
Differential inflammatory macrophage response to rutile and titanium particles
Upload
independentCategory
view
3download
0
Melissa H. Nagelin, Suseela Srinivasan, Jianyi Lee, Jerry L. Nadler and Catherine C. HedrickCassette Transporter G1
12/15-Lipoxygenase Activity Increases the Degradation of Macrophage ATP-Binding
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 2008 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/ATVBAHA.108.1679082008;28:1811-1819; originally published online July 17, 2008;Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/28/10/1811World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org/content/suppl/2008/07/17/ATVBAHA.108.167908.DC1.htmlData Supplement (unedited) at:
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. Question and AnswerPermissions and Rightspage under Services. Further information about this process is available in the
which permission is being requested is located, click Request Permissions in the middle column of the WebCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

12/15-Lipoxygenase Activity Increases the Degradation ofMacrophage ATP-Binding Cassette Transporter G1
Melissa H. Nagelin, Suseela Srinivasan, Jianyi Lee, Jerry L. Nadler, Catherine C. Hedrick
Objective—The purpose of this study was to evaluate the effect of 12/15-lipoxygenase (12/15LO) in macrophage ABCG1expression and function associated with cholesterol efflux.
Methods and Results—12/15LO was stably overexpressed in J774 macrophages. 12/15LO-overexpressing macrophageshad a 30% reduction in HDL-mediated cholesterol efflux, corresponding with significantly reduced ABCG1 proteinexpression. Treatment of 12/15LO-overexpressing macrophages with a 12/15LO ribozyme to reduce 12/15LO restoredHDL-mediated efflux and ABCG1 protein expression. Treating macrophages with 12/15LO unsaturated fatty acidsubstrates or eicosanoid products also reduced HDL-mediated cholesterol efflux. Additionally, both 12/15LOoverexpression in macrophages and incubation of macrophages with eicosanoids reduced ABCG1 protein, but notmRNA, expression. However, incubation of macrophages with linoleic or arachidonic acids significantly reduced bothABCG1 mRNA and protein expression, suggesting that 12/15LO substrates and eicosanoid products differentiallyregulate ABCG1 expression. 12/15LO fatty acids did not decrease ABCG1 translation; however, 12/15LO fatty acidsincreased ABCG1 degradation when blocked by cyclohexidmide. ABCG1 degradation may be regulated throughposttranslational modifications. Treatment with the 12/15LO eicosanoid product 12SHETE increased serine phosphor-ylation of ABCG1.
Conclusions—We conclude that serine phosphorylation may increase the degradation rate of ABCG1, and as a result causemacrophage cholesterol accumulation. These findings provide evidence that 12/15LO activity in the vessel wallcontributes to atherogenesis by impairing the macrophage ABCG1 cholesterol efflux pathway. (Arterioscler ThrombVasc Biol. 2008;28:1811-1819)
Key Words: lipoxygenase � ABCG1 � macrophage � fatty acid � eicosanoid
Atherosclerosis is a chronic inflammatory disease charac-terized by monocyte recruitment to the arterial wall and
their subsequent accumulation as lipid-laden macrophages, or“foam” cells. Oxidation of low-density lipoprotein (LDL)plays an essential role in the early events leading to foam cellformation, and the enzyme 12/15-lipoxygenase (12/15LO)has been implicated in LDL oxidation.1,2
12/15LO is a nonheme iron– containing dioxygenasewhich incorporates molecular oxygen stereospecifically toarachidonic and linoleic acid to produce the hydroperoxyfatty acids 12S- and 15S-hydroxyeicosatetranoic acids(12SHETE/15SHETE) and 13S-hydroxyoctadecadienoic acid(13SHODE), respectively.3,4 The 12/15-lipoxygenase familyconsists of “platelet-type” 12-lipoxygenase and “leukocyte-type” 12/15-lipoxygenase. Based on the ratios of 12S- and15SHETE produced from arachidonic acid, the enzyme isdesignated 15-lipoxygenase (15LO) in humans and leukocyte12-lipoxygenase in mice and pigs. Murine leukocyte 12/15LO and human 15LO proteins are highly homologous.5
12/15LO activation has been implicated in the pathogene-sis of atherosclerosis and diabetes. We and others have shown
that mice deficient in 12/15LO are protected from thedevelopment of atherosclerosis,6,7 whereas mice that overex-press the enzyme are more prone to lesion development.8 Themechanisms by which 12/15LO regulates atherogenesis re-main unknown, but may be attributable to inflammatoryactions of 12/15LO eicosanoids on the endothelium andmacrophages in the vessel wall or attributable to its ability tooxidize LDL.1,8,9 12/15LO protein has been localized withinatherosclerotic lesions in both rabbits and humans.10,11 Invitro data from Kuhn et al suggested a role for 12/15LO in theoxidation of LDL,12 whereas Cathcart and colleagues dem-onstrated that human monocytes generate superoxide thatmediates the oxidation of LDL in an enzyme-dependentmanner.2
Initially in lesion development monocytes are recruited tothe vessel wall, where they differentiate into macrophagesand upregulate scavenger receptors such as CD36 and SR-Ato facilitate the uptake of oxidized LDL.13 Conversely,differentiated macrophages also upregulate members of theATP binding cassette transporter family (ABC transporters),
Original received November 25, 2007; final version accepted July 8, 2008.From the Department of Pharmacology (M.H.N., C.C.H.), The Robert M. Berne Cardiovascular Research Center (M.H.N., S.S., J.L., C.C.H.), the
Division of Endocrinology and Metabolism (J.L.N.), and the Department of Internal Medicine (C.C.H.), University of Virginia, Charlottesville.Correspondence to Catherine C. Hedrick, Cardiovascular Research Center, University of Virginia, 415 Lane Rd, Bldg MR-5, Rm G123, P.O. Box
801394, Charlottesville, VA 22908. E-mail [email protected]© 2008 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.108.167908
1811 by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

namely ABCA1 and ABCG1, to mediate the transfer or effluxof modified cholesterol moieties to high density lipoproteins(HDL).14 HDL transports this cholesterol back to the liver forsubsequent conversion to bile and removal from the bodyduring the process of reverse cholesterol transport (RCT).ABCA1-mediated removal of cholesterol to lipid-poor HDLparticles has been extensively studied.15 Fatty acids canregulate ABCA1 expression and function. Oram and col-leagues have demonstrated that unsaturated fatty acids, suchas arachidonic and linoleic acid, inhibit cholesterol effluxfrom macrophages by enhancing the protein degradation andserine phosphorylation of ABCA1.16,17 Moreover, apoli-poprotein A-I (apoA-I) binds and cross-links ABCA1 tostabilize and protect the transporter from degradation.18
Much less is known about the regulation of ABCG1, andthe role of this transporter in the development of atheroscle-rosis remains controversial.19 ABCG1 mediates the removalof cholesterol to mature HDL particles and facilitates theclearance of esterified cholesterol from macrophages.20 Ue-hara et al have demonstrated that unsaturated fatty acidssuppress ABCG1 gene transcription.21 In the current study,we show that fatty acids of the 12/15LO pathway decreaseHDL-mediated cholesterol efflux from macrophages. Thisreduction in cholesterol efflux is associated with enhanceddegradation and serine phosphorylation of ABCG1. Thesenovel results support the concept that 12/15LO activity in thevessel wall impairs the clearance of excess cholesterol frommacrophages through regulation of ABCG1, thereby contrib-uting to the development of macrophage foam cell formation.
Materials and MethodsFor detailed methodology, please see the Data Supplement, availableonline at http://atvb.ahajournals.org. J774a.1 (J774) cells were cul-tured in DMEM containing 10% fetal bovine serum (FBS). J774cells stably overexpressing porcine leukocyte 12/15LO (Plox) andmock-transfected (Mock) J774 cells were kind gifts of Dr TanihiroYoshimoto (Kanazawa University School of Medicine, Kanazawa,Japan).22
Results12/15LO Activity Reduces HDL-MediatedCholesterol EffluxMacrophages in atherosclerotic lesions have been shown toexpress high levels of 12/15LO.23 Thus, we assessed the roleof 12/15LO and its eicosanoid products in regulating macro-phage function associated with RCT. We used mouse J774macrophages overexpressing porcine leukocyte 12/15LO(Plox) or mock-transfected (Mock) control cells. We havepreviously shown that Plox macrophages have increasedproduction of fatty acid hydroxy derivatives and a 3-foldincrease in 12SHETE production compared to Mock con-trols.9 In the current study, Mock and Plox macrophages werelabeled with [3H]cholesterol, and the effects of 12/15LOoverexpression on HDL- or apoA-I–mediated [3H]cholesterolefflux were measured. Tall and colleagues have reported thatHDL is the preferred cholesterol acceptor from ABCG1whereas lipid free apoA-I is the preferred cholesterol acceptorfrom ABCA1.24 12/15LO-overexpressing Plox macrophageshad a significant 30% reduction in HDL-mediated cholesterolefflux compared to Mock controls (Figure 1A). No significant
change in apoA-I–mediated cholesterol efflux between the 2groups was observed, suggesting that 12/15LO activity pref-erentially modulates ABCG1-mediated cholesterol efflux.
To further determine that 12/15LO activity is directlyresponsible for the reduction in cholesterol efflux, we inhib-ited expression of 12/15LO in Mock and Plox macrophagesusing an adenovirus expressing a ribozyme to 12/15LO(AdRz). We have previously used this ribozyme to reduce12/15LO expression by 60%.25,26 The 12/15LO ribozymerestored HDL-mediated cholesterol efflux in Plox macro-phages to levels similar to control (Figure 1A), indicating that12/15LO is able to regulate HDL-mediated cholesterol efflux.
12/15LO Activity Reduces ABCG1Cellular ExpressionBecause ABCG1 is known to regulate HDL-mediated cho-lesterol efflux, we assayed the effects of 12/15LO activity onABCG1 expression. We confirmed that 12/15LO protein wasoverexpressed in Plox macrophages (Figure 1B). Plox mac-rophages had a significant 60% reduction in ABCG1 proteinexpression (Figure 1B). No significant change in ABCA1protein expression was observed (Figure 1B). Inhibition of12/15LO in Plox macrophages with the 12/15LO ribozymeprevented the downregulation of ABCG1 protein expression(Figure 1C). These data suggest that the reduction in ABCG1expression and cholesterol efflux to HDL is regulated by12/15LO activity.
12/15LO Eicosanoids Reduce CholesterolEfflux to HDLWhen macrophages are presented with an amount of choles-terol that exceeds that which can be incorporated into cellularmembranes, the excess cholesterol is stored in macrophagesas cholesteryl esters. The enzyme acyl-coenzyme A (CoA):cholesterol acyltransferase (ACAT) esterifies excess choles-terol with long chain fatty acids, and the resulting cholesterylesters are stored in the cytoplasm as neutral lipid droplets. Toexamine whether fatty acid treatment enhanced cholesterylester formation, we treated J774 macrophages with arachi-donic or linoleic acid for 16 hours and measured esterifiedcholesterol accumulation (Wako Chemicals USA Inc). Treat-ment with arachidonic or linoleic acids did not significantlyalter macrophage cholesteryl ester content (data not shown).We next incubated J774 macrophages with fatty acid precur-sors and eicosanoid products of the 12/15LO pathway andmeasured cholesterol efflux. J774 macrophages were labeledwith [3H]cholesterol and incubated with arachidonic andlinoleic acids. Subsequently, HDL- and apoA-I–mediatedcholesterol efflux was measured. Incubation of J774 macro-phages with arachidonic or linoleic acids significantly re-duced HDL-mediated cholesterol efflux by 43% (Figure 1D),compared to control. Arachidonic and linoleic acids alsosignificantly reduced apoA-I–mediated cholesterol efflux by77% and 87%, respectively. These data are consistent withthe findings of Oram and colleagues indicating that unsat-urated fatty acids downregulate ABCA1 expression andfunction.16
To assess the role of 12/15LO eicosanoids on cholesterolefflux, J774 macrophages were labeled with [3H]cholesterol
1812 Arterioscler Thromb Vasc Biol October 2008
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from
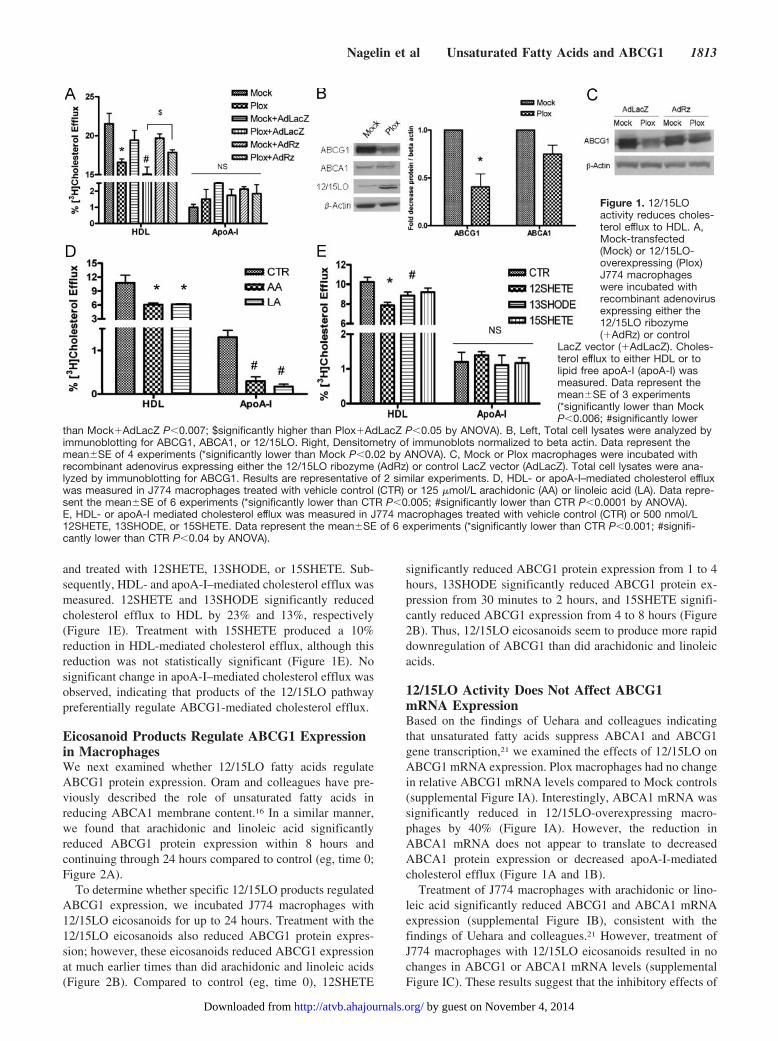
and treated with 12SHETE, 13SHODE, or 15SHETE. Sub-sequently, HDL- and apoA-I–mediated cholesterol efflux wasmeasured. 12SHETE and 13SHODE significantly reducedcholesterol efflux to HDL by 23% and 13%, respectively(Figure 1E). Treatment with 15SHETE produced a 10%reduction in HDL-mediated cholesterol efflux, although thisreduction was not statistically significant (Figure 1E). Nosignificant change in apoA-I–mediated cholesterol efflux wasobserved, indicating that products of the 12/15LO pathwaypreferentially regulate ABCG1-mediated cholesterol efflux.
Eicosanoid Products Regulate ABCG1 Expressionin MacrophagesWe next examined whether 12/15LO fatty acids regulateABCG1 protein expression. Oram and colleagues have pre-viously described the role of unsaturated fatty acids inreducing ABCA1 membrane content.16 In a similar manner,we found that arachidonic and linoleic acid significantlyreduced ABCG1 protein expression within 8 hours andcontinuing through 24 hours compared to control (eg, time 0;Figure 2A).
To determine whether specific 12/15LO products regulatedABCG1 expression, we incubated J774 macrophages with12/15LO eicosanoids for up to 24 hours. Treatment with the12/15LO eicosanoids also reduced ABCG1 protein expres-sion; however, these eicosanoids reduced ABCG1 expressionat much earlier times than did arachidonic and linoleic acids(Figure 2B). Compared to control (eg, time 0), 12SHETE
significantly reduced ABCG1 protein expression from 1 to 4hours, 13SHODE significantly reduced ABCG1 protein ex-pression from 30 minutes to 2 hours, and 15SHETE signifi-cantly reduced ABCG1 expression from 4 to 8 hours (Figure2B). Thus, 12/15LO eicosanoids seem to produce more rapiddownregulation of ABCG1 than did arachidonic and linoleicacids.
12/15LO Activity Does Not Affect ABCG1mRNA ExpressionBased on the findings of Uehara and colleagues indicatingthat unsaturated fatty acids suppress ABCA1 and ABCG1gene transcription,21 we examined the effects of 12/15LO onABCG1 mRNA expression. Plox macrophages had no changein relative ABCG1 mRNA levels compared to Mock controls(supplemental Figure IA). Interestingly, ABCA1 mRNA wassignificantly reduced in 12/15LO-overexpressing macro-phages by 40% (Figure IA). However, the reduction inABCA1 mRNA does not appear to translate to decreasedABCA1 protein expression or decreased apoA-I-mediatedcholesterol efflux (Figure 1A and 1B).
Treatment of J774 macrophages with arachidonic or lino-leic acid significantly reduced ABCG1 and ABCA1 mRNAexpression (supplemental Figure IB), consistent with thefindings of Uehara and colleagues.21 However, treatment ofJ774 macrophages with 12/15LO eicosanoids resulted in nochanges in ABCG1 or ABCA1 mRNA levels (supplementalFigure IC). These results suggest that the inhibitory effects of
Figure 1. 12/15LOactivity reduces choles-terol efflux to HDL. A,Mock-transfected(Mock) or 12/15LO-overexpressing (Plox)J774 macrophageswere incubated withrecombinant adenovirusexpressing either the12/15LO ribozyme(�AdRz) or control
LacZ vector (�AdLacZ). Choles-terol efflux to either HDL or tolipid free apoA-I (apoA-I) wasmeasured. Data represent themean�SE of 3 experiments(*significantly lower than MockP�0.006; #significantly lower
than Mock�AdLacZ P�0.007; $significantly higher than Plox�AdLacZ P�0.05 by ANOVA). B, Left, Total cell lysates were analyzed byimmunoblotting for ABCG1, ABCA1, or 12/15LO. Right, Densitometry of immunoblots normalized to beta actin. Data represent themean�SE of 4 experiments (*significantly lower than Mock P�0.02 by ANOVA). C, Mock or Plox macrophages were incubated withrecombinant adenovirus expressing either the 12/15LO ribozyme (AdRz) or control LacZ vector (AdLacZ). Total cell lysates were ana-lyzed by immunoblotting for ABCG1. Results are representative of 2 similar experiments. D, HDL- or apoA-I–mediated cholesterol effluxwas measured in J774 macrophages treated with vehicle control (CTR) or 125 �mol/L arachidonic (AA) or linoleic acid (LA). Data repre-sent the mean�SE of 6 experiments (*significantly lower than CTR P�0.005; #significantly lower than CTR P�0.0001 by ANOVA).E, HDL- or apoA-I mediated cholesterol efflux was measured in J774 macrophages treated with vehicle control (CTR) or 500 nmol/L12SHETE, 13SHODE, or 15SHETE. Data represent the mean�SE of 6 experiments (*significantly lower than CTR P�0.001; #signifi-cantly lower than CTR P�0.04 by ANOVA).
Nagelin et al Unsaturated Fatty Acids and ABCG1 1813
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

Figure 2. 12/15LO activity reduces cellular ABCG1. Cell lysateswere analyzed by immunoblotting for ABCG1 at each time point.Each time point is compared to control (CTR, eg, time 0) for sta-tistical evaluation. A, Top, J774 macrophages were incubatedwith vehicle control (CTR) or with arachidonic (AA) or linoleic(LA) acid (125 �mol/L) for up to 24 hours. Bottom, Densitometryof immunoblot normalized to beta actin. Data represent themean�SE of 3 experiments (*significantly lower than CTR P�0.04by ANOVA). B, Top, J774 macrophages were incubated with vehi-cle control (CTR) or with the indicated eicosanoids (500 nmol/L)for up to 24 hours. Bottom, Densitometry of immunoblot normal-ized to beta actin. Data represent the mean�SE of 3 experiments(*significantly lower than CTR P�0.05 by ANOVA).
1814 Arterioscler Thromb Vasc Biol October 2008
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from
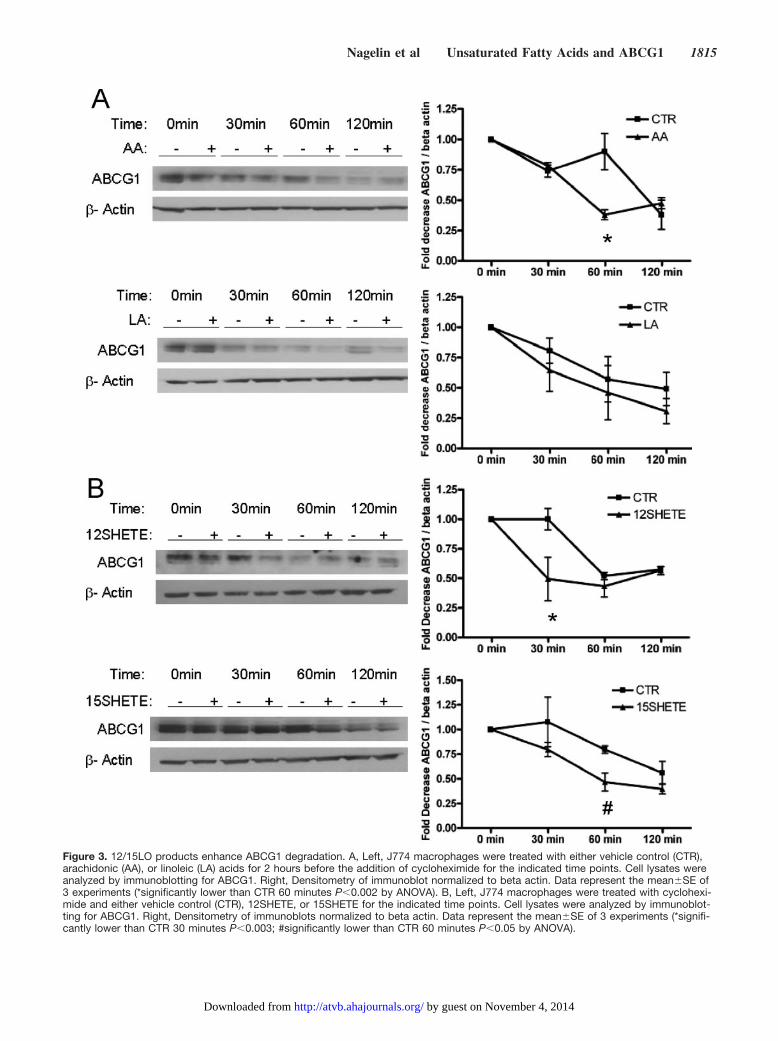
Figure 3. 12/15LO products enhance ABCG1 degradation. A, Left, J774 macrophages were treated with either vehicle control (CTR),arachidonic (AA), or linoleic (LA) acids for 2 hours before the addition of cycloheximide for the indicated time points. Cell lysates wereanalyzed by immunoblotting for ABCG1. Right, Densitometry of immunoblot normalized to beta actin. Data represent the mean�SE of3 experiments (*significantly lower than CTR 60 minutes P�0.002 by ANOVA). B, Left, J774 macrophages were treated with cyclohexi-mide and either vehicle control (CTR), 12SHETE, or 15SHETE for the indicated time points. Cell lysates were analyzed by immunoblot-ting for ABCG1. Right, Densitometry of immunoblots normalized to beta actin. Data represent the mean�SE of 3 experiments (*signifi-cantly lower than CTR 30 minutes P�0.003; #significantly lower than CTR 60 minutes P�0.05 by ANOVA).
Nagelin et al Unsaturated Fatty Acids and ABCG1 1815
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

12/15LO activity on HDL-mediated cholesterol efflux andABCG1 protein expression are not attributable to inhibitionof ABCG1 transcription or message stability. These data alsoindicate that unsaturated fatty acids and 12/15LO eicosanoidsregulate ABCG1 expression differently. Thus, unsaturatedfatty acids transcriptionally regulate ABCG1 expressionwhereas 12/15LO eicosanoids regulate ABCG1 expressionposttranscriptionally.
The 12/15LO product, 13SHODE, is a ligand for peroxi-some proliferator-activated receptor gamma (PPAR�).27
PPAR� has been shown to induce the expression of liver Xreceptor � (LXR�) thereby stimulating the expression ofABCA1 and ABCG1, leading to enhanced cholesterol ef-flux.28 Addition of 13SHODE to J774 macrophages re-sulted in a significant increase in LXR–� expression(supplemental Figure IC); however, 13SHODE treatmentdid not significantly induce ABC transporter expression,indicating that 13SHODE may regulate ABCA1 and ABCG1post-translationally.
12/15LO Products Enhance ABCG1 DegradationWe hypothesized that 12/15LO activity may reduce ABCG1protein expression through decreased translation or increasedprotein degradation. To measure the effect of 12/15LO ontranslation, J774 macrophages were treated with 12SHETE,arachidonic, or linoleic acids for 2 or 16 hours. Cells werelabeled with [35S]methionine for 20 minutes, and [35S]methi-onine incorporation into immunoprecipitated ABCG1 wasmeasured. Treatment with 12/15LO fatty acids did not changemethionine incorporation into ABCG1 (supplemental FigureII), indicating that reduced translation efficiency cannotaccount for the 12/15LO-mediated decrease in ABCG1expression.
To address the possibility that the 12/15LO pathwayregulates ABCG1 degradation, we treated J774 macrophageswith 12/15LO fatty acids in the presence of cycloheximide toarrest protein synthesis. We pretreated J774 macrophageswith arachidonic or linoleic acids for 2 hours before theaddition of cycloheximide. Arachidonic but not linoleic acidsignificantly reduced ABCG1 protein expression after treat-ment with cycloheximide for 60 minutes (Figure 3A). Toassess the role of the 12/15LO eicosanoids on ABCG1
degradation, we treated J774 macrophages with 12SHETE or15SHETE immediately before the addition of cycloheximide.12SHETE or 15SHETE also reduced ABCG1 protein expres-sion, but seemed to have a more rapid effect than arachidonicacid. 12SHETE and 15SHETE significantly reduced ABCG1expression after treatment with cycloheximide for 30 or 60minutes, respectively (Figure 3B). These data are consistentwith those of Figure 2, which shows rapid degradation ofABCG1 with 12/15LO eicosanoids compared to unsaturatedfatty acids.
12/15LO Products Enhance ABCG1Serine PhosphorylationWe hypothesized that enhanced degradation of ABCG1 maybe associated with posttranslational modifications. Oram andcolleagues determined that enhanced degradation of ABCA1is associated with serine phosphorylation.17 To measurephosphorylation changes in ABCG1, we used phosphoaminoacid-specific antibodies to determine whether 12/15LO eico-sanoids regulate serine, threonine, or tyrosine phosphoryla-tion. Immunoblots of immunoprecipitated ABCG1 revealedthat serine, threonine, and tyrosine phosphorylation occurs ata basal state (Figure 4). 12SHETE treatment significantlyenhanced serine, but not threonine or tyrosine, phosphoryla-tion (Figure 4). ABCG1 serine phosphorylation was inducedwithin 15 minutes after 12SHETE treatment and continuedthrough 1 hour of exposure. Thus, serine phosphorylationoccurs concurrently with 12/15LO eicosanoid-regulated deg-radation of ABCG1 (Figure 3B) and may account for theenhanced degradation rate.
Discussion12/15-lipoxygenase (12/15LO) is expressed in high levels inmacrophages found in early atherosclerotic lesions.10,11 12/15LO has been implicated in the pathogenesis of diabetes andatherosclerosis. Studies in mice have clearly shown that12/15LO activity contributes to the atherosclerotic burden.6,8
Moreover, studies have reported increased 12/15LO activityin patients with Type 2 diabetes, including increased renalexcretion 12/15LO eicosanoid metabolites.29 The 12/15LOpathway is upregulated by glucose, growth factors, andcytokines, so it is quite likely that this enzyme plays a
WB: p-Serine
WB: p-Threonine
WB: p-Tyrosine
IP: ABCG1 CTR 5min 15min 30min 60min
+12SHETE
CTR 5min 15min 30min 1hr0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5p-serinep-tyrosinep-threonine
Fold
cha
nge
phos
pho-
amin
o ac
id /
AB
CG
1
* *
WB: ABCG1
Figure 4. 12SHETE enhances ABCG1 serine phosphorylation. Left, J774 macrophages were treated with either vehicle control (CTR) or500 nmol/L 12SHETE for the indicated time points. ABCG1 was isolated by immunoprecipitation, and blots were probed with phospho-serine, phospho-threonine, phospho-tyrosine, or ABCG1 antibodies. Right, Densitometry of immunoblots normalized to ABCG1. Datarepresent the mean�SE of 3 experiments (*significantly higher than CTR P�0.05 by ANOVA).
1816 Arterioscler Thromb Vasc Biol October 2008
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

significant role in the cardiovascular complications associ-ated with diabetes. We have previously shown that 12/15LOactivity is upregulated in diabetic db/db mice,25 and that thesediabetic mice have reduced macrophage ABCG1 expressionand function.30 In the current study, we demonstrate for thefirst time that 12/15LO activity in macrophages increases thedegradation of ABCG1. ABCG1 is a key regulator of cho-lesterol efflux from peripheral cells to HDL in RCT.31 Thus,factors that reduce ABCG1 expression and function willimpact macrophage foam cell formation and cholesterolhomeostasis in the arterial wall, and may contribute to theaccelerated cardiovascular disease observed in patients withType 2 diabetes.
12/15LO overexpression in macrophages significantly re-duces HDL-mediated but not apoA-I–mediated cholesterolefflux, and this reduction in efflux corresponds with asignificant reduction in ABCG1, but not ABCA1, proteinexpression (Figure 1). Moreover, arachidonic and linoleicacids, and 12/15LO eicosanoid products also reduced HDL-mediated cholesterol efflux (Figure 1). Treatment of macro-phages with 12/15LO eicosanoids did not reduce cholesterolefflux to apoA-I, indicating a preferential role in targetingABCG1 (Figure 1).
Although the reduction in cholesterol efflux may appearmodest, we have previously found that a 20% reduction incholesterol efflux can lead to a 4-fold increase in cholesterylester accumulation.30 Rader and colleagues have demon-strated that knockdown of ABCG1 levels in J774 macro-phages by approximately 50% results in a significant reduc-tion in RCT.31 Thus, the changes in macrophage cholesterolefflux in response to fatty acids or 12/15LO products wouldhave a significant impact on RCT.
12/15LO activity in macrophages had no effect on ABCG1mRNA expression (supplemental Figure I). Uehara andcolleagues reported that unsaturated fatty acids can suppressABCG1 and ABCA1 gene expression.21 Consistent with theirfindings, we observed downregulation of ABCG1 andABCA1 mRNA expression by linoleic and arachidonic acids(supplemental Figure I). Interestingly, neither 12/15LO over-expression in macrophages nor incubation of macrophageswith 12/15LO eicosanoids produced significant changes inABCG1 mRNA expression (supplemental Figure I). BothABCA1 and ABCG1 gene expression are regulated by thenuclear hormone receptor, liver X receptor (LXR). It has beenpreviously shown that unsaturated fatty acids antagonize theactivation of LXR–� by oxysterols.32 Thus arachidonic andlinoleic acids might regulate ABCG1 and ABCA1 genetranscription through inhibition of LXR–� activation. Regu-lation of LXR–� is incompletely understood, and studieshave demonstrated important roles for phosphorylation andacetylation of LXR–� in regulating the activity of thistranscription factor.33,34 Posttranslational modifications ofLXR–� may account for upregulation of LXR–� expressionby 13SHODE with no corresponding upregulation of ABCtransporter mRNA expression (supplemental Figure I). Stud-ies to examine the effects of 12/15LO eicosanoids on LXRactivation are underway in the laboratory.
12/15LO could reduce ABCG1 protein expression throughdecreased message translation or enhanced protein degrada-
tion. 12/15LO fatty acids or eicosanoids did not alter trans-lation efficiency as determined by methionine incorporationinto de novo ABCG1 protein synthesis (supplemental FigureII). Treatment of J774 macrophages with 12SHETE in thepresence of cycloheximide indicated that protein degradationappears to be the likely mechanism by which 12/15LOproducts regulate ABCG1 levels (Figure 3). Surprisingly,unsaturated fatty acids and 12/15LO eicosanoids had differ-ential effects on ABCG1 protein expression. Incubation ofmacrophages with 12/15LO eicosanoids produced a rapidreduction in ABCG1 protein expression within 30 minutes to1 hour which continued through 4 to 8 hours, whereasincubation of macrophages with either arachidonic or linoleicacids reduced ABCG1 expression only after approximately 8hours of incubation (Figure 2). The time needed to convertfatty acid substrate into eicosanoid product might account forthe disparity in ABCG1 expression. Arachidonic and linoleicacid substrates also produced a more lasting effect in reduc-ing ABCG1 expression that continued 16 to 24 hours aftertreatment, whereas levels of ABCG1 returned to control (eg,time 0) levels by 16 to 24 hours of treatment with 12/15LOeicosanoids. Because cells rapidly metabolize eicosanoids,the return of ABCG1 levels to control levels is most likelyattributable to the metabolism of the eicosanoid and subse-quent loss of regulation. Alternatively, the return of ABCG1levels to control levels may be attributable to attempts of themacrophage to restore ABCG1 expression and cholesterolhomeostasis in a compensatory manner.
We anticipate that 12/15LO mediates posttranslationalmodifications of ABCG1, resulting in enhanced degradation.Indeed, we observed the presence of phosphorylated serine,threonine, and tyrosine residues in ABCG1 in the basal state(Figure 4). Treatment with 12SHETE enhanced ABCG1serine, but not threonine or tyrosine phosphorylation (Figure4). These data suggest that phosphorylation may be onepotential mechanism of regulation of ABCG1 protein expres-sion. Oram and colleagues have elegantly shown that unsat-urated fatty acids reduce ABCA1 expression, at least in partthrough phosphorylation that enhances degradation.17 Theseinvestigators did not examine the effects of 12/15LO eico-sanoids on ABCA1 in their study; however, our data (shownin Figure 1) indicate that 12/15LO overexpression does notsignificantly impact ABCA1 protein expression. We arecurrently working to identify the role that posttranslationalmodifications to ABCG1 play in regulating ABCG1 degra-dation by unsaturated fatty acids and eicosanoids. A directrole for serine phosphorylation in regulating ABCG1 degra-dation will need to be verified by mutational analysis.
Interestingly, Belkner et al demonstrated that 12/15LO-overexpressing macrophages have reduced intracellular lipidaccumulation and foam cell formation attributable to reduc-tions in SR-A expression and enhanced cholesteryl esterdegradation.35 However, these authors found that the 12/15LO products 12SHETE and 13SHODE failed to reduceintracellular cholesterol deposition and tended to enhancelipid accumulation.35 Moreover, cholesterol efflux to fetalcalf serum was not different between control and 12/15LO-overexpressing macrophages over 18 hours of incubation.35
Our study used specific acceptors (lipid free apoA-I or HDL)
Nagelin et al Unsaturated Fatty Acids and ABCG1 1817
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

to delineate the contribution of ABC transporters and shorterincubation time (4 hours), which may account for the differ-ences between the studies. Moreover, we demonstrated that12/15LO eicosanoids can directly regulate cholesterol effluxand ABCG1 expression. Thus, 12/15LO activity may regulateboth macrophage lipid uptake and efflux in a compensatorymanner, depending on the surrounding lipid environment inthe arterial wall. Additional mechanistic studies are needed todelineate the “proatherogenic” versus “antiatherogenic” prop-erties of this enzyme.
The findings of our current study have important implica-tions. Our data indicate that 12/15LO plays a role in regulat-ing cholesterol efflux from macrophages, thus providing anovel, important link between 12/15LO activity in the vesselwall and macrophage foam cell formation. Elevated levels ofplasma free fatty acids are prevalent among subjects withmetabolic syndrome and Type 2 diabetes, and these free fattyacids can accumulate in atherosclerotic lesions. The action ofthese fatty acids is 2-fold in the vessel wall. First, as shownby us and by Oram’s group, fatty acids have the ability todirectly regulate ABCG1 and ABCA1 expression and func-tion by enhancing ABC transporter degradation and inhibit-ing ABC transporter transcription. Second, these fatty acidsact as substrates for the 12/15LO enzyme to convert them intoeicosanoids, which cause ABCG1 degradation and reducedcholesterol efflux. We have previously reported regulation ofABCG1 transcription by chronic elevated glucose in Type 2diabetes.30 Taken together, our data indicate that ABCG1 isregulated by both glucose and free fatty acids, which hasimportant clinical implications for diabetic atherosclerosis.Understanding these novel mechanisms by which unsaturatedfatty acids and products of the 12/15LO pathway enhanceABCG1 degradation will be beneficial for developing newtherapies to regulate ABCG1 and ABCA1 expression inatherogenesis, especially in the setting of Type 2 diabetes.
AcknowledgmentsThe authors thank Dr John S. Parks (Wake Forest University) forrecombinant apoA-I and helpful advice regarding cholesterol efflux,and Dr David L. Brautigan (University of Virginia) for providinginvaluable advice regarding immunoprecipitation studies.
Sources of FundingThis work was supported by NIH P01 HL55798 (to J.L.N. andC.C.H.) and NIH R01 HL085790 (to C.C.H.).
DisclosuresNone.
References1. Sigari F, Lee C, Witztum JL, Reaven PD. Fibroblasts that overexpress
15-lipoxygenase generate bioactive and minimally modified LDL.Arterioscler Thromb Vasc Biol. 1997;17:3639 –3645.
2. McNally AK, Chisolm GM III, Morel DW, Cathcart MK. Activatedhuman monocytes oxidize low-density lipoprotein by a lipoxygenase-dependent pathway. J Immunol. 1990;145:254–259.
3. Yamamoto S, Suzuki H, Ueda N. Arachidonate 12-lipoxygenases. ProgLipid Res. 1997;36:23–41.
4. Kuhn H, Borngraber S. Mammalian 15-lipoxygenases. Enzymatic prop-erties and biological implications. Adv Exp Med Biol. 1999;447:5–28.
5. Yoshimoto T, Takahashi Y. Arachidonate 12-lipoxygenases. ProstaglandinsOther Lipid Mediat. 2002;68–69:245–262.
6. Cyrus T, Pratico D, Zhao L, Witztum JL, Rader DJ, Rokach J,Fitzgerald GA, Funk CD. Absence of 12/15-lipoxygenase expressiondecreases lipid peroxidation and atherogenesis in apolipoproteine-deficient mice. Circulation. 2001;103:2277–2282.
7. Bolick DT, Srinivasan S, Whetzel A, Fuller LC, Hedrick CC. 12/15lipoxygenase mediates monocyte adhesion to aortic endothelium in apo-lipoprotein E-deficient mice through activation of RhoA and NF-kappa B.Arterioscler Thromb Vasc Biol. 2006;26:1260–1266.
8. Reilly KB, Srinivasan S, Hatley ME, Patricia MK, Lannigan J, BolickDT, Vandenhoff G, Pei H, Natarajan R, Nadler JL, Hedrick CC. 12/15-lipoxygenase activity mediates inflammatory monocyte/endothelial inter-actions and atherosclerosis in vivo. J Biol Chem. 2004;279:9440–9450.
9. Wen Y, Gu J, Chakrabarti SK, Aylor K, Marshall J, Takahashi Y,Yoshimoto T, Nadler JL. The role of 12/15-lipoxygenase in theexpression of interleukin-6 and tumor necrosis factor-alpha in macro-phages. Endocrinology. 2007;148:1313–1322.
10. Yla-Herttuala S, Rosenfeld ME, Parthasarathy S, Sigal E, Sarkioja T,Witztum JL, Steinberg D. Gene expression in macrophage-rich humanatherosclerotic lesions. 15- lipoxygenase and acetyl low densitylipoprotein receptor messenger RNA colocalize with oxidation specificlipid-protein adducts. J Clin Invest. 1991;87:1146–1152.
11. Simon TC, Makheja AN, Bailey JM. Relationship of vascular 15-li-poxygenase induction to atherosclerotic plaque formation in rabbits.Prostaglandins Leukot Essent Fatty Acids. 1990;41:273–278.
12. Kuhn H, Belkner J, Suzuki H, Yamamoto S. Oxidative modification ofhuman lipoproteins by lipoxygenases of different positional specificities.J Lipid Res. 1994;35:1749–1759.
13. Kunjathoor VV, Febbraio M, Podrez EA, Moore KJ, Andersson L, KoehnS, Rhee JS, Silverstein R, Hoff HF, Freeman MW. Scavenger receptorsclass A-I/II and CD36 are the principal receptors responsible for theuptake of modified low density lipoprotein leading to lipid loading inmacrophages. J Biol Chem. 2002;277:49982–49988.
14. Vaughan AM, Oram JF. ABCA1 and ABCG1 or ABCG4 act sequentiallyto remove cellular cholesterol and generate cholesterol-rich HDL. J LipidRes. 2006;47:2433–2443.
15. Wang N, Silver DL, Costet P, Tall AR. Specific binding of ApoA-I,enhanced cholesterol efflux, and altered plasma membrane morphology incells expressing ABC1. J Biol Chem. 2000;275:33053–33058.
16. Wang Y, Oram JF. Unsaturated fatty acids inhibit cholesterol efflux frommacrophages by increasing degradation of ATP-binding cassette trans-porter A1. J Biol Chem. 2002;277:5692–5697.
17. Wang Y, Oram JF. Unsaturated fatty acids phosphorylate and destabilizeABCA1 through a phospholipase D2 pathway. J Biol Chem. 2005;280:35896–35903.
18. Arakawa R, Yokoyama S. Helical apolipoproteins stabilize ATP-bindingcassette transporter A1 by protecting it from thiol protease-mediateddegradation. J Biol Chem. 2002;277:22426–22429.
19. Curtiss LK. Is two out of three enough for ABCG1? Arterioscler ThrombVasc Biol. 2006;26:2175–2177.
20. Schmitz G, Langmann T, Heimerl S. Role of ABCG1 and other ABCGfamily members in lipid metabolism. J Lipid Res. 2001;42:1513–1520.
21. Uehara Y, Miura SI, von Eckardstein A, Abe S, Fujii A, Matsuo Y, RustS, Lorkowski S, Assmann G, Yamada T, Saku K. Unsaturated fatty acidssuppress the expression of the ATP-binding cassette transporter G1(ABCG1) and ABCA1 genes via an LXR/RXR responsive element.Atherosclerosis. 2007;191:11–21.
22. Sakashita T, Takahashi Y, Kinoshita T, Yoshimoto T. Essentialinvolvement of 12-lipoxygenase in regiospecific andstereospecific oxi-dation of low density lipoprotein by macrophages. Eur J Biochem. 1999;265:825–831.
23. Folcik VA, Nivar-Aristy RA, Krajewski LP, Cathcart MK. Lipoxygenasecontributes to the oxidation of lipids in human atherosclerotic plaques.J Clin Invest. 1995;96:504–510.
24. Wang N, Lan D, Chen W, Matsuura F, Tall AR. ATP-binding cassettetransporters G1 and G4 mediate cellular cholesterol efflux to high-densitylipoproteins. Proc Natl Acad Sci U S A. 2004;101:9774–9779.
25. Hatley ME, Srinivasan S, Reilly KB, Bolick DT, Hedrick CC. IncreasedProduction of 12/15 Lipoxygenase Eicosanoids Accelerates Monocyte/Endothelial Interactions in Diabetic db/db Mice. J Biol Chem. 2003;278:25369–25375.
26. Patricia MK, Natarajan R, Dooley AN, Hernandez F, Gu JL, Berliner JA,Rossi JJ, Nadler JL, Meidell RS, Hedrick CC. Adenoviral delivery of aleukocyte-type 12 lipoxygenase ribozyme inhibits effects of glucose andplatelet-derived growth factor in vascular endothelial and smooth musclecells. Circ Res. 2001;88:659–665.
1818 Arterioscler Thromb Vasc Biol October 2008
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

27. Nagy L, Tontonoz P, Alvarez JG, Chen H, Evans RM. Oxidized LDLregulates macrophage gene expression through ligand activation ofPPARgamma. Cell. 1998;93:229–240.
28. Chawla A, Boisvert WA, Lee CH, Laffitte BA, Barak Y, Joseph SB, LiaoD, Nagy L, Edwards PA, Curtiss LK, Evans RM, Tontonoz P. A PPARgamma-LXR-ABCA1 pathway in macrophages is involved in cholesterolefflux and atherogenesis. Mol Cell. 2001;7:161–171.
29. Antonipillai I, Nadler J, Vu EJ, Bughi S, Natarajan R, Horton R. A 12-li-poxygenase product, 12-hydroxyeicosatetraenoic acid, is increased in dia-betics with incipient and early renal disease. J Clin Endocrinol Metab.1996;81:1940–1945.
30. Mauldin JP, Srinivasan S, Mulya A, Gebre A, Parks JS, Daugherty A,Hedrick CC. Reduction in ABCG1 in type 2 diabetic mice increasesmacrophage foam cell formation. J Biol Chem. 2006;281:21216–21224.
31. Wang X, Collins HL, Ranalletta M, Fuki IV, Billheimer JT, Rothblat GH,Tall AR, Rader DJ. Macrophage ABCA1 and ABCG1, but not SR-BI,promote macrophage reverse cholesterol transport in vivo. J Clin Invest.2007;117:2216–2224.
32. Yoshikawa T, Shimano H, Yahagi N, Ide T, Amemiya-Kudo M, Mat-suzaka T, Nakakuki M, Tomita S, Okazaki H, Tamura Y, Iizuka Y,Ohashi K, Takahashi A, Sone H, Osuga J, Gotoda T, Ishibashi S, YamadaN. Polyunsaturated fatty acids suppress sterol regulatory element-bindingprotein 1c promoter activity by inhibition of liver X receptor (LXR)binding to LXR response elements. J Biol Chem. 2002;277:1705–1711.
33. Torra IP, Ismaili N, Feig JE, Xu CF, Cavasotto C, Pancratov R, RogatskyI, Neubert TA, Fisher EA, Garabedian MJ. Phosphorylation of liver Xreceptor alpha selectively regulates target gene expression in macro-phages. Mol Cell Biol. 2008;28:2626–2636.
34. Li X, Zhang S, Blander G, Tse JG, Krieger M, Guarente L. SIRT1deacetylates and positively regulates the nuclear receptor LXR. Mol Cell.2007;28:91–106.
35. Belkner J, Chaitidis P, Stender H, Gerth C, Kuban RJ, Yoshimoto T,Kuhn H. Expression of 12/15-lipoxygenase attenuates intracellular lipiddeposition during in vitro foam cell formation. Arterioscler Thromb VascBiol. 2005;25:797–802.
Nagelin et al Unsaturated Fatty Acids and ABCG1 1819
by guest on November 4, 2014http://atvb.ahajournals.org/Downloaded from

1
Data Supplement
Supplementary Figure Legends
FIGURE I. 12/15LO activity does not alter ABCG1 mRNA expression. Macrophage total
RNA was isolated and analyzed by quantitative real-time PCR for murine ABCG1, ABCA1,
LXRα, or LXRβ. Panel A. Mock and Plox macrophages. Data represent the mean +/- SE of 8
samples (*significantly lower than Mock p<0.006 by ANOVA). Panel B. J774 macrophages
were treated with either vehicle control (CTR) or with arachidonic (AA) or linoleic acids (LA).
Data represents mean +/- SE of 8 samples (*significantly lower than CTR p<0.0001;
#significantly lower than CTR p<0.002; $significantly lower than CTR p<0.0005 by ANOVA).
Panel C. J774 macrophages were treated with either vehicle control (CTR) or with 12SHETE,
13SHODE, or 15SHETE. Data represents mean +/- SE of 8 samples (*significantly higher than
CTR p<0.003 by ANOVA).
FIGURE II. 12/15LO fatty acids do not alter ABCG1 translation. J774 macrophages were
treated with either vehicle control (CTR), 12SHETE, arachidonic (AA), or linoleic acid (LA) for
2 or 16 hours. Cells were labeled with [35S]methionine for 20 minutes. ABCG1 was
immunoprecipitated, and 35S-labeled ABCG1 was detected by autoradiography. Results are
representative of 2 similar experiments.
Materials and Methods
Chemicals and Reagents. FBS was obtained from HyClone (Logan, UT, USA). HDL was from
Intracel (Frederick, MD, USA). NuPAGE 4-12% denaturing gels, MOPS running and transfer
buffers, and nitrocellulose was from Invitrogen (Carlsbad, CA, USA). Mouse anti-ABCG1
antibody was from Novus Biologicals, Inc. Mouse anti-ABCA1 antibody and human lipid-free
apoA-I were kind gifts from John Parks, Ph.D. at Wake Forest University. Mouse anti-beta actin
antibody was from Santa Cruz Biotechnology (Santa Cruz, CA, USA). RNeasy Mini kit was from

2
Qiagen (Valencia, CA, USA). Arachidonic acid, linoleic acid, 12SHETE, 13SHODE, and
15SHETE were purchased from BioMol (Plymouth Meeting, PA, USA). Cycloheximide was
from Sigma-Aldrich, Inc. (St. Louis, MO, USA).
Recombinant Adenoviral Delivery. Mock or Plox macrophages were infected at a multiplicity of
infection of 50 for 48 hours with the recombinant adenoviral vectors, AdRz (expresses 12/15LO
ribozyme) or AdLacZ control as we have described previously1.
Cellular Cholesterol Efflux Measurements. Cholesterol efflux assays were performed as
described, with minor modification2. J774 macrophages were plated in 12-well plates at a density
of 6x105 cells/well. To measure effects cholesterol efflux, cells were radiolabeled with 2 µCi/ml
of [3H]cholesterol for 16 hours in DMEM containing 10% fetal bovine serum (FBS). After
radiolabeling, cells were washed 3 times with PBS. Cells were incubated with 0.2% fatty acid
free BSA (FAFBSA) in DMEM in the presence of 125μM linoleic acid, arachidonic acid, or
vehicle control for 16 hours or the indicated eicosanoids (500nM) or vehicle control for 2 hours.
Cells were washed and allowed to equilibrate for an additional 2 hours in 0.2% FAFBSA.
Cholesterol efflux was conducted for 4 hours at 37°C in media containing: 1) 0.2% FAFBSA, 2)
0.2% FAFBSA + 15 µg/ml lipid-free human apolipoprotein A-I (apoA-I), or 3) 0.2% FAFBSA +
50µg protein/ml of human HDL. Human apoA-I and HDL were isolated as described
previously3,4. The efflux medium was then removed and a 100 µl aliquot was taken for 3H
radioactivity determination. Adherent cells were rinsed three times with cold PBS, cells were
dried, and isopropanol was added for overnight extraction at room temperature. A 100 µl aliquot
of the extract was taken for 3H radioactivity determination. Results are expressed as
[3H]cholesterol in medium/([3H]cholesterol in medium + cell) X 100%. Specific efflux to apoA-I
or HDL was calculated by subtracting non-specific efflux in the presence of 0.2% FAFBSA only.
Immunoblotting for ABCA1, ABCG1, and 12/15LO. J774 macrophages were incubated in 0.2%
FAFBSA in DMEM in the absence or presence of 125μM linoleic or arachidonic fatty acids,

3
500nM eicosanoids (HETEs and HODEs), or vehicle control as indicated in the Figure Legends.
RIPA buffer (50mMTrisHCl (pH 8.0), 150mM NaCl, 1% Igepal, 10mM NaF, 2mM Na3VO4
containing Sigma protease inhibitor cocktail) was added to cells after incubation to generate
whole cell lysates. Lysates were sonicated and protein was quantified via a protein assay kit
(BioRad, Hercules, CA, USA).
Proteins were separated by SDS-PAGE and transferred to nitrocellulose and blocked for
1 hour with 2.5% milk- Tris-buffered saline + 1% Tween 20 (TBST) at room temperature. Fifty
micrograms of whole cell lysate was used to detect all proteins. ABCA1 and ABCG1 antibodies
(1:500 dilution) were incubated with the blot at 4°C overnight in 2.5% Milk-TBST. The 12/15LO
antibody (1:1000 dilution) was incubated with the blot at 4°C overnight in 2.5% Milk-TBST.
ABCA1, ABCG1, and 12/15LO blots were then incubated with HRP conjugated anti-rabbit
secondary antibody (1:2000 dilution) in 2.5% Milk-TBST for 1 hour at room temperature. The
beta actin antibody (1:10000 dilution) was incubated with the blot for 1 hour at room temperature
in 2.5% Milk-TBST. Beta actin blots were then incubated with HRP conjugated anti-mouse
secondary antibody (1:5000 dilution) in 2.5% milk-TBST for 1 hour at room temperature.
Proteins were visualized using chemiluminescence and normalized to beta actin as gel loading
control. Densitometry was performed using Stratagene Zero D-Scan densitometry software.
Quantitative Real-Time PCR. J774 macrophages were treated with 0.2% FAFBSA with vehicle
control, 125μM arachidonic or linoleic acid for 8 hours, or 500nM 12/15LO eicosanoids (HETEs
and HODEs) for 4 hours. Total cellular RNA was collected from J774 macrophages using the
RNeasy Micro kit (Qiagen) following the manufacturer’s protocol. One microgram of cDNA was
synthesized using the Iscript cDNA synthesis kit (Bio-Rad). Total cDNA was diluted 1:10 in H2O
and 4μl were used for each real-time condition using a Bio-Rad MyIQ Single Color Real-Time
PCR Detection System and iQ SYBR Green supermix (Bio-Rad). Primer sequences are as
follows: ABCA1 Forward 5’-GGTTTGGAGATGGTTATACAATAGTTGT-3’ and Reverse 5’-

4
CCCGGAAACGCAAGTCC-3’; ABCG1 Forward 5’-TTCCCCTGGAGATGAGTGTC-3’ and
Reverse 5’-CAGTAGGCCACAGGGAACAT-3’; LXR alpha Forward 5’-
GGATAGGGTTGGAGTCAGCA-3’ and Reverse 5’-GGAGCGCCTGTTACACTGTT-3’; LXR
beta Forward 5’-GCTCAGGAGCTGATGATCCA-3’ and Reverse 5’-
GCGCTTGATCCTCGTGTAG-3’; Cyclophilin Forward 5’-
TGGAGAGCACCAAGACAGACA-3’ and Reverse 5’-TGCCGGAGTCGACAATGAT-3’.
Samples were normalized to cyclophilin using the ΔCt method.
Metabolic Labeling and Immunoprecipitation of ABCG1. J774 macrophages were incubated with
vehicle control, 12SHETE (500nM), arachidonic or linoleic acid (125μM) for 2 or 16 hours in
0.2% FAFBSA in DMEM. Cells were incubated with 200 μCi/mL [35S]methionine (New
England Nuclear) for 20 minutes at 37°C in 0.2% FAFBSA in DMEM. Cells were lysed with
RIPA buffer and lysates were incubated overnight at 4°C with antibody against ABCG1 (Novus
Biologicals). The antibody-antigen complex was isolated by protein A-sepharose beads
(Invitrogen) and resolved by SDS-PAGE. Each gel lane received equal amounts of
immunoprecipitated protein. The gels were dried, and [35S]methionine was detected by
autoradiography.
ABCG1 Degradation Rate. J774 macrophages were washed and incubated with 0.2% FAFBSA in
DMEM. Arachidonic or linoleic acid (125μM) or vehicle control was added 2 hours prior to the
addition of cycloheximide (20μg/mL) for the indicated time points, or 12SHETE (500nM) or
vehicle control was added immediately prior to the addition of cycloheximide (20μg/mL) for the
indicated time points. Cells were lysed with RIPA buffer and analyzed by SDS-PAGE and
immunoblotting for ABCG1.
Immunoprecipitation of ABCG1. J774 macrophages lysates were prepared with RIPA buffer.
Lysates were incubated overnight at 4°C with antibody against ABCG1 (Novus Biologicals). The
antibody-antigen complex was isolated by protein A-sepharose beads (Invitrogen) and resolved

5
by SDS-PAGE. Each gel lane received equal amounts of immunoprecipitated protein. ABCG1
phosphorylation was assayed by immunoblot analysis using phosphoserine (BioMol),
phosphothreonine, and phosphotyrosine (Santa Cruz Biotechnology) antibodies that recognize a
broad range of serine-, threonine-, and tyrosine-phosphorylated proteins.
Statistical Analysis. Data for all experiments comparisons between groups was performed using
analysis of variance (ANOVA) methods using the Statview 6.0 software program. Data is
graphically represented as the mean +/- SE, in which the mean consists of a minimum of three
experiments performed in triplicate. Comparisons between groups and tests of interactions were
performed assuming a two-factor analysis with the interaction term testing each main effect with
the residual error testing the interaction. All comparisons were made using Fisher’s least standard
difference procedure, so that multiple comparisons were performed at the alpha = 0.05 level only
if the overall F- test from the ANOVA is significant at p<0.055.
References
1. Patricia MK, Natarajan R, Dooley AN, Hernandez F, Gu JL, Berliner JA, Rossi JJ, Nadler JL, Meidell RS, Hedrick CC. Adenoviral delivery of a leukocyte-type 12 lipoxygenase ribozyme inhibits effects of glucose and platelet-derived growth factor in vascular endothelial and smooth muscle cells. Circ Res 2001;88:659-665.
2. Wang N, Silver DL, Thiele C, Tall AR. ATP-binding cassette transporter A1 (ABCA1) functions as a cholesterol efflux regulatory protein. J Biol Chem 2001;276:23742-23747.
3. Nichols AB, Ravenscroft C, Lamphiear DE, Ostrander LD, Jr. Independence of serum lipid levels and dietary habits. The Tecumseh study. JAMA 1976;236:1948-1953.
4. Parks JS, Rudel LL. Isolation and characterization of high density lipoprotein apoproteins in the non-human primate (vervet). J Biol Chem 1979;254:6716-6723.
5. Milliken GA. Methodologic issues. Stat Med 1995;14:701-702.

A B
C
Supplementary Figure I

CTR12
SHETE
AA LA CTR12
SHETE
AA LA
2 hours 16 hours
Supplementary Figure II
Copyright © 2022 FDOKUMEN




















