
1 Estructura trófica del ensamblaje de aves en tres ...
46
1 Estructura trófica del ensamblaje de aves en tres configuraciones del paisaje rural cafetero de Risaralda. Autora: Gloria Cristina Villabona Orozco Director: Gustavo Giraldo Quintero Trabajo de grado para optar al título de: Licenciada en Biología Universidad Distrital Francisco José de Caldas Facultad de Ciencias y Educación Proyecto Curricular de Licenciatura en Biología SILEAT Bogotá D.C. COLOMBIA 2018
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of 1 Estructura trófica del ensamblaje de aves en tres ...

1
Estructura trófica del ensamblaje de aves en tres configuraciones del paisaje rural
cafetero de Risaralda.
Autora:
Gloria Cristina Villabona Orozco
Director:
Gustavo Giraldo Quintero
Trabajo de grado para optar al título de:
Licenciada en Biología
Universidad Distrital Francisco José de Caldas
Facultad de Ciencias y Educación
Proyecto Curricular de Licenciatura en Biología
SILEAT
Bogotá D.C. COLOMBIA
2018

2
AGRADECIMIENTOS
El presente trabajo, se realizó gracias al apoyo del Grupo de Investigación Biología de la Conservación y Biotecnología de la Corporación Universitaria Santa Rosa de Cabal (UNISARC), responsable del Macroproyecto financiado por COLCIENCIAS (714/51270) titulado “Redes de Interacción entre vertebrados frugívoros y plantas en un paisaje rural heterogéneo de Risaralda”, con aspectos de logística (transporte, materiales y equipos), el diseño metodológico del macroproyecto en general, sin los cuales no habría sido posible llevar a cabo la investigación. Un agradecimiento especial a los docentes Jaime Andrés Carranza Quiceno y John Harold Castaño de la UNISARC quienes propusieron y ejecutaron este proyecto de investigación. A Dayro Leandro Rodriguez, Catalina Lopez Rico, Mariana Vélez Orozco de la UNISARC y Laura Melissa Romero Betancourt de la UDFJC integrantes del equipo de trabajo con quienes se llevaron a cabo extenuantes jornadas de campo y cuyo trabajo en equipo, solidaridad, constante ayuda, cariño y amistad fue posible la colección de datos para este trabajo, además de personal de apoyo como estudiantes del programa de biología de la UNISARC y la Universidad Javeriana quienes en ocasiones también brindaron su apoyo logístico, a todas las personas, campesinos y trabajadores quienes nos abrieron un espacio en sus fincas y en sus hogares, y con su gran hospitalidad que caracteriza a toda la gente linda de esta zona cafetera del país hicieron de la estancia en campo una experiencia inolvidable y maravillosa. Al docente Gustavo Giraldo Quintero director del Semillero de Investigación en Limnología, Ecología, Educación Ambiental y en Tecnología (SILEAT) del PCLB de la Universidad Distrital Francisco José de Caldas, director de este trabajo de tesis, por su constante apoyo con su tutoría, con la corrección y construcción de este documento. A mi familia, mi padre Jairo Alberto Villabona Vargas, mi madre en el cielo Gloria Mercedes Orozco Ríos, mi tía María de Jesús Villabona Vargas y mi hermano Leonardo Enrique Villabona Orozco, por ser mi constante apoyo económico, moral y emocional en toda mi vida y en mi formación académica. A la naturaleza con su grandeza y por supuesto a las aves, un grupo maravilloso envuelto en su infinidad de formas y colores, de plumas y cantos, que con su complejo universo pierde y enamora irremediablemente a quienes por un instante se detienen a observar y detallar su diseño perfecto. A todos aquellos ornitólogos que por décadas han caracterizado la avifauna de Colombia y que con sus trabajos científicos y guías de campo de alta calidad nos han permitido acercarnos al estudio de estos hermosos vertebrados.

3
DEDICATORIA
A mi padre Jairo Alberto Villabona Vargas quien con su esfuerzo, honestidad y tenacidad
me sacó adelante, me enseño a hacer grandes luchas en la vida, pero ante todo siempre
me enseño el respeto de la vida sobre todas las cosas, el amor por los animales, el
cuidado del agua y del medio ambiente, el amor por el saber y el conocimiento y el
aprecio por la causas justas y sociales, a él le debo todo lo que soy…
“Viajar es marcharse de casa, es dejar los amigos
es intentar volar
volar conociendo otras ramas recorriendo caminos
es intentar cambiar
Viajar es marcharse de casa,
es vestirse de loco
diciendo todo y nada con una postal,
Es dormir en otra cama,
sentir que el tiempo es corto,
viajar es regresar.”
Gabriel García Márquez

4
ÍNDICE
PORTADA…………………………………………………………………………………………1
AGRADECIMIENTOS……………………………………………………………………………2
DEDICATORIA……………………………………………………………………………………3
Resumen…………………………………………………………………………………………..6 Abstract…………………………………………………………………………………………….7 INTRODUCCIÓN GENERAL……………………………………………………………………8 1. Planteamiento del problema…………………………………………………………………..9
2. OBJETIVOS……………………………………………………………………………………10 2.1 Objetivo general……………………………………………………………………………...10
2.2 Objetivos específicos………………………………………………………………………..10 3. MARCO TEÓRICO…………………………………………………………………………….11 3.1 Generalidades de las aves…………………………………………………………………11 3.2 Estructura de Comunidades de Aves………………………………………………………11 3.3 Gremios tróficos……………………………………………………………………………..12 3.4 Pérdida y fragmentación del hábitat……………………………………………………….13 3.5 Índices ecológicos……………………………………………………………………………14 3.5.1 Índices de riqueza………………………………………………………………………....14 3.5.2 Índices de heterogeneidad…………………………………………………………….....14 3.5.3 Modelos de abundancia…………………………………………………………………15
3.5.4 Estimadores no paramétricos……………………………………………………………15 3.6 ANTECEDENTES………………………………………………………………………….16 4. METODOLOGÍA………………………………………………………………………………17 4.1 Área de estudio………………………………………………………………………………17
4.2 Caracterización del ensamblaje de aves………………………………………………….19
4.3 Muestras fecales……………………………………………………………………………..19

5
4.4 Colecta de datos…………………………………………………………………………….20
4.5 Categorización de los gremios alimenticios……………………………………………….20
4.5 Análisis de datos……………………………………………………………………………..20 5. RESULTADOS………………………………………………………………………………...22 5.1 Representatividad del muestreo …………………………………………………………...22 5.2 Composición del ensamblaje de aves…………………………………………………......23 5.3 Estructura trófica……………………………………………………………………………..25
6. DISCUSIÓN DE RESULTADOS……………………………………………………….……27
6.1 Representatividad del muestreo…………………………………………………………...27
6.2 Composición del ensamblaje de aves…………………………………………………..…28
6.3 Estructura trófica………………………………………………………………………….....29
7. CONCLUSIONES………………………………………………………………………….….31
8. RECOMENDACIONES……………………………………………………………………....32
9. REFERENCIAS BIBLIOGRAFICAS……………………………………………………...…33 10. ANEXOS……………………………………………….………………………………...…..39 10.1Hoja de campo para la toma de datos…………………………………………….….…..39
10.2 Lista de especies………………………………………………………………………..…39
10.3 Número de especies por cada tipo de escenario……………………………..……..…45
10.4 Número de individuos por cada tipo de escenario……………………………..…...….45
10.5 Especies endémicas para Colombia………………………………………….…….…....46
10.6 Especies migratorias…………………………………………………………………..…..46

6
Resumen
El objetivo de este proyecto fue describir la estructura trófica de la avifauna presente en
nueve localidades ubicadas en la zona rural de los municipios de Santa Rosa de Cabal,
Dos Quebradas y Marsella (Risaralda), las cuales fueron agrupadas en tres
configuraciones de paisaje con características estructurales distintas a escala local,
clasificadas como: agro ecosistema, fragmento de bosque rodeado de matrices
agroforestales o monocultivos y parche de bosque conservado. La metodología consistió
en caracterizar la composición del ensamblaje de aves a partir de muestreos que se
realizaron mediante redes de niebla y observación directa en campo. Posteriormente, se
colectaron las heces de las aves capturadas, con estos datos complementados con
literatura, se realizó la determinación taxonómica de las aves y la clasificación de los
gremios alimenticios presentes en el ensamblaje.
Se obtuvo como resultado: 19 Órdenes, 45 Familias, 18 subfamilias, 215 especies. Las
aves fueron categorizadas en función de su dieta de la siguiente manera; 5 gremios y 11
intergremios, siendo el gremio Insectívoro el más abundante con 60 especies, seguido del
intergremio Frugívoro-Insectívoro con 41 especies. Algunas aves poseen dietas muy
especializadas que las ubican en los gremios menos abundantes, especies como
Coereba flaveola, Chlorophanes spiza, Diglossa albilatera, Diglossa sittoides, que se
alimentan principalmente de néctar pero complementan su dieta con frutos e insectos en
menor medida.
En cuanto a las variaciones en los tres escenarios, el agro ecosistema representado por
áreas de pastizales y monocultivos, presentó una alta diversidad a pesar de la alta
intervención antrópica y transformación de áreas naturales, con respecto a los otros dos
escenarios no hay diferencias representativas, aunque a medida que el escenario esta
menos perturbado la riqueza aumenta levemente además de la presencia de especies
raras y endémicas. Las dietas predominantes en los tres escenarios son aquellas que se
basan principalmente en insectos, representadas en el gremio Insectívoro con el 27.9 %
del total de las especies del ensamblaje, seguido por el intergremio Frugívoro-Insectívoro
19.06 % del total de las especies y el más abundante de todo el ensamblaje en el
escenario de agroecosistema con 394 individuos registrados. El valor de diversidad para
la avifauna de esta zona cafetera, calculado con el método de Shannon-Wiener es:
Agroecosistema 4,27, Fragmento de bosque 4,58 y Bosque 4,48, para el índice de
diversidad Simpson el valor obtenido es de: Agroecosistema 0,9798, Fragmento de
bosque 0,9867 y Bosque 0,9838, lo que indica una diversidad alta para los tres
escenarios.
Este trabajo pretende aportar información acerca de la avifauna de esta zona cafetera y
aspectos de su ecología como los tipos de dieta presentes en el ensamblaje de aves. Con
el fin de analizar como la transformación de los sistemas naturales, la intensidad y el
modelo del cultivo del café y la extensión de los monocultivos sugieren una amenaza para
la diversidad de aves que habitan estas zonas boscosas. Vincular trabajos de
investigación con la participación de las comunidades en el seguimiento y conservación
de diversidad de aves en esta zona del eje cafetero es prioritario, para integrar la avifauna

7
como un componente importante en la conservación de áreas protegidas y el fomento del
aviturismo en la región.
Palabras claves: Dieta, ensamblaje gremios tróficos, paisaje transformado, región
cafetera.
Abstract
The objective of this project was to describe the trophic structure of the avifauna present in
nine localities located in the rural area of the municipalities of Santa Rosa de Cabal, Dos
Quebradas and Marsella (Risaralda), which were grouped into three landscape
configurations with characteristics different structural structures at local scale, classified
as: agro ecosystem, fragment of forest surrounded by agroforestry matrices or
monocultures and patch of conserved forest. The methodology consisted of characterizing
the composition of the birds' assemblage based on sampling conducted through fog nets
and direct observation in the field. Afterwards, the feces of the captured birds were
collected, with these data complemented with literature, the taxonomic determination of
the birds and the classification of the food guilds present in the assembly were made.
The result was: 19 Orders, 45 Families, 18 subfamilies, 215 species. The birds were
categorized according to their diet as follows; 5 guilds and 11 subcategories , with the
Insectívore guild being the most abundant with 60 species, followed by the Frugivore-
Insectivore intergrem with 41 species. Some birds have very specialized diets that place
them in the less abundant guilds, species such as Coereba flaveola, Chlorophanes spiza,
Diglossa albilatera, Diglossa sittoides, which feed mainly on nectar but supplement their
diet with fruits and insects to a lesser extent.
Regarding the variations in the three scenarios, the agro ecosystem represented by areas
of grasslands and monocultures, presented a high diversity despite the high anthropic
intervention and transformation of natural areas, with respect to the other two scenarios
there are no representative differences, although as the scenario is less disturbed, the
wealth increases slightly in addition to the presence of rare and endemic species. The
predominant diets in the three scenarios are those that are mainly based on insects,
represented in the Insectívoro guild with 27.9% of the total of the species of the assembly,
followed by the Frugivore-Insectivore intergremio 19.06% of the total of the species and
the most abundant of all the assembly in the agroecosystem scenario with 394 registered
individuals. The diversity value for the birds of this coffee zone, calculated with the
Shannon-Wiener method is: Agroecosystem 4.27, Forest fragment 4.58 and Forest 4.48,
for the diversity index Simpson the value obtained is of: Agroecosystem 0.9798, Fragment
of forest 0.9867 and Forest 0.9838, indicating a high diversity for the three scenarios.
This work aims to provide information about the avifauna of this coffee area and aspects of
its ecology as the types of diet present in the assembly of birds. In order to analyze how

8
the transformation of natural systems, the intensity and pattern of coffee cultivation and the
extension of monocultures suggest a threat to the diversity of birds that inhabit these forest
areas. Link research works with the participation of communities in the monitoring and
conservation of bird diversity in this area of the coffee axis is a priority, to integrate birdlife
as an important component in the conservation of protected areas and the promotion of
bird tourism in the region.
Keywords: Diet, assembly, trophic guilds, transformed landscape, coffee region.
INTRODUCCIÓN GENERAL
Los municipios de Santa Rosa de Cabal, Dos Quebradas y Marsella en el departamento de Risaralda (Colombia) se encuentran ubicados estratégicamente sobre una unidad de contexto regional, identificada como la Eco-región Eje Cafetero. Son municipios de vocación agrícola por excelencia, de acuerdo con datos regionales acerca de su cobertura y usos del suelo, los cultivos más predominantes son el café y el pasto manejado, así mismo, predominan el bosque secundario, la vegetación de páramo y el pasto natural. Esto permite inferir que tienen gran diversidad en flora y fauna, lo que se traduce en una riqueza paisajística que involucra escenarios de páramo, bosques y demás sitios que representan un potencial estratégico para su conservación (SIR, 2006).
El eje cafetero se presenta como una de las regiones con la mayor tasa de deforestación y pérdida de hábitat en Colombia, cerca del 78.09% del bosque andino ha sido transformado (CARDER 2004, Kattan et al, 2004) a sistemas productivos como la ganadería (para leche y carne) y cultivos (café, plátano, mixtos, guadua), donde la cobertura predominante es el pastizal (36.5%), seguida de los bosques (25.3%) y cultivos (19.8%) (Otero et al, 2005). Los paisajes de la región están constituidos por mosaicos de vegetación con una complejidad estructural diferente. Según lo anterior, la heterogeneidad paisajística se convierte en un factor determinante en la composición de su biodiversidad.
Colombia logra ser la nación del planeta con el mayor número de especies de aves y anfibios, y el segundo en especies de plantas, teniendo tan solo el 0,8% de la superficie del planeta Tierra. La región del Eje Cafetero no es una excepción, por el contrario, su variedad de climas, paisajes y ecosistemas albergan un gran número de especies de aves. Es difícil estimar la cantidad exacta en esta región, pero dicho número con seguridad supera las seiscientas cincuenta especies (Restrepo, 2016).
Teniendo en cuenta que las aves participan en procesos ecológicos relacionados con la polinización, la dispersión de semillas y el control biológico de insectos y/o plagas, detectar los cambios en las poblaciones de aves permite inferir el estado de los ecosistemas (Gómez & García, 2012).
Además de caracterizar la avifauna y su diversidad, analizar aspectos propios de su ecología permitirá entender más a fondo como se estructuran los conjuntos de aves en estos ecosistemas. En particular, la cuestión de cómo y dónde las distintas especies obtienen su alimento ha sido central en ecología de comunidades de aves por mucho tiempo (Lopez de Casenave, 2001). Varios estudios han mostrado la importancia del comportamiento de búsqueda de alimento y del uso de hábitat de alimentación en la

9
estructuración de los ensambles (Holmes, 1980), (Landres & MacMahon, 1983), sumando a esto el componente de transformación del hábitat a partir de procesos de fragmentación o desaparición total de las características originales de los ecosistemas, ya sea por procesos naturales de los mismos o por intervención del hombre, suponen una influencia importante en la disponibilidad de alimento y las formas de obtención del mismo por parte de los conjuntos de aves.
El conocimiento acerca de la biodiversidad de aves, su estructura funcional, para este caso interacciones de tipo trófico representadas en gremios alimenticios (redes mutualistas con plantas, control biológico de artropofauna, entre otros.) y la relación con la heterogeneidad de los ecosistemas dada por la trasformación del paisaje a partir de los sistemas productivos humanos se convierte en el objeto central de este trabajo.
Este trabajo fue realizado dentro del desarrollo del Macroproyecto de COLCIENCIAS (714/51270) titulado “Redes de Interacción entre vertebrados frugívoros y plantas en un paisaje rural heterogéneo de Risaralda”, cuyos muestreos fueron ejecutados entre Agosto del año 2016 y Octubre del 2017, siendo una de sus líneas de investigación el componente de aves. El análisis posterior de los datos obtenidos se realizó en el laboratorio del Semillero en Limnología, Ecología, Educación Ambiental y en Tecnología (SILEAT) del PCLB de la Universidad Distrital Francisco José de Caldas, bajo la tutoría del docente Gustavo Giraldo Quintero director del grupo, con la codirección del docente Jaime Andrés Carranza Quiceno; director del Grupo de Investigación Biología de la Conservación y Biotecnología de la Corporación Universitaria Santa Rosa de Cabal (UNISARC) y responsable del Macroproyecto. 1. Planteamiento del problema
La investigación acerca del papel que juegan las zonas rurales productivas del paisaje
cultural cafetero de Risaralda en la composición de la diversidad de la avifauna, se ha
venido incrementando en los últimos años; los listados, guías y libros de aves
presentados por distintas entidades estatales, privadas y académicas han permitido
comprender el impacto social y económico que tiene vincular estudios de diversidad a la
promoción del turismo como eje para la conservación de áreas protegidas. Sin embargo, y
a pesar de los esfuerzos, la biodiversidad está amenazada dada la eliminación de los
bosques, la transformación del modelo de cultivo del café y la transición a sistemas en
monocultivo, sistemas agroforestales, la ganadería extensiva, entre otros (Philptt et al,
2008).
Para realizar un análisis de la variabilidad que puede presentarse en la distribución espacio-temporal del ensamblaje de aves a partir de la caracterización de su organización gremial, se propone entonces; que las modificaciones al paisaje como resultado de la interacción dinámica entre factores ecológicos y procesos sociales humanos, que en conjunto forman sistemas socio-ecológicos, (Collins et al, 2010) juegan un papel determinante en la configuración de dicha variabilidad. La dinámica de estos sistemas, determina la forma en la que los grupos humanos se apropian de la naturaleza y las consecuencias ecológicas y sociales de dicha apropiación. Algunos de los grandes

10
elementos de investigación en los sistemas socio-ecológicos son la biodiversidad, las funciones de los ecosistemas y los servicios que éstos aportan a las sociedades.
El conocimiento acerca de la relación que existe entre la biodiversidad de aves presente
en estas zonas y su estructura funcional asociada a procesos ecológicos importantes para
los ecosistema es aún insuficiente; por lo tanto, con el propósito de aportar en este
campo, se propone las siguientes preguntas de investigación: ¿Cuál es la composición del
ensamblaje de aves presente en las zonas de estudio del paisaje rural cafetero en los
municipios de Santa Rosa de Cabal, Dos Quebradas y Marsella ubicados en el
departamento de Risaralda?, ¿Cuál es la estructura trófica en términos de gremios
alimenticios dentro del ensamblaje de aves presente en estas zonas?, y por último ¿Cuál
es la variación en la diversidad de la avifauna y la relación con su estructura trófica en los
tres escenarios de estudio los cuales presentan una composición paisajística distinta a
escala local?.
2. OBJETIVOS
2.1 Objetivo general
Caracterizar la estructura trófica del ensamblaje de aves presente en tres configuraciones
del paisaje rural cafetero de Risaralda.
2.2 Objetivos específicos
Identificar la composición del ensamblaje de aves en tres configuraciones del paisaje rural
cafetero de Risaralda.
Determinar los gremios tróficos presentes en el ensamblaje de aves en tres
configuraciones del paisaje rural cafetero de Risaralda.
Analizar la variación en la diversidad de la avifauna y su relación con la estructura trófica
en los tres escenarios de estudio.

11
3. MARCO TEÓRICO
3.1 Generalidades de las aves
Las aves representan un grupo diverso de interés científico, la diversidad de sus formas y conducta, la facilidad para su observación permite que sea uno de los grupos más ampliamente estudiados y su conocimiento ha permitido la generación de información sobre procesos ecológicos y biogeográficos relevantes (Sosa 2003, Perovic et al. 2008).
Se encuentran en casi todos los ecosistemas del mundo, debido a su capacidad de desplazamiento han desarrollado diferentes estrategias de vida logrando acceder a una amplia gama de recursos alimenticios, otorgándoles funciones importantes en los ecosistemas (polinización, depredación y control de plagas, eliminación de animales en descomposición, dispersión o propagación de semillas), constituyen parte de las cadenas tróficas como fuente alimenticia de otras especies animales y como depredadoras. Esta capacidad de movimiento les ha facilitado también desarrollar la habilidad de migración como técnica de sobrevivencia en diferentes gradientes ambientales (Sosa, 2003).
La biodiversidad en Colombia se cataloga como uno de sus grandes recursos, la cual le brinda servicios ecosistémicos invaluables, además de perfilar al país a nivel mundial como un destino ecoturístico de parajes naturales increíbles y de lugares completamente desconocidos para la investigación científica, uno de los grupos más representativos son las aves, siendo hasta ahora reconocido como el más numeroso y el primero a nivel mundial, de acuerdo con datos del Sistema de Información sobre Biodiversidad en Colombia (SIB) para el año 2017 el país posee alrededor de 1921 especies de aves registradas, de las cuales 79 solo se encuentran en nuestro territorio, ubicándolo como uno de los países más ricos en estas especies en Sudamérica. La región del Eje Cafetero con seguridad supera las 650 especies (Restrepo, 2016), distintas entidades oficiales (Red Nacional de Observadores de Aves de Colombia, la Corporación Autónoma de Risaralda, Sistema de Información en Biodiversidad de Risaralda (SIBIR) al año 2017 estiman que el departamento de Risaralda aproximadamente tiene el 80% de las aves registradas para el Eje Cafetero y el 44% de todo el territorio nacional, en el que apenas ocupa el 0.36 por ciento de su superficie.
3.2 Estructura de Comunidades de Aves La definición y delimitaciones acerca del concepto de comunidades biológicas han sido puntos focales de la ecología. Entre algunas definiciones; se puede considerar que las comunidades están formadas por un ensamblaje de poblaciones de diferentes especies que coexisten espacial y temporalmente y que durante la evolución han tenido procesos coevolutivos (Andraca 2010). La estructura de las comunidades puede estar determinada por factores no balanceados y estocásticos, por interacciones que se establecen entre las distintas especies (competencia, depredación, mutualismo y el parasitismo) y pueden ocurrir o no simultáneamente por procesos evolutivos (colonización, extinción y especiación) (Wilson 1989). Desde el enfoque de la ecología de comunidades, la diversidad de especies es esencialmente un concepto relacionado con la estructura de las comunidades, el cual

12
puede ser cuantificable: el recíproco de un promedio de las abundancias relativas (Hill, 1973). La estructura biológica de las comunidades está caracterizada por una única especie o pocas especies predominantes dentro de una comunidad, las que superan en número a las otras poseen mayor cantidad de biomasa y realizan la mayor contribución al flujo de energía. Las comunidades se pueden caracterizar por la diversidad de especies, que engloba dos componentes: la riqueza de especies, el número de especies dentro de una comunidad y equitatividad, es decir, cómo se reparten los individuos entre las distintas especies (Ricklefs, 1996). La composición de las comunidades de aves al igual que cualquier otro grupo biológico responde a distintas y numerosas variables geográficas, medioambientales, funcionales, de perturbación tanto natural como antrópica, entre otras; con las cuales la comunidad científica ha venido trabajando para intentar poner de manifiesto los patrones que determinan la riqueza de especies. Básicamente se ha tratado de documentar la importancia que tienen la complejidad estructural de la vegetación, elevación, latitud, predictibilidad de los recursos, etc., sobre la diversidad de las comunidades de aves en todo el mundo (Amat 1984). 3.3 Gremios tróficos Para contextualizar, la composición de la estructura gremial, cabe aclarar que sus definiciones son ampliamente discutidas, la controversia frente a este término, la definición y delimitación del concepto “gremio” se dan desde varios aspectos; teóricos, semánticos e incluso metodológicos, frente a los cuales los ecólogos han venido debatiendo fuertemente en las últimas décadas. Para fijar un punto de convergencia se parte entonces de que la estructura en gremios de una comunidad se define como la expresión de las similitudes y diferencias entre los patrones de uso de recursos de las especies que lo componen. En este contexto, determinar la estructura gremial consiste en separar a las especies en grupos sobre la base de su utilización de los recursos y analizar dicha estructura es explorar cómo utilizan los recursos las especies de cada grupo (Holmes & Recher, 1986); (López de Casenave, 2001). En 1967, Richard Root definió formalmente el término “gremio”; como un grupo de especies que explotan la misma clase de recursos ambientales de una manera similar (Root, 1967). Para Root la estructura en gremios de una comunidad es el resultado de la acción de dos fuerzas opuestas: las adaptaciones a determinados modos de vida o a determinados tipos de recurso compartidas por algunas especies actuarían agrupándolas en gremios, mientras que las interacciones competitivas tenderían a separarlas ecológicamente dentro de su gremio (Bradley RA & Bradley DW, 1985), (Hawkins & MacMahon, 1989), (Wiens,1989), (Simberloff & Dayan, 1991). Actualmente se ha tratado de “clasificar a los gremios”, estableciendo generalidades a partir de consensos entre las hipótesis construidas bajo cierta objetividad de autores que han trabajado este término, Gitay y Noble (1997) distinguen entre grupos basados en el uso de los recursos por parte de las especies (gremios sensu estricto) y grupos basados en la respuesta de las especies a modificaciones del ambiente (perturbaciones) (Gitay & Noble, 1997).

13
Wiens (1989) por su parte, basándose parcialmente en Jaksic (1981), reconoce dos aproximaciones generales mediante las cuales se han caracterizado gremios: la agremiación a priori y la agremiación a posteriori (Morrison et al, 1992). En el primer caso, la asignación de las especies a los gremios se realiza de manera subjetiva, ya que, si bien puede estar basada en información cualitativa o cuantitativa como observaciones de campo o datos publicados previamente, no se realiza una clasificación objetiva por medio de algún análisis estadístico de la misma. Por el contrario, la agremiación a posteriori está basada en análisis objetivos de observaciones cuantitativas en campo con la finalidad de agremiar a las especies. Debido a que la obtención de la información requerida para agremiar a posteriori es mucho más costosa, ésta ha sido menos utilizada, al menos en comunidades de aves. De acuerdo a la clasificación adaptada por Castaño y Patiño (2007) las subcategorías de los gremios tróficos pueden ser: Frugívoro, Frugívoro-Insectívoro, Frugívoro-Insectívoro-Nectarívoro, Insectívoro, Nectarívoro-Insectívoro y Rapaz. Con respecto a los gremios, se puede distinguir entre los procesos que actúan en el seno de los mismos (i.e., intragremiales) y aquellos que son responsables de la determinación de la estructura gremial de la comunidad (Lopez de Casenave, 2001). Dichos gremios se agrupan en categorías y subcategorías en función de las particularidades de la dieta de las aves. 3.4 Pérdida y fragmentación del hábitat. Las características del hábitat tienen influencia importante sobre las aves, por ejemplo, la estructura y composición florística de la vegetación pueden determinar, entre otros aspectos, la distribución y abundancia del alimento, la disponibilidad de perchas para cantar, de cobertura contra predadores y de sitios para nidificar (Santos & Tellería 2006; Cody, 1985), considerando el papel del hábitat, los estudios se han enfocado en analizar como las especies lo utilizan y que procesos podrían dar cuenta del patrón de selección de hábitat observado (interacciones competitivas, respuestas específicas de las especies) (Santos & Tellería 2006; Wiens, 1989). La configuración del paisaje de un lugar determinado puede estar constituido por un conjunto de ecosistemas variados, dicha composición está sujeta a perturbaciones o cambios a pequeña y gran escala que pueden darse de forma natural en la dinámica de los mismos o a partir de actividades antrópicas que tiene una incidencia directa en procesos ecológicos y por supuesto en la conformación y comportamiento de la diversidad allí presente. Principalmente en la actualidad los sistemas productivos humanos en distintos campos de la economía; desde la agricultura hasta la utilización para construcción de vivienda, entre otras numerosas actividades humanas, es uno de los factores que constituyen la transformación o la desaparición total de regiones naturales en el mundo. Con la fragmentación y destrucción de un hábitat se produce un cambio progresivo en la configuración del paisaje que puede definirse adecuadamente mediante las tendencias de variables paisajísticas, que cambian simultáneamente y que tienen en conjunto una incidencia perniciosa sobre la supervivencia de las especies afectadas (Santos & Tellería 2006; Saunders et al., 1991; Andrén, 1994; Fahrig, 2003) algunas de estas variables son: una pérdida regional en la cantidad de hábitat, una disminución del tamaño medio y un

14
aumento del número de los fragmentos de hábitat resultantes, un aumento de la distancia entre fragmentos, un aumento de la relación perímetro/superficie. En todos estos casos, las especies de los hábitats en retroceso ven mermar el territorio disponible a la vez que se enfrentan a una creciente atomización de sus poblaciones. Este proceso es tan antiguo como la expansión agrícola de la humanidad, solo que ahora se ha intensificado por una capacidad tecnológica que no conoce barreras. De esta forma, el hombre ha alterado en su propio beneficio la mayor parte de la tierra emergida útil (Santos & Tellería 2006;Loh y Wackernagel, 2004). En Colombia la tasa anual de deforestación en 2013 llegó a valores superiores a 300 mil hectáreas, también en la Ecorregión Cafetera, un territorio biodiverso que alberga al 7% de las especies de plantas y animales del país donde el paisaje estuvo dominado por bosques, ahora solo se conserva menos del 20% de dicha cobertura. Para el IDEAM, mientras la cifra entre 1990 y 2010 llegó a 310 mil hectáreas-año, y en el Chocó se pierde la batalla contra la deforestación: la Región Andina fue la zona más afectada, seguida de la Amazonía (Duque, 2014), lo que se traduce en una transformación del paisaje cafetero, de los ecosistemas naturales y sus áreas de conservación que juegan un papel altamente significativo en la composición de la avifauna de la zona. 3.5 Índices ecológicos La mayoría de los métodos estadísticos propuestos se inclinan a la evaluación de la diversidad dentro de las comunidades (alfa), en la cual se consideran métodos basados en la cuantificación de especies presentes (riqueza específica); y los basados en la estructura de la comunidad (distribución proporcional del valor de importancia de cada especie) (Hurtado, 2015). Según Garmendia y Samo (2005) existen varios índices para estimar la complejidad de una comunidad. Estos índices se pueden agrupar en: 3.5.1 Índices de riqueza Consideran únicamente la riqueza de especies, definidos como número de objetos contenidos en un determinado espacio (Ibáñez y Álvarez 2002). Suelen ser independientes del tamaño de la muestra, lo que dificulta la comparación entre comunidades (Garmendia y Samo 2005). Entre los más conocidos destacan los estimadores de Chao, los métodos de rarefacción, el denominado “Jacknife”. Entre los métodos no paramétricos está el propuesto por Smith y van Belle en 1984 y los más recientes por Southwood y Henderson (2000) entre otros ofrecen un sumario de los índices más utilizados para estimar la riqueza. 3.5.2 Índices de heterogeneidad Consideran tanto la uniformidad como la riqueza de especies, dentro de estos últimos se distinguen: los índices estadísticos que proceden de la teoría de la información y le conceden mayor peso a las especies raras. Un índice de este tipo es el de Shannon (H), que requiere el muestreo al azar de todos los individuos y que estén representadas todas las especies de la comunidad en la muestra. Éste índice proviene de la teoría de la información, donde se utiliza para estimar el máximo de información que puede llegar a contener un mensaje, la fórmula se describe a continuación:

15
Donde pi es la proporción de individuos de cada una de las especies en la comunidad. Esta proporción se estima a partir de n/N, que es la relación entre el número de individuos de la especie i (ni) y el número total de individuos de todas las especies (N). Los índices de dominancia, conceden mayor peso a las especies comunes, dentro de estos se encuentra el de Simpson, se determina la probabilidad de que al extraer dos individuos al azar de la comunidad, fuesen de la misma especie (D) el inverso de esta probabilidad es el índice de Simpson (S).
Donde pi es la proporción de individuos de cada una de las especies en la comunidad. El índice de similitud de Jaccard expresa el grado en que dos muestras son semejantes y es una medida inversa de la diversidad beta, que se refiere al cambio de especies entre dos muestras. El intervalo de valores para este índice es 0 cuando no hay especies compartidas entre ambos sitios, hasta 1 cuando los sitios tienen la misma composición de especies. A partir del valor de similitud (s) se puede calcular fácilmente la disimilitud (d) entre las muestras: d = 1 – s. Los lugares que se agrupan y tienen distancias menores entre ellos son los más similares en la composición de especies. Por el contrario, los que más se distancian son los que presentan menor similitud (Moreno 2001). 3.5.3 Modelos de abundancia Se ajustan mejor para describir la distribución de determinados tipos de objetos (Ibáñez y Álvarez 2002). Las diferencias en las abundancias de especies en las comunidades plantean dos problemas: primero, el número total de especies incluidas en una muestra, porque a medida que se capturan más individuos la probabilidad de encontrar especies raras aumenta. Por lo tanto, se puede comparar la diversidad entre áreas muestreadas con diferentes intensidades simplemente contando las especies. Segundo, no todas las especies deben contribuir por igual a la estimación de la diversidad total porque sus papeles funcionales en una comunidad varían, en cierto grado, en proporción a su abundancia (Garmendia y Samo 2005). Las curvas de rango de abundancia son posiblemente la mejor forma de representar la diversidad de una comunidad, puesto que se ve representada toda la distribución de las abundancias relativas por lo que es más fácil ver lo que está ocurriendo en la comunidad. 3.5.4 Estimadores no paramétricos Estos estimadores permiten hacer una comparación matemática, entre la riqueza estimada y la riqueza obtenida, para poder analizar la representatividad de los muestreos realizados. Algunos autores han considerado a los estimadores no paramétricos como el avance más importante en la medida de la biodiversidad en los últimos tiempos

16
(Magurran, 2004). Los modelos no paramétricos han sido llamados también libres de distribución (distribution-free) porque los datos no asumen un tipo de distribución particular ni una serie de supuestos a priori que los ajusten a un modelo determinado. El cálculo de los modelos no paramétricos es más sencillo y rápido, son más fáciles de entender y explicar, y son relativamente efectivos (Espinosa, 2003). Los principales modelos no paramétricos que se han empleado para la estimación de la riqueza, son: Bootstrap que estima la riqueza de especies a partir de la proporción de muestras que contienen a cada especie. ICE que se basa en el número de especies raras (las observadas en menos de 10 unidades de muestreo), Chao 1 basado en la abundancia; esto quiere decir que los datos que requiere se refieren a la abundancia de individuos que pertenecen a una determinada clase en una muestra, una muestra es cualquier lista de especies en un sitio, localidad, cuadrante, país, unidad de tiempo, trampa, etc. Chao 2 tiene en cuenta a las especies observadas en exactamente una y dos unidades de muestreo; Jack 1 es una función del número de especies presentes en sólo una unidad de muestreo, mientras que Jack 2 considera también a las especies presentes en dos unidades de muestreo (Oreja et al, 2010).
3.6 ANTECEDENTES En Colombia durante las últimas dos décadas, las asociaciones de investigadores y observadores han estimulado la investigación y observación de aves con el fin de adquirir información de las diferentes especies (Renjifo et al. 2000). Más de 2000 especies de aves, aproximadamente el 22% de todas las especies conocidas, habitan los Andes de Colombia, Ecuador, Perú y Bolivia, desde las estribaciones (500 msnm) hasta la nieve. Casi 600 especies (28%) son endémicas, lo que cataloga a esta región como una de las más diversas en la Tierra (Herzog & Kattan 2011). Los ecosistemas naturales de los Andes colombianos han sufrido grandes transformaciones por prácticas como la deforestación, la agricultura y la ganadería (Andrade 1993, Cavelier 1997, Kattan 1997; Kattan & Álvarez-López 1996). De tal manera que las investigaciones sobre diversidad en esta región han venido incrementándose como respuesta a la preocupación de la creciente desaparición de hábitats boscosos y áreas conservadas. La región del Eje Cafetero se localiza en el centro del occidente de Colombia y forma parte de la región andina, sobre las cordilleras Occidental y Central, entre los departamentos de Caldas, Risaralda, Quindío, la región norte del Valle del Cauca, y el noroccidente del Tolima; cuenta con gran diversidad climática, edáfica, topográfica y por lo tanto ecológica (Nadachowski & Valencia, 2009), acompañada por una amplia diversidad cultural, estas características se ven reflejadas en sus paisajes y ecosistemas los cuales albergan un gran número de especies de aves, lo que ha permitido que los estudios acerca de la composición en la diversidad de la avifauna presente en el paisaje cultural cafetero de Risaralda hayan ido incrementándose; los listados, guías y libros de aves presentados por distintas entidades estatales, privadas y académicas han permitido comprender el impacto social y económico que juegan las zonas rurales productivas y la importancia que tiene vincular estudios de diversidad a la promoción del turismo como eje para la conservación de áreas protegidas.

17
En el límite interdepartamental entre Risaralda, Quindío y Valle, el proceso de cambio de uso del suelo ha sido bastante fuerte; actualmente existen 41.158 ha bajo pasturas (Camargo et al. 2007) y siguen aumentando. La transformación de los sistemas naturales, la intensidad y el modelo del cultivo del café y la extensión de los monocultivos sugieren una amenaza para la diversidad de aves que habitan estas zonas (Borrero, 1986). A pesar de la transformación de los ecosistemas, el paisaje cultural cafetero se constituye como una zona con alta diversidad de aves, varios trabajos se han realizado caracterizando la composición y estructura de la avifauna de esta región, caracterización de dieta, su importancia ecológica e incluso algunos autores han enfocado y vinculado sus trabajos a la participación de las comunidades en el seguimiento y conservación de diversidad de aves: López et al.(2013), Londoño (2011), Lentijo & Kattan,(2005), Lentijo et al, (2005), Cardona et al, (2012), CORPOCALDAS & Fierro, (2010), Sáenz et al, (2013), Henao et al, (2014), Gómez & García, (2012), Restrepo,(2016), Botero & Lentijo (2014), Botero et al.( 2012), entre otros.
4. METODOLOGÍA 4.1 Área de estudio
El estudio se llevó a cabo en nueve localidades del paisaje rural cafetero, el área de estudio donde se ubican principalmente la mayoría de las localidades de muestreo, se encuentra en la zona rural del Municipio de Santa Rosa de Cabal en el departamento de Risaralda, localizado en el flanco occidental de la Cordillera Central entre alturas que van desde los 5.200 m.s.n.m. en el Nevado de Santa Isabel hasta los 1.400 en la vereda Las Mangas. Limita por el Norte con Chinchiná y Villamaría, por el Sur con Pereira y Dosquebradas, por el Oriente con el Departamento del Tolima y por el Occidente con Marsella y Dosquebradas. Santa Rosa de Cabal hace parte de la Subregión I con una extensión territorial aproximada de 544,3 km2.
La precipitación en este municipio, varía entre 1.700 y 3.100 mm/año, la precipitación media anual de todo el municipio es de 2.709,9 mm. En el municipio se identifican: Clima Medio, Clima Frío, Clima Muy Frío, Clima Extremadamente Frío, Clima SubNival, Clima Nival. (CARDER, 2004), es una zona con las condiciones propias del bosque húmedo premontano según la clasificación de Holdridge (1987). La localidad de la Nona se encuentra ubicada en los límites con el Municipio Marsella, las localidades de la Selva y Ángela María se encuentran ubicadas en los límites con el Municipio de Dos Quebradas. Estos tres municipios se encuentran ubicados estratégicamente sobre una unidad de contexto regional, identificada como la Eco-región Eje Cafetero.
Los sitios de muestreo se ubican en el rango de elevación entre 1600 y 2000 msnm. Las localidades fueron escogidas bajo características paisajísticas particulares, que se clasificaron de acuerdo a su composición vegetal, densidad de bordes boscosos, intervención de sistemas productivos (monocultivos de café u otros, sistemas agroforestales, etc.) teniendo en cuenta métricas del paisaje propuestas por Farina (2008) y categorías de coberturas en el Eje Cafetero; según el Sistema Regional de Áreas Protegidas del Eje Cafetero (SIRAP EC) para el 2011, las coberturas de tierra en el Eje

18
Cafetero se clasifican de la siguiente manera: Territorios artificializados (zonas urbanizadas, zonas industriales o comerciales y redes de comunicación, zonas de extracción minera y escombreras y zonas verdes artificializadas no agrícolas), Territorios agrícolas (cultivos transitorios, cultivos permanentes, pastos desnudos, áreas agrícolas heterogéneas) Bosques y áreas semi-naturales (Bosques, áreas con vegetación herbácea y/o arbustiva, áreas abiertas sin o con poca vegetación), Áreas húmedas (áreas húmedas continentales), Superficies de agua (Aguas continentales) y áreas sin información.
Las localidades fueron clasificadas en tres configuraciones de paisaje de la siguiente manera: agroecosistemas: Jazmín (1642 msnm), Leona (1819 msnm) y Manzanillo (1616 msnm), fragmento de bosque rodeado de matrices con sistemas productivos: Chorros (1820 msnm), Ángela María (1990 msnm) y Grecia (1790 msnm), por último y categorizándose como las zonas con mejor estado de conservación, Parches boscosos conservados: Chuzo (2000 msnm), Selva (1931 msnm) y Nona (1864 msnm).
FIGURA 1. Mapa, localización geográfica de las 9 localidades de estudio. Tres
configuraciones de paisaje: Agroecosistema (rojo), fragmento de bosque rodeado de
matrices con sistemas productivos (amarillo) y parches boscosos conservados (azul).

19
4.2 Caracterización del ensamblaje de aves.
Los muestreos fueron realizados entre los meses de Agosto de 2016 y Octubre de 2017, se llevaron a cabo 12 salidas de campo quincenales, con duración de cinco días por cada localidad. Después de haber realizado los muestreos en todas las nueve localidades escogidas, se realizaron tres réplicas a cada una de las localidades (repartidas en distintas épocas del año).
En cada finca se estableció un punto central a partir del cual se trazó un radio de un 1km donde se llevaron a cabo los muestreos por medio de redes de niebla: dimensiones de las redes:12-10 m largo y 2-3 m de ancho; de 7 a 10 redes en promedio, teniendo en cuenta la dificultad del terreno para su instalación, de las cuales 4 redes fueron elevadas aproximadamente a 8-10 m para obtener muestras en el dosel, las redes se mantuvieron abiertas durante 12 horas (06:00-18:00), y se revisaron con intervalos de 15 a 30 minutos de acuerdo con los picos de actividad de las aves, fueron cuatro días de muestreo en red por cada sitio.
Durante los recorridos de revisión, se llevaron a cabo observaciones ad libitum, por medio de binoculares (Bushnell, 10x45) y registro fotográfico para complementar el inventario de aves obtenido en red y registros de la dieta de las aves. Las observaciones se adelantaron en puntos fijos ubicados dentro del km a la redonda del área de muestreo donde se instalaron las redes.
Cada uno de los individuos capturados fue colocado en una bolsa de tela marcada con la ubicación, # consecutivo, la hora, la fecha y la altura de caída en la red, en donde permaneció aproximadamente durante 2 horas para tratar de obtener la muestra fecal, a excepción de los colibríes y mieleros, a los cuales se les tomaban las medidas correspondientes, el registro fotográfico y se liberaban inmediatamente. Posteriormente, se realizó la determinación taxonómica a nivel de especie por medio de las guías de campo “Aves de Colombia” (Hilty & Brown 2001; McMullan et al. 2011). La nomenclatura y arreglo taxonómico se presenta de acuerdo con el comité de la lista de aves de Sur América, la South American Classification Committe (SACC) actualizada a 2018; se realizaron mediciones morfológicas estándar. Por último, se procedió a alimentar el ave cuando fue necesario y posteriormente se liberaba.
4.3 Muestras fecales
Las heces colectadas en las bolsas de tela, fueron depositadas en bolsas plásticas transparentes resellables y marcadas con la correspondiente etiqueta con los siguientes datos: #consecutivo, especie, fecha y localidad. Posteriormente, en el laboratorio del Grupo de Investigación Biología de la Conservación y Biotecnología de la (UNISARC) se realizó el proceso de limpieza y adecuación de las muestras. Por medio de estereoscopio, microscopio, pinzas de punta fina, se clasificaron las muestras por medio de los siguientes ítems: insectos (fragmentos), semillas (distinta identidad taxonómica), cáscaras, pulpa, masas, entre otros.

20
4.4 Colecta de datos
Los datos obtenidos referentes a la captura, fueron consignados en una tabla: # de la
captura (consecutivo), sitio de captura (localidad, con su respectiva georeferenciación),
fecha, hora de captura, altura a la que el individuo cayó en la red (m), especie, familia y
orden, # de foto (si hay registro), sexo, longitud de: tarso, ala, cola, pico (largo y ancho)
(mm), peso (gr), colecta (si era necesario), contenido de las heces (análisis posterior en el
laboratorio) y observaciones particulares. Se complementaron los datos con registros
fotográficos y de observación directa. La hoja de campo para la toma de datos de las aves
se describe en el Anexo 10.1.
4.5 Categorización de los gremios alimenticios
Teniendo en cuenta que la categorización de los gremios a priori puede estar sujeta a distintos criterios, la clasificación de los gremios y los intergremios (subcategorías) dadas en este trabajo se realizaron con la adaptación de la categorización dada por (Gallo & Idrobo (2004). Adicionalmente, se hizo en correspondencia con los ítems encontrados y clasificados en las heces colectadas, las interacciones registradas en observaciones directas y complementadas con información bibliográfica (Tabla1).
GREMIOS INTERGREMIOS
Nectarívoro Nectarívoro-Frugívoro-Insectívoro
Granívoro Frugívoro-Nectarívoro-Insectívoro
Carnívoro Frugívoro-Insectívoro-Carnívoro
Frugívoros Insectívoro-Frugívoro-Nectarívoro
Insectívoros Insectívoro-Frugívoro
Insectívoro-Carnívoro
Frugívoro-Insectívoro
Granívoro-Insectívoro
Nectarívoro-Insectívoro
Carroñero-Carnívoro
Omnívoro
Tabla 1. Categorías de la clasificación de la dieta de las aves (GREMIOS, INTERGREMIOS). Adaptado de Gallo & Idrobo (2004).
4.5 Análisis de datos Para evaluar la representatividad de los resultados obtenidos mediante los recorridos de
observación y las aves capturadas en red, se realizaron curvas de acumulación de
especies para los dos tipos de métodos utilizados. Como unidad de esfuerzo de muestreo
se tuvo en cuenta el número de días, los datos obtenidos fueron aleatorizados 100 veces
para promediarlos y “suavizar” la curva, se utilizaron intervalos de confianza al 95% y se
comparó la curva con los siguientes estimadores no paramétricos: Bootstrap, ICE, Chao
2, Jacknife 2, basado en (Oreja et al, 2010), por medio del software EstimateS versión
9.1.0 (Colwell, 2000).

21
Luego se realizó el análisis estadístico y la estimación de parámetros estructurales como: abundancia, riqueza, índices de diversidad: Shannon como estimador de la equidad y Simpson para la dominancia. Garmendia y Samo (2005). La fórmula del índice de Shannon es:
La fórmula del índice de Simpson es:
Para estimar la similitud en la composición de las especies entre los tres tipos de configuración del paisaje propuestas (agro ecosistema, fragmento de bosque rodeado de matrices agroforestales o monocultivos y parche de bosque conservado), se usó el índice de similitud de Jaccard el cual expresa el grado en que dos muestras son semejantes y es una medida inversa de la diversidad beta, que se refiere al cambio de especies entre las dos muestras. Se construyó un dendograma en el cual se puede interpretar de manera sencilla el grado de similitud en la composición de especies entre los tipos de configuración del paisaje. La matriz de similitud fue obtenida usando la técnica del ligamiento promedio aritmético no ponderado (.UPGMA.), que es la que produce la menor distorsión en la representación de la matriz de similitud (Sneath y Sokal 1973). Para estimar los valores del índice y el dendrograma, estos fueron construidos mediante el programa PAST (Hammer et al. 2001).
22
5. RESULTADOS
5.1 Representatividad del muestreo
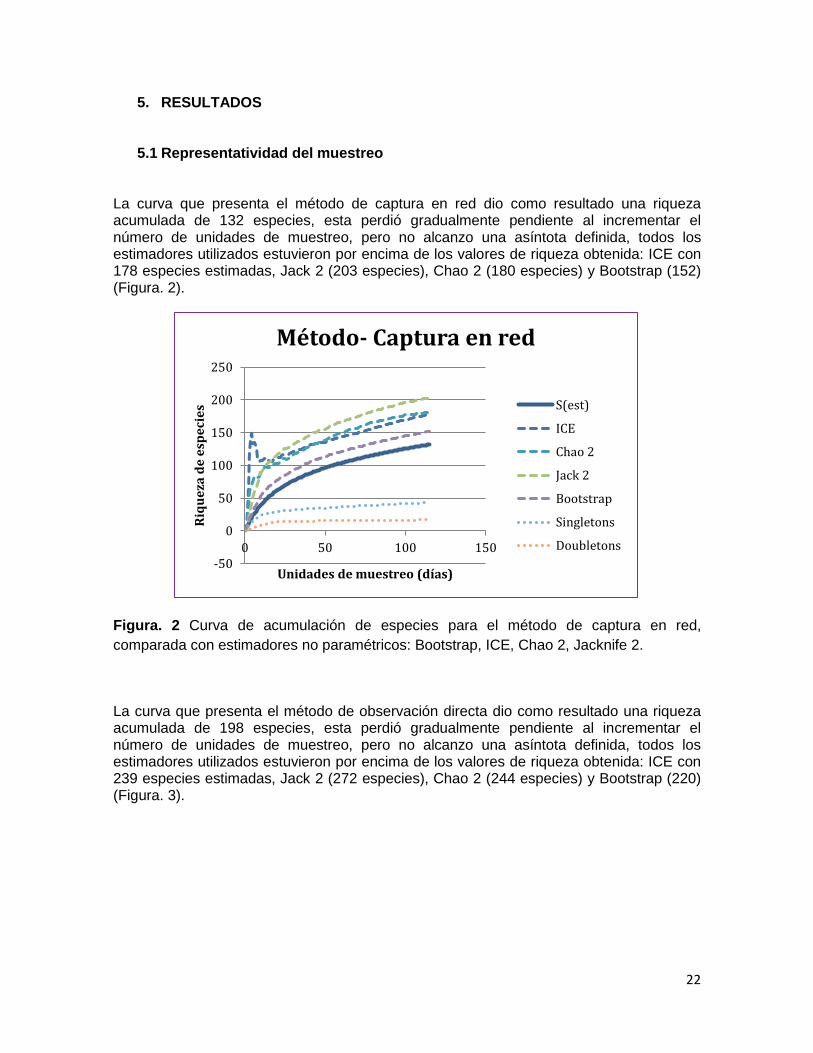
La curva que presenta el método de captura en red dio como resultado una riqueza acumulada de 132 especies, esta perdió gradualmente pendiente al incrementar el número de unidades de muestreo, pero no alcanzo una asíntota definida, todos los estimadores utilizados estuvieron por encima de los valores de riqueza obtenida: ICE con 178 especies estimadas, Jack 2 (203 especies), Chao 2 (180 especies) y Bootstrap (152) (Figura. 2).
Figura. 2 Curva de acumulación de especies para el método de captura en red,
comparada con estimadores no paramétricos: Bootstrap, ICE, Chao 2, Jacknife 2.
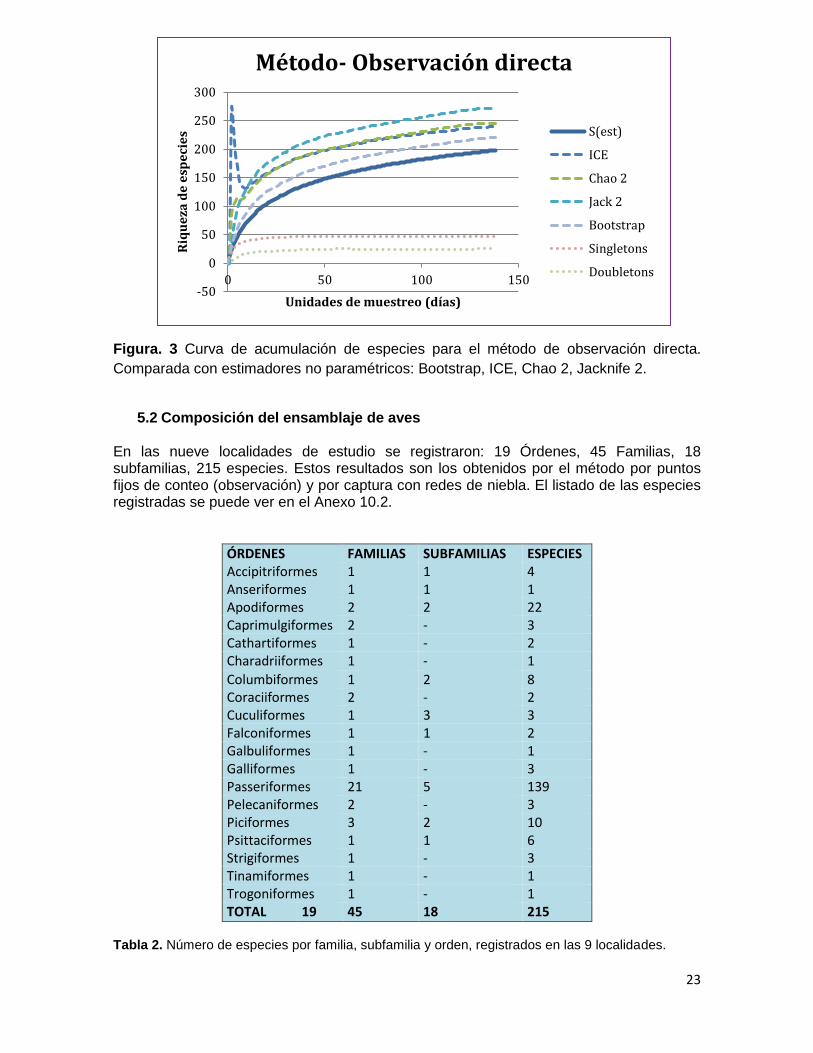
La curva que presenta el método de observación directa dio como resultado una riqueza acumulada de 198 especies, esta perdió gradualmente pendiente al incrementar el número de unidades de muestreo, pero no alcanzo una asíntota definida, todos los estimadores utilizados estuvieron por encima de los valores de riqueza obtenida: ICE con 239 especies estimadas, Jack 2 (272 especies), Chao 2 (244 especies) y Bootstrap (220) (Figura. 3).
-50
0
50
100
150
200
250
0 50 100 150
Riq
ue
za d
e e
spe
cie
s
Unidades de muestreo (días)
Método- Captura en red
S(est)
ICE
Chao 2
Jack 2
Bootstrap
Singletons
Doubletons
23
-50
0
50
100
150
200
250
300
0 50 100 150
Riq
ue
za d
e e
spe
cie
s
Unidades de muestreo (días)
Método- Observación directa
S(est)
ICE
Chao 2
Jack 2
Bootstrap
Singletons
Doubletons
Figura. 3 Curva de acumulación de especies para el método de observación directa.
Comparada con estimadores no paramétricos: Bootstrap, ICE, Chao 2, Jacknife 2.
5.2 Composición del ensamblaje de aves
En las nueve localidades de estudio se registraron: 19 Órdenes, 45 Familias, 18 subfamilias, 215 especies. Estos resultados son los obtenidos por el método por puntos fijos de conteo (observación) y por captura con redes de niebla. El listado de las especies registradas se puede ver en el Anexo 10.2.
ÓRDENES FAMILIAS SUBFAMILIAS ESPECIES Accipitriformes 1 1 4 Anseriformes 1 1 1 Apodiformes 2 2 22 Caprimulgiformes 2 - 3 Cathartiformes 1 - 2 Charadriiformes 1 - 1
Columbiformes 1 2 8 Coraciiformes 2 - 2 Cuculiformes 1 3 3 Falconiformes 1 1 2 Galbuliformes 1 - 1 Galliformes 1 - 3 Passeriformes 21 5 139 Pelecaniformes 2 - 3 Piciformes 3 2 10 Psittaciformes 1 1 6 Strigiformes 1 - 3 Tinamiformes 1 - 1 Trogoniformes 1 - 1 TOTAL 19 45 18 215
Tabla 2. Número de especies por familia, subfamilia y orden, registrados en las 9 localidades.

24
El orden más representativo fue el de los Passeriformes con 139 especies, seguido por el orden de los Apodiformes con 22 especies. La familia Thraupidae cuenta con el mayor número de especies (35), seguida por las familias Tyrannidae (27) y Trochilidae (20). El mayor número de individuos caídos en red corresponde a Tangara vitriolina, de la familia Thraupidae, seguida de Haplophaedia aureliae y Phaethornis guy de la familia Trochilidae. En cuanto al método de registros por observación directa la especie más abundante fue Tyrannus melancholicus de la familia Tyrannidae. En cuanto a la diversidad del ensamblaje evaluado para las tres configuraciones de paisaje distintas, no se encuentra una diferencia representativa entre los tres, aunque esta presenta un leve aumento en la medida en que el escenario está menos perturbado. El Agroecosistema con 131 especies (60.9 % del total de especies), Fragmento de bosque rodeado de matrices agroforestales 147 (68.3 %) y Bosque conservado 150 (69.7 %). (Anexo 10.3). Se obtuvo una totalidad de 2.754 individuos registrados tanto con el método de red de niebla como de observación directa, la abundancia para los tres escenarios se presenta de la siguiente manera: Agroecosistema con 1217 individuos (44.1 % del total de individuos), Fragmento de bosque rodeado de matrices agroforestales 740 (26.8 %) y Bosque conservado 797 (28.9 %) (Anexo 10.4). Siendo el Agroecosistema el que mayor
abundancia presenta, casi duplicando a los otros dos escenarios.
De las 215 especies registradas, 7 son endémicas para Colombia: Penelope perspicax, Ortalis columbiana, Picumnus granadensis, Cercomacroides parkeri, Dacnis hartlaubi, Chlorochrysa nitidissima y Habia cristata, según el Libro Rojo de Aves de Colombia se tienen como especies amenazadas a escala nacional; en categoría de En Peligro (EN) Penelope perspicax, Vulnerable (VU) Dacnis hartlaubi y Chlorochrysa nitidissima (Renjifo et al, 2016) (Anexo 10.5). Se registraron 16 especies migratorias, Ictinia plúmbea, transeúnte (Migratoria de Centroamérica; entre Julio-Abril), Buteo platypterus, Empidonax alnorum, Empidonax virescens, Contopus sordidulus, Myiarchus crinitus, Catharus ustulatus, Piranga rubra, Piranga olivácea, Pheucticus ludovicianus, Passerina cyanea, Leiothlypis peregrina, Mniotilta varia, Parkesia noveboracensis, Geothlypis philadelphia, Cardellina canadensis (Migratorias boreales; provenientes de Norteamérica, entre Septiembre-Abril), (Naranjo et al, 2012) (Anexo 10.6). El valor de diversidad para la avifauna de esta zona cafetera, calculado con el método de
Shannon-Wiener es: Agroecosistema 4,27, Fragmento de bosque 4,58 y Bosque 4,48 lo
que indica una diversidad alta de acuerdo con la interpretación del índice propuesta por.
Según el índice de diversidad Simpson el valor obtenido es de: Agroecosistema 0,9798,
Fragmento de bosque 0,9867 y Bosque 0,9838, estos valores altos indican que una o
algunas especies son dominantes en la comunidad (Magurran,1988, Moreno 2001).
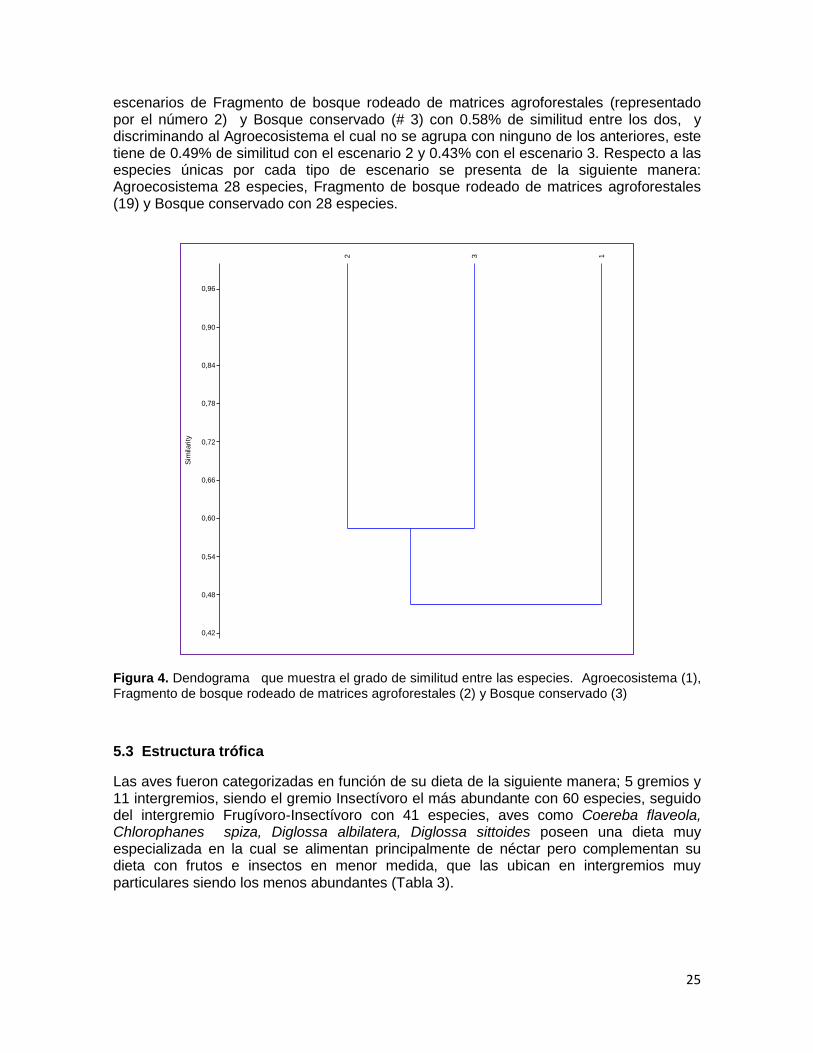
La estimación de la similitud en composición de especies entre los tres tipos de configuración del paisaje propuestas en este trabajo (agro ecosistema, fragmento de bosque rodeado de matrices agroforestales o monocultivos y parche de bosque conservado), se puede analizar a partir del dendograma obtenido en el Análisis de Agrupamiento (Fig. 2), el cual sintetiza las relaciones multi-dimensionales entre las especies en función de la configuración del paisaje, mostrando similitud entre los
25
escenarios de Fragmento de bosque rodeado de matrices agroforestales (representado por el número 2) y Bosque conservado (# 3) con 0.58% de similitud entre los dos, y discriminando al Agroecosistema el cual no se agrupa con ninguno de los anteriores, este tiene de 0.49% de similitud con el escenario 2 y 0.43% con el escenario 3. Respecto a las especies únicas por cada tipo de escenario se presenta de la siguiente manera: Agroecosistema 28 especies, Fragmento de bosque rodeado de matrices agroforestales (19) y Bosque conservado con 28 especies.
Figura 4. Dendograma que muestra el grado de similitud entre las especies. Agroecosistema (1), Fragmento de bosque rodeado de matrices agroforestales (2) y Bosque conservado (3)
5.3 Estructura trófica
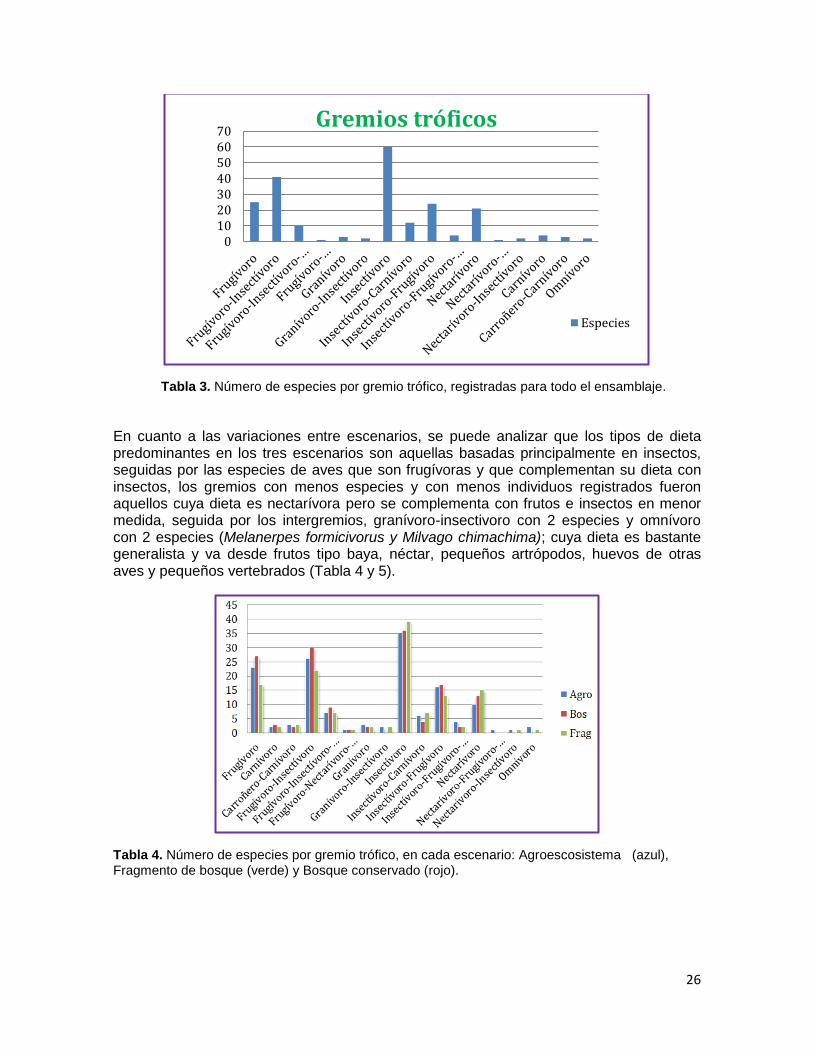
Las aves fueron categorizadas en función de su dieta de la siguiente manera; 5 gremios y 11 intergremios, siendo el gremio Insectívoro el más abundante con 60 especies, seguido del intergremio Frugívoro-Insectívoro con 41 especies, aves como Coereba flaveola, Chlorophanes spiza, Diglossa albilatera, Diglossa sittoides poseen una dieta muy especializada en la cual se alimentan principalmente de néctar pero complementan su dieta con frutos e insectos en menor medida, que las ubican en intergremios muy particulares siendo los menos abundantes (Tabla 3).
0,42
0,48
0,54
0,60
0,66
0,72
0,78
0,84
0,90
0,96
Sim
ilarity
2 3 1
26
Tabla 3. Número de especies por gremio trófico, registradas para todo el ensamblaje.
En cuanto a las variaciones entre escenarios, se puede analizar que los tipos de dieta predominantes en los tres escenarios son aquellas basadas principalmente en insectos, seguidas por las especies de aves que son frugívoras y que complementan su dieta con insectos, los gremios con menos especies y con menos individuos registrados fueron aquellos cuya dieta es nectarívora pero se complementa con frutos e insectos en menor medida, seguida por los intergremios, granívoro-insectivoro con 2 especies y omnívoro con 2 especies (Melanerpes formicivorus y Milvago chimachima); cuya dieta es bastante generalista y va desde frutos tipo baya, néctar, pequeños artrópodos, huevos de otras aves y pequeños vertebrados (Tabla 4 y 5).
Tabla 4. Número de especies por gremio trófico, en cada escenario: Agroescosistema (azul), Fragmento de bosque (verde) y Bosque conservado (rojo).
010203040506070
Gremios tróficos
Especies

27
Tabla 5. Número de individuos por gremio trófico, en cada escenario: Agroescosistema (azul), Fragmento de bosque (verde) y Bosque conservado (rojo).
6 DISCUSIÓN DE RESULTADOS
6.1 Representatividad del muestreo La totalidad de especies registradas con los dos métodos fue de 215 especies, de las cuales 132 fueron registradas por medio de captura con red de niebla y 198 por medio de observación directa. En cuanto a la representatividad de muestreo por tipo de metodología usada, el método de observación directa fue más eficiente ya que tuvo una riqueza acumulada más alta, esto puede deberse a que en escenarios con doseles de bosque altos las redes de niebla son casi ineficientes, a pesar de que fueron elevadas, es difícil capturar aves a estas alturas. Las curvas en general no alcanzaron una asíntota definida, de tal forma que se necesitarían más unidades de muestreo para complementar los inventarios. A pesar de que las curvas y los estimadores propuestos no fueron asintóticos, analizando las curvas que resultan a partir de los datos de los singletons y doubletons si se obtiene una asíntota, cuando estas son asintóticas o tienden a descender indican que se ha logrado un buen muestreo como lo sugiere (Villarreal, et al, 2006). El estimador que más se ajusta y se acerca a los valores de riqueza obtenidos en este trabajo, para los dos tipos de metodología fue Bootstrap que estima la riqueza de especies a partir de la proporción de muestras que contienen a cada especie, respecto a este se tuvo una representatividad con porcentajes de 90% para el método de observación directa y 86% para el método por captura en red. El estimador que menos se acercó fue Jack 2 que considera a las especies presentes en dos unidades de muestreo tuvo porcentajes de 72% para el método de observación directa y 65% para el método por captura en red. De igual forma a menos que la comunidad se haya muestreado de modo

28
exhaustivo, estas curvas subestimarán también la riqueza real de un modo desconocido (Magurran, 2004).
6.2 Composición del ensamblaje de aves
Los escenarios categorizados como agroecosistemas a pesar de considerarse zonas afectadas por las transformaciones antropogenicas, con un nivel de deforestación alto, áreas boscosas casi inexistentes y monocultivos de amplia extensión presentaron una alta diversidad tanto en número de especies como en número de individuos allí registrados, inclusive fue el escenario que tuvo mayor abundancia de aves tanto capturadas en red como observadas. De acuerdo al grado de complejidad estructural de los sistemas productivos (monocultivos, sistemas agroforestales, sistemas silvopastoriles, etc.) éstos pueden albergar una mayor diversidad y cumplir una función como corredores biológicos y como sitios para el mantenimiento de servicios ambientales (Weibull et al. 2003, CARDER 2004). El papel que puede jugar este tipo de paisajes agrícolas transformados como parte del mosaico de ecosistemas y como hábitat para distintas especies animales es complejo. Varios estudios han demostrado que muchas especies de aves pueden utilizar los recursos disponibles en las plantaciones y otros hábitats antropogénicos (Durán & Kattan 2005, Hayes & Samad 1998, Mitra & Sheldon 1993, Petit & Petit 2003, Renjifo 2001).
Algunos estudios realizados en cafetales y plantaciones forestales han encontrado que estos hábitats son más pobres cuando se comparan con los bosques naturales, los cuales tienen más especialistas de interior de bosque, más aves endémicas y mayor abundancia de insectívoros, mientras que las plantaciones presentan mayor abundancia de aves de áreas abiertas (Wunderle & Latta 1996). Sin embargo, las plantaciones pueden proveer hábitat para especies de bosque en regiones sometidas a deforestación extrema (Brash 1987, Ewel & Putz 2004, Haggar et al. 1997, Hayes & Samad 1998, Perfecto et al. 1996).
Por ejemplo, especies generalistas y de vulnerabilidades bajas, especies adaptadas o adaptables a las zonas abiertas y especies para las cuales sus recursos disponibles han aumentado o los enemigos o competidores naturales han disminuido, son especies cuyas poblaciones y distribución han aumentado. Algunas especies de loras, tórtolas, aves granívoras y frugívoras y los gallinazos, se podría decir que se han convertido en especies dominantes en el agroecosistema cafetero, debido a su alta abundancia y amplia distribución como lo asevera Botero & Lentijo (2014)
Sin embargo, este tipo de ecosistemas no brindaría los recursos adecuados para toda la avifauna local, teniendo en cuenta que especies raras de interior de bosque, que son especialistas con dietas muy específicas, y de zonas más conservadas que no se registraron en el agroecosistema se verían directamente afectadas si los ecosistemas menos intervenidos sufrieran algún tipo de transformación. Como también lo sugiere Botero & Lentijo (2014) respecto a la biodiversidad en zonas cafeteras en Colombia, las especies más vulnerables a los cambios en sus hábitats, las especialistas, las de mayores tamaños y las que requieren mayores extensiones boscosas para existir, han sido especialmente afectadas. También lo han sido aquellas susceptibles a la cacería y tráfico de fauna y flora, y aquellas susceptibles a la contaminación.

29
6.3 Estructura trófica
De las 16 variables de alimentación dadas en este trabajo para categorizar las dietas de las aves, el gremio insectívoro es el más representativo con el 27.9 % del total de las especies del ensamblaje, siendo más abundante en los escenarios de Fragmento de bosque y bosque conservado que en los categorizados como agroecosistemas, aves insectívoras que se encuentran asociadas en mayor medida a interior de bosque y fragmentos boscosos de grandes extensiones, especialistas pertenecientes a las familias Tyrannidae Picidae y Furnariidae, la predominancia de este tipo de dieta asociada a este tipo de hábitats concuerda con lo reportado en otros estudios realizados en la Cordillera Central de los andes colombianos (Gallo & Idrobo, 2004), (Castaño & Patiño, 2007). El intergremio Frugívoro-Insectívoro fue la siguiente categoría más abundante con el
19.06 % del total de las especies, el gremio de los frugívoros estrictos fue el siguiente con
el 11.6 % de especies, de este tipo de aves cuya alimentación se basa exclusivamente en
frutos; se registraron aproximadamente 127 morfotipos de plantas consumidas, de los
cuales se lograron identificar 90 especies pertenecientes a 37 familias en donde las
familias más representativas fueron Melastomataceae con 9 especies, Loranthaceae (7),
Urticaceae, Solanaceae y Poaceae (6), y Rubiaceae (4). Los géneros que fueron más
consumidos por las aves fueron Miconia y Cecropia (determinaciones realizadas por
Catalina López Rico y Dayro Leandro Rodríguez, UNISARC).
Las anteriores categorías de alimentación, en cuanto a número de especies por escenario
fueron registradas mayormente en bosque conservado, seguido por agroecosistema y
finalmente fragmento de bosque, especies pertenecientes mayormente a las familias
Thraupidae, Tyrannidae y Emberizidae. En cuanto a la abundancia la categoría de dieta
con el mayor número de registros fue la de frugívoro-insectívoro con 394 individuos en el
escenario de agroecosistema. Esto puede deberse a que las aves registradas en
agroecosistemas los cuales se conforman ampliamente de cafetales, otros cultivos
agrícolas y plantaciones forestales en extensión, no sean residentes teniendo en cuenta
que este tipo de ecosistemas transformados sirven como corredores y conectores que
permiten el paso de especies entre los bosques, y bordes de bosque en donde se da una
amplia variedad especies vegetales pioneras que ofrecen de manera constante frutos
(Castaño & Patiño, 2007).
En cuanto al gremio de los nectarívoros este se vio representado con el 9.7 % de las especies del ensamblaje total, cuya abundancia es mayor en el fragmento de bosque, seguida por el bosque conservado y en menor medida registrados en los agroecosistemas, lo que puede darse al presentarse una mayor disponibilidad de alimento representada en flores que están muy dispersas en el bosque y que producen volúmenes grandes de néctar concentrado según Murcia (2002), de igual forma también afirma que para los colibríes es más importante los recursos alimenticios que el elemento del paisaje en el cual estos se hallen a lo cual debe deberse que la abundancia en los tres tipos de configuración de paisaje no muestre diferencias muy representativas. Las categorías de alimentación con el menor número de especies son: carroñero-
carnívoro con 3 especies, granívoro (3), granívoro-insectívoro (2), y aquellas aves que

30
son nectarívoras y complementan su dieta con otro tipo de recursos; aves que poseen
dietas muy especializadas que las ubican en los gremios menos abundantes, especies
como Coereba flaveola, Chlorophanes spiza, Diglossa albilatera, Diglossa sittoides, que
se alimentan principalmente de néctar pero complementan su dieta con frutos e insectos
en menor medida.
En cuanto a la metodología utilizada para el análisis de la dieta las muestras de heces constituyen un método efectivo y no invasivo, a diferencia del análisis de contenidos estomacales y el uso de eméticos para colectar información sobre dieta de aves capturadas en redes de niebla (Wheelwright et al. 1984, Loiselle y Blake 1990). Los sesgos relacionados con la velocidad del paso por el tracto digestivo y con la digestión diferencial de los diferentes componentes de la dieta son algunas de las desventajas de este método (Rosenberg y Cooper 1990). Sin embargo, se ha demostrado una buena correspondencia entre muestras de heces y contenidos estomacales y no se encontró que hubiera un sesgo contra ítems pequeños o de cuerpo blando (Ralph et al. 1985)
Trabajos de investigación como este aportan a la información acerca de: la biodiversidad de aves presente en estas zonas del eje cafetero, particularidades de su ecología como la variedad de dietas que presentan los conjuntos locales de aves, como características particulares que conforman los “elementos del paisaje” en este tipo de ecosistemas transformados influyen directamente en la conformación de la avifauna en el departamento de Risaralda. De tal forma que se pueda comprender el impacto social, ambiental y económico que tiene vincular estudios de diversidad a la promoción del turismo como eje para la conservación de áreas protegidas, de fragmentos o remanentes de bosque como principal refugio para la biodiversidad original de las zonas cafeteras los cuales deben ser protegidos y restaurados urgentemente. Como lo presentan Botero & Lentijo (2014), los resultados de los estudios ornitológicos realizados por Cenicafé ilustran la importancia de los fragmentos de bosque para la biodiversidad original. De 390 especies de aves que han sido observadas en fragmentos de bosque, cerca del 28% se registró solamente en bosques y en ningún otro tipo de hábitat. En este gran grupo se encuentra el 91% de todas las especies de aves amenazadas, el 85% de las endémicas y el 84% de las migratorias , además en los bosques se ha encontrado el mayor número de especies de aves con vulnerabilidades alta y media, el mayor número de plantas nativas y de especies de importancia para la conservación.

31
7. CONCLUSIONES
La variabilidad en la estructuración del ensamblaje presente entre los tres tipos de
configuración de paisaje propuestos en este trabajo no es muy representativa, aun así se
observa que en la medida en que el escenario se clasifica como menos perturbado el
número de especies aumenta levemente.
A pesar de que la riqueza es mayor en zonas más conservadas, el agroecosistema es el
escenario con la mayor abundancia, casi duplicando en número de individuos a los otros
dos, además de haber registrado especies únicas que fueron observadas y capturadas
exclusivamente en este escenario (28 especies) que se compara con el mismo número de
especies únicas al escenario de bosque conservado, a pesar de considerarse
ecosistemas afectados por actividades antrópicas cumplen funciones como corredores
biológicos y como sitios con una gran diversidad de aves en el paisaje cultural cafetero.
Casi todas las categorías de gremios y las intergremiales estuvieron representadas en los
tres escenarios, para agroecosistemas estuvieron todas las 16, Fragmento de bosque
rodeado de matrices agroforestales solo un intergremio no estuvo presente y para Bosque
conservado 2 intergremios no estuvieron presentes; puesto que especies propias de
interior de bosque son un poco más especialistas, de este modo en la medida en que
estos ecosistemas estén más intervenidos especies endémicas y otras allí presentes se
ven directamente afectadas en la disposición de recursos alimenticios inmediatos.
El ensamblaje de aves en esta zona cafetera del oriente del departamento de Risaralda se caracteriza por una dominancia del gremio insectívoro, en especial hacia las zonas donde se han conservado áreas de bosque por la disponibilidad de insectos allí presente; en menor proporción integran la comunidad especies del gremio omnívoro, nectarivoro-insectivoro-frugivoro, granívoro-insectívoro, debido a la expansión de la frontera agrícola y de la explotación del suelo a partir de distintas actividades humanas, de igual forma los distintos tipos de dietas presentes cumplen en conjunto un papel fundamental en el mantenimiento del flujo de energía en los ecosistemas del paisaje rural cafetero. La transformación de los sistemas naturales, la intensidad y el modelo del cultivo del café
y la extensión de los monocultivos sugieren una amenaza para la diversidad de aves que
habitan estas zonas boscosas. Vincular trabajos de investigación con la participación de
las comunidades en el seguimiento y conservación de diversidad de aves en esta zona
del eje cafetero es prioritario, para integrar la avifauna como un componente importante
en la conservación de áreas protegidas y el fomento del aviturismo en la región.

32
8. RECOMENDACIONES
A pesar de que las muestras de heces constituyen un método efectivo y no invasivo (a
diferencia del análisis de dieta con fijación de estómago y el uso de eméticos) para
colectar información sobre dieta de aves capturadas en redes de niebla, como ya se
referencio anteriormente en otros trabajos, y que los items encontrados fueron eficientes
para su análisis y para los objetivos de este trabajo, para estudios más detallados acerca
de dieta como determinación taxonómica de semillas o artropofauna, peso específico de
cada ítem, estudios palinológicos, se recomienda complementar con otro tipo de métodos
como regurgitación, uso de eméticos, colecta de muestras de polen, combinación de las
categorías gremiales con estrategias y maniobras de obtención del alimento por parte de
las aves, entre otros.
En el territorio colombiano, está la región conocida como el triángulo del café, donde se encuentra el Departamento de Risaralda el cual se ha propuesto como “Bosque Modelo para el Mundo”, por tal motivo es un área representativa del paisaje cultural cafetero y del país, en la cual es importante el desarrollo de investigaciones en avifauna, ecología del paisaje, por lo que se debe tomar acciones de protección que garanticen su permanente conservación. Algunas de las localidades de estudio de este trabajo se encuentran cercanas a la cuenca del río Otún en los municipios de Santa Rosa de Cabal y Dosquebradas, también la microcuenca de La Nona en el municipio de Marsella que limita con el departamento de Caldas, los cuales son lugares de especial protección ya que hacen parte actualmente de zonas conservadas en el departamento.

33
9. REFERENCIAS BIBLIOGRAFICAS Amat J.A. 1984. Las poblaciones de aves acuáticas en las lagunas Andaluzas: composición y diversidad durante un ciclo anual. Ardeola 31: 61-79 pp. Andraca Collazo L. 2010. Reglas de ensamblaje y modelos de coexistencia de especies en las comunidades de aves cubanas. Tesis Biólogo. Facultad de Biología. Universidad de la Habana. Cuba. 64 p. ANDRADE, G. I. 1993. Paisaje y biodiversidad en las selvas de los Andes. Págs. 31-44 en: G. I. Andrade (ed.). Carpanta: selva nublada y páramo. Fundación Natura Colombia, The Nature Conservancy y Empresa de Acueducto y Alcantarillado de Bogotá, Santafé de Bogotá. Andrén, H. 1994. Effects of habitat fragmentation on birds and mammals in landscapes with different proportions of suitable habitat: a review. Oikos 71: 355-366. BORRERO, J.I. 1986. La sustitución de cafetales de sombrío por caturrales y su efecto negativo sobre la fauna de vertebrados. Caldasia, 15: 71-75. BOTERO, J. E., N. G. FRANCO-ROJAS, R. ESPINOSA-ALDANA Y A. M. LÓPEZ-LÓPEZ. 2012. Avifauna de la Reserva de Planalto. Revista Cenicafé 63(1): 41- 56. BOTERO, J.E., G.M. LENTIJO, L.M. SÁNCHEZ-CLAVIJO. 2014. Biodiversidad en zonas
cafeteras de Colombia. Principales lecciones. Avances Técnicos CENICAFE 444. 12p. Bradley RA & Bradley DW (1985) Do non-random patterns of species in niche space imply competition? Oikos 45:443- 446. Camargo, J.C.; Current, D.; Ibrahim, M.; Somarriba, E.; Finegan, B. 2000. Factores ecológicos y socioeconómicos que influyen en la regeneración natural del laurel en sistemas silvopastoriles del trópico húmedo y subhúmedo de Costa Rica. Agroforestería en las Américas 7(26): 46-49.
CARDER. 2004: Ecorregión eje cafetero: un territorio de oportunidades. Proyecto: Construcción de un ordenamiento territorial para el desarrollo sostenible en la Ecorregión del Eje Cafetero. Convenio CARDER-FONADE No 1068.
Cardona, W., Cano, T., Gil, R., & Gómez, D. (2012). CARACTERIZACIÓN DE FAUNA (RANAS Y AVES) Y FLORA EN SEIS HUMEDALES DEL DEPARTAMENTO DE RISARALDA.
Castaño Villa G.J y J.C. Patiño Zabala. 2007. Composición de la comunidad de aves en bosques fragmentados en la región de Santa Elena, andes centrales colombianos. Museo de Historia Natural. 11: 47 – 60 pp.
CAVELIER, J. 1997. Selvas y bosques montanos. Págs 38-55 en: M. E. Chaves & N. Arango (eds.). Informe nacional sobre el estado de la biodiversidad, Tomo I. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá

34
Cody M.L. (1985). Habitat selection in birds. Academic Press, Nueva York.
Collins S. L., Carpenter S. R., Swinton S. M., Orenstein D. E., Childers D. L., Gragson T. L., Grimm N. B., Grove J. M., Harlan S. L., Kaye J. P., Knapp A. K., Kofinas G. P., Magnuson J. J., McDowell W. H., Melack J. M., Ogden L. A., Robertson G. P., Smith M. D., and Whitmer A. C.,. (2010) “An integrated conceptual framework for long-term social–ecological research,” Front. Ecol. Environ., vol. 9, no. 6, pp. 351–357, Nov. 2010.
Colwell, R.K. 2000. EstimateS, Statistical Estimation of Species Richness and Shared.
Species from Samples. University of Connecticut.
Corpocaldas, A. C., & Fierro-Calderón, E. (2010). Estado de conocimiento de las aves en el departamento de Caldas: prioridades de conservación y vacíos de información.Corporación Autónoma Regional de Caldas, Manizales.
Duque Escobar, G. (2014). Gobernanza forestal para la ecorregión andina. Revista Civismo SMP Manizales.
Espinosa, T. E. (2003). ¿Cuántas especies hay. Los estimadores no paramétricos de Chao. Elementos, 52, 53-56.
Fahrig, L. 2003. Effects of habitat fragmentation on biodiversity. Annual Review of Ecology, Evolution and Systematics 34: 487–515.
Farina, A. (2008). Principles and Methods in Landscape Ecology: Towards a Science of the Landscape.
Gallo-Cajiao, E., & Idrobo-Medina, C. (2004). Fragmentos de bosque y conservación de aves: un estudio de caso en los Andes de Colombia. Memorias: Manejo de Fauna silvestre en Amazonia y Latinoamérica, 178-185.
Garmendia Salvador A. y A.J. Samo Lumbreras. 2005. Prácticas de ecología. Universidad Politécnica de Valencia. ES. 147 p. Gitay H & Noble IR (1997) What are functional types and how should we seek them? Pp. 3-19 en: Plant functional types. Their relevance to ecosystem properties and global change. Smith TM, Shugart HH y Woodward FI (eds). Cambridge University Press, Cambridge.
Gómez, E. L. S., & García, J. C. C. (2012). Diversidad de avifauna en paisajes rurales de la cuenca del río La Vieja, Eje Cafetero de Colombia. Recursos Naturales y Ambiente, (65-66).
Hammer, O., D. Harper y P. Ryan. 2001. PAST: Paleontological Statistics Software. Package for Education and Data Analysis. Paleontología Electrónica 4(1): 9.
Hawkins CP & MacMahon JA (1989) Guilds: the multiple meanings of a concept. Annual Review of Entomology 34:423-451

35
Henao-Isaza, J. R., Carranza, J. A., & Castaño, J. H. (2014) AVIFAUNA DEL CAMPUS UNIVERSITARIO EL JAZMIN: UN MOSAICO DE AGROECOSISTEMAS DIVERSOS EN EL PAISAJE CAFETERO DE RISARALDA. Boletín Invest Unisarc. 11(1-2): 42 – 54. 2014
Herzog, S. K., & Kattan, G. H. (2011). Patterns of diversity and endemism in the birds of the tropical Andes. Climate change and biodiversity in the tropical Andes. Montevideo, Inter-American Institute for Global Change Research (IAI) and Scientific Committee on Problems of the Environment (SCOPE), 245-259.
Hill, M. O. 1973. Diversity and evenness: a unifying notation and its consequences. Ecology 54:427–432.
HILTY, S.H. y BROWN, W.L. 2001. Guía de las Aves de Colombia. American Bird Conservancy. Colombia.
Holdridge, L. R. (1987). Ecología basada en zonas de vida (No. 83). Agroamérica.
Holmes RT (1980) Resource exploitation patterns and the structure of a forest bird community. Pp. 1056-1062 en: Actis XVII Congressus Internationalis Ornithologici. Nohring R (ed). Deutsche Ornithologen.Gesellschaft, Berlin
Holmes RT & Recher HF (1986) Determinants of guild structure in forest bird communities: an intercontinental comparison. Condor 88:427-439. Hurtado Veintimilla, N. I. (2015). Estructura trófica de una comunidad de aves y uso de especies paraguas en la conservación del bosque seco del cantón Zapotillo. Tesis de grado Ingenieria en Manejo y Conservación de Medio Ambiente. (Bachelor's thesis, Loja: Universidad Nacional de Loja). pp 5-6. Ibáñez Martí J.J. y A. García Álvarez. 2002. Diversidad: biodiversidad edáfica y geodiversidad. Edafología 9(3): 329-385 pp. Jaksic FM (1981) Abuse and misuse of the term guild in ecological studies. Oikos 37:397-400. KATTAN, G. H. & H. ALVAREZ-LÓPEZ. 1996. Preservation and management of biodiversity in fragmented landscapes in the Colombian Andes. Págs. 3-18 en: J. Schelhas & R. Greenberg (eds.). Forest remnants in the tropical landscape. Island Press, Washington, D. C. KATTAN, G. H. 1997. Transformación del paisaje y fragmentación del hábitat: ecosistemas terrestres. Págs. 76-87 en: M. E. Chaves & N. Arango (eds.). Informe nacional sobre el estado de la biodiversidad, Tomo II. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá.
Kattan, G., Valderrama, C., Trujillo, A.F., Rojas-Díaz, V. 2004: Análisis de representatividad en el SIRAP EC. En: Análisis de representatividad y biodiversidad para la construcción del sistema regional de áreas protegidas del eje cafetero. Fundación EcoAndina, programa Colombia de Wildlife Conservation Society. World Wildlife Fund-Colombia.

36
Landres PB & MacMahon JA (1983) Community organization of arboreal birds in some oak woodlands of western North America. Ecological Monographs 53:183-208
Lentijo, G. M., & Kattan, G. H. (2005). Estratificación vertical de las aves en una plantación mono específica y en bosque nativo en la cordillera central de Colombia. Ornitología Colombiana, 3, 51-61.
Lentijo, G. M., Arbeláez, D., Castellanos, O., Franco, N. G., López, A. M., & Botero, J. E. (2008). Enfoques participativos en investigación como una herramienta de conservación de las aves en zonas cafeteras de Colombia. Ornitologıa Neotropical, 19, 567-574.
Loh, J. y Wackernagel, M. (eds.). 2004. Informe Planeta Vivo 2004. WWF, Gland.
Londoño-Betancourth, J. C. (2011). Una mirada a la diversidad ornitológica de Pereira. Bol Cient Mus Hist Nat, 15, 84-103.
Lopez de Casenave, J. N. (2001). Estructura gremial y organización de un ensamble de aves del desierto del Monte (Tesis Doctoral, Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires). Biblioteca digital FCEN-UBA. www.digital.bl.fcen.uba.ar, pp 15.
López-Ordóñez P. J., Cortés-Herrera, O. J., Paez-Ortíz, A. C., & González-Rojas, F. M. (2013). Nuevos registros y comentarios sobre la distribución de algunas especies de aves en los Andes Occidentales de Colombia.Ornitología Colombiana, (13).
Magurran, A. (1988). Ecological diversity and its measurement. Princeton University
Press, New Jersey, 179 pp.
Magurran, A. E., 2004. Measuring Biological Diversity. Blackwell, Malden
MCMULLAN, M.; QUEVEDO, A. y DONEGAN, T.M. 2011. Guía de campo de las Aves de Colombia. Fundación ProAves. Colombia.
Moreno, C. E. 2001. Manual de métodos para medir la diversidad. Universidad Veracruzana. Xalapa, México. 49p.
Morrison ML, Marcot BG & Mannan RW (1992) Wildlife.habitat relationships. Concepts and applications. University of Wisconsin Press, Madison.
MURCIA, C., 2002. - Frugivoría: 531-558 (en) GUARIGUATA, M.R. & KATTAN, G.H. (eds). Ecología y Conservación de bosques. Ediciones LUR, Cartago, Costa Rica.
Nadachowski Chávarro, E., & Valencia Valencia, M. Y. (2009). Sistema regional de áreas naturales protegidas en el Eje Cafetero de Colombia: un esfuerzo colectivo para la conservación de nuestro territorio.
Naranjo, L. G., J. D. Amaya, D. Eusse-González y Y. Cifuentes-Sarmiento (Editores). 2012. Guía de las Especies Migratorias de la Biodiversidad en Colombia. Aves. Vol. 1. Ministerio de Ambiente y Desarrollo Sostenible / WWF Colombia. Bogotá, D.C. Colombia. 708 p.

37
Oreja, J. G., de la Fuente-Díaz-Ordaz, A. A., Hernández-Santín, L., Buzo-Franco, D., & Bonache-Regidor, C. (2010). Evaluación de estimadores no paramétricos de la riqueza de especies. Un ejemplo con aves en áreas verdes de la ciudad de Puebla, México. Animal biodiversity and conservation, 33(1), 31-45.
Otero, J., Quiceno, M.P., Vieira, M.I., Cárdenas, K., Cabrera, E. 2005: Las Cercas Vivas: Oportunidades de uso y conservación de biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt. Bogotá, D.C.
Perovic P., C. Trucco, A. Tálamo, V. Quiroga, D. Ramallo, A. Lacci, A. Baungardner y F. Mohr. 2008. Guía técnica para el monitoreo de la biodiversidad. Programa de Monitoreo de Biodiversidad, Parque Nacional Copo, Parque y Reserva Provincial Copo, y Zona de amortiguamiento. APN/GEF/BIRF. Salta, Argentina.
Philptt, S. M. ,Arendt W. J., I. Armnbretch I., Bichier P.,Diestch T. V,Gorgon C., R. Greenberg R., Perfecto I., R. Reynoso-Santos, Soto-Pinto L., Tejeda-Cruz C., Williams-Linera G., Valenzuela J., and Zolotoff J. M.,. (2008). “Biodiversity Loss in Latin American Coffee Landscapes: Review of the Evidence on Ants, Birds, and Trees,” Conserv. Biol., vol. 22, no. 5, pp. 1093–1105, 2008.
Renjifo, L. M., A. M. Franco-Maya, J. D. Amaya-Espinel, G. H. Kattan y B. López-Lanús (eds). 2002. Libro rojo de aves de Colombia. Serie Libros Rojos de Especies Amenazadas de Colombia. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt y Ministerio del Medio Ambiente. Bogotá, Colombia.
Renjifo L.M., Amaya-Villarreal, A.M., Velásquez-Tibatá, Burbano-Girón J. (2016). Libro rojo de aves de Colombia. Volumen II: Ecosistemas abiertos, secos, insulares, acuáticos continentales, marinos, tierras altas del Darién y Sierra Nevada de Santa Marta y bosques húmedos del centro, norte y oriente del país. Editorial Pontificia Universidad Javeriana e Instituto Alexander von Humboldt. Bogotá, D.C. Colombia. (v. 2).
Restrepo, D. U. (2016). Algunas de las aves emblemáticas del Eje Cafetero. Boletín Cultural y Bibliográfico, 50(91), 4-20.
Ricklefs R.E. 1996. Invitación a la ecología: la economía de la naturaleza. Trad. DS Klajn. 4 ed. Madrid, ES. Panamericana. 692 p.
Root RB (1967) The niche exploitation pattern of the Blue. gray Gnatcatcher. Ecological Monographs 37:317-350. Sáenz, J. C., Villatoro, F., Ibrahim, M., Fajardo, D., & Pérez, M. (2013). Relación entre las comunidades de aves y la vegetación en agropaisajes dominados por la ganadería en Costa Rica, Nicaragua y Colombia. Santos, T., & Tellería, J. L. (2006). Pérdida y fragmentación del hábitat: efecto sobre la conservación de las especies. Revista Ecosistemas, 15(2).

38
Saunders, D.A., Hobbs, R.J. y Margules, C.R.. 1991. Biological consequences of ecosystem fragmentation: A review. Conservation Biology 5: 18-32 Simberloff D & Dayan T (1991) The guild concept and the structure of ecological communities. Annual Review of Ecology and Systematics 22:115-143
SISTEMA DE INFORMACIÓN REGIONAL. Cobertura y usos del suelo. Municipio de Santa Rosa de Cabal. 2006.
Smith, E. P., & van Belle, G. (1984). Nonparametric estimation of species richness. Biometrics, 119-129.
Sneath PHA y Sokal RR (1973) Numerical taxonomy. The principle and practice of numerical classification. W. H. Freeman & Co., San Francisco.
Sosa, N. (2003). Las aves: riqueza, diversidad y patrones de distribución espacial. Las ENSEÑANZAS DE San Juan, 257. South American Classification Committee (SACC). 2018. A classification of the bird species of South America. Version (en línea). Southwood, T. R. E., & Henderson, P. A. (2000). Ecological Methods–Blackwell Science. SUÁREZ, C., TRUJILLO, A., GIRALDO, J., RÍOS-FRANCO, C., CUADROS, L., DORADO, J., & NARANJO, L. (2011). Clasificación de Ecosistemas Naturales Terrestres del Eje Cafetero (en Preparación). Santiago de Cali, Colombia: WWF, Sirap EC, WCS, CARDER, CVC, CRQ, CORPOCALDAS, CORTOLIMA. Villarreal, H., Álvarez, M., Córdoba, S., Escobar, F., Fagua, G., Gast, F. & Umaña, A. M. (2006). Métodos para el análisis de datos: una aplicación para resultados provenientes de caracterizaciones de biodiversidad. Manual de Métodos Para el Desarrollo de Inventarios de Biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt, Bogotá, Colombia, 185-226. Weibull, A.C., Östnann, Ö., Granqvist, A. 2003: Species richness in agroecosystems: the
effect of landscape, habitat and farm management. Biodiversity and Conservation 12,
1335-1355.
Wilson J.B. 1989. A null model of guild proportionality, applied to stratification of New Zealand temperate rain forest. Oecología 80: 263-67 pp. Wiens JA (1989) The ecology of bird communities. Volume 1. Foundations and patterns. Cambridge University Press, Cambridge WWF-Colombia, Wildlife Conservation Society, The Nature Conservancy. SIRAP Eje
Cafetero, (2013). Clasificación de Ecosistemas Naturales Terrestres del Eje Cafetero
Análisis de Representatividad del Sistema Regional de Áreas Protegidas. Santiago de
Cali, Colombia.
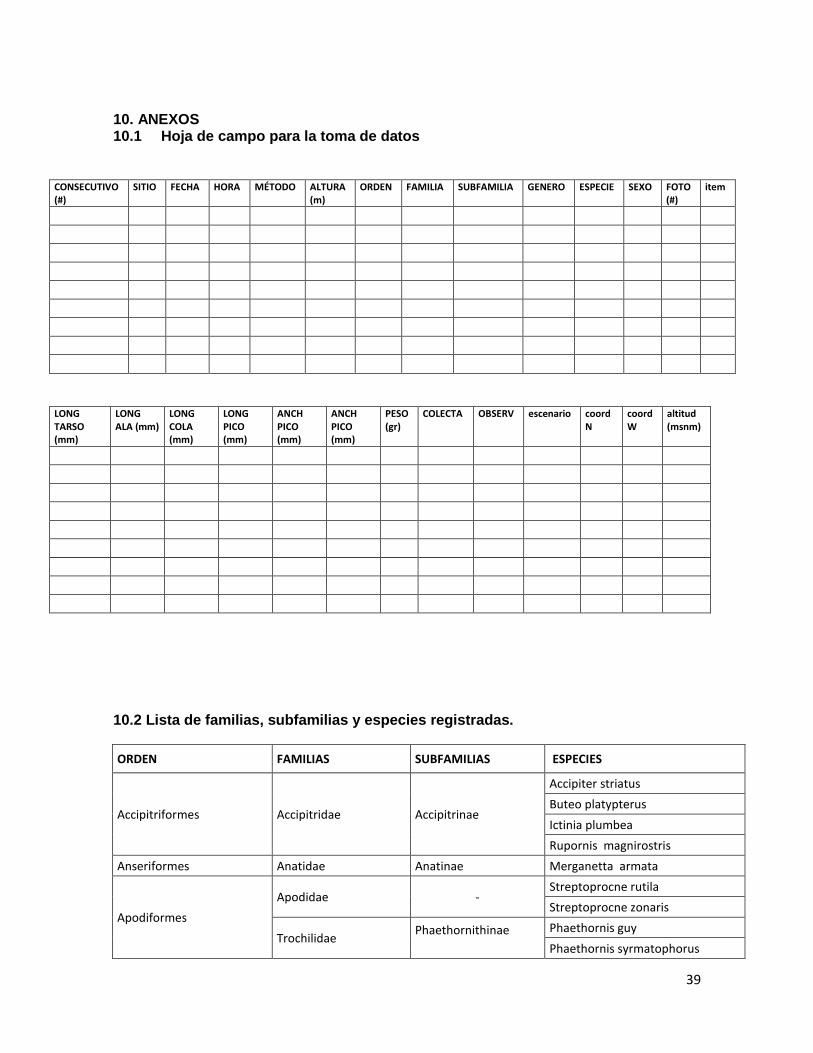
39
10. ANEXOS 10.1 Hoja de campo para la toma de datos
CONSECUTIVO (#)
SITIO FECHA HORA MÉTODO ALTURA (m)
ORDEN FAMILIA SUBFAMILIA GENERO ESPECIE SEXO FOTO (#)
item
LONG TARSO (mm)
LONG ALA (mm)
LONG COLA (mm)
LONG PICO (mm)
ANCH PICO (mm)
ANCH PICO (mm)
PESO (gr)
COLECTA OBSERV escenario
coord N
coord W
altitud (msnm)
10.2 Lista de familias, subfamilias y especies registradas.
ORDEN FAMILIAS SUBFAMILIAS ESPECIES
Accipitriformes Accipitridae Accipitrinae
Accipiter striatus
Buteo platypterus
Ictinia plumbea
Rupornis magnirostris
Anseriformes Anatidae Anatinae Merganetta armata
Apodiformes
Apodidae - Streptoprocne rutila
Streptoprocne zonaris
Trochilidae Phaethornithinae
Phaethornis guy
Phaethornis syrmatophorus
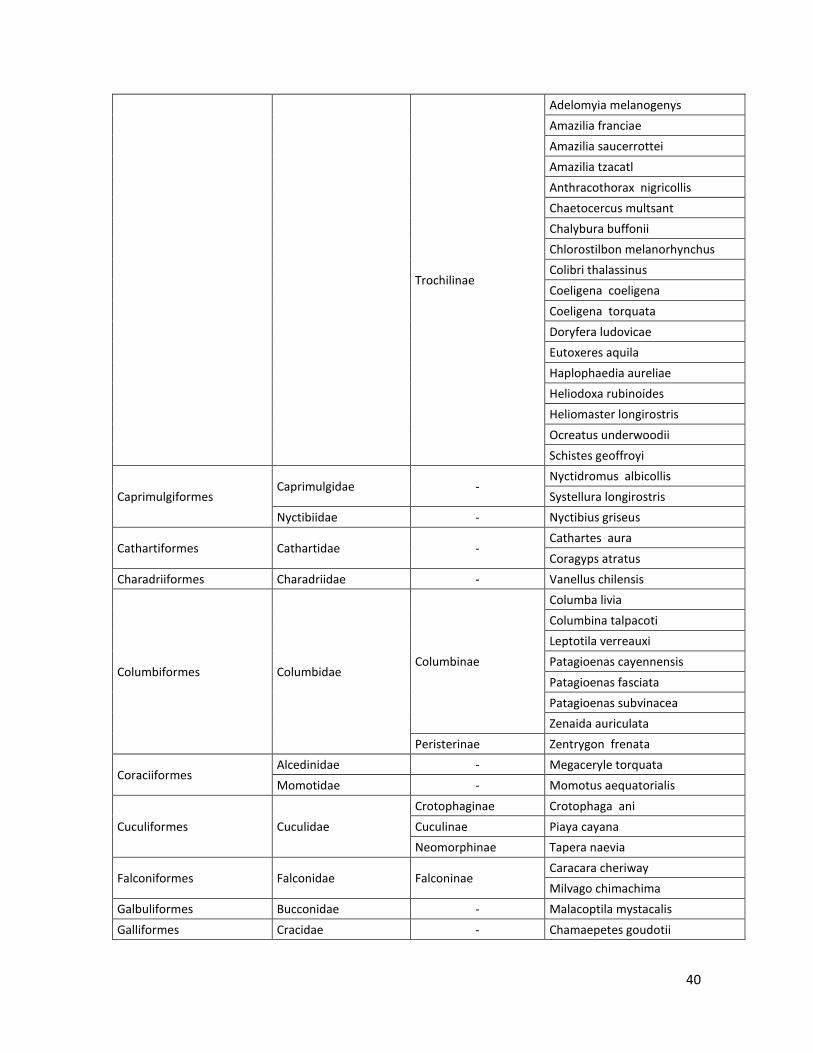
40
Trochilinae
Adelomyia melanogenys
Amazilia franciae
Amazilia saucerrottei
Amazilia tzacatl
Anthracothorax nigricollis
Chaetocercus multsant
Chalybura buffonii
Chlorostilbon melanorhynchus
Colibri thalassinus
Coeligena coeligena
Coeligena torquata
Doryfera ludovicae
Eutoxeres aquila
Haplophaedia aureliae
Heliodoxa rubinoides
Heliomaster longirostris
Ocreatus underwoodii
Schistes geoffroyi
Caprimulgiformes Caprimulgidae -
Nyctidromus albicollis
Systellura longirostris
Nyctibiidae - Nyctibius griseus
Cathartiformes Cathartidae - Cathartes aura
Coragyps atratus
Charadriiformes Charadriidae - Vanellus chilensis
Columbiformes Columbidae Columbinae
Columba livia
Columbina talpacoti
Leptotila verreauxi
Patagioenas cayennensis
Patagioenas fasciata
Patagioenas subvinacea
Zenaida auriculata
Peristerinae Zentrygon frenata
Coraciiformes Alcedinidae - Megaceryle torquata
Momotidae - Momotus aequatorialis
Cuculiformes Cuculidae
Crotophaginae Crotophaga ani
Cuculinae Piaya cayana
Neomorphinae Tapera naevia
Falconiformes Falconidae Falconinae Caracara cheriway
Milvago chimachima
Galbuliformes Bucconidae - Malacoptila mystacalis
Galliformes Cracidae - Chamaepetes goudotii
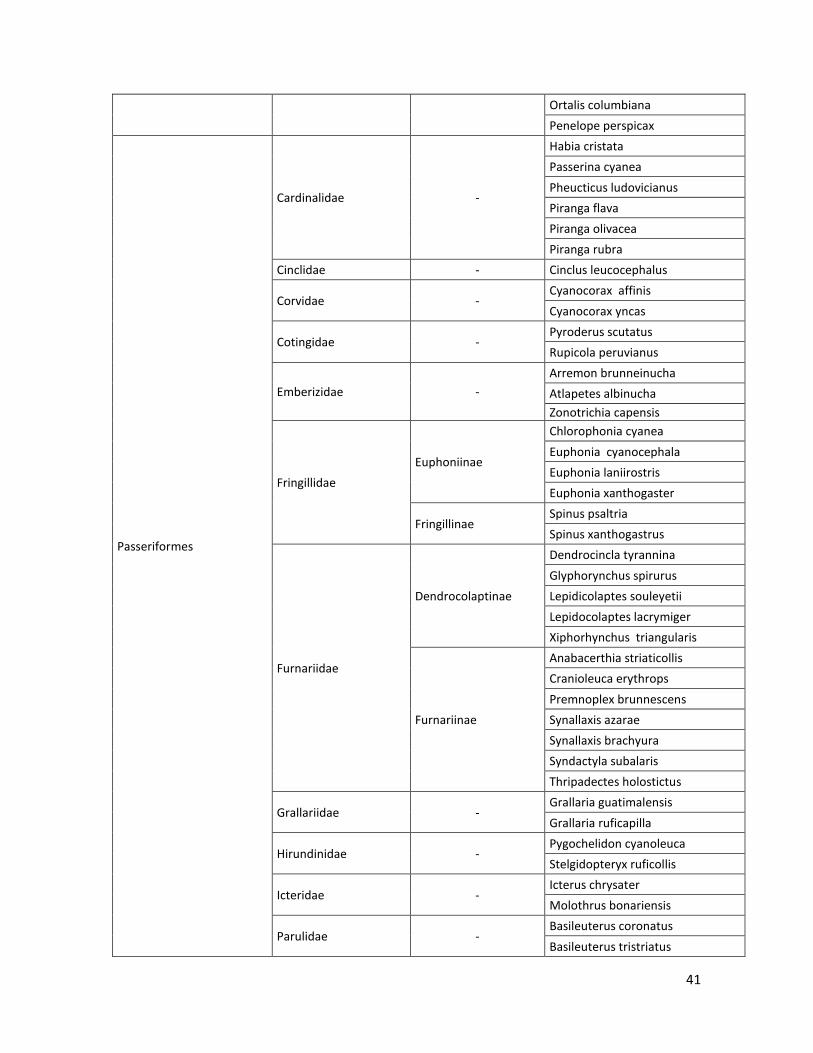
41
Ortalis columbiana
Penelope perspicax
Passeriformes
Cardinalidae -
Habia cristata
Passerina cyanea
Pheucticus ludovicianus
Piranga flava
Piranga olivacea
Piranga rubra
Cinclidae - Cinclus leucocephalus
Corvidae - Cyanocorax affinis
Cyanocorax yncas
Cotingidae - Pyroderus scutatus
Rupicola peruvianus
Emberizidae -
Arremon brunneinucha
Atlapetes albinucha
Zonotrichia capensis
Fringillidae
Euphoniinae
Chlorophonia cyanea
Euphonia cyanocephala
Euphonia laniirostris
Euphonia xanthogaster
Fringillinae Spinus psaltria
Spinus xanthogastrus
Furnariidae
Dendrocolaptinae
Dendrocincla tyrannina
Glyphorynchus spirurus
Lepidicolaptes souleyetii
Lepidocolaptes lacrymiger
Xiphorhynchus triangularis
Furnariinae
Anabacerthia striaticollis
Cranioleuca erythrops
Premnoplex brunnescens
Synallaxis azarae
Synallaxis brachyura
Syndactyla subalaris
Thripadectes holostictus
Grallariidae - Grallaria guatimalensis
Grallaria ruficapilla
Hirundinidae - Pygochelidon cyanoleuca
Stelgidopteryx ruficollis
Icteridae - Icterus chrysater
Molothrus bonariensis
Parulidae - Basileuterus coronatus
Basileuterus tristriatus
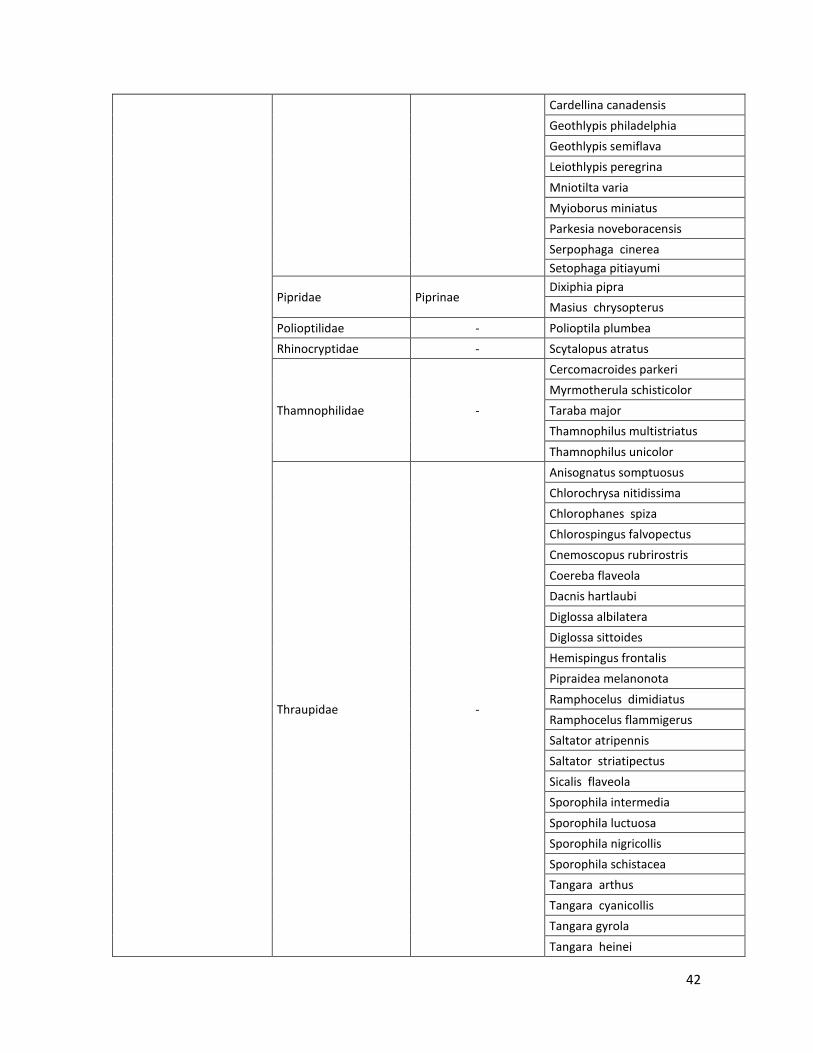
42
Cardellina canadensis
Geothlypis philadelphia
Geothlypis semiflava
Leiothlypis peregrina
Mniotilta varia
Myioborus miniatus
Parkesia noveboracensis
Serpophaga cinerea
Setophaga pitiayumi
Pipridae Piprinae Dixiphia pipra
Masius chrysopterus
Polioptilidae - Polioptila plumbea
Rhinocryptidae - Scytalopus atratus
Thamnophilidae -
Cercomacroides parkeri
Myrmotherula schisticolor
Taraba major
Thamnophilus multistriatus
Thamnophilus unicolor
Thraupidae -
Anisognatus somptuosus
Chlorochrysa nitidissima
Chlorophanes spiza
Chlorospingus falvopectus
Cnemoscopus rubrirostris
Coereba flaveola
Dacnis hartlaubi
Diglossa albilatera
Diglossa sittoides
Hemispingus frontalis
Pipraidea melanonota
Ramphocelus dimidiatus
Ramphocelus flammigerus
Saltator atripennis
Saltator striatipectus
Sicalis flaveola
Sporophila intermedia
Sporophila luctuosa
Sporophila nigricollis
Sporophila schistacea
Tangara arthus
Tangara cyanicollis
Tangara gyrola
Tangara heinei
43
Tangara vitriolina
Tangara labradorides
Tangara nigroviridis
Tangara parzudakii
Tangara ruficervix
Tangara xanthocephala
Thraupis episcopus
Thraupis palmarum
Tiaris obscurus
Tiaris olivaceus
Volatinia jacarina
Tityridae -
Pachyramphus cinnamomeus
Pachyramphus polychopterus
Pachyramphus versicolor
Troglodytae -
Cantorchilus nigricapillus
Cinnycerthia olivascens
Cyphorhinus thoracicus
Henicorhina leucophrys
Microcerculus marginatus
Pheugopedius mistacalis
Troglodytes aedon
Turdidae -
Catharus aurantiirostris
Catharus ustulatus
Myadestes ralloides
Turdus fuscater
Turdus serranus
Turdus ignobilis
Tyrannidae -
Contopus cinereus
Contopus sordidulus
Elaenia flavogaster
Empidonax alnorum
Empidonax virescens
Leptopogon rufipectus
Leptopogon superciliaris
Lophotricus pileatus
Mionectes oleagineus
Mionectes striaticollis
Myiarchus cephalotes
Myiarchus crinitus
Myiarchus tuberculifer
Myiodynastes chrysocephalus
44
Myiopagis viridicata
Myiophobus flavicans
Myiophobus phoenicomitra
Myiozetetes cayanensis
Phyllomyias cinereiceps
Phylloscartes ophthalmicus
Pitangus sulphuratus
Pyrocephalus rubinus
Sayornis nigricans
Serpophaga cinerea
Todirostrum cinereum
Tyrannus melancholicus
Zimmerius chrysops
Vireonidae -
Pachysylvia semibrunnea
Vireo leucophrys
Vireo olivaceus
Pelecaniformes
Ardeidae - Bubulcus ibis
Threskiornithidae - Phimosus infuscatus
Theristicus caudatus
Piciformes
Capitonidae - Eubucco bourcierii
Picidae Picinae
Campephilus melanoleucos
Colaptes rubiginosus
Dryocopus lineatus
Melanerpes formicivorus
Melanerpes rubricapillus
Picumninae Picumnus granadensis
Ramphastidae -
Andigena nigrirostris
Aulacorhynchus haematopygus
Aulacorhynchus prasinus
Psittaciformes Psittacidae Arinae
Amazona amazonica
Amazona mercenarius
Forpus conspicillatus
Pionus chalcopterus
Pionus menstruus
Psittacara wagleri
Strigiformes Strigidae -
Ciccaba virgata
Megascops choliba
Pulsatrix perspicillata
Tinamiformes Tinamidae - Nothocercus bonapartei
Trogoniformes Trogonidae - Trogon collaris
TOTAL 19 45 18 215
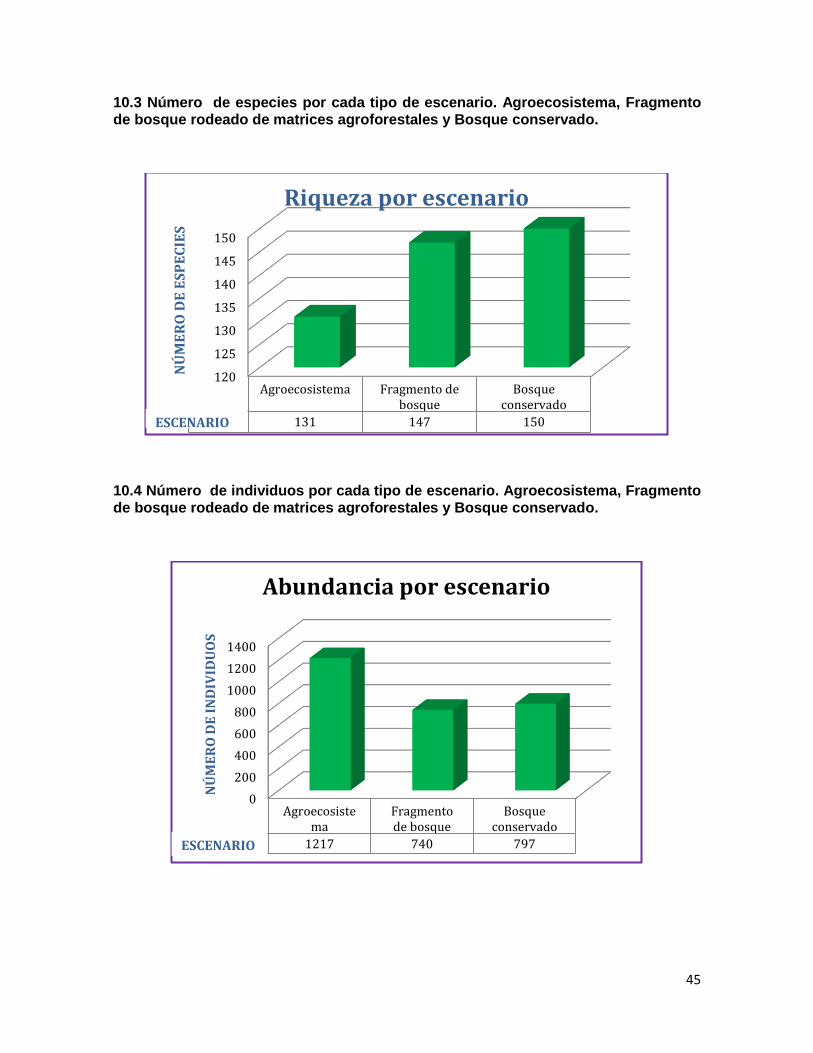
45
10.3 Número de especies por cada tipo de escenario. Agroecosistema, Fragmento de bosque rodeado de matrices agroforestales y Bosque conservado.
10.4 Número de individuos por cada tipo de escenario. Agroecosistema, Fragmento de bosque rodeado de matrices agroforestales y Bosque conservado.
120
125
130
135
140
145
150
Agroecosistema Fragmento debosque
Bosqueconservado
Series1 131 147 150
NÚ
ME
RO
DE
ES
PE
CIE
S
Riqueza por escenario
ESCENARIO
0
200
400
600
800
1000
1200
1400
Agroecosistema
Fragmentode bosque
Bosqueconservado
Series1 1217 740 797
NÚ
ME
RO
DE
IN
DIV
IDU
OS
Abundancia por escenario
ESCENARIO

46
10.5 Especies endémicas registradas para Colombia con su respectiva categoría de
amenaza a escala nacional. (Renjifo et al, 2016)
ESPECIES Grado de amenaza
Penélope perspicax En Peligro (EN)
Ortalis columbiana
Picumnus granadensis
Cercomacroides parkeri,
Dacnis hartlaubi Vulnerable (VU)
Chlorochrysa nitidissima Vulnerable (VU)
Habia cristata
10. 6 Especies migratorias registradas, con su respectiva categoría de migración (Naranjo et al, 2012).
ESPECIES Migración
Ictinia plúmbea Migratoria de Centroamérica; entre Agosto-Marzo (CA)
Buteo platypterus Migratorias boreales; provenientes de Norteamérica, entre Septiembre-Abril (MB)
Empidonax alnorum MB
Empidonax virescens MB
Contopus sordidulus MB
Myiarchus crinitus MB
Catharus ustulatus MB
Piranga rubra MB
Piranga olivácea MB
Pheucticus ludovicianus MB
Passerina cyanea MB
Leiothlypis peregrina MB
Mniotilta varia MB
Parkesia noveboracensis MB
Geothlypis philadelphia MB
Cardellina canadensis MB




















