
Pertambahan bobot badan, status fisiologis, komposisi ... · Barbados Cross (BC) atau domba...
25
TINJAUAN PUSTAKA Domba Barbados Domba barbados berasal dari Kepulauan Karibia, tepatnya di Pulau Barbados, yang merupakan hasil persilangan antara domba afrika dengan domba daerah dingin (Suparyanto 1999). Domba ini merupakan domba tipe rambut dengan tujuan produksi daging, bobot badan dewasa betina 35-50 kg dan jantan 50-80 kg dan pertambahan bobot badan 104-109 g/hari (Rastogi 2001). Pada kondisi pakan yang baik, domba barbados beranak pertama kali pada umur 12-13 bulan, sedangkan pada kondisi pakan yang kurang baik pertama kali beranak dicapai pada umur 14-15 bulan, dengan frekuensi kelahiran anak kembar berkisar antara 56-71% (Suparyanto 1999). Domba barbados yang disilangkan dengan domba priangan memiliki bobot lepas sapih 26.63±6.83 kg/induk, bobot lepas sapih individu untuk jantan adalah 15.09±1.97 kg/ekor dan untuk betina 12.96±1.58 kg/ekor (Rahmat et al. 2007). Domba barbados yang digunakan pada studi adalah domba barbados hasil persilangan dengan domba sumatera dengan komposisi genetik 50% domba barbados dan 50% domba sumatera. Domba ini biasa dikenal dengan domba Barbados Cross (BC) atau domba barbados silangan. Pola perkawinan domba persilangan barbados adalah disajikan pada Gambar 2. Gambar 2 Pola perkawinan domba persilangan barbados (BC) (Subandriyo et al. 1996). X Barbados Blackbelly (B) 100% Sumatera (S) 100% X Barbados Cross (BC) 50%B, 50%S Barbados Cross (BC) 50%B, 50%S Barbados Cross (BC) 50%B, 50%S
Transcript of Pertambahan bobot badan, status fisiologis, komposisi ... · Barbados Cross (BC) atau domba...

5
TINJAUAN PUSTAKA
Domba Barbados
Domba barbados berasal dari Kepulauan Karibia, tepatnya di Pulau
Barbados, yang merupakan hasil persilangan antara domba afrika dengan domba
daerah dingin (Suparyanto 1999). Domba ini merupakan domba tipe rambut
dengan tujuan produksi daging, bobot badan dewasa betina 35-50 kg dan jantan
50-80 kg dan pertambahan bobot badan 104-109 g/hari (Rastogi 2001). Pada
kondisi pakan yang baik, domba barbados beranak pertama kali pada umur 12-13
bulan, sedangkan pada kondisi pakan yang kurang baik pertama kali beranak
dicapai pada umur 14-15 bulan, dengan frekuensi kelahiran anak kembar berkisar
antara 56-71% (Suparyanto 1999). Domba barbados yang disilangkan dengan
domba priangan memiliki bobot lepas sapih 26.63±6.83 kg/induk, bobot lepas
sapih individu untuk jantan adalah 15.09±1.97 kg/ekor dan untuk betina
12.96±1.58 kg/ekor (Rahmat et al. 2007).
Domba barbados yang digunakan pada studi adalah domba barbados hasil
persilangan dengan domba sumatera dengan komposisi genetik 50% domba
barbados dan 50% domba sumatera. Domba ini biasa dikenal dengan domba
Barbados Cross (BC) atau domba barbados silangan. Pola perkawinan domba
persilangan barbados adalah disajikan pada Gambar 2.
Gambar 2 Pola perkawinan domba persilangan barbados (BC) (Subandriyo et al.
1996).
X
Barbados Blackbelly (B)
100%
Sumatera (S)
100%
X
Barbados Cross (BC)
50%B, 50%S
Barbados Cross (BC)
50%B, 50%S
Barbados Cross (BC)
50%B, 50%S

6
(a) (b)
Gambar 3 (a) Domba barbados peranakan penelitian (BC); (b) Domba barbados
asli di Pulau Barbados
Domba Komposit Sumatera
Domba komposit sumatera atau domba sei putih adalah salah satu domba
bentukan baru dari Balai Penelitian Ternak, Badan Litbang Pertanian, Ciawi-
Bogor yang memiliki beberapa keunggulan. Keunggulan tersebut ialah dapat
dikawinkan sepanjang tahun dan beradaptasi dengan baik pada pakan sederhana.
Domba komposit sumatera adalah domba unggul hasil persilangan antara bibit
domba lokal Sumatera dengan bibit domba St. Croix asal Virgin Islands, Amerika
Serikat dan domba barbados blackbelly asal Bardados Islands. Penelitian
pembentukan domba komposit sumatera telah berlangsung dua dekade lebih
hingga menghasilkan domba yang dapat dikembangkan dalam kondisi
pemeliharaan semi intensif (BPATP 2008).
Gambar 4 Domba komposit sumatera penelitian di kandang individu
7
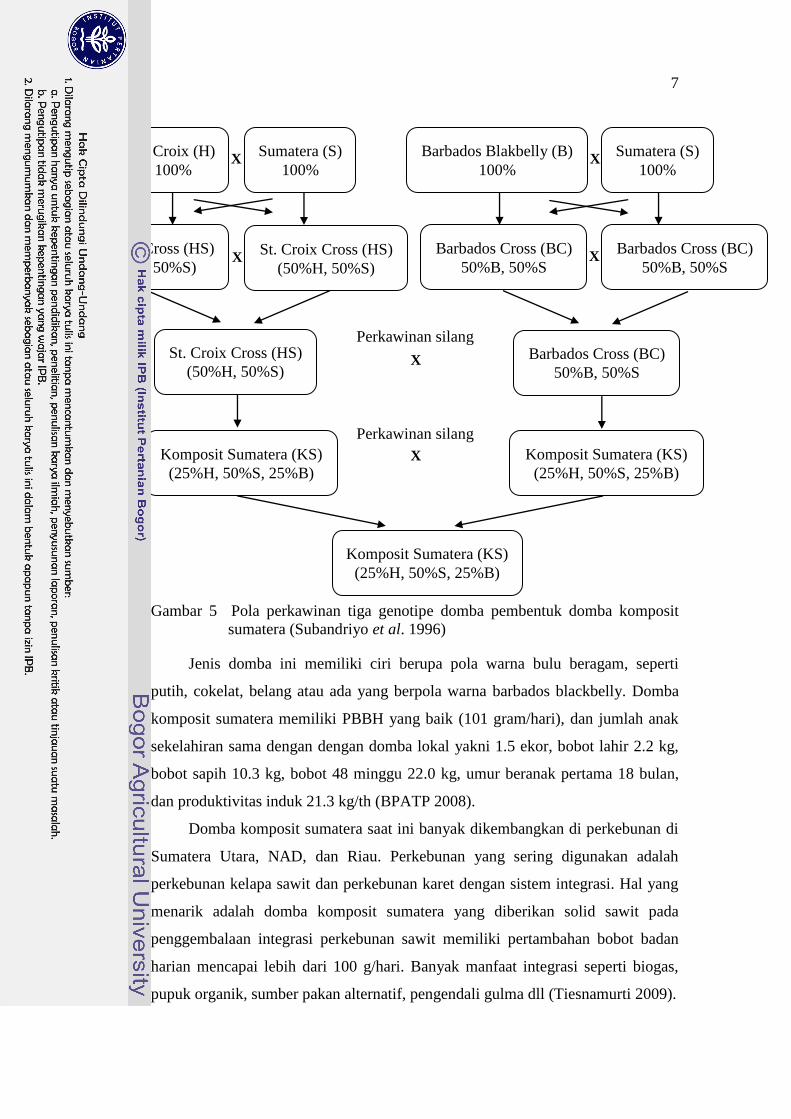
Gambar 5 Pola perkawinan tiga genotipe domba pembentuk domba komposit
sumatera (Subandriyo et al. 1996)
Jenis domba ini memiliki ciri berupa pola warna bulu beragam, seperti
putih, cokelat, belang atau ada yang berpola warna barbados blackbelly. Domba
komposit sumatera memiliki PBBH yang baik (101 gram/hari), dan jumlah anak
sekelahiran sama dengan dengan domba lokal yakni 1.5 ekor, bobot lahir 2.2 kg,
bobot sapih 10.3 kg, bobot 48 minggu 22.0 kg, umur beranak pertama 18 bulan,
dan produktivitas induk 21.3 kg/th (BPATP 2008).
Domba komposit sumatera saat ini banyak dikembangkan di perkebunan di
Sumatera Utara, NAD, dan Riau. Perkebunan yang sering digunakan adalah
perkebunan kelapa sawit dan perkebunan karet dengan sistem integrasi. Hal yang
menarik adalah domba komposit sumatera yang diberikan solid sawit pada
penggembalaan integrasi perkebunan sawit memiliki pertambahan bobot badan
harian mencapai lebih dari 100 g/hari. Banyak manfaat integrasi seperti biogas,
pupuk organik, sumber pakan alternatif, pengendali gulma dll (Tiesnamurti 2009).
St. Croix (H)
100%
Sumatera (S)
100%
Barbados Blakbelly (B)
100%
Sumatera (S)
100% X X
St. Croix Cross (HS)
(50%H, 50%S) St. Croix Cross (HS)
(50%H, 50%S)
Barbados Cross (BC)
50%B, 50%S
Barbados Cross (BC)
50%B, 50%S X X
St. Croix Cross (HS)
(50%H, 50%S) Barbados Cross (BC)
50%B, 50%S
Komposit Sumatera (KS)
(25%H, 50%S, 25%B)
Komposit Sumatera (KS)
(25%H, 50%S, 25%B)
X
X
Komposit Sumatera (KS)
(25%H, 50%S, 25%B)
Perkawinan silang
Perkawinan silang
8
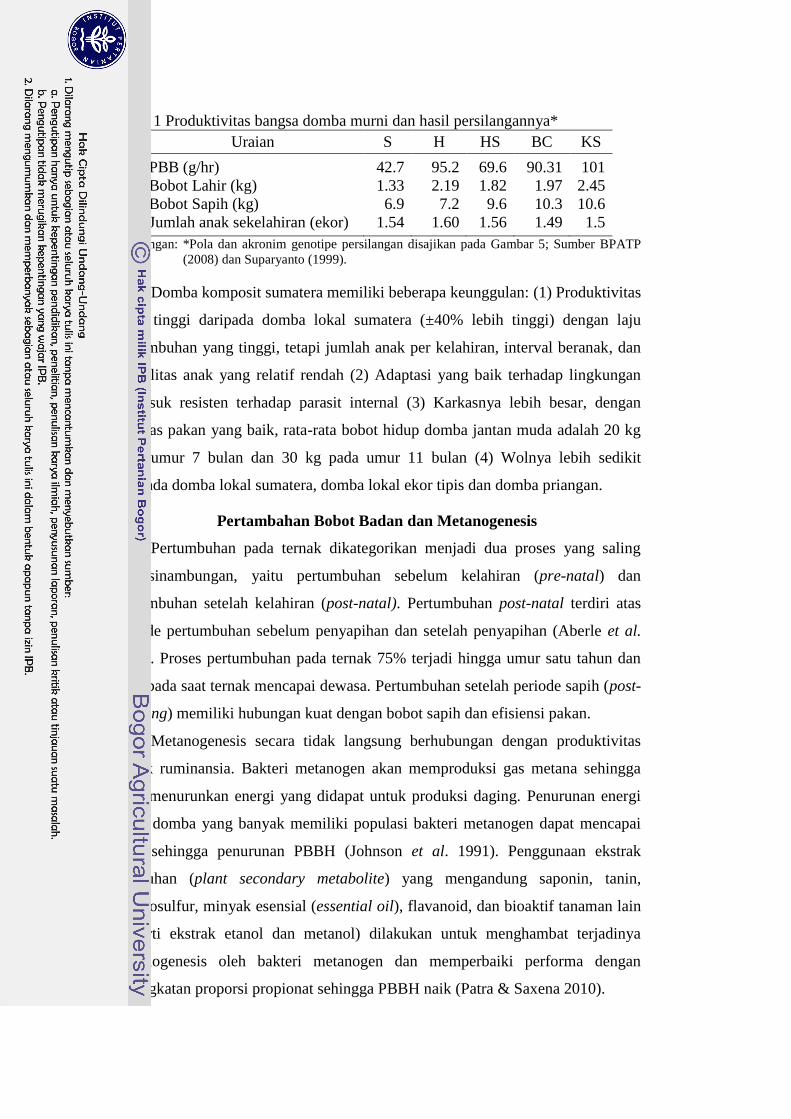
Tabel 1 Produktivitas bangsa domba murni dan hasil persilangannya*
Uraian S H HS BC KS
PBB (g/hr) 42.7 95.2
69.6
90.31
101
Bobot Lahir (kg) 1.33
2.19
1.82 1.97
2.45
Bobot Sapih (kg) 6.9
7.2
9.6
10.3
10.6
Jumlah anak sekelahiran (ekor) 1.54
1.60
1.56
1.49
1.5
Keterangan: *Pola dan akronim genotipe persilangan disajikan pada Gambar 5; Sumber BPATP
(2008) dan Suparyanto (1999).
Domba komposit sumatera memiliki beberapa keunggulan: (1) Produktivitas
lebih tinggi daripada domba lokal sumatera (±40% lebih tinggi) dengan laju
pertumbuhan yang tinggi, tetapi jumlah anak per kelahiran, interval beranak, dan
mortalitas anak yang relatif rendah (2) Adaptasi yang baik terhadap lingkungan
termasuk resisten terhadap parasit internal (3) Karkasnya lebih besar, dengan
kualitas pakan yang baik, rata-rata bobot hidup domba jantan muda adalah 20 kg
pada umur 7 bulan dan 30 kg pada umur 11 bulan (4) Wolnya lebih sedikit
daripada domba lokal sumatera, domba lokal ekor tipis dan domba priangan.
Pertambahan Bobot Badan dan Metanogenesis
Pertumbuhan pada ternak dikategorikan menjadi dua proses yang saling
berkesinambungan, yaitu pertumbuhan sebelum kelahiran (pre-natal) dan
pertumbuhan setelah kelahiran (post-natal). Pertumbuhan post-natal terdiri atas
periode pertumbuhan sebelum penyapihan dan setelah penyapihan (Aberle et al.
2001). Proses pertumbuhan pada ternak 75% terjadi hingga umur satu tahun dan
25% pada saat ternak mencapai dewasa. Pertumbuhan setelah periode sapih (post-
weaning) memiliki hubungan kuat dengan bobot sapih dan efisiensi pakan.
Metanogenesis secara tidak langsung berhubungan dengan produktivitas
ternak ruminansia. Bakteri metanogen akan memproduksi gas metana sehingga
akan menurunkan energi yang didapat untuk produksi daging. Penurunan energi
bruto domba yang banyak memiliki populasi bakteri metanogen dapat mencapai
15% sehingga penurunan PBBH (Johnson et al. 1991). Penggunaan ekstrak
tumbuhan (plant secondary metabolite) yang mengandung saponin, tanin,
organosulfur, minyak esensial (essential oil), flavanoid, dan bioaktif tanaman lain
(seperti ekstrak etanol dan metanol) dilakukan untuk menghambat terjadinya
metanogenesis oleh bakteri metanogen dan memperbaiki performa dengan
peningkatan proporsi propionat sehingga PBBH naik (Patra & Saxena 2010).

9
Komposisi Tubuh
Komposisi tubuh adalah suatu nilai yang menunjukkan proporsi dari
komponen penyusun tubuh, antara lain air, protein, dan lemak. Komposisi tubuh
sangat dipengaruhi oleh spesies, tingkat kegemukan, atau bobot tubuh temak
(Parakkasi, 1981). Komposisi tubuh domba disajikan dalam.
Komposisi tubuh yang diukur dengan teknik menginjeksikan tracer (dilution
technique) ke dalam tubuh ternak yang dilaporkan oleh beberapa peneliti disajikan
dalam Tabel 1. Data tersebut menunjukkan bahwa komposisi tubuh sangat
bervariasi bergantung pada umur dan bobot tubuh. Komposisi tubuh dapat
dipengaruhi oleh berbagai macam faktor, yaitu bobot potong, umur potong,
bangsa, dan jenis kelamin. Bobot potong merupakan peubah yang paling
berpengaruh pada komposisi tubuh (Berg & Butterfield 1976).
Soeparno (1992) menyatakan bahwa nutrisi, umur, dan bobot tubuh
merupakan faktor yang saling berhubungan erat, dan dapat secara bebas atau
secara bersama mempengaruhi komposisi tubuh ternak atau karkas. Variasi
komposisi tubuh sebagian besar didominasi oleh variasi bobot tubuh, dan
sebagian kecil dipengaruhi oleh umur dan bobot potong.
Metode Pendugaan Komposisi Tubuh
Menurut Nonaka (2002) ada dua cara untuk mengetahui komposisi tubuh
ternak yaitu cara langsung dan cara tidak langsung. (1) Cara langsung (direct
method), yaitu dengan memotong ternak dan kemudian memisahkan dan
menimbang daging dan lemaknya. Cara ini adalah yang paling akurat, namun
dalam pelaksanaannya, biaya dan tenaga kerja menjadi faktor pembatas. Ada tiga
macam cara yang termasuk dalam cara tidak langsung (indirect method), yaitu
metode bobot karkas, metode kesetimbangan energi dan protein, dan metode
injeksi. Metode bobot jenis karkas masih dilakukan dengan memotong ternak dan
kemudian diceburkan ke dalam kolam untuk kemudian dihitung bobot jenisnya.
Bobot jenis inilah yang kemudian digunakan untuk menentukan perbandingan
komposisi daging dan lemak ternak. Cara ini banyak digunakan pada ternak babi
dan mempunyai akurasi yang sangat tinggi. Kendala cara ini adalah biaya tinggi,
ternak tidak dapat digunakan berulang kali (sebuah cara yang biasa digunakan
untuk mengetahui perbedaan komposisi tubuh berdasarkan fase pertumbuhannya).

10
Pada metode kesetimbangan energi dan nitrogen (protein) kita dapat mengetahui
pembongkaran protein atau lemak tubuh serta dapat menghitung deposisi yang
terjadi. Keuntungan metode ini adalah ternak dapat digunakan berulang kali,
murah, akan tetapi diperlukan banyak tenaga dalam pengerjaannya terutama
dalam pengumpulan feses dan urine, sehingga sulit dilaksanakan pada jumlah
ternak yang besar. Pada metode menginjeksikan (tracer dilution technique) ke
dalam tubuh ternak, konsentrasi tracer akan berkurang karena beredar (melalui
darah) ke seluruh tubuh sesuai dengan bobot dan komposisi tubuhnya. Teknik ini
yang paling sederhana, karena hanya menggunakan sampel darah, sedikit tenaga
kerja, penggunaan ternak dapat dalam jumlah besar dan berulang kali, biaya jauh
lebih rendah dan mempunyai akurasi yang cukup tinggi.
Teknik penyuntikan tracer ini dikembangkan berdasarkan pemikiran bahwa
proporsi protein konstan, sementara lemak dan air tubuh berhubungan terbalik.
Dari pemikiran ini maka apabila air tubuh dapat diketahui maka komposisi lemak
dan daging dapat ditentukan. Tracer yang sering digunakan dalam metode ini ada
tiga, yaitu tritium, deuterium, dan urea. Dari ketiga traser tersebut, traser urea
sangat mudah didapat, mudah, dan analisisnya hanya membutuhkan alat
spektrofotometer. Apabila gagal dapat segera diulang karena dalam waktu sekitar
dua hari pengaruh urea yang disuntikkan akan menghilang. Urea yang disuntikkan
akan memasuki pool tubuh, oleh karena akan terjadi pelarutan (pemerataan) urea
dalam tubuh dan terjadi perbedaan antara urea sebelum dan sesudah penyuntikan.
Berdasarkan basil penelitian Astuti dan Sastradipradja (1999) pendugaan
kandungan air, lemak, dan protein tubuh domba priangan dengan teknik "urea
space" menunjukkan tingkat akurasi yang tinggi dengan r masing-masing 0.95;
0.98 dan 0.96 sedangkan hasil penelitian Panaretto dan Till (1963) diperoleh r
masing-masing 0.99; 0.93 dan 0.98.
Panas Tubuh dan Konsumsi Nutrien
Paparan panas suhu lingkungan akan mengakibatkan penurunan bobot
badan, penurunan PBBH, laju pertumbuhan, dan bentuk fisik tubuh (Marai et al.
2000). Selain itu, paparan suhu lingkungan yang tinggi menyebabkan peningkatan
laju respirasi, suhu tubuh, konsumsi air dan penurunan konsumsi bahan kering
(Marai et al. 2007). Studi oleh Monty et al. (1991), Nardon et al. (1991) bahwa

11
domba st. croix, karakul, dan rambouillet mengalami penurunan konsumsi bahan
kering ketika mengalami stres panas. Apabila konsumsi bahan kering menurun
oleh stres panas, konsumsi protein kasar (PK), energi bruto (GE) dan energi
metabolis (EM) juga akan menurun (Marai et al. 2001).
Fisiologis Pertumbuhan Domba
Perubahan fungsi kerja biologi domba yang mengalami cekaman panas
lingkungan dan peningkaatan suhu tubuh dapat disebabkan oleh: (1) penurunan
konsumsi dan kecernaan pakan; (2) gangguan metabolisme pada air tubuh, energi,
dan keseimbangan mineral; (3) reaksi enzimatis, sekresi hormon, dan metabolit
darah (Marai et al. 2007).
Termoregulasi
Termoregulasi domba erat kaitanya dengan evaporasi dan disipasi panas
karena peran keringat berkurang dengan adanya lapisan bulu-bulu wol (Marai et
al. 2007). Termoregulasi adalah pengaturan suhu tubuh yang bergantung pada
produksi panas melalui metabolisme dan pelepasan panas tersebut ke lingkungan
(Esmay 1982). Panas adalah sebuah bentuk energi yang ditransmisikan dari suatu
tubuh ke yang lainnya karena adanya perbedaan temperatur. Temperatur mengacu
pada kemampuan tubuh untuk menyerap panas. Energi didefinisikan sebagai
kapasitas untuk melakukan kerja (Esmay 1982). Menurut Etgen (1987), energi
dibutuhkan untuk mendukung fungsi normal tubuh ternak seperti respirasi,
pencernaan, dan metabolisme untuk pertumbuhan dan produksi susu. Pada hewan
yang lebih aktif, lebih banyak energi yang dikeluarkan untuk mendukung
aktivitasnya dan faktor intrinsik yang paling besar mempengaruhi metabolisme
adalah sumber pakan (Scheer 1963).
Suhu Rektal
Suhu tubuh menunjukkan kemampuan tubuh untuk melepas dan menerima
panas (Esmay,1982). Pengukuran suhu tubuh pada dasarnya sulit dilakukan,
karena pengukuran suhu tubuh merupakan resultan dari berbagai pengukuran di
berbagai tempat (Schmidt-Nielsen 1997). Suhu tubuh atau suhu inti (core
temperature) dapat dihitung pada beberapa lokasi. Lokasi yang biasa digunakan
adalah rektum, karena cukup mewakili dan kondisinya stabil. Suhu inti

12
mendominasi penentuan suhu tubuh. Temperatur rektum dan kulit saat siang hari
meningkat akibat dehidrasi, dan frekuensi respirasi dan temperatur tubuh
berfluktuasi lebih besar saat dehidrasi. Menurut Kelly (1984), suhu tubuh yang
diukur dengan termometer klinis bukan indikasi dari jumlah total yang diproduksi,
tetapi hanya merefleksikan keseimbangan antara suhu yang diproduksi dengan
suhu yang dilepaskan. Suhu rektum sering digunakan sebagai ukuran representatif
suhu tubuh (Marai et al. 2007). Suhu rektum domba pada zona nyaman adalah
38.3-39.9°C (Marai et al. 2007). Zona nyaman (thermoneutral zone) pada domba
adalah 22-31°C untuk beraktivitas dan reproduksi (Yousef 1985).
Denyut Jantung
Laju denyut jantung merupakan refleksi utama dari proses homeostatis
sirkulasi darah sepanjang status metabolisme yang umum (Marai et al. 2007).
Faktor fisiologis yang mempengaruhi denyut jantung pada hewan normal adalah
spesies, ukuran tubuh, umur, kondisi fisik, jenis kelamin, rangsangan, tahap
laktasi, rangsangan, posisi tubuh, aktivitas sistem pencernaan, ruminasi, dan
temperatur lingkungan (Frandson 1992). Menurut Schmidt-Nielsen (1997),
jantung memiliki suatu kapasitas yang kompleks untuk berkontraksi tanpa
stimulus eksternal. Denyut jantung domba normal menurut Duke’s (1995) adalah
60-120 denyut per menit. Cara untuk mendeteksi denyut jantung adalah dengan
meraba arteri menggunakan jari hingga denyutan terasa atau pada bagian dada
kiri atas (dekat lengan) dekat tulang axilla sebelah kiri dengan menggunakan
stetoskop. Selama musim panas, laju denyut jantung signifikan lebih tinggi
dibandingkan dengan musim dingin pada domba Barki di Timur Tengah (Ismail et
al. 1995). Laju denyut jantung pada waktu pagi hari (pukul 8.00) akan lebih
rendah dibandingkan dengan pada siang hari pukul (15.00) (Marai et al. 2007).
Respirasi
Dua fungsi utama sistem respirasi adalah menyediakan oksigen untuk darah,
dan mengambil karbondioksida dari dalam darah. Fungsi-fungsi yang bersifat
sekunder meliputi membantu dalam regulasi keasaman cairan ekstraseluler dalam
tubuh, membantu pengendalian suhu, eliminasi air dan pembentukan suara. Laju
respirasi dapat menjadi indikator terjadinya stres panas (Habeeb et al. 1992).

13
Domba akan melepaskan 20% panas tubuh melalui respirasi pada suhu 12°C dan
meningkat menjadi 60% dari total panas tubuh pada suhu 35°C (Thompson 1985).
Aktivitas respirasi ditandai dengan pergerakan tulang rusuk, tulang dada,
dan perut (merespons kontraksi paru-paru dan pergerakan diafragma), observasi
aktivitas respirasi lebih diutamakan saat ternak dalam posisi berdiri, karena posisi
berbaring akan mempengaruhi respirasi terlebih lagi pada ternak yang sedang
sakit. Pengontrolan frekuensi respirasi dengan cara berdiri pada salah satu sisi
ternak, lalu mengamati daerah dada dan perut, disarankan untuk mengobservasi
ternak dari kedua sisi, untuk mengetahui similaritas pergerakan kedua sisi
(Kelly1984).
Menurut Hecker (1983), laju respirasi untuk domba normal adalah 15-40
helaan/menit atau 30-80 respirasi/menit, sedangkan menurut Frandson (1992)
adalah 26-32 helaan/menit = 52-64 respirasi/menit. Mekanisme respirasi dikontrol
di medula yang sensitif terhadap CO2 dan tekanan darah. Jika tekanan meningkat
sedikit, pernapasan menjadi lebih dalam dan cepat (Esmay 1982). Peningkatan
frekuensi respirasi terjadi ketika ada peningkatan permintaan oksigen, yaitu ketika
terpapar ke suhu lingkungan dan kelembapan relatif yang tinggi (Kelly1984).
Konsumsi pakan berdasarkan jenis nutrisi akan mempengaruhi laju respirasi
pada kondisi stres panas (Bluett et al. 2001). Laju respirasi pada ternak yang
terpapar suhu lingkungan yang tinggi dapat mencapai 400 respirasi/menit dan
ketika terjadi penurunan menuju suhu yang rendah maka laju respirasi menjadi
lebih lama durasinya pada 155-200 respirasi/menit hingga menuju laju respirasi
normal dan nyaman (Marai et al. 2007). Laju respirasi akan meningkat pada siang
hari (15.00; 12.00; dan 16.00) dibandingkan dengan waktu pagi hari (8.00) pada
musim panas terhadap bangsa domba Mesir (Marai et al. 2008).
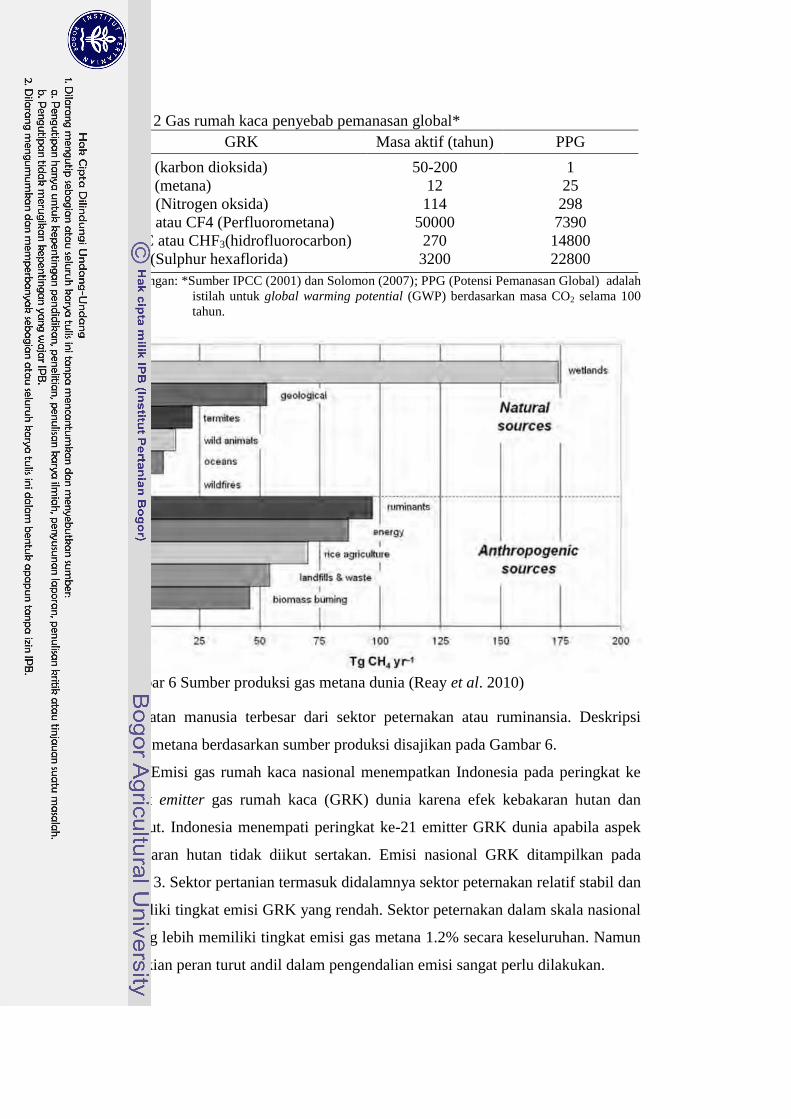
Emisi Gas Metana
Gas metana (CH4) merupakan salah satu gas yang menyebabkan pengaruh
gas rumah kaca atau GRK (CO2, CH4, N2O, PFC, HFC, dan SF6) sehingga terjadi
pemanasan global (Vlaming 2008). Gas metana memiliki potensi pemanasan
global (PPG) 25 kali lipat dibandingkan dengan gas CO2 (Tabel 2). Sumber emisi
gas metana berasal dari alam (natural resources), manusia (anthropogenic
resources), dan sinks (Reay et al. 2010). Sumber emisi metana yang berasal dari
14
Tabel 2 Gas rumah kaca penyebab pemanasan global*
GRK Masa aktif (tahun) PPG
CO2 (karbon dioksida) 50-200 1
CH4 (metana) 12 25
N2O (Nitrogen oksida) 114 298
PFC atau CF4 (Perfluorometana) 50000 7390
HFC atau CHF3(hidrofluorocarbon) 270 14800
SF6 (Sulphur hexaflorida) 3200 22800
Keterangan: *Sumber IPCC (2001) dan Solomon (2007); PPG (Potensi Pemanasan Global) adalah
istilah untuk global warming potential (GWP) berdasarkan masa CO2 selama 100
tahun.
Gambar 6 Sumber produksi gas metana dunia (Reay et al. 2010)
perbuatan manusia terbesar dari sektor peternakan atau ruminansia. Deskripsi
emisi metana berdasarkan sumber produksi disajikan pada Gambar 6.
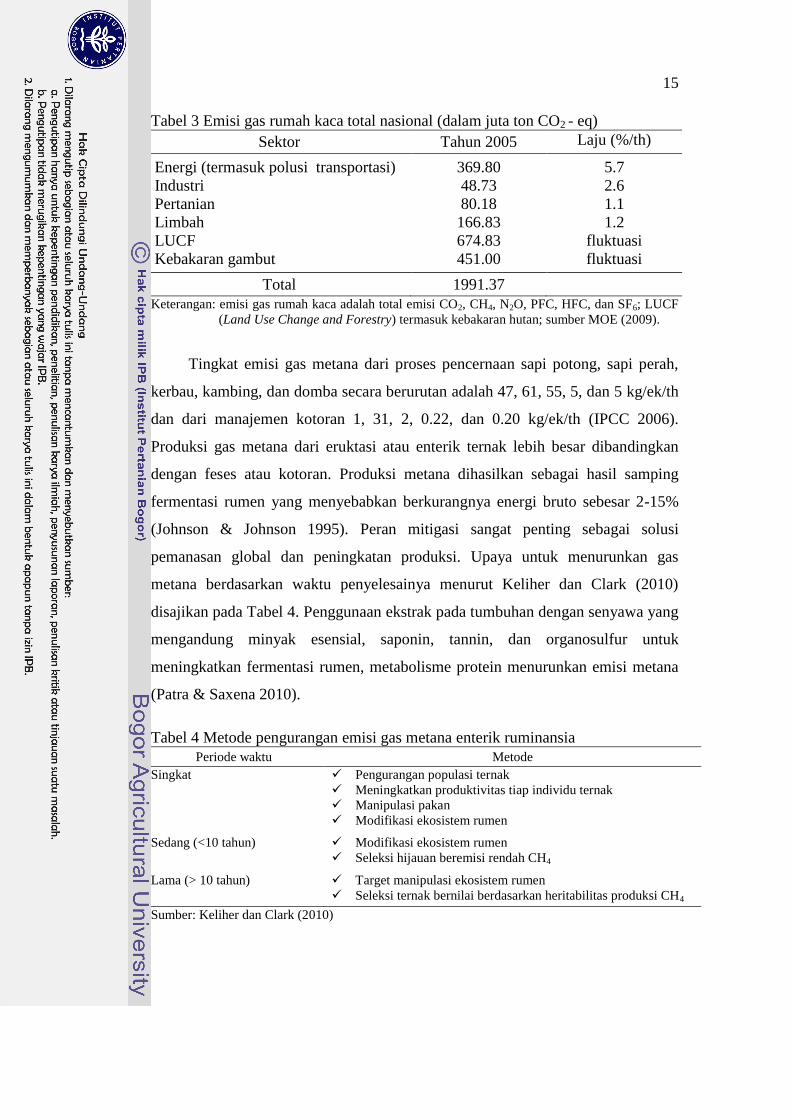
Emisi gas rumah kaca nasional menempatkan Indonesia pada peringkat ke
empat emitter gas rumah kaca (GRK) dunia karena efek kebakaran hutan dan
gambut. Indonesia menempati peringkat ke-21 emitter GRK dunia apabila aspek
kebakaran hutan tidak diikut sertakan. Emisi nasional GRK ditampilkan pada
Tabel 3. Sektor pertanian termasuk didalamnya sektor peternakan relatif stabil dan
memiliki tingkat emisi GRK yang rendah. Sektor peternakan dalam skala nasional
kurang lebih memiliki tingkat emisi gas metana 1.2% secara keseluruhan. Namun
demikian peran turut andil dalam pengendalian emisi sangat perlu dilakukan.
15
Tabel 3 Emisi gas rumah kaca total nasional (dalam juta ton CO2 - eq)
Sektor Tahun 2005 Laju (%/th)
Energi (termasuk polusi transportasi) 369.80 5.7
Industri 48.73 2.6
Pertanian 80.18 1.1
Limbah 166.83 1.2
LUCF 674.83 fluktuasi
Kebakaran gambut 451.00 fluktuasi
Total 1991.37
Keterangan: emisi gas rumah kaca adalah total emisi CO2, CH4, N2O, PFC, HFC, dan SF6; LUCF
(Land Use Change and Forestry) termasuk kebakaran hutan; sumber MOE (2009).
Tingkat emisi gas metana dari proses pencernaan sapi potong, sapi perah,
kerbau, kambing, dan domba secara berurutan adalah 47, 61, 55, 5, dan 5 kg/ek/th
dan dari manajemen kotoran 1, 31, 2, 0.22, dan 0.20 kg/ek/th (IPCC 2006).
Produksi gas metana dari eruktasi atau enterik ternak lebih besar dibandingkan
dengan feses atau kotoran. Produksi metana dihasilkan sebagai hasil samping
fermentasi rumen yang menyebabkan berkurangnya energi bruto sebesar 2-15%
(Johnson & Johnson 1995). Peran mitigasi sangat penting sebagai solusi
pemanasan global dan peningkatan produksi. Upaya untuk menurunkan gas
metana berdasarkan waktu penyelesainya menurut Keliher dan Clark (2010)
disajikan pada Tabel 4. Penggunaan ekstrak pada tumbuhan dengan senyawa yang
mengandung minyak esensial, saponin, tannin, dan organosulfur untuk
meningkatkan fermentasi rumen, metabolisme protein menurunkan emisi metana
(Patra & Saxena 2010).
Tabel 4 Metode pengurangan emisi gas metana enterik ruminansia
Periode waktu Metode
Singkat Pengurangan populasi ternak
Meningkatkan produktivitas tiap individu ternak
Manipulasi pakan
Modifikasi ekosistem rumen
Sedang (<10 tahun) Modifikasi ekosistem rumen
Seleksi hijauan beremisi rendah CH4
Lama (> 10 tahun) Target manipulasi ekosistem rumen
Seleksi ternak bernilai berdasarkan heritabilitas produksi CH4
Sumber: Keliher dan Clark (2010)

16
Gas metana adalah salah satu jenis gas rumah kaca utama yang berpengaruh
pada pemanasan global. Kontribusi CH4 terhadap pemanasan global diperkirakan
sebesar 18%, nomor dua setelah CO2 (49%), kemudian CFC 14%, N2O 6%, dan
lainnya 13%. Meskipun jumlahnya di atmosfer jauh lebih rendah dibandingkan
dengan gas CO2, namun gas metana memiliki kemampuan menyerap panas (radiasi
infra merah) yang dipancarkan oleh bumi 21 kali lebih tinggi dari CO2. Gas
metana memiliki waktu tinggal di atmosfer lebih singkat yaitu selama 12 tahun,
sedangkan CO2 selama 100 tahun. Gas metana mampu menembus sampai lapisan
ionosfer di mana terdapat senyawa radikal O3 (ozon) yang berfungsi sebagai
pelindung bumi dari serangan radiasi gelombang pendek ultra violet. Kehadiran
gas metana pada lapisan ini berpengaruh pada reaksi oksidasi yang menghasilkan
CO2, sehingga menurunkan kandungan O3. Jadi, metana adalah salah satu gas
yang yang menyebabkan penipisan ozon bumi (IPCC 1994).
Keseimbangan metana di atmosfer diatur melalui keseimbangan antara
sumber dan saluran pembuangannya. Saluran metana yang utama adalah atmosfer
yang mengandung ion hidrogen bebas pada lapisan troposfer. Metana bereaksi
dengan ion hidroksil (OH) membentuk uap air dan CO2. Saluran kedua adalah
tanah yang mengandung bakteri yang dapat menyerap metana dan mengoksidasi-
nya. Akan tetapi, bila konsentrasi metana melebihi hilangnya melalui kedua
saluran ini maka konsentrasinya di atmosfer menjadi meningkat dan mengakibat-
kan efek rumah kaca yang berlebihan dan akhirnya berpengaruh pada pemanasan
global (Reay et al. 2010).
Konsentrasi metana di atmosfer pada saat ini sudah mencapai lebih dari dua
kali lipat, yaitu dari 700 ppb menjadi 1785 ppb. Meningkatnya jumlah metana di
atmosfer berkaitan dengan meningkatnya populasi manusia, karena berkorelasi
dengan kebutuhan pangan yang dihasilkan oleh ternak dan padi sawah. Di
samping itu, juga disebabkan oleh meningkatnya kebutuhan energi dan perusakan
hutan serta lahan gambut. Meningkatnya metana di atmosfer akan berpengaruh
pada perubahan iklim dan suhu global yang dikenal dengan pemanasan global
(Moss 2000).
Terjadinya pemanasan global sebagai akibat dari meningkatnya gas-gas
rumah kaca di atmosfer akan berpengaruh pada lingkungan dan kehidupan di

17
bumi. Pemanasan global diperkirakan mengakibatkan bumi lebih panas 1-2oC
dibanding sekarang pada tahun 2030, dan air laut naik 5-44 cm akibat
mencairnya es di kutub. Naiknya permukaan air laut membuat daratan semakin
sempit dan terjadi krisis air bersih terutama di perkotaan akibat intrusi air laut.
Dampak lainnya adalah meningkatknya penyakit yang ditularkan melalui
nyamuk, menurunnya produktivitas pertanian akibat perubahan suhu dan pola
hujan yang tidak teratur, dan punahnya sejumlah keragaman hayati akibat
peningkatan suhu bumi. Oleh karena itu, mengurangi emisi metana sangat
penting untuk menjaga kelangsungan hidup di bumi ini (Moss et al. 2000).
Mitigasi Emisi Gas Metana Enterik
Metana enterik dari ruminansia sekitar 11-17% dari metana global atau 17-
30% dari total metana antropogenik (Beauchemin et al. 2007). Teknologi
menurunkan emisi gas metana enterik telah banyak diteliti, dikembangkan dan
diadopsi oleh masyarakat, namun masih perlu diinventarisasi dan diseleksi untuk
dapat dipilih agar dapat diterapkan pada peternakan rakyat. Mara et al. (2008)
merekomendasikan beberapa teknologi alternatif untuk menurunkan produksi
metana akibat fermentasi pakan di dalam saluran pencernaan, yaitu meningkatkan
frekuensi pemberian pakan, meningkatkan jumlah konsentrat di dalam ransum,
memilih pakan yang mengandung karbohidrat non-struktural lebih tinggi,
penggunaan legum, perbaikan manajemen padang pengembalaan, penggunaan
biji-bijian pada silase, memanfaatkan tanaman yang mengandung saponin dan
tanin sebagai pendefaunasi protozoa di dalam rumen, dan penggunaaan feed aditif.
Prinsip penggunaan feed aditif dapat dilakukan dengan berbagai macam zat
kimia dengan beberapa tipe mekanisme, antar lain berdasarkan sifat toksik
terhadap bakteri metanogen, seperti senyawa-senyawa metana terhalogenasi
(Boccazzi dan Patterson 1995), berdasarkan pada reaksi hidrogenasi, seperti
senyawa asam-asam lemak rantai panjang tak jenuh, berdasarkan senyawa-
senyawa kimia yang afinitasnya terhadap hidrogen lebih tinggi daripada CO2,
seperti ion ferri dan sulfat, dan berdasarkan defaunasi atau penekanan populasi
mikroba seperti senyawa saponin dan tanin (Thalib 2004; Beauchemin et al.
2007), dan bakteri asetogenik (Thalib 2008). Rata-rata penurunan persentase
produksi metana (pengujian in vitro) oleh feed additive berkisar 10-40%,

18
bergantung pada level pemberian dalam satuan bobot feed additive per bobot
substrat yang difermentasi, dan daya kerjanya sebagai inhibitor metanogenesis.
Beberapa jenis pakan lokal yang telah dicoba dan berpotensi untuk
mengurangi emisi gas metana pada saluran pencernaan ternak ruminansia antara
lain ekstrak buah lerak ditambah metanaol (aksapon SR), daun kembang sepatu,
daun randu, daun waru, dan leguminosa akibat adanya kandungan senyawa
saponin dan tanin dalam tanaman tersebut.
Sistem Pencernaan Domba
Domba adalah termasuk ternak ruminansia yang proses pencernaannya
relatif lebih kompleks bila dibandingkan dengan proses pencernaan pada ternak
nonruminansia. Sistem pencernaan pada hewan ruminansia terdiri atas organ-
organ yang langsung berhubungan dengan penerimaan dan penyerapan zat-zat
makanan. Organ-organ ini dikelompokkan menjadi dua golongan, yaitu saluran
pencernaan dan organ-organ tambahan seperti gigi, lidah, kelenjar saliva, hati, dan
pankreas. Saluran pencernaan ternak ruminansia dibagi atas empat bagian penting,
yaitu mulut, perut, usus halus, dan organ pencernaan bagian belakang (hind gut).
Perut ternak ruminansia dibagi menjadi empat bagian, yaitu retikulum (perut jala),
rumen (perut beludru), omasum (perut buku) dan abomasum (perut sejati). Rumen
merupakan bagian lambung yang terbesar, permukaan bagian dalamnya tidak
halus, tetapi terdapat tonjolan-tonjolan yang tidak halus yang disebut papilae yang
berfungsi untuk memperbesar luas permukaan dinding rumen, sehingga absorbsi
produk fermentasi (VFA) lebih besar. Rumen dan retikulum dihuni oleh mikroba
dan merupakan alat pencernaan fermentatif dengan kondisi anaerob. Suhu di
dalam rumen berkisar 38-42oC dan pH-nya 6,8. pH tersebut dapat dipertahankan
tetap oleh adanya serapan VFA dan amonia serta saliva yang masuk dalam rumen.
Dari seluruh VFA yang diproduksi, 85% diserap melalui epitel retikulo-rumen
(Arora 1995).
Di dalam rumen terdapat sejumlah mikroba yang terdiri atas bakteri,
protozoa, dan sejumlah kecil fungi dan virus. Bakteri merupakan penghuni
terbesar di dalam rumen dan dikelompokkan menjadi tiga, yaitu (1) bakteri yang
hidup bebas dalam cairan rumen, yang jumlahnya sekitar 30% dari total bakteri,
(2) bakteri yang menempel pada partikel makanan yang jumlahnya sekitar 70%

19
dari total bakteri, dan (3) sebagian kecil kelompok bakteri yang melekat pada
dinding epitel rumen dan ada juga dalam jumlah kecil yang melekat pada protozoa
yaitu bersifat metanogenik.
Berdasarkan macam substrat yang disukainya, bakteri rumen dapat
dikelompokkan sebagai bakteri pencerna selulosa (misalnya Ruminococcus
albus), pencerna hemiselulosa (Butyrivibrio fibrisolvens), pencerna pati
(Bacteroides amylophilus), pencerna gula (Lactobacillus ruminus), dan bakteri
pengguna produk sekunder (pemakai laktat). Sekitar 30% bakteri rumen memiliki
aktivitas proteolitik.
Protozoa dalam rumen lebih sedikit populasinya, tetapi dari segi jumlah
biomasanya ternyata cukup besar. Spesies protozoa dipengaruhi oleh jenis
makanan. Populasi protozoa dalam rumen sapi atau domba yang memakan pakan
berserat dan mengandung gula terlarut yang rendah adalah sangat rendah berkisar
105/ml. Pada ternak yang pakannya mengandung gula atau tepung, populasi
protozoa meningkat mencapai 40 x 105/ml cairan rumen. Protozoa lebih menyukai
substrat yang fermentabel (pati, gula, dan bakteri). Produk fermentasi yang
dihasilkan adalah asam asetat, butirat, laktat, CO2, dan H2. Protozoa sangat peka
terhadap situasi asam, bila pH diturunkan maka jumlahnya dalam rumen akan
menurun.
Mikroba rumen yang lain adalah fungi yang bersifat anaerob dan ditemukan
pada beberapa jenis hewan herbivora, termasuk biri-biri, domba, sapi, dan
beberapa famili rusa (Arora 1995). Fungi mempunyai fungsi dalam mencerna
pakan ternak yaitu sebagai pembuka jalan agar bagian-bagian dinding sel tanaman
yang semula tidak dapat dicerna akhirnya dapat dicerna oleh ternak ruminansia.
Fungi adalah mikroba rumen yang pertama menyerang dan mencerna komponen
tanaman. Retikulum ukurannya lebih kecil dari rumen. Sering disebut dengan
retikulo-rumen karena bersama-sama rumen berfungsi sebagai alat pencernaan
fermentatif. Permukaan dalam retikulum menyerupai bentuk sarang tawon, yang
berfungsi mendorong pakan padat dan ingesta ke dalam rumen atau mengalirkan
ingesta selama ruminasi. Pola fermentasi di dalam organ ini serupa dengan yang
terjadi di dalam rumen (Arora 1995).

20
Tabel 5 Bakteri rumen, sumber energi, dan produk-produk fermentasi
Spesies Sumber energi Produk utama fermentasi
Bacteroides succinogenes Glukosa, selulosa,
selobiosa, pati
Asetat, suksinat, format
Ruminococcus albus Glukosa, selulosa, silan Asetat, laktat, format,
etanol, CO2, H2
Ruminococcus flavivacilus Glukosa, selulosa, silan Asetat, butirat, laktat,
format, etanol, CO2, H2
Butyrivibrio fibrisolvans Glukosa, selulosa, silan,
pati
Asetat, butirat, laktat,
format, etanol, CO2, H2
Bacteroides ruminicola Glukosa, silan, pati Asetat, propionat suksinat,
format
Bacteroides amylophilus Pati, maltose Asetat, suksinat, format
Selenomonas ruminantium Glukosa, pati, laktat,
gliserol,suksinat
Asetat, propionat, laktat,
format, CO2
Streptococcus bovis Glukosa,pati Laktat
Lachnospira Glukosa, pati, pektin Asetat, laktat, format,
etanol, CO2, H2
Succinivibrio Glukosa, dekstrin Asetat, suksinat, format
Vibrio spesies (lipolitik) Gliserol Propionat
Methanobacterium
ruminantum Format, H2 Metana
Sumber : Arora (1995)
Bagian ketiga dari lambung yang terletak di sebelah kanan rumen disebut
omasum. Dinding bagian dalammnya ditaburi lamina sehingga menambah luas
permukaannya (Arora 1995). Fungsi abomasum dalam proses pencernaan adalah
membantu memperkecil ukuran partikel pakan dan mengendalikan aliran ingesta
ke dalam perut bagian belakang serta absorbsi nutrien.
Bagian yang terakhir disebut dengan abomasum atau perut sejati,
merupakan tempat pertama terjadinya pencernaan pakan secara kimiawi atau
enzimatik karena adanya sekresi getah lambung. Abomasum mempunyai fungsi
yang sama dengan perut nonruminansia yaitu tempat pencernaan pakan oleh
enzim yang dihasilkan sel-sel dalam tubuh hewan maupun berupa getah-getah
pencernaan serta penyerapan nutrien.
Usus halus merupakan tempat utama hasil akhir pencernaan nutrien, seperti
asam amino, glukosa, asam-asam lemak, monogliserida, mineral, dan vitamin. Di
dalam lumen usus terdapat getah pankreas, getah usus, dan empedu untuk
mengubah pakan hasil akhir fermentasi mikroba rumen menjadi nutrien yang siap
diserap. Di dalam usus halus juga terdapat enzim-enzim proteolitik, seperti

21
tripsinogen, kemotripsinogen, prokarboksipeptidase, dan aminopeptidase. Selain
itu, juga terdapat enzim lipase, nukleosidase, enterokinase, dan gastrin.
Enterokinase dan gastrin adalah enzim yang terlibat dalam pengaktifan enzim-
enzim inaktif atau proses-proses sekresi (Arora 1995).
Termasuk organ pencernaan bagian belakang adalah sekum, kolon, dan
rektum. Sekum dan kolon adalah tempat fermentasi sisa bahan pakan yang tidak
dapat dicerna dalam rumen dan usus halus. Sisa pakan difermentasi oleh mikroba
sekum menjadi bentuk karbohidrat terlarut, nitrogen, dan sulfur yang kemudian
digunakan oleh mikroba untuk berbiak. Proses fermentasi VFA sama dengan di
rumen, namun jumlah bakterinya 10-1000 kali lebih sedikit daripada rumen
(Arora 1995).
Pencernaan dan Metabolisme Karbohidrat
Pakan ternak ruminansia sebanyak 60-70% terdiri atas karbohidrat berupa
selulosa, hemiselulosa, pati, pektin, dan karbohidrat yang mudah larut. Selulosa
dan hemiselulosa tidak dicerna oleh enzim yang dihasilkan ternak ruminansia,
tetapi dicerna oleh enzim-enzim yang dihasilkan oleh mikroba rumen yang juga
mencerna pati dan karbohidrat yang larut dalam air. Oleh karena itu, ternak
ruminansia bergantung sepenuhnya pada peranan mikroba rumen dalam mencerna
fraksi serat pada pakan. Laju pertumbuhan mikroba dalam rumen sangat
bergantung pada ketersediaan karbohidrat, karena merupakan sumber energi dan
sebagai kerangka karbon dalam sintesis protein mikroba. Oleh karena itu, laju
pencernaan karbohidrat merupakan salah satu faktor penentu produksi protein
mikroba.
Pencernaan karbohidrat di dalam rumen berlangsung melalui kerja sama
mikroba rumen, terutama fungi yang bekerja sama dengan bakteri selulolitik,
amilolitik, serta protozoa. Fermentasi karbohidrat dalam rumen terjadi melalui
dua tahap. Tahap pertama adalah pemecahan karbohidrat kompleks menjadi gula
sederhana. Pemecahan ini terjadi oleh pengaruh enzim ekstraseluler dari mikro-
organisme (Tilman et al. 1989). Urutan pola fermentasi dalam rumen adalah
glukosa, xylosa, pati, dan selulosa (Arora 1995). Selulosa dipecah oleh satu atau
lebih glukosidase menjadi selobiosa yang kemudian diubah menjadi glukosa.

22
Pada tahap kedua, glukosa yang terbentuk pada tahap pertama akan diserap
oleh sel mikroba dan segera mengalami metabolisme intraseluler menjadi piruvat
melalui lintasan Embden Meyerhorf dan lintasan Pentosa Fosfat. Piruvat adalah
bentuk intermedier yang segera dimetabolisme melalui proses fermentasi untuk
membentuk produk utama pencernaan fermentatif dalam rumen, yaitu asam-asam
lemak rantai pendek yang biasa disebut VFA. VFA utama yang dihasilkan adalah
asam asetat (CH3COOH), propionat (CH3CH2COOH), dan butirat
(CH3(CH2)2COOH). Di samping VFA, proses fermentasi karbohidrat juga
menghasilkan produk sampingan berupa gas metana (CH4), CO2, dan H2. Gas-gas
tersebut dikeluarkan dari dalam rumen melalui proses eruktasi. Banyaknya VFA
yang dihasilkan di dalam rumen sangat bervariasi antara 200-1500 mg/100 ml
cairan rumen, bergantung pada jenis ransum yang dikonsumsi. Menurut
Suryapratama (1999), kisaran konsentrasi VFA total yang layak bagi
kelangsungan hidup ternak adalah 80–160 mM, dengan titik optimum 110 mM.
Stoikiometri reaksi fermentasi karbohidrat menjadi tiga produk fermentasi
utama di dalam rumen dapat disederhanakan sebagai berikut (Orskov & Ryle
1990) :
C6H12O6 + 2H2O 2CH3COOH + 2CO2 + 4H2
C6H12O6 + 2H+
2 2CH3CH2COOH + 4 H2O
C6H12O6 2CH3(CH2)2COOH + 2CO2 + 2 H+
2
4H+2 + CO2 CH4 + H2O
Dari stoikiometri reaksi tersebut, tampak bahwa pada proses sintesis asam
asetat banyak dihasilkan gas hidrogen, demikian pula pada sintesis asam butirat.
Sebaliknya pada proses sintesis asam propionat, gas hidrogen banyak yang
digunakan. Gas hidrogen bersama dengan gas CO2 merupakan prekursor untuk
sintesis gas metana. Gas metana sesungguhnya tidak bermanfaat bagi hewan
induk semang, sehingga pola fermentasi rumen yang mengarah pada sintesis asam
propionat jelas akan lebih menguntungkan dari segi efisiensi penggunaan energi
pakan. Konsentrasi asam asetat dalam rumen berhubungan dengan kandungan
serat kasar dalam pakan, sedangkan propionat berhubungan dengan konsentrat.
VFA yang dihasilkan dalam fermentasi rumen diserap oleh darah dari
dinding rumen dan dibawa ke hati. Asam propionat diubah menjadi glukosa

23
melalui asetil Ko-A dan menjadi bagian cadangan glukosa hati. Sebagian dari
glukosa tersebut diubah menjadi glikogen dan disimpan di dalam hati atau diubah
menjadi alfa-gliserolfosfat dan digunakan untuk sintesis trigliserida. Sisa glukosa
akan masuk ke peredaran darah menuju pelbagai jaringan tubuh dan digunakan
sebagai sumber energi, sumber koenzim pereduksi dalam sintesis asam lemak, dan
glikogen otot. Asam asetat dan butirat diabsorbsi seperti halnya asam propionat,
namun asam butirat diubah menjadi asam beta-hidroksi-butirat (BHBA) oleh
jaringan dinding rumen. Asam asetat dan BHBA dari hati disalurkan ke sistem
sirkulasi dan dipakai oleh jaringan sebagai sumber energi dan untuk sintesis asam
lemak.
Pencernaan dan Metabolisme Protein
Protein bahan makanan terdiri atas protein murni dan senyawa-senyawa
nitrogen bukan protein (NPN). Pada ternak ruminansia, sebagian protein yang
masuk ke dalam rumen mengalami perombakan (degradasi) oleh enzim proteolitik
yang diproduksi oleh mikroba rumen. Enzim tersebut terdiri atas eksopeptidase
dan endopeptidase. Beberapa bakteri rumen yang paling tinggi aktivitas
proteolitiknya ialah Fibrobacter, Butyrivibrio, dan selenomonas.
Perombakan protein oleh enzim proteolitik di dalam rumen menghasilkan
peptida dan asam-asam amino. Sebagian mikroba rumen dapat memanfaatkan
oligopeptida untuk membuat protein tubuhnya. Mikroba rumen tidak dapat
memanfaatkan asam amino secara langsung karena tidak mempunyai sistem
transpor untuk mengangkut asam amino ke dalam tubuhnya. Oleh karena itu,
asam amino didegradasi melalui deaminasi menjadi asam-asam organik, amonia
dan CO2. Amonia yang terbentuk dari deaminasi dapat dikombinasikan dengan
asam organik alfa-keto membentuk asam amino baru yang dipakai untuk sintesis
protein mikroba, terutama dari kelompok bakteri.
Amonia merupakan sumber nitrogen yang relatif banyak dibutuhkan oleh
mikroba rumen untuk membentuk protein mikroba bersama dengan kerangka
karbon yang berasal dari sumber energi. Menurut Arora (1995), bakteri rumen
menggunakan amonia sampai 5 mM (8.5 mg/100ml), sedangkan kelompok ciliata
tidak dapat menggunakan amonia seperti halnya bakteri, sehingga protozoa ini
memakan bakteri untuk kebutuhan zat makanannya. Di samping dimanfaatkan

24
oleh mikroba rumen, amonia yang dihasilkan juga diserap oleh darah melalui
dinding rumen di bawa ke hati dan diubah menjadi urea. Sebagian besar urea
difiltrasi oleh ginjal dan dikeluarkan bersama urine, dan sebagian lagi masuk
peredaran darah dan dikembalikan ke rumen melalui saliva. Di dalam rumen, urea
diubah oleh urease mikroba menjadi CO2 dan amonia. Konsentrasi amonia rumen
merupakan suatu besaran yang sangat penting untuk dikendalikan, karena sangat
menentukan optimasi pertumbuhan mikroba rumen. Konsentrasi optimum NH3
untuk menunjang pertumbuhan mikroba rumen berkisar 85–300 mg/l atau 6–21
mM dengan titik optimum 8 mM.
Protein makanan yang lolos dari degradasi rumen dan protein mikroba
selanjutnya mengalami pencernaan dan penyerapan di usus halus. Protein ini
kebanyakan berupa protein mikroba dan sedikit berasal dari makanan. Agar
protein bahan makanan lebih bnyak tersedia bagi ternak maka ada dua cara yang
dapat dilakukan, yaitu melalui by pass protein dan pemberian NPN. Mengingat
sifat mikroba rumen yang lebih mudah menggunakan amonia dibanding asam
amino, maka pemberian NPN akan lebih tepat dari by pass protein.
Pencernaan protein di usus halus menghasilkan asam-asam amino yang
selanjutnya diserap oleh darah dibawa ke hati. Dari hati, asam-asam amino
tersebut disalurkan ke jaringan tubuh lainnya termasuk kelenjar susu untuk
membentuk protein jaringan dan protein susu. Sumbangan protein mikroba rumen
terhadap kebutuhan asam-asam amino ternak ruminansia mencapai 40-80%. Sisa-
sisa pencernaan protein berupa protein dan asam amino bahan makanan yang
tidak tercerna dan terabsorbsi akan dikeluarkan dari tubuh melalui feses.
Metanogenesis di Rumen
Ruminansia sangat bergantung pada mikroba rumen untuk mencerna dan
memfermentasi dinding sel polisakarida tanaman menjadi sumber energi,
seperti VFA dan asam organik lainnya. Ternak juga mencerna mikroba yang
lolos dari rumen sebagai sumber protein. Melalui Simbiosis mikroba di dalam
retikulo-rumen, karbohidrat tanaman dihidrolisis menjadi gula sederhana,
dengan hasil akhir berupa volatile fatty acid (asam asetat, propionat, dan
butirat), dan gas CH4, H2, dan CO2 (Hobson 1997).

25
Untuk mengurangi akumulasi H2 di dalam rumen, bakteri metanogen
terutama dari golongan Archaea, memanfaatkan gas tersebut menjadi metana
dan digunakan sebagai sumber energi. Persamaan reaksinya adalah : CO2 +
4H2 CH4 + 2H2O. Meskipun proses ini sangat diperlukan dalam fermentasi
rumen, namun secara energetika merugikan ternak karena tidak bisa digunakan
sebagai sumber energi. Sebagian besar metana dikeluarkan melalui eruktasi
yaitu sekitar 83% dan sisanya melalui pernapasan dan anus masing-masing
16%, dan 1%. Hilangnya energi pakan sebagai metana menyebabkan
rendahnya efisiensi dan berpengaruh pada produktivitas ternak.
Proses metanogenesis sebagian besar terjadi di dalam rumen (83-94%)
dan hanya sedikit di pencernaan bagian belakang (6-13%) oleh bakteri
metanogen. Koloni mikroba pengguna hidrogen (metanogen) di dalam rumen
mulai muncul setelah hewan ruminansia mendapat susu induknya, sedangkan
koloni asetogen muncul 24 jam pertama setelah lahir. Konsentrasi metanogen
bertambah banyak setelah 1-3 hari setelah lahir dengan konsentrasi sekitar 104 per
gram isi rumen dan meningkat secara eksponensial menjadi 108-10
9 per gram isi
rumen, setelah umur 3 minggu. Sejak metanogen menghuni rumen, konsentrasi
asetogen menurun. Beberapa jenis bakteri metanogen yang telah diisolasi dari
rumen dan substrat yang digunakan untuk metanogenesis tercantum pada Tabel 6
dan 7.
Terbentuknya metana di dalam rumen merupakan hasil kerja sama melalui
simbiosis dari beberapa mikroba untuk memperoleh kebutuhan energinya.
Sebagian besar mikroba rumen memenuhi kebutuhan energinya dari gula
sederhana hasil degradasi dan pencernaan bahan pakan yang selanjutnya
dioksidasi melalui siklus asam sitrat. Selama siklus asam sitrat, elektron-elektron
Tabel 6 Metanogen yang telah diisolasi dari rumen
Organisme Sumber energi
Methanobrevibacter ruminantium H2, format
Methanobrevibacter sp. H2,format
Methanosarcina barkeri H2, methanol, methylamine, asetat
Methanosarcina mazei H2, methanol, methylamine, asetat
Methanobacterium formicicum H2, format
Methanomicrobium mobile H2, format
Sumber : McAllister dan Cheng (1996)

26
Tabel 7 Substrat untuk metanogenesis
Substrat Persamaan Reaksi
H2 dan CO2 4H2 +CO2 CH4 + 2H2O
Format 4HCO2H CH4 + 3CO2 + 2H2O
Metanol 4CH3OH 3CH4 + CO2 + 2H2O
Metanol dan H2 CH3OH + H2 CH4 + H2O
Metilamine 4CH2NH2Cl + 2H2O 3CH4 + CO2 + 4NH4Cl
Dimetilamine 2(CH3)2NHCl + 2H2O 3CH4 + CO2 + 4NH4Cl
Trimetilamine 4(CH3)3NCl + 6H2O 9CH4 + 3CO2 + 4NH4Cl
Asetat CH3COOH CH4 + CO2
Sumber : Hobson (1997)
lepas kembali sebagai hidrogen bebas (H2), yang selanjutnya diambil oleh
koenzim khusus pembawa elektron intraseluler (NAD), dan direduksi menjadi
NADH. Elektron kemudian ditransfer ke aseptor elektron seperti CO2, sulfat,
nitrat, atau fumarat menghasikan NAD dan fermentasi gula menjadi sempurna
(Immig 1996; Moss et al. 2000). Fermentasi mikroba menghasilkan H2 dan
metanogen menggunakan hidrogen untuk mereduksi CO2 menjadi CH4.
Satu-satunya sumber energi bagi metanogen berasal dari reduksi CO2
menjadi CH4 menggunakan H2, format, metanaol, metilamine, atau asetat melalui
reduksi intermediate formyl-methanofuran, methenyl-tetrahydromethanopterin,
methylenyl-tetrahydromethanopterin dan methyl-tetrahydromethanopterin (Mc-
Allister & Cheng 1996). Metanogen memiliki 3 koenzim yang tidak ditemukan
pada mikroorganisme lainnya, yaitu koenzim 420 yang terlibat dalam transfer
elektron, koenzim N terlibat dalam transfer metil, dan faktor B yang terlibat dalam
pembentukan enzimatik CH4 dari metil ko enzim M.Enzim-enzim spesifik
tersebut memungkinkan metanogen memproduksi CH4. Transfer H2 dari fer-
mentasi mikroba ke metanogen dibantu sifat hidrofobik yang dimilikinya
sehingga memudahkan mereka menempel pada partikel pakan dan permukaan
protozoa. Pengambilan H2 dan produksi CH4 menjadi lebih banyak (Boadi et al.
2004). Hal ini membantu mencegah peningkatan konsentrasi H2 di rumen dan
mengurangi akumulasi nukleotida (NADH) dan menyebabkan fermentasi bahan
tanaman oleh bakteri dan fungi lebih besar (5-28%) dibanding tidak adanya
metanogen (McAllister & Cheng 1996).

27
Gambar 7 Fermentasi bahan tanaman oleh mikroba rumen dan beberapa
bakteri yang berperan, serta metanogen yang mengubah hidrogen
menjadi metana (McAllister & Cheng 1996)
Produksi metana di dalam saluran pencernaan (enteric fermentation)
dipengaruhi oleh beberapa faktor seperti tingkat konsumsi pakan, jenis
karbohidrat, pengelolaan hijauan, penambahan lemak dan manipulasi mikroflora
rumen. Tiga faktor yang paling utama adalah laju fermentasi bahan organik, tipe
VFA yang diproduksi karena berkaitan dengan jumlah hidrogen yang diproduksi
dan efisiensi biosintesis mikroba. Laju fermentasi bahan organik sangat
dipengaruhi oleh tingkat konsumsi pakan dan karakteristik degradasi karbohidrat.
Lambatnya laju pencernaan dan meningkatnya retensi rumen menyebabkan
penurunan konsumsi pakan dan lebih banyak CH4 yang diproduksi per kg dray
matter intake (DMI). Meningkatnya tingkat serat dalam ransum menghasilkan
rasio asetat propionat dan produksi CH4 yang lebih tinggi (Johnson & Johnson
1995).
Produksi propionat dari piruvat membutuhkan hidrogen, sedangkan
produksi asetat menghasilkan hidrogen, yang digunakan oleh metanogen untuk
membentuk CH4. Oleh karena itu, produksi metana adalah berbanding terbalik
dengan produksi propionat dan sebanding dengan rasio asetat propionat.
Karbohidrat yang mudah difermentasi dan pH yang rendah menyebabkan rasio
28
asetat propionat dan CH4 yang dihasilkan lebih rendah, dan ini terjadi pada pakan
yang tinggi biji-bijian (Johnson & Johnson 1995; Moss et al. 2000).
Complete Rumen Modifier (CRM) dan Cassapon
Complete rumen modifier (CRM) merupakan feed additives yang
kandungannya terdiri atas (1) Senyawa saponin dari buah lerak (Sapindus rarak)
yang disiapkan menurut prosedur Thalib et al. (1994), tepung daun Albizia
falcutaria dan Sesbania grandiflora; (2) isolat bakteri dengan media asetogenik
pengguna hidrogen-karbondioksida yaitu: Asetoanaerobium notrae yang
disiapkan menurut prosedur Thalib (2008); dan (3) faktor pertumbuhan mikroba
(FPM) yang disiapkan menurut prosedur Thalib et al. (1998).
Cassapon merupakan produk dari Balai Penelitian Ternak terdiri dari
kombinasi CRM dan onggok yang berfungsi untuk menurunkan emisi gas metana
enterik dan sekaligus meningkatkan produktivitas ternak ruminansia. Selain itu,
peranan penambahan onggok adalah substrat bagi konsorsium mikroba selulolitik
untuk produksi enzim selulosa. Prinsip kerja bahan ini adalah mensuplai nutrisi
untuk pertumbuhan mikroba, menghambat proses metanogenesis dengan cara
menumbuhkan bakteri asetogenik pengguna hidrogen, dan menekan populasi
protozoa (defaunasi). Tabel proksimat studi sebelumnya mengenai CRM disajikan
pada Tabel 8.
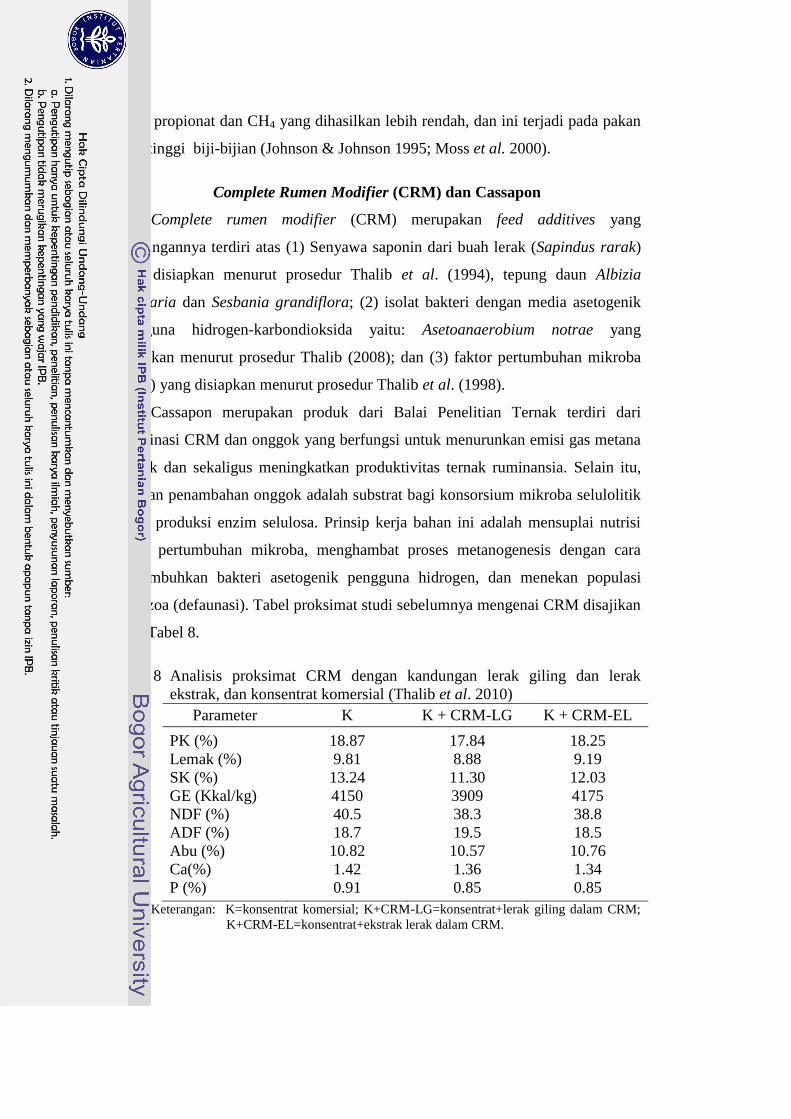
Tabel 8 Analisis proksimat CRM dengan kandungan lerak giling dan lerak
ekstrak, dan konsentrat komersial (Thalib et al. 2010)
Parameter K K + CRM-LG K + CRM-EL
PK (%) 18.87 17.84 18.25
Lemak (%) 9.81 8.88 9.19
SK (%) 13.24 11.30 12.03
GE (Kkal/kg) 4150 3909 4175
NDF (%) 40.5 38.3 38.8
ADF (%) 18.7 19.5 18.5
Abu (%) 10.82 10.57 10.76
Ca(%) 1.42 1.36 1.34
P (%) 0.91 0.85 0.85
Keterangan: K=konsentrat komersial; K+CRM-LG=konsentrat+lerak giling dalam CRM;
K+CRM-EL=konsentrat+ekstrak lerak dalam CRM.

29
Penelitian pendahuluan mengenai bahan-bahan penyusun CRM ini sudah
dilakukan, namun masih secara tersendiri. Menurut hasil penelitian Thalib (2008),
pemberian bakteri asetogenik (Acetanoanaerobium noterae) mampu memperbaiki
performans domba dan menurunkan produksi gas metana enterik dan hasilnya
lebih nyata bila dikombinasikan dengan defaunator berupa aksafon SR.
Defaunator berfungsi untuk menekan populasi protozoa karena disamping
memakan bakteri pencerna serat, protozoa juga berhubungan dengan populasi
metanogen. Metanogen (Archaea) biasanya melekat pada protozoa terutama dari
golongan ciliata. Bahan yang biasa digunakan sebagai defaunator adalah bahan-
bahan yang mengandung saponin, salah satu di antaranya adalah buah lerak.
Menurut Thalib et al. (1996) buah lerak sangat efektif digunakan sebagai
defaunator protozoa. Secara kimia, saponin memiliki diversifikasi struktur yang
luas dan senyawa-senyawa saponin tertentu dengan sifat surfaktannya dapat
menyebabkan terjadinya lisis pada dinding sel protozoa.




















