Setelah Proses Penyerapan Melalui Dinding Usus Halus
18
Click here to load reader
-
Upload
galuh-panji-rakasiwi -
Category
Documents
-
view
86 -
download
2
Transcript of Setelah Proses Penyerapan Melalui Dinding Usus Halus

MAAF TELAT
Setelah proses penyerapan melalui dinding usus halus, sebagian besar monosakarida di bawa oleh aliran darah ke hati. Di dalam hati, monosakarida mengalami proses sintesis menghasilkan glikogen, oksidasi menjadi CO2 dan H2O, atau dilepaskan untuk dibawa dengan aliran darah ke bagian tubuh yang memerlukannya. Sebagian lain monosakarida dibawa langsung ke sel jaringan organ tertentu dan mengalami proses metabolisme lebih lanjut.
Menurut Drh Kadek Rachmawati M.Kes ketika kadar glukosa dalam darah tinggi maka akan memacu jalur metabolisme karbohidrat berikut :
1. Glikolisis 1.1 Glikolisis aerob1.2 Glikolisis anaerob
2. Glikogenesis 3. HMP Shunt4. Oksidasi Piruvat 5. Siklus Asam Sitrat 6. Sisa ditimbun sbg lemak
Sedangkan ketika kadar glukosa dalam darah rendah ( kelaparan ) maka akan memacu jalur metabolisme karbohidrat berikut :
7. Glikogenolisis 8. Glukoneogenesis

Menurut Muhamad Wiradikusumah dalam bukunya yang berjudul biokimia proses penguraian glukosa menjadi piruvat, alcohol, laktat, atau CO2 dan air dapat berlangsung melalui beberapa jalan metabolism, tergantung dari keadaan lingkungan, keadaan dalam sel, atau macam jasadnya.
1. Glikolisis
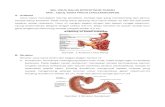
1.1 GlikolisisDengan adanya oksigen ( dalam suasana aerob), glikolisis berlangsung menghasilkan
piruvat, atau tanpa oksigen (glikolisis anaerob) menghailkan laktat. Glikolisis menghasilkan dua senyawa karbohidrat beratom tiga dari satu senyawa beratom enam. Pada proses ini terjadi sintesis ATP dari ADP + Pi. Gambar dibawah menunjukkan proses glikolisis secara keseluruhan.

Bila glikolisis dumulai dari glukosa , reaksi pertamanya adalah pemasukan gugus fosfat ke dalam molekul glukosa menghasilkan glukosa 6-fosfat. Reaksi ini dikatalis oleh glukokinase. Sedangkan gugus fosfat dan energy yang diperlukan didapat dari penguraian ATP menjadi ADP. Reaksi tahap dua adalah merupakan isomerasi glukosa 6-fosfat menjadi fruktosa 6-fosfat, dikatalis oleh fosfoheksoisomerase yang juga mengkatalis reaksi kebalikannya.
Reaksi tahap ketiga adalah pemasukan gugus fosfat dari ATP, dikatalis oleh fosfofruktosakinase dengan ion MG2+ sebagai kofaktor menghasilkan fruktosa 1,6- difosfat. Keempat merupakan pemecahan enam menjadi 2 senyawa beratom tiga, gliseraldehid 3-fosfat dan dehidroksiaseton fosfat. Selanjutnya pada reaksi tahap kelima merupakan perubahan gliseraldehida 3-fosfat menjadi asam 1,3 -difosfogliserat, yang melibatkan reaksi pemasukan satu gugus fosfat dari asam fosfat.
Selanjutnya pada reaksi tahap keenam dikatalis oleh fosfo gliserat kinase ( dengan ion magnesium sebagai kofaktor), menghasilkan asam 3-fosfogliserat. Reaksi tahap kelima dalam glikolisis merupakan reaksi pertama yang menghasilkan energy. Reaksi tahap ketujuh adalah isomirasi asam gliserat 3-fosfat menjadi asam gliserat 2-fosfat, dikatalis oleh fosfogliserat mutase. Selanjutnya dealam reaksi tahap kedelapan enzim enolase melepaskan satu molekul H20 dari asam fosfoenol piruvat. Dan tahap yang terakhir dari glikolis aerob adalah pembentukan asam piruvat dari asam fosfoenolpiruvat melalui senyawa antara asam enolpiruvat. Pada akhir reaksi glikolisis aerob didapatkan 10 mol ATP dikurangi 2 ATP sebagai reaksi menjadi 8 mol ATP.
1.2 Glikolisis anaerob
Dalam keadaan tanpa oksigen respirasi terhenti karena proses pengangkutan electron yang dirangkaikan dengan fosforolasi bersifat oksidasi melalui rantai pernafasan yang menggunakan molekul oksigen sebagai penerima electron terakhir, tidak berjalan. Akibatnya jalur metabolism lingkar trikarboksilat ( daur Krebs ) akan terhenti pula sehingga piruvat tidak lagi masuk ke dalam daur krebs melainkan dialihkan pemakainnya yaitu diubah menjadi asam laktat oleh laktat dehidrogenase. Dalam glikolisis anaerob hanya dhasilkan 4 ATP dikurangi 2 ATP menjadi 2 ATP saja.
2. Glikogenesis
Glikogenesis adalah lintasan metabolisme yang mengkonversi glukosa menjadi glikogen untuk disimpan di dalam hati. Lintasan diaktivasi di dalam hati, oleh hormon insulin sebagai respon terhadap rasio gula darah yang meningkat, misalnya karena kandungan karbohidrat setelah makan; atau teraktivasi pada akhir siklus Cori.
Proses glikogenesis adalah sebagai berikut :
1) Glukosa mengalami fosforilasi menjadi glukosa 6-fosfat (reaksi yang lazim terjadi juga pada lintasan glikolisis). Di otot reaksi ini dikatalisir oleh heksokinase sedangkan di hati oleh glukokinase.

2) Glukosa 6-fosfat diubah menjadi glukosa 1-fosfat dalam reaksi dengan bantuan katalisator enzim fosfoglukomutase. Enzim itu sendiri akan mengalami fosforilasi dan gugus fosfo akan mengambil bagian di dalam reaksi reversible yang intermediatnya adalah glukosa 1,6-bifosfat. Enz-P + Glukosa 1-fosfat↔Enz + Glukosa 1,6-bifosfat↔Enz-P + Glukosa 6-fosfat
3. Selanjutnya glukosa 1-fosfat bereaksi dengan uridin trifosfat (UTP) untuk membentuk uridin difosfat glukosa (UDPGlc). Reaksi ini dikatalisir oleh enzim UDPGlc pirofosforilase. UDPGlc + PPi↔UTP + Glukosa 1-fosfat
4. Hidrolisis pirofosfat inorganic berikutnya oleh enzim pirofosfatase inorganik akan menarik reaksi kearah kanan persamaan reaksi.
5. Atom C1 pada glukosa yang diaktifkan oleh UDPGlc membentuk ikatan glikosidik dengan atom C4 pada residu glukosa terminal glikogen, sehingga membebaskan uridin difosfat. Reaksi ini dikatalisir oleh enzim glikogen sintase. Molekul glikogen yang sudah ada sebelumnya (disebut glikogen primer) harus ada untuk memulai reaksi ini. Glikogen primer selanjutnya dapat terbentuk pada primer protein yang dikenal sebagai glikogenin. UDP + (C6)n+1◊ UDPGlc + (C6)n
uridinitrifosfat dengan glukosa-1-P. Siklus pengubahan glukosa, asam laktat dan glikogen disebut dengan siklus cori.3. HMP SHUNT
Biasanya HMP-Shunt Disebut juga jalur pentosa fosfat / heksosa monofosfat. Jalur ini menghasilkan NADPH dan ribosa di luar mitokondria. NADPH diperlukan untuk biosintesis; asam lemak, kolesterol, dan steroid lain. Ribosa untuk biosintesis asam nukleat. Kepentingan lain HMP-shunt berlangsung dalam jaringan; hepar, lemak, korteks adrenal, tiroid, eritrosit, kelenjar mammae sedang laktasi. NADPH juga penting dalam; detoksifikasi obat oleh monooksigenase, reduksi glutation. HMP-shunt terdiri dari fase: 1. Oksidatif (irreversible); glukosa 6-fosfat --->

ribulosa 5-fosfat2. Non-oksidatif (reversible); ribulosa 5-fosfat ---> ribosa 5-fosfat . Jalur ini aktif dalam hepar, jaringan adiposa (lemak), adrenal korteks, glandula tiroid, sel darah merah, testis dan payudara yang sedang menyusui. Dalam otot aktivitas jalur ini rendah sekali.
Fungsi utama jalur ini adalah untuk menghasilkan NADPH, yaitu dengan mereduksi NADP+. NADPH diperlukan untuk proses anabolik di luar mitokhondria, seperti sintesis asam lemak dan steroid. Fungsi yang lain adalah menghasilkan ribosa-5-fosfat untuk sintesis nukleotida dan asam nukleat.
-D-glukosa 6-fosfat mengalami oksidasi menjadi 6-fosfoglukonolakton. Enzimnya adalah glukosa 6-fosfat dehidrogenase (G6PD). Reaksi ini memerlukan Mg++ atau Ca++ , memakai NADP+ dan menghasilkan NADPH. Insulin meningkatkan sintesis enzim ini.
Selanjutnya 6-fosfoglukonolakton diubah menjadi 6-fosfoglukonat. Reaksi ini juga memer-lukan Mg++, Mn++ atau Ca++. Enzimnya glukono-lakton hidrolase. Satu molekul air (H2O) terpakai, ikatan cincin terlepas. 6-fosfoglukonat selanjutnya mengalami dekarboksilasi dan berubah menjadi riboluse-5-fosfat.
Sebelum dekarboksilasi 6-fosfoglukonat dioksidasi menjadi semyawa antara 3-keto 6-fosfoglukonat. Ion Mg++, Mn++ atau Ca++ diperlukan. NADP+ bertindak sebagai hidrogen ekseptor menjadi NADPH. Enzim yang mengkatalisis reaksi ini adalah 6-fosfoglukonat dehidrogenase. Aktivitas enzim ini tergantung adanya NADP+. Seperti halnya enzim G6PD enzim 6-fosfoglukonat dehidrogenase sintesisnya dirangsang oleh insulin. Selanjutnya Ribulosa 5-fosfat dapat menjadi dua substrat dari dua enzim yaitu:
1. Ribulosa 5-fosfat epimerase, yang membentuk suatu epimer pada karbon ketiga, yaitu xylulose 5-fosfat (xylulose 5-phosphate).
2. Ribosa 5-fosfat ketoisomerase, yang merubah ribulosa 5-fosfat menjadi ribosa 5-fosfat.
Proses selanjutnya akan melibatkan suatu enzim transketolase, yang dapat memindah dua unit karbon ( C1 dan C2 ) dari suatu ketosa pada aldehida dari aldosa. Dalam reaksi ini diperlukan suatu koenzim, tiamin difosfat dan ion Mg++. Dua karbon dari xylulose 5-fosfat dipindah pada ribosa 5-fosfat, menghasilkan suatu ketosa dengan tujuh karbon yaitu sedoheptulosa 7-fosfat dan aldosa dengan tiga karbon gliseraldehida 3-fosfat. Sedoheptulosa 7-fosfat dan gliseraldehida 3-fosfat akan bereaksi dengan bantuan enzim transaldolase dan membentuk fruktosa 6-fosfat dan eritrosa 4-fosfat.Dalam reaksi ini, transaldolase memindah tiga karbon "active dihydroxy acetone" (C1-C3) dari keto dengan tujuh karbon pada aldosa dengan tiga karbon.
Reaksi selanjutnya kembali melibatkan enzim transketolase, dimana xylulose 5-fosfat menjadi donor "active glycoaldehyde" (C1-C2). Eritrosa 4-fosfat yang terbentuk dari reaksi sebelumnya, akan bertindak sebagai akseptor (penerima) C1-C2. Reaksi ini memerlukan tiamin dan ion Mg++ sebagai ko-enzim dan menghasilkan fruktosa 6-fosfat dan gliseraldehida 3-fosfat. Agar glukosa dapat dioksidasi secara sempurna menjadi CO2, diperlukan enzim yang dapat mengubah gliseraldehide 3-fosfat menjadi glukosa 6-fosfat. Untuk ini diperlukan enzim Embden-Meyerhof (glikolisis) yang bekerja kearah yang berlawanan. Selain itu, juga diperlukan enzim fruktosa 1,6-difosfatase. Enzim ini mengubah fruktosa 1,6-difosfat menjadi fruktosa 6-fosfat.
Secara keseluruhan proses ini dapat dianggap suatu oksidasi tiga molekul glukosa 6-fosfat menjadi tiga molekul CO2 dan tiga molekul pentosa fosfat. Tiga molekul pentosa fosfat diubah menjadi dua molekul glukosa fosfat dan satu molekul gliseraldehida 3-fosfat. Karena dua molekul gliseraldehide 3-fosfat dapat diubah menjadi satu molekul glukosa 6-fosfat melalui jalur

kebalikan glikolisis, maka HMP Shunt dapat dikatakan suatu oksidasi glukosa yang komplit (sempurna).
Enzim 6-fosfoglukonat dehidrogenase mengontrol HMP Shunt. Enzim ini dapat dihambat oleh NADPH. Reaksi yang dikatalisis enzim ini tidak akan berjalan apabila NADPH tidak dipakai atau dengan kata lain konsentrasinya tidak menurun. Perlu diingat bahwa produksi ribosa 5-fosfat tidak tergantung pada oksidasi glukosa, tapi dapat melewati kebalikan jalur glikolisis. NADPH yang terbentuk berguna dalam sintesis asam lemak, steroid dan sintesis asam amino. Sintesis asam amino melalui glutamat dehidrogenase. Adanya lipogenesis yang aktif ,maka NADPH diperlukan, hal ini mungkin akan merangsang oksidasi glukosa lewat HMP Shunt. "Fed state", suatu keadaan dimana seseorang baru saja makan, mungkin dapat menginduksi sintesis enzim-enzim glukosa 6-fosfat dehidro-genase dan 6-fosfoglukonat dehidrogenase.
HMP Shunt dalam eritrosit berguna sebagai penghasil suatu reduktor (NADPH). NADPH dapat mereduksi glutation yang telah mengalami oksidasi ( G-S-S-G ) menjadi glutation yang tereduksi (2 G-SH). Enzim yang mengkatalisis reaksi ini adalah glutation reduktase. Selanjutnya glutation yang tereduksi dapat membebaskan eritrosit dari H2O2 dengan suatu reaksi yang dikatalisis oleh enzim glutation peroksidase.
2G-SH + H2O2 ® G-S-S-G + 2 H2O
Reaksi ini penting sebab penimbunan H2O2 memperpendek umur eritrosit. Telah dibuktikan adanya korelasi terbalik antara aktivitas enzim glukosa 6-fosfat dehidrogenase dengan fragilitas sel darah merah. Pada beberapa orang yang mengalami mutasi dimana enzim ini berkurang, maka mereka akan lebih mudah mengalami hemolisis sel darah merah apabila diberi suatu oksidan seperti primaquin, aspirin, sulfonamid atau apabila diberi makan "fava bean".
HMP Shunt akan menghasilkan suatu pentosa untuk sintesis nukleotida dan asam nukleat. Ribosa 5-fosfat akan bereaksi dengan ATP menjadi 5-fosforibosil-1-pirofosfat (PRPP). Dalam otot enzim glukosa 6-fosfat dehidro-genase dan 6-fosfoglukonat dehidrogenase hanya sedikit sekali, namun otot dapat membuat ribosa 5-fosfat, yaitu dengan kebalikan HMP-Shunt.
(Lehninger A.L., Nelson D.L and Cox M.M : Principles of Biochemistry. Second Ed. Worth Publ. Inc. New York. 1993)
4. OKSIDASI PIRUVATDalam jalur ini, piruvat dioksidasi (dekarboksilasi oksidatif) menjadi Asetil-KoA, yang
terjadi didalam mitokondria sel. Reaksi ini dikatalisir oleh berbagai enzim yang berbeda yang bekerja secara berurutan di dalam suatu kompleks multi enzim yang berkaitan dengan membran dalam mitokondria. Secara kolektif, enzim tersebut diberi nama kompleks piruvat dehidrogenase
dan analog dengan kompleks ∝-ketoglutarat dehidrogenase pada siklus asam sitrat. Jalur ini merupakan penghubung antara glikolisis dan siklus Kreb’s. Rangkaian reaksi kimia yang terjadi dalam lintasan oksidasi piruvat adalah sebagai berikut:
1. Piruvat mengalami dekarboksilasi oleh komponen piruvat dehidrogenase pada kompleks enzimtersebut menjadi turunan hidroksi etil cincin tiazol tiamin difosfat (yang terikat enzim).
2. Tiamin difosfat bereaksi dengan lipoamida teroksidasi, yakni gugus prostetik pada dihidrolipoiltrans asetilase membentuk asetil lipoamida.
3. Asetil lipoamida bereaksi dengan koenzim A membentuk asetil-koA dan lipoamida tereduksi.Reaksi ini tuntas apabila lipoamida yang tereduksi tersebut

direoksidasi oleh suatu flavoprotein,yaitu dihidrolipoil dehydrogenase, yang mengandung FAD.
4. Flavo protein terduksi mengalami oksidasi oleh NAD+, memindahkan ekivalen pereduksi kerantai respiratorik.
Piruvat + NAD+ +KoA → Asetil-koA
Jadi, oksidasi piruvat akan menghasilkan Asetil-koA, NADH, dan COPiruvat dehidrogense dihambat oleh produknya sendiri yaitu asetil-koA dan NADH. Enzim
ini juga diatur melalui fosforilasi oleh suatu kinase tiga residu serin pada komponen piruvat dehidrogenase kompleks multi enzim sehingga akivitas enzim menurun dan menyebabkan peningkatan aktivitas melalui defosforilasi oleh satu fostatase. Kinase diaktifkan oleh peningkatan rasio [ATP] / [ADP], [asetil-koA] / [ko-A], dan [NADH] / [NAD+]. Karenaitu, piruvat dehidrogense, demikian juga dengan glikolisis dihambat jika tersedia ATP dalam jumlah memadai dan jika asam lemak teroksidasi. Dalam keadaan puasa, ketika konsentrasi asam lemak bebas meningkat, terjadi penurunan proporsi enzim tersebut dalam bentuk aktif sehingga karbohidrat dihemat.
5. SIKLUS ASAM SITRAT
Siklus Krebs adalah proses utama kedua dalam reaksi pernafasan sel. Siklus Krebs ini ditemukan oleh Hans Krebs (1900-1981). Reaksi pernafasan sel tersebut disebut juga sebagai daur asam sitrat atau daur asam trikarboksilat.
Tahapan Reaksi dalam Siklus Krebs
Siklus Krebs terjadi di mitokondria dengan menggunakan bahan utama berupa asetil-CoA, yang dihasilkan dari proses dekarboksilasi oksidatif. Ada delapan tahapan utama yang terjadi selama siklus Krebs.

1. Kondensasi
Kondensasi merupakan reaksi penggabungan molekul asetil-CoA dengan oksaloasetat membentuk asam sitrat. Enzim yang bekerja dalam reaksi ini adalah enzim asam sitrat sintetase.
2. Isomerase sitrat
Tahapan ini dibantu oleh enzim aconitase, yang menghasilkan isositrat.
3. Produksi CO2
Dengan bantuan NADH, enzim isositrat dehidrogenase akan mengubah isositra menjadi alfa-ketoglutarat. Satu molekul CO2 dibebaskan setiap satu reaksi.
4. Dekarboksilasi oksidatif kedua
Tahapan reaksi ini mengubah alfa-ketoglutara menjadi suksinil-CoA. Reaksi dikatalisasi oleh enzim alfa-ketoglutarat dehidrogenase.
5. Fosforilasi tingkat substrat
Respirasi seluler juga menghasilkan ATP dari tahapan ini. Reaksi pembentukan ATP inilah yang dinamakan dengan fosforilasi, karena satu gugus posfat akan ditambahkan ke ADP menjadi ATP. Pada awalnya, suksinil-CoA akan diubah menjadi suksinat, dengan mengubah GDP + Pi menjadi GTP. GTP tersebut akan digunakan untuk membentuk ATP.
6. Dehidrogenasi
Suksinat yang dihasilkan dari proses sebelumnya akan didehidrogenasi menjadi fumarat dengan bantuan enzim suksinat dehidrogenase.
7. Hidrasi dan regenerasi oksaloasetat
Dua tahapan ini merupakan akhir dari Siklus Krebs. Hidrasi merupakan penambahan atom hidrogen pada ikatan ganda karbon (C=C) yang ada pada fumarat sehingga menghasilkan malat. Malat dehidrogenase mengubah malat menjadi oksaloasetat. Oksaloasetat yang dihasilkan berfungsi untuk menangkap asetil-CoA, sehingga siklus Krebs akan terus berlangsung. Adapun hasil dari Siklus Krebs adalah ATP, FADH2, NADH dan CO2. Siklus akan menghasilkan 2 molekul CO2, yang dilepaskan. Jumlah molekul NADH yang dihasilkan adalah 6 molekul, sedangkan FADH adalah 2 molekul. ATP yang diproduksi secara langsung ada sebanyak 2 molekul, yang merupakan hasil dari reaksi fosforilasi tingkat substrat. FADH2 dan NADH adalah molekul yang digunakan dalam tahapan transpor elektron. Setiap molekul NADH akan dioksidasi lewat transpor elektron sehingga menghasilkan 3 ATP per molekul, sedangkan satu molekul FADH2 menghasilkan 2 molekul ATP
6. Sisa ditimbun sebagai lemakGlukosa yang tidak terpakai akan disimpan sebagai lemak sebagai cadangan energy.
7. Glikogenolisis

Glikogenolisis adalah lintasan metabolisme yang digunakan oleh tubuh, selain glukoneogenosis, untuk menjaga keseimbangan kadar glukosa di dalam plasma darah untuk menghindari simtoma hipoglisemia. Pada glikogenolisis, glikogen digradasi berturut-turut dengan 3 enzim, glikogen fosforilase, glukosidase, fosfoglukomutase, menjadi glukosa. Hormon yang berperan pada lintasan ini adalah glukagon dan adrenalin.
Tahap pertama penguraian glikogen adalah pembentukan glukosa 1-fosfat. Berbeda dengan reaksi pembentukan glikogen, reaksi ini tidak melibatkan UDP-glukosa, dan enzimnya adalah glikogen fosforilase. Selanjutnya glukosa 1-fosfat diubah menjadi glukosa 6-fosfat oleh enzim yang sama seperti pada reaksi kebalikannya (glikogenesis) yaitu fosfoglukomutase.
Tahap reaksi berikutnya adalah pembentukan glukosa dari glukosa 6-fosfat. Berbeda dengan
reaksi kebalikannya dengan glukokinase, dalam reaksi ini enzim lain, glukosa 6-fosfatase, melepaskan gugus fosfat sehigga terbentuk glukosa. Reaksi ini tidak menghasilkan ATP dari ADP dan fosfat.
Glukosa yang terbentuk inilah nantinya akan digunakan oleh sel untuk respirasi sehingga menghasilkan energi, yang energi itu terekam / tersimpan dalam bentuk ATP.
8. GLOKONEOGENESIS
Pada dasarnya glukoneogenesis adalah sintesis glukosa dari senyawa bukan karbohidrat, misalnya asam laktat dan beberapa asam amino. Proses glukoneogenesis berlangsung terutama dalam hati. Asam laktat yang terjadi pada proses glikolisis dapat dibawa oleh darah ke hati. Di sini asam laktat diubah menjadi glukosa kembali melalui serangkaian reaksi dalam suatu proses yaitu glukoneogenesis (pembentukan gula baru).
Glukoneogenesis yang dilakukan oleh hati atau ginjal, menyediakan suplai glukosa yang tetap. Kebanyakan karbon yang digunakan untuk sintesis glukosa akhirnya berasal dari katabolisme asam amino. Laktat yang dihasilkan dalam sel darah merah dan otot dalam keadaan

anaerobik juga dapat berperan sebagai substrat untuk glukoneogenesis. Glukoneogenesis mempunyai banyak enzim yang sama dengan glikolisis, tetapi demi alasan termodinamika dan pengaturan, glukoneogenesis bukan kebalikan dari proses glikolisis karena ada tiga tahap reaksi dalam glikolisis yang tidak reversibel, artinya diperlukan enzim lain untuk reaksi kebalikannya.
enzim glukokinaseGlukosa + ATP Glukosa-6-fosfat + ADP
enzim fosfofruktokinase Fruktosa-6-fosfat + ATP fruktosa-1,6-difosfat + ADP
enzim piruvatkinaseFosfenol piruvat + ADP asam piruvat + ATP
Enzim glikolitik yang terdiri dari glukokinase, fosfofruktokinase, dan piruvatkinase mengkatalisis reaksi yang ireversibel sehingga tidak dapat digunakan untuk sintesis glukosa. Dengan adanya tiga tahap reaksi yang tidak reversibel tersebut, maka proses glukoneogenesis berlangsung melalui tahap reaksi lain. Reaksi tahap pertama glukoneogenesis merupakan suatu reaksi kompleks yang melibatkan beberapa enzim dan organel sel (mitokondrion), yang diperlukan untuk mengubah piruvat menjadi malat sebelum terbentuk fosfoenol piruvat.
Tiga reaksi pengganti yang pertama mengubah piruvat menjadi fosfoenolpiruvat (PEP), jadi membalik reaksi yang dikatalisis oleh piruvat kinase. Perubahan ini dilakukan dalam 4 langkah. Pertama, piruvat mitokondria mengalami dekarboksilasi membentuk oksaloasetat. Reaksi ini memerlukan ATP (adenosin trifosfat) dan dikatalisis oleh piruvat karboksilase. Seperti banyak enzim lainnya yang melakukan reaksi fiksasi CO2, pada reaksi ini memerlukan biotin untuk aktivitasnya. Oksaloasetat direduksi menjadi malat oleh malat dehidrogenase mitokondria. Pada reaksi ini, glukoneogenesis secara singkat mengalami overlap (tumpang tindih) dengan siklus asam sitrat. Malat meninggalkan mitokondria dan dalam sitoplasma dioksidasi membentuk kembali oksaloasetat. Kemudian oksaloasetat sitoplasma mengalami dekarboksilasi membentuk PEP pada reaksi yang tidak memerlukan GTP (guanosin trifosfat) yang dikatalisis oleh PEP karboksikinase.
Reaksi pengganti kedua dan ketiga dikatalisis oleh fosfatase. Fruktosa-1,6-bisfosfatase mengubah fruktosa-1,6-bisfosfat menjadi fruktosa-6-fosfat, jadi membalik reaksi yang dikatalisis oleh fosfofruktokinase. Glukosa-6-fosfatase yang ditemukan pada permulaan metabolisme glikogen, mengkatalisis reaksi terakhir glukoneogenesis dan mengubah glukosa-6-fosfat menjadi glukosa bebas.
Dengan penggantian reaksi-reaksi pada glikolisis yang secara termodinamika ireversibel, glukoneogenesis secara termodinamika seluruhnya menguntungkan dan diubah dari lintasan yang menghasilkan energi menjadi lintasan yang memerlukan energi. Dua fosfat berenergi tinggi

digunakan untuk mengubah piruvat menjadi PEP. ATP tambahan digunakan untuk melakukan fosforilasi 3-fosfogliserat menjadi 1,3-bisfosfogliserat. Diperlukan satu NADH pada perubahan 1,3-bisfosfogliserat menjadi gliseraldehida-3-fosfat. Karena 2 molekul piruvat digunakan pada sintesis satu glukosa, maka setiap molekul glukosa yang disintesis dalam glukoneogenesis, sel memerlukan 6 ATP dan 2 NADH. Glikolisis dan glukoneogenesis tidak dapat bekerja pada saat yang sama. Oleh karena itu, ATP dan NADH yang diperlukan pada glukoneogenesis harus berasal dari oksidasi bahan bakar lain, terutama asam lemak.
Walaupun lemak menyediakan sebagian besar energi untuk glukoneogenesis, tetapi lemak hanya menyumbangkan sedikit fraksi atom karbon yang digunakan sebagai substrat. Ini sebagai akibat struktur siklus asam sitrat. Asam lemak yang paling banyak pada manusia yaitu asam lemak dengan jumlah atom karbon genap didegradasi oleh enzim -oksidasi menjadi asetil-KoA. Asetil KoA menyumbangkan fragmen 2-karbon ke siklus asam sitrat, tetapi pada permulaan siklus 2 karbon hilang sebagai CO2. Jadi, metabolisme asetil KoA tidak mengakibatkan peningkatan jumlah oksaloasetat yang tersedia untuk glukoneogenesis. Bila oksaloasetat dihilangkan dari siklus dan tidak diganti, kapasitas pembentukan ATP dari sel akan segera membahayakan. Siklus asam sitrat tidak terganggu selama glukoneogenesis karena oksaloasetat dibentuk dari piruvat melalui reaksi piruvat karboksilase.Kebanyakan atom karbon yang digunakan pada sintesis glukosa disediakan oleh katabolisme asam amino. Beberapa asam amino yang umum ditemukan mengalami degradasi menjadi piruvat. Oleh karena itu masuk ke proses glukoneogenesis melalui reaksi piruvat karboksilase. Asam amino lainnya diubah menjadi zat antara 4 atau 5 karbon dari siklus asam sitrat sehingga dapat membantu meningkatkan kandungan oksaloasetat dan malat mitokondria. Dari 20 asam amino yang sering ditemukan dalam protein, hanya leusin dan lisin yang seluruhnya didegradasi menjadi asetil-KoA yang menyebabkan tidak dapat menyediakan substrat untuk glukoneogenesis.
Dalam pengaturan glukoneogenesis hati dapat membuat glukosa melalui glukoneogenesis dan menggunakan glukosa melalui glikolisis sehingga harus ada suatu sistem pengaturan yang mencegah agar kedua lintasan ini bekerja serentak.Sistem pengaturan juga harus menjamin bahwa aktivitas metabolik hati sesuai dengan status gizi tubuh yaitu pembentukan glukosa selama puasa dan menggunakan glukosa saat glukosa banyak. Aktivitas glukoneogenesis dan glikolisis diatur secara terkoordinasi dengan cara perubahan jumlah relatif glukagon dan insulin dalam sirkulasi.
`Bila kadar glukosa dan insulin darah turun, asam lemak dimobilisasi dari cadangan jaringan adipose dan aktivitas -oksidasi dalam hati meningkat. Hal ini mengakibatkan peningkatan konsentrasi asam lemak dan asetil-KoA dalam hati. Karena asam amino secara serentak dimobilisasi dari otot, maka juga terjadi peningkatan kadar asam amino terutama alanin. Asam amino hati diubah menjadi piruvat dan substrat lain glukoneogenesis. Peningkatan kadar asam lemak, alanin, dan asetil-KoA semuanya memegang peranan mengarahkan substrat masuk ke glukoneogenesis dan mencegah penggunaannya oleh siklus asam sitrat. Asetil-KoA secara alosterik mengaktifkan piruvat karboksilase dan menghambat piruvat dehidrogenase. Oleh

karena itu, menjamin bahwa piruvat akan diubah menjadi oksaloasetat. Piruvat kinase dihambat oleh asam lemak dan alanin, jadi menghambat pemecahan PEP yang baru terbentuk menjadi piruvat.
Pengaturan hormonal fosfofruktokinase dan fruktosa-1,6-bisfosfatase diperantarai oleh senyawa yang baru ditemukan yaitu fruktosa 2,6-bisfosfat. Pembentukan dan pemecahan senyawa pengatur ini dikatalisis oleh enzim-enzim yang diatur oleh fosforilasi dan defosforilasi. Perubahan konsentrasi fruktosa-2,6-bisfosfat sejajar dengan perubahan untuk glukosa dan insulin yaitu konsentrasinya meningkat bila glukosa banyak dan berkurang bila glukosa langka. Fruktosa-2,6- bisfosfat secara alosterik mengaktifkan fosfofruktokinase dan menghambat fruktosa 1,6-bisfosfatase. Jadi, bila glukosa banyak maka glikolisis aktif dan glukoneogenesis dihambat. Bila kadar glukosa turun, peningkaan glukagon mengakibatkan penurunan konsentrasi fruktosa-2,6-bisfosfat dan penghambatan yang sederajat pada glikolisis dan pengaktifan glukoneogenesis.
Daftar Pustaka
Poedjiadi,A.2005.Dasar-dasarBiokimia.UI-Press,Jakarta. Wirahadikusumah,M.1985.Biokimia:metabolisme energi, karbohidrat, dan lipid. Penerbit ITB, Bandung.