Tugas Ento Hormon Juvenil
10
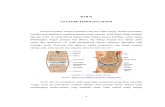
JUVENILE HORMON MENGONTROL PERTUMBUHAN DAN PERKEMBANGAN SERANGGA Metamorfosis serangga dikendalikan oleh JH. Regulasi JH mempunyai peranan yang penting dalam mengendalikan metamorfosis. Proses dimana JH berperan dalam mengatur pertumbuhan dan perkembangan serangga, dimulai dari sel neurosecretory yang ada pada otak akan menghasilkan allatotropin yang digunakan untuk menstimulasi corpora allata untuk memproduksi JH (Li et al.,2005). Proses selanjutnya JH akan dikeluarkan oleh corpora allata ke dalam hemolymph. JH yang berada pada hemolymph akan diikat oleh juvenile hormon binding protein (JHBP) yang berfungsi untuk memudahkan larut dalam hemolymph dan didistribusikan pada sel epidermis. JHBP kemudian akan terdistribusi pada sel epidermis yang kemudian akan terjadi moulting. Konsentrasi JH dalam hemolymph menentukan apakah larva akan moulting pada fase berikutnya atau akan berubah bentuk menjadi pupa demikian juga menentukan apakah pupa akan berubah bentuk menjadi dewasa. Jika dalam hemolymph larva konsentrasi JH tinggi maka larva akan melakukan moulting tetapi jika konsentrasi JH rendah sedangkan hormon 20-hydroxyecdysone rendah makan akan memberi signal larva untuk berubah menjadi pupa. Proses pengaturan JH pada serangga dapat dilihat pada gambar 4 (Gilbert et al.,1980). Ewer et al. (1997) memberi gambaran bagaimana pengaktifan hormon mempengaruhi perilaku yaitu proses ecdysis larva Manduca sexta. Ecdysis merupakan pergantian kulit dari
-
Upload
asmidyah-dwi-rahayu-part-ii -
Category
Documents
-
view
34 -
download
6
description
Tugas entomologi tentang hormon juvenil
Transcript of Tugas Ento Hormon Juvenil
JUVENILE HORMON MENGONTROL PERTUMBUHAN DAN PERKEMBANGAN SERANGGA
Metamorfosis serangga dikendalikan oleh JH. Regulasi JH mempunyai peranan yang penting dalam mengendalikan metamorfosis. Proses dimana JH berperan dalam mengatur pertumbuhan dan perkembangan serangga, dimulai dari sel neurosecretory yang ada pada otak akan menghasilkan allatotropin yang digunakan untuk menstimulasi corpora allata untuk memproduksi JH (Liet al.,2005). Proses selanjutnya JH akan dikeluarkan oleh corpora allata ke dalam hemolymph. JH yang berada pada hemolymph akan diikat olehjuvenile hormon binding protein(JHBP) yang berfungsi untuk memudahkan larut dalam hemolymph dan didistribusikan pada sel epidermis. JHBP kemudian akan terdistribusi pada sel epidermis yang kemudian akan terjadi moulting. Konsentrasi JH dalam hemolymph menentukan apakah larva akan moulting pada fase berikutnya atau akan berubah bentuk menjadi pupa demikian juga menentukan apakah pupa akan berubah bentuk menjadi dewasa. Jika dalam hemolymph larva konsentrasi JH tinggi maka larva akan melakukan moulting tetapi jika konsentrasi JH rendah sedangkan hormon 20-hydroxyecdysone rendah makan akan memberi signal larva untuk berubah menjadi pupa. Proses pengaturan JH pada serangga dapat dilihat pada gambar 4 (Gilbertet al.,1980).
Eweret al.(1997) memberi gambaran bagaimana pengaktifan hormon mempengaruhi perilaku yaitu proses ecdysis larvaManduca sexta. Ecdysis merupakan pergantian kulit dari kulit lama pada saat moulting, proses ini tergantung positive feedback antara hormon eclosion dan JH. Pelepasan hormon neuropeptide dari sel neurosecretory dalam sistem syaraf pusat menyebabkan peripheral yang terletak pada kelenjar epitracheal melepaskan hormon yang akan memicu ecdysis.
Gambar 4. A. Peranan JH dalam Mengatur Pertumbuhan Serangga, B. Proses Pelepasan JH untuk mengontrol proses moulting pada serangga (Gilbertet al.,1980).
JUVENILE HORMON MEMPENGARUHI SINTESIS VITELLOGENIN
Perkembangan dan reproduksi tergantung dari JH dan ecdysteroids. Pada sebagian besar serangga, JH merupakan hormon yang berperan besar dalam proses regulasi sintesis dan pengambilan vitellogenin, tetapi faktor ecdysteroid juga diperlukan dalam proses ini (Engelman, 1984, Koeppeet al.,1985dalamLorenzet al.,1998).
Martinezet al.,(2007) juga melaporkan bahwa JH merupakan hormon yang mempunyai peranan penting dalam mengatur perkembanganprevitellogonic ovarian. Bukti yang menunjukkan bahwa JH mengatur perkembanganprevitellogonic ovarianyaitu penelitian yang dilakukan oleh Martinezet al.,(2007) terhadap nyamuk. JH di dalamAe. aegyptijumlahnya sedikit pada saat eclosion, dan meningkat pada hari pertama setelah imago muncul. Jumlah JH yang naik pada saat awal sangat penting untuk menyempurnakan organ reproduksi serangga betina. Kecapatan biosintesis JH oleh corpora allata secara in vitro mencerminkan tingkat JH dalam nyamuk, biosintesis JH sangat rendah pada serangga betina baru yang muncul dan meningkat drastis selama 24 jam setelah eclosion (Liet al., 2003). Aktivitas corpora allata nyamuk dikendalikan oleh faktor-faktor yang terdapat di kepala (Liet al., 2004), dan signal nutrisi akan mempengaruhi aktivasi sintesis JH atau menghambat sintesis JH.. Pemenggalan kepala dalam 1 h dariemergenceatau penghilangan CA setelah eclosion mencegah pertumbuhan ovarian previtellogenic.
Menurut Hagedornet al.,(1997dalamCarociet al.,2004) mekanisme JH mempengaruhi sintesis vitellogenin pada nyamukA. aegyptiyaitu neurosecretory pada otak akan menghasilkan allatotropin yang selanjutnya memerintah corpora allata untuk menghasilkan JH. JH yang sudah dihasilkan oleh corpora allata akan menstimuli fat body dari incompetence menjadi competence untuk menghasilkan vitellogenin. Pada kondisi ini JH hanya menstimuli fat body menjadi competence (siap untuk menghasilkan vitellogenin), JH tidak memerintah fat body untuk menghasilkan vitellogenin. JH juga mempengaruhi ovary dari immature ovary menjadi ovary yang mature tetapi inaktif (keadaan ovary siap untuk menjalankan perintah berikutnya). JH juga mempengaruhi perilaku mating dan feeding serangga, setelah nyamuk menghisap darah maka otak akan menyuruh neurosecretory sel untuk menghasilkan Egg development neurohormone (EDNH) dan selanjutnya akan dilepaskan dalam hemolymp. EDNH dalam hemolym kemudian akan diterima oleh ovary yang inaktif (resting stage ovary) dan menstimuli sel folikel untuk menghasilkan ecdysteroid. Ecdysteroid selanjutnya akan memerintah fat body yang sudah competence untuk menghasilkan vitellogenin. Vitellogenin kemudian akan diambil oleh ovary untuk menyusun kuning telur, dan selanjutnya akan menghasilkan telur
JUVENILE HORMON MENGINDUKSI POLYPHENISM DAN POLYMORPHISM
Suzuki dan Nijhout (2006)dalamVerma (2007) mendefiniskan polyphenism adalah adaptasi di mana suatu genom dihubungkan dengan fenotipe alternatif dengan ciri-ciri tertentu pada lingkungan yang berbeda. Polifenisme dapat juga definisikan sebagai adaptasi suatu genotipe untuk menghasilkan fenotipe yang berbeda pada kondisi lingkungan yang berbeda.
Polymorphisme diakibatkan oleh bermacam-macam penyebab antara lain : morfologi yang berbeda kemungkinan dikode dengan genotipe yang berbeda (geneticpolymorphism), diinduksi/dipicu oleh lingkungan yang berbeda (environment polymorphism) atau yang dihasilkan oleh variasi genetik dan faktor lingkungan. Contoh-contoh penting fase polymorphism antara lain polymorphism pada belalang, polymorphism kasta pada serangga sosial, polymorphisme seasonal pada kupu-kupu dan polymorphism sayap pada berbagai jenis serangga. Polymorphism pada umumnya mempunyai peranan integral dalam siklus hidup suatu spesies sebagai contoh menghasilkan penyebaran serangga, menghasilkan fenotipe untuk menyesuaikan diri dengan musim tertentu atau habitat (Emlen dan Nijhout, 2000).
Emlen dan Nijhout (2000) telah mengembangkan model konseptual untuk menjelaskan bagaimana JH mengontrol polymorphism dan polyphenism.Polymorphism dan polyphenis mempunyai karakteristik dengan bentuk yang mencolok dari variasi intraspesifik pada perilaku, morfologi, dan fisiologi. Polymorphism biasanya menjelaskan variasi antara individual yang berasosiasi dengan variasi genotype dan polyphenism menjelaskan variasi persamaan antara genotypically individu yang dipengaruhi faktor lingkungan. Nijhout dan Wheeler menyatakan bahwa JH mengontrol polymorphism dan polyphenism dengan mempengaruhi gen expression selama periode sensitif JH yang terjadi secara periodik selama serangga hidup. JH diketahui untuk mengatur ekspresi dari variasi gen, tetapi analisis ini belum diperluas untuk polymorphism atau polyphenisms.
DAFTAR PUSTAKA
Caroci, A.S., Y. Li., F. G. Noriega. 2004. Reduced juvenile hormone synthesis in mosquitoes with low teneral reserves reduces ovarian previtellogenic development inAedes aegypti.The Journal of Experimental Biology 207: 2685-2690Diana E. Wheeler dan H. F. Nijhout. 2000. A perspective for understdaning the modes of juvenile hormon action as a lipid signaling system.BioEssays 25:9941001, 2003.Elekonich, M.M dan G.E. Robinson. 2000. Organizational and activational effects of hormones on insect behavior.Journal of Insect Physiology 46(2000) : 15091515Emlen, D.J dan H.F. Nijhout. 2000. Hormonal control of male horn length dimorphism in the dung beetleOnthophagus taurus(Coleoptera: Scarabaeidae).Journal of Insect Physiology 45 (1999) : 4553Emlen, D.J dan H.F. Nijhout. 1999. The Development Dan Evolution Of Exaggerated Morphologies In Insects.Annu. Rev. Entomol. 2000. 45:661708.Gilbert, J Tata, P Atkison, eds. 1980. Metamorphosis: post-embryonic reprogramming of gene expression in amphibian dan insectcells.SanDiego: Academic Press, pp 59-107.Li, S., Y.C. Ouyang, E. Ostrowski, D.W. Borst. 2005. Allatotropin regulation of juvenile hormone synthesis by the corpora allata from the lubber grasshopper,Romalea microptera.Peptides 26 (2005) 6372Lorenz, M.W., K.H.Hoffmann, G.G.de. 1998. Juvenile hormone biosynthesis in larval and adult stick insects,Carausius morosus.Journal of Insect Physiology 45(1999) : 443452Martinez, S. H, J. G. Mayoral, Y. Li, F. G. Noriega. 2007. Role of juvenile hormon dan allatotropin on nutrient allocation, ovarian development dan survivorship in mosquitoes.Journal of Insect Physiology 53(2007) : 230234Nijhout, H.F. 2003. The control of body size in insects.Developmental Biology 261 (2003) : 19Robinson, G.E dan E. L. Vargo. 1997. Juvenile Hormone in Adult Eusocial Hymenoptera: Gonadotropin and Behavioral Pacemaker.Insect Biochemistry and Physiology 35:559583Sen, S.E., A.E. Sperry, M. Childress, D.E. Hannemann. 2003. Juvenile hormon biosynthesis in moths: synthesis dan evaluation of farnesol homologs as alternate substrates of farnesol oxidase.Insect Biochemistry dan Molecular Biology 33 (2003) 601607Shelby, J.A, R. Madewell, A.P. Moczek. 2007. Juvenile Hormone Mediates Sexual Dimorphism in Horned Beetles.J. Exp. Zool. (Mol. Dev. Evol.) DOI 10.1002/jez.: 1-11Smith SL. 1985. Regulation of ecdysteroid titre: synthesis.InGA Kerkut, LI Gilbert, eds.Comprehensive insect physiology, biochemistry dan pharmacology. Vol 8.Oxford: Pergamon Press, pp 295-341.Verma, K.K. 2007. Polyphenism in insects and the juvenile hormone.J. Biosci.32(2) (2007) : 415420.
TUGAS TERSTRUKTURMATA KULIAH ENTOMOLOGIDosen Pengampu Mata Kuliah: Tita Widjayanti, SP., MP.
Disusun oleh:Nama: Asmidyah Dwi RahayuNim: 125040201111019Kelas: C
JURUSAN HAMA DAN PENYAKIT TUMBUHANPROGRAM STUDI AGROEKOTEKNOLOGIFAKULTAS PERTANIANUNIVERSITAS BRAWIJAYAMALANG2015