was used as a positive control for this cell line. However, in HT-29cells co-treated with CN-BA, CN-BA-Me and b-CDODA-Me plusthe PPARc antagonist T007, induction of KLF4 was significantlydecreased only for b-CDODA-Me. In contrast, CN-BA and CN-BA-Me did not induce KLF4 expression in SW480 cells, whereasb-CDODA-Me treatment enhanced KLF4 mRNA as described pre-viously (28). The differences between CN-BA/CN-BA-Me and b-CDODA-Me as inducers of KLF4 mRNA levels in colon cancer cellsclearly distinguished between two classes of structurally relatedPPARc agonists derived from triterpenoid acids and confirm thatCN-BA/CN-BA-Me are a novel class of PPARc antagonists. In addi-tion, we also confirmed that BA/CN-BA induced apoptosis in SW480
Fig. 5. Induction of caveolin-1 expression in colon cancer cells. HT-29 (A), HCT-15 (B) and SW480 (C) cells were treated with DMSO, different concentrationsof BA, CN-BA or CN-BA-Me for 72 h. Caveolin-1 expression was determined by western blot analysis as described in Materials and methods. Similar results wereobserved in replicate experiments. (D) Effects of T007 on induction of caveolin-1. HCT-15 or HT-29 cells were treated with DMSO or different concentrations ofCN-BA and CN-BA-Me alone or in combination with 5 lM T007 and caveolin-1 expression was determined by western blot analysis as described in Materials andmethods.
2-Cyano betulinic acid analogs activate PPARg
2343
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from
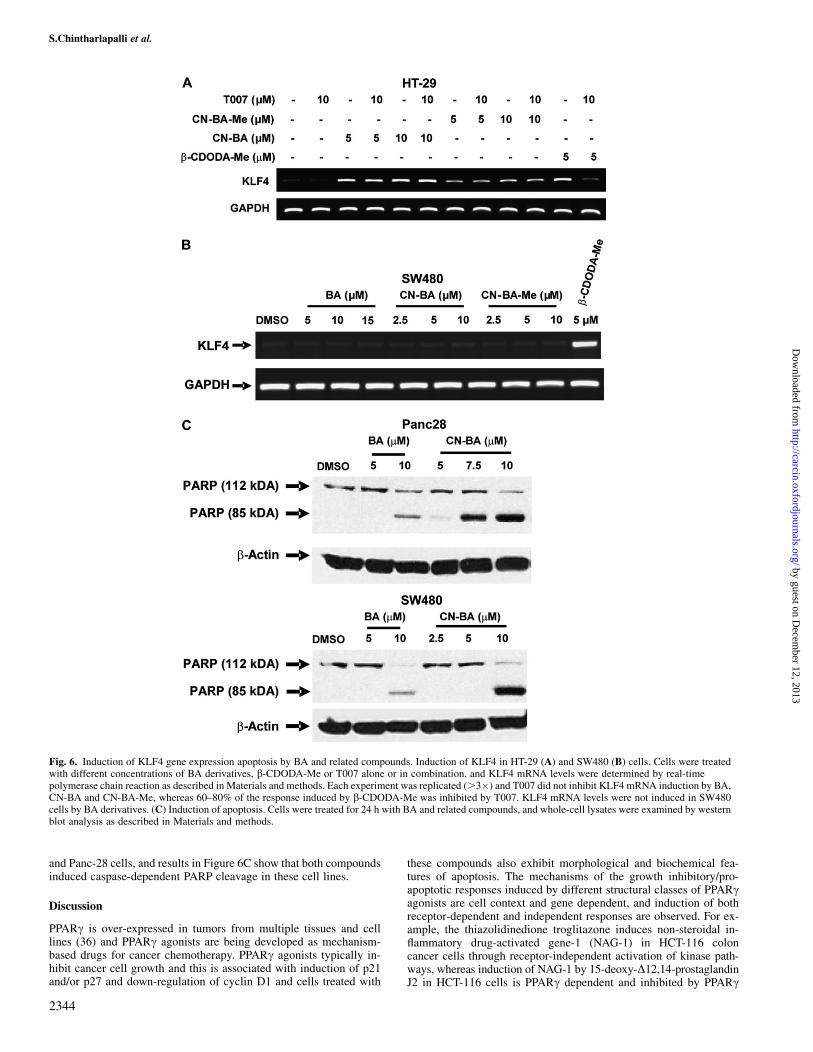
and Panc-28 cells, and results in Figure 6C show that both compoundsinduced caspase-dependent PARP cleavage in these cell lines.
Discussion
PPARc is over-expressed in tumors from multiple tissues and celllines (36) and PPARc agonists are being developed as mechanism-based drugs for cancer chemotherapy. PPARc agonists typically in-hibit cancer cell growth and this is associated with induction of p21and/or p27 and down-regulation of cyclin D1 and cells treated with
these compounds also exhibit morphological and biochemical fea-tures of apoptosis. The mechanisms of the growth inhibitory/pro-apoptotic responses induced by different structural classes of PPARcagonists are cell context and gene dependent, and induction of bothreceptor-dependent and independent responses are observed. For ex-ample, the thiazolidinedione troglitazone induces non-steroidal in-flammatory drug-activated gene-1 (NAG-1) in HCT-116 coloncancer cells through receptor-independent activation of kinase path-ways, whereas induction of NAG-1 by 15-deoxy-D12,14-prostaglandinJ2 in HCT-116 cells is PPARc dependent and inhibited by PPARc
Fig. 6. Induction of KLF4 gene expression apoptosis by BA and related compounds. Induction of KLF4 in HT-29 (A) and SW480 (B) cells. Cells were treatedwith different concentrations of BA derivatives, b-CDODA-Me or T007 alone or in combination, and KLF4 mRNA levels were determined by real-timepolymerase chain reaction as described in Materials and methods. Each experiment was replicated (.3�) and T007 did not inhibit KLF4 mRNA induction by BA,CN-BA and CN-BA-Me, whereas 60–80% of the response induced by b-CDODA-Me was inhibited by T007. KLF4 mRNA levels were not induced in SW480cells by BA derivatives. (C) Induction of apoptosis. Cells were treated for 24 h with BA and related compounds, and whole-cell lysates were examined by westernblot analysis as described in Materials and methods.
S.Chintharlapalli et al.
2344
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from

antagonist (37–39). PPARc-active C-DIMs induce caveolin-1 expres-sion in HT-29 and HCT-15 colon cancer cells, whereas rosiglitazoneinduced caveolin-1 only in the former cell lines (33). The inductionresponses by both compounds were inhibited by PPARc antagonistsand cell context-dependent differences of C-DIMs and rosiglitazonein HCT-15 cells were associated with expression of mutant PPARc(K422Q) in this cell line (34) and the mutant receptor was insensitiveto rosiglitazone.
CDDO and its related methyl ester and imidazole derivatives arePPARc agonists (27,35) and are potent anticancer drugs currentlyundergoing clinical trials. These triterpenoid acid derivatives of ole-anolic acid, a phytochemical used in traditional medicine induce mul-tiple receptor-independent and some receptor-dependent responsesincluding the receptor-dependent induction of caveolin-1 in coloncancer cells (35). Although oleanolic acid is only weakly cytotoxicto cancer cells, the introduction of the 2-cyano-substituted 1-en-3-oxomoiety into the A-ring of oleanolic acid greatly enhanced the cyto-toxicity of the resulting 2-cyano derivatives including CDDO that alsohas an enone system in the C-ring (24–26) and similar results wereobserved for the corresponding 2-cyano derivatives of glycyrrhetinicacid, namely a-CDODA and b-CDODA-Me (28). Moreover, in stud-ies with glycyrrhetinic acid analogs, it was shown that the 2-cyanogroup was required for PPARc agonist activity.
The major structural differences between BA and oleanolic andglycyrrhetinic acids are their 5- and 6-member E-rings, respectively(Figure 1), and the position of substituents in this ring. However,despite these structural differences, introduction of the 2-cyano-1-ene-3-oxo system into the A-ring of BA gave CN-BA, which activatedPPARc in both SW480 and Panc-28 cell lines (Figures 2 and 3) andenhanced the cytotoxicity of these compounds compared with BA(Figure 1). Surprisingly, CN-BA-Me activated PPARc-GAL4/pGAL4and PPRE-luc in SW480 but was much less effective in activatingthese constructs in Panc-28 cells, and these cell context-dependentdifferences in CN-BA and CN-BA-Me suggest that these compoundsmay be selective PPARc agonists or modulators.
Selective receptor modulators exhibit tissue-selective receptor ag-onist activities and differences between diverse structural classes ofthese compounds can be discerned in mammalian two-hybrid assaysusing VP-PPAR and GAL4-coactivator chimeras (28,33,35). This as-say has some relevance for identifying selective receptor modulatorssince differences between selective receptor modulators may be due,in part, to their interaction with coactivator proteins. Induction ofluciferase activity in colon cancer cells transfected with VP-PPARc/GAL4-coactivator constructs is dependent on the coactivator andstructure of the PPARc agonist (28,33,35). C-DIM PPARc agonistsinduce transactivation in cells transfected with GAL4-PGC-1 (33),whereas CDDO and CDDO-Me are active in cells transfected withGAL4-chimeras containing SRC-1, SRC-2 (TIFII), SRC-3 (A1B1),TRAP220, PGC-1 and CARM-1 (35). b-CDODA and a-CDODA-Me activate GAL4-chimeras containing PGC-1 and SRC-1 andPGC-1 and SRC-2, respectively (28). CN-BA activates GAL4-chimeras containing PGC-1 and SRC-1 in SW480 cells and resemblesb-CDODA-Me. CN-BA-Me activates PGC-1, SRC-1 and CARM-2(Figure 3D), whereas these compounds did not activate GAL4-chimeras containing SRC-2, TRAP220 or SMRT (data not shown).The unique pattern for CN-BA-Me in the mammalian two-hybridassay highlights differences that are dependent only on methylationof the 20-carboxyl group in the E-ring, and these results are con-sistent with differences between CN-BA and CN-BA-Me in theiractivation of transfected PPARc-responsive constructs in Panc-28cells (Figure 3C).
The PPARc agonist activities of CN-BA and CN-BA-Me and theirrole as selective receptor modulators were further investigated usingfour receptor-mediated responses, namely (i) the induced differentia-tion of 3T3-L1 adipocytes, (ii) induction of the cyclin-dependentkinase inhibitor p21 in Panc-28 cells and the induction of (iii) cav-eolin-1 and (iv) KLF4 in colon cancer cells. Both CN-BA and CN-BA-Me induced differentiation of 3T3-L1 adipocytes and this wascharacterized by accumulation of fat droplets, which are visualized
by Oil red O staining (Figure 2D). Previous studies showed thatPPARc-active C-DIMs induced p21 expression in Panc-28 cells andthis response was associated with interactions of PPARc with theproximal GC-rich region of the p21 promoter (32). Similar resultswere obtained for both CN-BA and CN-BA-Me that induced p21expression in Panc-28 cells and reporter gene activity in cells trans-fected with p21-luc(F1), and both responses were inhibited after co-treatment with PPARc antagonist T007 (Figure 4A). Deletion analysisof the p21 promoter suggested that for CN-BA-Me, GC-rich sites 5and 6 were required for activation of p21, whereas CN-BA also in-duced activity with p21-luc(60) that only contained GC-rich sites 5and 6. Nevertheless, sites 3 and 4 appear to play an important role forboth CN-BA-Me and CN-BA, and these same sites were also requiredfor PPARc-dependent activation of p21 by C-DIMs (32). It has alsobeen reported previously that progesterone receptor and androgenreceptor agonists induce p21 through receptor-Sp protein interactionswith GC-rich sites 3 and 4 and site 3, respectively (40,41), suggestingthat these GC-rich sites in the p21 promoter are important targetsfor nuclear receptors. We also showed that BA induced PPARc-independent activation of p21, and the differences between BA versusCN-BA/CN-BA-Me are evident not only after treatment with thePPARc antagonist T007 (Figure 4A and B) but also in the recruitmentof PPARc to the p21 promoter by CN-BA/CN-BA-Me but not BA ina ChIP assay in Panc-28 cells (Figure 4D).
In a recent study with the 18a and 18b isomers of CDODA-Me, weshowed that both compounds induced caveolin-1 in HT-29 and HCT-15 cells, whereas only a-CDODA-Me induced caveolin-1 in SW480cells (28). These results suggested that in SW480 cells, the stereo-chemistry at C-18 of CDODA that influences the confirmation of theE-ring also differentially affected PPARc-dependent activation ofcaveolin-1. CN-BA and CN-BA-Me but not BA induced caveolin-1in HT-29 and HCT-15 cells and co-treatment with the PPARc antag-onist T007 inhibited the induction response (Figure 6). In contrast,CN-BA and CN-BA-Me do not induce caveolin-1 in SW480 cells asobserved previously for 18b isomer of CDODA; however, the stereo-chemistry at C-18 for the BA derivatives is a, suggesting that the cellcontext-dependent activation of caveolin-1 by the cyano-substitutedtriterpenoid acids is dependent not only on the stereochemistry atC-18 but also on the structure of the E-ring. The presence of the5-membered E-ring with carboxy and isopropyl substituents (Figure 1)resulted in loss of PPARc-dependent induction of caveolin-1 inSW480 cells by CN-BA and CN-BA-Me and this cell context-dependentresponse was consistent with the activity of these compounds asselective receptor modulators. The cytotoxicity of BA and CN-BAderivatives (Figure 2) was due not only to growth inhibition but alsoto induction of apoptosis (Figure 6C), which was not inhibited byPPARc antagonists (data not shown), suggesting a receptor-independentpro-apoptotic pathway which is currently being investigated.
The CDODA-Me compound induced the tumor suppressor geneKLF4 in SW480 and HT-29 colon cancer cells, and this response wasinhibited by T007 (28). In contrast, CN-BA/CN-BA-Me did not induceKLF4 mRNA in SW480 cells and induction of this gene in HT-29 cellswas receptor independent. These data, coupled with the effects of thecyano-substituted compounds on transactivation in the mammaliantwo-hybrid and reporter gene assays, adipocyte differentiation, p21and caveolin-1 expression, demonstrate that CN-BA and CN-BA-Merepresent a novel class of selective PPARc agonists in colon and pan-creatic cancer cells. The concentration-dependent differences in theactivation of p21 and PPARc-GAL4/PPRE3-luc (�2.5 lM) and induc-tion of Oil red O staining and caveolin-1 (�0.5 lM) may be due, in part,to relatively short (24 h) and longer (72–120 h) treatment times, re-spectively. However, differences in gene responsiveness may also bedue to other nuclear proteins and competition by receptor complexesbound to response elements on different gene promoters for commonnuclear cofactors. The activities of CN-BA and CN-BA-Me coupledwith their cytotoxicity (Figure 2) suggest that the receptor-dependentand independent responses induced by these compounds will be advan-tageous for further development of these compounds for clinical appli-cations in the treatment of colon and pancreatic cancer.
2-Cyano betulinic acid analogs activate PPARg
2345
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from

Funding
National Institutes of Health (ES09106, CA108718 and CA112337);the M.D.Anderson Cancer Center Pancreatic Cancer Spore (P20-CA10193) and the Texas Agricultural Experiment Station.
Acknowledgements
Conflict of interest statement: None declared.
References
1.Sami,A. et al. (2006) Pharmacological properties of the ubiquitous naturalproduct betulin. Eur. J. Pharmacol. Sci., 29, 1–13.
2.Yogeeswari,P. et al. (2005) Betulinic acid and its derivatives: a review ontheir biological properties. Curr. Med. Chem., 12, 657–666.
3.Pisha,E. et al. (1995) Discovery of betulinic acid as a selective inhibitor ofhuman melanoma that functions by induction of apoptosis. Nat. Med., 1,1046–1051.
4.Fulda,S. et al. (1998) Activation of mitochondria and release of mitochon-drial apoptogenic factors by betulinic acid. J. Biol. Chem., 273, 33942–33948.
5.Tan,Y. et al. (2003) Betulinic acid-induced programmed cell death in hu-man melanoma cells involves mitogen-activated protein kinase activation.Clin. Cancer Res., 9, 2866–2875.
6.Fulda,S. et al. (1997) Betulinic acid triggers CD95 (APO-1/Fas)- and p53-independent apoptosis via activation of caspases in neuroectodermal tu-mors. Cancer Res., 57, 4956–4964.
7.Galgon,T. et al. (2005) Betulinic acid induces apoptosis in skin cancer cellsand differentiation in normal human keratinocytes. Exp. Dermatol., 14,736–743.
8.Ehrhardt,H. et al. (2004) Betulinic acid-induced apoptosis in leukemiacells. Leukemia, 18, 1406–1412.
9.Noda,Y. et al. (1997) Enhanced cytotoxicity of some triterpenes towardleukemia L1210 cells cultured in low pH media: possibility of a new modeof cell killing. Chem. Pharm. Bull. (Tokyo), 45, 1665–1670.
10.Mukherjee,R. et al. (2004) Betulinic acid and its derivatives as anti-angio-genic agents. Bioorg. Med. Chem. Lett., 14, 2181–2184.
11. Jeremias,I. et al. (2004) Cell death induction by betulinic acid, ceramideand TRAIL in primary glioblastoma multiforme cells. Acta Neurochir.(Wien.), 146, 721–729.
12.Fulda,S. et al. (2005) Sensitization for anticancer drug-induced apoptosisby betulinic Acid. Neoplasia, 7, 162–170.
13.Fulda,S. et al. (1998) Molecular ordering of apoptosis induced by antican-cer drugs in neuroblastoma cells. Cancer Res., 58, 4453–4460.
14.Kasperczyk,H. et al. (2005) Betulinic acid as new activator of NF-kappaB:molecular mechanisms and implications for cancer therapy. Oncogene, 24,6945–6956.
15.Fulda,S. et al. (2004) Cooperation of betulinic acid and TRAIL to induceapoptosis in tumor cells. Oncogene, 23, 7611–7620.
16.Zuco,V. et al. (2002) Selective cytotoxicity of betulinic acid on tumor celllines, but not on normal cells. Cancer Lett., 175, 17–25.
17.Kim,J.Y. et al. (2001) Development of C-20 modified betulinic acid deriv-atives as antitumor agents. Bioorg. Med. Chem. Lett., 11, 2405–2408.
18.Sarek,J. et al. (2003) New lupane derived compounds with pro-apoptoticactivity in cancer cells: synthesis and structure-activity relationships.J. Med. Chem., 46, 5402–5415.
19.You,Y.J. et al. (2003) Synthesis and cytotoxic activity of A-ring modifiedbetulinic acid derivatives. Bioorg. Med. Chem. Lett., 13, 3137–3140.
20.Sawada,N. et al. (2004) Betulinic acid augments the inhibitory effects ofvincristine on growth and lung metastasis of B16F10 melanoma cells inmice. Br. J. Cancer, 90, 1672–1678.
21. Jeong,H.J. et al. (1999) Preparation of amino acid conjugates of betulinicacid with activity against human melanoma. Bioorg. Med. Chem. Lett., 9,1201–1204.
22.Liu,W.K. et al. (2004) Apoptotic activity of betulinic acid derivatives onmurine melanoma B16 cell line. Eur. J. Pharmacol., 498, 71–78.
23.Kim,D.S. et al. (1998) Synthesis of betulinic acid derivatives with activityagainst human melanoma. Bioorg. Med. Chem. Lett., 8, 1707–1712.
24.Honda,T. et al. (2000) Novel synthetic oleanane and ursane triterpenoidswith various enone functionalities in ring A as inhibitors of nitric oxideproduction in mouse macrophages. J. Med. Chem., 43, 1866–1877.
25.Honda,T. et al. (1998) Design and synthesis of 2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid, a novel and highly active inhibitor of nitric oxide productionin mouse macrophages. Bioorg. Med. Chem. Lett., 8, 2711–2714.
26.Honda,T. et al. (1997) New enone derivatives of oleanolic acid and ursolicacid as inhibitors of nitric oxide production in mouse macrophages. Bioorg.Med. Chem. Lett., 7, 1623–1628.
27.Wang,Y. et al. (2000) A synthetic triterpenoid, 2-cyano-3,12-dioxooleana-1,9-dien-28-oic acid (CDDO), is a ligand for the peroxisome proliferator-activated receptor g. Mol. Endocrinol., 14, 1550–1556.
28.Chintharlapalli,S. et al. (2007) Structure-dependent activity of glycyrrhe-tinic acid derivatives as peroxisome proliferator-activated receptor g(PPARg) agonists in colon cancer cells. Mol. Cancer Therap, 6, 1588–1598.
29.Escher,P. et al. (2000) Peroxisome proliferator-activated receptors: insightinto multiple cellular functions. Mutat. Res., 448, 121–138.
30.Fajas,L. et al. (2001) Peroxisome proliferator-activated receptor-g: fromadipogenesis to carcinogenesis. J. Mol. Endocrinol., 27, 1–9.
31.Lee,G. et al. (2002) T0070907, a selective ligand for peroxisome prolifer-ator-activated receptor g, functions as an antagonist of biochemical andcellular activities. J. Biol. Chem., 277, 19649–19657.
32.Hong,J. et al. (2004) Peroxisome proliferator-activated receptor g-dependent activation of p21 in Panc-28 pancreatic cancer cells involvesSp1 and Sp4 proteins. Endocrinology, 145, 5774–5785.
33.Chintharlapalli,S. et al. (2004) 1,1-Bis(3#-indolyl)-1-(p-substituted phe-nyl)methanes induce peroxisome proliferator-activated receptor g-mediated growth inhibition, transactivation and differentiation markers incolon cancer cells. Cancer Res., 64, 5994–6001.
34.Chintharlapalli,S. et al. (2006) 1,1-Bis(3#-indolyl)-1-(p-substituted phe-nyl)methanes inhibit colon cancer cell and tumor growth throughPPARg-dependent and PPARg-independent pathways. Mol. Cancer Ther.,5, 1362–1370.
35.Chintharlapalli,S. et al. (2005) 2-Cyano-3,12-dioxoolean-1,9-dien-28-oicacid and related compounds inhibit growth of colon cancer cells throughperoxisome proliferator-activated receptor g-dependent and -independentpathways. Mol. Pharmacol., 68, 119–128.
36. Ikezoe,T. et al. (2001) Mutational analysis of the peroxisome proliferator-activated receptor g gene in human malignancies. Cancer Res., 61, 5307–5310.
37.Baek,S.J. et al. (2003) Troglitazone, a peroxisome proliferator-activatedreceptor g (PPARg) ligand, selectively induces the early growth re-sponse-1 gene independently of PPARg. A novel mechanism for its anti-tumorigenic activity. J. Biol. Chem., 278, 5845–5853.
38.Baek,S.J. et al. (2004) Expression of NAG-1, a transforming growth factor-b superfamily member, by troglitazone requires the early growth responsegene EGR-1. J. Biol. Chem., 279, 6883–6892.
39.Gupta,R.A. et al. (2003) Peroxisome proliferator-activated receptorg-mediated differentiation: a mutation in colon cancer cells reveals diver-gent and cell type-specific mechanisms. J. Biol. Chem., 278, 22669–22677.
40.Owen,G.I. et al. (1998) Progesterone regulates transcription of the p21WAF1
cyclin-dependent kinase inhibitor gene through Sp1 and CBP/p300. J. Biol.Chem., 273, 10696–10701.
41.Lu,S. et al. (2000) Androgen induction of cyclin-dependent kinase inhibitorp21 gene: role of androgen receptor and transcription factor Sp1 complex.Mol. Endocrinol., 14, 753–760.
Received June 8, 2007; revised August 13, 2007; accepted August 13, 2007
S.Chintharlapalli et al.
2346
by guest on Decem
ber 12, 2013http://carcin.oxfordjournals.org/
Dow
nloaded from




















