
PENGEMBANGAN METODE ORGANOGENESIS DAN … · Oleh karena itu, teknologi kultur in vitro dipandang...
185
PENGEMBANGAN METODE ORGANOGENESIS DAN EMBRIOGENESIS SOMATIK PADA NENAS (Ananas comosus (L.) Merr.) SERTA DETEKSI DINI UNTUK MEREDUKSI KERAGAMAN SOMAKLONAL IKA ROOSTIKA TAMBUNAN SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR BOGOR 2012
Transcript of PENGEMBANGAN METODE ORGANOGENESIS DAN … · Oleh karena itu, teknologi kultur in vitro dipandang...

PENGEMBANGAN METODE ORGANOGENESIS DAN EMBRIOGENESIS SOMATIK PADA NENAS (Ananas comosus
(L.) Merr.) SERTA DETEKSI DINI UNTUK MEREDUKSI KERAGAMAN SOMAKLONAL
IKA ROOSTIKA TAMBUNAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2012

PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Pengembangan Metode Organogenesis dan Embriogenesis Somatik pada Nenas (Ananas comosus (L.) Merr.) serta Deteksi Dini untuk Mereduksi Keragaman Somaklonal adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini. Bogor, 26 Juli 2012 Ika Roostika Tambunan NRP A263090111

ABSTRACT IKA ROOSTIKA TAMBUNAN. Development of Organogenesis and Somatic Embryogenesis Regeneration of Pineapple (Ananas comosus (L.) Merr.) and Early Detection to Reduce Somaclonal Variation. Supervised by NURUL KHUMAIDA, GUSTAAF ADOLF WATTIMENA, and IKA MARISKA SOEDHARMA. Pineapple is a unique plant belongs to facultative CAM (Crassulacean Acid Metabolism) photosynthesis. It is highly potential to be planted on suboptimal land. Conventionally, it can be propagated from many propagules but their reproductive time is not uniform and this availability is commonly limited in cultivar Smooth Cayenne. Today micropropagation is being used commercially in the pineapple industry abroad but in Indonesia, the industries survive with conventional vegetative propagation because of somaclonal variation. The main goal of the study was to obtain the effective regeneration method of pineapple with high level of seedling production and low level of somaclonal variation, included the artificial seed formation method, the early detection method of somaclonal variation for eliminating undesirable traits during micropropagation, and the method to reduce the level of somaclonal variation during micropropagation. This study was devided into six chapters, included the study of morhogenetic system (with solid and liquid media); direct organogenesis induced by auxin and cytokinin (IAA, IBA, NAA, and BA); indirect organogenesis and somatic embryogenesis induced by 2,4-D, picloram, adenine sulfate, and N-organic compounds; encapsulation and minimal growth induced by paclobutrazol, mannitol, and reduced temperature; morphological and molecular characterization by using RAPD markers; and early detection to reduce of somaclonal variation. Generally, this study has some novelities in the in vitro characterization, the information of the non dead-end mechanism of rooted pineapple explants, the information of the complete events of somatic embryogenesis, the conservation method by minimal growth, and the large number of variants. The result showed that the direct and indirect organogenesis generated high shoot multiplication level. The use of mannitol (4%) was better than paclobutrazol for storing the encapsulated explants for 4 months. The morphological characterization could easily differentiate the normal and the abnormal variants (off-types). The direct organogenesis showed the lower level of variation (1.6%) compare to the indirect organogenesis (2%) and somatic embryogenesis (31.1%). The RAPD analysis strengthened the occurence of somaclonal variation. The use of the new in vitro cultures could reduce somaclonal variation. In conclusion, the direct organogenesis was the effective regeneration method for mass propagation. The morphological characterization of in vitro cultures can be applied as guidance for early detection and elimination of undesirable variants in mass propagation. The in vitro conservation method by mannitol can be applied to store pineapple cultures for medium-term period. The use of the new in vitro population could reduce the level of somaclonal variation. Keywords: Organogenesis, somatic embryogenesis, synthetic seed, in vitro
conservation, somaclonal variation.

RINGKASAN IKA ROOSTIKA TAMBUNAN. Pengembangan Metode Organogenesis dan Embriogenesis Somatik pada Nenas (Ananas comosus (L.) Merr.) serta Deteksi Dini untuk Mereduksi Keragaman Somaklonal. Dibimbing oleh NURUL KHUMAIDA, GUSTAAF ADOLF WATTIMENA, dan IKA MARISKA SOEDHARMA. Nenas (Ananas comosus L. Merr.) merupakan tanaman yang unik karena
memiliki tipe fotosintesis CAM (Crassulacean Acid Metabolism) fakultatif sehingga
tergolong sebagai tanaman masa depan yang prospektif untuk dikembangkan secara
luas dalam rangka revitalisasi tanaman pertanian untuk mendukung era Revolusi
Hijau Lestari, terutama pada lahan marginal. Tanaman nenas memiliki propagul yang
beraneka ragam, tetapi umur panennya bervariasi sehingga kurang ideal sebagai
bahan perbanyakan tanaman secara masal. Kultivar Smooth Cayenne bahkan hanya
memiliki jenis dan jumlah propagul yang terbatas, yaitu berupa mahkota dan sucker.
Oleh karena itu, teknologi kultur in vitro dipandang sebagai solusi yang terbaik untuk
diterapkan. Sayangnya, isu keragaman somaklonal masih menjadi kendala utama
sehingga pengguna, khususnya pihak industri masih enggan menerapkan teknologi
tersebut. Tujuan utama penelitian ini adalah untuk memperoleh metode regenerasi
yang efektif (dengan tingkat multiplikasi tinggi dan tingkat keragaman rendah),
termasuk metode pembentukan benih sintetik, deteksi dini dan reduksi keragaman
somaklonal. Penelitian terbagi atas enam bagian, mencakup (1) studi morfogenesis
eksplan secara organogenesis langsung yang diinduksi oleh auksin dan sitokinin
(IAA, IBA, NAA, dan BA), (2) studi organogenesis dan embriogenesis tidak
langsung yang diinduksi oleh 2,4-D, (3) studi embriogenesis somatik tidak langsung
yang diinduksi oleh pikloram, (4) studi pembentukan benih sintetik dan konservasi in
vitro secara pertumbuhan minimal yang diinduksi oleh paklobutrazol atau manitol
dan modifikasi suhu penyimpanann, (5) studi karakterisasi morfologi dan molekuler
dengan penanda RAPD (10 primer) terhadap biakan dan bibit nenas, serta (6) studi
evaluasi keragaman bibit, deteksi dini, dan reduksi keragaman somaklonal. Hasil
penelitian menunjukkan bahwa metode organogenesis langsung dan tidak langsung
mempunyai tingkat multiplikasi tunas yang lebih tinggi daripada embriogenesis
somatik tidak langsung. Pikloram lebih baik dalam menginduksi embriogenesis
somatik daripada NAA dan 2,4-D dengan tingkat multiplikasi 17 embrio/eksplan

selama 2 bulan dari struktur ETs (embryogenic tissues) atau 14 embrio/eksplan
selama 2.5 bulan dari struktur FETs (friable embryogenic tissues). Penggunaan
manitol (4%) lebih baik daripada penggunaan paklobutrazol maupun penurunan suhu
dalam menyimpan eksplan nenas dalam kondisi terenkapsulasi, dengan periode
simpan hingga 4 bulan. Terdapat 21 varian yang terdeteksi dari populasi biakan
induk, di mana 8 varian menunjukkan fenomena epigenetik, 8 varian menunjukkan
fenomena khimera, dan 7 varian menunjukkan kestabilan fenotipe di rumah kaca.
Karakterisasi secara morfologi membedakan varian dan tanaman kontrol dengan
koefisien kemiripan 0.49-0.91 (r=0.83). Karakterisasi molekuler memperkuat dugaan
terjadinya keragaman somaklonal. Berdasarkan karakterisasi tersebut, tanaman
kontrol dan varian memiliki koefisien kemiripan sebesar 0.32-0.61 (r=0.97). Ketidak-
stabilan fenotipe pada karakter duri pada daun diduga kuat disebabkan oleh
retrotransposon karena pola terbentuknya duri sama sekali tidak beraturan, bahkan
pada bibit yang dihasilkan dari metode embriogenesis somatik yang terbukti bersifat
unicellular origin. Metode embriogenesis somatik tidak langsung menghasilkan
keragaman fenotipe yang tertinggi (31.1%), diikuti dengan metode organogenesis
tidak langsung (2%) dan organogenesis langsung (1.6%). Populasi biakan in vitro
yang baru memiliki koefisien kemiripan genetik yang lebih tinggi (0.65-0.85)
daripada populasi lama, baik sebelum (0.32-0.61) maupun sesudah diekstraksi
dengan menggunakan 3 macam metode regenerasi (0.34-0.63). Hal ini
mengindikasikan bahwa keragaman somaklonal dapat direduksi dengan
menggunakan bahan tanaman yang baru. Disimpulkan bahwa organogenesis
langsung merupakan metode terbaik untuk mikropropagasi tanaman nenas.
Penggunaan manitol lebih baik daripada paklobutrazol untuk penyimpanan in vitro
dengan periode simpan 4 bulan. Metode karakterisasi morfologi dapat diterapkan
pada biakan in vitro untuk eliminasi varian yang tidak diharapkan selama proses
mikropropagasi secara dini. Teknik RAPD mengkonfirmasi dugaan terjadinya variasi
somaklonal pada kultur in vitro nenas.
Kata kunci: Organogenesis, embriogenesis somatik, benih sintetik, konservasi in vitro, karakterisasi morfologi, karakterisasi molekuler, keragaman somaklonal.

© Hak Cipta milik IPB, tahun 2012 Hak Cipta dilindungi oleh Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis dalam bentuk apapun tanpa izin IPB

PENGEMBANGAN METODE ORGANOGENESIS DAN EMBRIOGENESIS SOMATIK PADA NENAS (Ananas comosus
(L.) Merr.) SERTA DETEKSI DINI UNTUK MEREDUKSI KERAGAMAN SOMAKLONAL
IKA ROOSTIKA TAMBUNAN
Disertasi sebagai salah satu syarat untuk memperoleh gelar
Doktor pada Program Studi Pemuliaan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2012

Judul : Pengembangan Metode Organogenesis dan Embriogenesis Somatik pada Nenas (Ananas comosus (L.) Merr.) serta Deteksi Dini untuk Mereduksi Keragaman Somaklonal
Nama : Ika Roostika Tambunan
NRP : A263090111
Disetujui, Komisi Pembimbing
Dr. Ir. Nurul Khumaida, M.S.Ketua
Prof. Dr. Ir. G.A. Wattimena, M.Sc. Prof. (R) Dr. Ika Mariska, APU Anggota Anggota
Diketahui,
Ketua Mayor Dekan Sekolah Pascasarjana Pemuliaan Bioteknologi Tanaman Dr. Ir. Tri Koesoemaningtyas, M.Sc. Dr. Ir. Dahrul Syah, M.Agr. Tanggal Ujian: 26 Juli 2012 Tanggal lulus: 9 Agustus 2012

Penguji pada Ujian Tertutup: Prof. Dr. Ir. Slamet Susanto, MAgr
Dr. Ir. Darda Efendi, MSi
Penguji pada Ujian Terbuka: Dr. Ir. Sobir, MS
Dr. Ir. Yusdar Hilman, MSc

PRAKATA
Syukur Alhamdulillah penulis panjatkan ke hadirat Allah SWT atas segala petunjuk dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Disertasi ini mengangkat topik tentang Pengembangan Metode Organogenesis dan Embriogenesis Somatik pada Nenas (Ananas comosus (L.) Merr.) serta Deteksi Dini untuk Mereduksi Keragaman Somaklonal. Penelitian tersebut dilaksanakan sejak Februari 2010 hingga April 2012.
Disertasi ini memuat enam bab yang merupakan pengembangan dari naskah artikel yang diajukan ke jurnal dan pertemuan ilmiah. Bab 2 telah diterbitkan pada Jurnal Agrobiogen Vol.8 No.1 Th. 2012 dengan judul Indirect organogenesis and somatic embryogenesis of pineapple induced by dichlorophenoxy acetic acid. Bab 3 sedang dalam proses penerbitan pada Indonesian Journal of Agricultural Science dengan judul The effect of picloram and light on somatic embryogenesis regeneration of pineapple. Bab 4 sedang diajukan ke Jurnal Hortikultura dengan judul Pembentukan Benih Sintetik Nenas dan Konservasi In Vitro secara Pertumbuhan Minimal. Bab 5 masih dalam proses pendampingan publikasi pada jurnal internasional (Plant Cell Tissue and Organ Culture) dengan judul Morphological characterization of pineapple in vitro cultures and acclimated seedlings. Bab 6 akan disampaikan pada International Seminar on Agriculture Adaptation in The Tropics dengan judul Molecular characterization of pineapple in vitro cultures for detecting plant off-types during micropropagation.
Penghargaan yang setinggi-tingginya disampaikan kepada Badan Litbang Pertanian yang telah memberikan kesempatan kepada penulis untuk melanjutkan studi S3 dan atas dukungan dana penelitian melalui Program KKP3T 2011. Terima kasih yang tidak terhingga penulis ucapkan kepada Dr. Nurul Khumaida, Prof. Dr. G.A. Wattimena dan Prof. (R) Dr. Ika Mariska sebagai komisi pembimbing. Ucapan terima kasih juga disampaikan kepada Dr. Sutrisno, Dr. Karden Mulya (Kepala BB-Biogen), Dr. Sobir, Dr. Wicaksono, Prof. Dr. Sudarsono, Prof. Dr. Wasmen Manalu, Dr. Tri Koesoemaningtyas, Dr. Endah Retno Palupi, Prof. Dr. Bambang S. Purwoko, dan Dr. Made Tasma yang telah memberikan dukungan sebelum dan semasa penelitian serta pemahaman, saran dan masukan dalam penelitian maupun penyusunan naskah publikasi ilmiah. Kepada penguji luar komisi, Dr. Agus Purwito dan Dr. Sintho W. Ardie (pada ujian Prakualifikasi Doktor), Prof. Dr. Slamet Susanto dan Dr. Darda Efendi (pada Ujian Tertutup) serta Dr. Yusdar Hilman, APU dan Dr. Sobir (pada Ujian Terbuka), disampaikan terima kasih. Terima kasih juga ditujukan kepada teman-teman sejawat dan rekan-rekan teknisi di Kelti BSJ BB-Biogen serta rekan-rekan mahasiswa pascasarjana IPB. Rasa cinta dan terima kasih juga disampaikan kepada Ibunda dan Ayahanda (Ibu Suhaesih dan Alm. Djamian Tambunan), suami (Faleh Setia Budi, MT.), ananda (Furaida Alya Muna) serta keluarga besar dan seluruh pihak yang telah memberikan dukungan moril, dan spirituil. Besar harapan penulis akan seluas-luasnya manfaat dari hasil karya ini. Bogor, Juli 2012 Ika Roostika Tambunan

RIWAYAT HIDUP
Penulis dilahirkan di Magelang pada tanggal 12 September 1972 sebagai anak kedua dari pasangan Djamian Tambunan dan Suhaesih. Pendidikan sarjana ditempuh pada Jurusan Hama dan Penyakit Tumbuhan Program Studi Fitopatologi, Fakultas Pertanian Universitas Gadjah Mada, lulus pada tahun 1997. Pada tahun 1999, penulis diterima sebagai mahasiswa pascasarjana pada Program Studi Agronomi dengan peminatan Pemuliaan Non Konvensional Institut Pertanian Bogor, lulus pada tahun 2003. Kesempatan untuk melanjutkan studi ke program doktor pada Program Studi Pemuliaan Bioteknologi Tanaman IPB diperoleh pada tahun 2009, dengan beasiswa dari Badan Litbang Pertanian.
Sejak lulus sebagai sarjan hingga saat ini, penulis bekerja di Balai Besar Penelitian dan Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian, Bogor. Bidang keahlian adalah Bioteknologi Pertanian dengan spesifikasi kultur in vitro, terutama pada aspek konservasi in vitro (pertumbuhan minimal dan kriopreservasi) serta variasi somaklonal.
Selama mengikuti program S3, penulis mengikuti beberapa macam pelatihan, yaitu penulisan publikasi pada jurnal internasional, perancangan percobaan, dan bahasa Inggris. Karya ilmiah yang berjudul Indirect organogenesis and somatic embryogenesis of pineapple induced by dichlorophenoxy acetic acid telah diterbitkan pada Jurnal Agrobiogen Vol.8 No.1 Th. 2012. Artikel lainnya yang berjudul The effect of picloram and light on somatic embryogenesis regeneration of pineapple menunggu penerbitannya pada Indonesian Journal of Agricultural Science (Jurnal IJAS), sedangkan naskah yang berjudul Morphological characterization of pineapple in vitro cultures and acclimated seedlings masih dalam proses pendampingan publikasi pada jurnal internasional (Plant Cell Tissue and Organ Culture). Naskah yang berjudul Pembentukan Benih Sintetik Nenas dan Konservasi In Vitro secara Pertumbuhan Minimal sedang diajukan ke Jurnal Hortikultura dan naskah yang berjudul Molecular characterization of pineapple in vitro cultures for detecting plant off-types during micropropagation akan disampaikan pada International Seminar on Agriculture Adaptation in The Tropics. Karya-karya tersebut merupakan bagian dari program S3 penulis.

DAFTAR ISI
Halaman DAFTAR TABEL ……………………………………………………….. viii DAFTAR GAMBAR ……………………………………………………. x PENDAHULUAN
Latar Belakang …………………………………………………… 1Tujuan Penelitian ………………………………………………… 3Manfaat Penelitian ……………………………………………….. 4Ruang Lingkup Penelitian ……………………………………….. 5
TINJAUAN PUSTAKA Taksonomi, Morfologi, Anatomi, dan Nilai Penting Nenas …………….. 6Perbanyakan In Vitro …………………………………………………….. 9Konservasi In Vitro ………………………………………………........... 12Fenomena Keragaman Somaklonal ……………………………………… 13Deteksi Keragaman Somaklonal …………………………………………. 14 STUDI MORFOGENESIS EKSPLAN NENAS YANG DIINDUKSI OLEH AUKSIN DAN SITOKININ
Abstrak ………………………………………………………….... 16Abstract …………………………………………………………... 16Pendahuluan ……………………………………………………… 17Bahan dan Metode ……………………………………………….. 19Hasil dan Pembahasan …………………………………………… 20Simpulan …………………………………………………………. 29Daftar Pustaka ……………………………………………………. 30
ORGANOGENESIS DAN EMBRIOGENESIS SOMATIK TIDAK LANGSUNG PADA NENAS YANG DIINDUKSI OLEH 2,4-D
Abstrak ………………………………………………………….... 32Abstract …………………………………………………………... 32Pendahuluan ……………………………………………………… 33Bahan dan Metode ……………………………………………….. 35Hasil dan Pembahasan …………………………………………… 38Simpulan …………………………………………………………. 48Daftar Pustaka ……………………………………………………. 48
INDUKSI EMBRIOGENESIS SOMATIK TIDAK LANGSUNG PADA NENAS MELALUI PENGGUNAAN PIKLORAM
Abstrak ………………………………………………………….... 51Abstract …………………………………………………………... 52Pendahuluan ……………………………………………………… 52Bahan dan Metode ……………………………………………….. 54Hasil dan Pembahasan …………………………………………… 57Simpulan …………………………………………………………. 70Daftar Pustaka ……………………………………………………. 71

PEMBENTUKAN BENIH SINTETIK NENAS DAN KONSERVASI IN VITRO SECARA PERTUMBUHAN MINIMAL
Abstrak ………………………………………………………….... 74Abstract …………………………………………………………... 74Pendahuluan ……………………………………………………… 75Bahan dan Metode ……………………………………………….. 76Hasil dan Pembahasan …………………………………………… 79Simpulan …………………………………………………………. 90Daftar Pustaka ……………………………………………………. 90
KARAKTERISASI MORFOLOGI DAN MOLEKULER BIAKAN IN VITRO DAN BIBIT NENAS
Abstrak ………………………………………………………….... 93Abstract …………………………………………………………... 94Pendahuluan ……………………………………………………… 94Bahan dan Metode ……………………………………………….. 96Hasil dan Pembahasan …………………………………………… 99Simpulan …………………………………………………………. 112Daftar Pustaka ……………………………………………………. 113
EVALUASI KERAGAMAN BIBIT NENAS DARI POPULASI LAMA DAN PEMBENTUKAN POPULASI BARU SERTA DETEKSI DINI KERAGAMAN SOMAKLONAL
Abstrak ………………………………………………………….... 116Abstract …………………………………………………………... 116Pendahuluan ……………………………………………………… 117Bahan dan Metode ……………………………………………….. 119Hasil dan Pembahasan …………………………………………… 122Simpulan …………………………………………………………. 133Daftar Pustaka ……………………………………………………. 134
PEMBAHASAN UMUM ……………………………………………….. 136 SIMPULAN DAN SARAN ……………………………………………… 149 DAFTAR PUSTAKA ……………………………………………………. 150

DAFTAR TABEL
Halaman
1 Status penelitian kultur in vitro nenas di mancanegara ........................ 10
2 Status penelitian kultur in vitro nenas di Indonesia dan peluang risetnya ………………………………………………………............. 11
3 Data produksi buah-buahan di Indonesia tahun 1995-2009 …………. 18
4 Penampilan kalus embriogenik nenas kultivar Smooth Cayenne pada beberapa macam formulasi media, 2 bulan setelah subkultur .............. 45
5 Pengaruh media dasar dan adenin sulfat terhadap pertumbuhan FETs nenas kultivar Smooth Cayenne ……………………………………... 45
6 Respon kalus nenas kultivar Smooth Cayenne terhadap media induksi kalus embriogenik pada dua macam kondisi inkubasi yang berbeda, 1 bulan setelah subkultur …………………………………………….. 61
7 Perkembangan ETs nenas kultivar Smooth Cayenne pada media MS yang mengandung Kn 1 mg l-1, 2 bulan setelah subkultur 65
8 Pengaruh pencahayaan dan formulasi media terhadap pembentukan suspensi sel nenas kultivar Smooth Cayenne ....................................... 68
9 Tingkat pencoklatan FETs nenas kultivar Smooth Cayenne pada media regenerasi, 1 bulan setelah subkultur …………………………. 69
10 Tingkat regenerasi FETs nenas kultivar Smooth Cayenne pada media regenerasi, 1 bulan setelah subkultur …………………………. 69
11 Jumlah embrio somatik dewasa yang terbentuk dari FETs nenas kultivar Smooth Cayenne, 1 bulan setelah subkultur ………………... 70
12 Pengaruh suhu penyimpanan terhadap pertumbuhan biakan tunas nenas kultivar Smooth Cayenne yang terenkapsulasi dalam Na-alginat 3% ………………………………………………………......... 83

13 Pengaruh suhu penyimpanan dan taraf manitol terhadap daya tembus biakan nenas kultivar Smooth Cayenne yang terenkapsulasi dengan Na-alginat 3% ………………………………………………………... 88
14 Pengaruh suhu penyimpanan dan taraf manitol terhadap daya hidup biakan nenas kultivar Smooth Cayenne yang terenkapsulasi dengan Na-alginat 3% ………………………………………………………... 89
15 Karakter dan sub karakter yang diamati pada fenotipe normal dan varian dari biakan in vitro nenas kultivar smooth Cayenne (umur 4 tahun) ………………………………………………………................ 98
16 Ciri-ciri spesifik dari varian yang muncul dari populasi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun ........................ 103
17 Proporsi varian yang muncul dalam populasi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun ……………………………... 104
18 Stabilitas fenotipe setiap individu varian nenas kultivar Smooth Cayenne dan regenerannya selama subkultur dan aklimatisasi ............ 107
19 Jumlah pita DNA yang teramplifikasi dan yang bersifat polimorfik
dalam analisis RAPD populasi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun …………………………………………….... 109
20 Proporsi variasi fenotipe dari populasi biakan in vitro nenas kultivar
Smooth Cayenne yang dihasilkan dari metode organogenesis (langsung dan tidak langsung serta embriogenesis somatik tidak langsung................................................................................................ 125
21 Rekapitulasi tingkat variasi dari biakan in vitro yang dihasilkan dari
tiga macam metode regenerasi dibandingkan dengan populasi biakan induk.....………………………………………………………............. 126
22 Proporsi bibit yang tidak berduri, berduri khimera, dan berduri
lengkap yang dihasilkan dari metode organogenesis langsung yang diinduksi oleh BA dan NAA................................................................ 126
23 Proporsi bibit nenas kultivar Smooth Cayenne dengan daun tidak
berduri, berduri khimera, dan berduri lengkap yang dihasilkan dari metode organogenesis tidak langsung yang diinduksi oleh 2,4-D...................................................................................................... 127
24 Proporsi bibit nenas kultivar Smootha Cayenne dengan daun tidak
berduri, berduri khimera, dan berduri lengkap yang dihasilkan dari metode embriogenesis somatik yang diinduksi oleh pikloram ........…. 128

DAFTAR GAMBAR
Halaman
1 Diagram alir tahapan penelitian dalam studi pengembangan metode organogenesis dan embriogenesis somatik pada nenas serta deteksi dini untuk mereduksi keragaman somaklonal ……………………….... 6
2 Berbagai macam organ vegetatif tanaman nenas……………………… 8
3 Keragaan kultur nenas kultivar Smooth Cayenne yang berasal dari eksplan basal daun (1 bulan setelah tanam) …………………………... 21
4 Pengaruh tipe eksplan dan ZPT serta pengaruh interaksi antara tipe
eksplan dan ZPT terhadap morfogenesis eksplan basal daun nenas kultivar Smooth Cayenne pada media padat.............................……….. 23
5 Penampilan kalus nenas pasca-perlakuan NAA dan tipe sel yang
menyusunnya ………………………………………………………….. 25 6 Pembelahan sel secara asimetris dari sel embriogenik yang diiduksi
oleh NAA …………………………………………………………....... 25 7 Pengaruh kombinasi sitokinin dan auksin pada media cair terhadap
pembentukan tunas, nodul, dan akar kultur in vitro nenas kultivar Smooth Cayenne yang berasal dari eksplan daun utuh dan potongan basal daun, umur 1.5 bulan …………………………………………… 27
8 Pengaruh kombinasi sitokinin dan auksin pada media cair terhadap
jumlah tunas kultur in vitro nenas kultivar Smooth Cayenne yang berasal dari eksplan daun utuh dan potongan basal daun ...... ................
28
9 Tahapan morfogenesis dari eksplan daun nenas kultivar Smooth
Cayenne pada media cair …………………………………………….... 29
10 Tahapan analisis histologi kalus embriogenik nenas, modifikasi metode Kiernan (1990) ………………………………………………... 37
11 Tahapan induksi kalus dari basal daun nenas kultivar Smooth Cayenne 38
12 Pengaruh 2,4-D terhadap pembentukan kalus dan bobot basah kalus
nenas kultivar Smooth Cayenne ………………………………………. 39
13 Respon basal daun nenas kultivar Smooth Cayenne terhadap perlakuan 2,4-D ..................................................... …………………… 40

14 Penampilan tunas in vitro nenas kultivar Smooth Cayenne hasil perlakuan 2,4-D pada tahap regenerasi dan elongasi ............................. 41
15 Pengaruh 2,4 terhadap persentase pembentukan tunas normal, tunas
abnormal, dan konversi dari tunas abnormal ke tunas normal.....………………………………………………......................... 42
16 Penampilan jaringan embriogenik nenas kultivar Smooth Cayenne
pada berbagai macam media ………………………………………….. 43
17 Tahapan embriogenesis somatik nenas kultivar Smooth Cayenne ........ 44
18 Pengaruh faktor tunggal dan pengaruh interaksi antara media induksi kalus embriogenik dan media regenerasi terhadap regenerasi FETs nenas kultivar Smooth Cayenne ............................................................. 46
19 Perbedaan perkembangan embrio somatik nenas kultivar Smooth
Cayenne umur 8 bulan pada media regenerasi ………………………... 47
20 Pengaruh pikloram terhadap pembentukan kalus dan bobot basah dari eksplan basal daun nenas kultivar Smooth cayenne …………………... 58
21 Respon basal daun nenas kultivar Smooth Cayenne terhadap
perlakuan pikloram dan pengamatan mikroskopisnya ………………... 59
22 Pengamatan mikroskopis dari ECCs nenas kultivar Smooth Cayenne setelah ditanam pada media induksi kalus embriogenik ……………… 60
23 Penampilan ETs dan FETs nenas kultivar Smooth Cayenne ...……….. 62
24 Pengaruh faktor tunggal (media dan pencahayaan) terhadap tingkat pencoklatan, persentase pembentukan nodul, persentase pembentukan embrio, dan jumlah embrio prematur (2 bulan setelah subkultur) ......... 64
25 Regenerasi ETs nenas kultivar Smooth Cayenne pada media MS
dengan penambahan Kn 1 mg l-1 ……………………………………… 66
26 Pengaruh faktor pencahayaan dan media induksi kalus embriogenik serta pengaruh interaksinya terhadap pembentukan embrio somatik nenas kultivar Smooth Cayenne ……………………..……………...… 66
27 Tahapan perkembangan embrio somatik nenas kultivar Smooth
Cayenne dari struktur FETs yang diinduksi oleh pikloram .................... 67
28 Analisis histologi embrio somatik dari kultur suspensi sel nenas kultivar Smooth Cayenne ....................................................................... 68

29 Pengaruh BA dan NAA terhadap pertumbuhan eksplan batang semu nenas kultivar Smooth Cayenne yang dienkapsulasi dalam Na-alginat 3%, 1 bulan masa inkubasi …………………………………………… 80
30 Keragaan eksplan batang semu nenas kultivar Smooth Cayenne yang
dienkapsulasi dengan Na-alginat 3% (1 bulan masa inkubasi) .............. 81
31 Proporsi pembentukan planlet, akar, dan nodul dari eksplan basal daun nenas kultivar Smooth Cayenne yang diinduksi oleh BA 0.5 mg l-1 dan NAA 0.5 mg l-1, 2 bulan periode inkubasi ............................................. 82
32 Pengaruh kombinasi BA dan NAA terhadap pertumbuhan eksplan
basal daun nenas kultivar Smootha Cayenne yang terenkapsulasi dalam Na-alginat 3% ………………………………………………….. 82
33 Penampilan tunas nenas kultivar Smooth Cayenne yang dienkapsulasi
dengan alginat 3% yang mengandung paklobutrazol dan disimpan pada suhu yang berbeda ………………………………………………. 84
34 Daya hidup, daya regenerasi dan jumlah tunas nenas kultivar Smooth
Cayenne pada tahap pemulihan pasca penyimpanan dengan paklobutrazol .......................................................................................... 86
35 Penampilan tunas nenas kultivar Smooth Cayenne pada tahap
pemulihan (1 bulan) pasca-penyimpanan dengan paklobutrazol ........... 86
36 Penampilan kapsul embrio somatik nenas kultivar Smooth Cayenne (4 bulan periode simpan) yang disimpan dengan menggunakan manitol pada suhu penyimpanan yang berbeda ...................................... 89
37 Keragaan varian dari populasi biakan in vitro nenas kultivar Smooth
Cayenne umur 4 tahun ………………………………………………… 100
38 Dendogram hasil karakterisasi morfologi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun, berdasarkan metode (SAHN)-UPGMA (r=0.83) …………………………………………................... 102
39 Kualitas DNA hasil isolasi sampel daun nenas kultivar Smooth
Cayenne dengan metode CTAB ………………………………………. 107
40 Profil DNA varian nenas kultivar Smooth Cayenne, hasil amplifikasi PCR dengan penanda RAPD ………………………………………….. 111
41 Dendogram hasil karakterisasi molekuler biakan in vitro nenas
kultivar Smooth Cayenne umur 4 tahun, menggunakan metode RAPD, berdasarkan analisis (SAHN)-UPGMA (r=0.97) ……………. 112

42 Tingkat keragaman bibit nenas kultivar Smooth Cayenne yang dihasilkan dari tiga metode regenerasi ………………………………... 123
43 Profil DNA biakan nenas kultivar Smooth Cayenne yang diekstraksi
dari populasi biakan induk ………………………... ............................. 129
44 Dendogram hasil evaluasi keragaman genetik (dengan penanda RAPD, r=0.98) bibit nenas kultivar Smooth Cayenne populasi lama yang diperbanyak secara organogenesis langsung, organogenesis tak langsung, dan embriogenesis somatik ……………………………….. 130
45 Profil DNA dari populasi baru biakan in vitro nenas kultivar Smooth
Cayenne ……………………………………………………………….. 132
46 Dendogram hasil deteksi dini keragaman somaklonal biakan in vitro nenas kultivar Smooth Cayenne dari populasi baru dengan penanda RAPD (r=0.96) ………………………………………………………... 133
47 Tahapan lengkap embriogenesis somatik nenas kultivar Smooth
Cayenne ……………………………………………………………… 140
48 Tahapan lengkap organogenesis nenas kultivar Smooth Cayenne …... 141

DAFTAR ISTILAH DAN SINGKATAN
Tanaman CAM fakultatif : tanaman yang berfotosintesis tipe CAM ketika lingkungan tumbuhnya suboptimal dan berfotosintesis tipe C3 atau C4 ketika lingkungan tumbuhnya optimal (nenas bertipe C3-CAM)
Propagul : bagian tanaman yang dapat digunakan sebagai bahan perbanyakan
Mikropropagasi : teknik perbanyakan mikro untuk produksi bibit
Aklimatisasi : proses penyesuaian fisiologi bibit tanaman dari kondisi in vitro ke kondisi ex vitro
Multiplikasi : tingkat penggandaan tunas atau bibit
Proliferasi tunas : teknik perbanyakan in vitro melalui penggandaan tunas aksilar
Organogenesis : proses pembentukan organ yang bersifat unipolar (satu kutub) berupa tunas atau akar
Embriogenesis somatik : proses pembentukan embrio yang bersifat bipolar (dua kutub), diregenerasikan dari sel-sel somatik
Suspensi sel : kultur sel yang terdispersi di dalam media cair yang selalu digoyang
Pertumbuhan minimal : suatu teknik konservasi in vitro untuk penyimpanan jangka menengah melalui modifikasi unsur hara, sumber karbon, suhu penyimpanan, penggunaan zat penghambat tumbuh dan retardan
Keragaman somaklonal : keragaman yang terjadi pada sel-sel somatik, disebabkan oleh mutasi atau perubahan sekuens DNA, aktivasi atau pembungkaman gen
Syncarp : sejumlah bakal buah yang membentuk satu ruang, daun fertil pendukung makrospora berupa bakal biji (ovulum) yang secara kolektif membentuk putik
Eksplan : bahan tanaman yang digunakan dalam penanaman secara in vitro
BA : benzyl adenine, sitokinin turunan purin
Kn : kinetin, sitokinin turunan purin
TDZ : thidiazuron, secara fisiologis fungsinya sama dengan sitokinin, turunan fenilurea
IAA : indole acetic acid, auksin alami
NAA : naphthalene acetic acid
IBA : indole butyric acid

Gln : glutamine, asam amino
Arg : arginine, asam amino
Gly : glycine, asam amino
CH : casein hydrolysate, senyawa organik kompleks
AdS : adenine sulfate, senyawa organik sumber nitrogen
2,4-D : 2,4-dichlorophenoxy acetic acid
Pikloram : 4-amino 3,5,6-trichloropicolinic acid
Sel embriogenik : sel yang akan berdiferensiasi membentuk embrio melalui pembelahan yang bersifat asimetris
Pro-embrio : calon embrio yang terbentuk dari sel embriogenik
PEM : pre-embryogenic mass, kumpulan atau massa proembrio
ETs : embryogenic tissues
FETs : friable embryogenic tissues
Planlet : tanaman utuh yang telah lengkap mempunyai tunas dan akar, dihasilkan dari regenerasi secara in vitro
Nekrosis : gejala matinya sel dan berasosiasi dengan degradasi klorofil yang disebabkan oleh oksidasi fenol
Paklobutrazol : senyawa sintetik golongan triazol yang berfungsi sebagai retardan
Manitol : gula alkohol yang dapat berfungsi sebagai regulator osmotik (pengatur tekanan osmotikum) media
Amplifikasi : proses penggandaan molekul DNA
EtBr : ethidium bromide
CTAB : cetyl trimethyl amonium bromide
PCR : polymerase chain reaction, proses polimerisasi DNA secara in vitro
Denaturasi : proses pemisahan DNA dari ikatan ganda menjadi utas tunggal
Elektroforesis : teknik pemisahan molekul berdasarkan gerakan yang berbeda pada medan listrik
RAPD : randomly amplified polymorphism DNA
Epigenetik : perubahan sifat yang tidak disebabkan oleh perubahan sekuens DNA, bersifat dapat balik
Monomorfik : pola pita DNA yang sama antar individu
Polimorfik : pola pita DNA yang berbeda antar individu

PENDAHULUAN
Latar Belakang
Nenas (Ananas comosus L. Merr.) merupakan tanaman penting di daerah
tropis, termasuk di Indonesia. Berdasarkan data produksi buah-buahan, komoditas
nenas menempati peringkat ketiga atau keempat setelah pisang, mangga, dan jeruk
(BPS 2009). Tanaman ini bersifat toleran terhadap cekaman kekeringan dan
kemasaman sehingga berpeluang besar dikembangkan di lahan-lahan marginal,
termasuk lahan gambut (Sagiman 2007).
Menurut Ochse et al. (1961) serta Coppens d’Eeckenbrugge dan Leal (2003),
tanaman nenas pada umumnya diperbanyak secara vegetatif dengan menggunakan
butt atau stump, ratoon, sucker, basal slips, hapas, crown slips, dan crown atau
mahkota. Sayangnya, jenis dan jumlah propagul tersebut sangat terbatas pada
kultivar Smooth Cayenne (sucker dan mahkota) sehingga perbanyakan secara
konvensional perlu dukungan teknologi lainnya, terutama ketika diperlukan bibit
dalam jumlah yang besar. Penerapan teknologi kultur in vitro akan berguna bagi
penyediaan materi genetik calon varietas unggul maupun perbanyakan varietas
unggul yang baru saja dilepas, serta pembukaan lahan baru untuk pengembangan
komoditas tersebut secara luas.
Di mancanegara, beberapa kultivar nenas telah dikembangkan secara
komersial, antara lain Cayenne (Champaka, Giant Kew, Sarawak, Maipuri,
Esmeralda, Claire, dan Typhon), Queen, Spanish (Red Spanish dan Singapore
Spanish), Perola (Pernambuco atau Abacaxi), Manzana, dan Perolera (atau Motilana)
(IBPGR 1991; Chan et al. 2003). Di antara beberapa kultivar tersebut, Queen dan
Smooth Cayene merupakan kultivar yang banyak dikembangkan di Indonesia.
Teknik mikropropagasi nenas telah diterapkan secara komersial oleh pihak
industri di mancanegara (Smith et al. 2003), tetapi industri di Indonesia masih
bertahan pada teknik perbanyakan konvensional. Teknik mikropropagasi masih
dipandang perlu diterapkan untuk menyediakan bibit dalam jumlah besar, terutama
untuk varietas baru hasil persilangan, seleksi, mutasi, dan rekayasa genetika

(Firoozabady dan Moy 2004; Nursandi et al. 2005). Secara praktis, mikropropagasi
digunakan untuk memantapkan blok multiplikasi bagi penyediaan bahan tanaman
perbanyakan konvensional dalam skala yang lebih luas. Selain itu, teknik
mikropapagasi dapat diterapkan untuk skrining varian sebelum diterapkannya
metode perbanyakan secara konvensional (Smith et al 2003). Dalam rangka
peningkatan produksi, populasi tanaman akan ditingkatkan dari 40.000 tanaman/ha
menjadi 100.000 tanaman/ha sehingga keperluan penyediaan bibit akan semakin
tinggi (Suminar 2010).
Dalam teknik kultur in vitro, terjadinya keragaman genetik atau keragaman
somaklonal (off-type) kadang-kadang tidak dapat dihindari. Keragaman somaklonal
merupakan kejadian abnormalitas yang terjadi oleh karena mutasi atau perubahan
sekuens, aktivasi gen, dan pembungkaman gen (Kaeppler et al. 2000). Keragaman
tersebut dapat dideteksi melalui karakterisasi morfologi (Noor et al. 2009; Somsri et
al. 2009; Zhao et al. 2005; Podwyszyńska 2005), sitologi (Al-Zahim et al. 1999),
biokimia, fisiologi (Perez et al. 2011), dan molekuler (Chen et al. 1998; Al-Zahim et
al. 1999; Soniya et al. 2001; Perez et al. 2011).
Wakasa (1979) telah melakukan pengamatan bibit nenas hasil kultur in vitro
1.5 tahun dalam periode in vitro, 14-22 bulan pada tahap aklimatisasi, dan 1 tahun
periode pertumbuhan tanaman melalui pengamatan karakter morfologi, yaitu duri
pada daun, warna daun, sekresi lilin pada permukaan daun bawah, dan corak daun.
Diperlukan waktu sekitar 4 tahun untuk mendeteksi keragaman yang terjadi pada
bibit hasil perbanyakan secara in vitro tersebut. Nursandi (2006) melaporkan bahwa
perlakuan in vitro dapat menimbulkan keragaman tanaman nenas hingga 88% untuk
karakter daun varigata, roset, dan kerdil, namun sebagian besar tanaman tersebut
mampu berubah menjadi normal setelah umur tertentu (25 minggu setelah tanam),
yang diduga sebagai epigenetik.
Untuk efisiensi waktu, biaya, dan area maka evaluasi stabilitas sifat tersebut
sebaiknya dilakukan secara dini, sejak biakan masih di dalam botol sebelum tanaman
tersebut ditanam di lapang. Selain itu, penggunaan penanda molekuler menjadi
sangat penting dalam membantu upaya mendeteksi keragaman somaklonal secara
dini.

Pada tanaman nenas, deteksi penyimpangan secara molekuler banyak
diterapkan dengan menggunakan penanda RAPD (Random Amplified Polymorphism
DNA). Soneji et al. (2002) telah mengkarakterisasi bibit nenas kultivar Queen untuk
sifat daun berduri dan tidak berduri dengan penanda RAPD. Feuser et al. (2003) juga
menganalisis ketepatan genetik planlet nenas kultivar Amarelinho (grup Perola) dan
Santos et al. (2008) melakukan evaluasi keragaman genetik dari biakan in vitro nenas
hias (Ananas comosus var. bracteatus) dengan menggunakan penanda RAPD.
Hingga saat ini, belum terdapat laporan tentang analisis keragaman somaklonal
terhadap biakan dan bibit nenas hasil perbanyakan in vitro yang dikulturkan dalam
periode yang panjang.
Tujuan Penelitian
Tujuan umum penelitian ini adalah untuk memperoleh metode regenerasi yang
efektif (tingkat multiplikasi tinggi dan tingkat keragaman rendah), termasuk metode
pembentukan benih sintetik tanaman nenas, metode deteksi dini keragaman
somaklonal untuk eliminasi varian yang tak diharapkan, dan metode reduksi
keragaman somaklonal. Tujuan khusus penelitian ini adalah:
1. Mempelajari morfogenesis eksplan nenas kultivar Smooth Cayenne dalam
merespon zat pengatur tumbuh (ZPT) dan konsistensi media
2. Memperoleh informasi tentang pengaruh kombinasi sitokinin (benzyl adenine/
BA) dan auksin (naphthalene acetic acid/NAA, indole acetic acid/IAA, indole
butiric acid/IBA) terhadap regenerasi nenas kultivar Smooth Cayenne secara
organogenesis langsung
3. Mempelajari pengaruh auksin non herbisida NAA dan auksin herbisida 2,4-
dichlorophenoxyacetic acid (2,4-D) atau 4-amino 3,5,6-trichloropicolinic acid
(pikloram) terhadap induksi organogenesis dan embriogenesis somatik tidak
langsung pada nenas kultivar Smooth Cayenne
4. Mempelajari pengaruh suhu dan paklobutrazol atau manitol terhadap daya hidup,
daya simpan, dan daya regenerasi eksplan nenas kultivar Smooth Cayenne yang
dienkapsulasi dengan natrium alginat 3%
5. Mempelajari karakter morfologi dan molekuler biakan in vitro dan bibit nenas
kultivar Smooth Cayenne

6. Mengetahui tingkat keragaman biakan dan bibit nenas kultivar Smooth Cayenne
yang dihasilkan dari metode organogenesis langsung dan tidak langsung, serta
embriogenesis somatik tidak langsung
7. Mendeteksi keragaman molekuler biakan in vitro dari populasi yang baru dibentuk
untuk menentukan metode perbanyakan yang paling efektif.
Manfaat Penelitian
Hasil penelitian ini tidak hanya bermanfaat bagi perkembangan ilmu dasar,
namun juga bagi perkembangan ilmu terapan. Pada aspek ilmu dasar, penelitian ini
berhasil mengantisipasi fenomena dead-end pada eksplan yang berakar sehingga
mampu meregenerasikan tunas. Penelitian ini juga berhasil menjelaskan proses
embriogenesis somatik tanaman monokotil (khususnya tanaman nenas) secara
lengkap serta asal-usul pembentukan embrio somatik. Beberapa tahapan penting
dalam teori embriogenesis somatik berhasil terekam atau tervisualisasi (mulai dari
sel tunggal hingga terbentuknya planlet yang siap diaklimatisasi) sehingga akan
memudahkan pengguna dalam memahami proses tersebut maupun dalam menyusun
strategi penelitian yang berkaitan dengan proses tersebut. Penelitian ini juga
berkontribusi dalam upaya menguak misteri asal-usul terjadinya keragaman
somaklonal pada tanaman nenas. Pada aspek ilmu terapan, hasil penelitian ini
bermanfaat bagi upaya penyediaan metode perbanyakan bibit yang efektif untuk
perbanyakan materi pemuliaan (calon varietas baru) maupun produksi bibit secara
masal setelah pelepasan varietas baru serta untuk penyediaan bibit yang memadai
bagi pembukaan lahan-lahan baru. Metode embriogenesis somatik yang telah
dihasilkan dapat dimanfaatkan untuk melakukan rekayasa seluler (berbasis pada
teknik kultur in vitro) dan molekuler (berbasis pada teknik transformasi genetik)
untuk mendukung program pemuliaan tanaman nenas. Metode pembentukan benih
sintetik akan berguna bagi penyediaan benih yang aplikatif dan untuk konservasi in
vitro (secara pertumbuhan minimal) dalam upaya pelestarian koleksi plasma nutfah
dan nomor-nomor hasil persilangan sehingga terhindar dari resiko hilangnya
genotipe oleh cekaman lingkungan biotik dan abiotik di lapang serta dapat
menghemat tempat, waktu, biaya, dan tenaga. Manfaat lainnya adalah mendapatkan
metode deteksi keragaman somaklonal tanaman nenas secara dini untuk eliminasi

varian yang tidak diharapkan, sebelum bibit ditanam di lapang sehingga dapat
meningkatkan efisiensi produksi bibit. Selain itu, varian-varian yang dihasilkan
dalam penelitian ini juga dapat dimanfaatkan sebagai bahan pemuliaan tanaman atau
bahkan sebagai kandidat klon superior. Secara umum, hasil penelitian ini akan
bermanfaat bagi pengembangan komoditas nenas di Indonesia.
Ruang Lingkup Penelitian
Untuk mencapai tujuan penelitian tersebut maka dibuat suatu strategi dengan
serangkaian kegiatan penelitian secara komprehensif. Penelitian ini diawali dengan
studi karakterisasi morfologi dan molekuler dari populasi biakan in vitro lama
(berumur 4 tahun) yang selanjutnya digunakan sebagai biakan induk. Evaluasi
keragaman fenotipe bibit di rumah kaca juga dilakukan terhadap populasi tanaman
dari biakan induk tersebut. Penelitian dilanjutkan dengan melakukan studi
morfogenesis eksplan pada media padat dan cair secara organogenesis langsung
(yang diinduksi oleh auksin dan sitokinin), studi regenerasi secara organogenesis dan
embriogenesis somatik secara tidak langsung dengan menggunakan eksplan dari
biakan induk (yang diinduksi oleh auksin herbisida). Studi morfogenesis dilakukan
pula terhadap eksplan (batang semu, basal daun, tunas in vitro, dan embrio somatik)
yang dienkapsulasi dengan natrium alginat untuk pembentukan benih sintetik dan
konservasi in vitro secara pertumbuhan minimal melalui penerapan modifikasi suhu
penyimpanan dan penggunaan paklobutrazol atau manitol. Selain itu, juga dilakukan
evaluasi fenotipe dan stabilitas genetik biakan in vitro dan bibit aklimatisasi hasil
ekstraksi dari biakan induk menggunakan 3 macam metode regenerasi. Setelah itu,
dilakukan pembentukan populasi biakan in vitro yang baru (0 tahun) serta
pendeteksian keragaman somaklonal secara morfologi dan molekuler. Secara umum,
ruang lingkup kegiatan penelitian ini dijelaskan pada diagram alir penelitian
(Gambar 1).



Karakterisasi morfologidan molekuler biakanin vitro dan bibit nenas
Karakterisasi morfologibiakan in vitro
Evaluasi fenotipe bibit dirumah kaca
Karakterisasi molekulerdenganmetodeRAPD
Subkulturdan evaluasifenotipe individu varian
Studi morfogenesiseksplan
Organogenesis langsung padamedia
padat
Induksi akardanaklimatisasi di rumah
kaca
Evaluasi fenotipe dananalisis molekuler
denganmetodeRAPD
Organogenesis langsung padamedia
cair
Pemilihan metodeterbaik
Studi organogenesis danembriogenesis tidak langsung
menggunakan 2,4-D
Induksi kalus
Induksi akar danaklimatisasi di rumah
kaca
Evaluasi fenotipe dananalisis molekuler
denganmetodeRAPD
Regenerasi kalusmenjadi tunas melaluijalur organogenesis
Induksi kalusembriogenik
Regenerasi kalusembriogenikmenjadi
embrio somatik
Aklimatisasi di rumahkaca
Evaluasi fenotipe dananalisis molekuler
denganmetodeRAPD
Studi embriogenesissomatik tidak langsungmenggunakan pikloram
Induksi kalusembriogenik
Regenerasi kalusembriogenikmenjadi
embrio somatik
Aklimatisasi di rumahkaca
Proliferasi kalusembriogenik
Evaluasi fenotipe dananalisis molekuler
denganmetodeRAPD
Pembentukan benihsintetik dan
pertumbuhan minimal
Studi enkapsulasi: pengaruh jenis eksplan
dan pra-perlakuan
Pertumbuhan minimal denganmanitol dan
modifikasi suhu
Pemulihandanregenerasi
Pertumbuhan minimal denganpaklobutrazoldan modifikasi suhu
Induksi akardanaklimatisasi di rumah
kaca
Evaluasi keragaman bibitdari 3 metode regenerasidari populasi lama (induk)
Evaluasi keragaman bibithasil organogenesis
langsung
Evaluasi keragaman bibithasil embriogenesis somatik
tidak langsung
Evaluasi ketepatangenetikdenganmetodeRAPD
Evaluasi keragaman bibithasil organogenesis tidak
langsung
Sterilisasi dan inisiasitunas in vitro
Regenerasi secaraorganogenesis
langsung
Evaluasi fenotipebiakan in vitro
Isolasi eksplanbasal daun
Pembentukan populasi barubiakan in vitro dan deteksi
dini keragaman somaklonal
Deteksi dini secaramolekulerdengan
metode RAPD
Populasi lama biakan in vitro (umur 4 tahun)
Gambar 1. Diagram alir tahapan penelitian dalam studi pengembangan metode organogenesis dan embriogenesis pada nenas serta deteksi dini untuk
mereduksi keragaman somaklonal.


TINJAUAN PUSTAKA
Taksonomi, Morfologi, Anatomi, dan Nilai Penting Nenas
Ananas comosus (L.) Merr. termasuk dalam monocotyledonae, ordo
Bromeliales, famili Bromeliaceae, dan subfamily Bromeliadeae. Nenas
merupakan tanaman herbaseus perenial. Batangnya memiliki internodus yang
sangat pendek (1-10 mm tergantung pada posisinya) sehingga tampak kompak
dan roset. Jaringan pembuluh yang padat memisahkan korteks dan stele yang
terutama terdiri atas xilem dengan sedikit floem. Tanaman nenas mempunyai
filotaksi yang bervariasi antara 3/8 hingga 5/13 dan 8/21 ketika berbunga. Daun
memiliki lapisan kutikula yang tebal dan jaringan penyimpan air pada bagian
epidermis atas. Bagian abaksial daun banyak mengandung trikoma yang
memberikan warna keperakan dan mampu meningkatkan pantulan cahaya. Selain
itu, densitas stomata daun agak rendah, yaitu sekitar 80 stomata mm-2.
Keberadaan kutikula, jaringan penyimpan air, jarangnya stomata, dan banyaknya
trikoma menyebabkan tanaman nenas bersifat ekonomis dalam menggunakan air
(Coppens d’Eeckenbrugge dan Leal 2003).
Nenas merupakan tanaman penting di daerah tropis. Berdasarkan data
produksi buah-buahan di Indonesia, komoditas nenas menempati peringkat ketiga
atau keempat setelah pisang, mangga, dan jeruk (BPS 2009). Sebagian besar buah
nenas dikonsumsi sebagai buah segar (buah meja) dan buah kaleng. Buah nenas
adalah sumber berbagai macam mineral dan vitamin seperti Mn, Cu, Ca, N, P, Fe,
vitamin C, B1 (tiamin), B2 (riboflavin), B3 (niasin), B6 (piridoksin), asam
askorbat, karoten dan serat. Nenas juga mengandung enzim bromelain yang
berfungsi untuk berbagai macam pengobatan seperti detoksifikasi liver, antitumor
dan antiradang. Selain itu, daun nenas juga dapat dimanfaatkan sebagai sumber
untuk kertas dan tekstil (Baez et al. 2007; Department of Health and Ageing 2008;
Jacques et al. 2008).
Menurut Ochse et al. (1961) dan Bartholomew et al. (2003), tanaman nenas
pada umumnya diperbanyak secara vegetatif. Beberapa macam organ vegetatif
yang digunakan sebagai bahan perbanyakan adalah: (1) butt atau stump, yaitu

batang utama dari tanaman dewasa; (2) ratoon, yaitu tunas yang muncul pada
pangkal batang; (3) sucker, yaitu tunas-tunas pada ketiak daun; (4) basal slips,
yaitu cabang-cabang yang muncul dari dasar buah; (5) hapas, yaitu tunas yang
muncul dari dasar tangkai buah, (6) crown slips, yaitu cabang-cabang yang
muncul dari ujung buah; dan (7) crown atau mahkota, tunas yang muncul dari
bagian atas buah (Gambar 2).
6
5
4
3
21
Sumber: Ochse et al. (1961)
Gambar 2. Berbagai macam organ vegetatif tanaman nenas: batang pokok (1), ratoon (2), aerial sucker (3), basal slip (4), crown slip (5), dan mahkota (6).
Perbanyakan dengan cara konvensional dilaporkan dapat menghasilkan bibit
dalam jumlah yang terbatas dengan ukuran yang bervariasi dan umur panen yang
bervariasi pula sehingga pemanenan tidak dapat dilakukan secara serempak.
Coppens d’Eckenbrugge dan Leal (2003) serta Ochse et al. (1961) melaporkan
bahwa bibit nenas yang berasal dari sucker memiliki umur panen 18–20 bulan,
ratoon 12–14 bulan, slip 20 bulan, crown 22–24 bulan, dan collar 14-16 bulan.

Perbanyakan in Vitro
Di mancanegara, teknik kultur in vitro tanaman nenas telah berkembang
(Tabel 1) mulai dari proliferasi tunas in vitro (Wakasa 1979; Firoozabady dan
Gutterson 2003; Khan et al. 2004; Danso et al. 2008), organogenesis (Sripaoraya
et al. 2003; Firoozabady dan Moy 2004; Hamasaki et al. 2005), embriogenesis
somatik (Sripaoraya et al. 2003; Firoozabady dan Moy 2004; Silva et al. 2008),
hingga kultur suspensi sel (Sripaoraya et al. 2003) dan teknologi benih sintetik
untuk konservasi jangka pendek (Gangopadhyay et al. 2004). Di Indonesia, teknik
perbanyakan in vitro tanaman nenas masih terbatas dalam hal teknik proliferasi
tunas nenas kultivar Queen (Nursandi et al. 2003; Purnamaningsih et al 2009),
teknik etiolasi nenas kultivar Smooth Cayenne (Nursandi et al. 2005), dan teknik
mutagenesis nenas kultivar Smooth Cayenne (Suminar 2010), sedangkan teknik
embriogenesis somatik dan benih sintetik masih belum dikaji (Tabel 2).
Benih sintetik didefinisikan sebagai embrio somatik yang berada di dalam
mantel (kapsul) sehingga sifatnya mirip dengan benih zigotik (Redenbaugh 1992).
Mantel tersebut dapat bertindak sebagai endosperma yang mengandung sumber
karbon, nutrisi, zat pengatur tumbuh, dan agen anti mikroba. Dewasa ini, definisi
benih sintetik dikembangkan lebih lanjut karena eksplan yang digunakan tidak
terbatas pada embrio somatik melainkan juga tunas terminal, tunas aksilar,
segmen buku, dan jaringan meristematik lainnya. Selain untuk tujuan perbanyakan
bibit, teknologi benih sintetik juga dapat dimanfaatkan untuk tujuan konservasi
(Rai et al. 2009).

Tabel 1. Status penelitian kultur in vitro nenas di mancanegara
Aspek/Varietas Eksplan Hasil Pustaka
Proliferasi tunas Smooth Cayenne Syncarp, slip,
mahkota, tunas aksilar
Keragaman bibit dari syncarp 100%, slip 98%, mahkota 7%, dan tunas aksilar 34%
Wakasa 1979
Smooth Cayenne Basal daun 6000–8000 tunas dalam 6 bulan (dengan bioreaktor) dari 2 tunas inisial
Firoozabady & Gutterson (2003)
MD2 Clump (kumpulan 2–3 tunas)
Keragaman 4% Danso et al. (2008)
Organogenesis
Phuket Basal daun dari sucker
Keragaman 5.8% Sripaoraya et al. (2003)
Smooth Cayenne Basal daun & core (batang semu) dari mahkota
NAA dan BA menyebabkan organogenesis langsung; TDZ dan IBA menyebabkan organogenesis tak langsung
Firoozabady & Moy (2004)
Smooth Cayenne Basal daun Pemaparan glutamin berkorelasi dengan respon organogenik
Hamasaki et al (2005)
Moris dan Josepine
MGB (meristematic globular bodies)
NAA dapat menginisiasi dan memproliferasi kalus
Silva et al. (2008)
Embriogenesis somatik
Phuket Basal daun MS + pikloram 3 mg/L (12.4 µM) dapat mengiduksi kalus embriogenik dan embrio somatik
Sripaoraya et al. (2003)
Smooth Cayenne Basal daun & batang semu
Pikloram dapat menginduksi kalus embriogenik (15–30%) dalam 3 bulan; menghasilkan keragaman 21%
Firoozabady & Moy (2004)
Suspensi sel Phuket Sel embriogenik Dengan menggunakan
pikloram, sifat alami sel embriogenik terpelihara hingga 6 bulan (frekuensi subkultur setiap 7 hari)
Sripaoraya et al. (2003)
Penyimpanan secara pertumbuhan minimal Elite (varietas lokal Jailpaiguri, Bengal Barat)
Tunas in vitro MS bebas ZPT dengan suhu inkubasi 8 0C dapat menyimpan biakan selama 45 hari
Gangopadhyay et al. (2004)

Tabel 2. Status penelitian kultur in vitro nenas di Indonesia dan peluang risetnya
Aspek/Varietas Eksplan Hasil Pustaka
Proliferasi tunas
Queen Tunas in vitro MS + BA 4.4 uM + NAA 1.6 µM menghasilkan multiplikasi tunas tertinggi (26.3 tunas/eksplan); penggunaan TDZ menghasilkan struktur noduler; tanaman roset dapat tumbuh normal setelah 30 MST
Nursandi et al. (2003 dan 2005)
Smooth Cayenne Tunas in vitro Perlakuan paklobutrazol 0.1 mg/L tidak nyata dalam menurunkan variasi
Purnama-ningsih et al. (2009)
Etiolasi Smooth Cayenne Tunas in vitro MS + BA 4 mg/L menghasilkan
2–3 planlet/eksplan dalam 10 minggu
Nursandi et al. (2005)
Mutagenesis
Golden Pineapple
Kalus Iradiasi sinar gamma 15 Gy menginduksi variasi pertumbuhan tunas, fenotipe dan karakter molekuler dengan penanda ISSR.
Suminar (2010)
Peluang riset dan kebaruan
Studi organogenesis (tahapan lengkap organogenesis tanaman nenas)
Studi embriogenesis somatik tanaman monokotil (tahapan lengkap embriogenesis somatik tanaman nenas)
Pembentukan benih sintetik untuk perbanyakan dan penyimpanan in vitro (metode enkapsulasi eksplan tanaman nenas)
Penyimpanan secara pertumbuhan minimal biakan in vitro nenas secara enkapsulasi (metode penyimpanan in vitro dengan modifikasi suhu dan paklobutrazol atau manitol)
Karakterisasi morfologi biakan in vitro nenas umur 4 tahun (metode deteksi dini untuk eliminasi varian yang tak diharapkan selama proses mikropropagasi)
Reduksi keragaman somaklonal biakan in vitro nenas
Penelusuran asal-usul keragaman somaklonal pada tanaman nenas

Konservasi In Vitro
Teknik konservasi in vitro dapat dibedakan menjadi tiga macam: (1) teknik
penyimpanan pada media tumbuh, (2) teknik pertumbuhan minimal, dan (3)
teknik kriopreservasi (Mariska et al. 1996; Leunufna 2004). Teknik pertumbuhan
minimal disarankan untuk diterapkan untuk working collection atau active
collection, sedangkan teknik kriopreservasi diterapkan untuk base collection
(Withers 1985).
Pada teknik penyimpanan dengan media tumbuh, tidak diperlukan
penambahan zat penghambat tumbuh. Penyimpanan dengan cara tersebut akan
memerlukan tindakan subkultur yang frekuentif sehingga kurang menghemat
tenaga, waktu, dan biaya serta beresiko terhadap kontaminasi (Mariska et al.
1996). Selain itu, subkultur yang frekuentif juga beresiko terhadap timbulnya
keragaman somaklonal (Eeuwens et al. 2002).
Pada teknik pertumbuhan minimal, beberapa modifikasi media dan
lingkungan dapat diterapkan, antara lain penurunan temperatur lingkungan dan
intensitas cahaya (Hu dan Wang 1983; Withers 1985; Keller et al. 2006),
penggunaan regulator osmotik seperti sukrosa dan manitol (Withers 1985;
Bessembinder et al. 1993), penurunan taraf beberapa faktor esensial seperti
pengenceran media (Desbrunais et al. 1992), serta penggunaan zat penghambat
tumbuh seperti paclobutrazol, cycocel, dan ancymidol (Withers 1985). Dengan
penerapan teknik pertumbuhan minimal maka biakan dapat disimpan dalam
jangka menengah (bulanan hingga tahunan).
Di Indonesia, teknik pertumbuhan minimal tanaman nenas belum pernah
dilaporkan. Di mancanegara, dilaporkan bahwa teknik enkapsulasi tunas in vitro
mampu menyimpan biakan nenas selama 45 hari melalui aplikasi suhu 8 0C
dengan menggunakan media MS tanpa zat pengatur tumbuh (Gangopadhyay et al.
2004).

Fenomena Keragaman Somaklonal
Dalam teknik kultur in vitro, terjadinya keragaman genetik atau
keragaman somaklonal kadang-kadang tidak dapat dihindari. Keragaman
somaklonal merupakan kejadian abnormalitas yang terjadi secara sitologi, mutasi,
perubahan sekuens, aktivasi gen, dan silencing atau pembungkaman gen
(Kaeppler et al. 2000). Dari berbagai kajian pustaka, dilaporkan bahwa off types
dapat disebabkan oleh variasi dari tanaman donor (pre-existing variation)
(Wakasa 1979), periode in vitro yang panjang (Koornneef 1991; Masoud and
Hamta 2008), zat pengatur tumbuh (Bairu et al. 2009), subkultur yang frekuentif
(Eeuwens et al. 2002), dan tergantung pada genotipe tanaman (Zucchi et al.
2002). Untuk menghindari tingginya keragaman somaklonal maka diperlukan
strategi untuk memproduksi bibit dengan kesamaan identitas genetik yang tinggi.
Dengan demikian, metode deteksi keragaman somaklonal memegang peranan
penting dalam pengujian tersebut.
Melalui pengamatan karakter morfologi, Wakasa (1979) menemukan tingkat
keragaman genetik yang berbeda-beda pada tanaman nenas hasil perbanyakan
secara in vitro ketika digunakan sumber eksplan yang berbeda pula. Berdasarkan
penelitian tersebut, diketahui bahwa syncarp menghasilkan keragaman sebesar
100%, slip 98%, crown 7%, dan tunas aksilar 34%. Sripaoraya et al. (2001)
menemukan fenomena keragaman somaklonal sebesar 5.7-7% ketika
menggunakan eksplan berupa tunas aksilar dan tunas terminal sucker nenas
kultivar Phuket. Soneji et al. (2002a) melaporkan terjadinya keragaman sebesar
7.6% dari bibit hasil perbanyakan in vitro dengan menggunakan mata tunas
aksilar dorman yang berasal dari mahkota nenas kultivar Queen. Dengan
menggunakan jenis eksplan yang sama, Santos et al. (2008) menemukan kejadian
keragaman genetik sebesar 38% pada nenas hias. Melalui jalur organogensis,
Firoozabady dan Moy (2004) menghasilkan keragaman sebesar 4% sedangkan
melalui jalur embriogenesis somatik dihasilkan variasi sebesar 21%.
Penekanan keragaman somaklonal (off-type) ditujukan untuk mendapatkan
bibit yang serupa dengan induknya. Beberapa faktor yang berpengaruh terhadap
variasi somaklonal adalah sumber eksplan, periode pengkulturan, jumlah

subkultur, zat pengatur tumbuh, genotipe, komposisi media, tingkat ploidi, dan
khimera (Zucchi et al. (2002). Dari bebagai jenis eksplan nenas yang digunakan,
mahkota menghasilkan tanaman dengan tingkat keragaman yang paling rendah
dibandingkan dengan syncarp, slip, dan tunas aksilar (Wakasa 1979). Sripaoraya
et al. (2003) mensinyalir bahwa keragaman somaklonal dapat ditekan dengan
menggunakan jaringan somatik yang masih muda. Regenerasi secara langsung
dari basal daun terbukti menghasilkan tanaman nenas yang true-to-type. Di lain
pihak, Masoud dan Hamta (2008) menemukan fenomena bahwa makin panjang
periode in vitro kultur cengkeh maka frekuensi keragaman makin tinggi pula.
Bairu et al. (2006) melaporkan bahwa perbanyakan pisang Cavendish dengan
menggunakan BA pada taraf rendah (2,5 ppm) dan jumlah subkultur yang rendah
(maksimal 6 kali) menghasilkan peluang off type yang lebih rendah, yaitu 10%.
Ketika taraf BA ditingkatkan menjadi 7.5 ppm dan frekuensi subkultur dinaikkan
menjadi 10 kali maka peluang off type dapat melebihi 70%. Menurut Eeuwens et
al. (2002), tingginya tingkat abnormalitas bibit kelapa sawit hasil kultur in vitro
diduga karena intensifnya penggunaan auksin dalam konsentrasi yang tinggi,
frekuensi subkultur yang tinggi (interval waktu subkultur yang singkat), dan umur
embrioid.
Deteksi Keragaman Somaklonal
Perbanyakan bibit dan konservasi menghendaki stabilitas genetik bahan
tanaman yang diperlakukan. Deteksi penyimpangan bibit yang dihasilkan dari
teknik kultur in vitro dapat dilakukan melalui karakterisasi morfologi (Wakasa
1979; Noor et al. 2009; Somsri et al. 2009 ; Zhao et al. 2005; Podwyszyńska
2005; Suminar 2010), sitologi (Al-Zahim et al. 1999; Joachimiak dan Ilnicki
2003), biokimia (Feuser et al. 2003), fisiologi (Perez et al. 2011), dan molekuler
(Chen et al. 1998; Al-Zahim et al. 1999; Soniya et al. 2001; Soneji et al. 2002;
Feuser et al. 2003; Santos et al. 2008; Perez et al. 2011).
Wakasa (1979) mengamati adanya keragaman somaklonal pada nenas secara
morfologi, yaitu karakter daun berduri, daun hijau gelap, sekresi lilin pada
permukaan daun bawah, dan albino dengan daun belang-belang putih. Nursandi
(2006) juga melakukan evaluasi keragaman somaklonal terhadap bibit nenas hasil

kultur in vitro melalui pengamatan karakter morfologi. Hasil penelitian tersebut
menunjukkan bahwa perlakuan in vitro dapat menimbulkan keragaman tanaman
hingga 88% untuk karakter daun variegata, roset, dan kerdil, namun sebagian
besar tanaman tersebut mampu berubah menjadi normal setelah umur tertentu (25
MST).
Dalam analisis molekuler, Popluechai et al. (2007) menerapkan analisis
RAPD (Random Amplified Polymorphism DNA) untuk mengetahui hubungan
kekerabatan beberapa aksesi nenas. Penelitian tersebut telah berhasil
membuktikan bahwa 17 primer, yaitu OPA2, OPA3, OPA4, OPA9, OPA13,
OPC2, OPC5, OPC8, OPC9, OPC10, OPC11, OPC12, OPC14, OPC15, OPC16,
OPC19, dan OPC20 (Operon Tech. Alameda, USA) bersifat polimorfik (hingga
12 pita). Soneji et al. (2002b) juga menerapkan RAPD untuk menganalisis varian
nenas kultivar Queen yang berduri dan yang tidak berduri dengan menggunakan
primer OPA1, OPA2, OPA3, OPA4, OPA7, OPA8, OPA9, OPA11, OPA12,
OPA13, OPA15, OPA16, OPA17, OPA18, OPA19, OPF1, OPF4, OPF5, OPF6,
OPF8, OPF10, OPF13, OPJ4, OPJ5, OPJ7, OPJ8, OPJ9, OPJ10, OPJ11, OPJ12,
OPJ13, OPJ14, OPJ16, OPJ18, dan OPJ19. Santos et al. (2008) juga
menggunakan teknik RAPD untuk mengevaluasi variabilitas regeneran nenas hias
dan menghasilkan pita yang polimorfik dengan menggunakan primer OPA1,
OPA20, OPB1, OPB19, OPC19, OPF13, OPL17, OPM13, OPP16, OPT7,
OPV19, dan OPX3. Selain itu, marka ISSR (Inter Simple Sequence Repeat), yaitu
(CAG)5, (CAA)5, dan (GACA)4 (disintesis oleh Isogen) juga pernah diterapkan
pada tanaman nenas untuk mengevaluasi biakan pasca-penyimpanan in vitro
(Gangopadhyay et al. 2004).
Analisis molekuler terhadap tanaman nenas juga telah dilakukan di
Indonesia. Nursandi (2006) melaporkan bahwa perlakuan BA 2.22-17.76 µM
menghasilkan tanaman nenas normal yang dideteksi dengan menggunakan
penanda RAPD pimer OPE7 dan OPG2. Namun demikian, tanaman varigata tidak
dapat dideteksi dengan menggunakan primer OPG2. Penggunaan primer OPE7
dilaporkan hanya dapat membedakan sebagian tanaman varigata dengan tanaman
normalnya (6 dari 8 tanaman varigata).

STUDI MORFOGENESIS EKSPLAN NENAS YANG DIINDUKSI OLEH AUKSIN DAN SITOKININ
Abstrak
Morfogenesis eksplan perlu dipelajari sebelum penerapan metode regenerasi untuk perbanyakan masal, konservasi in vitro, dan transformasi genetik. Tujuan penelitian ini adalah untuk mengetahui pengaruh auksin dan sitokinin serta jenis eksplan terhadap morfogenesis eksplan nenas kultivar Smooth Cayenne pada media padat dan cair. Pada tahap pertama, digunakan media MS padat dengan penambahan zat pengatur tumbuh secara tunggal berupa BA atau NAA (0, 1, 2, dan 3 mg l-1) dengan eksplan berupa basal daun muda dan basal daun tua. Pada tahap kedua, digunakan media MS cair dengan kombinasi perlakuan sitokinin BA (0.5 atau 1.0 mg l-1) dan auksin (NAA atau IAA atau IBA pada taraf 0.5 mg l-1) dengan menggunakan eksplan berupa daun utuh dan potongan basal daun. Hasil percobaan pertama menunjukkan bahwa eksplan basal daun hanya memberikan respon ketika diberi perlakuan sitokinin ataupun auksin. Perlakuan terbaik untuk morfogenesis eksplan basal daun pada media padat adalah BA 3 mg l-1 yang menghasilkan persentase pertunasan sebasar 30% dan jumlah tunas sebesar 10 tunas/eksplan dalam waktu 2 bulan. Hal yang menarik adalah munculnya sel embriogenik dan pro-embrio dari eksplan yang diberi perlakuan NAA, namun embrio somatik gagal terbentuk. Hasil percobaan kedua menunjukkan bahwa kombinasi BA 0.5 mg l-1 dan IBA 0.5 mg l-1 dengan eksplan potongan basal daun merupakan perlakuan yang terbaik untuk morfogenesis pada media cair dengan persentase pertunasan sebesar 18% dan jumlah tunas sebesar 13 tunas/eksplan dalam waktu 2 bulan. Secara umum, disimpulkan bahwa morfogenesis eksplan basal daun nenas menggunakan media padat lebih baik daripada menggunakan media cair.
Kata kunci: Morfogenesis, nenas, kultur in vitro, media padat, media cair, embriogenesis somatik
Abstract
Morphogenetic system of explants should be firstly studied before establishing the efficient regeneration method for several applications, such as clonal mass propagation, in vitro conservation, and genetic transformation. The objective of the research was to study the effect of auxin, cytokinin and explant type to the morphogenesis of pineapple on solid media and in liquid media. On the first step, it was used MS solid media containing of a single plant growth regulator of BA or NAA at the rate of 0, 1, 2, and 3 mg l-1 respectively. On the second step, it was used MS liquid media containing of BA (0.5 or 1.0 mg l-1) combined with NAA or IAA or IBA at the rate of 0.5 mg l-1 respectively by using whole leaves and leaf bases as the explants. The result of the first experiment showed that leaf base explants showed responses only when they were treated by cytokinin or auxin. The leaf bases planted on solid MS medium with addition of 3 mg l-1 BA yielded 30% shoot formation and 10 shoots/explant in 2 months. Interestingly, embryogenic cells and pro-embryos were detected from the NAA

treated explants but they failed to form somatic embryos. The result of the second experiment showed that the basal leaves grown in MS liquid medium containing of 0.5 mg l-1 BA and 0.5 mg l-1 NAA yielded 18% shoot formation and 13 shoots/explant in 2 months. In conclusion, morphogenesis of pineapple leaf bases on solid media was beter than that in liquid media.
Keywords: Morphogenetic system, pineapple, in vitro culture, solid media, liquid media, and somatic embryogenesis
Pendahuluan
Nenas (Ananas comosus (L.) Merr.) merupakan tanaman penting di daerah
tropis dan subtropis. Berdasarkan data produksi buah-buahan, nenas menempati
peringkat ketiga atau keempat setelah pisang, mangga, dan jeruk (Tabel 3).
Penanaman bibit nenas sebanyak 40.000 tanaman/ha akan diarahkan menjadi
100.000 tanaman/ha (Suminar 2010) karena saat ini peningkatan densitas tanaman
dapat meningkatkan produksi. Konsekuensinya, keperluan penyediaan bibit nenas
menjadi sangat tinggi pada masa mendatang.
Menurut Coppens d’Eckenbrugge dan Leal (2003), secara umum organ
vegetatif nenas terdiri dari crown (mahkota), sucker (tunas yang berasal dari
ketiak daun), butt atau stump (merupakan tanaman utuh setelah pemanenan),
hapas (tunas yang diproduksi pada bagian dasar tangkai buah), ratoon (tunas yang
muncul dari bagian batang yang tertimbun di dalam tanah), dan slip (tunas yang
timbul dari tangkai buah pada bagian persis di bawah buah). Khusus untuk
kultivar Smooth Cayenne, organ vegetatif tersebut terbatas pada mahkota dan
sucker, sedangkan slip jarang sekali dihasilkan.
Perbanyakan tanaman nenas secara konvensional menghasilkan bibit dalam
jumlah yang terbatas dengan ukuran yang bervariasi dan umur panen yang
bervariasi pula sehingga pemanenan tidak dapat dilakukan secara serempak.
Coppens d’Eckenbrugge dan Leal (2003) melaporkan bahwa bibit nenas yang
berasal dari sucker memiliki umur panen 18–20 bulan, ratoon 12–14 bulan, slip
20 bulan, crown 22–24 bulan, dan collar 14-16 bulan. Walaupun mahkota
dilaporkan mempunyai tingkat keseragaman yang lebih tinggi daripada sucker
(Wakasa 1979), organ vegetatif tesebut tidak dapat digunakan sebagai alat
perbanyakan, khususnya untuk varietas nenas yang ditujukan sebagai buah meja
karena mahkota harus selalu terikut bersama buahnya pada saat dijual.
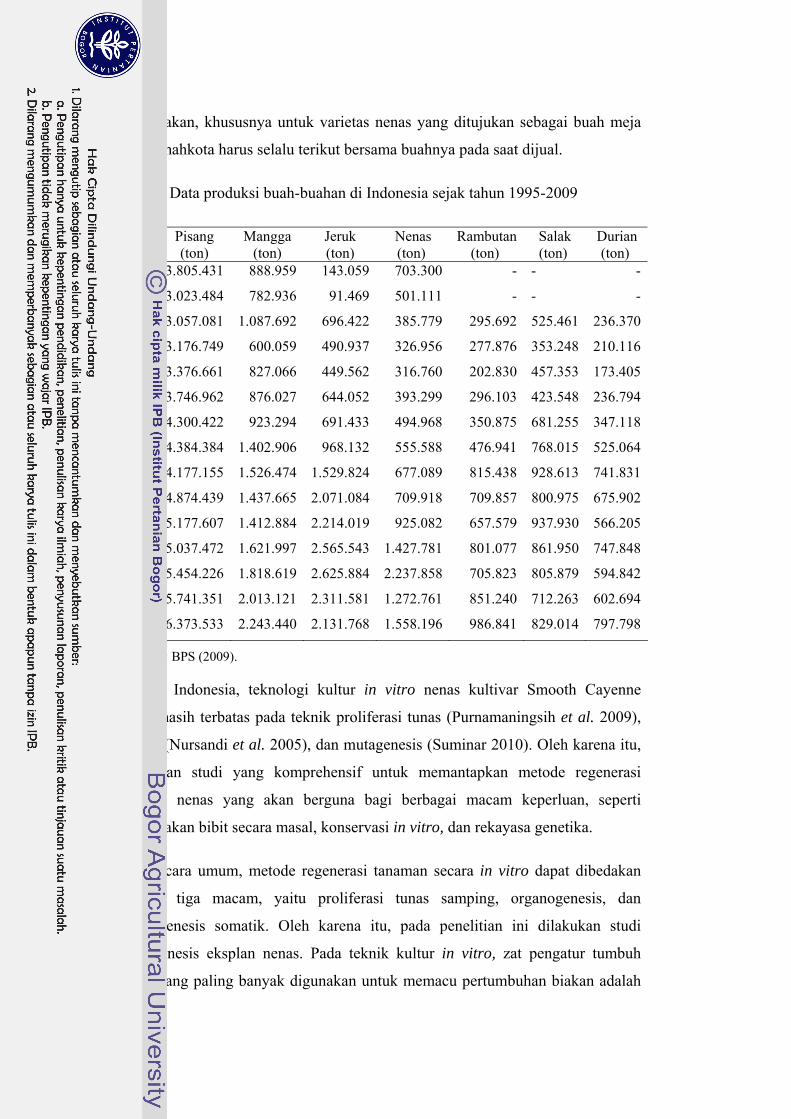
Tabel 3. Data produksi buah-buahan di Indonesia sejak tahun 1995-2009
Tahun Pisang (ton)
Mangga (ton)
Jeruk (ton)
Nenas (ton)
Rambutan(ton)
Salak (ton)
Durian (ton)
1995 3.805.431 888.959 143.059 703.300 - - -
1996 3.023.484 782.936 91.469 501.111 - - -
1997 3.057.081 1.087.692 696.422 385.779 295.692 525.461 236.370
1998 3.176.749 600.059 490.937 326.956 277.876 353.248 210.116
1999 3.376.661 827.066 449.562 316.760 202.830 457.353 173.405
2000 3.746.962 876.027 644.052 393.299 296.103 423.548 236.794
2001 4.300.422 923.294 691.433 494.968 350.875 681.255 347.118
2002 4.384.384 1.402.906 968.132 555.588 476.941 768.015 525.064
2003 4.177.155 1.526.474 1.529.824 677.089 815.438 928.613 741.831
2004 4.874.439 1.437.665 2.071.084 709.918 709.857 800.975 675.902
2005 5.177.607 1.412.884 2.214.019 925.082 657.579 937.930 566.205
2006 5.037.472 1.621.997 2.565.543 1.427.781 801.077 861.950 747.848
2007 5.454.226 1.818.619 2.625.884 2.237.858 705.823 805.879 594.842
2008 5.741.351 2.013.121 2.311.581 1.272.761 851.240 712.263 602.694
2009 6.373.533 2.243.440 2.131.768 1.558.196 986.841 829.014 797.798
Sumber: BPS (2009).
Di Indonesia, teknologi kultur in vitro nenas kultivar Smooth Cayenne
relatif masih terbatas pada teknik proliferasi tunas (Purnamaningsih et al. 2009),
etiolasi (Nursandi et al. 2005), dan mutagenesis (Suminar 2010). Oleh karena itu,
diperlukan studi yang komprehensif untuk memantapkan metode regenerasi
tanaman nenas yang akan berguna bagi berbagai macam keperluan, seperti
perbanyakan bibit secara masal, konservasi in vitro, dan rekayasa genetika.
Secara umum, metode regenerasi tanaman secara in vitro dapat dibedakan
menjadi tiga macam, yaitu proliferasi tunas samping, organogenesis, dan
embriogenesis somatik. Oleh karena itu, pada penelitian ini dilakukan studi
morfogenesis eksplan nenas. Pada teknik kultur in vitro, zat pengatur tumbuh
(ZPT) yang paling banyak digunakan untuk memacu pertumbuhan biakan adalah

sitokinin dan auksin. Kedua senyawa tersebut berperan penting dalam pembelahan
dan elongasi sel (Srivastava 2002). Oleh karena itu, kedua jenis ZPT tersebut
digunakan dalam penelitian ini. Tujuan penelitian ini adalah untuk mengetahui
pengaruh auksin dan sitokinin serta jenis eksplan terhadap morfogenesis eksplan
nenas kultivar Smooth Cayenne pada media padat dan cair.
Bahan dan Metode
Bahan tanaman yang digunakan adalah kultur in vitro tanaman nenas
kultivar Smooth Cayenne yang berasal dari Subang, Jawa Barat. Biakan tanaman
induk tersebut dipelihara pada media MS (Murashige dan Skoog 1962) dengan
penambahan benzyl adenine (BA) 0.5 mg l-1 (2.2 µM) dan kinetin (Kn) 1 mg l-1
(4.6 µM). Inkubasi dilakukan di ruang kultur dengan suhu 25±2 0C dan
pencahayaan 800–1000 lux dengan fotoperiodisitas 16 jam. Penelitian dibagi atas
dua tahap percobaan, yaitu morfogenesis eksplan pada media padat dan
morfogenesis eksplan pada media cair.
Morfogenesis eksplan pada media padat
Percobaan disusun secara faktorial dalam lingkungan Rancangan Acak
Lengkap. Faktor pertama adalah jenis eksplan, yaitu basal daun muda dan basal
daun tua. Faktor kedua adalah zat pengatur tumbuh (ZPT), yaitu BA dan
naphthalene acetic acid (NAA) masing-masing pada taraf 0, 1, 2, dan 3 mg l-1.
Daun muda diambil dari daun ke-4 hingga ke-8 dari pucuk dengan ciri khusus,
yaitu bagian pangkalnya berwarna putih kehijauan dan tebal. Daun tua diambil
dari daun ke-3 hingga ke-5 dari pangkal batang dengan ciri khusus, yaitu bagian
pangkalnya transparan, tipis, dan agak melingkar. Daun diisolasi dari tunas in
vitro dengan cara melepaskan daun-daunnya satu persatu secara hati-hati
kemudian bagian basalnya dipotong (berukuran 0.3-0.5 cm). Eksplan ditanam
pada media MS dengan penambahan ZPT sesuai perlakuan dan agar 8 g l-1. Pada
setiap botol ditanami sepuluh eksplan dan perlakuan diulang sebanyak 4 kali.
Eksplan diinkubasikan pada ruang kultur dengan suhu 25±2 0C, fotoperiodisitas
16 jam terang dan intensitas cahaya 800–1000 lux. Respon yang diamati adalah
persentase pembentukan akar, tunas, kalus, jumlah tunas, dan penampilan visual

biakan. Persentase pembentukan akar atau tunas atau kalus adalah jumlah eksplan
yang berakar atau bertunas atau berkalus dibagi dengan jumlah total eksplan yang
diamati dikalikan 100%.
Morfogenesis eksplan pada media cair
Pada tahap ini, terdapat dua percobaan. Percobaan pertama menggunakan
eksplan daun utuh, sedangkan percobaan kedua menggunakan eksplan potongan
basal daun. Pada kedua percobaan tersebut digunakan Rancangan Acak Lengkap.
Helaian daun in vitro dilepaskan secara hati-hati dari batangnya dengan bantuan
skalpel lalu basal daun diisolasi dengan mata pisau tajam. Perlakuan yang diujikan
adalah media MS cair (25 ml) yang mengandung BA pada taraf 0.5 atau 1.0 mg l-1
(2.2 atau 4.4 µM) dengan atau tanpa penambahan NAA, indole butyric acid
(IBA), dan indole acetic acid (IAA) pada taraf 0.5 mg l-1. Ke dalam setiap botol,
dimasukkan sepuluh eksplan, dan perlakuan diulang sebanyak 4 kali.
Penggoyangan atau pemutaran dilakukan pada rotary shaker dengan kecepatan 60
rpm secara periodik 16 jam selama 3 minggu kemudian kecepatan ditingkatkan
menjadi 70 rpm. Fotoperiodisitas diterapkan selama 16 jam terang dengan
intensitas cahaya 800–1000 lux. Inkubasi dilakukan pada suhu 25±2 0C selama 1.5
bulan. Respon yang diamati adalah persentase pembentukan akar, tunas, kalus,
jumlah tunas, dan penampilan visual biakan. Persentase pembentukan akar atau
tunas atau kalus adalah jumlah eksplan yang berakar atau bertunas atau berkalus
dibagi dengan jumlah total eksplan yang diamati dikalikan 100%.
Hasil dan Pembahasan
Morfogenesis eksplan pada media padat
Hasil percobaan menunjukkan bahwa eksplan basal daun tidak memberikan
respon apapun ketika ditumbuhkan pada media bebas ZPT, namun memberikan
respon ketika diberi perlakuan sitokinin ataupun auksin (Gambar 3 dan 4). Respon
tersebut dihasilkan dari bagian pangkal daun. Menurut Firoozabady and Moy
(2004), basal daun mempunyai area meristematis di mana sel-selnya membelah
dengan pesat sehingga bagian tersebut paling respon terhadap perlakuan kultur in
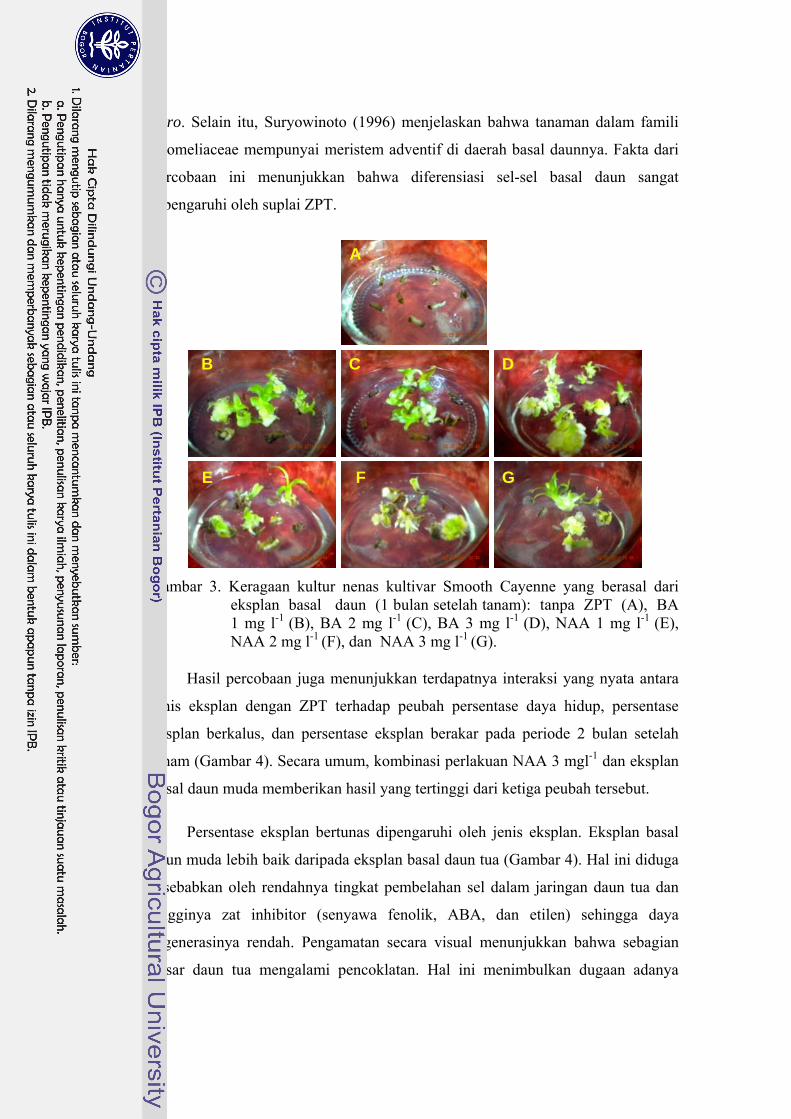
vitro. Selain itu, Suryowinoto (1996) menjelaskan bahwa tanaman dalam famili
Bromeliaceae mempunyai meristem adventif di daerah basal daunnya. Fakta dari
percobaan ini menunjukkan bahwa diferensiasi sel-sel basal daun sangat
dipengaruhi oleh suplai ZPT.
A
CB D
GFE
Gambar 3. Keragaan kultur nenas kultivar Smooth Cayenne yang berasal dari
eksplan basal daun (1 bulan setelah tanam): tanpa ZPT (A), BA 1 mg l-1 (B), BA 2 mg l-1 (C), BA 3 mg l-1 (D), NAA 1 mg l-1 (E), NAA 2 mg l-1 (F), dan NAA 3 mg l-1 (G).
Hasil percobaan juga menunjukkan terdapatnya interaksi yang nyata antara
jenis eksplan dengan ZPT terhadap peubah persentase daya hidup, persentase
eksplan berkalus, dan persentase eksplan berakar pada periode 2 bulan setelah
tanam (Gambar 4). Secara umum, kombinasi perlakuan NAA 3 mgl-1 dan eksplan
basal daun muda memberikan hasil yang tertinggi dari ketiga peubah tersebut.
Persentase eksplan bertunas dipengaruhi oleh jenis eksplan. Eksplan basal
daun muda lebih baik daripada eksplan basal daun tua (Gambar 4). Hal ini diduga
disebabkan oleh rendahnya tingkat pembelahan sel dalam jaringan daun tua dan
tingginya zat inhibitor (senyawa fenolik, ABA, dan etilen) sehingga daya
regenerasinya rendah. Pengamatan secara visual menunjukkan bahwa sebagian
besar daun tua mengalami pencoklatan. Hal ini menimbulkan dugaan adanya

aktivitas polifenol oksidase yang menghasilkan quinon sehingga eksplan menjadi
coklat.
Selain persentase pembentukan tunas, hingga periode 2 bulan, peubah
jumlah tunas juga dipengaruhi oleh jenis eksplan walaupun pada periode 4 bulan,
ZPT memberikan pengaruh yang nyata terhadap jumlah tunas. Pada periode 2
bulan, sebanyak 30% eksplan beregenerasi membentuk tunas (Gambar 4) dengan
jumlah tunas sebanyak 10 tunas/eksplan (Gambar 4). Pada periode 4 bulan,
perlakuan BA 3 mg l-1 menghasilkan jumlah tunas tertinggi (18 tunas/eksplan)
dibandingkan perlakuan lainnya. Diduga bahwa sitokinin terakumulasi di dalam
jaringan biakan dengan bertambahnya periode kultur in vitro sehingga
pengaruhnya nyata pada 4 bulan setelah tanam.
Secara visual terdapat perbedaan respon yang mencolok antara eksplan yang
diberi perlakuan BA dan NAA. Penggunaan BA pada awalnya memacu
pertumbuhan ke arah pertunasan dan selanjutnya menginduksi pembentukan kalus
setelah pembentukan tunas (Gambar 4). Diduga bahwa kandungan sitokinin
endogen menurun seiring dengan bertambahnya periode in vitro sehingga tarafnya
menjadi seimbang dengan auksin endogen yang disintesis oleh pucuk tunas dan
berakibat pada terbentuknya kalus. Di lain pihak, NAA memacu pertumbuhan
akar dan mampu menginduksi pertumbuhan kalus setelah pembentukan akar dan
menginduksi pembentukan tunas setelah kalus terbentuk. Terbentuknya kalus oleh
perlakuan NAA diduga karena akar yang terbentuk mampu segera mensintesis
sitokinin lalu sitokinin tersebut ditranslokasikan secara akropetal sehingga ketika
rasio antara keduanya seimbang maka terbentuk kalus dan ketika sitokinin
endogen lebih tinggi tarafnya maka terbentuk tunas.
Terbentuknya organ tunas dan akar secara terpisah menunjukkan bahwa
sifatnya unipolar (satu kutub). Proses regenerasi demikian disebut sebagai
regenerasi secara organogenesis (Phillips et al. 1995). Hasil percobaan ini
mengindikasikan bahwa basal daun nenas mampu beregenerasi secara
organogenesis, baik langsung (tanpa melalui fase kalus) maupun tak langsung
(melalui fase kalus).
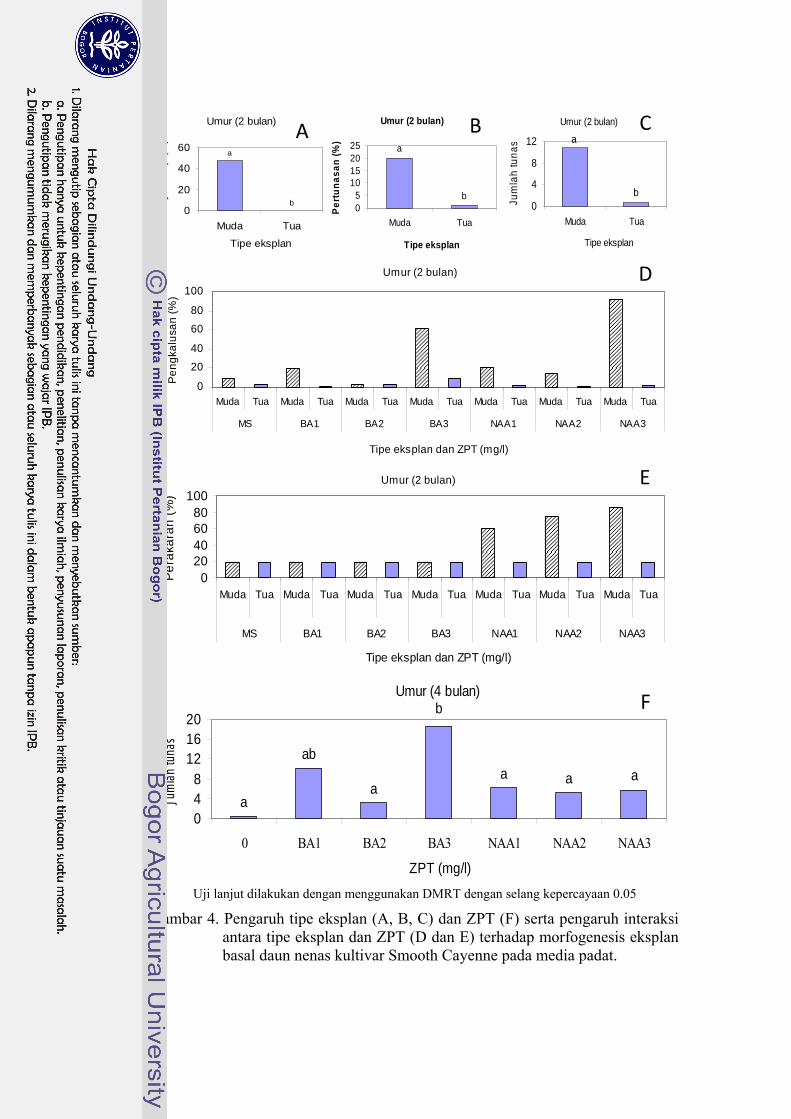
Umur (2 bulan)
a
b0
4
8
12
Muda Tua
Tipe eksplan
Jum
lah
tuna
s
Umur (2 bulan)
a
b0
20
40
60
Muda Tua
Tipe eksplan
Day
a hi
dup
(%)
Umur (2 bulan)
0
20
40
60
80
100
Muda Tua Muda Tua Muda Tua Muda Tua Muda Tua Muda Tua Muda Tua
MS BA1 BA2 BA3 NAA1 NAA2 NAA3
Tipe eksplan dan ZPT (mg/l)
Pen
gkal
usan
(%)
Umur (2 bulan)
020406080
100
Muda Tua Muda Tua Muda Tua Muda Tua Muda Tua Muda Tua Muda Tua
MS BA1 BA2 BA3 NAA1 NAA2 NAA3
Tipe eksplan dan ZPT (mg/l)
Pera
kara
n (%
)
Umur (2 bulan)
b
a
05
10152025
Muda Tua
Tipe eksplan
Pertu
nasa
n (%
)
Umur (4 bulan)
aaa
b
a
ab
a048
121620
0 BA1 BA2 BA3 NAA1 NAA2 NAA3
ZPT (mg/l)
Jum
lah tu
nas
A B C
D
E
F
Uji lanjut dilakukan dengan menggunakan DMRT dengan selang kepercayaan 0.05
Gambar 4. Pengaruh tipe eksplan (A, B, C) dan ZPT (F) serta pengaruh interaksi antara tipe eksplan dan ZPT (D dan E) terhadap morfogenesis eksplan basal daun nenas kultivar Smooth Cayenne pada media padat.

Hasil percobaan ini juga menunjukkan bahwa kalus yang terbentuk dari
perlakuan BA bersifat kompak sedangkan kalus yang terbentuk dari perlakuan
NAA bersifat remah. Terbentuknya kalus kompak diduga disebabkan oleh
tingginya kandungan sitokinin endogen yang diinduksi oleh perlakuan sitokinin
eksogen berupa BA. Terbentuknya kalus remah menimbulkan dugaan adanya sel-
sel induced embryogenic determined cell (IEDC) pada daerah basal daun nenas
dan berpotensi untuk induksi embriogenesis somatik. NAA merupakan salah satu
jenis auksin yang mempunyai daya aktivitas kuat, mudah ditranslokasikan, dan
mempunyai persistensi yang tinggi sehingga memacu pembentukan akar dan
mampu menginduksi terbentuknya kalus. Dengan demikian, dapat dikatakan
bahwa eksplan basal daun nenas juga berpeluang tinggi untuk diregenerasikan
secara embriogenesis somatik.
Untuk meyakinkan dugaan tersebut maka kalus remah tersebut diamati di
bawah mikroskop. Hasil pengamatan secara mikroskopis menunjukkan bahwa
kalus terdiri atas kumpulan sel yang bersifat non embriogenik (Gambar 5) dan
embriogenik (Gambar 6). Sel yang non embriogenik ditandai dengan
pembelahannya yang bersifat simetris sedangkan sel yang embriogenik ditandai
dengan pembelahan sel yang asimetris. Menurut Abrash dan Bergmann (2009),
pembelahan asimetris menghasilkan sel-sel anakan yang berbeda secara
morfologi. Pada penelitian ini, teramati bahwa pembelahan asimetris didahului
dengan polaritas sel (Gambar 6A) kemudian pembentukan sel apikal dan basal
(Gambar 6B) lalu dilanjutkan dengan pembelahan sel secara transversal (Gambar
6C) dan longitudinal (Gambar 6D) hingga terbentuk sel yang linier (Gambar 6E)
dan struktur oktan (Gambar 6F). Sayangnya, sel embriogenik yang telah
dihasilkan gagal membentuk embrio somatik. Kegagalan tersebut kemungkinan
disebabkan oleh kurangnya akumulasi amilum, protein, dan lipid di dalam sel-sel
yang menyusun pro-embrio sehingga pro-embrio tersebut tidak mampu tumbuh
dewasa untuk membentuk embrio somatik. Oleh karena itu, perlu digunakan tipe
auksin lainnya.

A
B C
Gambar 5. Penampilan kalus nenas pasca-perlakuan NAA dan tipe sel yang
menyusunnya: kalus remah (A), pembelahan sel secara simetris (B), dan kumpulan sel non embriogenik (C).
apikal
Basal transversal
longitudinal
A B
C
D
E F Gambar 6. Pembelahan sel secara asimetris dari sel embriogenik yang diiduksi
oleh NAA: sel yang terpolarisasi (A), sel apikal dan basal (B), sel yang mengalami pembelahan transversal (C) dan longitudinal (D), sel yang tersusun linier (E), stadium oktan atau 8 sel (F).

Morfogenesis eksplan pada media cair
Berdasarkan hasil percobaan pertama diketahui bahwa eksplan daun nenas
memberikan respon yang baik pada media padat yang ditambah dengan ZPT.
Diduga bahwa eksplan tersebut akan memberikan respon yang lebih baik pada
media cair karena kontak eksplan dengan media cair lebih intensif dibandingkan
dengan media padat. Oleh karena itu, taraf ZPT pada percobaan ini diturunkan
dan digunakan kombinasi perlakuan antara sitokinin dan auksin. Selain itu, jenis
eksplan (daun utuh dan potongan basal daun) juga dipelajari pengaruhnya
terhadap daya morfogenesis pada media cair.
Hasil percobaan menunjukkan bahwa semua formulasi media tidak
memberikan pengaruh yang nyata terhadap persentase pertunasan tetapi
berpengaruh nyata terhadap persentase pembentukan nodul dan akar dari eksplan
daun utuh (Gambar 7). Berbeda dengan daun utuh, eksplan basal daun mempunyai
respon yang lebih baik karena beregenerasi hampir pada semua formulasi media.
Persentase pembentukan tunas dan akar tidak dipengaruhi secara nyata oleh
perlakuan yang diberikan. Perlakuan kombinasi antara BA 0.5 mgl-1 dan IAA 0.5
mgl-1, BA 0.5 mgl-1 dan IBA 0.5 mgl-1, serta BA 1 mgl-1 dan IBA 0.5 mgl-1
memberikan persentase pembentukan nodul yang berbeda nyata daripada
perlakuan lainnya (Gambar 7). Selanjutnya, peubah jumlah tunas, tidak terdapat
perbedaan yang nyata antar formulasi media yang diujikan (Gambar 8). Jumlah
tunas tertinggi diperoleh dari perlakuan BA 0.5 mgl-1 dan IBA 0.5 mgl-1 serta
perlakuan IBA 1 mgl-1. Dengan demikian, dapat disimpulkan bahwa perlakuan
BA 0.5 mgl-1 dan IBA 0.5 mgl-1 merupakan perlakuan terbaik untuk morfogenesis
eksplan nenas pada media cair.
Berdasarkan Gambar 9 tampak bahwa respon awal dari eksplan daun nenas
adalah menggembungnya bagian basal daun sehingga warnanya menjadi
keputihan. Diduga bahwa respon awal tersebut diperankan oleh auksin. Secara
visual, tampak bahwa daun menggembung di daerah abaksial (permukaan bawah).
Hal ini menunjukkan bahwa elongasi sel lebih pesat pada daerah abaksial daripada
daerah adaksial (permukaan atas). Srivastava (2002) mengemukakan bahwa
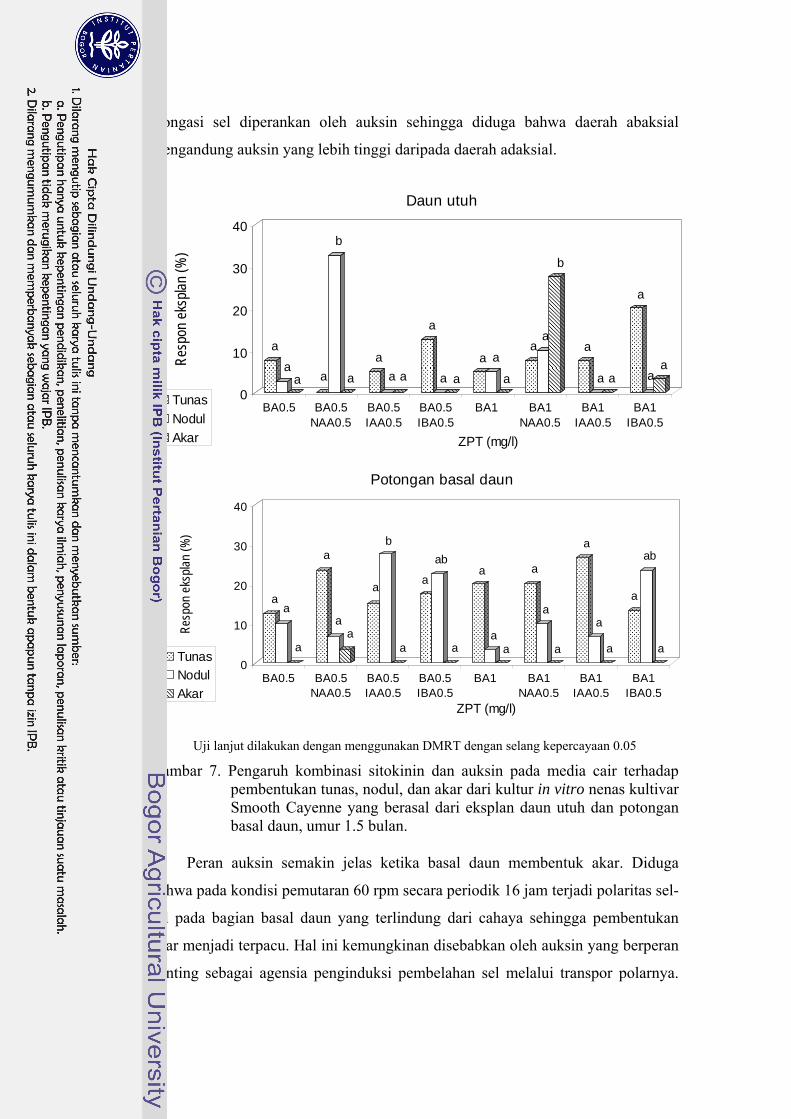
elongasi sel diperankan oleh auksin sehingga diduga bahwa daerah abaksial
mengandung auksin yang lebih tinggi daripada daerah adaksial.
aa
a a
b
aa
a a
a
a aa a
a
aa
b
a
a a
a
aa
0
10
20
30
40
Resp
on e
kspla
n (%
)
BA0.5 BA0.5NAA0.5
BA0.5IAA0.5
BA0.5IBA0.5
BA1 BA1NAA0.5
BA1IAA0.5
BA1IBA0.5
ZPT (mg/l)
Daun utuh
TunasNodulAkar
aa
a
a
aa
a
b
a
a
ab
a
a
aa
a
a
a
a
a
a
a
ab
a0
10
20
30
40
Resp
on e
kspla
n (%
)
BA0.5 BA0.5NAA0.5
BA0.5IAA0.5
BA0.5IBA0.5
BA1 BA1NAA0.5
BA1IAA0.5
BA1IBA0.5
ZPT (mg/l)
Potongan basal daun
TunasNodulAkar
Uji lanjut dilakukan dengan menggunakan DMRT dengan selang kepercayaan 0.05
Gambar 7. Pengaruh kombinasi sitokinin dan auksin pada media cair terhadap pembentukan tunas, nodul, dan akar dari kultur in vitro nenas kultivar Smooth Cayenne yang berasal dari eksplan daun utuh dan potongan basal daun, umur 1.5 bulan.
Peran auksin semakin jelas ketika basal daun membentuk akar. Diduga
bahwa pada kondisi pemutaran 60 rpm secara periodik 16 jam terjadi polaritas sel-
sel pada bagian basal daun yang terlindung dari cahaya sehingga pembentukan
akar menjadi terpacu. Hal ini kemungkinan disebabkan oleh auksin yang berperan
penting sebagai agensia penginduksi pembelahan sel melalui transpor polarnya.

Menurut Vanneste and Frim (2009), kendali distribusi auksin menentukan suatu
sel dari jaringan untuk melaksanakan perubahan spesifik dalam program
perkembangannya, mencakup organogenesis dan tropisme. Srivastava 2002 juga
melaporkan bahwa auksin berperan penting sebagai pemandu arah dalam berbagai
proses perkembangan tanaman yang melibatkan polaritas. Menariknya, ketika
pemutaran ditingkatkan menjadi 70 rpm secara kontinyu, eksplan mampu bertunas
dalam waktu yang singkat (1 minggu). Hal ini membuktikan bahwa polaritas
dipengaruhi oleh kecepatan dan periodisitas pemutaran. Terbentuknya tunas dari
ekplan yang berakar menunjukkan bahwa mekanisme dead-end tidak terjadi pada
kultur nenas kultivar Smooth Cayenne, suatu fenomena yang tidak umum terjadi
pada berbagai macam tanaman lainnya.
Daun utuh
0
10
20
30
40
BA0.5 BA0.5NAA0.5
BA0.5IAA0.5
BA0.5IBA0.5
BA1 BA1NAA0.5
BA1IAA0.5
BA1IBA0.5
ZPT (mg/L)
Juml
ah tu
nas 1.5 bulan
2 bulan
Potongan basal daun
0
5
10
15
20
BA0.5 BA0.5NAA0.5
BA0.5IAA0.5
BA0.5IBA0.5
BA1 BA1NAA0.5
BA1IAA0.5
BA1IBA0.5
ZPT (mg/l)
Juml
ah tu
nas
1.5 bulan
2 bulan
Tidak berbeda nyata pada uji DMRT dengan selang kepercayaan 0.05
Gambar 8. Pengaruh kombinasi sitokinin dan auksin pada media cair terhadap jumlah tunas dari kultur in vitro nenas kultivar Smooth Cayenne yang berasal dari eksplan daun utuh dan potongan basal daun.

Perbedaan respon juga terlihat pada daerah abaksial dan adaksial. Daerah
abaksial cenderung membentuk akar sedangkan tunas muncul dari daerah adaksial
(Gambar 9). Hal ini menunjukkan bahwa distribusi dan rasio ZPT pada kedua
daerah tersebut juga berbeda, di mana sitokinin lebih dominan pada daerah
adaksial sehingga memacu pertumbuhan tunas, sedangkan auksin lebih dominan
pada daerah abaksial sehingga memacu pertumbuhan akar.
A
C
B
D
Gambar 9. Tahapan morfogenesis dari eksplan daun nenas kultivar Smooth
Cayenne pada media cair: respon awal eksplan oleh perlakuan kombinasi sitokinin dan auksin (A), pembentukan nodul, akar, dan tunas (B), elongasi tunas (C), dan pembentukan planlet (D).
Simpulan
Basal daun nenas merupakan area yang meristematis sehingga mampu
berdiferensiasi. Pemberian ZPT (berupa sitokinin atau auksin) mutlak diperlukan
untuk menginduksi proses diferensiasi tersebut. Pemberian BA memacu ke arah
pembentukan tunas dan kalus kompak. Pemberian NAA memacu ke arah
pembentukan akar yang diikuti dengan pembentukan kalus remah yang tersusun
atas sel-sel non embriogenik dan embriogenik. Terdapat interaksi yang nyata
antara jenis eksplan dan ZPT terhadap persentase daya hidup, persentase eksplan
berkalus, dan persentase eksplan berakar sedangkan jumlah tunas yang terbentuk
dipengaruhi secara nyata oleh ZPT. Perlakuan terbaik untuk organogenesis
langsung pada media padat adalah BA 3 mg l-1 dan eksplan berupa basal daun
muda dengan persentase pertunasan sebesar 30% dan tingkat multiplikasi tunas

sebesar 10 tunas/eksplan dalam waktu 2 bulan. Perlakuan terbaik untuk
organogenesis langsung pada media cair adalah BA 0.5 mg l-1 dan IBA 0.5 mg l-1
dengan eksplan berupa potongan basal daun dengan dengan persentase pertunasan
sebesar 18% dan tingkat multiplikasi tunas sebesar 13 tunas/eksplan dalam waktu
2 bulan. Secara umum dapat disimpulkan bahwa organogenesis langsung basal
daun nenas pada media padat lebih baik daripada organogenesis langsung pada
media cair.
Daftar Pustaka
Abrash, E.B. and D.C. Bergmann. 2009. Asymmetric cell divisions: A view from plant development. Developmental Cell 16: 783-796.
[BPS] Badan Pusat Statistik Republik Indonesia. 2009. Produksi buah-buahan di Indonesia. Jakarta
Coppens d’Eeckenbrugge G, Leal F. 2003. Morphology, anatomy, and taxonomy. Pp. 13-32. In: Bartholomew DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany: Production and Uses. CABI Publishing.Wallingford.
Department of Health and Ageing. 2008. The Biology of Ananas comosus var comosus (pineapple). Australian Government. 43pp.
Firoozabady E, Moy Y. 2004. Regeneration of pineapple via somatic embryogenesis and organogenesis. In vitro Cell Dev Biol_Plant 40:67-74.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15: 473-497.
Nursandi F, Poerwanto R, Sobir, Sujiprihati S. 2003. Perbanyakan in vitro nenas (Ananas comosus (L.) Merr.) dengan menggunakan BAP dan TDZ. Dalam: Simposium Nasional dan Konggres VIII. Bandar Lampung.
Nursandi F, Sobir, Murtini. 2005. Perbanyakan tanaman nenas (Ananas comosus (L.) Merr.) cv. Smooth Cayenne dengan teknik etiolasi secara in vitro. Tropika 13(1):15-24.
Phillips GC, Hubstenberger JF, Hansen EE. 1995. Plant regeneration from callus and cell suspension cultures by somatic embryogenesis. Pp. 81 - 90. In: Gamborg OL, Phillips GC, editor. Plant Cell Tissue and Organ Culture: Fundamental Methods. Springer. Berlin.
Purnamaningsih R, Mariska I, Supriati Y. 2009. Penggunaan ABA dan paklobutrazol dalam perbanyakan nenas Simadu melalui kultur in vitro. Ber Bio 9(6): 751-758.
Srivastava LM. 2002. Plant Growth and Development. Academic Press. New York. 772p.

Suminar E. 2010. Induksi keragaman genetik dengan mutagen sinar gamma pada nenas (Ananas comosus (L.) Merr.) secara in vitro [seminar]. Sekolah Pascasarjana, Institut Pertanian Bogor. Bogor.
Suryowinoto M. 1996. Pemuliaan Tanaman secara In Vitro. Pusat Antar Universitas-Bioteknologi UNiversitas Gadjah Mada. Yogyakarta. 252 hal.
Vanneste S, Frim J. 2009. Auxin: A trigger for change in plant development. Cell 136: 1005-1016.
Wakasa K. 1979. Variation in the plants differentiated from the tissue culture of pineapple. Japan J Breed 29(1): 13-22.

ORGANOGENESIS DAN EMBRIOGENESIS SOMATIK TIDAK LANGSUNG PADA NENAS YANG DIINDUKSI OLEH
2,4-D
Abstrak
Penelitian ini bertujuan: (1) menentukan taraf 2,4-D yang terbaik untuk induksi kalus dan regenerasi secara organogenesis, (2) mengetahui pengaruh interaksi antara media dan AdS terhadap induksi kalus embriogenik, (3) menentukan formulasi media terbaik untuk regenerasi kalus embriogenik. Induksi kalus dilakukan dengan perlakuan 2,4-D (21, 41, dan 62 µM) dengan penambahan TDZ 9 µM. Sel-sel yang non embriogenik diregenerasikan secara organogenesis dengan memindahkannya ke media yang mengandung Kn 4.65 µM. Induksi kalus embriogenik dilakukan pada media MS atau Bac yang diperkaya dengan senyawa N-organik (Gln 6.8 µM, CH 500 mg l-1, Arg 0.69 mM, dan Gly 0.027 µM) dengan penambahan AdS (0, 0,05 dan 0,1 µM). Kalus embriogenik diregenerasikan pada dua macam media. Media pertama adalah media MS modifikasi dengan penambahan IBA 0.9 µM, BA 1.1 µM, GA3 0.09 µM sedangkan yang kedua adalah media MS dengan penambahan BA 18 µM. Hasil penelitian membuktikan bahwa penggunaan 2,4-D secara tunggal tidak mampu menginduksi sel embriogenik sehingga sebagian kalus non embriogenik diregenerasikan secara organogenesis. Media terbaik untuk organogenesis adalah 2,4-D 21 µM karena menghasilkan persentase pembentukan kalus yang tinggi (80%), bobot basah kalus tertinggi (0.2 g/eksplan), dan jumlah tunas tertinggi (sekitar 25 tunas/eksplan/2 bulan). Kalus embriogenik terbentuk ketika kalus non embriogenik diperlakukan dengan senyawa N-organik. Media regenerasi merupakan faktor yang berpengaruh nyata terhadap tingkat pencoklatan. Media MS dengan penambahan BA 18 µM mampu menekan pencoklatan tersebut. Terdapat interaksi yang nyata antara media induksi kalus embriogenik dengan media regenerasi terhadap jumlah embrio somatik dewasa. Media MS yang diperkaya dengan senyawa N-organik dan AdS 0.05 µM yang dikombinasikan dengan media MS dengan penambahan BA 18 µM merupakan perlakuan terbaik untuk induksi kalus embriogenik dan regenerasi embrio somatik nenas (17 embrio dewasa/eksplan) dalam 6.5 bulan.
Kata kunci: Organogenesis, embriogenesis somatik, nenas, 2,4-D, adenin sulfat
Abstract
This research was aimed to: (1) determine the best rate of 2,4-D for callus induction and regeneration through organogenesis, (2) study the interaction effect of media and AdS to induce embryogenic callus formation, and (3) obtain the best treatment for regeneration of embryogenic calli. Callus formation was induced by 2,4-D (21, 41, and 62 µM) with addition of 9 µM TDZ. Non embryogenic calli were regenerated through organogenesis method by transferring them onto MS medium containing of 4.65 µM Kn. Embryogenic callus formation was induced on MS or Bac basal media consisted of N-organic compounds (6.8 µM Gln + 500 mg l-1 CH + 0.69 mM Arg + 0.027 µM Gly) with addition of AdS (0, 0.05 and 0.1

µM). There were two different media for embryogenic calli regeneration. The first medium was MS medium supplemented with 0.9 µM IBA, 1.1 µM BA, 0.09 µM GA3. The second medium was modified MS with addition of 18 µM BA. The result showed that the single auxin of 2,4-D could induce callus formation but it could not induce embryogenic cells. The best treatment for callus induction was 21 µM 2,4-D which provided high level of callus formation (80%), highest level of fresh weight (0.2 g/explant), and highest number of shoot (about 25 shoots/explant/2 months). The embryogenic calli were formed when non embryogenic calli were treated by N-organic compounds. The regeneration medium significantly affected the level of browning, where 18 µM BA containing medium could inhibit that occurrence. There was a significant interaction between callus induction media and regeneration media to the number of mature somatic embryos. The MS medium enriched by N-organic compounds and 0.05 µM AdS combined with the MS containing of 18 µM BA was the best combination which yielded 17 mature somatic embryos/explant in 6.5 months.
Keywords: Organogenesis, somatic embryogenesis, pineapple, dichlorophenoxy acetic acid, adenine sulphate
Pendahuluan
Nenas adalah buah tropis komersial yang produksinya menempati peringkat
ketiga atau keempat setelah pisang, mangga, dan jeruk (BPS 2009), sangat
berpotensi dikembangkan lebih luas di Indonesia. Secara konvensional, tanaman
tersebut mempunyai banyak propagul (IBPGR 1991; Coppens d’Eeckenbrugge
and Leal 2003), seperti mahkota, sucker, stump, hapas, ratoon, dan slip, namun
umur reproduktifnya tidak serempak. Pada kultivar Smooth Cayenne, ketersediaan
propagul tersebut bahkan terbatas sehingga metode perbanyakan konvensional
perlu didukung teknologi lainnya, terutama untuk produksi bibit skala masal.
Teknik mikropropagasi nenas diterapkan secara komersial oleh pihak
industri di mancanegara (Smith et al. 2003) tetapi industri di Indonesia masih
bertahan pada teknik perbanyakan konvensional. Namun demikian, teknik
mikropropagasi masih dipandang perlu diterapkan untuk menyediakan bibit dalam
jumlah besar, terutama untuk kultivar-kultivar baru hasil persilangan, seleksi,
mutasi, dan rekayasa genetika (Firoozabady dan Moy 2004; Nursandi et al. 2005).
Secara praktis, mikropropagasi digunakan untuk memantapkan blok multiplikasi
bagi penyediaan bahan tanaman perbanyakan konvensional dalam skala yang
lebih luas. Selain itu, teknik mikropapagasi dapat diterapkan untuk skrining varian

sebelum diterapkannya metode perbanyakan secara konvensional (Smith et al
2003).
Di Indonesia, studi kultur in vitro nenas kultivar Smooth Cayenne masih
terbatas pada pembahasan teknik proliferasi tunas (Purnamaningsih et al. 2009),
etiolasi tunas (Nursandi et al. 2003), dan mutagenesis (Suminar 2010). Hingga
saat ini, pembahasan tentang organogenesis masih terbatas dan studi
embriogenesis somatik nenas kultivar Smooth Cayenne bahkan belum pernah
dilakukan.
Organogenesis adalah pembentukan tunas adventif secara langsung dari
eksplan yang menghasilkan struktur unipolar, sedangkan embriogenesis somatik
adalah proses regenerasi sel somatik yang membelah dan berkembang menjadi
struktur bipolar atau embrio. Struktur unipolar terdiri atas tunas atau akar,
sedangkan struktur bipolar terdiri atas meristem tunas dan akar (Phillips et al.
1995; Arnold et al 2002).
Hingga saat ini, di dunia belum terdapat laporan tentang tahapan lengkap
embriogenesis somatik dari tanaman nenas. Pemahaman tentang embriogenesis
penting diketahui untuk memperoleh sistem morfogenik dengan cara mengamati
proses seluler yang mendasari proses diferensiasi (Corredoira et al. 2006).
Informasi tersebut akan berguna untuk menyediakan metode embriogenesis
somatik yang mantap bagi propagasi masal, kriopreservasi, dan transformasi.
Di antara jenis auksin, dichlorophenoxy acetic acid (2,4-D) biasanya
digunakan dalam konsentrasi rendah. 2,4-D adalah auksin herbisida yang
memiliki fungsi hormonal yang menyerupai auksin alami (Kelley dan Riechers
2007). Selanjutnya Srivastava (2002) mengemukakan bahwa senyawa tersebut
efektif sebagai penginduksi proliferasi sel dalam kultur sel dan jaringan.
Penelitian ini bertujuan untuk mengetahui pengaruh 2,4-D, adenine sulfate (AdS),
dan media dasar terhadap regenerasi nenas kultivar Smooth Cayenne melalui jalur
organogenesis dan embriogenesis somatik, serta untuk mempelajari tahapan
lengkap embriogenesis somatik yang diawali dari inisiasi sel embriogenik hingga
pembentukan planlet yang siap diaklimatisasi.

Bahan dan Metode
Bahan tanaman adalah tunas in vitro tanaman nenas kultivar Smooth
Cayenne klon Simadu dari Subang. Kultur tersebut diperoleh dari isolasi dan
inisiasi mata tunas yang terdapat pada mahkota nenas. Pemeliharaan kultur
dilakukan dengan menggunakan media MS (Murashige dan Skoog 1962) dengan
penambahan benzyl adenine (BA) 2.21 µM (0.5 mg l-1) dan kinetin (Kn) 4.65 µM
(1 mg l-1). Kemasaman media (pH ) ditera pada nilai 5.7±0.1 sebelum dimasukkan
ke dalam otoklaf. Inkubasi dilakukan pada ruang kultur dengan suhu 25 0C,
fotoperiodisitas 16 jam terang dengan intensitas penyinaran 800–1000 lux.
Regenerasi secara Organogenesis
Tunas in vitro sekitar 4–6 cm dipotong secara longitudinal menjadi dua
bagian, kemudian ditanam secara horizontal pada media induksi kalus (10 bagian
per botol) dan diinkubasikan pada kondisi remang-remang (di bawah 500 lux).
Media induksi kalus adalah media MS yang mengandung 2,4-D pada taraf 21, 41,
dan 62 µM (4.6, 9, dan 13.7 mg l-1) dengan penambahan thidiazuron (TDZ) pada
taraf 9 µM (2 mg l-1). Eksplan diinkubasikan pada suhu 25±2 0C dalam keadaan
gelap. Setelah eksplan membesar (3 minggu), helaian daun diisolasi secara
individual dan kemudian ditanam pada media yang sama. Inkubasi juga dilakukan
pada kondisi yang sama dengan tahap sebelumnya. Rancangan percobaan adalah
Rancangan Acak Lengkap dengan 5 ulangan (botol). Setiap botol terdiri atas 15
eksplan. Pembentukan kalus diamati pada minggu ke-3, 6, 8, dan 12. Bobot basah
kalus diukur pada akhir pengamatan. Data dianalisis dengan program SPSS V.19
(IBM Company) dan perbedaan antar rerata dibandingkan dengan uji Duncans’s
multiple range tests (DMRT) pada tingkat kepercayaan 0.05. Penampilan visual
kalus juga diamati (kompak atau remah, putih atau kuning atau hijau, mudah
dipisahkan atau tidak, berair atau vigor). Pengamatan secara mikroskopis
dilakukan untuk menentukan tipe pembelahan sel. Jika pembelahan sel terjadi
secara asimetris maka kalus dipindahkan ke media regenerasi embrio somatik,
namun jika kalus membelah secara simetris maka kalus dipindahkan ke media
regenerasi tunas, yaitu media MS dengan penambahan Kn 4.65 µM (1 mg l-1).
Selanjutnya dihitung jumlah tunas yang normal dan abnormal. Abnormalitas

ditentukan dengan mengamati keseimbangan aksialitas. Data ditampilkan dalam
bentuk rerata dan standar deviasi.
Regenerasi secara Embriogenesis Somatik
Pertama kali, eksplan basal daun ditanam pada media MS dengan
penambahan 2,4-D 21 µM (4.6 mg l-1). Kalus yang terbentuk kemudian
disubkultur pada media induksi kalus embriogenik. Rancangan percobaan disusun
secara faktorial dalam lingkungan Rancangan Acak Lengkap dengan 10 ulangan.
Faktor pertama adalah media dasar (MS dan Bac) sedangkan faktor kedua adalah
adenine sulphate (AdS) pada taraf 0, 0.05, dan 0.1 µM (0, 20, dan 40 mg l-1).
Kedua macam media dasar tersebut mengandung senyawa N-organik, yaitu
glutamine (Gln) 6.8 µM (1 mg l-1), casein hydrolysate (CH) 500 mg l-1, arginine
(Arg) 0.69 mM (120 mg l-1), dan glycine (Gly) 0.027 µM (2 mg l-1). Media Bac
mirip dengan media MS terkecuali dalam komposisi sumber nitrogennya, dalam
bentuk KNO3 2528 mg l-1, NH4Cl 535 mg l-1 (Firoozabady dan Moy 2004). Kalus
diinkubasikan dalam ruang kultur bersuhu 25±2 0C, dalam kondisi gelap selama 3
minggu. Peubah yang diamati adalah diameter kalus yang terpendek dan
terpanjang. Pengamatan secara mikroskopis dan analisis histologi (Gambar 10)
dilakukan untuk mengetahui tipe pembelahan dan perkembangan embrio somatik.
Analisis histologi dilakukan dengan menerapkan modifikasi metode Kiernan
(1990).
Pada tahap regenerasi embrio somatik, rancangan percobaan yang
digunakan adalah faktorial dalam lingkungan Rancangan Acak Lengkap. Faktor
pertama adalah media induksi kalus embriogenik dan faktor kedua adalah media
regenerasi. Media induksi kalus embriogenik adalah media MS atau Bac yang
diperkaya dengan senyawa N-organik serta ditambah dengan AdS, seperti
dijelaskan sebelumnya. Media regenerasi yang pertama diberi kode Reg A, yaitu
media MS modifikasi (tanpa nicotinic acid dan pyridoxine HCl) dengan
penambahan IBA 0.9 µM (0.18 mg l-1), BA 1.1 µM (0.25 mg l-1), GA3 0.09 µM
(0.03 mg l-1) menurut Perez et al. (2009). Media regenerasi yang kedua diberi
kode Reg B, yaitu media MS dengan penambahan BA 18 µM (4 mg l-1) menurut
Firoozabady dan Moy (2004). Perlakuan diulang sebanyak 6 kali. Eksplan

diinkubasikan pada ruang kultur dengan 16 jam pencahayaan 1000 lux dan suhu
25±2 0C. Peubah yang diamati adalah persentase pencoklatan dan jumlah embrio
somatik dewasa.
Sampel
Perendaman dalam formalin 4% (3 x 24 jam)
Dehidrasi dengan alkohol 70% (semalam – 3 minggu)
Perendaman dalam alkohol 80% (4 – 24 jam)
Perendaman dalam alkohol 90% ((4 – 24 jam)
Perendaman dalam alkohol 95% (16 – 24jam)
Perendaman dalam alkohol absolut I (1 jam), alkohol absolut II (1 jam), alkohol absolut III (1jam)
Perendaman dalam xilol I (1 jam), xilol II (1 jam), xilol III (30 menit pada suhu ruang dan 30 menit pada suhu 650C)
Inf iltrasi dengan paraf in pada suhu 650C 3kali (1 jam, 40 menit, dan kemudian 40 menit)
Pembuatan blok paraf in
Pemotongan blok dengan microtom 8 µm dan perendalam dalam penangas 380C selama 5 menit
Inkubasi dalam inkubator pada suhu 450C selama 3 hari
Rehidrasi: (jenis dan konsentrasi cairan berkebalikan dengan tahap dehidrasi tetapi waktunya masing-masing 5 menit dan kemudian perendaman dalam air kran selama 10 menit dan dalam akuades selama 5 menit
Pewarnaan dengan haematoxilin selama 2 menit, dan kemudian perendaman dalam air kran selama 10 menit dan dalam akuades selama 5 menit dilanjutkan dengan pewarnaan eosin selama 5 menit
Dehidrasi (alkohol 70 – 90% masing-masing selama 3 detik) dan dalam alkohol absolut I, II, III dan Xilol I, II, III masing-masing selama 1 menit
Penutupan preparat dengan gelas penutup dan entelan
Pengamatan di bawah mikroskop binokular
Gambar 10. Tahapan analisis histologi kalus embriogenik nenas, modifikasi metode Kiernan (1990).

Hasil dan Pembahasan
Regenerasi secara Organogenesis
Setelah 1-2 minggu, potongan tunas menggembung (Gambar 11A), dan
kemudian beberapa daun membuka, sebagai tanda adanya aktivitas ZPT (Gambar
11B). Kalus terbentuk 3 minggu setelah penanaman eksplan (Gambar 11C).
Sebagian besar kalus terinisiasi dari daerah basal dan sebagian kecil terinisiasi
dari daerah luka di sekitar basal daun (Gambar 11D-E). Firoozabady dan Moy
(2004) menduga bahwa area basal daun merupakan area yang meristematik atau
area yang memiliki pembelahan sel yang cepat sehingga mempengaruhi
morfogenesis jaringan. Suryowinoto (1996) juga menyatakan bahwa
Bromeliaceae mempunyai meristem adventif pada area basal daunnya. Munculnya
kalus pada daerah luka kemungkinan disebabkan oleh TDZ dalam media induksi
kalus. TDZ termasuk senyawa serupa sitokinin tipe fenilurea yang mempunyai
aktivitas fisiologi yang tinggi (Sakakibara 2004).
A CB
D E
Gambar 11. Tahapan induksi kalus dari basal daun nenas kultivar Smooth
Cayenne: belahan tunas (A), eksplan basal daun (B), inisiasi kalus (C), kalus terinduksi dari bagian jaringan yang terluka (D), dan kalus terinduksi dari bagian basal daun (D).
Gambar 12 memperlihatkan bahwa formasi kalus semakin meningkat sejak
3 hingga 8 minggu, kemudian statis hingga 12 minggu. Formasi kalus tertinggi
(90%) dihasilkan dari perlakuan 2,4-D 41 µM, namun tidak berbeda nyata dengan
yang dihasilkan dari perlakuan 2,4-D 21 µM. Bobot basah kalus tertinggi

dihasilkan dari perlakuan 2,4-D 21 µM, dan berbeda secara nyata dibandingkan
dengan perlakuan lainnya. Hasil ini lebih baik jika dibandingkan dengan
penelitian sebelumnya yang dilakukan oleh Sripaoraya et al (2003), yang
menghasilkan 58% formasi kalus nenas kultivar Phuket dan juga yang dilakukan
oleh Firoozabady and Moy (2004) yang menghasilkan 55% untuk kultivar Smooth
Cayenne.
aaa
a
a
a
b
ab
a
0
20
40
60
80
100
3 6 8 12Periode inkubasi (minggu)
Pem
bent
ukan
kal
us (%
)
2,4-D21 2,4-D41 2,4-D62
a
a
b
0.00
0.05
0.10
0.15
0.20
0.25
21 41 622,4-D (µM)
Bobo
t bas
ah (g
)
Uji lanjut dilakukan dengan DMRT pada selang kepercayaan 0.05
Gambar 12. Pengaruh 2,4-D terhadap pembentukan kalus dan bobot basah kalus nenas kultivar Smooth Cayenne.
Secara visual, 2,4-D menginduksi kalus kompak dan berwarna kehijau-
hijauan (Gambar 13A-B) daripada kalus friabel dan berwarna kekuning-kuningan
yang biasa dijumpai pada kalus embriogenik. Fenomena pencoklatan juga
ditemukan pada kalus dari perlakuan 2,4-D 41 dan 62 µM (Gambar 13C).
Konsentrasi 2,4-D yang tinggi diduga menyebabkan nekrosis sel karena senyawa
tersebut merupakan herbisida.
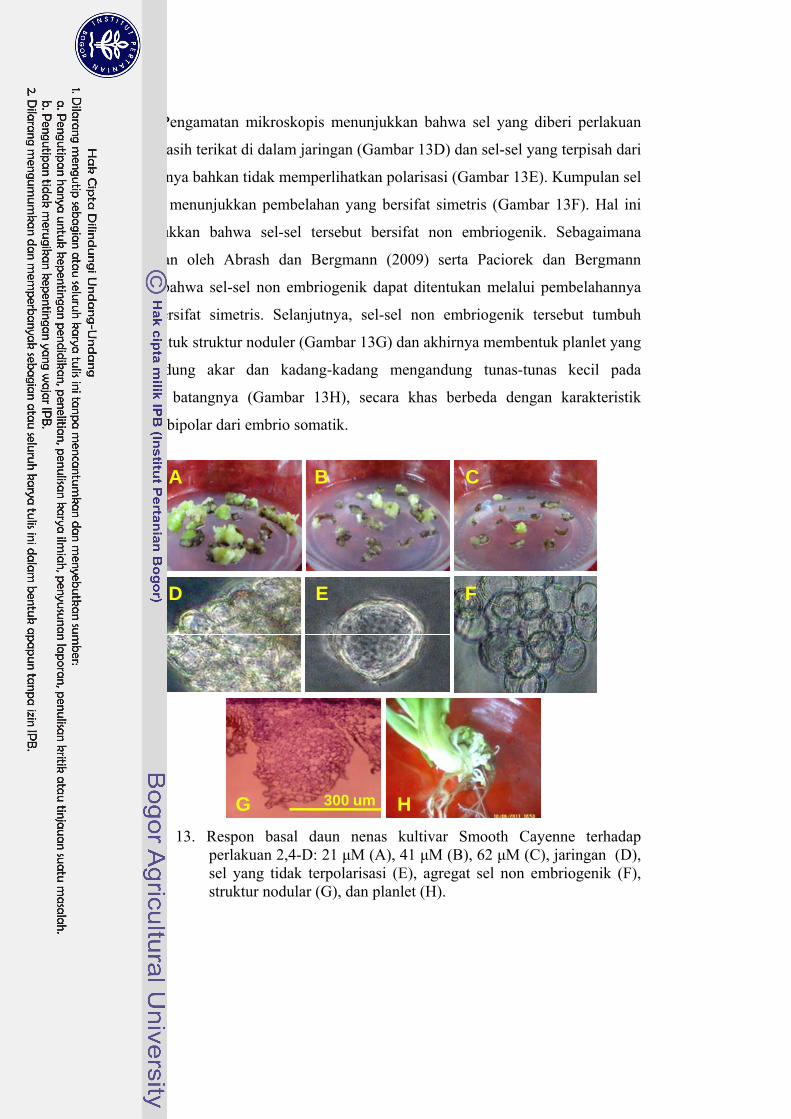
Pengamatan mikroskopis menunjukkan bahwa sel yang diberi perlakuan
2,4-D masih terikat di dalam jaringan (Gambar 13D) dan sel-sel yang terpisah dari
jaringannya bahkan tidak memperlihatkan polarisasi (Gambar 13E). Kumpulan sel
tersebut menunjukkan pembelahan yang bersifat simetris (Gambar 13F). Hal ini
menunjukkan bahwa sel-sel tersebut bersifat non embriogenik. Sebagaimana
dijelaskan oleh Abrash dan Bergmann (2009) serta Paciorek dan Bergmann
(2010) bahwa sel-sel non embriogenik dapat ditentukan melalui pembelahannya
yang bersifat simetris. Selanjutnya, sel-sel non embriogenik tersebut tumbuh
membentuk struktur noduler (Gambar 13G) dan akhirnya membentuk planlet yang
mengandung akar dan kadang-kadang mengandung tunas-tunas kecil pada
pangkal batangnya (Gambar 13H), secara khas berbeda dengan karakteristik
struktur bipolar dari embrio somatik.
FED
300 umG H
CBA
Gambar 13. Respon basal daun nenas kultivar Smooth Cayenne terhadap
perlakuan 2,4-D: 21 µM (A), 41 µM (B), 62 µM (C), jaringan (D), sel yang tidak terpolarisasi (E), agregat sel non embriogenik (F), struktur nodular (G), dan planlet (H).

Pada tahap regenerasi, terdapat perbedaan respon kalus. Peningkatan taraf
2,4-D selama induksi kalus menyebabkan penurunan regenerasi tunas (Gambar
14A-F). Jaringan yang mengalami nekrosis tidak dapat tumbuh lebih lanjut,
seperti pada kalus dari perlakuan 2,4-D 62 µM (Gambar 14C). Tingkat regenerasi
tunas yang tinggi (sekitar 25 tunas/eksplan dalam 2 bulan) dihasilkan dari
perlakuan 2,4-D 21 µM (Gambar 15A).
A B C
D E F
Gambar 14. Penampilan tunas in vitro nenas kultivar Smooth Cayenne hasil
perlakuan 2,4-D pada tahap regenerasi dan elongasi (baris atas dan bawah): 21 µM 2,4-D (A dan D), 41 µM 2,4-D (B dan E), 62 µM 2,4-D (C dan F).
Gambar 15 juga menunjukkan adanya pengaruh negatif dari 2,4-D bertaraf
tinggi, khususnya taraf 62 µM yang menyebabkan tumbuhnya tunas abnormal
(Gambar 15F dan Gambar 15B). Abnormalitas tersebut dapat diamati secara
visual, di mana aksialitas tunas menunjukkan ketidak-seimbangan atau bagian atas
helaian daun membelah menjadi dua bagian (Gambar 15I). Namun demikian,
abnormalitas tersebut bersifat reversible (dapat balik). Tunas-tunas abnormal
tersebut memerlukan tambahan waktu inkubasi untuk tumbuh menjadi normal,
yaitu 3.5 bulan. Lebih dari 90% tunas abnormal berubah menjadi normal (Gambar
15C).

0
5
10
15
20
25
21 41 62
2,4-D (µM)
Juml
ah tu
nas
Normal Abnormal
0
20
40
60
80
100
21 41 622,4-D (µM)
Reg
ener
asi t
unas
(%) Normal Abnormal
0
20
40
60
80
100
21 42 62
2,4-D (µM)
Konv
ersi
tuna
s ab
norm
ake
nor
mal
(%)
Gambar 15. Pengaruh 2,4-D terhadap persentase pembentukan tunas normal,
tunas abnormal, dan konversi dari tunas abnormal ke tunas normal.
Regenerasi secara Embriogenesis Somatik
Karena 2,4-D terbukti tidak mampu menginduksi sel embriogenik maka
selanjutnya kalus disubkultur ke media yang mengandung N-organik, baik dengan
atau tanpa AdS. Firoozabady dan Moy (2004) juga menggunakan formulasi media
yang sama kecuali penggunaan AdS.
Gambar 16 menunjukkan bahwa embrio globular berhasil terbentuk, namun
struktur tersebut saling terikat satu dengan lainnya dan tidak terpisah secara
sempurna. Hal ini menunjukkan bahwa kalus embriogenik terbentuk di dalam
kelompok. Hal serupa juga dilaporkan oleh Firoozabady dan Moy (2004). Struktur
demikian disebut sebagai friable embryogenic tissue (FET) atau jaringan
embriogenik friabel karena individu struktur globular mengelompok dalam
matriks.
Pengamatan mikroskopis dan analisis histologi menunjukkan lemahnya
interaksi antar sel dari perlakuan AdS (Gambar 17A). Kejadian tersebut
menunjukkan dimulainya tahapan embriogenesis. Kejadian yang sama juga
dilaporkan oleh Cunha dan Fernandez-Ferreira (2012). Pelemahan interaksi sel
tersebut mengindikasikan dimulainya inisiasi embriogenesis somatik. Dugaan

tersebut semakin diperkuat dengan dijumpainya sel-sel yang bersifat isodiametrik
dan meristematik yang berwarna merah tua sebagai respon diserapnya
haematoxylin-eosin. Pekatnya warna tersebut disebabkan oleh pekatnya
sitoplasma, besarnya ukuran nukleus (tingginya rasio nukleoplasma), dan kecilnya
ukuran vakuola. Sel-sel meristematik tersebut membentuk agregat yang disebut
sebagai proembryogenic mass (PEM) atau massa proembrio. Pembesaran sel
embriogenik sebesar 400 kali memperlihatkan permukaan yang granular (Gambar
17B) dan tipe pembelahan sel yang asimetris (Gambar 17C). Granular kecokaltan
tersebut kemungkinan disebabkan oleh kandungan pati yang tinggi sebagai tanda
khusus yang biasa dijumpai pada sel embriogenik. Selanjutnya, sel-sel tersebut
berkembang menjadi embrio globular (Gambar 17D). Gambar 17E diduga sebagai
tahap awal dari tahap skutelar karena memiliki struktur yang melancip pada salah
satu ujungnya, namun fase koleoptilar tidak dapat diamati (Gambar 17F). Setelah
5 bulan, sel-sel embriogenik mencoklat dan bercampur dengan sel-sel non
embriogenik. Fakta ini menunjukkan bahwa kompetensi embriogenik hilang
seiring dengan bertambahnya periode in vitro.
A
D E F
B C
Gambar 16. Penampilan jaringan embriogenik nenas kultivar Smooth Cayenne
pada berbagai macam media: MS + N-organik (A), MS + N-organik + Adenin sulfat 0.05 µM (B), MS + N-organik + Adenin sulfat 0.1 µM (C), Bac + N-organik (D), Bac + N-organik + Adenin sulfat 0.05 µM (E), dan Bac + N-organik + Adenin sulfat 0.1 µM (F).
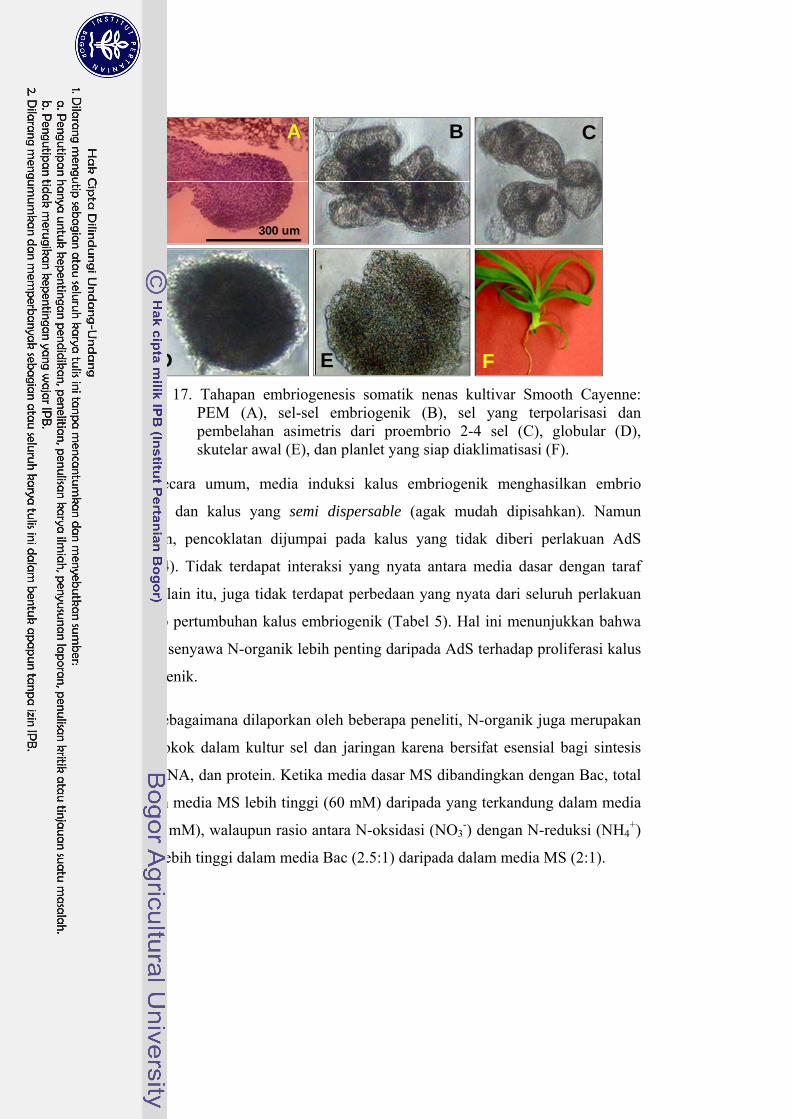
300 um
B CA
E FD
Gambar 17. Tahapan embriogenesis somatik nenas kultivar Smooth Cayenne: PEM (A), sel-sel embriogenik (B), sel yang terpolarisasi dan pembelahan asimetris dari proembrio 2-4 sel (C), globular (D), skutelar awal (E), dan planlet yang siap diaklimatisasi (F).
Secara umum, media induksi kalus embriogenik menghasilkan embrio
globular dan kalus yang semi dispersable (agak mudah dipisahkan). Namun
demikian, pencoklatan dijumpai pada kalus yang tidak diberi perlakuan AdS
(Tabel 4). Tidak terdapat interaksi yang nyata antara media dasar dengan taraf
AdS. Selain itu, juga tidak terdapat perbedaan yang nyata dari seluruh perlakuan
tergadap pertumbuhan kalus embriogenik (Tabel 5). Hal ini menunjukkan bahwa
peranan senyawa N-organik lebih penting daripada AdS terhadap proliferasi kalus
embriogenik.
Sebagaimana dilaporkan oleh beberapa peneliti, N-organik juga merupakan
unsur pokok dalam kultur sel dan jaringan karena bersifat esensial bagi sintesis
DNA, RNA, dan protein. Ketika media dasar MS dibandingkan dengan Bac, total
N dalam media MS lebih tinggi (60 mM) daripada yang terkandung dalam media
Bac (35 mM), walaupun rasio antara N-oksidasi (NO3-) dengan N-reduksi (NH4
+)
sedikit lebih tinggi dalam media Bac (2.5:1) daripada dalam media MS (2:1).
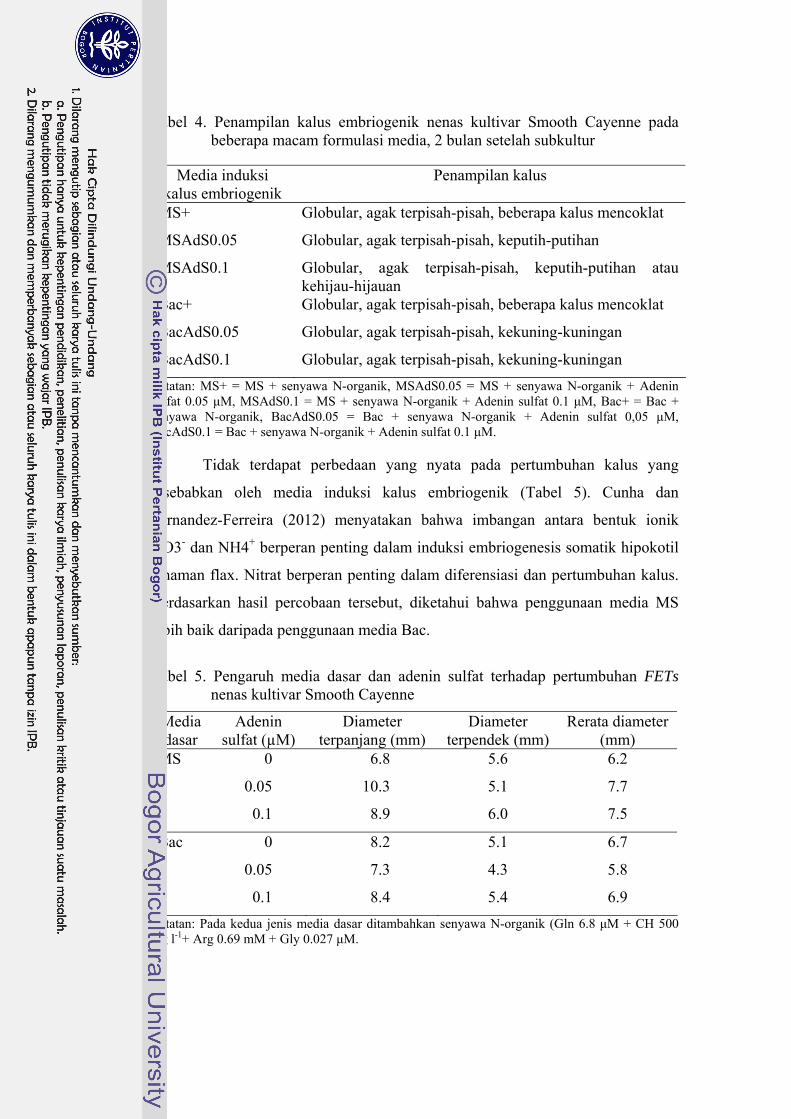
Tabel 4. Penampilan kalus embriogenik nenas kultivar Smooth Cayenne pada beberapa macam formulasi media, 2 bulan setelah subkultur
Media induksi kalus embriogenik
Penampilan kalus
MS+ Globular, agak terpisah-pisah, beberapa kalus mencoklat
MSAdS0.05 Globular, agak terpisah-pisah, keputih-putihan
MSAdS0.1 Globular, agak terpisah-pisah, keputih-putihan atau kehijau-hijauan
Bac+ Globular, agak terpisah-pisah, beberapa kalus mencoklat
BacAdS0.05 Globular, agak terpisah-pisah, kekuning-kuningan
BacAdS0.1 Globular, agak terpisah-pisah, kekuning-kuningan Catatan: MS+ = MS + senyawa N-organik, MSAdS0.05 = MS + senyawa N-organik + Adenin sulfat 0.05 µM, MSAdS0.1 = MS + senyawa N-organik + Adenin sulfat 0.1 µM, Bac+ = Bac + senyawa N-organik, BacAdS0.05 = Bac + senyawa N-organik + Adenin sulfat 0,05 µM, BacAdS0.1 = Bac + senyawa N-organik + Adenin sulfat 0.1 µM.
Tidak terdapat perbedaan yang nyata pada pertumbuhan kalus yang
disebabkan oleh media induksi kalus embriogenik (Tabel 5). Cunha dan
Fernandez-Ferreira (2012) menyatakan bahwa imbangan antara bentuk ionik
NO3- dan NH4+ berperan penting dalam induksi embriogenesis somatik hipokotil
tanaman flax. Nitrat berperan penting dalam diferensiasi dan pertumbuhan kalus.
Berdasarkan hasil percobaan tersebut, diketahui bahwa penggunaan media MS
lebih baik daripada penggunaan media Bac.
Tabel 5. Pengaruh media dasar dan adenin sulfat terhadap pertumbuhan FETs nenas kultivar Smooth Cayenne
Media dasar
Adenin sulfat (µM)
Diameter terpanjang (mm)
Diameter terpendek (mm)
Rerata diameter (mm)
MS 0 6.8 5.6 6.2
0.05 10.3 5.1 7.7
0.1 8.9 6.0 7.5
Bac 0 8.2 5.1 6.7
0.05 7.3 4.3 5.8
0.1 8.4 5.4 6.9 Catatan: Pada kedua jenis media dasar ditambahkan senyawa N-organik (Gln 6.8 µM + CH 500 mg l-1+ Arg 0.69 mM + Gly 0.027 µM.
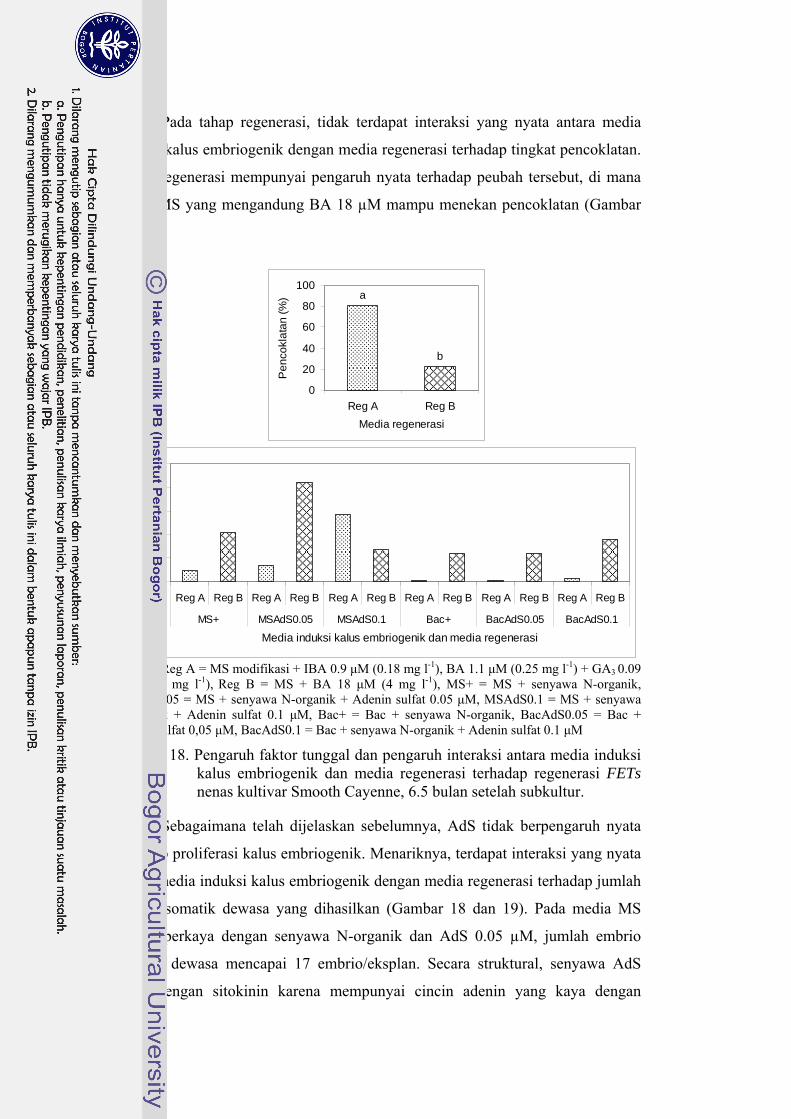
Pada tahap regenerasi, tidak terdapat interaksi yang nyata antara media
induksi kalus embriogenik dengan media regenerasi terhadap tingkat pencoklatan.
Media regenerasi mempunyai pengaruh nyata terhadap peubah tersebut, di mana
media MS yang mengandung BA 18 µM mampu menekan pencoklatan (Gambar
18).
b
a
0
20
40
60
80
100
Reg A Reg BMedia regenerasi
Pen
cokl
atan
(%)
0
4
8
12
16
20
Reg A Reg B Reg A Reg B Reg A Reg B Reg A Reg B Reg A Reg B Reg A Reg B
MS+ MSAdS0.05 MSAdS0.1 Bac+ BacAdS0.05 BacAdS0.1
Media induksi kalus embriogenik dan media regenerasi
Jum
lah e
mbr
io so
mat
ik de
wasa
Catatan: Reg A = MS modifikasi + IBA 0.9 µM (0.18 mg l-1), BA 1.1 µM (0.25 mg l-1) + GA3 0.09 µM (0.03 mg l-1), Reg B = MS + BA 18 µM (4 mg l-1), MS+ = MS + senyawa N-organik, MSAdS0.05 = MS + senyawa N-organik + Adenin sulfat 0.05 µM, MSAdS0.1 = MS + senyawa N-organik + Adenin sulfat 0.1 µM, Bac+ = Bac + senyawa N-organik, BacAdS0.05 = Bac + Adenin sulfat 0,05 µM, BacAdS0.1 = Bac + senyawa N-organik + Adenin sulfat 0.1 µM
Gambar 18. Pengaruh faktor tunggal dan pengaruh interaksi antara media induksi kalus embriogenik dan media regenerasi terhadap regenerasi FETs nenas kultivar Smooth Cayenne, 6.5 bulan setelah subkultur.
Sebagaimana telah dijelaskan sebelumnya, AdS tidak berpengaruh nyata
terhadap proliferasi kalus embriogenik. Menariknya, terdapat interaksi yang nyata
antara media induksi kalus embriogenik dengan media regenerasi terhadap jumlah
embrio somatik dewasa yang dihasilkan (Gambar 18 dan 19). Pada media MS
yang diperkaya dengan senyawa N-organik dan AdS 0.05 µM, jumlah embrio
somatik dewasa mencapai 17 embrio/eksplan. Secara struktural, senyawa AdS
mirip dengan sitokinin karena mempunyai cincin adenin yang kaya dengan

nitrogen. Senyawa AdS kemungkinan berperan sebagai sumber nitrogen dalam
sintesis RNA, DNA, dan protein untuk pembelahan sel. Oleh karena itu, senyawa
tersebut mempengaruhi perkembangan embrio somatik.
A
G
FED
CB
IH
KJ L
Gambar 19. Perbedaan perkembangan embrio somatik nenas kultivar Smooth
Cayenne umur 8 bulan pada media Reg A ( dua baris atas) dan Reg B (dua baris bawah): MS + senyawa N-organik (A dan G), MS + senyawa N-organik + Adenin sulfat 0.05 µM (B dan H), MS + senyawa N-organik + Adenin sulfat 0.1 µM (C dan I), Bac + senyawa N-organik (D dan J), Bac + senyawa N-organik + Adenin sulfat 0.05 µM (E and K), dan Bac + senyawa N-organik + Adenin sulfat 0.1 µM (F dan L).

Simpulan
Penggunaan 2,4-D secara tunggal tidak mampu menginduksi sel
embriogenik sehingga sebagian kalus non embriogenik diregenerasikan secara
organogenesis. Media terbaik untuk organogenesis adalah 2,4-D 21 µM karena
menghasilkan persentase pembentukan kalus yang tinggi (80%), bobot basah
kalus tertinggi (0.2 g/eksplan), dan jumlah tunas tertinggi (sekitar 25
tunas/eksplan/2 bulan). Kalus embriogenik terbentuk ketika kalus non
embriogenik diperlakukan dengan senyawa N-organik. Media regenerasi
merupakan faktor yang berpengaruh nyata terhadap tingkat pencoklatan. Media
MS dengan penambahan BA 18 µM mampu menekan pencoklatan tersebut.
Terdapat interaksi yang nyata antara media induksi kalus embriogenik dengan
media regenerasi terhadap jumlah embrio somatik dewasa. Media MS yang
diperkaya dengan senyawa N-organik dan AdS 0.05 µM yang dikombinasikan
dengan media MS dengan penambahan BA 18 µM merupakan perlakuan terbaik
untuk induksi kalus embriogenik dan regenerasi embrio somatik nenas (17 embrio
dewasa/eksplan) dalam waktu 6.5 bulan.
Daftar Pustaka
Abrash, E.B. and D.C. Bergmann. 2009. Asymmetric cell divisions: A view from plant development. Developmental Cell 16: 783-796.
Arnold S, Sabala I, Bozhkov P, Dyachok J, Filonova L. 2002. Developmental pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult 69: 233-249.
Coppens d’Eeckenbrugge G, Leal F. 2003. Morphology, anatomy, and taxonomy. p. 13-32. In: Bartholomew DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany: Production and Uses. CABI Publishing.Wallingford.
Corredoira E, Valladares S, Vieitez AM. Morphohistological analysis of the origin and development of somatic embryos from leaves of mature Quercus robur. In Vitro Cell Dev Biol_Plant 42:525-533.
Cunha A, Fernandez-Ferreira M. 2012. Influence of medium parameters on somatic embryogenesis from hypocotyl explants of flax (Linum usitatissimum L.): Effect of carbon source, total inorganic nitrogen and balance between ionic forms and interaction between calcium and zeatin. http://dx.doi.org/10.1016/S0176-1617(99)80059-5.

Firoozabady E, Moy Y. 2004. Regeneration of pineapple via somatic embryogenesis and organogenesis. In vitro Cell Dev Biol-Plant 40:67-74.
[IBPGR] International Board for Plant Genetic Resources. 1991. Descriptors for Pineapple. IBPGR Headquarters, Rome
Kelley KB, Riechers DE. 2007. Recent development in auxin biology and new properties for auxinic herbicide research. Pesticide Biochemistry and Physiology 89: 1-11.
Kiernan JA. 1990. Histological and Histochemical Methods: Theory and Practice. 2nd edition. Pergamon Press.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15: 473-497.
Nursandi F, Poerwanto R, Sobir, Sujiprihati S. 2003. Perbanyakan in vitro nenas (Ananas comosus (L.) Merr.) dengan menggunakan BAP dan TDZ. Dalam: Simposium Nasional dan Konggres VIII. Bandar Lampung.
Nursandi F, Sobir, Murtini. 2005. Perbanyakan tanaman nenas (Ananas comosus (L.) Merr.) cv. Smooth Cayenne dengan teknik etiolasi secara in vitro. Tropika 13(1):15-24.
Paciorek T, Bergmann DC. 2010. The secret to life is being different: asymmetric divisions in plant development. Current Opinion in Plant Biology 13:661–669
Perez G, Yanes E, Isidron M, Lorenzo JC. 2009. Phenotypic and AFLP characterization of two new pineapple somaclones derived from in vitro culture. Plant Cell Tissue Organ Cult 96:113-116.
Phillips GC, Hubstenberger JF, Hansen EE. 1995. Plant regeneration from callus and cell suspension cultures by somatic embryogenesis. Pp. 81 - 90. In: Gamborg OL, Phillips GC, editor. Plant Cell Tissue and Organ Culture: Fundamental Methods. Springer. Berlin.
Purnamaningsih R, Mariska I, Supriati Y. 2009. Penggunaan ABA dan paklobutrazol dalam perbanyakan nenas Simadu melalui kultur in vitro. Ber Bio 9(6): 751-758.
Sakakibara. H. 2004. Cytokinin biosynthesis and metabolism. Pp. 95-114. In. Davies, P.J. (Ed.). Plant Hormones: Biosynthesis, Signal, Action. 3rd Edition. Kluwer Academic Publishers. London.

Smith MK, Ko H-L, Hamil SD, Sanewski GM, Graham MW. 2003. Biotechnology. Pp. 57-68. In: Bartholomew DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany: Production and Uses. CABI Publishing.Wallingford.
Sripaoraya S, Marchamt R, Power JB, Davey MR. 2003. Plant regeneration by somatic embryogenesis and organogenesis in commercial pineapple (Ananas comosus L.). In Vitro Cell Dev Biol_Plant 39:450-454.
Srivastava LM. 2002. Plant Growth and Development. Academic Press. New York. 772p.
Suminar E. 2010. Induksi keragaman genetik dengan mutagen sinar gamma pada nenas (Ananas comosus (L.) Merr.) secara in vitro [seminar]. Sekolah Pascasarjana, Institut Pertanian Bogor. Bogor.
Suryowinoto M. 1996. Pemuliaan Tanaman secara In Vitro. Pusat Antar Universitas-Bioteknologi UNiversitas Gadjah Mada. Yogyakarta. 252 hal.

INDUKSI EMBRIOGENESIS SOMATIK TIDAK LANGSUNG PADA NENAS MELALUI PENGGUNAAN PIKLORAM
Abstrak
Embriogenesis somatik merupakan teknik yang banyak digunakan untuk perbanyakan tanaman secara klonal dan masal. Tujuan penelitian ini adalah: (1) menentukan taraf pikloram yang terbaik untuk induksi kalus, (2) mengetahui pengaruh interaksi antara perlakuan media induksi kalus embriogenik dan pencahayaan terhadap induksi kalus embriogenik, (3) mengetahui perlakuan terbaik untuk pembentukan suspensi sel, dan (4) menentukan perlakuan kombinasi yang terbaik untuk regenerasi kalus embriogenik. Induksi kalus dilakukan dengan menggunakan pikloram (21, 41, dan 62 µM) dengan penambahan TDZ 9 µM. Selanjutnya, kalus dipindahkan ke media MS atau Bac yang mengandung senyawa N-organik (Gln 6.8 µM + CH 500 mg l-1 + Arg 0.69 mM + Gly 0.027 µM) dengan atau tanpa penambahan pikloram 21 µM pada kondisi tanpa pencahayaan atau dengan pencahayaan untuk induksi kalus embriogenik. Sebagian kalus embriogenik dari perlakuan MS atau Bac yang ditambah pikloram 21 µM disuspensikan dalam media cair. Kalus yang semi kompak dipindahkan ke media MS yang mengandung Kn 1 mg l-1 untuk regenerasi dan elongasi embrio prematur, sedangkan kalus yang remah dipindahkan ke media MS dengan penambahan BA 18 µM atau media MS modifikasi yang ditambah dengan BA 1.1 µM, IBA 0.9 µM, dan GA3 0.09 µM untuk perkembangan embrio. Hasil penelitian menunjukkan bahwa embriogenesis somatik berasal dari sel tunggal. Perlakuan terbaik untuk induksi kalus adalah pikloram 21 µM. Tidak terdapat interaksi yang nyata antara media induksi kalus embriogenik dan kondisi inkubasi terhadap pencoklatan serta pembentukan nodul dan embrio. Media terbaik untuk induksi dan proliferasi kalus embriogenik adalah media MS yang diperkaya dengan senyawa N-organik dan pikloram 21 µM sedangkan kondisi inkubasi terbaik untuk induksi dan proliferasi kalus embriogenik adalah kondisi tanpa pencahayaan. Regenerasi ETs yang terbaik dihasilkan dari media Bac yang diperkaya dengan senyawa N-organik tanpa penambahan pikloram dan diinkubasikan pada kondisi dengan pencahayaan. Jumlah embrio somatik dewasa dari ETs mencapai 17 embrio/eksplan dalam waktu 2 bulan dan lebih dari 30 embrio/eksplan dalam waktu 4 bulan. Regenerasi FETs yang terbaik dihasilkan dari media MS dengan penambahan BA 18 µM, dengan perlakuan asal berupa media MS yang diperkaya dengan senyawa N-organik dan pikoram 21 µM yang diinkubasikan pada kondisi dengan pencahayaan. Jumlah embrio somatik dewasa dari FETs adalah 14 embrio/eksplan dalam waktu 2.5 bulan.
Kata kunci: Embriogenesis somatik, nenas, pikloram, senyawa N-organik, pencahayaan

Abstract
Somatic embryogenesis is a desirable method for plant regeneration, especially for large-scale clonal propagation. The objectives of this research were: (1) to determine the best concentration of picloram for callus induction, (2) to know the interaction effect of embryogenic callus induction media and lighting, (3) to know the best treatment for cell suspension culture, and (4) to obtain the best combination treatment for embryogenic callus regeneration. Callus formation was induced by picloram (21, 41, and 62 µM) with addition of 9 µM TDZ. The calli were transferred onto N-enriched MS or Bac medium (6.8 µM Gln + 500 mg l-1 CH + 0.69 mM Arg + 0.027 µM Gly) with or without addition of 21 µM picloram under light or without light condition for embryogenic callus formation. Some of the embryogenic calli from MS or Bac media with addition of 21 µM picloram were then cultured in liquid media to form cell suspension. The semi compact calli were subcultured onto MS medium supplemented with 1 mg l-1 Kn for regeneration and elongation of premature somatic embryos while the friable calli were transferred onto MS medium supplemented with 18 µM BA or modified MS medium with addition of 1.1 µM BA, 0.9 µM IBA, 0.09 µM GA3 for embryo development. The result showed that somatic embryogenesis was unicellular origin. The best medium for callus induction was MS media enriched with 21 µM picloram. There was no interaction between embryogenic callus induction media with condition of incubation. The best treatment for embryogenic callus induction and proliferation was MS medium enriched with N-organic compounds and supplemented with 21 µM picloram. The best condition for embryogenic callus induction was without light condition. There was interaction between embyogenic callus induction media and condition of incubation to the regeneration of ETs. The best regeneration of ETs resulted from Bac medium enriched with N-organic compounds and incubated under light condition. The number of somatic embryos resulted from that treatment was up to 17 embryos/explant in 2 months or more than 30 embryos/explant in 4 months. The best treatment for regenerating FETs was MS medium containing of BA 18 µM. Those embryos (14 embryos/explant in 2.5 months) were yielded from MS medium enriched with N-organic compounds and 21 µM picloram under light condition.
Keywords: Somatic embryogenesis, pineapple, picloram, N-organic compounds, lighting
Pendahuluan
Embriogenesis somatik merupakan metode perbanyakan masal yang banyak
diterapkan pada berbagai macam tanaman, seperti kelapa (Buffard-Morel 1995),
pisang (Sidha et al 2007), stroberi (Kordestani and Karami 2008), kopi (Oktavia
et al 2003; Gatica et al 2008), dan jahe (Lincy et al 2009). Teknik tersebut juga
telah diterapkan pada tanaman nenas (Sripaoraya et al. 2003; Firoozabady and

Moy 2004), namun informasi tentang tahapan lengkap dari proses embriogenesis
somatik tersebut masih terbatas, khususnya pembahasan pada tahapan awal dari
inisiasi sel embriogenik. Menurut Corredoira et al (2006), pemahaman tentang
embriogenesis tanaman penting diketahui, yaitu melalui pengamatan proses
seluler yang mendasari diferensiasi. Hal tersebut akan bermanfaat untuk
memperoleh metode yang mantap untuk aplikasi perbanyakan bibit secara masal,
kriopreservasi, dan transformasi genetik.
Di Indonesia, teknik kultur in vitro culture tanaman nenas kultivar Smooth
Cayenne masih cukup terbatas. Beberapa penelitian membahas topik proliferasi
tunas (Nursandi et al. 2003; Purnamaningsih et al. 2009), etiolasi tunas (Nursandi
et al. 2003), dan mutagenesis (Suminar 2010).
Dilaporkan bahwa auksin memberikan pengaruh pada perkembangan
tanaman pada tingkat seluler, termasuk mempengaruhi proses embriogenesis
(Kelley and Riechers 2007; Vanneste and Frim 2009). Pada dosis yang rendah,
auksin herbisida dilaporkan memiliki fungsi hormonal yang mirip dengan auksin
alami (Kelley and Riechers 2007).
Pada percobaan sebelumnya telah digunakan senyawa dichlorophenoxy
acetic acid (2,4-D). Hasil percobaan tersebut menunjukkan bahwa secara tunggal,
2,4-D tidak mampu menginisiasi sel embriogenik. Induksi sel dan kalus
embriogenik baru dapat dilakukan setelah ditambahkan beberapa macam senyawa
nitrogen organik (N-organik), seperti glutamin, arginin, kasein hidolisat, dan
glisin.
Selain 2,4-D, 4-amino 3,5,6-trichloropicolinic acid (pikloram) juga
merupakan auksin herbisida. Senyawa tersebut merupakan auksin dari kelompok
piridin dan biasa digunakan pada konsentrasi rendah sebagai zat pengatur tumbuh
(ZPT).
Selain auksin, kondisi pencahayaan juga memegang peranan penting dalam
regenerasi embriogenesis somatik. Uozumi et al. (19933) melaporkan bahwa
pencahayaan mempengaruhi regenerasi dan perkembangan embrio seledri. Gatica

et al. (2008) juga melaporkan peranan penting dari pencahayaan selama proses
embriogenesis kopi. Tujuan penelitian ini adalah: (1) menentukan taraf pikloram
yang terbaik untuk induksi kalus, (2) mengetahui pengaruh interaksi antara
perlakuan media induksi kalus embriogenik dan pencahayaan terhadap induksi
kalus embriogenik, (3) menentukan perlakuan kombinasi yang terbaik untuk
regenerasi kalus embriogenik, dan (4) mengetahui perlakuan terbaik untuk
pembentukan suspensi sel.
Bahan dan Metode
Bahan Tanaman dan Pemeliharaan
Bahan tanaman yang digunakan dalam penelitian ini adalah tunas in vitro
tanaman nenas kultivar Smooth Cayenne klon Simadu dari Subang. Kultur
tersebut diperoleh dari isolasi dan inisiasi mata tunas yang terdapat pada mahkota
nenas. Pemeliharaan kultur dilakukan dengan menggunakan media MS
(Murashige and Skoog 1962) dengan penambahan benzyl adenine (BA) 2.21 µM
(0.5 mg l-1) dan kinetin (Kn) 4.65 µM (1 mg l-1). Kemasaman media (pH ) ditera
pada nilai 5.7±0.1 sebelum dimasukkan ke dalam otoklaf. Inkubasi dilakukan
pada ruang kultur dengan suhu 25 0C, fotoperiodisitas 16 jam dengan
pencahayaan dengan intensitas penyinaran 800 – 1000 lux. Penelitian dibagi atas
tiga tahapan, yaitu (1) Induksi kalus, (2) Induksi kalus embriogenik, dan (3)
Pembentukan suspensi sel, dan Perkembangan embrio somatik (4).
Induksi Kalus
Tunas in vitro sekitar 4–6 cm dipotong secara longitudinal menjadi dua
bagian, kemudian ditanam secara horizontal pada media induksi kalus (10 bagian
per botol) dan diinkubasikan pada kondisi remang-remang (di bawah 500 lux).
Media induksi kalus adalah media MS yang mengandung pikloram pada taraf 21,
41, dan 62 µM (5, 10, dan 15 mg l-1) dengan penambahan thidiazuron (TDZ) pada
taraf 9 µM (2 mg l-1). Eksplan diinkubasikan pada suhu 25±2 0C dalam keadaan
tanpa pencahayaan. Setelah eksplan membesar (3 minggu), helaian daun diisolasi
secara individual dan kemudian ditanam pada media yang sama. Inkubasi juga

dilakukan pada kondisi yang sama dengan tahap sebelumnya. Rancangan
percobaan adalah Rancangan Acak Lengkap dengan 5 ulangan (botol). Setiap
botol terdiri atas 15 eksplan. Pembentukan kalus diamati pada minggu ke-3, 6, 8,
dan 12. Bobot basah kalus diukur pada akhir pengamatan. Data dianalisis dengan
program SPSS V.19 (IBM Company) dan perbedaan antar rerata dibandingkan
dengan uji Duncans’s multiple range tests (DMRT) pada tingkat kepercayaan
0.05. Penampilan visual kalus juga diamati (kompak atau remah, putih atau
kuning atau hijau, mudah dipisahkan atau tidak, berair atau vigor). Pengamatan
secara mikroskopis dilakukan untuk menentukan tipe pembelahan sel (simetris
atau asimetris).
Induksi dan Proliferasi Kalus Embriogenik
Eksplan yang digunakan adalah kalus yang dihasilkan dari perlakuan
pikloram 21 µM. Kalus tersebut disubkultur pada berbagai macam formulasi
media untuk menginduksi kalus embriogenik, yaitu 4 perlakuan. Rancangan
percobaan disusun secara faktorial dalam lingkungan Rancangan Acak Lengkap
dengan 8 ulangan. Faktor pertama adalah pikoram (tanpa dan penambahan
pikloram 21 µM) sedangkan faktor kedua adalah media dasar (MS dan Bac).
Media Bac mirip dengan media MS, kecuali pada komposisi dan sumber
nitrogennya, yatiu KNO3 bertaraf 2528 mg l-1, dan NH4Cl 535 bertaraf mg l-1
(Firoozabady dan Moy (2004). Seluruh media mengandung senyawa N-organik,
yaitu glutamine (Gln) 6.8 µM (1 mg l-1), casein hydrolysate (CH) 500 mg l-1,
arginine (Arg) 0.69 mM (120 mg l-1), dan glycine (Gly) 0.027 µM (2 mg l-1).
Kalus diinkubasi dilakukan pada suhu 25±2 0C dalam kondisi dengan atau tanpa
pencahayaan (800-1000 lux) selama 3 minggu. Peubah yang diamati adalah
pembentukan kalus embriogenik, pembentukan akar, dan penampilan kalus
(kompak atau remah, putih atau kuning atau hijau, mudah dipisahkan atau sulit
dipisahkan, berair atau tidak, dan nodular atau globular. Data dianalisis dengan
program SPSS V.19 (IBM Company). Pengamatan mikroskopis dilakukan dengan
menggunakan converted stereomicroscope untuk mengetahui tipe pembelahan sel
(simetris atau asimetris). Analisis histologi dilakukan dengan menerapkan
modifikasi metode Kiernan (1990).

Pembentukan Suspensi Sel
Eksplan yang digunakan adalah kalus embriogenik yang dihasilkan dari
perlakuan pada tahap sebelumnya. Kalus tersebut disubkultur ke media cair
dengan formulasi media sesuai dengan tahapan sebelumnya, yaitu media MS dan
Bac dengan penambahan pikloram 21 µM. Inkubasi dilakukan pada kondisi tanpa
pencahayaan dan dengan pencahayaan (16 jam dengan intensitas cahaya 800 lux)
dengan suhu 25±2 0C. Peubah yang diamati adalah persentase pencoklatan,
persentase pembentukan suspensi, dan jumlah embrio somatik. Data ditampilkan
dalam bentuk rerata dan standar deviasi.
Perkembangan Embrio Somatik
Pada tahap regenerasi, kalus kompak dipindahkan ke media MS dengan
penambahan Kn 4.65 µM (1 mg l-1) untuk regenerasi dan elongasi embrio
prematur. Kalus remah dipindahkan ke dua macam media. Rancangan percobaan
yang digunakan adalah faktorial dalam lingkungan Rancangan Acak Lengkap.
Faktor pertama adalah media induksi kalus embriogenik dan faktor kedua adalah
media regenerasi. Media induksi kalus embriogenik adalah media MS atau Bac
yang diperkaya dengan senyawa N-organik serta ditambah dengan AdS, seperti
dijelaskan sebelumnya. Media regenerasi yang pertama diberi kode Reg A, yaitu
media MS modifikasi (tanpa nicotinic acid dan pyridoxine HCl) dengan
penambahan IBA 0.9 µM (0.18 mg l-1), BA 1.1 µM (0.25 mg l-1), GA3 0.09 µM
(0.03 mg l-1) menurut Perez et al. (2009). Media regenerasi yang kedua diberi
kode Reg B, yaitu media MS dengan penambahan BA 18 µM (4 mg l-1) menurut
Firoozabady dan Moy (2004). Perlakuan diulang sebanyak 8 kali. Eksplan
diinkubasikan pada ruang kultur dengan 16 jam pencahayaan 1000 lux dan suhu
25±2 0C. Peubah yang diamati adalah persentase pencoklatan, persentase eksplan
yang beregenerasi, dan jumlah embrio somatik dewasa serta penampilan visual
kalus. Data dianalisis dengan program SPSS V.19 (IBM Company).

Hasil dan Pembahasan
Induksi Kalus
Pembentukan kalus dimulai pada minggu ke-3 setelah tanam. Hampir semua
eksplan membentuk kalus pada daerah basal daun. Firoozabady dan Moy (2004)
menduga bahwa area basal daun merupakan area yang meristematik atau area
yang memiliki pembelahan sel secara cepat sehingga mempengaruhi morfogenesis
jaringan. Suryowinoto (1996) juga menyatakan bahwa Bromeliaceae mempunyai
meristem adventif pada area basal daunnya.
Hasil penelitian menunjukkan bahwa kalus tumbuh hingga minggu ke-8, dan
pertumbuhannya berhenti pada periode inkubasi berikutnya hingga 12 minggu.
Tingkat pembentukan kalus yang tertinggi (90%) diperoleh dari perlakuan
pikloram 21 µM walaupun tidak berbeda nyata dengan perlakuan lainnya
(Gambar 20). Demikian pula, bobot basah kalus tidak berbeda nyata secara
statistik (Gambar 20). Dengan demikian, dapat disimpulkan bahwa penggunaan
pikoram 21 µM merupakan perlakuan yang paling efisien. Hasil ini lebih baik jika
dibandingkan dengan penelitian sebelumnya yang dilakukan oleh Sripaoraya et al
(2003), yang menghasilkan 58% formasi kalus nenas kultivar Phuket dan juga
yang dilakukan oleh Firoozabady and Moy (2004) yang menghasilkan 55% untuk
kultivar Smooth Cayenne. Respon eksplan terhadap pikloram bahkan lebih baik
daripada responnya terhadap 2,4-D (pada Bab sebelumnya). Pada taraf yang tinggi
(62 µM), pikloram mampu menginduksi kalus dengan persentase sekitar 80%. Hal
ini menunjukkan bahwa fitotoksisitas pikloram lebih rendah daripada 2,4-D.
Secara visual tampak bahwa eksplan membentuk kalus yang globuler dan
berwarna kekuning-kuningan, yang terbentuk dari daerah basal daun. Pengamatan
secara mikroskopis menunjukkan bahwa kalus tersebut merupakan campuran
antara sel-sel embriogenik dan non embriogenik. Sel-sel embriogenik
memperlihatkan polaritas yang diikuti dengan pembelahan asimetris menjadi
bagian apikal dan basal (Gambar 21). Abrash dan Bergmann (2009) melaporkan
bahwa pembelahan asimetris menghasilkan sel-sel anakan yang berbeda secara
morfologi. Selanjutnya, Paciorek dan Bergmann (2010) menggambarkan proses
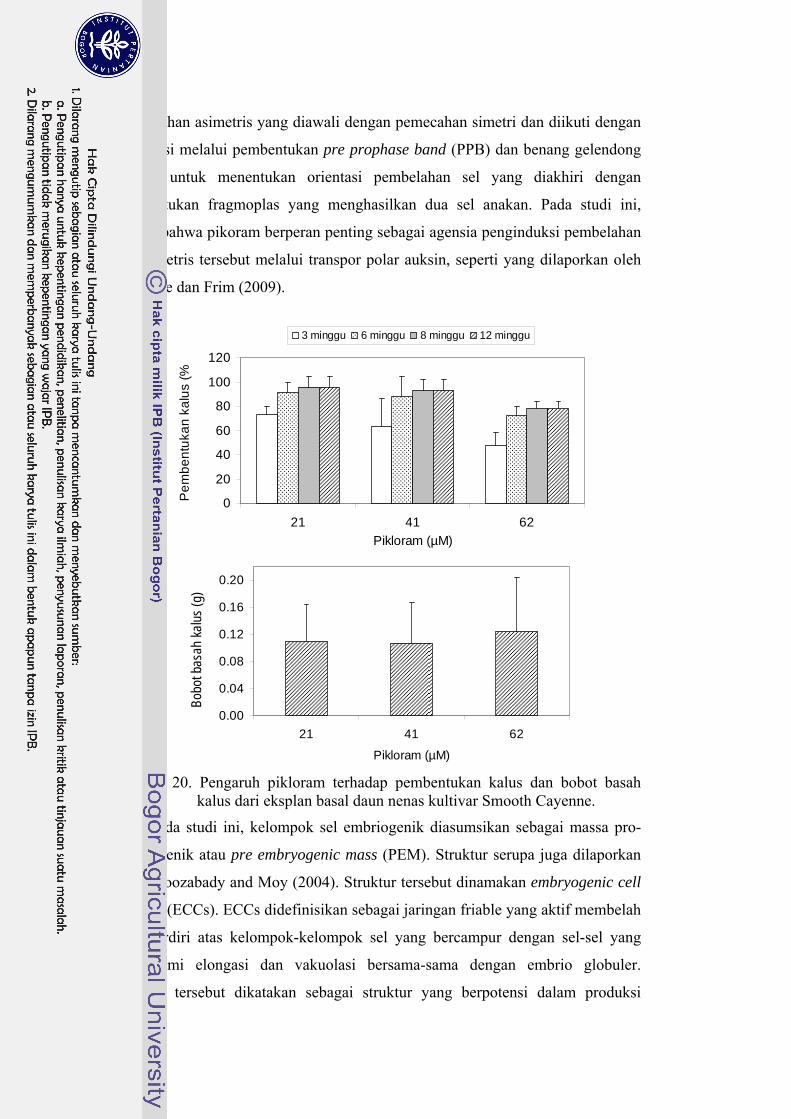
pembelahan asimetris yang diawali dengan pemecahan simetri dan diikuti dengan
polarisasi melalui pembentukan pre prophase band (PPB) dan benang gelendong
mitotic untuk menentukan orientasi pembelahan sel yang diakhiri dengan
pembentukan fragmoplas yang menghasilkan dua sel anakan. Pada studi ini,
diduga bahwa pikoram berperan penting sebagai agensia penginduksi pembelahan
sel asimetris tersebut melalui transpor polar auksin, seperti yang dilaporkan oleh
Vanneste dan Frim (2009).
0
20
40
60
80
100
120
21 41 62Pikloram (µM)
Pem
bent
ukan
kal
us (%
3 minggu 6 minggu 8 minggu 12 minggu
0.00
0.04
0.08
0.12
0.16
0.20
21 41 62
Pikloram (µM)
Bobo
t bas
ah ka
lus (g
)
Gambar 20. Pengaruh pikloram terhadap pembentukan kalus dan bobot basah
kalus dari eksplan basal daun nenas kultivar Smooth Cayenne.
Pada studi ini, kelompok sel embriogenik diasumsikan sebagai massa pro-
embriogenik atau pre embryogenic mass (PEM). Struktur serupa juga dilaporkan
oleh Firoozabady and Moy (2004). Struktur tersebut dinamakan embryogenic cell
clusters (ECCs). ECCs didefinisikan sebagai jaringan friable yang aktif membelah
yang terdiri atas kelompok-kelompok sel yang bercampur dengan sel-sel yang
mengalami elongasi dan vakuolasi bersama-sama dengan embrio globuler.
Struktur tersebut dikatakan sebagai struktur yang berpotensi dalam produksi

embrio somatik. Sayangnya, struktur tersebut hilang ketika dibiarkan lebih lama
(2 bulan) dalam media yang mengandung pikoram secara terus-menerus.
A B C
D E F
Gambar 21. Respon basal daun nenas kultivar Smooth Cayenne terhadap
perlakuan pikloram dan pengamatan mikroskopisnya: 21 µM (A), 41 µM (B), 62 µM (C), kelompok sel non embriogenik (D), sel yang terpolarisasi dan yang belum terpolarisasi (E), dan sel embriogenik yang terdiri atas bagian basal dan apikal (F). Tanda panah menunjukkan tahap perkembangan lanjut dari masing-masing sel yang ditunjuk.
Induksi dan Proliferasi Kalus Embriogenik
Karakter sel embriogenik dapat distimulir kembali dengan memindahkan
kalus ke media yang mengandung senyawa N-organik. Pada awalnya, polaritas sel
terjadi yang diikuti dengan pembelahan asimteris dan dilanjutkan dengan elongasi
sel, sebagai bukti terjadinya regenerasi secara embriogenesis somatik. Arnold et
al. (2002) menyatakan bahwa mekanisme penting selama proses embriogenesis
somatik adalah pembelahan sel secara asimetris dan elongasi sel. Kejadian
tersebut dipicu oleh pikloram, yang mampu mengubah polaritas sel dengan cara
menginterfensi melalui gradien pH atau medan listrik di sekitar sel. Selain itu,
ekspansi sel berasosiasi dengan polisakarida dinding sel dan bersesuaian dengan
ensim hidrolitik.
Tahapan embriogenesis somatik yang didahului dengan pembentukan pro-
embrio dapat diamati secara mikroskopis. Gambar 22 menunjukkan bahwa sel-sel
embriogenik membelah secara periklinal selama proses diferensiasi awal (1-4 sel).
Hal ini sekaligus memperlihatkan bahwa embriogenesis somatik pada tanaman
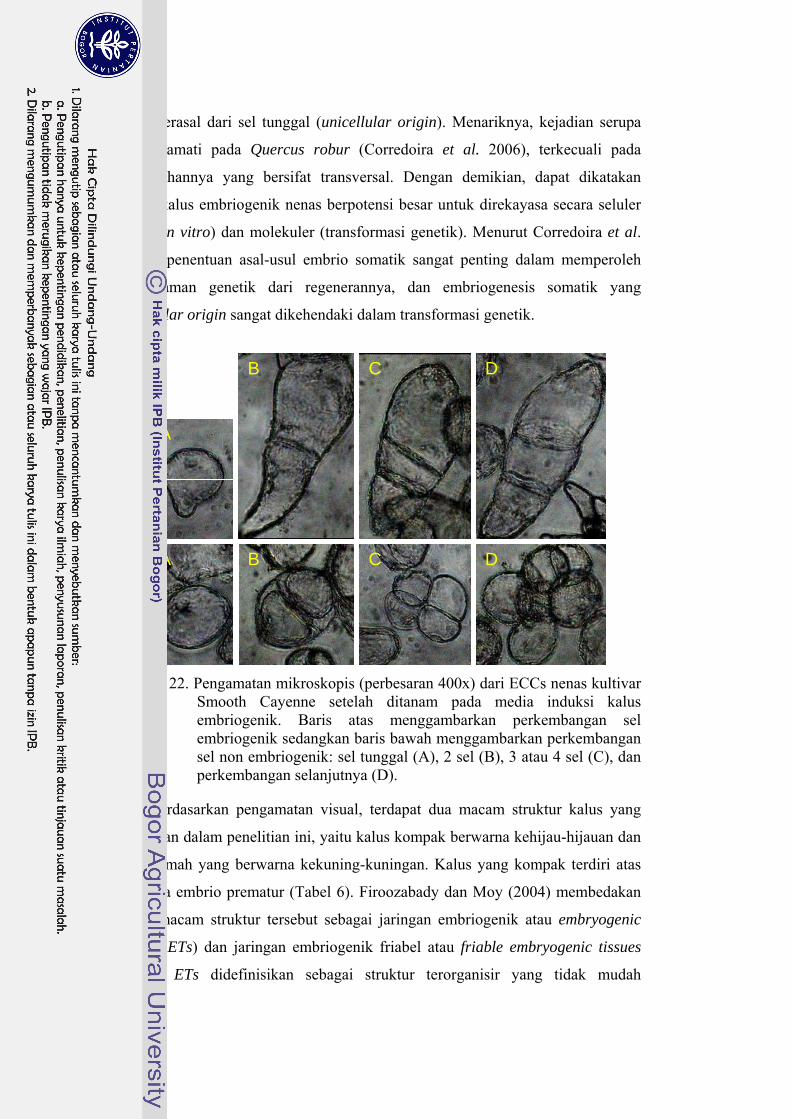
nenas berasal dari sel tunggal (unicellular origin). Menariknya, kejadian serupa
juga teramati pada Quercus robur (Corredoira et al. 2006), terkecuali pada
pembelahannya yang bersifat transversal. Dengan demikian, dapat dikatakan
bahwa kalus embriogenik nenas berpotensi besar untuk direkayasa secara seluler
(kultur in vitro) dan molekuler (transformasi genetik). Menurut Corredoira et al.
(2006), penentuan asal-usul embrio somatik sangat penting dalam memperoleh
keseragaman genetik dari regenerannya, dan embriogenesis somatik yang
unicellular origin sangat dikehendaki dalam transformasi genetik.
B C D
A
A B C D
Gambar 22. Pengamatan mikroskopis (perbesaran 400x) dari ECCs nenas kultivar
Smooth Cayenne setelah ditanam pada media induksi kalus embriogenik. Baris atas menggambarkan perkembangan sel embriogenik sedangkan baris bawah menggambarkan perkembangan sel non embriogenik: sel tunggal (A), 2 sel (B), 3 atau 4 sel (C), dan perkembangan selanjutnya (D).
Berdasarkan pengamatan visual, terdapat dua macam struktur kalus yang
dihasilkan dalam penelitian ini, yaitu kalus kompak berwarna kehijau-hijauan dan
kalus remah yang berwarna kekuning-kuningan. Kalus yang kompak terdiri atas
beberapa embrio prematur (Tabel 6). Firoozabady dan Moy (2004) membedakan
kedua macam struktur tersebut sebagai jaringan embriogenik atau embryogenic
tissues (ETs) dan jaringan embriogenik friabel atau friable embryogenic tissues
(FETs). ETs didefinisikan sebagai struktur terorganisir yang tidak mudah
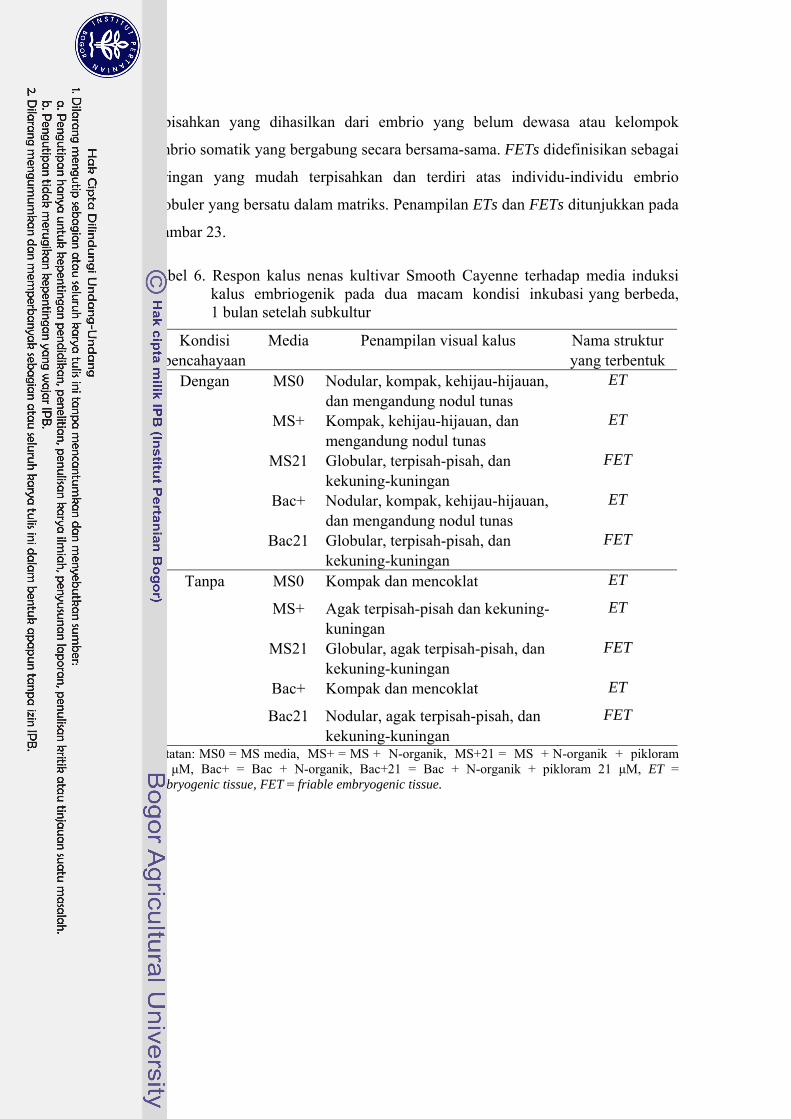
dipisahkan yang dihasilkan dari embrio yang belum dewasa atau kelompok
embrio somatik yang bergabung secara bersama-sama. FETs didefinisikan sebagai
jaringan yang mudah terpisahkan dan terdiri atas individu-individu embrio
globuler yang bersatu dalam matriks. Penampilan ETs dan FETs ditunjukkan pada
Gambar 23.
Tabel 6. Respon kalus nenas kultivar Smooth Cayenne terhadap media induksi kalus embriogenik pada dua macam kondisi inkubasi yang berbeda, 1 bulan setelah subkultur
Kondisi pencahayaan
Media Penampilan visual kalus Nama struktur yang terbentuk
Dengan MS0 Nodular, kompak, kehijau-hijauan, dan mengandung nodul tunas
ET
MS+ Kompak, kehijau-hijauan, dan mengandung nodul tunas
ET
MS21 Globular, terpisah-pisah, dan kekuning-kuningan
FET
Bac+ Nodular, kompak, kehijau-hijauan, dan mengandung nodul tunas
ET
Bac21 Globular, terpisah-pisah, dan kekuning-kuningan
FET
Tanpa MS0 Kompak dan mencoklat ET
MS+ Agak terpisah-pisah dan kekuning-kuningan
ET
MS21 Globular, agak terpisah-pisah, dan kekuning-kuningan
FET
Bac+ Kompak dan mencoklat ET
Bac21 Nodular, agak terpisah-pisah, dan kekuning-kuningan
FET
Catatan: MS0 = MS media, MS+ = MS + N-organik, MS+21 = MS + N-organik + pikloram 21 µM, Bac+ = Bac + N-organik, Bac+21 = Bac + N-organik + pikloram 21 µM, ET = embryogenic tissue, FET = friable embryogenic tissue.
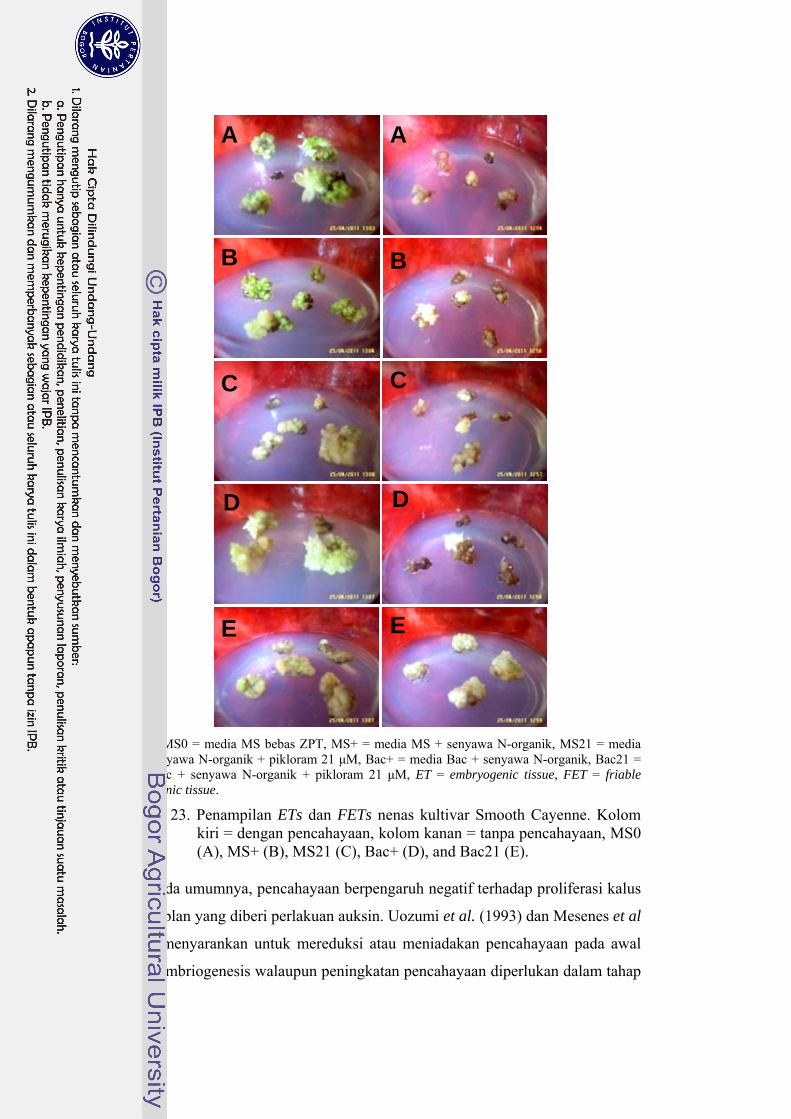
A
B
A
B
C
D
C
D
E E
Catatan: MS0 = media MS bebas ZPT, MS+ = media MS + senyawa N-organik, MS21 = media MS + senyawa N-organik + pikloram 21 µM, Bac+ = media Bac + senyawa N-organik, Bac21 = media Bac + senyawa N-organik + pikloram 21 µM, ET = embryogenic tissue, FET = friable embryogenic tissue.
Gambar 23. Penampilan ETs dan FETs nenas kultivar Smooth Cayenne. Kolom kiri = dengan pencahayaan, kolom kanan = tanpa pencahayaan, MS0 (A), MS+ (B), MS21 (C), Bac+ (D), and Bac21 (E).
Pada umumnya, pencahayaan berpengaruh negatif terhadap proliferasi kalus
dari eksplan yang diberi perlakuan auksin. Uozumi et al. (1993) dan Mesenes et al
(2005) menyarankan untuk mereduksi atau meniadakan pencahayaan pada awal
proses embriogenesis walaupun peningkatan pencahayaan diperlukan dalam tahap

perkembangan akhir embrio. Hasil serupa juga disarankan oleh Gatica et al
(2008), di mana kondisi tanpa pencahayaan lebih sesuai untuk produksi embrioid
kopi. Namun demikian, penelitian ini menunjukkan fenomena yang berbeda, di
mana kondisi tanpa pencahayaan justru menyebabkan penghambatan
pertumbuhan kalus dan menyebabkan nekrosis pada semua perlakuan media
(Gambar 23) walaupun tidak berbeda nyata secara statistik. Hal ini diduga
disebabkan oleh adanya akumulasi pikloram dalam kalus yang diinkubasikan pada
kondisi tanpa pencahayaan. Karena pikloram merupakan agensia herbisida maka
akumulasi senyawa tersebut akan bersifat fitotoksik terhadap biakan dan
menyebabkan pertumbuhan biakan terhambat. Dilaporkan bahwa pikloram
bersifat stabil pada kondisi tanpa pencahayaan dan mengalami degradasi pada
kondisi dengan pencahayaan (Hagen et al. 1991). Arteca (1996) juga melaporkan
bahwa pikloram bersifat persisten di dalam sel ketika terakumulasi dan tidak dapat
dimetabolismekan oleh sel tersebut. Fenomena pencoklatan yang disertai dengan
nekrosis dilaporkan disebabkan oleh oksidasi fenol setelah disintegrasi selular
yang berakibat pada kematian. Proses tersebut dikatalisis secara non enzimatis dan
enzimatis oleh fenol oksidase atau peroksidase yang menghasilkan senayawa
quinon yang berwarna coklat tanpa pencahayaan. Peroksidase tersebut berasosiasi
dengan degradasi klorofil pada jaringan yang senesens (Laukkanen et al. 2000).
Berdasarkan analisis statistik, tidak terdapat interaksi yang nyata antara
formulasi media dengan kondisi inkubasi untuk persentase pencoklatan,
persentase pembentukan nodul, dan persentase pembentukan embrio prematur.
Respon tersebut lebih dominan dipengaruhi oleh formulasi media induksi kalus
embriogenik. Persentase pencoklatan yang terendah, persentase nodul yang
tertinggi, persentase embrio prematur yang terendah, dan jumlah embrio yang
terendah dihasilkan dari kalus yang diberi perlakuan pikloram 21 µM. Hal ini
mungkin disebabkan oleh tingginya proliferasi sel daripada diferensiasi sel pada
kalus yang diberi perlakuan pikloram. Menurut Arteca (1996), pikloram
merupakan senyawa yang siap diserap oleh semua bagian tanaman dan siap
ditransportasikan ke seluruh area pertumbuhan. Penggunaan pikloram telah
dilaporkan Sudarmonowati dan Henshaw (1996) untuk menginduksi
embriogenesis somatik beberapa kultivar singkong yang gagal diinduksi dengan
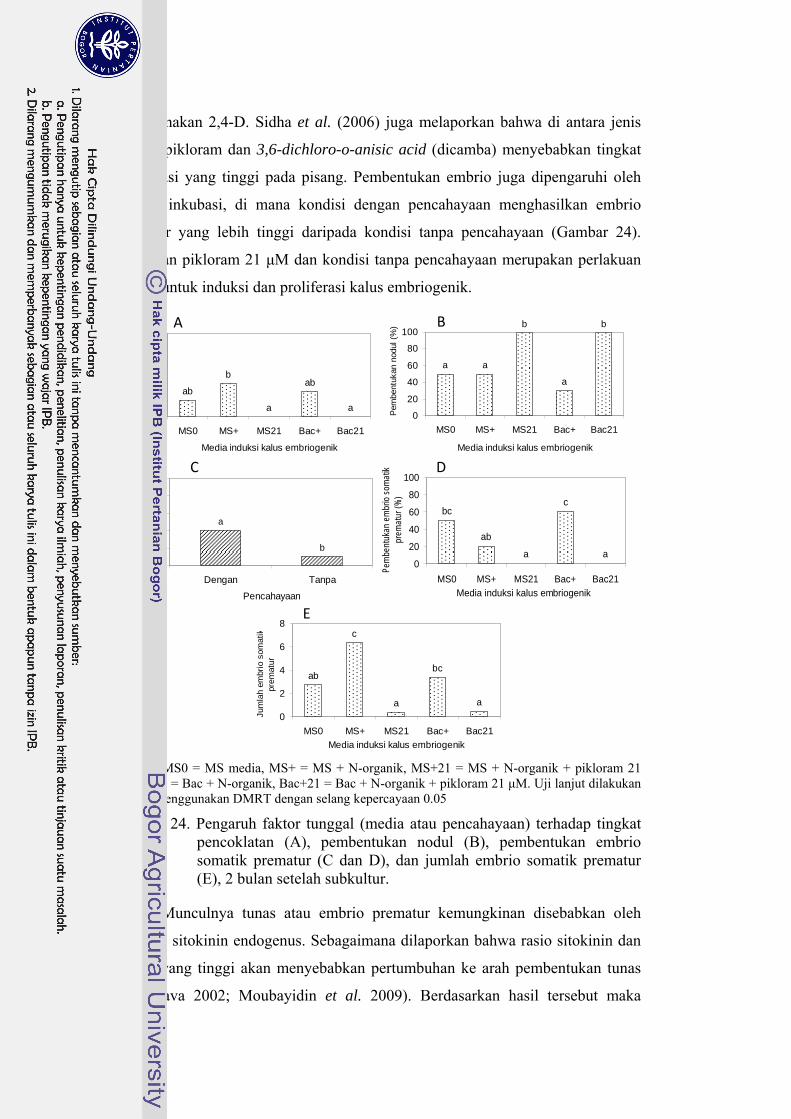
menggunakan 2,4-D. Sidha et al. (2006) juga melaporkan bahwa di antara jenis
auksin, pikloram dan 3,6-dichloro-o-anisic acid (dicamba) menyebabkan tingkat
proliferasi yang tinggi pada pisang. Pembentukan embrio juga dipengaruhi oleh
kondisi inkubasi, di mana kondisi dengan pencahayaan menghasilkan embrio
prematur yang lebih tinggi daripada kondisi tanpa pencahayaan (Gambar 24).
Perlakuan pikloram 21 µM dan kondisi tanpa pencahayaan merupakan perlakuan
terbaik untuk induksi dan proliferasi kalus embriogenik.
ab
b
a
ab
a0
20
40
60
80
100
MS0 MS+ MS21 Bac+ Bac21
Media induksi kalus embriogenik
Ting
kat p
enco
klata
n (%
)
a a
b
a
b
0
20
40
60
80
100
MS0 MS+ MS21 Bac+ Bac21
Media induksi kalus embriogenik
Pem
bent
ukan
nod
ul (%
)
bc
ab
a
c
a0
20
40
60
80
100
MS0 MS+ MS21 Bac+ Bac21Media induksi kalus embriogenik
Pem
bent
ukan
em
brio
som
atik
prem
atur
(%)
a
b
0
20
40
60
80
100
Dengan Tanpa
Pencahayaan
Pemb
entuk
an em
brio s
omati
k prem
atur
(%)
ab
c
a
bc
a0
2
4
6
8
MS0 MS+ MS21 Bac+ Bac21Media induksi kalus embriogenik
Jum
lah
embr
io s
omat
ikpr
emat
ur
A B
C D
E
Catatan: MS0 = MS media, MS+ = MS + N-organik, MS+21 = MS + N-organik + pikloram 21 µM, Bac+ = Bac + N-organik, Bac+21 = Bac + N-organik + pikloram 21 µM. Uji lanjut dilakukan dengan menggunakan DMRT dengan selang kepercayaan 0.05 Gambar 24. Pengaruh faktor tunggal (media atau pencahayaan) terhadap tingkat
pencoklatan (A), pembentukan nodul (B), pembentukan embrio somatik prematur (C dan D), dan jumlah embrio somatik prematur (E), 2 bulan setelah subkultur.
Munculnya tunas atau embrio prematur kemungkinan disebabkan oleh
aktivitas sitokinin endogenus. Sebagaimana dilaporkan bahwa rasio sitokinin dan
auksin yang tinggi akan menyebabkan pertumbuhan ke arah pembentukan tunas
(Srivastava 2002; Moubayidin et al. 2009). Berdasarkan hasil tersebut maka

struktur ETs dan FETs dipindahkan ke media yang berbeda. ETs dipindahkan ke
media MS dengan penambahan Kn 1 mg l-1 untuk regenerasi dan elongasi embrio
premature sedangkan FETs dipindahkan ke media Reg A atau Reg B untuk
perkembangan embrio.
Perkembangan Embrio Somatik
Regenerasi ETs (Embryogenic Tissues)
Hasil percobaan menunjukkan bahwa daya renegenerasi ETs tergolong
tinggi, yaitu sebesar 77.8-100% (Tabel 7). Performan ETs sebelum dan setelah
regenerasi ditampilkan pada Gambar 25. Pada gambar tersebut tampak bahwa
biakan kalus yang ditumbuhkan pada media Bac yang diperkaya dengan senyawa
N-organik mempunyai struktur yang mudah terpisahkan. Terdapat interaksi yang
nyata antara media induksi kalus embriogenik dengan kondisi inkubasi untuk
peubah jumlah total embrio umur 1 bulan dan jumlah embrio ukuran besar umur 4
bulan (Gambar 26). Perlakuan terbaik adalah media Bac dengan penambahan
senyawa N-organik pada kondisi dengan pencahayaan. Perlakuan tersebut
menghasilkan sekitar 17 embrio/eksplan dalam waktu 1 bulan dan lebih dari 30
embrio/eksplan dalam waktu 4 bulan. Hasil tersebut lebih baik daripada hasil
penelitian sebelumnya (Sripaoraya et al. 2003) yang dilakukan pada nenas
kultivar Phuket (65.7% dengan 3 embrio/eksplan) dan kultivar Champaka yang
dilaporkan oleh Firoozabady dan Moy (2004) yang menghasilkan 63% regenerasi.
Tabel 7. Perkembangan ETs nenas kultivar Smooth Cayenne pada media MS yang mengandung Kn 1 mg l-1, 2 bulan setelah subkultur
Kondisi inkubasi pada tahap pengkalusan
Media induksi kalus embriogenik
Pembentukan embrio somatik dewasa (%)
Dengan pencahayaan MS0 87.5
MS+ 100
Bac+ 100
Tanpa pencahayaan MS0 77.8
MS+ 100
Bac+ 100 Catatan: MS0 = MS media, MS+ = MS + senyawa N-organik, Bac+ = Bac + senyawa N-organik

A B C
A B C
Terang
Gelap
Gambar 25. Regenerasi ETs nenas kultivar Smooth Cayenne pada media MS
dengan penambahan Kn 1 mg l-1: media MS tanpa ZPT (kiri), media MS yang diperkaya dengan senyawa N-organik (tengah), dan media Bac yang diperkaya dengan senyawa N-organik (kanan).
Umur (1 bulan)a
b
0
4
8
12
16
20
Dengan TanpaPencahayaan
Jum
lah
embr
io s
omat
ikuk
uran
bes
ar
Umur (1 bulan)
a
b
b
0
5
10
15
20
MS0 MS+ Bac+
Media induksi kalus embriogenik
Jum
lah
embr
io s
omat
ikuk
uran
bes
ar
Umur (1 bulan)
abb
a
0
4
8
12
MS0 MS+ Bac+
Media induksi kalus embriogenik
Jum
lah
embr
io
som
atik
ukr
an k
eci
Umur (4 bulan)
0
10
20
30
40
Dengan Tanpa Dengan Tanpa Dengan Tanpa
MS+ MS0 Bac+Pencahayaan dan media induksi kalus embriogenik
Juml
ah em
brio s
omati
k uk
uran b
esar
Umur (1 bulan)
048
121620
Dengan Tanpa Dengan Tanpa Dengan Tanpa
MS+ MS0 Bac+Pencahayaan dan media induksi kalus embriogenik
Jum
lah
tota
l em
brio
som
atik
A B
C D
E
Catatan: MS0 = MS media, MS+ = MS + N-organik, Bac+ = Bac + N-organik. Uji lanjut dilakukan dengan menggunakan DMRT dengan selang kepercayaan 0.05 Gambar 26. Pengaruh faktor pencahayaan (A) dan media induksi kalus
embriogenik (B dan C) serta pengaruh interaksi antara media induksi kalus embriogenik dengan pencahayaan (D dan E) terhadap pembentukan embrio somatik nenas kultivar Smooth Cayenne umur 1 dan 4 bulan.

Secara visual, struktur globular dan koleoptilar dapat diamati secara
langsung pada biakan, namun struktur skutelar tidak dapat terdeteksi secara visual
karena embrio somatik prematur bergabung dalam bentuk ETs atau embrio
somatik bersatu sebagai matriks dalam bentuk FETs. Menariknya, akar terbentuk
secara spontan tanpa induksi dan strukturnya sangat spesifik (Gambar 27)
sebagaimana yang terbentuk pada embrio zigotik. Kordestani dan Karami (2008)
melaporkan hal serupa, di mana tumbuhnya akar dan primordia tunas yang
sempurna menyebabkan embrio somatik stroberi dapat berkecambah dengan
mudah membentuk planlet tanpa tambahan tahapan perakaran. Jimenez (2005)
menjelaskan bahwa perkembangan embrio tanaman monokotil melalui beberapa
tahapan, yaitu globular, skutelar, dan koleoptilar.
A B C D
Gambar 27. Tahapan perkembangan embrio somatik nenas kultivar Smooth
Cayenne dari struktur FETs yang diinduksi oleh pikloram: globular (A), koleoptilar akhir (B), embrio somatik dewasa (C), dan planlet yang siap diaklimatisasi (D).
Struktur skutelar (Gambar 28C) baru dapat diamati melalui analisis histologi
terhadap embrio somatik yang dihasilkan dari proses suspensi sel. Percobaan
suspensi sel menunjukkan bahwa kombinasi perlakuan tanpa pencahayaan dan
media Bac yang diperkaya dengan senyawa N-organik dan pikloram 21 µM
merupakan perlakuan yang terbaik karena menghasilkan suspensi dengan
persentase yang lebih tinggi daripada perlakuan lainnya, juga mampu menekan
pencoklatan hingga 100% (Tabel 8).
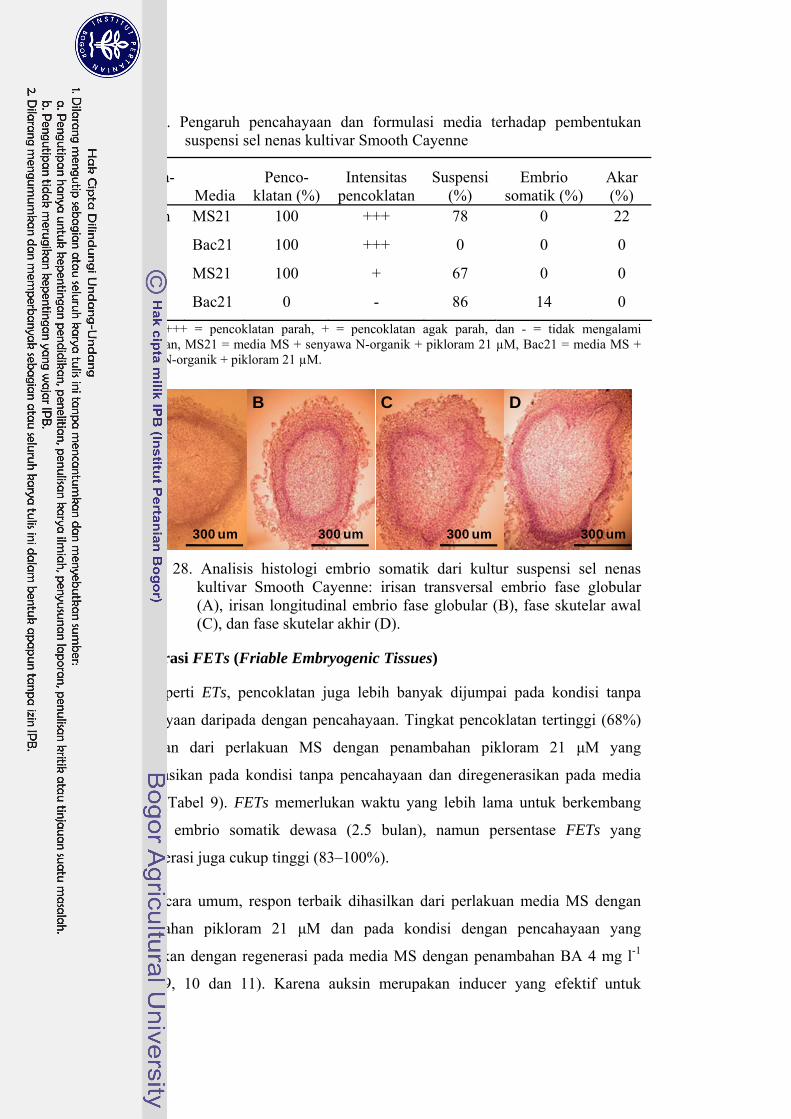
Tabel 8. Pengaruh pencahayaan dan formulasi media terhadap pembentukan suspensi sel nenas kultivar Smooth Cayenne
Pencaha-yaan Media
Penco-klatan (%)
Intensitas pencoklatan
Suspensi (%)
Embrio somatik (%)
Akar (%)
Dengan MS21 100 +++ 78 0 22
Bac21 100 +++ 0 0 0
Tanpa MS21 100 + 67 0 0
Bac21 0 - 86 14 0
Catatan: +++ = pencoklatan parah, + = pencoklatan agak parah, dan - = tidak mengalami pencoklatan, MS21 = media MS + senyawa N-organik + pikloram 21 µM, Bac21 = media MS + senyawa N-organik + pikloram 21 µM.
A B C D
300 um 300 um300 um 300 um
Gambar 28. Analisis histologi embrio somatik dari kultur suspensi sel nenas kultivar Smooth Cayenne: irisan transversal embrio fase globular (A), irisan longitudinal embrio fase globular (B), fase skutelar awal (C), dan fase skutelar akhir (D).
Regenerasi FETs (Friable Embryogenic Tissues)
Seperti ETs, pencoklatan juga lebih banyak dijumpai pada kondisi tanpa
pencahayaan daripada dengan pencahayaan. Tingkat pencoklatan tertinggi (68%)
dihasilkan dari perlakuan MS dengan penambahan pikloram 21 µM yang
diinkubasikan pada kondisi tanpa pencahayaan dan diregenerasikan pada media
Reg B (Tabel 9). FETs memerlukan waktu yang lebih lama untuk berkembang
menjadi embrio somatik dewasa (2.5 bulan), namun persentase FETs yang
beregenerasi juga cukup tinggi (83–100%).
Secara umum, respon terbaik dihasilkan dari perlakuan media MS dengan
penambahan pikloram 21 µM dan pada kondisi dengan pencahayaan yang
dilanjutkan dengan regenerasi pada media MS dengan penambahan BA 4 mg l-1
(Tabel 9, 10 dan 11). Karena auksin merupakan inducer yang efektif untuk
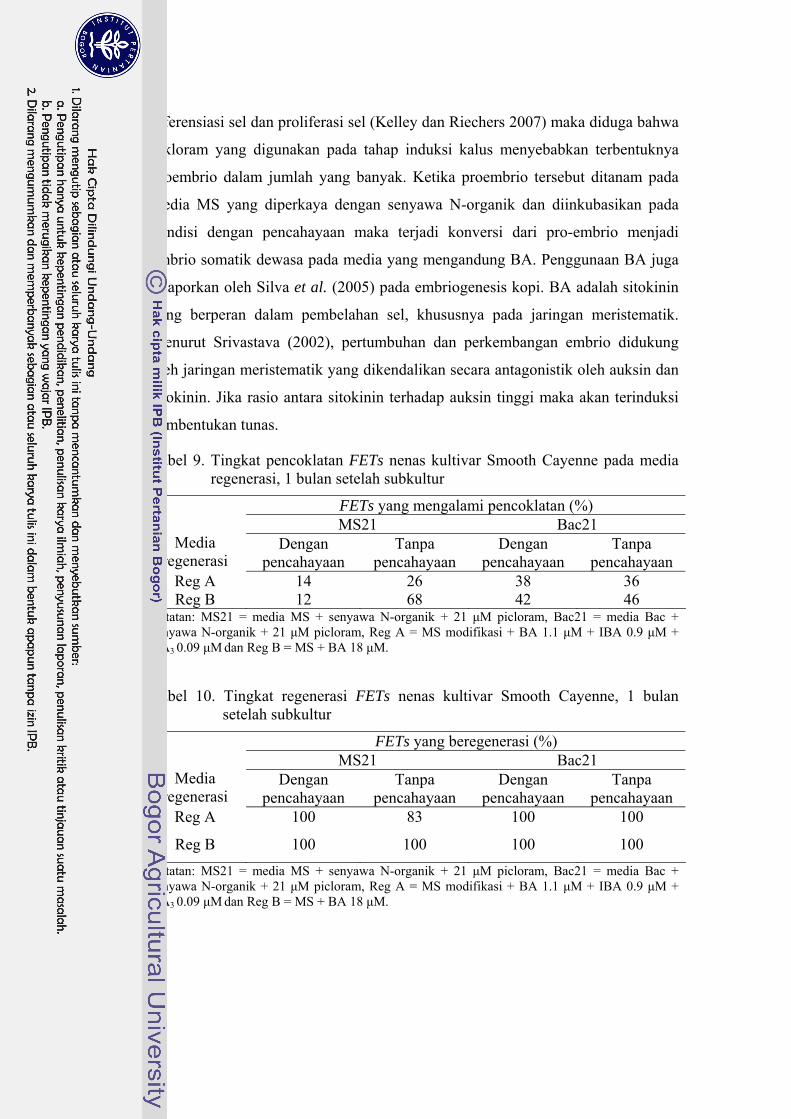
diferensiasi sel dan proliferasi sel (Kelley dan Riechers 2007) maka diduga bahwa
pikloram yang digunakan pada tahap induksi kalus menyebabkan terbentuknya
proembrio dalam jumlah yang banyak. Ketika proembrio tersebut ditanam pada
media MS yang diperkaya dengan senyawa N-organik dan diinkubasikan pada
kondisi dengan pencahayaan maka terjadi konversi dari pro-embrio menjadi
embrio somatik dewasa pada media yang mengandung BA. Penggunaan BA juga
dilaporkan oleh Silva et al. (2005) pada embriogenesis kopi. BA adalah sitokinin
yang berperan dalam pembelahan sel, khususnya pada jaringan meristematik.
Menurut Srivastava (2002), pertumbuhan dan perkembangan embrio didukung
oleh jaringan meristematik yang dikendalikan secara antagonistik oleh auksin dan
sitokinin. Jika rasio antara sitokinin terhadap auksin tinggi maka akan terinduksi
pembentukan tunas.
Tabel 9. Tingkat pencoklatan FETs nenas kultivar Smooth Cayenne pada media regenerasi, 1 bulan setelah subkultur
FETs yang mengalami pencoklatan (%) MS21 Bac21
Media regenerasi
Dengan pencahayaan
Tanpa pencahayaan
Dengan pencahayaan
Tanpa pencahayaan
Reg A 14 26 38 36 Reg B 12 68 42 46
Catatan: MS21 = media MS + senyawa N-organik + 21 µM picloram, Bac21 = media Bac + senyawa N-organik + 21 µM picloram, Reg A = MS modifikasi + BA 1.1 µM + IBA 0.9 µM + GA3 0.09 µM dan Reg B = MS + BA 18 µM.
Tabel 10. Tingkat regenerasi FETs nenas kultivar Smooth Cayenne, 1 bulan setelah subkultur
FETs yang beregenerasi (%) MS21 Bac21
Media regenerasi
Dengan pencahayaan
Tanpa pencahayaan
Dengan pencahayaan
Tanpa pencahayaan
Reg A 100 83 100 100
Reg B 100 100 100 100 Catatan: MS21 = media MS + senyawa N-organik + 21 µM picloram, Bac21 = media Bac + senyawa N-organik + 21 µM picloram, Reg A = MS modifikasi + BA 1.1 µM + IBA 0.9 µM + GA3 0.09 µM dan Reg B = MS + BA 18 µM.

Tabel 11. Jumlah embrio somatik dewasa yang terbentuk dari FETs nenas kultivar Smooth Cayenne, 1 bulan setelah subkultur
Jumlah embrio somatik dewasa MS21 Bac21
Media regenerasi
Dengan pencahayaan
Tanpa pencahayaan
Dengan pencahayaan
Tanpa pencahayaan
Reg A 12.7±6.8 10.5±6.0 2.5±1.3 11.4±5.5
Reg B 14.8±6.0 8.8±3.9 9.2±3.3 4.8±2.6 Catatan: MS21 = media MS + senyawa N-organik + 21 µM picloram, Bac21 = media Bac + senyawa N-organik + 21 µM picloram, Reg A = MS modifikasi + BA 1.1 µM + IBA 0.9 µM + GA3 0.09 µM dan Reg B = MS + BA 18 µM.
Simpulan
Tahapan embriogenesis somatik tanaman nenas kultivar Smooth Cayenne
dapat diamati secara mikroskopis dan visual. Embriogenesis somatik tersebut
merupakan unicellular origin atau berasal dari satu sel. Tahapan embriogenesis
somatik yang teramati adalah polarisasi sel, pembelahan sel secara asimetris,
pembentukan ETs dan FETs, perkembangan embrio (globular, skutelar, dan
koleoptilar). Perlakuan terbaik untuk induksi kalus adalah media MS dengan
penambahan pikloram 21 µM. Perlakuan terbaik untuk induksi dan proliferasi
kalus embriogenik adalah kombinasi perlakuan media MS dengan penambahan
pikloram 21 µM dan kondisi tanpa pencahayaan. Media Bac dengan penambahan
senyawa N-organik dan pikloram 21 µM pada kondisi tanpa pencahayaan
merupakan perlakuan terbaik untuk pembentukan suspensi sel nenas (86%).
Perlakuan media Bac yang diperkaya dengan senyawa N-organik dan inkubasi
pada kondisi dengan pencahayaan merupakan perlakuan terbaik untuk regenerasi
ETs (17 embrio/eksplan dalam 2 bulan dan lebih dari 30 embrio/eksplan dalam 4
bulan). Kombinasi perlakuan media MS dengan penambahan pikloram 21 µM dan
kondisi dengan pencahayaan serta media regenerasi Reg B (MS + BA 18 µM)
merupakan perlakuan terbaik untuk regenerasi FETs (14 embrio/eksplan dalam
2.5 bulan).

Daftar Pustaka
Abrash, E.B. and D.C. Bergmann. 2009. Asymmetric cell divisions: A view from plant development. Developmental Cell 16: 783-796.
Arnold S, Sabala I, Bozhkov P, Dyachok J, Filonova L. 2002. Developmental pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult 69: 233-249.
Arteca RN. 1996. Plant Growth Substances. Chapman and Hall. New York.332p.
Buffard-Morel JL et al. 1995. Initiation of somatic embryogenesis in coconut (Cocos nucifera L.). Pp. 217-223. In: Oropeza C et al., editor. Lethal Yellowing: Research and Practical Aspects. Kluwer Academic Publishers, Netherlands.
Corredoira E, Valladares S, Vieitez AM. Morphohistological analysis of the origin and development of somatic embryos from leaves of mature Quercus robur. In Vitro Cell Dev Biol_Plant 42:525-533.
Firoozabady E, Moy Y. 2004. Regeneration of pineapple via somatic embryogenesis and organogenesis. In vitro Cell Dev Biol_Plant 40:67-74.
Gatica AM, Arrieta G, Espinoza AM. 2008. Direct somatic embryogenesis in Coffea arabica L.. cvs. Caturra and Catuaí: effect of triacontanol, light condition, and medium consistency. Agronomía Costarricense 32(1): 139-147.
Hagen SR, Muneta P, Augustin J, Tourneau D Le. 1991. Stability and utilization of picloram, vitamins, and sucrose in a tissue culture medium. Plant Cell Tissue and Organ Cult 25(1):45-48.DOI: 10.1007/BF00033911.(Abstract)
Jimenez VM. 2005. Involvement of plant hormone and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regulation 47: 91-110.
Kelley B, Riechers DE. 2007. Recent development in auxin biology and new opportunities for auxinic herbicide research. Pesticide Biochemistry and Physiology 89: 1-11.
Kiernan JA. 1990. Histological and Histochemical Methods: Theory and Practice. 2nd edition. Pergamon Press.
Kordestani GK, Karami O. 2008. Picloram-induced somatic embryogenesis in leaves of strawberry (Fragaria ananassa L.). Acta Biologica Cracoviensia Series Botanica 50(1): 69–72.

Laukkanen, H., L. Rautiainen, E. Taulavuori, and A. Hohtola. 2000. Changes in cellular structures and enzymatic activities during browning of Scots pine callus derived from mature buds. Tree Physiology 20: 467–475.
Lincy AK, Remashree AB, Sasikumar B. 2009. Indirect and direct somatic embryogenesis from aerial stem explants of ginger (Zingiber officinale Rosc.). Acta Bot Croat 68 (1): 93–103.
Meneses A, Flores D, Muñoz M, Arriet G, Espinoza AM. 2005. Effect of 2,4-D, hydric stress and light on indica rice (Oryza sativa) somatic embryogenesis. Int J Trop Biol 53 (3-4): 361-368.
Moubayidin L, Mambro RD, Sabatini S. 2009. Cytokinin–auxin crosstalk. Trends in Plant Science 14 (10): 557-562.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15: 473-497.
Nursandi F, Poerwanto R, Sobir, Sujiprihati S. 2003. Perbanyakan in vitro nenas (Ananas comosus (L.) Merr.) dengan menggunakan BAP dan TDZ. Dalam: Simposium Nasional dan Konggres VIII. Bandar Lampung.
Oktavia F, Siswanto, Budiani A, Sudarsono. 2003. Direct somatic embryogenesis and regeneration of arabica coffee plantlets (Coffea arabica) from different explant. Menara Perkebunan 71(2): 44-55.
Paciorek T, Bergmann DC. 2010. The secret to life is being different: asymmetric divisions in plant development. Current Opinion in Plant Biology 13:661–669.
Perez G, Yanes E, Isidron M, Lorenzo JC. 2009. Phenotypic and AFLP characterization of two new pineapple somaclones derived from in vitro culture. Plant Cell Tissue Organ Cult 96:113-116.
Purnamaningsih R, Mariska I, Supriati Y. 2009. Penggunaan ABA dan paklobutrazol dalam perbanyakan nenas Simadu melalui kultur in vitro. Ber Bio 9(6): 751-758.
Sidha M, Suprasanna P, Bapat VA, Kulkarni UG, Shinde BN. 2007. Developing somatic embryogenic culture system and plant regeneration in banana. Founder’s Day 285: 153-161.
Sripaoraya S, Marchamt R, Power JB, Davey MR. 2003. Plant regeneration by somatic embryogenesis and organogenesis in commercial pineapple (Ananas comosus L.). In Vitro Cell Dev Biol_Plant 39:450-454.
Srivastava LM. 2002. Plant Growth and Development. Academic Press. New York. 772p.

Sudarmonowati E, Henshaw GG. 1996. The use of picloram and dicamba to induce somatic embryogenesis in cassava. Annales Bogoriensis 4 (1):27-34.
Suminar E. 2010. Induksi keragaman genetik dengan mutagen sinar gamma pada nenas (Ananas comosus (L.) Merr.) secara in vitro [seminar]. Sekolah Pascasarjana, Institut Pertanian Bogor. Bogor.
Suryowinoto M. 1996. Pemuliaan Tanaman secara In Vitro. Pusat Antar Universitas-Bioteknologi UNiversitas Gadjah Mada. Yogyakarta. 252 hal.
Uozumi N, Kohketsu K, Okamoto A, Kobayashi T. 1993. Light dependency in celery somatic embryogenesis and plantlet development in suspension culture. Plant Tissue Culture Letter 10(1): 25-32.
Vanneste S, Frim J. 2009. Auxin: A trigger for change in plant development. Cell 136: 1005-1016.

PEMBENTUKAN BENIH SINTETIK NENAS DAN KONSERVASI IN VITRO SECARA PERTUMBUHAN
MINIMAL
Abstrak
Benih sintetik merupakan alat perbanyakan secara vegetatif dan sekaligus sebagai alat penyimpanan. Melalui teknik pertumbuhan minimal, biakan dapat disimpan dalam waktu yang relatif lama. Tujuan penelitian ini adalah untuk mengetahui pengaruh kombinasi auksin dan sitokinin terhadap morfogenesis eksplan nenas yang terenkapsulasi, mengetahui pengaruh interaksi antara suhu penyimpanan dengan taraf paklobutrazol atau manitol terhadap pertumbuhan tunas nenas yang terenkapsulasi. Penelitian terbagi atas 3 percobaan, yaitu enkapsulasi eksplan, pertumbuhan minimal dengan menggunakan paklobutrazol dan pertumbuhan minimal dengan menggunakan manitol. Enkapsulasi dilakukan dengan menggunakan Na-alginat 3% yang berisi media MS dengan penambahan BA (0, 1, 2, dan 3 mg l-1) yang dikombinasikan dengan NAA (0, 1, 2, dan 3 mg l-
1). Pertumbuhan minimal dengan paklobutrazol dilakukan pada taraf 0, 1, 2, dan 3 mg l-1 atau manitol pada taraf 0, 1, 2, 3, 4, dan 5% dengan suhu penyimpanan 15 dan 25 0C. Hasil penelitian menunjukkan bahwa basal daun nenas tidak mampu bertahan hidup dalam kondisi terenkapsulasi dengan Na-alginat 3%. Tidak terdapat interaksi yang nyata antara taraf paklobutrazol dengan suhu penyimpanan terhadap daya hidup dan daya tembus kapsul tunas nenas. Biakan tersebut hanya dapat disimpan selama 3 bulan. Interaksi yang nyata juga tidak dijumpai antara taraf manitol dan suhu penyimpanan terhadap daya hidup dan daya tembus kapsul embrio somatik nenas. Dengan menggunakan manitol 4% maka penyimpanan dapat diperpanjang hingga 4 bulan. Manitol dapat menggantikan suhu rendah sehingga biaya konservasi akan dapat diturunkan.
Kata kunci: Benih sintetik, pertumbuhan minimal, paklobutrazol, manitol, Ananas comosus (L.) Merr.
Abstract
Artificial seeds can be used as propagules for vegetative propagation and as storage materials. Through minimal growth, the artificial seeds can be stored for long time. The objectives of the study were to know the effect of combination treatment between auxin and cytokinin to the morphogenesis of encapsulated pineapple cultures, to know the effect of paclobutrazol, manitol, and temperature of storage to the growth of encapsulated pineapple cultures. The study was divided into 3 steps which were encapsulation, minimal growth by using paclobutrazol and reduced temperature, and minimal growth by using manitol and reduced temperature. Encapsulation was conducted by using 3% Na-alginat containing of MS medium with addition of BA (0, 1, 2, and 3 mg l-1) combined with NAA (0, 1, 2, and 3 mg l-1). Paklobutrazol was used at the rate of 0, 1, 2, and 3 mg l-1 in the temperature of 15 and 25 0C. Manitol was used at the rate of 0, 1, 2, 3, 4, and 5% in the temperature of 15 and 25 0C. The result showed that encapsulated leaf bases of pineapple could not survive. There was no interaction

between paclobutrazol and temperature to the survival rate and emergence rate of the encapsulated cultures. The encapsulated shoots could be stored for 3 months. There was also no interaction between mannitol and temperature to the survival rate and emergence rate of the encapsulated cultures. By using somatic embryos and 4% mannitol, the storage of pineapple cultures could be prolonged for 4 months. Mannitol could substitute the use of low temperature so that the cost of conservation can be minimized.
Keywords: Artificial seed, minimal growth, paclobutrazol, mannitol, Ananas comosus (L.) Merr.
Pendahuluan
Benih sintetik didefinisikan sebagai embrio somatik yang berada di dalam
mantel (kapsul) sehingga sifatnya mirip dengan benih zigotik (Redenbaugh 1992).
Mantel tersebut berperan sebagai endosperma yang mengandung sumber karbon,
nutrisi, zat pengatur tumbuh, dan agensia anti mikroba. Dewasa ini, definisi benih
sintetik dikembangkan lebih lanjut karena eksplan yang digunakan tidak terbatas
pada embrio somatik melainkan juga tunas terminal, tunas aksilar, nodus, dan
jaringan meristematik lainnya. Selain untuk tujuan perbanyakan bibit, teknologi
benih sintetik juga dapat dimanfaatkan untuk tujuan konservasi (Rai et al., 2009).
Teknik konservasi in vitro dapat dibedakan menjadi tiga macam: (1)
penyimpanan pada media tumbuh, (2) penyimpanan secara pertumbuhan minimal,
dan (3) penyimpanan secara kriopreservasi (Mariska et al. 1996; Leunufna 2004).
Teknik pertumbuhan minimal disarankan diterapkan untuk koleksi aktif (working
collection atau active collection), sedangkan teknik kriopreservasi diterapkan
untuk koleksi dasar (base collection) (Withers 1985).
Pada teknik penyimpanan dengan media tumbuh, tidak diperlukan
penambahan zat penghambat tumbuh. Penyimpanan dengan cara tersebut akan
memerlukan tindakan subkultur yang frekuentif sehingga kurang menghemat
tenaga, waktu, dan biaya serta beresiko terhadap kontaminasi (Mariska et al.
1996). Selain itu, subkultur yang frekuentif juga beresiko terhadap timbulnya
keragaman somaklonal (Eeuwens et al. 2002).

Pada teknik pertumbuhan minimal, beberapa modifikasi media dan
lingkungan dapat diterapkan, antara lain penurunan temperatur lingkungan dan
intensitas cahaya (Hu dan Wang 1983; Withers 1985; Keller et al. 2006),
penggunaan regulator osmotik seperti sukrosa dan manitol (Withers 1985;
Bessembinder et al. 1993), penurunan taraf beberapa faktor esensial seperti
pengenceran media (Desbrunais et al. 1992), serta penggunaan zat penghambat
tumbuh seperti paclobutrazol, cycocel, dan ancymidol (Withers 1985). Dengan
penerapan teknik pertumbuhan minimal maka biakan dapat disimpan dalam
jangka menengah (bulanan hingga tahunan).
Di Indonesia, teknik pertumbuhan minimal tanaman nenas belum pernah
dilaporkan. Di mancanegara, dilaporkan bahwa teknik enkapsulasi tunas in vitro
mampu menyimpan biakan nenas selama 1.5 bulan melalui aplikasi suhu 8 0C
dengan menggunakan media MS tanpa zat pengatur tumbuh (Gangopadhyay et al.
2004). Penyimpanan pada suhu sangat rendah memerlukan energi listrik yang
cukup besar biayanya sehingga diperlukan metode lain yang mampu menghambat
pertumbuhan biakan supaya dapat disimpan dalam waktu yang lebih lama tanpa
menurunkan daya regenerasinya pasca-penyimpanan.
Secara umum, penelitian ini ditujukan untuk pembentukan benih sintetik
dan penyimpanan eksplan nenas dengan metode pertumbuhan mininal. Secara
khusus, tujuan penelitian ini adalah: (1) mengetahui pengaruh kombinasi auksin
dan sitokinin terhadap morfogenesis eksplan nenas yang terenkapsulasi, (2)
mengetahui pengaruh taraf paklobutrazol dan suhu penyimpanan terhadap
pertumbuhan tunas nenas yang terenkapsulasi, (3) mengetahui pengaruh taraf
manitol dan suhu penyimpanan terhadap pertumbuhan embrio somatik nenas yang
terenkapsulasi.
Bahan dan Metode
Sumber bahan tanaman yang digunakan adalah tunas in vitro tanaman nenas
Smooth Cayenne yang berasal dari Subang, Jawa Barat. Tunas in vitro tersebut
dipelihara pada media MS dengan penambahan benzyl adenine (BA) 0,5 mg l-1
dan kinetin (Kn) 1 mg l-1. Inkubasi dilakukan di ruang kultur dengan suhu 25 0C

dan pencahayaan 800–1000 lux dengan fotoperiodisitas 16 jam. Penelitian dibagi
atas tiga tahap percobaan, yaitu (1) Enkapsulasi eksplan, (2) Pertumbuhan
minimal dengan menggunakan paklobutrazol, dan (2) Pertumbuhan minimal
dengan menggunakan manitol.
Enkapsulasi eksplan
Eksplan yang digunakan adalah basal daun dan batang semu (core). Eksplan
dienkapsulasi dengan natrium alginat 3% yang berisi media MS dengan
penambahan BA (0, 1, 2, dan 3 mg l-1) yang dikombinasikan dengan naphthalene
acetic acid (NAA) pada taraf 0, 1, 2, dan 3 mg l-1. Proses enkapsulasi dilakukan
dengan metode tetes ke dalam larutan CaCl2.2H2O 100 mM dan direndam selama
15 menit dengan penggojokan hingga membentuk gel atau kapsul. Setiap
perlakuan diulang sebanyak 2 kali (botol) dan setiap botol terdiri atas 10 kapsul.
Kapsul-kapsul tersebut direndam dalam akuades steril dengan volume 25 ml.
Inkubasi dilakukan pada suhu 25 0C, fotoperiodisitas 16 jam terang dengan
intensitas 800–1000 lux. Respon yang diamati adalah persentase biakan yang
hidup dan persentase biakan yang menembus kapsul.
Pertumbuhan minimal dengan menggunakan paklobutrazol
Eksplan yang digunakan adalah tunas in vitro yang berukuran kecil (tinggi
sekitar 2 cm). Beberapa daun tuanya dibuang dan disisakan daun mudanya sekitar
5 helai. Pucuk dan pangkal dipangkas sedemikian rupa sehingga diperoleh eksplan
dengan ukuran kurang dari 0.5 cm. Percobaan disusun secara faktorial dalam
lingkungan Rancangan Acak Lengkap. Faktor pertama adalah taraf paklobutrazol
(0, 1, 2, dan 3 mg l-1) sedangkan faktor kedua adalah suhu penyimpanan (15 dan
25 0C). Setiap perlakuan diulang sebanyak 4 kali (botol) dan setiap botol terdiri
atas 10 kapsul. Eksplan dienkapsulasi dengan natrium alginat 3% yang berisi
media MS dengan penambahan BA 1 mg l-1dan indole butyric acid (IBA) 0.5 mg
l-1 dengan penambahan 1-[4-chlorophenyl]-4,4-dimethyl-2-[1,2,4-triazol-1-yl]
pentan-3-ol (paklobutrazol). Proses enkapsulasi dilakukan dengan metode tetes ke
dalam larutan CaCl2.2H2O 100 mM dan direndam selama 15 menit dengan
penggojokan hingga membentuk kapsul. Inkubasi pada suhu 15 0C dilakukan di

dalam growth chamber dan inkubasi pada suhu 25 0C dilakukan di ruang kultur
dengan fotoperiodisitas 16 jam terang dengan intensitas 800–1000 lux. Respon
yang diamati adalah persentase daya hidup, persentase daya regenerasi, dan
persentase biakan yang menembus kapsul. Biakan yang masih bertahan hidup
kemudian dipindah ke media MS padat yang mengandung BA 0.5 mg l-1 dan Kn 1
mg l-1 untuk pemulihan dan regenerasi. Respon yang diamati adalah persentase
biakan yang hidup dan persentase biakan yang beregenerasi, jumlah tunas dan
jumlah daun.
Pertumbuhan minimal dengan menggunakan manitol
Eksplan yang digunakan adalah embrio somatik prematur. Induksi kalus
dilakukan dengan menggunakan 4-amino-3,5,6-trichloropicolinic acid (pikloram)
pada taraf 21 µM dengan penambahan thidiazuron (TDZ) 9 µM. Inkubasi
dilakukan pada suhu 25 0C dalam keadaan gelap selama 3 minggu. Setelah itu,
daun diisolasi dan dipotong pada bagian basal daunnya. Eksplan selanjutnya
ditanam pada media yang sama dan diinkubasikan pada kondisi gelap.
Selanjutnya, kalus disubkultur pada media Bac yang diperkaya dengan senyawa
N-organik (glutamin 1 mg l-1, kasein hidrolisat 500 mg l-1, arginin 120 mg l-1, dan
glisin 2 mg l-1) dan diinkubasikan pada kondisi terang (800–1000 lux) selama 16
jam terang. Kalus embriogenik dipindahkan ke media MS dengan Kn 1 mg l-1.
Embrio somatik yang belum membuka daunnya (kurang dari 0.5 cm) digunakan
sebagai eksplan untuk dienkapsulasi.
Percobaan disusun secara faktorial dalam lingkungan Rancangan Acak
Lengkap. Faktor pertama adalah taraf manitol (0, 1, 2, 3, 4, dan 5%) sedangkan
faktor kedua adalah suhu penyimpanan (15 dan 25 0C). Setiap perlakuan diulang
sebanyak 4 kali (botol) dan setiap botol terdiri atas 10 kapsul. Eksplan
dienkapsulasi dengan natrium alginat 3% yang berisi media MS yang
mengandung BA 1 mg l-1 dan IBA 0,5 mg l-1 dengan penambahan manitol. Proses
enkapsulasi dilakukan dengan metode tetes ke dalam larutan CaCl2.2H2O 100 mM
dan direndam selama 15 menit dengan penggojokan hingga membentuk kapsul.
Inkubasi pada suhu 15 0C dilakukan di dalam growth chamber dan inkubasi pada

suhu 25 0C dilakukan di ruang kultur dengan fotoperiodisitas 16 jam terang
dengan intensitas 800–1000 lux. Respon yang diamati adalah persentase daya
hidup, persentase daya regenerasi, dan persentase biakan yang menembus kapsul.
Biakan yang masih bertahan hidup kemudian dipindah ke media MS padat yang
mengandung BA 0,5 mg l-1 dan Kn 1 mg l-1 untuk pemulihan dan regenerasi.
Respon yang diamati adalah persentase biakan yang hidup dan persentase biakan
yang menembus kapsul, jumlah tunas dan jumlah daun.
Hasil dan Pembahasan
Enkapsulasi
Berdasarkan hasil percobaan sebelumnya telah diketahui bahwa basal daun
nenas memberikan respon yang baik pada media padat dan media cair. Pada tahap
ini ingin diketahui apakah respon yang sama juga diperoleh ketika eksplan
dienkapsulasi dengan kapsul alginat untuk pembentukan benih sintetik dan
penyimpanannya secara pertumbuhan minimal. Oleh karena itu, digunakan
eksplan berupa basal daun dan sebagai pembandingnya digunakan batang semu.
Hasil percobaan menunjukkan bahwa basal daun tidak menunjukkan respon
pertumbuhan walaupun kapsul alginat mengandung sitokinin dan auksin. Setelah
1 bulan, semua eksplan basal daun mengalami pencoklatan. Sebaliknya, batang
semu mampu beregenerasi membentuk tunas dengan persentase mencapai 60%
(Gambar 29 dan 30). Diduga bahwa eksplan basal daun mengalami hambatan
dalam proses respirasi atau sel-sel meristematis pada area basal daun tersebut
menerima tekanan mekanis yang cukup kuat dalam kapsul alginat sehingga tidak
mampu bertahan hidup dan tumbuh lebih lanjut hingga akhirnya mati. Sebaliknya,
batang semu mempunyai kemampuan tumbuh yang lebih baik karena
mengandung mata tunas aksilar yang secara struktural lebih terorganisir
dibandingkan dengan sel-sel meristematis pada area basal daun. Namun demikian,
tunas-tunas yang tumbuh dari batang semu tersebut terlalu cepat menembus
kapsul (dalam waktu 1 bulan) sehingga eksplan ini kurang ideal digunakan dalam
pembentukan benih sintetik karena periode simpannya sangat singkat. Selain itu,
jumlah batang semu lebih terbatas sehingga akan memerlukan bahan tanaman
induk yang lebih banyak.
0
10
20
30
40
50
60
Eksp
lan ya
ng tu
mbuh
(%)
MS N1 N2 N3 B1 B1N1 B1N2 B1N3 B2 B2N1 B2N2 B2N3 B3 B3N1 B3N2 B3N3
ZPT (mg/l)
MS = tanpa ZPT, N1 = NAA 1mg l-1, N2 = NAA 2 mg l-1, N3 = NAA 3 mg l-1, B1 = BA 1 mg l-1, B1N1 = BA 1 mg l-1 + NAA 1 mg l-1, B1N2 = BA 1 mg l-1 + NAA 2 mg l-1, B1N3 = BA 1 mg l-1 + NAA 3 mg l-1, B2 = BA 2 mg l-1, B2N1 = BA 2 mg l-1 + NAA 1 mg l-1, B2N2 = BA 2 mg l-1 + NAA 2 mg l-1, B2N3 = BA 2 mg l-1 + NAA 3 mg l-1, B3 = BA 3 mg l-1, B3N1 = BA 3 mg l-1 + NAA 1 mg l-1, B3N2 = BA 3 mg l-1 + NAA 2 mg l-1, dan B3N3 = BA 3 mg l-1 + NAA 3 mg l-1
Gambar 29. Pengaruh BA dan NAA terhadap pertumbuhan eksplan batang semu nenas kultivar Smooth Cayenne yang dienkapsulasi dalam Na-alginat 3%, 1 bulan masa inkubasi.
Untuk menginduksi diferensiasi sel-sel pada area basal daun maka diduga
perlu adanya pra-perlakuan basal daun sebelum enkapsulasi untuk memberikan
peluang bagi sel-sel di daerah basal mengalami diferensiasi. Oleh karena itu,
dilakukan pra-perlakuan basal daun dengan menggunakan media MS dengan
penambahan BA 0.5 mg l-1 dan NAA 0.5 mg l-1 yang merupakan media terbaik
untuk menginduksi pembentukan nodul berdasarkan hasil penelitian sebelumnya.
Pada tahap ini, pembentukan nodul lebih dikehendaki daripada pembentukan
tunas supaya biakan tidak terlalu cepat menembus kapsul karena penelitian ini
diarahkan untuk konservasi in vitro.
Hasil pra-perlakuan terhadap 650 helai basal daun menunjukkan bahwa
lebih dari 50% eksplan memberikan respon dan didominasi dengan pembentukan
nodul lebih dari 30% (Gambar 31). Basal yang mengandung nodul tersebut
kemudian digunakan dalam enkapsulasi karena lebih ideal daripada basal daun
yang mengandung akar atau tunas. Hasil enkapsulasi basal daun yang
mengandung nodul tersebut menunjukkan bahwa eksplan memiliki respon yang
rendah, di mana akar lebih mudah terbentuk daripada tunas dengan persentase

masing-masing sebesar 30 dan 5% (Gambar 32). Selain itu, warna daun eksplan
mengalami pemudaran dan diikuti dengan menurunnya daya hidup hingga
mengalami kematian. Dengan demikian, disimpulkan bahwa eksplan basal daun
yang mengandung nodul (hasil pra-perlakuan selama 2 bulan) masih belum layak
digunakan dalam pembentukan benih sintetik melalui teknik enkapsulasi sehingga
perlu digunakan jenis eksplan lainnya, misalnya tunas.
A
KJI
HGFE
DCB
L
NM O P
Gambar 30. Keragaan eksplan batang semu nenas kultivar Smooth Cayenne yang
dienkapsulasi dengan Na-alginat 3% (1 bulan masa inkubasi): kontrol (A), NAA 1mg l-1 (B), NAA 2 mg l-1 (C), NAA 3 mg l-1 (D), BA 1 mg l-1 (E), BA 1 mg l-1 + NAA 1 mg l-1 (F), BA 1 mg l-1 + NAA 2 mg l-1 (G), BA 1 mg l-1 + NAA 3 mg l-1 (H), BA 2 mg l-1 (I), BA 2 mg l-1 + NAA 1 mg l-1 (J), BA 2 mg l-1 + NAA 2 mg l-1 (K), BA 2 mg l-1 + NAA 3 mg l-1 (L), BA 3 mg l-1 (M), BA 3 mg l-1 + NAA 1 mg l-1 (N), BA 3 mg l-1 + NAA 2 mg l-1 (O), dan BA 3 mg l-1 + NAA 3 mg l-1 (P).

0 10 20 30 40 50 60
Tidak berespon
Nodul
Akar
Planlet
Respon eksplan (%) Gambar 31. Proporsi pembentukan planlet, akar, dan nodul dari eksplan basal
daun nenas kultivar Smooth Cayenne yang diinduksi oleh BA 0.5 mg l-1 dan NAA 0.5 mg l-1, 2 bulan periode inkubasi (data diperoleh dari total 650 eksplan basal daun).
0
20
40
60
80
100
B0N0 B0.5N0 B0.5N0.5 B1N0 B1N1ZPT (mg/l)
Resp
on e
kspla
n (%
)
Hijau Akar Tunas
Gambar 32. Pengaruh kombinasi BA dan NAA terhadap respon eksplan basal
daun nenas kultivar Smooth Cayenne yang terenkapsulasi dalam Na-alginat 3%.
Pertumbuhan minimal dengan menggunakan paklobutrazol
Ketika diketahui bahwa eksplan basal daun bernodul masih kurang sesuai
untuk dienkapsulasi maka pada tahap ini digunakan eksplan berupa tunas in vitro
dengan ukuran kurang dari 0.5 cm. Hasil percobaan menunjukkan tidak terdapat
interaksi yang nyata antara suhu inkubasi dengan taraf paklobutrazol terhadap
daya tembus eksplan umur 1 bulan. Daya tembus eksplan pada umur simpan 1
bulan tersebut dipengaruhi secara nyata oleh faktor tunggal, yaitu taraf
paklobutrazol ataupun suhu penyimpanan (Tabel 12).
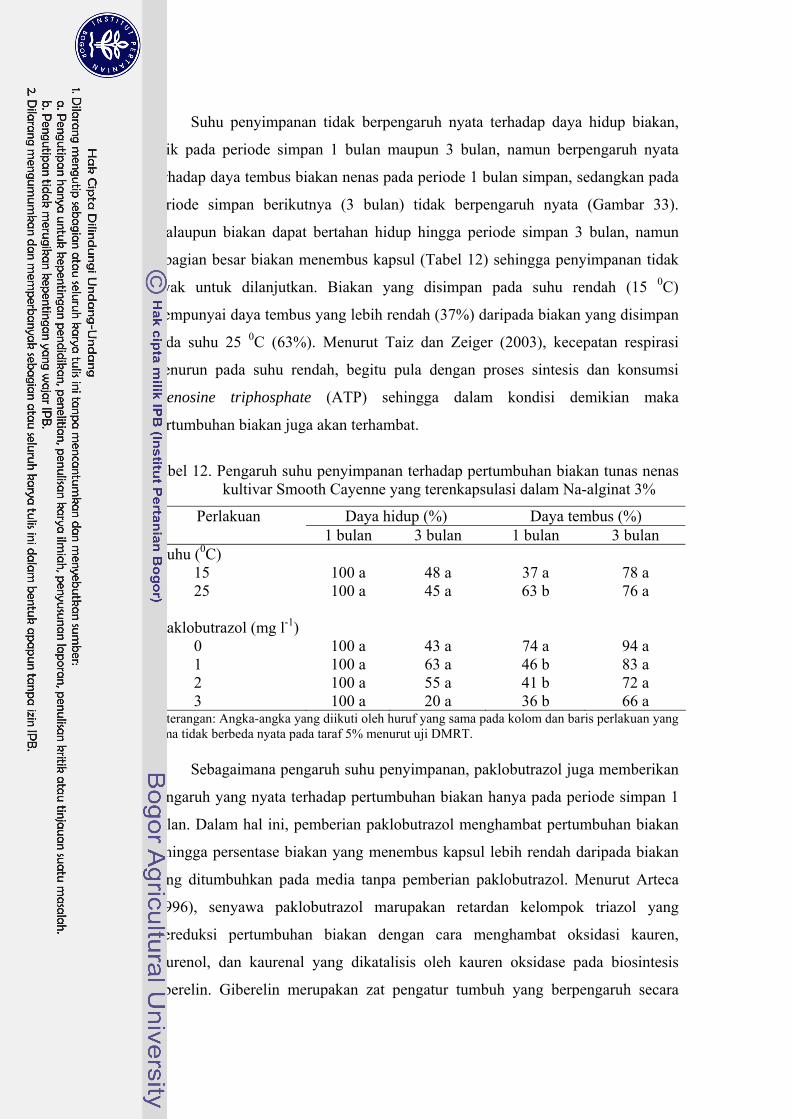
Suhu penyimpanan tidak berpengaruh nyata terhadap daya hidup biakan,
baik pada periode simpan 1 bulan maupun 3 bulan, namun berpengaruh nyata
terhadap daya tembus biakan nenas pada periode 1 bulan simpan, sedangkan pada
periode simpan berikutnya (3 bulan) tidak berpengaruh nyata (Gambar 33).
Walaupun biakan dapat bertahan hidup hingga periode simpan 3 bulan, namun
sebagian besar biakan menembus kapsul (Tabel 12) sehingga penyimpanan tidak
layak untuk dilanjutkan. Biakan yang disimpan pada suhu rendah (15 0C)
mempunyai daya tembus yang lebih rendah (37%) daripada biakan yang disimpan
pada suhu 25 0C (63%). Menurut Taiz dan Zeiger (2003), kecepatan respirasi
menurun pada suhu rendah, begitu pula dengan proses sintesis dan konsumsi
adenosine triphosphate (ATP) sehingga dalam kondisi demikian maka
pertumbuhan biakan juga akan terhambat.
Tabel 12. Pengaruh suhu penyimpanan terhadap pertumbuhan biakan tunas nenas kultivar Smooth Cayenne yang terenkapsulasi dalam Na-alginat 3%
Daya hidup (%) Daya tembus (%) Perlakuan 1 bulan 3 bulan 1 bulan 3 bulan
Suhu (0C) 15 25 Paklobutrazol (mg l-1) 0 1 2 3
100 a 100 a
100 a 100 a 100 a 100 a
48 a 45 a
43 a 63 a 55 a 20 a
37 a 63 b
74 a 46 b 41 b 36 b
78 a 76 a
94 a 83 a 72 a 66 a
Keterangan: Angka-angka yang diikuti oleh huruf yang sama pada kolom dan baris perlakuan yang sama tidak berbeda nyata pada taraf 5% menurut uji DMRT.
Sebagaimana pengaruh suhu penyimpanan, paklobutrazol juga memberikan
pengaruh yang nyata terhadap pertumbuhan biakan hanya pada periode simpan 1
bulan. Dalam hal ini, pemberian paklobutrazol menghambat pertumbuhan biakan
sehingga persentase biakan yang menembus kapsul lebih rendah daripada biakan
yang ditumbuhkan pada media tanpa pemberian paklobutrazol. Menurut Arteca
(1996), senyawa paklobutrazol marupakan retardan kelompok triazol yang
mereduksi pertumbuhan biakan dengan cara menghambat oksidasi kauren,
kaurenol, dan kaurenal yang dikatalisis oleh kauren oksidase pada biosintesis
giberelin. Giberelin merupakan zat pengatur tumbuh yang berpengaruh secara

fisiologis pada pembentukan meristem sub-apikal sehingga penghambatannya
dapat menyebabkan tanaman menjadi roset.
A B C D
E F G H
Gambar 33. Penampilan tunas nenas kultivar Smooth Cayenne yang dienkapsulasi
dengan alginat 3% yang mengandung paklobutrazol dan disimpan pada suhu yang berbeda: kontrol pada suhu 15 0C (A), paklobutrazol 1 mg l-1 suhu 15 0C (B), paklobutrazol 2 mg l-1 suhu 15 0C (C), paklobutrazol 3 mg l-1 suhu 15 0C (D), tanpa paklobutrazol suhu 25 0C (E), paklobutrazol 1 mg l-1 suhu 25 0C (F), paklobutrazol 2 mg l-1 suhu 25 0C (G), paklobutrazol 3 mg l-1 suhu 25 0C (H). Tanda lingkaran menunjukkan tunas yang telah menembus dan keluar dari kapsul sehingga mengapung.
Pada periode simpan berikutnya (3 bulan), paklobutrazol tidak memberikan
pengaruh yang nyata terhadap daya tembus dan daya hidup biakan (Tabel 12). Hal
ini menunjukkan bahwa pengaruh dari paklobutrazol tersebut berlangsung dalam
waktu yang sangat singkat. Pengaruh retardan yang cukup lama dapat teramati
pada ubi jalar (Roostika dan Sunarlim 2001; Sunarlim dan Roostika 2003),
gembili (Sunarlim et al. 2004), pule, pulasari, daun dewa (Lestari dan Mariska
1997), serta purwoceng (Roostika et al. 2009) dengan ciri-ciri visual terbentuknya
biakan roset atau pemendekan ruas sehingga biakan dapat disimpan dalam waktu
yang cukup lama (12-18 bulan). Walaupun demikian, tidak semua biakan
memberikan ciri-ciri visual tersebut, seperti yang terjadi pada biakan kentang
hitam (Roostika dan Sunarlim 2005). Kurangnya efek penghambatan
pertumbuhan biakan nenas kemungkinan disebabkan oleh rendahnya taraf
paklobutrazol atau jenis retardan dan jenis eksplan yang digunakan dalam
penelitian ini kurang sesuai sehingga perlu diganti.

Pada tahap pemulihan dan regenerasi pasca-penyimpanan, daya hidup dan
daya regenerasi biakan dari perlakuan suhu 25 0C lebih tinggi daripada biakan dari
perlakuan suhu 15 0C biakan nenas. Perlakuan tanpa paklobutrazol dan suhu 15 0C
menghasilkan daya regenerasi yang tertinggi setelah 1 bulan masa pemulihan
(Gambar 34). Pada masa pemulihan 5 bulan, pertumbuhan biakan dari perlakuan
suhu 15 0C jauh lebih pesat daripada biakan yang berasal dari perlakuan suhu 25 0C, di mana paklobutrazol 1 mg l-1 dan suhu 15 0C menghasilkan jumlah tunas
yang tertinggi (Gambar 34). Secara visual, biakan yang berasal dari perlakuan
paklobutrazol lebih tegar daripada biakan yang tidak diberi perlakuan
paklobutrazol (Gambar 35). Hal ini menunjukkan pengaruh positif dari
paklobutrazol yang dapat memacu pertumbuhan biakan. Secara fisiologis,
retardan dilaporkan dapat mendukung terbentuknya klorofil sehingga kultur
tampak lebih tegar (Cathey, 1975; Bessembinder et al. 1993). Arteca (1996)
mengatakan bahwa selain memblokir biosintesis giberelin, paklobutrazol juga
dapat mereduksi absisic acid (ABA), etilen, dan indole-3-acetic acid (IAA), dan
dapat meningkatkan kandungan sitokinin. Banyaknya klorofil yang terbentuk
dapat meningkatkan efisiensi fotosintesis sehingga pertumbuhan kultur menjadi
lebih tegar dan terpacu.
Secara umum dapat dikatakan bahwa penggunaan paklobutrazol 1 mg l-1
merupakan perlakuan terbaik daripada perlakuan lainnya. Perlakuan tersebut
sebaiknya diaplikasikan untuk penyimpanan kapsul tunas nenas dalam jangka
waktu yang pendek (1 bulan) atau untuk keperluan transportasi benih sintetik
dalam jarak dekat.
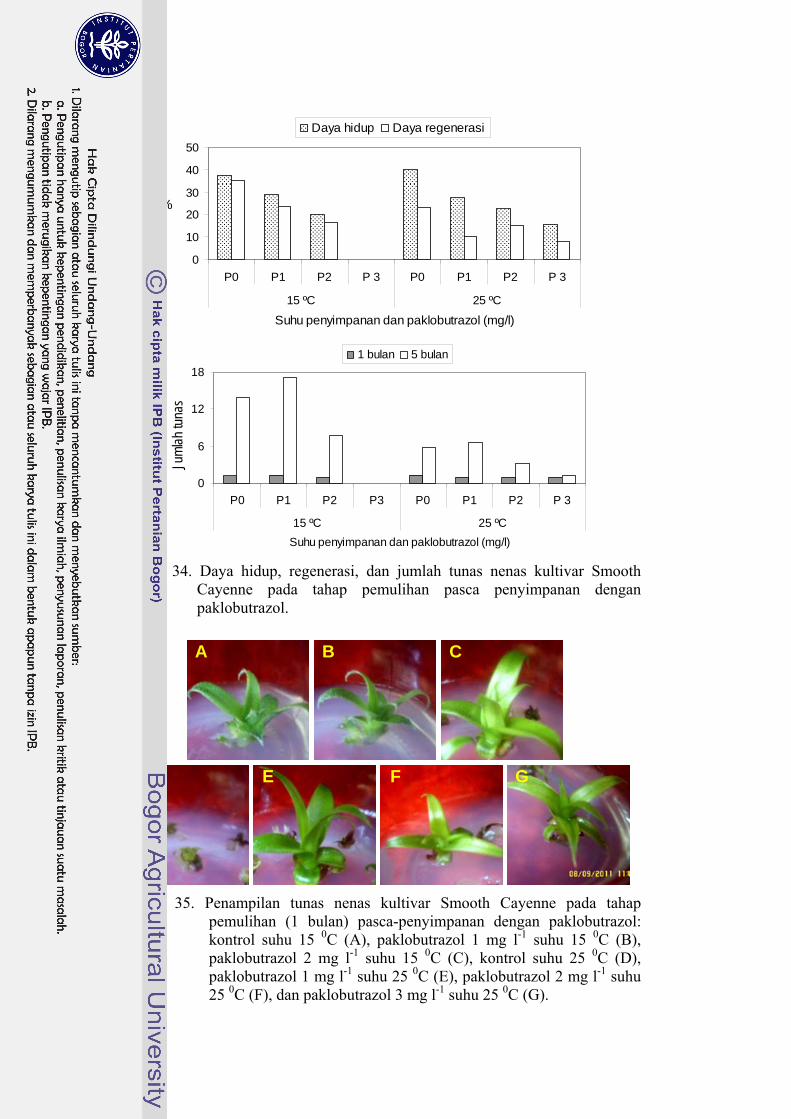
0
10
20
30
40
50
P0 P1 P2 P 3 P0 P1 P2 P 3
15 ºC 25 ºC
Suhu penyimpanan dan paklobutrazol (mg/l)
%
Daya hidup Daya regenerasi
0
6
12
18
P0 P1 P2 P3 P0 P1 P2 P 3
15 ºC 25 ºCSuhu penyimpanan dan paklobutrazol (mg/l)
Juml
ah tu
nas
1 bulan 5 bulan
Gambar 34. Daya hidup, regenerasi, dan jumlah tunas nenas kultivar Smooth
Cayenne pada tahap pemulihan pasca penyimpanan dengan paklobutrazol.
A B C
D E F G
Gambar 35. Penampilan tunas nenas kultivar Smooth Cayenne pada tahap
pemulihan (1 bulan) pasca-penyimpanan dengan paklobutrazol: kontrol suhu 15 0C (A), paklobutrazol 1 mg l-1 suhu 15 0C (B), paklobutrazol 2 mg l-1 suhu 15 0C (C), kontrol suhu 25 0C (D), paklobutrazol 1 mg l-1 suhu 25 0C (E), paklobutrazol 2 mg l-1 suhu 25 0C (F), dan paklobutrazol 3 mg l-1 suhu 25 0C (G).

Pertumbuhan Minimal dengan Manitol
Jika paklobutrazol menghambat pertumbuhan melalui pemblokiran
biosintesis giberelin (Arteca 1996), manitol merupakan gula alkohol yang
berdifusi di bagian ekstraseluler namun tidak dapat memasuki sel (Taiz dan Zeiger
2003) sehingga berfungsi sebagai regulator osmotik. Penggunaan regulator
osmotik telah terbukti dapat menyimpan biakan ubi jalar selama 10 bulan
(Roostika et al. 2001), biakan kentang hitam selama 3 bulan (Roostika dan
Sunarlim 2005), biakan purwoceng selama 7 bulan (Roostika et al. 2008), dan
biakan jeruk besar selama 5 bulan (Dewi et al. 2010).
Hasil penelitian menunjukkan tidak terdapat interaksi yang nyata antara
suhu penyimpanan dengan taraf manitol terhadap daya tembus biakan. Daya
tembus biakan tersebut sangat rendah hingga akhir periode simpan (5 bulan). Hal
ini menunjukkan bahwa manitol mampu menekan pertumbuhan embrio somatik
nenas dalam kondisi terenkapsulasi dengan Na-alginat 3%. Data tersebut perlu
didukung dengan data daya hidup biakan sehingga dapat ditentukan perlakuan
yang terbaik untuk penyimpanan.
Hasil penelitian ini menunjukkan tidak terdapat interaksi yang nyata antara
suhu penyimpanan dengan taraf manitol terhadap daya hidup biakan. Dalam hal
ini, pertumbuhan embrio somatik nenas yang terenkapsulasi dengan Na-alginat
3% dipengaruhi secara nyata oleh faktor tunggal yang diujikan, yaitu suhu
penyimpanan atau taraf manitol. Pada periode simpan 1 bulan, taraf manitol
berpengaruh nyata terhadap daya hidup biakan. Taraf manitol 2, 3, dan 4% justru
meningkatkan daya hidup biakan, namun pengaruh tersebut tidak berbeda nyata
dengan bertambahnya periode simpan hingga 5 bulan (Tabel 13). Dari tabel
tersebut tampak bahwa daya hidup biakan menurun drastis dari periode simpan 4
bulan ke periode simpan 5 bulan. Manitol merupakan osmoregulator yang dapat
meningkatkan tekanan osmotik media sehingga nutrisi mengalir secara perlahan
ke dalam jaringan. Taraf manitol yang sangat tinggi dapat menyebabkan tekanan
osmotik yang sangat tinggi pula sehingga menyebabkan seolah-olah nutrisi tidak
tersedia dan memungkinkan terjadinya dehidrasi jaringan. Oleh karena itu, biakan
yang disimpan dengan manitol pada taraf yang tinggi tidak dapat bertahan hidup
lebih lanjut. Dengan demikian, dapat dikatakan bahwa biakan nenas yang
terenkapsulasi dengan Na-alginat 3% sebaiknya disimpan dengan manitol 4%
tidak lebih dari 4 bulan.
Tabel 13. Pengaruh suhu penyimpanan dan manitol terhadap daya tembus biakan nenas kultivar Smooth Cayenne yang terenkapsulasi dengan Na-alginat 3%
Daya tembus (%) Perlakuan
1 bulan 2 bulan 3 bulan 4 bulan 5 bulan Manitol (%)
0 3 3 3 8 9 1 1 1 3 3 12 2 5 5 10 20 25 3 6 6 11 26 29 4 13 13 23 25 32 5 0 0 8 8 9
Suhu (0C) 15 0.6 a 0.6 a 4.3 a 6.9 a 10.6 a 25 7.7 b 7.7 b 13.7 a 19.8 a 25.1 b
Keterangan: Angka-angka yang diikuti oleh huruf yang sama pada kolom dan baris perlakuan yang sama tidak berbeda nyata pada taraf 5% menurut uji DMRT.
Berbeda dengan pengaruh manitol, suhu penyimpanan memberikan
pengaruh yang nyata hanya pada 5 bulan periode penyimpanan (Tabel 14), dengan
daya hidup yang sangat rendah (3 dan 27%) yang ditandai dengan banyaknya
biakan yang mencoklat (Gambar 36). Sebagaimana dijelaskan sebelumnya,
kecepatan respirasi menurun pada suhu rendah, begitu pula dengan proses sintesis
dan konsumsi adenosine triphosphate (ATP) sehingga dalam kondisi tersebut
maka pertumbuhan biakan akan sangat terhambat dan berakibat pada kematian
(Taiz dan Zeiger 2003). Oleh karena itu, penyimpanan dengan penurunan suhu
tersebut tidak disarankan untuk dilakukan. Selain daya hidup yang sangat rendah,
penyimpanan dengan penurunan suhu memerlukan energi listrik dengan biaya
yang lebih tinggi dibandingkan dengan menggunakan manitol.
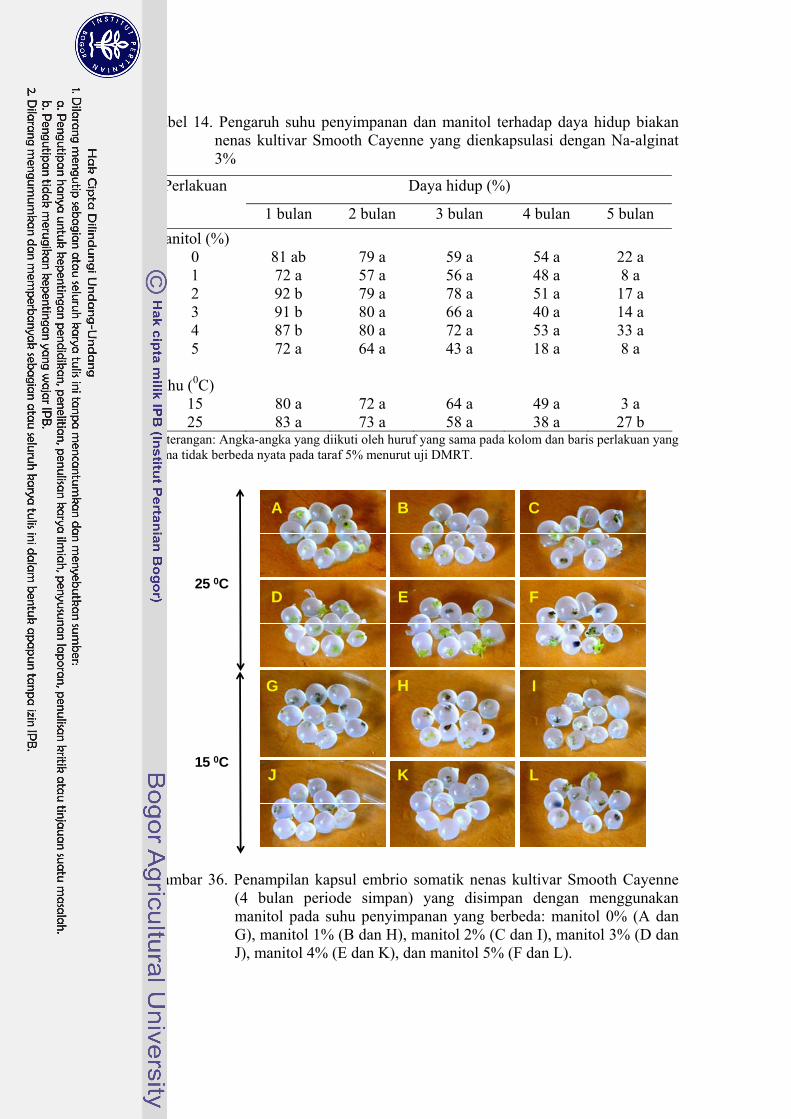
Tabel 14. Pengaruh suhu penyimpanan dan manitol terhadap daya hidup biakan nenas kultivar Smooth Cayenne yang dienkapsulasi dengan Na-alginat 3%
Daya hidup (%) Perlakuan
1 bulan 2 bulan 3 bulan 4 bulan 5 bulan Manitol (%)
0 81 ab 79 a 59 a 54 a 22 a 1 72 a 57 a 56 a 48 a 8 a 2 92 b 79 a 78 a 51 a 17 a 3 91 b 80 a 66 a 40 a 14 a 4 87 b 80 a 72 a 53 a 33 a 5 72 a 64 a 43 a 18 a 8 a
Suhu (0C) 15 80 a 72 a 64 a 49 a 3 a 25 83 a 73 a 58 a 38 a 27 b
Keterangan: Angka-angka yang diikuti oleh huruf yang sama pada kolom dan baris perlakuan yang sama tidak berbeda nyata pada taraf 5% menurut uji DMRT.
15 0C
25 0C
A
IHG
FED
CB
LJ K
Gambar 36. Penampilan kapsul embrio somatik nenas kultivar Smooth Cayenne
(4 bulan periode simpan) yang disimpan dengan menggunakan manitol pada suhu penyimpanan yang berbeda: manitol 0% (A dan G), manitol 1% (B dan H), manitol 2% (C dan I), manitol 3% (D dan J), manitol 4% (E dan K), dan manitol 5% (F dan L).

Penurunan suhu (2–10 0C) telah diterapkan pada beberapa macam tanaman,
seperti kentang, bawang, dan mentha dengan periode simpan 12 hingga 18 bulan
(Keller et al. 2006). Namun demikian, pada penelitian ini penurunan suhu
penyimpanan hingga 15 0C tidak mampu mempertahankan daya hidup dan
memperpanjang masa simpan biakan nenas. Sebaliknya penggunaan manitol lebih
baik daripada penurunan suhu penyimpanan. Hasil penelitian ini lebih baik
daripada hasil penelitian sebelumnya yang dilaporkan oleh Gangopadhyay et al.
(2004) yang hanya mampu menyimpan kapsul nenas selama 1.5 bulan pada suhu
8 0C.
Simpulan
Basal daun nenas tidak mampu bertahan hidup dalam kondisi terenkapsulasi
dengan Na-alginat 3%. Tidak terdapat interaksi yang nyata antara taraf
paklobutrazol dengan suhu penyimpanan terhadap daya hidup dan daya tembus
kapsul tunas nenas. Biakan tersebut hanya dapat disimpan selama 1 bulan.
Interaksi yang nyata juga tidak dijumpai antara taraf manitol dan suhu
penyimpanan terhadap daya hidup dan daya tembus kapsul embrio somatik nenas.
Dengan menggunakan manitol 4% maka penyimpanan dapat diperpanjang hingga
4 bulan. Penurunan suhu penyimpanan tidak disarankan karena terbukti tidak
mampu memperpanjang masa simpan baikan nenas dan memerlukan energi listrik
serta biaya yang lebih tinggi.
Daftar Pustaka
Arteca RN. 1996. Plant Growth Substances. Chapman and Hall. New York.332p.
Bessembinder JJE, Staritsky G, Zandvoort EA. 1993. Longterm in vitro storage of Colocasia esculenta under minimal growth conditions. Plant Cell Tiss Org Cult 33:121-127.
Cathey HM. 1975. Comparative plant growth-retarding activities of ancymidol with ACPC, phosfon, clhormequat, and SADH on ornamental plant species. Hort Science 10(3):204-215.
Desbrunais AB, Noirot M, Charrier A. 1992. Slow growth in vitro conservation of coffee (Coffea spp.). Plant Cell Tiss Org Cult 31:105-110.

Dewi IS, Jawak G, Roostika I, Sabda M, Purwoko BS, Adil WH. 2010. Konservasi in vitro tanaman jeruk besar (Citrus maxima (Burn.) Merr.) kultivar Srinyonya menggunakan osmotikum dan retardan. Jurnal Agrobiogen 6(2):84-90
Eeuwens CJ et al. 2002. Effects of tissue culture conditions during embryoid multiplication on the incidence of “mantled” flowering in clonally propagated oil palm. Plant Cell Tiss Org Cult 70:311-323.
Gangopadhyay G, Bandyopadhyay T, Poddar R, Gangopadhyay SB, Mukherjee KK. 2005. Encapsulation of micro shoots in alginate beads for temporary storage. Curr Res 88(6): 972-977.
Hu CY, Wang PJ. 1983. Meristem, shoot tip, and bud culture. Pp. 177-227. In. Evans DA, Sharp WR, Amiroto PV, Yamada Y, editor. Handbook of Plant Cell Culture Vol. I. Techniques for Propagation and Breeding. McMilan Publishing, New York.
Keller ERJ, Senula A, Leunufna S, Grube M. 2006. Slow growth storage and cryopreservation-tools to facilitate germplasm maintenance of vegetatively propagated crops in living plant collections. Int J of Refr 29:411-417.
Lestari EG, Mariska I. 1997. Kultur in vitro sebagai metode pelestarian tumbuhan obat langka. Buletin Plasma Nutfah 2(1): 1-8.
Leunufna S. 2004. Improvement of the in vitro maintenance and cryopreservation of yams (Dioscorea spp.). Dissertation Martin-Luther–Universitat Halle-Wittenberg. Berlin. 12p. Unpublished.
Mariska I, Suwarno, Damardjati DS. 1996. Pengembangan konservasi in vitro sebagai salah satu bentuk pelestarian plasma nutfah di dalam bank gen. Dalam: Seminar Penyusunan Konsep Pelestarian Ex Situ Plasma Nutfah Pertanian. Bogor. 18 Desember 1996. Balitbio Bogor.
Rai MK, Asthana P, Singh SK, Jaiswal VS, Jaiswal U. 2009. The encapsulation technology in fruit plants – A review. Biotech Adv 27: 671-679.
Redenbaugh K. 1992. Synseeds: Application of synthetic seeds to crop improvement. CRC Press. 481 p.
Roostika I, Sunarlim N. 2001. Penyimpanan in vitro tunas ubi jalar dengan penggunaan paclobutrazol dan ancymidol. PP 20 (3):48-56.
Roostika I, Sunarlim N, Arief VN. 2005. Penyimpanan kentang hitam (Coleus tuberosus) secara kultur in vitro. PP 24 (1):46-52
Roostika I, Purnamaningsih R, Darwati I. 2009. Penyimpanan in vitro tanaman purwoceng (Pimpinella pruatjan Molk.) melalui aplikasi pengenceran media dan paklobutrazol. Jurnal Littri 15(2): 84-90

Roostika I, Purnamaningsih R, Arief VN. 2008. Pengaruh sumber karbon dan kondisi inkubasi terhadap pertumbuhan kultur in vitro purwoceng (Pimpinella pruatjan Molk.). AgroBiogen 4(2): 65-69
Sunarlim N, Roostika I, Arief VN. 2004. Penyimpanan in vitro gembili melalui pertumbuhan minimal. Dalam: Prosiding Seminar Kinerja Penelitian Mendukung Agribisnis Kacang-kacangan dan Umbi-umbian. BALITKABI. Malang.
Sunarlim N, Roostika I. 2003. Penggunaan zat penghambat tumbuh dan regulator osmotik manitol dalam penyimpanan ubi-ubian secara kultur jaringan. Dalam: Prosiding Seminar Teknologi Inovatif Agribisnis Kacang-kacangan dan Umbi-umbian untuk Mendukung Ketahan Pangan. BALITKABI. Malang.
Taiz L, Zeiger E. 2003. Plant Physiology-on line 3rd Ed. Sunderland: Sinauer Associates. doi:10.1093/aob/mcg079.
Withers LA. 1985. Cryopreservation and storage of germplasm. In: Dixon DA, editor. Plant Cell Culture. IRL Press, Washington. Pp.169-190.

KARAKTERISASI MORFOLOGI DAN MOLEKULER BIAKAN IN VITRO DAN BIBIT NENAS
Abstrak
Variasi somaklonal menjadi perhatian yang serius pada mikropropagasi tanaman nenas. Oleh karena itu, diperlukan metode yang mampu mendeteksi dan mengontrol kelainan biakan in vitro sebelum planlet yang dihasilkan ditanam di lapang. Tujuan penelitian adalah untuk mengetahui tipe dan tingkat keragaman morfologi serta keragaman molekuler yang terjadi pada biakan in vitro nenas yang dikulturkan dalam periode yang panjang. Pada penelitian ini dilakukan karakterisasi morfologi dan molekuler biakan dan bibit nenas kultivar Smooth Cayenne umur 4 tahun dan evaluasi kestabilan fenotipe varian pada tahap in vitro dan ex vitro. Karakterisasi morfologi dilakukan secara kualitatif sedangkan karakterisasi molekuler dilakukan dengan menggunakan penanda RAPD dengan 10 primer. Data diskor secara biner kemudian dianalisis dengan menggunakan program NTSYS-pc 2.02. Tingkat kemiripan dihitung dengan menggunakan koefisien Dice. Pengelompokan dilakukan dengan berdasarkan UPGMA. Evaluasi kestabilan fenotipe varian pada tahap in vitro dilakukan dengan mengisolasi tiap-tiap varian secara individual dan melakukan subkultur sebanyak 3 kali sedangkan evaluasi pada tahap ex vitro dilakukan dengan mengamati perubahan karakter spesifik setiap varian di rumah kaca. Hasil penelitian memperlihatkan 21 varian terdeteksi dari populasi biakan in vitro nenas umur 4 tahun. Delapan varian menunjukkan fenomena epigenetik selama isolasi varian secara individual, 8 varian menunjukkan fenomena khimera selama proses subkultur, dan 7 varian menunjukkan kestabilan fenotipenya selama di rumah kaca. Karakterisasi morfologi tersebut menghasilkan dendogram dengan koefisien kemiripan sebesar 0.49-0.91 (r=0.83). Karakterisasi molekuler tersebut menghasilkan dendogram dengan koefisien kemiripan sebesar 0.32-0.91, di mana varian dan tanaman kontrolnya memiliki kemiripan sebesar 0.32-0.61 (r=0.97). Pola duri pada daun dari bibit hasil aklimatisasi di rumah kaca menampakkan pola yang sama sekali tidak beraturan karena karakter daun halus dan berduri (1 sisi atau 2 sisi) terjadi secara silih berganti sepanjang pertumbuhan tanaman. Selanjutnya, diduga kuat adanya elemen loncat pada tanaman nenas yang diamati. Dugaan tersebut diperkuat oleh penemuan sebelumnya yang melaporkan bahwa sekuens mirip retrotransposon telah terintegrasi ke dalam genom tanaman nenas. Metode karakterisasi morfologi yang dihasilkan dari penelitian ini dapat diterapkan untuk deteksi dini dan eliminasi varian yang tak diharapkan selama proses mikropropagasi.
Kata kunci: Karakterisasi morfologi, RAPD, biakan in vitro, Ananas comosus (L.) Merr., epigenetik, khimera, variasi somaklonal

Abstract
Somaclonal variation still becomes a concern in propagation of pineapple plants. Reliable methods are needed to control and to detect deleterious off-types produced during in vitro culture, especially before the plants are grown in the field. The objective of the study was to determine the type and the level of variation of long period in vitro cultures of pineapple. Morphological and molecular characterization of 4 year-old pineapple in vitro cultures and evaluation of phenotypic stability during in vitro and ex vitro culture were conducted in this research. Morphological characterizations were conducted qualitatively and molecular characterizations were conducted by using RAPD markers with 10 primers. The data were scored binary and then they were analyzed by using NTSYS-pc 2.02 program. The similarity degrees were calculated according to the Dice coefficient. Groupings were carried out using UPGMA cluster analysis. Twenty one aberrant phenotypes were observed in cultures derived from the shoot proliferation method, of which the 8 variants showed epigenetic phenomena and the 8 variants showed polysomic cells, indicated chimera tissues. Phenotypic evaluations showed that 7 variants were stable during acclimatization in glass house. The morphological characterization yielded dendogram with the similarity coefficient range from 0.49 to 0.91 (r=0.83). The molecular characterization yielded dendogram with the similarity coefficient range from 0.32 to 0.91 where separated variant and the control plant within the range of 0.32 to 0.61 (r=0.97). Unexpectedly, the observation of spininess showed that there was irregular pattern (the spines sometimes emerged from the young leaves, sometimes along one side of the leaves or along both side of the leaves with different density and sharpness). Therefore, it strongly suggested the presence of retrotransposon. The morphological characterization resulted in this research can be used as guidance for early detection and elimination of undesirable variants in micropropagation.
Keywords: Morphological characterization, RAPD, in vitro culture, Ananas comosus (L.) Merr, epigenetic, chimera, somaclonal variation
Pendahuluan
Teknik kultur in vitro merupakan teknologi alternatif yang tepat untuk
produksi bibit secara masal karena mampu memproduksi bibit dalam jumlah yang
besar, dalam waktu yang singkat, dan dengan keseragaman yang tinggi. Menurut
Smith et al. (2003), teknik mikropropagasi tanaman nenas dapat diterapkan untuk
pemantapan blok multiplikasi yang dapat menyediakan bahan tanaman
konvensional untuk blok produksi. Hal ini disebabkan oleh penyimpangan (off
types) yang terjadi pada bibit nenas hasil kultur in vitro. Wakasa (1979)
melaporkan bahwa bibit nenas yang dihasilkan dari eksplan berupa mahkota
mempunyai tingkat keragaman yang lebih rendah (7%) daripada syncarp (100%),
tunas aksilar (34%), dan slip (98%).

Dari berbagai kajian pustaka, dapat diketahui bahwa off types dapat
disebabkan oleh berbagai macam faktor, yaitu variasi dari tanaman donor (pre-
existing variation) (Wakasa 1979), periode in vitro yang panjang (Koornneef
1991; Masoud and Hamta 2008), zat pengatur tumbuh (Bairu et al. 2009),
subkultur yang frekuentif (Eeuwens et al. 2002), dan tergantung pada genotipe
tanaman (Zucchi et al. 2002). Untuk menghindari tingginya keragaman
somaklonal dan untuk mengefektifkan metode mikropropagasi maka diperlukan
strategi untuk memproduksi bibit dengan kesamaan identitas genetik. Oleh karena
itu, diperlukan metode karakterisasi yang mampu mendeteksi penyimpangan
tersebut secara dini, sebelum tanaman ditanam di lapang.
Deteksi penyimpangan bibit yang dihasilkan dari teknik kultur in vitro
dapat dilakukan melalui karakterisasi morfologi (Noor et al. 2009 ; Somsri et al.
2009; Zhao et al. 2005; Podwyszyńska 2005), sitologi (Al-Zahim et al. 1999;
Joachimiak dan Ilnicki 2003), biokimia (Feuser et al. 2003), fisiologi (Perez et al.
2011), dan molekuler (Chen et al. 1998; Al-Zahim et al. 1999; Soniya et al.
2001). Pada tanaman nenas, karakterisasi telah dilakukan secara morfologi,
biokimia, dan molekuler. Soneji et al. (2002) telah mengkarakterisasi bibit nenas
kultivar Queen untuk sifat daun berduri dan tidak berduri dengan penanda RAPD.
Feuser et al. (2003) juga menganalisis ketepatan genetik planlet nenas kultivar
Amarelinho (grup Perola) dengan menggunakan penanada RAPD. Demikian pula,
Santos et al. (2008) melakukan evaluasi keragaman genetik dari biakan in vitro
nenas hias (Ananas comosus var. bracteatus) dengan menggunakan penanda yang
sama. Wakasa (1979) telah melakukan evaluasi karakter morfologi bibit nenas
kultivar Smooth Cayenne hasil kultur in vitro dengan periode 1.5 tahun,
aklimatisasi selama 14-22 bulan, dan 1 tahun periode pertumbuhan tanaman.
Suminar (2010) telah melakukan karakterisasi morfologi biakan in vitro nenas
Golden Pineapple (kultivar Smooth Cayenne) hasil radiasi sinar gamma. Namun
hingga saat ini, belum terdapat penelitian yang melaporkan karakterisasi secara
morfologi biakan in vitro nenas yang dikulturkan dalam periode yang panjang.
Tujuan penelitian ini adalah mengkarakterisasi biakan in vitro umur 4 tahun dan
bibit nenas aklimatisasi dengan menggunakan penanda morfologi dan molekuler.

Bahan dan Metode
Persiapan Bahan Tanaman
Bahan tanaman adalah biakan in vitro nenas kultivar Smooth Cayenne Klon
Simadu dari Subang yang telah berumur 4 tahun pada periode in vitro. Bahan
tanaman awal adalah mahkota yang disterilisasi dengan menggunakan alkohol dan
natrium hipoklorit. Eksplan ditanam pada media MS (Murashige dan Skoog 1962)
dengan penambahan gula 3%, agar 0.8% dengan penambahan benzyl adenine
(BA) 2 mg l-1 dan kinetin (Kn) 2 mg l-1. Kultur diinkubasikan pada kondisi terang
16 jam dengan intensitas cahaya 800-1000 lux, suhu 25±2 0C. Setelah 4 tahun
periode in vitro, biakan dipelihara pada media MS dengan penambahan BA 0.5
mg l-1 dan Kn 1 mg l-1.
Karakterisasi Morfologi
Pengamatan terhadap kelainan atau penyimpangan biakan in vitro nenas
dilakukan secara visual. Karakterisasi secara morfologi dilakukan dengan
menggunakan deskripsi yang dikeluarkan oleh IBPGR (1991) dan Suminar (2010)
dengan beberapa modifikasi (Tabel 15). Data dari karakter morfologi diskor
secara biner. Data tersebut dianalisis dengan menggunakan program SIMQUAL,
SAHN, dan TREE dari Numerical Taxonomy and Multivariate Analysis System
version 2.02 (NTSYS-pc 2.02). Tingkat kemiripan dihitung dengan menggunakan
koefisien Dice. Pengelompokan dilakukan dengan metode klastering berdasarkan
Unweighted Pair Group Method and Arithmetic Average (UPGMA).
Evaluasi Stabilitas Varian
Untuk mengetahui frekuensi biakan normal dan abnormal, tiap-tiap varian
yang muncul dari populasi biakan in vitro nenas umur 4 tahun dihitung
jumlahnya. Untuk mengetahui kestabilan fenotipe tersebut maka setiap individu
varian diisolasi pada media BA 0.5 mg l-1 dan Kn 1 mg l-1. Subkultur dilakukan
sebanyak 3 kali dengan menggunakan media yang sama. Stabilitas sifat
dikategorikan menjadi 3 macam, yaitu stabil (S) jika tidak terjadi perubahan
fenotipe, agak stabil (R) jika anakan atau regeneran yang dihasilkan mempunyai

fenotipe yang sesuai dengan fenotipe awalnya dan fenotipe yang lain dengan
fenotipe awalnya, tidak stabil (N) jika terjadi perubahan fenotipe. Uji stabilitas
fenotipe dilakukan ketika isolasi varian secara individual, selama proses
subkultur, dan selama pertumbuhan bibit di rumah kaca. Kestabilan fenotipe
ditentukan secara kualitatif dengan menitikberatkan pada karakter spesifik yang
dimiliki oleh tiap-tiap varian berdasarkan karakterisasi morfologi. Selain
perubahan fenotipe, jumlah tunas juga dihitung untuk mengetahui tingkat
multiplikasi tunas dari masing-masing varian selama proses subkultur. Masing-
masing tunas dari varian diinduksi perakarannya dengan menggunakan indole
butyric acid (IBA) 3 mg l-1 dan naphthalene acetic acid (NAA) 1 mg l-1 (5.4 µM).
Planlet yang diperoleh tersebut kemudian diaklimatisasi di rumah kaca dengan
menggunakan media tanah dan kompos dengan perbandingan 1:1.
Karakterisasi Molekuler
Sebanyak 10 varian dipilih untuk karakterisasi secara molekuler. Varian
yang telah diaklimatisasi di rumah kaca diambil daunnya sebagai sampel, kecuali
varian climb posture yang diambil sampel daunnya dari biakan in vitro. Sampel
daun diekstraksi dengan metode cetyl trimethyl ammonium bromide (CTAB).
Karakterisasi molekuler dilakukan dengan menggunakan metode Randomly
Amplified Polymorphism DNA (RAPD), modifikasi metode Soneji et al. (2002).
DNA diamplifikasi dengan mesin Polymerase Chain Reaction (PCR). Masing-
masing reaksi mengandung 25 ng/l DNA genomik 1 µl, 1 µl primer (Operon
Tech. Alameda, USA) pada 10 ng/µl, 11 µl ddH2O, dan 12 µl PCR mix, dalam
total reaksi 25 µl. Siklus PCR terdiri atas pra-denaturasi pada suhu 94 0C selama 4
menit; 40 siklus pada 94 0C selama 30 detik (denaturasi); 36 0C selama 1 menit
(penempelan primer) dan 1 menit pada 72 0C (ekstensi); pasca-ekstensi pada 72 0C selama 5 menit, serta ekstensi akhir pada 40 0C selama 4 menit. Produk PCR
dipisahkan pada agarosa 1.2% (Promega) dan dielektroforesis pada voltase
konstan 50 V selama 50 menit, kemudian pita-pita diwarnai dengan ethidium
bromide (EtBr) serta divisualisasi di bawah UV transiluminator. Sebagai standar
molekuler digunakan DNA ladder 1 kb.

Tabel 15. Karakter dan sub karakter yang diamati pada biakan in vitro nenas kultivar smooth Cayenne (umur 4 tahun)
No. Karakter Sub karakter 1 Postur tunas Tegak/membengkok/agak kerdil/kerdil/tinggi
2 Tegakan daun Tegak lurus/agak vertikal/terbuka/menjuntai 3 Warna daun Hijau/hijau gelap/hijau terang/putih
keperakan/varigata 4 Panjang daun Normal/pendek/panjang 5 Lebar daun Normal/lebar/sempit 6 Pola pada daun Normal/bergaris putih/bergaris hijau/berbintik di
bagian adaksial/berbintik di bagian abaksial 7 Permukaan daun Normal/mengkilap/berlilin 8 Helaian daun Normal/keriting/berpilin/bergelombang 9 Konsistensi daun Normal/kaku/lunglai 10 Duri Ada/tidak ada 11 Panjang batang semu Normal/lebih pendek/lebih tinggi
12 Panjang nodus Normal/lebih pendek/lebih panjang 13 Filotaksi Normal/lebih rapat/lebih jarang/tak beraturan 14 Jumlah anakan Normal/banyak/jarang/hampir tidak ada
15 Asal tumbuh anakan Basal tunas/basal daun/nodus/tak diketahui 16 Dominanasi apikal Ada/tidak ada
Sampel diskor berdasarkan ada dan tidak adanya pita pada ukuran yang
sama, di mana 1 berarti ada sedangkan 0 berarti tidak ada. Untuk skrining primer,
digunakan sampel DNA tanaman nenas kultivar Smooth Cayenne klon Simadu
dari lapang dan varian needle leaf dengan menggunakan 20 primer kemudian
dipilih 10 primer yang menghasilkan pita yang jelas. Data tersebut dianalisis
dengan menggunakan program SIMQUAL, SAHN, dan TREE dari NTSYS-pc
2.02. Tingkat kemiripan dihitung dengan menggunakan koefisien Dice.
Pengelompokan dilakukan dengan metode klastering berdasarkan UPGMA.

Hasil dan Pembahasan
Karakterisasi Morfologi
Untuk membedakan tipe normal dan varian maka diperlukan informasi
tentang karakter asli dari tipe liar tanaman nenas. Wakasa (1979) telah melakukan
pengamatan morfologi bibit nenas selama aklimatisasi dan di lapang. Berdasarkan
laporan tersebut maka diketahui bahwa fenotipe asli dari tanaman nenas kultivar
Smooth Cayenne adalah daun berwarna hijau gelap, tidak ada varigata, bagian
abaksial daun ditutupi oleh lapis lilin, dan duri muncul di sekitar ujung daun
(penciri spesifik dari kultivar tersebut).
Berdasarkan karakterisasi morfologi, teramati 21 varian dari populasi
biakan in vitro nenas umur 4 tahun. Tabel 16 dan Gambar 37 memperlihatkan tipe
variasi dan ciri-ciri spesifik dari tiap-tiap varian. Tipe varian yang dihasilkan pada
penelitian ini lebih beragam dibandingkan dengan hasil penelitian Wakasa (1979)
yang memperoleh 6 varian, yaitu (spines leaf, lighter green leaf, denser
phyllotaxy, powdery leaf, wider leaf, dan albino stripe leaf). Banyaknya varian
yang muncul pada penelitian ini kemungkinan disebabkan oleh periode in vitro
yang lebih panjang. Wakasa (1979) menerapkan kultur in vitro selama 1.5 tahun
sedangkan pada penelitian ini digunakan biakan in vitro yang telah berumur 4
tahun. Masoud dan Hamta (2008) juga mengatakan bahwa periode in vitro dapat
menyebabkan induksi variasi somaklonal. Joachimiak dan Ilnicki (2003) juga
menjelaskan bahwa selama periode kultur in vitro, jaringan tanaman dapat
mengalami perubahan struktural, antara lain tingkat ploidi, ukuran telomer,
eliminasi kromatin yang berasosiasi dengan ukuran nukleus dan kandungan
deoxyribo nucleic acid (DNA).

A B C
D E F
G H I
J K L
M N O
P Q R
S T U
W Gambar 37. Keragaan varian nenas kultivar Smooth Cayenne: normal (a),
branched shoot (b). needle leaf (c). long core (e). climb posture (f), whitish leaf (g), glossy stiff leaf (h), spiness spot leaf (i), narrow leaf (j), bending posture (k), albino stripes leaf (l), curly leaf (m), wider leaf (n), dwarf posture (o), powdery leaf (p), ultra dwarf posture (q), wave laef (r), darker leaf (s), lighter leaf (t), denser foliage (u), reddish leaf (v), erect posture (w).

Jumlah varian yang dihasilkan dalam penelitian ini bahkan lebih banyak
jika dibandingkan dengan hasil radiasi sinar gamma yang dilakukan oleh Suminar
(2010). Tampaknya pengaruh periode kultur in vitro lebih besar dalam
menginduksi terbentuknya varian, terutama ketika dosis radiasi yang diujikan
masih belum optimal. Suminar (2010) mengatakan bahwa kalus yang
mendapatkan perlakuan radiasi sebagian besar mempunyai pertumbuhan yang
baik dan tidak menunjukkan gejala kerusakan fisiologis.
Analisis pengelompokan berdasarkan 16 karakter dan 58 sub karakter
(Tabel 15) menghasilkan koefisien kemiripan sebesar 0.49-0.91 (Gambar 38).
Analisis pengelompokan tersebut memiliki nilai korelasi matriks Rohlf sebesar
0.83 (r=0.83). Hal ini berarti bahwa dendogram yang diperoleh sudah sesuai
dalam menggambarkan pengelompokan berbagai macam varian atau fenotipe
tersebut. Pada koefisien kemiripan 0.62, populasi tersebut terpisah atas 3
kelompok.
Hasil penelitian ini juga memperlihatkan bahwa varian bending posture
memiliki proporsi paling tinggi (16.6%), diikuti dengan needle-like leaf (15.1%),
spines spot leaf (9.8%), glossy stiff leaf (8.6%), narrow leaf (5.4%), dan curly leaf
(5.3%) (Tabel 16). Proprosi tersebut kemungkinan berkaitan dengan tingkat
multiplikasi tunas dari tiap-tiap varian (Tabel 17). Selain itu, terdapat 3 varian
yang kehilangan dominansi apikalnya yang berasosiasi dengan tingkat
multiplikasi tunas yang tinggi.
Evaluasi Stabilitas Varian
Zucchi et al. (2002) melaporkan bahwa stress selama kultur in vitro dapat
menyebabkan terjadinya metilasi di dalam genom tanaman. Diduga bahwa
perubahan epigenetik dalam tingkat metilasi DNA memegang peranan penting
dalam mekanisme adaptasi tanaman terhadap stres lingkungan. Menurut
Koornneef (1991), diduga bahwa propagasi vegetatif yang stabil dari suatu varian
dapat dipertimbangkan sebagai indikasi bahwa varian tersebut mutan atau bukan
mutan. Oleh karena itu, pada penelitian ini dilakukan subkultur sebanyak 3 kali.
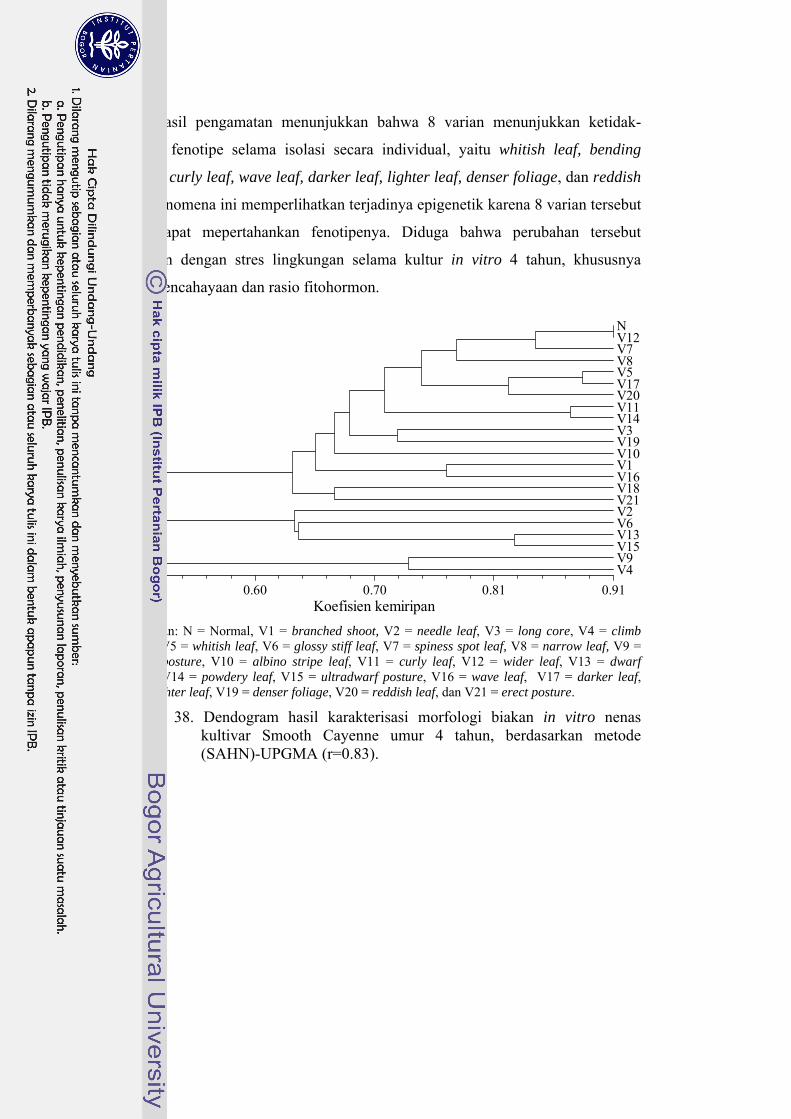
Hasil pengamatan menunjukkan bahwa 8 varian menunjukkan ketidak-
stabilan fenotipe selama isolasi secara individual, yaitu whitish leaf, bending
posture, curly leaf, wave leaf, darker leaf, lighter leaf, denser foliage, dan reddish
leaf. Fenomena ini memperlihatkan terjadinya epigenetik karena 8 varian tersebut
tidak dapat mepertahankan fenotipenya. Diduga bahwa perubahan tersebut
berkaitan dengan stres lingkungan selama kultur in vitro 4 tahun, khususnya
faktor pencahayaan dan rasio fitohormon.
Koefisien kemiripan0.49 0.60 0.70 0.81 0.91
N V12 V7 V8 V5 V17 V20 V11 V14 V3 V19 V10 V1 V16 V18 V21 V2 V6 V13 V15 V9 V4
Keterangan: N = Normal, V1 = branched shoot, V2 = needle leaf, V3 = long core, V4 = climb posture, V5 = whitish leaf, V6 = glossy stiff leaf, V7 = spiness spot leaf, V8 = narrow leaf, V9 = bending posture, V10 = albino stripe leaf, V11 = curly leaf, V12 = wider leaf, V13 = dwarf posture, V14 = powdery leaf, V15 = ultradwarf posture, V16 = wave leaf, V17 = darker leaf, V18 = lighter leaf, V19 = denser foliage, V20 = reddish leaf, dan V21 = erect posture.
Gambar 38. Dendogram hasil karakterisasi morfologi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun, berdasarkan metode (SAHN)-UPGMA (r=0.83).

Tabel 16. Ciri-ciri spesifik dari varian yang muncul dari populasi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun
No. Varian Karakter spesifik
1 Branched shoot (V1)
Banyak tunas, dominansi apikal hilang, daun normal atau terkulai
2 Needle leaf (V2) Daun agak terbuka, pendek, dan sempit, kaku, seperti jarum, mempunyai banyak anakan, tidak ada dominansi apikal
3 Long core (V3) Batang semu lebih pajang dan besar tetapi nodus tidak memanjang, daun terakumulasi di bagian atas tunas
4 Climb posture (V4)
Tunas tinggi, tampak seperti memanjat, daun pendek dan sempit, normal atau terkulai, nodus lebih panjang
5 Whitish leaf (V5)
Daun keperakperakan dan berduri
6 Glossy stiff leaf (V6)
Daun mengkilat dan kaku, elongasi tunas agak terbatas
7 Spines spot leaf (V7)
Duri muncul pada sepanjang tepi daun, permukaan daun berbintik-bintik putih
8 Narrow leaf (V8)
Daun lebih sempit dan ramping, menjuntai ke bawah atau tidak
9 Bending posture (V9)
Nodus lebih panjang, tunas membelok-belok, tidak roset
10 Albino stripe leaf (V10)
Daun bergaris-garis putih dan hijau atau albino
11 Curly leaf (V11) Daun melengkung dengan ujung melekuk ke bawah, keriting atau spiral
12 Wider leaf (V12) Daun lebih lebar 13 Dwarf posture
(V13) Tunas kerdil, lebar daun normal, warna daun lebih gelap
14 Powdery leaf (V14)
Bagian abaksial dan adaksial daun seperti bertaburan tepung dipenuhi lapis lilin atau kutikula
15 Ultradwarf posture (V15)
Tunas kerdil, daun lebih kecil dan lebih pendek
16 Wave laf (V16) Struktur daun bergelombang dengan filotaksi tak beraturan
17 Darker leaf (V17)
Daun berwarna hijau tua
18 Lighter leaf (V18)
Daun berwarna hijau muda
19 Denser foliage (V19)
Tata letak daun lebih rapat
20 Reddish leaf (V20)
Bagian sekitar tulang daun berwarna kemerah-merahan
21 Erect posture (V21)
Tunas tegak dengan daun agak tegak lurus

Tabel 17. Proporsi varian yang muncul dalam populasi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun
Varian Jumlah tunas Proporsi (%) Normal 134 17.7 Branched shoot (V1) 4 0.5 Needle leaf (V2) 114 15.1 Long core (V3) 4 0.5 Climb posture (V4) 17 2.3 Whitish leaf (V5) 5 0.7 Glossy stiff leaf (V6) 65 8.6 Spines spot leaf (V7) 74 9.8 Narrow leaf (V8) 41 5.4 Bending posture (V9) 141 18.7 Albino stripe leaf (V10) 4 0.5 Curly leaf (V11) 40 5.3 Wider leaf (V12) 14 1.9 Dwarf posture (V13) 30 4.0 Powdery leaf (V14) 25 3.3 Ultradwarf posture (V15) 21 2.8 Wave laf (V16) 8 1.1 Darker leaf (V17) 2 0.3 Lighter leaf (V18) 1 0.1 Denser foliage (V19) 6 0.8 Reddish leaf (V20) 3 0.4 Erect posture (V21) 2 0.3 Total 755 100
Terjadinya varian bending posture kemungkinan disebabkan oleh upaya
biakan dalam mengakses cahaya. Srivastava (2002) mengatakan bahwa
pertumbuhan yang asimetris sebagian diperantarai oleh auksin. Menurut Taiz dan
Zeiger (2002), pertumbuhan ke arah cahaya demikian disebut sebagai
fototropisme. Perolehan cahaya yang tidak seimbang menyebabkan ketidak-
seimbangan konsentrasi auksin pada daerah yang terkena cahaya dan yang
ternaungi sehingga menyebabkan pertumbuhan yang tidak seimbang atau
membelok. Asumsi ini dikonfirmasi dengan kenyataan yang menunjukkan bahwa
tunas-tunas yang bengkok tersebut berukuran kecil dan selalu bergerombol.
Alasan yang serupa juga ditujukan pada varian whitish leaf, lighter leaf, darker
leaf, dan reddish leaf, di mana cahaya yang tidak mencukupi atau bahkan
berlebihan menyebabkan munculnya varian-varian tersebut.

Varian curly leaf dan wave leaf kemungkinan berkaitan dengan taraf auksin
endogenus. Menurut Srivastava (2002), hilangnya homeostasis IAA dapat
menyebabkan perubahan taraf IAA bebas sehingga tanaman memperlihatkan daun
yang epinastik oleh karena pertumbuhan yang asimetris. Pada penelitian ini,
varian wave leaf berubah menjadi curly leaf. Ini menunjukkan bahwa pada
kenyataannya, kedua varian tersebut adalah sama.
Fenotipe branched shoot, needle leaf, dan dwarf posture kemungkinan
berkaitan dengan overproduksi sitokinin. Semua varian tersebut mempunyai
tingkat multiplikasi tunas yang tinggi (Tabel 18) dan kehilangan dominansi
apikalnya. Srivastava (2002) meriviu bahwa produksi sitokinin yang berlebihan
akan menghasilkan kelainan yang bermacam-macam, dan kelaianan yang paling
konsisten adalah meningkatnya percabangan atau pembentukan tunas dan
hilangnya dominansi apikal. Sebaliknya, dominansi apikal pada varian climb
posture sangat tinggi. Pada varian ini, pertunasan terjadi pada pada bagian apikal
sehingga semakin memperkuat dugaan tersebut.
Varian dwarf posture dan climb posture juga berbeda dengan fenotipe
normal dalam hal tinggi tanaman atau ukuran daunnya. Hal ini kemungkinan
berhubungan dengan respon tanaman terhadap giberelin. Taiz dan Zeiger (2003)
menyatakan bahwa terdapat tiga macam mutasi yang mempengaruhi tinggi
tanaman, yaitu gibberellin-insensitive dwarf, gibberellin-deficient mutant, dan
mutan dengan respon gibberellin yang konstitutif. Pada kasus penelitian ini,
varian dwarf posture diduga sebagai mutan gibberellin-insensitive dwarf yang
mirip dengan mutan ga-1 (gibberelin insensitive-1), di mana mutasi terjadi pada
daerah regulatory domain. Varian climb posture diduga sebagai gibberellin-
deficient mutant karena ukurannya tinggi (sifat roset hilang). Mutan demikian
mirip dengan mutan rga (repressor of ga1-3), di mana mutasi terjadi pada daerah
repressor domain sehingga tanaman tetap tinggi walaupun tanpa pemberian GA.
Munculnya varian albino stripes leaf kemungkinan berkaitan dengan
kelainan pada kloroplas. Collins (1951) melaporkan bahwa albino dikendalikan
oleh karakter tunggal. Taiz dan Zeiger (2002) juga mengatakan bahwa sektor

albino berhubungan dengan diferensiasi kloroplas, di mana daun albino tidak
memiliki klorofil sehingga tidak mampu berfotosintesis, namun daun yang
berwarna hijau dapat mengekspor produk fotosintatnya ke sektor albino (Taiz dan
Zeiger 2002) sehingga varian albino stripe leaf dapat bertahan hidup. Terlebih
lagi ketika varian tersebut dalam kondisi heterotrof, sektor albino dapat
memperoleh sumber energi (karbon atau gula) dari media tumbuhnya. Pada
penelitian ini, pertumbuhan varian albino stripe leaf akan lebih baik ketika daerah
hijau daun mendominasi sektor albinonya. Sebaliknya, jika sektor albino
mendominasi sektor hijau maka tanaman tidak dapat bertahan hidup ketika
diaklimatisasi di rumah kaca.
Varian lainnya memperlihatkan kestabilan fenotipe selama isolasi secara
individual, tetapi menghasilkan dua macam atau lebih anakan selama proses
subkultur, yaitu fenotipe yang sama dan berbeda dengan asalnya (Tabel 18).
Fenomena ini jelas mengindikasikan adanya jaringan polisomik di dalam biakan
nenas sehingga menimbulkan khimera yang jelas tampak pada regeneran atau
anakan yang dihasilkannya. Dugaan tersebut semakin kuat dengan ditemukannya
variasi pada populasi tanaman nenas di lapang (Subang), yaitu adanya duri pada
satu atau dua sisi daun, daun klorosis, daun yang berwarna kemerah-merahan, dan
tanaman yang melimpah anakannya. Variasi yang terdapat pada tanaman donor
juga dilaporkan oleh Wakasa (1979). Collins (1936) juga melaporkan adanya
khimera dalam jaringan tanaman nenas yang disebabkan oleh mutasi somatik.
Karakter duri pada daun dilaporkan dikendalikan oleh gen tunggal, demikian pula
untuk karakter daun berlapis lilin (Collins 1951). Karakter daun berduri pada
bagian ujung daun merupakan karakter asli dari tanaman nenas kultivar Smooth
Cayenne. Karakter tersebut bersifat dominan sehingga munculnya karakter daun
berduri diduga sebagai hasil dari mutasi dominan dari alel S menjadi s (Collins
1960) sedangkan karakter waxy dan semi-waxy disebabkan oleh tiadanya atau
berkurangnya trikoma (Collins 1951).


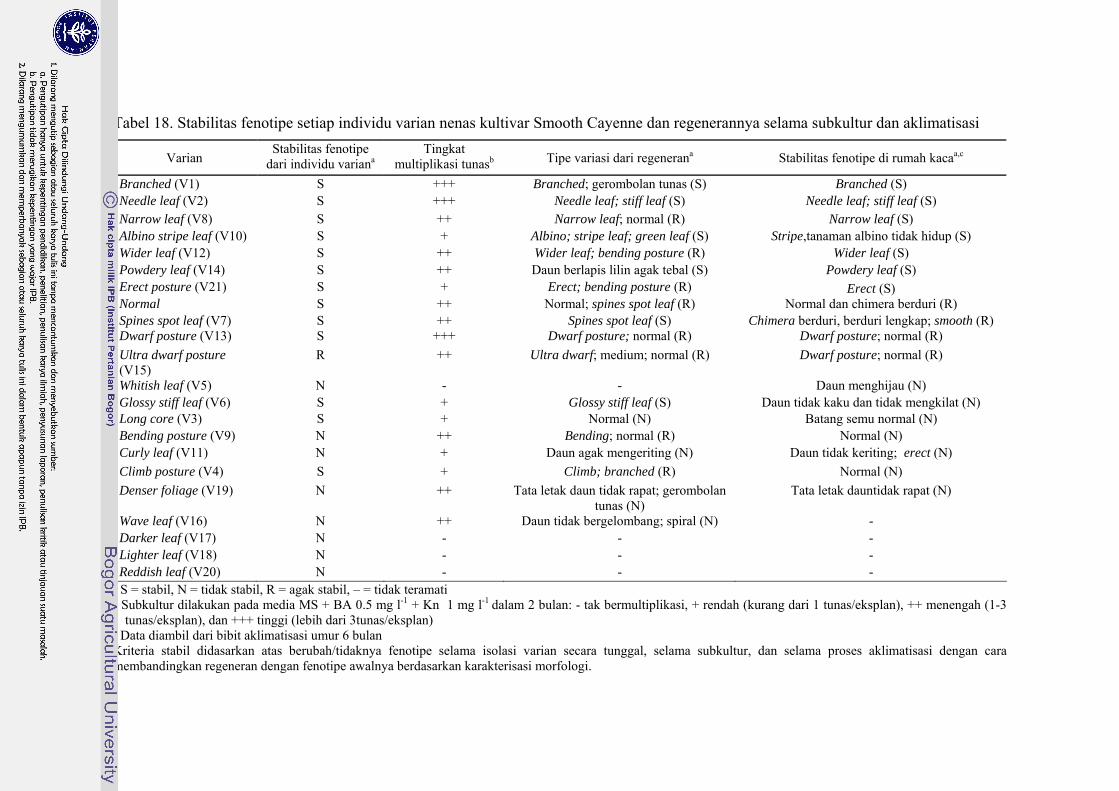
Tabel 18. Stabilitas fenotipe setiap individu varian nenas kultivar Smooth Cayenne dan regenerannya selama subkultur dan aklimatisasi
Varian Stabilitas fenotipe
dari individu varianaTingkat
multiplikasi tunasb Tipe variasi dari regenerana Stabilitas fenotipe di rumah kacaa,c
Branched (V1) S +++ Branched; gerombolan tunas (S) Branched (S) Needle leaf (V2)
S +++ Needle leaf; stiff leaf (S) Needle leaf; stiff leaf (S) Narrow leaf (V8) S ++ Narrow leaf; normal (R) Narrow leaf (S) Albino stripe leaf (V10) S + Albino; stripe leaf; green leaf (S) Stripe,tanaman albino tidak hidup (S) Wider leaf (V12) S ++ Wider leaf; bending posture (R) Wider leaf (S) Powdery leaf (V14) S ++ Daun berlapis lilin agak tebal (S) Powdery leaf (S) Erect posture (V21) S + Erect; bending posture (R) Erect (S) Normal S ++ Normal; spines spot leaf (R) Normal dan chimera berduri (R) Spines spot leaf (V7) S ++ Spines spot leaf (S) Chimera berduri, berduri lengkap; smooth (R) Dwarf posture (V13) S +++ Dwarf posture; normal (R) Dwarf posture; normal (R) Ultra dwarf posture (V15)
R ++ Ultra dwarf; medium; normal (R) Dwarf posture; normal (R)
Whitish leaf (V5) N - - Daun menghijau (N) Glossy stiff leaf (V6) S + Glossy stiff leaf (S) Daun tidak kaku dan tidak mengkilat (N) Long core (V3) S + Normal (N) Batang semu normal (N) Bending posture (V9) N ++ Bending; normal (R) Normal (N) Curly leaf (V11) N + Daun agak mengeriting (N) Daun tidak keriting; erect (N) Climb posture (V4) S + Climb; branched (R) Normal (N)Denser foliage (V19) N ++ Tata letak daun tidak rapat; gerombolan
tunas (N) Tata letak dauntidak rapat (N)
Wave leaf (V16) N ++ Daun tidak bergelombang; spiral (N)
- Darker leaf (V17) N - - -Lighter leaf (V18) N - - - Reddish leaf (V20) N - - -
a S = stabil, N = tidak stabil, R = agak stabil, – = tidak teramati b Subkultur dilakukan pada media MS + BA 0.5 mg l-1 + Kn 1 mg l-1 dalam 2 bulan: - tak bermultiplikasi, + rendah (kurang dari 1 tunas/eksplan), ++ menengah (1-3
tunas/eksplan), dan +++ tinggi (lebih dari 3tunas/eksplan) c Data diambil dari bibit aklimatisasi umur 6 bulan Kriteria stabil didasarkan atas berubah/tidaknya fenotipe selama isolasi varian secara tunggal, selama subkultur, dan selama proses aklimatisasi dengan cara membandingkan regeneran dengan fenotipe awalnya berdasarkan karakterisasi morfologi.


Pada tahap aklimatisasi, 7 varian menunjukkan kestabilan fenotipenya, yaitu
branched shoot, needle leaf, narrow leaf, stripe leaf, wider leaf, powdery leaf dan erect
posture. Dengan demikian, timbul dugaan bahwa variasi yang terjadi pada populasi
tersebut merupakan variasi somaklonal (menyentuh pada tingkat DNA). Oleh karena
itu, dilakukan analisis molekuler untuk mengkonfirmasi dugaan tersebut.
Karakterisasi Molekuler
Isolasi sampel daun menghasilkan DNA dengan kualitas yang cukup baik, di
mana pita DNA yang dihasilkan tidak berbeda dengan lamda (Gambar 38), diperkirakan
mempunyai kuantitas sekitar 100 ng. Kualitas DNA yang tinggi diharapkan akan
menjamin keberhasilan dalam proses PCR.
Pada tahap skrining primer, beberapa primer tidak menghasilkan produk
amplifikasi yang jelas. Oleh karena itu, dipilih 10 primer dengan amplikon yang jelas
dan digunakan untuk analisis pada tahap berikutnya. Hasil karakterisasi molekuler
dengan penanda RAPD menunjukkan tingginya polimorfisme pita DNA yang
dihasilkan dari populasi biakan in vitro nenas yang dikulturkan selama 4 tahun (Gambar
40). Sebanyak 73 pita DNA teramplifikasi dan 70 pita (96%) bersifat polimorfik (Tabel
19). Profil pita-pita DNA dari tiap-tiap varian ditampilkan pada Gambar 40.
L N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
Gambar 39. Kualitas DNA hasil isolasi sampel daun nenas kultivar Smooth Cayenne
dengan metode CTAB: lamda (L), tanaman kontrol dari lapang (N), dwarf posture (V13), albino stripe leaf (V10), narrow leaf (V8), spiness spot leaf (V7), erect posture (V21), wider leaf (V12), powdery leaf (V14), climb posture (V4), needle leaf (V2), dan ultra dwarf posture (V15).
Analisis RAPD pernah digunakan untuk mendeteksi ketepatan genetik dan
variasi somaklonal yang terjadi pada regeneran nenas A. comosus var Queen (Soneji et
al. 2002), A. comosus var Amarelinho (Feuser et al 2003), A. comosus var bracteatus

(Santos et al 2008), dan klon harapan Golden Pineapple (Suminar 2010). Teknik
tersebut mempunyai beberapa kelebihan dibandingkan dengan teknik lainnya, antara
lain sedikitnya jumlah DNA yang diperlukan untuk analisis polimorfisme secara cepat,
tidak diperlukannya informasi tentang sekuens DNA, dan tidak melibatkan
radioaktivitas (Williams et al. 1990).
Tabel 19. Jumlah pita DNA yang teramplifikasi dan yang bersifat polimorfik dalam analisis RAPD populasi biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun
No Primer Jumlah pita yang
teramplifikasi Jumlah pita polimorfik
1 OPA2 8 8 2 OPA3 7 6 3 OPA7 8 8 4 OPA9 7 6 5 OPA13 10 10 6 OPA16 13 13 7 OPA18 8 8 8 OPA19 6 6 9 OPJ11 2 1 10 OPJ13 4 4
Jumlah total pita 73 70
Hasil analisis RAPD menghasilkan dendogram dengan koefisien kemiripan
sebesar 0.32-0.91, di mana varian dan tanaman kontrolnya (normal) memiliki kemiripan
sebesar 0.32-0.61 (Gambar 41). Hal ini membuktikan bahwa variasi somaklonal telah
terjadi pada populasi biakan in vitro nenas umur 4 tahun yang digunakan dalam
penelitian ini. Pengelompokan yang dihasilkan dari analisis tersebut memiliki nilai
korelasi matriks Rohlf yang sangat sesuai (r=0.97), namun pengelompokan tersebut
sangat berbeda dengan pengelompokan yang dihasilkan dari metode karakterisasi secara
morfologi.
Evaluasi stabilitas fenotipe bibit di rumah kaca menunjukkan berubahnya
fenotipe daun halus menjadi berduri (baik khimera maupun berduri lengkap 100%) dan
sebaliknya fenotipe daun berduri menjadi halus. Jumlah tanaman dengan daun yang
berduri lebih banyak daripada yang tidak berduri dan jumlah yang khimera lebih banyak
daripada yang berduri lengkap. Coppens d’Eeckenbrugge dan Sanewski (2003)
menduga bahwa munculnya daun berduri tersebut merupakan karakter yang tidak stabil

dan frekuensinya tinggi. Lebih lanjut dijelaskan bahwa mutasi dari gen dominan daun
tak berduri manjadi gen resesif daun berduri dapat terjadi kapan saja sepanjang
pertumbuhan tanaman, dan tergantung pada tahap perkembangan tanaman ketika mutasi
tersebut terjadi dan tergantung pada posisi sel yang mengalami mutasi sehingga
menimbulkan khimera atau terbentuknya daun berduri secara lengkap.
Fenotipe daun berduri masih menyisakan pertanyaan yang belum terjawab,
mengapa fenotipe tersebut dapat berubah sepanjang pertumbuhan tanaman.
Permasalahan tentang asal-usul jaringan khimera juga masih belum terpecahkan.
Diyakini bahwa stres lingkungan berperan penting dalam perubahan tersebut. Uniknya,
duri pada daun kadang-kadang muncul pada satu sisi dan kadang-kadang muncul pada
dua sisi dengan pola yang tidak beraturan sehingga daun yang tidak berduri bercampur
dengan daun yang berduri pada satu tanaman. Dugaan berikutnya adalah adanya gen
yang mengatur ekspresi karakter tersebut, di mana cis acting element dari gen tersebut
berinteraksi sangat kuat dengan lingkungan. Selanjutnya, diduga kuat adanya elemen
loncat pada tanaman nenas yang diamati. Fenomena tersebut menimbulkan dugaan kuat
akan adanya elemen loncat dalam genom tanaman nenas. Thomson et al. (1998)
melaporkan bahwa sekuens mirip retrotransposon terintegrasi dalam genom tanaman
nenas secara berulang dan bahkan dua sekuens di antaranya tidak memiliki stop kodon
sehingga berpeluang untuk berpindah ke bagian lainnya di dalam genom tanaman nenas
tersebut. Dilaporkan oleh Kumar dan Benetzen (1999) bahwa retrotransposon diaktivasi
oleh stres biotik dan abiotik karena retrotransposon memiliki sekuens cis-acting element
yang terdapat di dalam daerah U3 pada bagian 5’ long terminal repeat (LTR) yang
bersebelahan dengan daerah yang tidak ditranslasikan (untranslated region). Menurut
Kaeppler et al. (2000) serta Okamoto dan Hirochika (2001), variasi metilasi DNA
dihipotesiskan sebagai faktor yang mendasari mutagenesis yang terjadi pada biakan in
vitro yang berkaitan dengan aktivasi elemen loncat.

OPA2 OPA3
OPA9OPA7
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
OPA13 OPA16
OPA18 OPA19
OPJ11 OPJ13
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15 M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15 M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
M N V13 V10 V8 V7 V21 V12 V14 V4 V2 V15
2000 bp
1000 bp 500 bp 250 bp
Gambar 40. Profil DNA varian nenas kultivar Smooth Cayenne, hasil amplifikasi PCR dengan penanda RAPD. M = kb ladder, N = tanaman kontrol (klon Simadu) dari lapang, V2 = needle leaf, V4 = climb posture, V7 = spiness spot leaf, V8 = narrow leaf, V10 = albino stripe leaf, V12 = wider leaf, V13 = dwarf posture, V14 = powdery leaf, V15 = ultradwarf posture, dan V21= erect posture.
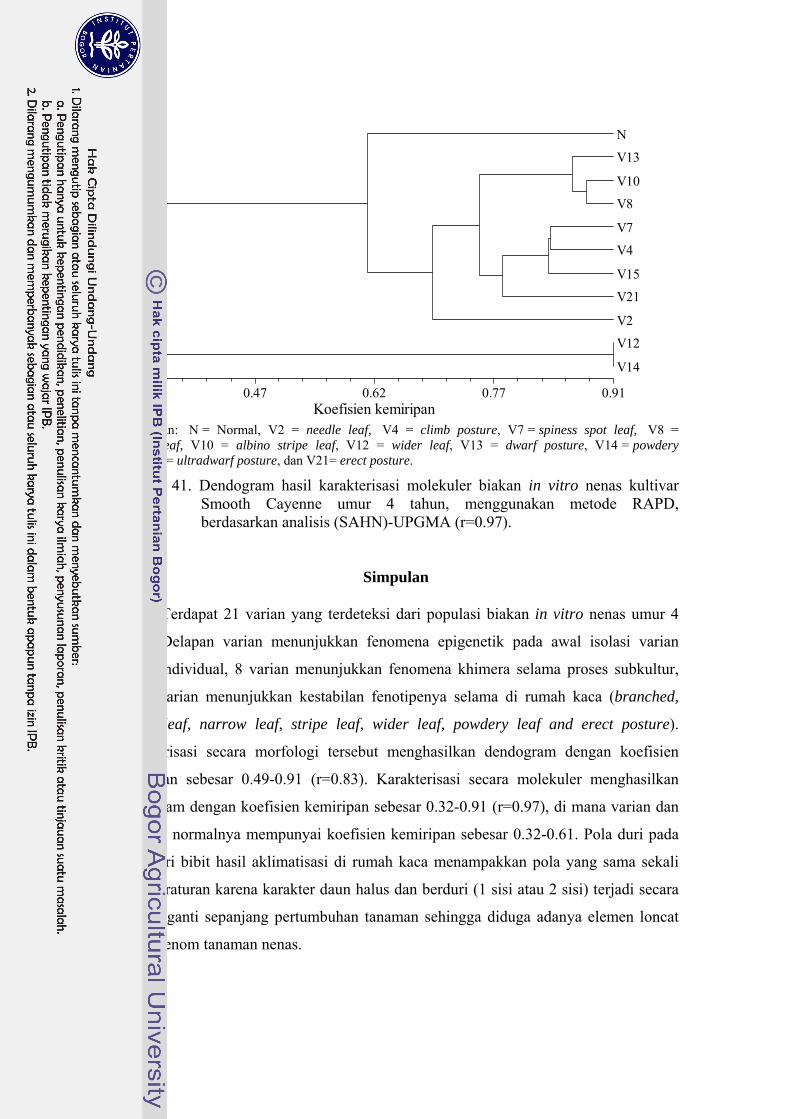
Koefisien kemiripan0.32 0.47 0.62 0.77 0.91
N
V13
V10
V8
V7
V4
V15
V21
V2
V12
V14
Keterangan: N = Normal, V2 = needle leaf, V4 = climb posture, V7 = spiness spot leaf, V8 = narrow leaf, V10 = albino stripe leaf, V12 = wider leaf, V13 = dwarf posture, V14 = powdery leaf, V15 = ultradwarf posture, dan V21= erect posture.
Gambar 41. Dendogram hasil karakterisasi molekuler biakan in vitro nenas kultivar Smooth Cayenne umur 4 tahun, menggunakan metode RAPD, berdasarkan analisis (SAHN)-UPGMA (r=0.97).
Simpulan
Terdapat 21 varian yang terdeteksi dari populasi biakan in vitro nenas umur 4
tahun. Delapan varian menunjukkan fenomena epigenetik pada awal isolasi varian
secara individual, 8 varian menunjukkan fenomena khimera selama proses subkultur,
dan 7 varian menunjukkan kestabilan fenotipenya selama di rumah kaca (branched,
needle leaf, narrow leaf, stripe leaf, wider leaf, powdery leaf and erect posture).
Karakterisasi secara morfologi tersebut menghasilkan dendogram dengan koefisien
kemiripan sebesar 0.49-0.91 (r=0.83). Karakterisasi secara molekuler menghasilkan
dendogram dengan koefisien kemiripan sebesar 0.32-0.91 (r=0.97), di mana varian dan
tanaman normalnya mempunyai koefisien kemiripan sebesar 0.32-0.61. Pola duri pada
daun dari bibit hasil aklimatisasi di rumah kaca menampakkan pola yang sama sekali
tidak beraturan karena karakter daun halus dan berduri (1 sisi atau 2 sisi) terjadi secara
silih berganti sepanjang pertumbuhan tanaman sehingga diduga adanya elemen loncat
dalam genom tanaman nenas.

Daftar Pustaka
Al-Zahim MA, Ford-Lloyd BV, Newbury HJ. 1999. Detection of somaclonal variation in garlic (Allium sativumL.) using RAPD and cytological analysis. Plant CellRep18: 473-477.
Bairu MW, Fennell CW, Staden J van. 2006. The effect of plant growth regulators on somaclonal variation in Cavendish banana (Musa AAA cv. ‘Zelig’). Sci Hort 108:347–351.
Chen WH, Chen TM, Fu YM, Hsieh RM, Chen WS. 1998. Studies on somaclonal variation in Phalenopsis. Plant Cell Rep18: 7-13.
Collins JL. 1951. Notes on the origin, history, and genetic nature of the Cayenne pineapple. Pacific Science 5:3-17
Collins JL. 1960. The Pineapple Botany, Cultivation, and Utilization. Leonard Hill, London
Coppens d’Eeckenbrugge G, Leal F. 2003. Morphology, anatomy and taxonomy. In: Bartholomew, DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany, production and uses. Wallingford: CABI Publishing. pp 13-32.
Eeuwens CJ et al. 2002. Effects of tissue culture conditions during embryoid multiplication on the incidence of “mantled” flowering in clonally propagated oil palm. Plant Cell Tissue Organ Cult 70:311-323.
Feuser S, Meler K, Daquinta M, Guerra MP. 2003. Genotype fidelity of micropropagated pineapple (Ananas comosus) plantlets assessed by isozymes and RAPD markers. Plant Cell Tiss Org Cult 72:221-227.
[IBPGR] International Board for Plant Genetic Resources. 1991. Descriptors for Pineapple. Rome: IBPGR Headquarters.
Joachimiak A, Ilnicki T. 2003. Nuclear morphology, polyploidy, and chromatin elimination in tissue culture of Allium fistulosum L. Acta Societatis Botanicorum Poloniae 77(1):11-17.
Kaeppler SM, Kaeppler HF, Rhee Y. 2000. Epigenetic aspects of somaclonal variation in plants. Plant Mol Biol 43:179-188.
Koornneef M. 1991. Variation and mutant selection in plant cell and tissue culture. In: Jones L, editor. Biotechnological Innovation in Crop Improvement. London: Butterworth-Heinemann Ltd. pp 100-115.
Kumar A, Bennetzen JZ. 1999. Plant retrotransposon. Annu. Rev. Genet 33: 479-532.

Masoud S, Hamta A. 2008. Cytogenetic analysis of somaclonal variation in regenerated plants of berseem clover (Trifolium alexandrium L.). Cryologia 61(4):392-396.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15:473-497.
Noor NM, Clyde MM, Rao VR, Jeevamoney J. 2009. Radiosensitivity and in vitro studies of Citrus suhuiensis. Pp. 17-32. In: IAEA. Induced Mutation in Tropical Fruit Trees. Vienna.
Okamoto H, Hirochika H. 2001. Silencing of transposable elements in plants. Trends in Plant Sci 6(11):527-534.
Podwyszyńska M. 2005. Somaclonal variationin micropropagated tulips based on phenotype observation. Journal of Fruit and Ornamental Plant Research 13:109-122.
Santos MDM, Buso GCS, Torres AC. 2008. Evaluation of genetic variability in micropropagated propagules of ornamental pineapple (Ananas comosus var. bracteatus (Lindley) Coppens and Leal) using RAPD markers. Gen Mol Res 7(4):1097-1105.
Smith MK, Ko H-L, Hamill SD, Sanewski GM, Graham MW. 2003. Biotechnology. In: Bartholomew, DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany: production and uses. Wallingford: CABI Publishing. pp 57-68.
Somsri S, et al. 2009. Development of seedless fruits mutants in citrus including tangerine (C. reticulate) and pummelo (C. grandis) through induced mutations and biotechnology. Pp. 33-40. In: IAEA. Induced Mutation in Tropical Fruit Trees. Vienna.
Soneji JR, Rao PS, Mhatre M. 2002. Suitability of RAPD for analyzing spined and spineless variant regenerants of pineapple (Ananas comosus L., Merr.). Plant Mol Biol Rep 20:307a-307i.
Soniya EV, Banerjee NS. Das MR. 2001. Genetic analysis of somaclonal variation among callus-derived plants of tomato. Current Science 80(9):1213-1215.
Srivastava LM. 2002. Plant Growth and Development. Academic Press. New York. 772p.
Suminar E. 2010. Induksi keragaman genetik dengan mutagen sinar gamma pada nenas (Ananas comosus (L.) Merr.) secara in vitro [seminar]. Sekolah Pascasarjana, Institut Pertanian Bogor. Bogor.
Taiz L, Zeiger E. 2003. Plant Physiology-on line 3rd Ed. Sunderland: Sinauer Associates. doi:10.1093/aob/mcg079.

Thomson KG, Thomas JE, Dietzgen RG. 1998. Retrotransposon-like sequence intregated into genome of pineapple (Ananas comosus). Plant Mol Biol 38:461-465.
Wakasa K. 1979. Variation in the plants differentiated from the tissue culture of pineapple. Japan J Breed 29(1):13-22.
Williams JG, Kubelik AR, Livak KJ, Rafalski JA. 1990. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res 18:6531-6535.
Zhao Y, Grout BWW, Crisp P. 2005. Variation in morphology and disease susceptibility of micropropagated rhubarb (Rheum rhaponticum) PC49, compared to conventional plants. Plant Cell Tissue Organ Cult 82:357-361.
Zucchi MI, Arizono H, Morais VA, Fungaro MHP, Vieira MLC. 2002. Genetic instability of sugarcane plants derived from meristem cultures. Gen Mol Biol 25(1):91-96.

EVALUASI KERAGAMAN BIBIT DARI POPULASI LAMA DAN PEMBENTUKAN POPULASI BARU SERTA DETEKSI DINI KERAGAMAN SOMAKLONAL
Abstrak
Perbanyakan tanaman memerlukan stabilitas sifat yang tinggi. Untuk menghindari tingginya keragaman somaklonal dan untuk mengefektifkan metode mikropropagasi maka diperlukan strategi untuk memproduksi bibit dengan kesamaan identitas genetik yang tinggi. Penelitian ini bertujuan untuk: (1) mengevaluasi keragaman fenotipe bibit hasil ekstraksi dari populasi lama dengan menggunakan penanda morfologi; (2) mengevaluasi keragaman genetik bibit hasil ekstraksi dari populasi lama dengan menggunakan penanda RAPD; (3) mengevaluasi keragaman fenotipe bibit di rumah kaca, dan (4) mendeteksi keragaman somaklonal dari populasi biakan in vitro yang baru. Evaluasi keragaman fenotipe dari bibit di rumah kaca dititik-beratkan pada karakter duri pada daun sedangkan evaluasi ketepatan genetik dan deteksi keragaman somaklonal dilakukan terhadap sampel curah (bulk) dari daun biakan in vitro. Hasil penelitian menunjukkan bahwa metode embriogenesis somatik menghasilkan tingkat keragaman fenotipe yang tinggi (31.1%), diikuti dengan metode organogenesis tak langsung (2%) dan organogenesis langsung (1.6%). Evaluasi keragaman genetik dengan teknik RAPD menunjukkan bahwa dendogram dari bibit hasil ekstraksi dari populasi lama memiliki koefisien kemiripan genetik sebesar 0.32-0.99 di mana varian dengan tanaman kontrolnya memiliki kemiripan sebesar 0.32-0.62. Populasi yang baru memiliki koefisien kemiripan genetik sebesar 0.65-1.0, di mana varian dengan tanaman kontrolnya memiliki kemiripan sebesar 0.65-0.85, mengindikasikan bahwa tingkat keragaman somaklonal menurun. Metode organogenesis langsung menghasilkan bibit berdaun halus sebesar 48%, diikuti dengan metode organogenesis tak langsung (41.4%), dan embriogenesis somatik (27.1%). Disimpulkan bahwa metode organogenesis langsung dapat diterapkan untuk perbanyakan tanaman nenas secara masal.
Kata kunci: Keragaman somaklonal, organogenesis langsung, organogenesis tak langsung, embriogenesis somatik, RAPD
Abstract
Genetic stability is very important in clonally mass propagation. A strategy to avoid the high level of somaclonal variation and to streamline micropropagation method is needed for producing seedlings with the same genetic identity. The objectives of this study were: (1) to evaluate phenotypic variation of seedlings derived from the old in vitro cultures by using morphological marker; (2) to evaluate genetic variation of seedlings derived from the old in vitro cultures by using RAPD markers; (3) to evaluate phenotypic variation of acclimated seedlings in glass house; and (4) to detect somaclonal variation of the new population of in vitro cultures. The evaluation of phenotypic variation of acclimated seedlings was focused on the leaf spininess whereas the evaluation of genetic fidelity was conducted to the bulk of leaves of in vitro cultures. The result showed that somatic embryogenesis yielded the high level of phenotypic variation (31.1%) which yielded 6 stable variants, followed by indirect organogenesis

(2%) which yielded 4 stable variants, and direct organogenesis (1.6%) which yielded 1 stable variant. The molecular analysis revealed that the coefficient of similarity of the seedlings derived from the old population ranged from 0.32 to 0.99 where separated the variants and its control range from 0.32 to 0.62. The new population revealed a higher coefficient of similarity (0.65-1.0) where separated the variants and its control range from 0.65 to 0.85, indicated the decrease of somaclonal variation. The direct organogenesis yielded the higher level of smooth plant proportion (48%), followed by indirect organogenesis (41.4%) and somatic embryogenesis (27.1%). It was concluded that direct organogenesis is high potential to be applied for mass propagation.
Keywords: somaclonal variation, direct organogenesis, indirect organogenesis, somatic embryogenesis, RAPD
Pendahuluan
Teknik kultur in vitro merupakan teknologi alternatif yang tepat untuk produksi
bibit secara masal karena mampu memproduksi bibit dalam jumlah yang besar, dalam
waktu yang singkat, dan dengan keseragaman yang tinggi. Namun demikian, kejadian
off types (penyimpangan atau kelainan) masih menjadi isu yang cukup serius di
Indonesia sehingga pengguna, khususnya pihak swasta masih enggan untuk
memanfaatkan teknik tersebut.
Dari berbagai kajian pustaka, dilaporkan bahwa off types dapat disebabkan oleh
variasi dari tanaman donor (pre-existing variation) (Wakasa 1979), periode in vitro
yang panjang (Koornneef 1991; Masoud and Hamta 2008), zat pengatur tumbuh (Bairu
et al. 2009), subkultur yang frekuentif (Eeuwens et al. 2002), dan tergantung pada
genotipe tanaman (Zucchi et al. 2002). Untuk menghindari tingginya keragaman
somaklonal dan untuk mengefektifkan metode mikropropagasi maka diperlukan strategi
untuk memproduksi bibit dengan kesamaan identitas genetik yang tinggi. Oleh karena
itu, evaluasi stabilitas sifat bibit perlu dilakukan untuk menentukan metode perbanyakan
yang efektif bagi penerapannya secara masal. Dengan demikian, metode deteksi
keragaman somaklonal memegang peranan penting dalam pengujian tersebut.
Keragaman somaklonal dari bibit yang dihasilkan dari teknik kultur in vitro dapat
dideteksi melalaui karakterisasi morfologi (Noor et al. 2009; Somsri et al. 2009 ; Zhao
et al. 2005; Podwyszyńska 2005), sitologi (Al-Zahim et al. 1999), biokimia, fisiologi
(Perez et al. 2011), dan molekuler (Chen et al. 1998; Al-Zahim et al. 1999; Soniya et al.
2001; Perez et al. 2011). Untuk efisiensi waktu, biaya, dan area maka evaluasi stabilitas

sifat tersebut sebaiknya dilakukan secara dini, sejak biakan masih di dalam botol
sebelum tanaman tersebut ditanam di lapang. Wakasa (1979) telah melakukan
pengamatan bibit nenas hasil kultur in vitro 1.5 tahun dalam periode in vitro, 14-22
bulan pada tahap aklimatisasi, dan 1 tahun periode pertumbuhan tanaman. Dengan
demikian, diperlukan waktu sekitar 4 tahun untuk mendeteksi keragaman yang terjadi
pada bibit hasil perbanyakan secara kultur in vitro tersebut.
Pada hasil penelitian sebelumnya telah dilakukan karakterisasi morfologi dan
molekuler biakan in vitro nenas umur 4 tahun pada periode in vitro yang diperbanyak
secara proliferasi tunas. Hasil penelitian tersebut memperlihatkan berbagai macam
varian yang muncul selama proses kultur in vitro tanaman nenas. Hasil penelitian
sebelumnya juga telah memperoleh metode terbaik untuk induksi organogenesis
langsung, organogenesis tak langsung, dan embriogenesis somatik. Metode karakterisasi
morfologi biakan in vitro, primer-primer terpilih, dan bibit nenas yang dihasilkan dari
penelitian tersebut, digunakan pada penelitian ini.
Secara khusus, penelitian ini bertujuan untuk: (1) mengevaluasi keragaman
fenotipe biakan in vitro nenas yang dihasilkan dari metode organogenesis langsung dan
tidak langsung, serta embriogenesis somatic tidak langsung dengan menggunakan
penanda morfologi; (2) mengevaluasi keragaman genetik biakan in vitro nenas yang
dihasilkan dari ketiga metode tersebut dengan menggunakan penanda RAPD; (3)
mengevaluasi stabilitas fenotipe bibit di rumah kaca, dan (4) mendeteksi keragaman
somaklonal dari populasi biakan yang baru. Secara umum, penelitian ini bertujuan
untuk melakukan deteksi dini terhadap keragaman somaklonal dari bibit yang dihasilkan
dari teknik kultur in vitro sekaligus untuk menentukan metode perbanyakan yang paling
efektif, yaitu dengan tingkat multiplikasi tunas yang tinggi namun dengan tingkat
keragaman yang rendah.

Bahan dan Metode
Persiapan Bahan Tanaman
Bahan tanaman yang digunakan adalah biakan in vitro nenas (umur 5 tahun pada
periode in vitro) kultivar Smooth Cayenne klon Simadu dari Subang, Jawa Barat.
Biakan yang dipilih adalah yang mempunyai fenotipe normal. Penelitian ini terdiri atas
5 tahapan, yaitu: (1) induksi organogenesis langsung, (2) induksi organogenesis tidak
langsung, (3) induksi embriogenesis somatik tidak langsung, (4) evaluasi stabilitas
fenotipe, (5) evaluasi ketepatan genetik secara molekuler, dan (6) deteksi dini
keragaman somaklonal dari populasi baru.
Induksi Organogenesis Langsung
Helaian daun in vitro dilepaskan secara hati-hati dari batangnya dengan bantuan
skalpel lalu basal daun diisolasi dengan mata pisau tajam. Eksplan ditumbuhkan pada
media MS cair (25 ml) yang mengandung benzyl adenine (BA) 0.5 mg l-1 (2.2 µM)
dengan penambahan naphthalene acetic acid (NAA) 0.5 mg l-1 (2.7 µM). Botol biakan
tersebut diletakkan di atas shaker dengan kecepatan 70 rpm dan diinkubasikan pada
kondisi terang selama 16 jam dengan intensitas cahaya 800-1000 lux pada suhu 25±2 0C. Dua bulan berikutnya, biakan dipindahkan ke media MS pada dengan penambahan
kinetin (Kn) 1 mg l-1 (4.6 µM) untuk regenerasi dan elongasi tunas. Deteksi keragaman
fenotipe dilakukan dengan menggunakan metode karakterisasi morfologi yang telah
diperoleh pada penelitian sebelumnya. Setelah itu, tunas diinduksi perakarannya pada
media MS padat dengan penambahan indole butyric acid (IBA) 3 mg l-1 dan NAA 1 mg
l-1 (5.4 µM) selama 2-3 minggu untuk pembentukan planlet.
Induksi Organogenesis Tidak Langsung
Tunas in vitro sekitar 4–6 cm dipotong secara longitudinal menjadi dua bagian,
kemudian ditanam secara horizontal pada media induksi kalus dan diinkubasikan pada
kondisi remang-remang (di bawah 500 lux). Induksi kalus dilakukan pada media MS
yang mengandung dichlorophenoxy acetid acid (2,4-D) pada taraf 21, 41, dan 62 µM
(4.6, 9, dan 13.7 mg l-1) dan thidiazuron (TDZ) pada taraf 9 µM (2 mg l-1). Eksplan
diinkubasikan pada suhu 25±2 0C dalam keadaan gelap selama 3 minggu. Setelah

eksplan membesar, helaian daun diisolasi secara individual dan kemudian ditanam pada
media yang sama. Inkubasi juga dilakukan pada kondisi yang sama dengan tahap
sebelumnya. Kalus yang terbentuk selanjutnya dipindahkan ke media regenerasi tunas,
yaitu media MS dengan penambahan Kn 1 mg l-1 (4.65 µM) untuk regenerasi dan
elongasi tunas. Selanjutnya, dilakukan deteksi keragaman fenotipe dan keragaman
genetik sebagaimana dijelaskan pada tahapan sebelumnya. Induksi perakaran dilakukan
pada media MS padat dengan penambahan IBA 3 mg l-1 dan NAA 1 mg l-1 (5.4 µM)
selama 2-3 minggu.
Induksi Embriogenesis Somatik Tidak Langsung
Tunas in vitro sekitar 4–6 cm dipotong secara longitudinal menjadi dua bagian,
kemudian ditanam secara horizontal pada media induksi kalus dan diinkubasikan pada
kondisi remang-remang (di bawah 500 lux). Media induksi kalus adalah media MS yang
mengandung pikloram 21 µM (5 mg l-1) dengan penambahan TDZ 9 µM (2 mg l-1).
Eksplan diinkubasikan pada suhu 25±2 0C dalam keadaan gelap selama 3 minggu.
Setelah eksplan membesar, helaian daun diisolasi secara individual dan kemudian
ditanam pada media yang sama. Kalus yang terbentuk selanjutnya disubkultur pada
media induksi kalus embriogenik, yaitu media MS atau Bac dengan penambahan
senyawa N-organik, yaitu glutamine (Gln) 6.8 µM (1 mg l-1), casein hydrolysate (CH)
500 mg l-1, arginine (Arg) 0.69 mM (120 mg l-1), dan glycine (Gly) 0.027 µM (2 mg l-1).
Kalus diinkubasi dilakukan pada suhu 25±2 0C dalam kondisi terang (800-1000 lux)
selama 3 minggu. Regenerasi dilakukan pada media MS dengan penambahan Kn 1 mg l-
1 (4.65 µM) untuk regenerasi dan elongasi tunas. Selanjutnya, dilakukan deteksi
keragaman fenotipe sebagaimana dijelaskan pada tahapan sebelumnya.
Evaluasi Stabilitas Fenotipe Bibit di Rumah Kaca
Planlet yang telah dihasilkan dari ketiga metode tersebut kemudian diaklimatisasi
di rumah kaca dengan mengggunakan media tanah dan kompos dengan perbandingan
1:1. Setiap klon yang berasal dari 1 basal daun dikelompokkan secara terpisah dari klon
lainnya. Setelah itu, bibit yang berumur 7-11 bulan dievaluasi fenotipenya, khususnya
untuk karakter duri pada daun. Karakter duri pada daun dikategorikan menjadi 3
macam, yaitu halus (jika sedikitnya 10% daun berduri), khimera (jika lebih dari 10%

atau kurang dari 90% daun berduri, baik pada salah satu atau kedua sisinya), dan berduri
lengkap (jika 100% daun berduri hanya pada kedua sisinya).
Evaluasi Ketepatan Genetik secara Molekuler
Deteksi keragaman genetik dilakukan dengan metode yang telah dihasilkan dari
penelitian sebelumnya. Pertama kali, sampel daun dikoleksi dari beberapa tunas atau
embrio somatik (sekitar 10 buah). Seluruh sampel daun dari tiap-tiap metode regenerasi
diekstraksi dengan metode CTAB hingga DNA murni. Deteksi dilakukan dengan
menggunakan metode RAPD, modifikasi metode Soneji et al. (2002). DNA
diamplifikasi dengan mesin PCR. Masing-masing reaksi mengandung 25 ng/l DNA
genomik 1 µl, 1 µl primer (Operon Tech. Alameda, USA) pada 10 ng/µl, 11 µl ddH2O,
dan 12 µl PCR mix, dalam total reaksi 25 µl. Siklus PCR terdiri atas pre-denaturasi pada
suhu 94 0C selama 4 menit, 40 siklus pada 94 0C selama 30 detik (denaturasi); 36 0C
selama 1 menit (penempelan primer) dan 1 menit pada 72 0C (ekstensi); pasca-ekstensi
pada 72 0C selama 5 menit, serta ekstensi akhir pada 40 0C selama 4 menit. Produk PCR
dipisahkan pada agarosa 1.2% (Promega) dan dielektroforesis pada voltase konstan 50
V selama 50 menit, kemudian pita-pita diwarnai dengan ethidium bromide (EtBr) serta
divisualisasi di bawah UV transiluminator. Sebagai standar molekuler digunakan DNA
ladder 1 kb. Sebanyak 10 primer digunakan dalam tahap ini, yaitu primer OPA2, OPA3,
OPA7, OPA9, OPA13, OPA16, OPA18, OPA19, OPJ11, and OPJ13.
Sampel diskor berdasarkan ada dan tidak adanya pita pada ukuran pita yang sama,
di mana 1 berarti ada sedangkan 0 berarti tidak ada. Untuk skrining primer, digunakan
sampel DNA tanaman nenas kultivar Smooth Cayenne Klon Simadu dari lapang dan
varian needle leaf dengan menggunakan 20 primer. Data tersebut dianalisis dengan
menggunakan program SIMQUAL, SAHN, dan TREE dari NTSYS-pc 2.02. Tingkat
kemiripan dihitung dengan menggunakan koefisien Dice. Pengelompokan dilakukan
dengan metode klastering berdasarkan UPGMA.
Deteksi Dini Keragaman Somaklonal dari Populasi Baru
Populasi biakan in vitro yang baru dibentuk dengan cara mengisolasi ulang bahan
tanaman (mahkota) nenas kultivar Smooth Cayenne klon Simadu dan non Simadu.

Regenerasi dilakukan teknik proliferasi tunas dan organogenesis langsung. Deteksi dini
dilakukan secara molekuler dengan menggunakan metode RAPD seperti pada tahapan
sebelumnya, demikian pula dengan analisisnya. Sebagai kontrol, digunakan tanaman
nenas klon Simadu yang berasal dari lapang dan bibit hasil perbanyakan in vitro dengan
fenotipe normal (tepi daun halus atau tidak berduri).
Hasil dan Pembahasan
Ketika planlet normal yang berumur 5 tahun diseleksi dan kemudian basal
daunnya diregenerasikan secara organogenesis langsung, dihasilkan tunas-tunas dengan
fenotipe yang sesuai dengan biakan donornya true-to-type) pada awal pengamatan (2
bulan). Namun demikian, ketika periode inkubasi dilanjutkan hingga 4 bulan maka
terjadi variasi. Hal ini menunjukkan bahwa lama periode in vitro turut menentukan
terjadinya variasi (Tabel 20), sesuai dengan yang dilaporkan oleh Koornneef (1991) dan
Podwyszyńska (2005) yang mengatakan bahwa periode in vitro yang lama dapat
meningkatkan resiko terjadinya mutasi. Varian yang dihasilkan dari metode tersebut
adalah sebanyak 4, yaitu 1 varian bersifat stabil (powdery leaf), 1 varian bersifat agak
stabil (spines spots leaf) dan 2 varian bersifat tidak stabil (curly leaf dan glossy stiff
leaf). Dengan menitik-beratkan pada varian yang bersifat stabil maka secara umum
metode organogenesis langsung pada penelitian ini menghasilkan variasi yang rendah,
yaitu 1.6% (Tabel 21). Hal ini menunjukkan bahwa metode tersebut berpotensi tinggi
untuk diterapkan dalam perbanyakan tanaman nenas secara klonal dan masal.
Keseragaman bibit nenas hasil perbanyakan secara organogenesis langsung ditampilkan
pada Gambar 42A.
Dari metode organogenesis tidak langsung, diperoleh sebanyak 9 varian (Table
20), yaitu 4 varian yang bersifat stabil (branched shoot, narrow leaf, albino stripe leaf,
dan wider leaf), 2 varian yang bersifat agak stabil (spines spot leaf dan dwarf posture)
serta 3 varian yang bersifat tidak stabil (bending posture, glossy stiff leaf dan denser
foliage). Jika dititik-beratkan pada varian yang bersifat stabil maka metode
organogenesis tak langsung tersebut menghasilkan variasi sebesar 2% (Table 21). Bibit
nenas hasil perbanyakan secara organogenesis tidak langsung memperlihatkan
pertumbuhan yang relatif seragam (Gambar 42B).

A
CB
Gambar 42. Keragaman pertumbuhan bibit nenas kultivar Smooth Cayenne yang
dihasilkan dari tiga metode regenerasi: organogenesis langsung (A), organogenesis tak langsung (B), dan embriogenesis somatik (C). Tanda lingkaran menunjukkan pertumbuhan bibit yang lambat.
Berbeda dengan kedua metode sebelumnya, embriogenesis somatik tidak
langsung menghasilkan jenis dan tingkat variasi yang lebih tinggi. Sebanyak 14 varian
dihasilkan dari teknik tersebut (Tabel 20), meliputi 6 varian yang bersifat stabil
(branched, needle leaf, narrow leaf, albino stripe leaf, wider leaf dan powdery leaf), 2
varian yang bersifat agak stabil (ultra dwarf posture dan spiness spot leaf), dan 6 varian
yang bersifat tidak stabil (glossy stiff leaf, bending posture, curly leaf, denser foliage,
lighter leaf, dan piping). Dengan pertimbangan yang sama, teknik embriogenesis
somatik menghasilkan variasi sebesar 31.1% (Tabel 21), labih tinggi dibandingkan
dengan variasi yang terjadi pada biakan donornya (27%). Bibit nenas hasil perbanyakan
secara embriogenesis somatik tidak langsung memperlihatkan pertumbuhan yang
kurang seragam (Gambar 42C).
Untuk mengkonfirmasi variasi yang dihasilkan dari ketiga metode tersebut maka
dilakukan evaluasi fenotipe terhadap bibit nenas di rumah kaca. Hasil pengamatan
menunjukkan bahwa metode organogenesis langsung menghasilkan bibit dengan

karakter daun halus sebesar 48.3% (Tabel 22). Hasil tersebut lebih tinggi daripada yang
dihasilkan dari metode organogenesis tidak langsung yang menghasilkan bibit berdaun
halus sebesar 41.4% (Tabel 23) dan embriogenesis somatik yang menghasilkan bibit
berdaun halus sebesar 27.1% (Tabel 24). Sebaliknya, ketiga metode tersebut secara
berturut-turut menghasilkan daun berduri lengkap sebesar 27.2% (organogenesis
langsung), 42.1% (organogenesis tidak langsung), dan 58.7% (embriogenesis somatik
tidak langsung).
Rendahnya tingkat variasi yang dihasilkan dari metode organogenesis langsung
diduga karena metode tersebut memberikan tekanan atau stres yang paling rendah
terhadap biakan nenas dibandingkan dengan kedua metode regenerasi lainnya. Hal ini
kemungkinan disebabkan oleh tipe dan taraf ZPT yang lebih sederhana (BA dan NAA)
serta periode in vitro yang lebih singkat (4 bulan) untuk meregenerasikan biakan jika
dibandingkan dengan penggunaan 2,4-D dengan rentang waktu yang sama (pada
metode organogenesis tak langsung) atau pikloram dengan rentang waktu yang lebih
lama, yaitu 6.5 bulan (pada metode embriogenesis somatik tidak langsung).


Tabel 20. Proporsi variasi fenotipe dari populasi biakan in vitro nenas kultivar Smooth Cayenne yang dihasilkan dari metode organogenesis (langsung dan tidak langsung), serta embriogenesis somatik tidak langsung
Organogenesis langsungb Organogenesis tidak langsungc
Embriogenesis somatik tidak langsungd
No.
Varian Stabilitas
fenotipea
Jumlah tunas Proporsi (%) Jumlah tunas Proporsi (%) Jumlah embrio Proporsi (%)1 Branched (V1) S 0 0 4 0.3 1 0.3 2 Needle leaf (V2) S 0 0 0 0 3
(V14) S 2 1.6 0 0 14
0 0 0 0 9
3) N 0 0 0 0 0 0
) N 1 0.8 0 0 17
18) - 0 0 0 0 5(V22) - 0 0 0 0 1
0.8 3 Narrow leaf (V8) S 0 0 13 0.8 82 22.84 Albino stripe leaf
(V10) S
0 0 10 0.7 3 0.8 5 Wider leaf (V12) S 0 0 4 0.3 9 2.5 6 Powdery leaf 3.9 7 Normal R 119 93.0 1415 92.4 121 33.68 Spines spot leaf (V7) R 5 3.9 21 1.4 48 13.39 Dwarf posture (V13) R 0 0 6 0.4 0 0 10 Ultra dwarf posture
(V15) R
2.5 11 Glossy stiff leaf (6)
(VN 1 0.8 5 0.3 26 7.2
12 Long core 13 Bending posture (V9)
(V11N 0 0 48 3.1 15 4.2
14 Curly leaf 4.7 15 Denser foliage (V19)
(VN 0 0 5 0.3 6 1.7
16 Lighter leaf 1.4 17
Piping leaf
0.3
Total 128 100 1531 100 360 100a Stabil, N tidak stabil, R agak stabil, and – tidak teramati (biakan sangat rendah tingkat multiplikasi tunasnya) b Organogenesis langsung, diinduksi oleh BA 0.5 mg l-1 dan NAA 0.5 mg l-1
c Organogenesis tak langsung, diinduksi oleh 2,4-D pada taraf 4.6-13.7 mg l-1 d Embriogenesis somatik, diinduksi oleh pikloram dan senyawa N-organik (Gln 1 mg l-1, CHb500 mg l-1, Arg 120 mg l-1, dan Gly 2 mg l-1).

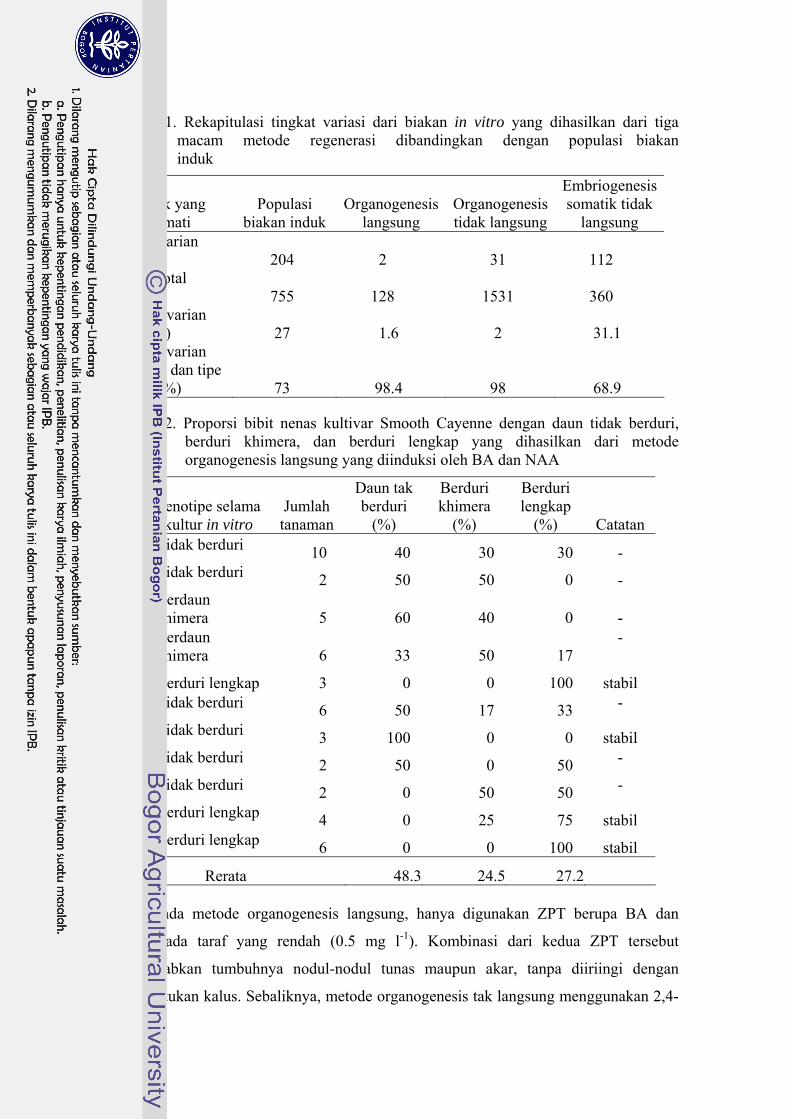
Tabel 21. Rekapitulasi tingkat variasi dari biakan in vitro yang dihasilkan dari tiga macam metode regenerasi dibandingkan dengan populasi biakan induk
Obyek yang
diamati Populasi
biakan induk Organogenesis
langsung Organogenesis tidak langsung
Embriogenesis somatik tidak
langsung Jumlah varian stabil 204 2 31 112 Jumlah total biakan 755 128 1531 360 Proporsi varian stabil (%) 27 1.6 2 31.1 Proporsi varian tak stabil dan tipe normal (%) 73 98.4 98 68.9
Tabel 22. Proporsi bibit nenas kultivar Smooth Cayenne dengan daun tidak berduri, berduri khimera, dan berduri lengkap yang dihasilkan dari metode organogenesis langsung yang diinduksi oleh BA dan NAA
No.klon
Fenotipe selama kultur in vitro
Jumlah tanaman
Daun tak berduri
(%)
Berduri khimera
(%)
Berduri lengkap
(%) Catatan
2 Tidak berduri 10 40 30 30 -
4 Tidak berduri 2 50 50 0 -
7 Berdaun khimera 5 60 40 0 -
8 Berdaun khimera 6 33 50 17
-
13 Berduri lengkap 3 0 0 100 stabil
14 Tidak berduri 6 50 17 33 -
15 Tidak berduri 3 100 0 0 stabil
16 Tidak berduri 2 50 0 50 -
17 Tidak berduri 2 0 50 50 -
18 Berduri lengkap 4 0 25 75 stabil
20 Berduri lengkap 6 0 0 100 stabil
Rerata 48.3 24.5 27.2
Pada metode organogenesis langsung, hanya digunakan ZPT berupa BA dan
NAA pada taraf yang rendah (0.5 mg l-1). Kombinasi dari kedua ZPT tersebut
menyebabkan tumbuhnya nodul-nodul tunas maupun akar, tanpa diiriingi dengan
pembentukan kalus. Sebaliknya, metode organogenesis tak langsung menggunakan 2,4-

D pada taraf yang cukup tinggi (4.6-13.7 mg l-1) untuk induksi kalus. Demikian pula
dengan metode embriogenesis somatik, menggunakan pikloram pada taraf yang cukup
tinggi pula (5 mg l-1) untuk induksi kalus. Kedua macam ZPT tersebut merupakan
agensia herbisida yang pada konsentrasi rendah dapat menginduksi pembentukan kalus.
Kalus merupakan sekelompok sel yang tidak terorganisir dan belum berdiferensiasi
sehingga peluang terjadinya keragaman lebih tinggi daripada struktur yang telah
mengalami diferensiasi, seperti tunas.
Tabel 23. Proporsi bibit nenas kultivar Smooth Cayenne dengan daun tidak berduri, berduri khimera, dan berduri lengkap yang dihasilkan dari metode organogenesis tak langsung yang diinduksi oleh 2,4-D
Klon Jumlah tanaman Daun tidak berduri (%)
Berduri khimera (%)
Daun berduri lengkap %)
1 10 20 0 80
2 10 22 11 67
3 5 60 20 20
4 10 60 10 30
5 8 38 50 12
6 10 30 10 60
7 10 60 0 40
Rerata 63 41.4 14.4 44.1
Hasil evaluasi keragaman fenotipe selanjutnya dikonfirmasi dengan secara
molekuler. Dari berbagai teknik molekuler yang tersedia, metode RAPD mempunyai
kelebihan dibandingkan dengan teknik lainnya, antara lain sedikitnya jumlah DNA yang
diperlukan untuk analisis polimorfisme secara cepat, tidak diperlukannya informasi
tentang sekuens DNA, dan tidak melibatkan radioaktivitas (Williams et al. 1990).
Metode tersebut pernah digunakan untuk mendeteksi ketepatan genetik dan variasi
somaklonal yang terjadi pada regeneran nenas A. comosus var Queen (Soneji et al.
2002), A. comosus var Amarelinho (Feuser et al 2003), A. comosus var bracteatus
(Santos et al 2008), dan klon harapan Golden Pineapple (Suminar 2010), bahkan juga

untuk mendeteksi stabilitas genetik hasil mikropropagasi tanaman tebu yang memiliki
tingkat ploidi yang tinggi dan kromosom yang mosaik (Tawar et al 2008).
Tabel 24. Proporsi bibit nenas kultivar Smooth Cayenne dengan daun tidak berduri, berduri khimera, dan berduri lengkap yang dihasilkan dari metode embriogenesis somatik yang diinduksi oleh pikloram
Klon Jumlah tanaman Daun tidak berduri (%)
Berduri khimera (%)
Daun berduri lengkap %)
1 7 14 28 58 2 5 40 20 40 3 4 50 0 50 4 3 67 0 33 5 3 67 0 33 6 3 0 0 100 7 2 0 0 100 8 3 0 0 100 9 2 0 50 50 10 3 33 33 33
Rerata 35 27.1 13.1 59.7
Pada penelitian ini, profil DNA yang dihasilkan dari metode RAPD dengan
menggunakan 10 primer menunjukkan polimorfisme yang cukup tinggi (Gambar 43).
Analisis pola pita DNA menghasilkan dendogram dengan rentang kemiripan genetik
sebesar 0.34-0.99 (r=0.98). Gambar 44 menunjukkan bahwa bibit yang dihasilkan dari
metode embriogenesis somatik yang diinduksi oleh pikloram mempunyai kemiripan
genetik yang paling rendah (0.34) dibandingkan dengan bibit yang dihasilkan dari
metode organogenesis langsung, organogenesis tak langsung dan embriogenesis
somatik yang diinduksi oleh 2,4-D (0.61). Rani dan Raina (2000) mengatakan bahwa
bibit yang dihasilkan dari mikropropagasi yang melalui fase kalus akan mempunyai
ketidak-stabilan genetik. Selain melalui fase kalus, teknik embriogenesis somatik dalam
penelitian ini memerlukan waktu yang lama sehingga memperbesar peluang terjadinya
perubahan genetik selama kultur in vitro. Polimorfisme yang cukup tinggi (35/84 pita)
juga dilaporkan oleh Soniya et al (2001) pada tanaman tomat yang dikulturkan dengan
menggunakan pikloram.

OPA9 OPA13
OPA2 OPA7
OPA16 OPA18
OPA3 OPA9
OPJ11 OPJ13M S BN D21 D41 D62 ESD ESP S BN D21 D41 D62 ESD ESP M
M S BN D21 D41 D62 ESD ESP S BN D21 D41 D62 ESD ESP M
M S BN D21 D41 D62 ESD ESP S BN D21 D41 D62 ESD ESP M
M S BN D21 D41 D62 ESD ESP S BN D21 D41 D62 ESD ESP M
M S BN D21 D41 D62 ESD ESP S BN D21 D41 D62 ESD ESP M
2000 bp
1000 bp 500 bp 250 bp
Gambar 43. Profil DNA biakan nenas kultivar Smooth Cayenne yang diekstraksi dari biakan induk: M = kb ladder, S = klon Simadu dari lapang, BN = biakan hasil organogenesis langsung, D21, D41 dan D62 = biakan hasil organogenesis tidak langsung masing-masing diinduksi oleh 2,4-D 21, 41 dan 62 µM, ESD serta ESP = biakan hasil embriogenesis somatik masing-masing diinduksi oleh 2,4-D dan pikloram. Panah mencotohkan pita-pita yang polimorfis.
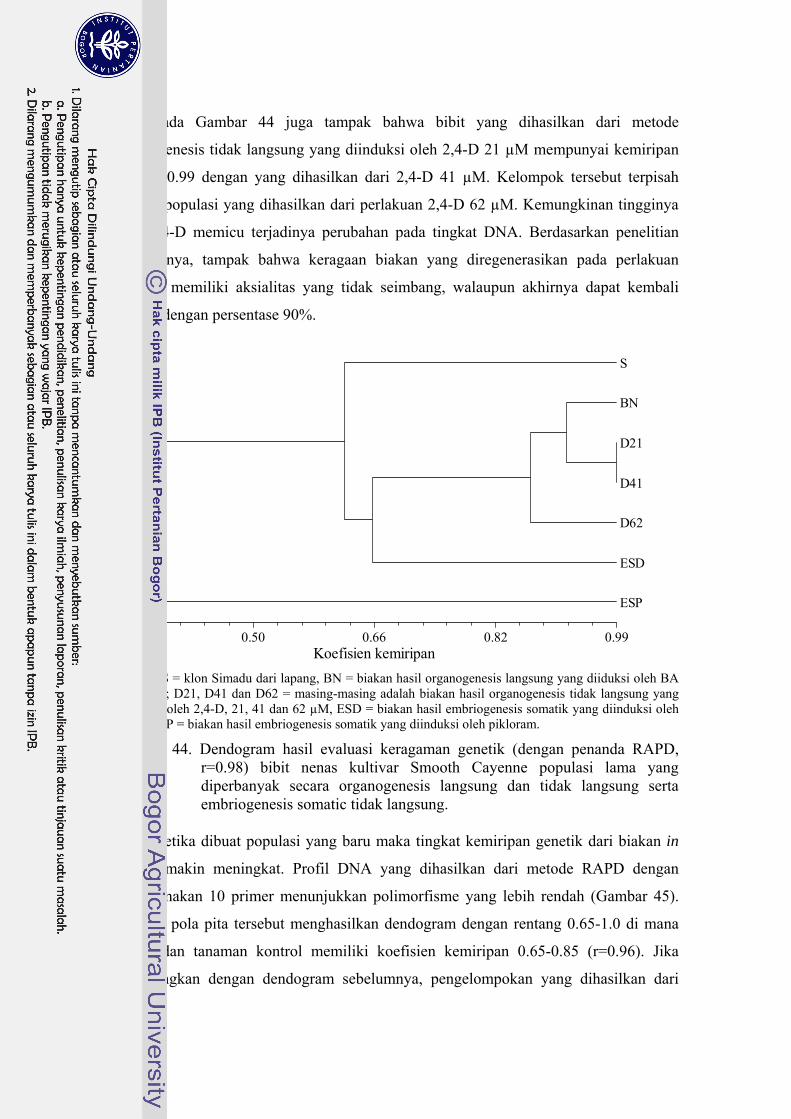
Pada Gambar 44 juga tampak bahwa bibit yang dihasilkan dari metode
organogenesis tidak langsung yang diinduksi oleh 2,4-D 21 µM mempunyai kemiripan
sebesar 0.99 dengan yang dihasilkan dari 2,4-D 41 µM. Kelompok tersebut terpisah
dengan populasi yang dihasilkan dari perlakuan 2,4-D 62 µM. Kemungkinan tingginya
taraf 2,4-D memicu terjadinya perubahan pada tingkat DNA. Berdasarkan penelitian
sebelumnya, tampak bahwa keragaan biakan yang diregenerasikan pada perlakuan
tersebut memiliki aksialitas yang tidak seimbang, walaupun akhirnya dapat kembali
normal dengan persentase 90%.
Koefisien kemiripan0.34 0.50 0.66 0.82 0.99
S
BN
D21
D41
D62
ESD
ESP
Catatan: S = klon Simadu dari lapang, BN = biakan hasil organogenesis langsung yang diiduksi oleh BA dan NAA; D21, D41 dan D62 = masing-masing adalah biakan hasil organogenesis tidak langsung yang diinduksi oleh 2,4-D, 21, 41 dan 62 µM, ESD = biakan hasil embriogenesis somatik yang diinduksi oleh 2,4-D, ESP = biakan hasil embriogenesis somatik yang diinduksi oleh pikloram.
Gambar 44. Dendogram hasil evaluasi keragaman genetik (dengan penanda RAPD, r=0.98) bibit nenas kultivar Smooth Cayenne populasi lama yang diperbanyak secara organogenesis langsung dan tidak langsung serta embriogenesis somatic tidak langsung.
Ketika dibuat populasi yang baru maka tingkat kemiripan genetik dari biakan in
vitro semakin meningkat. Profil DNA yang dihasilkan dari metode RAPD dengan
menggunakan 10 primer menunjukkan polimorfisme yang lebih rendah (Gambar 45).
Analisis pola pita tersebut menghasilkan dendogram dengan rentang 0.65-1.0 di mana
varian dan tanaman kontrol memiliki koefisien kemiripan 0.65-0.85 (r=0.96). Jika
dibandingkan dengan dendogram sebelumnya, pengelompokan yang dihasilkan dari

dendogram tersebut bergeser ke kanan yang mengindikasikan bahwa tingkat kemiripan
genetik dari populasi baru ini lebih tinggi dibandingkan dengan populasi yang lama (4
tahun).
Gambar 46 memperlihatkan bahwa klon non Simadu mempunyai perbedaan
genetik sebesar 0.16 dengan klon bukan Simadu. Hal ini menunjukkan bahwa kedua
klon tersebut mempunyai komposisi genetik yang berbeda. Kemungkinan klon Simadu
merupakan mutan dari klon non Simadu, di mana semua ciri-ciri tanamannya mirip
kecuali pada sifat buah yang dihasilkan.
Hasil penelitian juga menunjukkan bahwa bibit hasil organogenesis langsung
yang dibentuk dari dari populasi lama mempunyai kesamaan genetik (koefisien
kemiripan 100%) dengan klon Simadu yang berasal dari lapang. Walaupun memiliki
kesamaan genetik, bibit yang dihasilkan dari teknik organogenesis langsung tersebut
diduga belum tentu akan menghasilkan buah nenas Simadu karena sifat buah tersebut
merupakan fenotipe, yaitu hasil interaksi antara genotipe dan lingkungan. Sebagaimana
telah dikaji pada Bab sebelumnya, interaksi genotipe dan lingkungan sangat tinggi
dalam mempengaruhi fenotipe (morfologi) tanaman nenas. Terlebih lagi, beberapa
kriteria buah nenas Simadu (daging buah berwarna kuning oranye, rasanya sangat
manis, memiliki kadar air yang tinggi, dan mengeluarkan bunyi ketukan yang sangat
khas) merupakan indigenous knowledge dari petani setempat (Subang) dan masih belum
dapat dijelaskan secara scientific sehingga prediksinya akan semakin sulit dilakukan.
Selain itu, metode deteksi yang digunakan dalam penelitian ini adalah metode RAPD
yang bersifat dominant marker sehingga tidak mampu mendeteksi perbedaan antara
genotipe homosigot dominan dengan genotipe heterosigot. Jumlah primer yang
digunakan dalam analisis ini juga terbatas, yaitu hanya 10 primer.

OPA2 OPA7
OPA3 OPA19
OPA9 OPA13
OPA16 OPA18
OPJ13 OPJ11
M S NS NI PS PNS OS ONS S NS NI PS PNS OS ONS M
M S NS NI PS PNS OS ONS S NS NI PS PNS OS ONS M
M S NS NI PS PNS OS ONS S NS NI PS PNS OS ONS M
M S NS NI PS PNS OS ONS S NS NI PS PNS OS ONS M
M S NS NI PS PNS OS ONS S NS NI PS PNS OS ONS M
2000 bp 1000 bp
500 bp 250 bp
Gambar 45. Profil DNA dari populasi baru biakan in vitro nenas kultivar Smooth Cayenne: M= kb ladder, S = klon Simadu dari lapang, NS = klon non Simadu dari lapang, NI = bibit berfenotipe normal hasil organogenesis langsung dari populasi lama, PS = biakan klon Simadu hasil proliferasi tunas, PNS = biakan klon non Simadu hasil proliferasi tunas, OS = biakan klon Simadu hasil organogenesis langsung, ONS = biakan klon non Simadu hasil organogenesis langsung. Panah mencontohkan pita yang polimorfis.

Koefisien kemiripan0.32 0.49 0.66 0.83 1.00
S
NI
NS
PNS
PS
OS
ONS
Catatan: S = klon Simadu dari lapang, NS = klon non Simadu dari lapang, PS = biakan klon Simadu hasil proliferasi tunas, PNS = biakan klon non Simadu hasil proliferasi tunas, OS = biakan klon Simadu hasil organogenesis langsung, ONS = biakan klon non Simadu hasil organogenesis langsung, NI = bibit berfenotipe normal hasil organogenesis langsung dari populasi lama.
Gambar 46. Dendogram hasil deteksi dini keragaman somaklonal biakan in vitro nenas kultivar Smooth Cayenne dari populasi baru (dengan penanda RAPD, r=0.96).
Simpulan
Metode embriogenesis somatik menghasilkan tingkat keragaman fenotipe yang
tinggi (31.1%), diikuti dengan metode organogenesis tak langsung (2%) dan
organogenesis langsung (1.6%). Evaluasi keragaman genetik dengan teknik RAPD
menunjukkan bahwa bibit hasil ekstraksi dari populasi lama memiliki koefisien
kemiripan genetik sebesar 0.32-0.99 di mana varian dan tanaman kontrol memiliki
koefisien kemiripan sebesar 0.32-0.62. Populasi yang baru memiliki koefisien
kemiripan sebesar 0.65-1.0, di mana varian dan tanaman kontrol memiliki koefisien
kemiripan sebesar 0.65-0.85. Metode organogenesis langsung menghasilkan bibit
berdaun halus sebesar 48%, diikuti dengan metode organogenesis tak langsung (41.4%),
dan embriogenesis somatik (27.1%). Disimpulkan bahwa metode organogenesis
langsung dapat diterapkan untuk perbanyakan tanaman nenas secara masal.

Daftar Pustaka
Al-Zahim MA, Ford-Lloyd BV, Newbury HJ. 1999. Detection of somaclonal variation in garlic (Allium sativumL.) using RAPD and cytological analysis. Plant CellRep18: 473-477.
Bairu MW, Fennell CW, Staden J van. 2006. The effect of plant growth regulators on somaclonal variation in Cavendish banana (Musa AAA cv. ‘Zelig’). Sci Hort 108:347–351.
Chen WH, Chen TM, Fu YM, Hsieh RM, Chen WS. 1998. Studies on somaclonal variation in Phalenopsis. Plant Cell Rep18: 7-13.
Eeuwens CJ et al. 2002. Effects of tissue culture conditions during embryoid multiplication on the incidence of “mantled” flowering in clonally propagated oil palm. Plant Cell Tissue Organ Cult 70:311-323.
Feuser S, Meler K, Daquinta M, Guerra MP. 2003. Genotype fidelity of micropropagated pineapple (Ananas comosus) plantlets assessed by isozymes and RAPD markers. Plant Cell Tiss Org Cult 72:221-227
Koornneef M. 1991. Variation and mutant selection in plant cell and tissue culture. In: Jones L, editor. Biotechnological Innovation in Crop Improvement. London: Butterworth-Heinemann Ltd. pp 100-115.
Masoud S, Hamta A. 2008. Cytogenetic analysis of somaclonal variation in regenerated plants of berseem clover (Trifolium alexandrium L.). Cryologia 61(4):392-396.
Noor NM, Clyde MM, Rao VR, Jeevamoney J. 2009. Radiosensitivity and in vitro studies of Citrus suhuiensis. Pp. 17-32. In: IAEA. Induced Mutation in Tropical Fruit Trees. Vienna.
Podwyszyńska M. 2005. Somaclonal variationin micropropagated tulips based on phenotype observation. Journal of Fruit and Ornamental Plant Research 13:109-122.
Rani V, Raina SN. 2000. Genetic fidelity of organized meristem-derived micropropagated plants: a critical Reappraisal. In Vitro Cell Dev Biol_Plant 36:319-330.
Santos MDM, Buso GCS, Torres AC. 2008. Evaluation of genetic variability in micropropagated propagules of ornamental pineapple (Ananas comosus var. bracteatus (Lindley) Coppens and Leal) using RAPD markers. Gen Mol Res 7(4):1097-1105.

Somsri S, et al. 2009. Development of seedless fruits mutants in citrus including tangerine (C. reticulate) and pummelo (C. grandis) through induced mutations and biotechnology. Pp. 33-40. In: IAEA. Induced Mutation in Tropical Fruit Trees. Vienna.
Soneji JR, Rao PS, Mhatre M. 2002. Suitability of RAPD for analyzing spined and spineless variant regenerants of pineapple (Ananas comosus L., Merr.). Plant Mol Biol Rep 20:307a-307i
Soniya EV, Banerjee NS. Das MR. 2001. Genetic analysis of somaclonal variation among callus-derived plants of tomato. Current Science 80(9):1213-1215.
Suminar E. 2010. Induksi keragaman genetik dengan mutagen sinar gamma pada nenas (Ananas comosus (L.) Merr.) secara in vitro [seminar]. Sekolah Pascasarjana, Institut Pertanian Bogor. Bogor.
Tawar PN et al. 2008. An assessment of somaclonal variation in micropropagated plants of sugarcane by RAPD markers. Sugar Tech 10(2):124-127.
Wakasa K. 1979. Variation in the plants differentiated from the tissue culture of pineapple. Japan J Breed 29(1):13-22.
Williams JG, Kubelik AR, Livak KJ, Rafalski JA. 1990. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res 18:6531-6535
Zhao Y, Grout BWW, Crisp P. 2005. Variation in morphology and disease susceptibility of micropropagated rhubarb (Rheum rhaponticum) PC49, compared to conventional plants. Plant Cell Tissue Organ Cult 82:357-361
Zucchi MI, Arizono H, Morais VA, Fungaro MHP, Vieira MLC. 2002. Genetic instability of sugarcane plants derived from meristem cultures. Gen Mol Biol 25(1):91-96.

PEMBAHASAN UMUM
Tanaman nenas merupakan tanaman yang unik karena merupakan tanaman CAM
fakultatif, artinya berperilaku sebagai tanaman C3 ketika lingkungannya optimal,
namun berperilaku sebagai tanaman CAM ketika lingkungannya suboptimal. Kelebihan
lain dari tanaman ini adalah memiliki akar terestrial, bukan akar tempel (epifit) seperti
tanaman CAM pada umumnya. Wattimena (2012) menggolongkan tanaman C3-CAM
sebagai tanaman masa depan yang prospektif dikembangkan secara luas dalam rangka
revitalisasi tanaman pertanian untuk mendukung era Revolusi Hijau Lestari (Wattimena
2012), terutama pada lahan-lahan marginal yang kurang sesuai diterapkan untuk
tanaman pangan sehingga produktivitas lahan pertanian dapat ditingkatkan.
Suatu tinjauan melaporkan bahwa tanah liat dengan kandungan bahan organik
yang tinggi dengan pH 4.5-6.5 merupakan kondisi yang ideal bagi budidaya tanaman
nenas. Dengan demikian, diperkirakan bahwa tanaman tersebut akan sesuai
dikembangkan di lahan-lahan gambut yang tersebar di berbagai wilayah di Indonesia.
Sagiman (2007) melaporkan bahwa tanaman nenas merupakan tanaman yang sangat
cocok ditanam pada tanah gambut karena dapat memberikan hasil yang baik sekalipun
tidak dilakukan pemupukan dan pengendalian hama penyakit. Nenas bahkan
merupakan hasil komoditas pertanian tertinggi pada lahan gambut di Malaysia dengan
hasil mencapai 40-48 ton/ha/th mengalahkan ubi kayu (35-49 ton/ha/th) dan sawit (20-
25 ton/ha/th).
Penelitian ini mencakup studi morfogenesis tanaman nenas secara in vitro yang
meliputi organogenesis (langsung dan tidak langsung) serta embriogenesis somatik.
Teknik tersebut digabungkan dengan teknik enkapsulasi untuk pembentukan benih
sintetik yang dapat diterapkan untuk produksi bibit maupun konservasi in vitro.
Selanjutnya, biakan dan bibit nenas yang dihasilkan dari ketiga metode regenerasi
tersebut dievaluasi stabilitas sifatnya secara morfologi maupun molekuler. Deteksi dini
keragaman somaklonal juga dilakukan terhadap populasi biakan in vitro yang baru.
Dengan demikian, dapat ditentukan metode regenerasi tanaman nenas yang efektif,
yaitu dengan tingkat multiplikasi yang tinggi, dalam waktu yang singkat, dan dengan
tingkat keragaman yang rendah.

Berdasarkan studi morfogenesis eksplan pada Bab I, diketahui bahwa
morfogenesis eksplan pada media padat menghasilkan persentase tunas sebesar 30%
dan jumlah tunas sebanyak 10 tunas/eksplan. Metode organogenesis langsung tersebut
memiliki potensi produksi bibit yang tinggi, yang diformulasikan dengan Y = M x S x
(D x B x T)n x K x A, di mana Y potensi produksi bibit, M jumlah mahkota
(diasumsikan 1 buah), S jumlah stok induk in vitro (diasumsikan 10 mata
tunas/mahkota), D jumlah eksplan (diasumsikan 7 basal daun/tunas), B persentase
bertunas (diasumsikan 30%), T jumlah tunas/eksplan, N jumlah subkultur (diasumsikan
3 kali), K persentase bebas kontaminasi (diasumsikan 80%), A persentase keberhasilan
aklimatisasi (diasumsikan 80%), maka Y = 1 x 10 x (7 x 30% x 10 )3 x 80% x 80% =
59.270 bibit/tahun. Dibandingkan dengan metode tersebut, organogenesis tidak
langsung memiliki potensi produksi bibit yang lebih tinggi, yaitu Y = M x S x (D x B x
T)n x K x A, di mana Y potensi produksi bibit, M jumlah mahkota (diasumsikan 1
buah), S jumlah stok induk in vitro (diasumsikan 10 mata tunas/mahkota), D jumlah
eksplan (diasumsikan 7 basal daun/tunas), B persentase bertunas (diasumsikan 80%), T
jumlah tunas/eksplan, N jumlah subkultur (diasumsikan 2 kali), K persentase bebas
kontaminasi (diasumsikan 80%), A persentase keberhasilan aklimatisasi (diasumsikan
80%), maka Y = 1 x 10 x (7 x 80% x 25 )2 x 80% x 80% = 125.440 bibit/tahun. Metode
embriogenesis somatik memiliki potensi produksi bibit yang lebih rendah daripada
kedua metode tersebut, yaitu Y = M x S x D x C x E x R x T x K x A, di mana Y
potensi produksi bibit, M jumlah mahkota (diasumsikan 1 buah), S jumlah stok induk in
vitro (diasumsikan 10 mata tunas/mahkota), D jumlah eksplan (diasumsikan 7 basal
daun/tunas), C persentase pembentukan kalus (diasumsikan 90%), E persentase
pembentukan kalus embriogenik (diasumsikan 100%), R persentase regenerasi
(diasumsikan 100%), T jumlah embrio dewasa (diasumsikan 30 embrio/eksplan dalam 4
bulan), K persentase bebas kontaminasi (diasumsikan 80%), A persentase keberhasilan
aklimatisasi (diasumsikan 80%), maka Y = 1 x 10 x 7 x 90% x 100% x 30 x 80% x 80%
= 1.209 bibit/tahun. Potensi produksi bibit dari ketiga macam metode regenerasi
tersebut dapat ditingkatkan dengan cara memperbanyak bahan tanaman induk (lebih
dari 1 mahkota).
Tingginya potensi produksi bibit perlu diiringi dengan rendahnya tingkat
keragaman somaklonal dari bibit yang dihasilkan. Oleh karena itu, studi evaluasi

kestabilan sifat bibit menjadi sangat penting dilakukan dalam penentuan metode yang
terbaik untuk diterapkan.
Pada studi embriogenesis somatik, penggunaan pikloram terbukti lebih baik
daripada 2,4-D karena pikloram dapat menginisiasi pembentukan sel embriogenik
secara langsung sedangkan penggunaan 2,4-D secara tunggal gagal membentuk sel
embriogenik, bahkan tidak mempengaruhi polaritas sel. Pada embriogenesis nenas,
jaringan embriogenik lebih mudah terbentuk daripada kalus embriogenik karena kalus
embriogenik tersebut terbentuk dalam kelompok sel yang saling berhubungan satu
dengan lainnya. Terdapat dua macam jaringan embriogenik yang dijumpai pada
penelitian ini, yaitu ETs (strukturnya semi friabel) dan FETs (strukturnya friabel). Pada
perlakuan 2,4-D, FETs dapat terbentuk ketika kalus diberi perlakuan senyawa N-
organik sedangkan pada perlakuan pikloram, struktur ETs dapat terbentuk hanya dengan
memindahkan kalus ke media MS tanpa ZPT sehingga embrio somatik dari perlakuan
pikloram lebih cepat terbentuk dan berkembang dewasa. Penggunaan pikloram 62 µM
tidak memperlihatkan penghambatan pertumbuhan kalus, namun penggunaan 2,4-D 62
µM berpengaruh secara nyata dalam menghambat pembentukan kalus dan menimbulkan
nekrosis. Hal ini mengindikasikan bahwa fitotoksisitas 2,4-D lebih tinggi daripada
pikloram terhadap jaringan nenas. Selain itu, jumlah embrio somatik yang dihasilkan
dari perlakuan pikloram lebih banyak (30 embrio/eksplan dalam waktu 4 bulan)
daripada yang diperoleh dari perlakuan 2,4-D (17 embrio/eksplan dalam waktu 6.5
bulan). Dengan demikian, dapat disimpulkan bahwa auksin pikloram lebih sesuai untuk
induksi embriogenesis somatik tanaman nenas daripada auksin 2,4-D dan NAA. Hasil
penelitian ini lebih baik daripada hasil penelitian sebelumnya yang menghasilkan 58%
formasi kalus nenas kultivar Phuket (Sripaoraya et al. 2003) dan 55% untuk kultivar
Smooth Cayenne (Firoozabady dan Moy 2004).
Jika NAA berhasil membentuk struktur proembrio oktan maka embriogenesis
somatik yang diinduksi oleh 2,4-D dan pikloram mampu membentuk struktur embrio
bipolar (embrio somatik). Tahapan lengkap embriogenesis somatik nenas dapat
diketahui dengan mengurutkan setiap tahapan yang diperoleh dari ketiga percobaan
tersebut. Secara keseluruhan, tahapan embriogenesis somatik tidak langsung pada nenas
kultivar Smooth Cayenne (Gambar 47) diawali dengan inisiasi sel embriogenik yang

berbentuk isodiametrik yang terkumpul sebagai struktur PEM, lalu masing-masing sel
tersebut mengalami polarisasi, pembelahan asimetris secara transversal dari 2 sel hingga
6 sel, pembelahan longitudinal hingga terbentuk proembrio 8 sel (oktan), dan
dilanjutkan dengan perkembangan embrio somatik yang didahului dengan pembentukan
struktur globular, skutelar, koleoptilar, dan diakhiri dengan perkecambahan embrio
membentuk planlet yang secara visual tampak sebagai struktur bipolar (mengandung
tunas dan akar). Tahapan tersebut sangat berbeda dengan tahapan organogenesis yang
dirangkumkan pada Gambar 48. Berdasarkan pengamatan mikroskopis dan analisis
histologi maka dapat diketahui bahwa embrio somatik nenas berkembang dari sel
tunggal (unicellular origin) sehingga berpotensi besar untuk diterapkan dalam rakayasa
genetika.
Morfogenesis eksplan yang mantap melalui metode proliferasi tunas,
organogenesis dan embriogenesis somatik diperlukan dalam pembentukan benih
sintetik. Redenbaugh (1993) menjelaskan bahwa benih sintetik dapat dibentuk dari
jaringan meristematik, seperti embrio somatik, tunas aksilar, tunas terminal, dan nodus.
Panaia (2008) menggolongkan benih sintetik menjadi tipe basah (dehydrated) dan tipe
kering (desiccated), baik dengan atau tanpa enkapsulasi. Pada penelitian ini, diterapkan
benih sintetik tipe basah dengan menggunakan hidrogel berupa Na-alginat 3%.

A B C D
E F
LJ K
G H
I
Gambar 47. Tahapan lengkap embriogenesis somatik nenas kultivar Smooth Cayenne: sel-sel isodiamterik dalam PEM, sitoplasma pekat, nukleus besar, dan menyerap warna (A), polarisasi sel (B), pembelahan asimetris (C), 2 sel yang terdiri atas sel apikal dan basal hasil pembelahan secara transversal (D), 4 sel kuadran (E), 6 sel hasil pembelahan secara longitudinal (F), 8 sel atau oktan (G), embrio globular (H), embrio skutelar (I), embrio koleoptilar (J), embrio somatik dewasa (K), dan planlet yang siap diaklimatisasi (L).

A B C
E F G
D
H
Gambar 48. Tahapan lengkap organogenesis nenas kultivar Smooth Cayenne: sel-sel non embriogenik yang berbentuk tidak beraturan, sitoplasma tidak pekat, vakuola besar, dan kurang menyerap warna (A), sel tidak terpolarisasi (B), 2 sel hasil pembelahan simetris (C), 4 sel (D) dan 6 sel (E) hasil pembelahan simetris, kumpulan sel non embriogenik tidak membentuk struktur yang spesifik (F), planlet dengan beberapa tunas (G), dan planlet dengan beberapa akar (H).
Berbeda dengan morfogenesis basal daun pada media padat dan cair, basal daun
yang terenkapsulasi dengan Na-alginat 3% tidak mampu bertahan hidup. Diduga bahwa
kapsul alginat memberikan tekanan mekanis terhadap eksplan dan proses respirasi
eksplan mengalami penghambatan pada kondisi terenkapsulasi. Redenbaugh et al.
(1993) menduga rendahnya respirasi oleh karena aliran gas yang terbatas menyebabkan
rendahnya pertumbuhan biakan yang terenkapsulasi. Dugaan penghambatan proses
respirasi juga dilaporkan oleh Alatar dan Faisal (2012) pada enkapsulasi tunas Rauvolfia
tetraphylla. Oleh karena itu, pada tahap selanjutnya diharapkan bahwa induksi nodul
dari basal daun melalui pra-perlakuan eksplan dengan BA dan NAA akan meningkatkan
daya hidup dan regenerasi eksplan terenkapsulasi. Sayangnya, pra-perlakuan tidak
mampu memacu pertumbuhan eksplan terenkapsulasi dan hanya mampu menginduksi
pertunasan sebesar 5% serta tidak mampu mempertahankan daya hidup lebih dari 1
bulan. Oleh karena itu, pada tahapan berikutnya (percobaan penyimpanan in vitro)
digunakan eksplan berupa tunas dan embrio somatik.
Sebagaimana fungsi benih sebagai bahan perbanyakan dan sekaligus sebagai
bahan penyimpanan maka pada penelitin ini juga dilakukan percobaan pembentukan
benih sintetik dan penyimpanan in vitro. Hasil penelitian menunjukkan bahwa suhu

tidak memberikan pengaruh yang nyata sedangkan pemberian paklobutrazol dan
manitol berpengaruh nyata terhadap penghambatan pertumbuhan biakan yang
terenkapsulasi. Pemberian paklobutrazol mampu menghambat pertumbuhan biakan
sehingga daya tembusnya lebih rendah daripada biakan yang ditumbuhkan pada media
tanpa pemberian paklobutrazol. Menurut Arteca (1996), senyawa paklobutrazol
merupakan retardan kelompok triazol yang mampu mereduksi pertumbuhan biakan
dengan cara menghambat oksidasi kauren, kaurenol, dan kaurenal yang dikatalisis oleh
kauren oksidase pada biosintesis giberelin. Secara visual, pemberian paklobutrazol juga
menyebabkan ketegaran biakan. Cathey (1975) dan Bessembinder et al. (1993)
melaporkan bahwa secara fisiologis, retardan dapat mendukung terbentuknya klorofil
sehingga kultur tampak lebih tegar. Arteca (1996) juga menjelaskan bahwa selain
memblokir biosintesis giberelin, paklobutrazol juga dapat mereduksi ABA dan etilen,
namun dapat meningkatkan kandungan sitokinin. Banyaknya klorofil yang terbentuk
dapat meningkatkan efisiensi fotosintesis sehingga pertumbuhan kultur menjadi lebih
tegar dan terpacu. Penggunaan paklobutrazol 1 mg l-1 dapat diaplikasikan untuk
penyimpanan kapsul tunas nenas dalam jangka waktu yang pendek (1 bulan) atau
untuk keperluan transportasi benih sintetik dalam jarak dekat. Untuk penyimpanan yang
lebih lama maka diperlukan modifikasi metode pertumbuhan minimal.
Penggunaan embrio somatik dan pengggunaan manitol terbukti dapat
memperlama masa simpan biakan nenas hingga 4 bulan dengan daya hidup 50%. Di
antara perlakuan manitol, konsentrasi manitol 4% dan suhu 25 0C merupakan perlakuan
yang terbaik karena perlakuan tersebut dapat mempertahankan daya hidup eksplan
pasca penyimpanan. Manitol 4% dapat digunakan untuk penyimpanan jangka
menengah. Berdasarkan efektivitas penyimpanan, hasil penelitian ini lebih baik
daripada penelitian sebelumnya (Gangopadhyay et al. 2004) yang mampu menyimpan
biakan nenas selama 1.5 bulan melalui aplikasi suhu 8 0C dengan menggunakan media
MS tanpa zat pengatur tumbuh. Berdasarkan efisiensi, penyimpanan dengan
menggunakan manitol 4% akan lebih hemat daripada penyimpanan dengan penurunan
suhu yang memerlukan energi listrik yang lebih banyak dan biaya yang lebih tinggi.
Pada masa mendatang, teknologi benih sintetik perlu diteliti lebih lanjut untuk
pengembangannya secara aplikatif. Penggunaan teknik tetes, benam, dan celup dengan

menggunakan bahan hidrofilik (alginat, karagenan, gelatin) dan hidrofobik (etil
selulosa, hidroksietil selulosa, matriks bebas polimer) dapat diujikan untuk menentukan
metode enkapsulasi yang terbaik. Demikian pula, penggunaan bahan-bahan lain (busa,
serat, ijuk) perlu dicobakan untuk mendukung proses konversi biakan menjadi planlet
dan proses transplanting bahan tanaman ke lapang. Teknologi benih sintetik dalam skala
masal atau skala industri dapat diterapkan dengan menggunakan mesin enkapsulasi.
Sebagaimana telah dijelaskan sebelumnya, efektivitas teknik regenerasi tidak
hanya ditentukan oleh tingkat multiplikasi tunas yang tinggi namun juga ditentukan oleh
stabilitas sifat yang tinggi dari bibit yang dihasilkannya. Oleh karena itu, uji stabilitas
sifat perlu dilakukan sebelum metode regenerasi tersebut diterapkan, terutama untuk
bibit nenas hasil kultur in vitro yang mudah mengalami penyimpangan. Uji stabilitas
sifat dapat dilakukan lebih efektif jika diketahui metode deteksi penyimpangan tersebut.
Deteksi penyimpangan bibit yang dihasilkan dari teknik kultur in vitro dapat
dilakukan melalui karakterisasi morfologi (Wakasa 1979; Noor et al. 2009; Somsri et al.
2009 ; Zhao et al. 2005; Podwyszyńska 2005; Suminar 2010), sitologi (Al-Zahim et al.
1999; Joachimiak dan Ilnicki 2003), biokimia (Feuser et al. 2003), fisiologi (Perez et al.
2011), dan molekuler (Chen et al. 1998; Al-Zahim et al. 1999; Soniya et al. 2001;
Soneji et al. 2002; Feuser et al. 2003; Santos et al. 2008; Perez et al. 2011).) Hingga
saat ini, belum terdapat penelitian yang melaporkan karakterisasi secara morfologi
biakan in vitro nenas yang dikulturkan dalam periode yang panjang (tahunan).
Hasil penelitian menunjukkan tingginya keragaman fenotipe biakan in vitro nenas
yang dikulturkan dalam waktu yang panjang (4 tahun). Varian yang diperoleh jauh lebih
banyak (21 varian) dibandingkan dengan hasil penelitian sebelumnya yang dilaporkan
oleh Wakasa (1979) yang menghasilkan 6 varian. Jumlah varian yang dihasilkan dalam
penelitian ini bahkan lebih banyak jika dibandingkan dengan hasil iradiasi sinar gamma
yang dilakukan oleh Suminar (2010) sehingga diduga bahwa periode in vitro yang
panjang lebih dominan dalam membentuk keragaman daripada perlakuan iradiasi,
terutama ketika dosis yang diterapkan adalah dosis suboptimal.
Keragaman yang teramati pada studi karakterisasi morfologi selanjutnya menjadi
pedoman dalam deteksi penyimpangan morfologi biakan yang dihasilkan dari 3 macam

metode regenerasi, yaitu organogenesis langsung, organogenesis dan embriogenesis
somatik tak langsung. Demikian pula, penanda RAPD yang telah terseleksi dan diujikan
pada populasi biakan umur 4 tahun tersebut selanjutnya digunakan untuk deteksi dini
biakan in vitro yang dihasilkan dari 3 macam metode regenerasi tersebut.
Ketika masing-masing varian diisolasi, beberapa varian berubah fenotipenya
sehingga diduga bahwa varian tersebut terjadi karena epigenetik, yaitu perubahan
fenotipe terjadi tanpa perubahan pada sekuens DNA. Sebagian varian memperlihatkan
sifat yang agak stabil selama proses subkultur karena menghasilkan anakan dengan
fenotipe yang sama dan yang berbeda dengan varian induknya sehingga diduga bahwa
nenas mempunyai jaringan polisomik yang menyebabkan khimera. Dugaan tersebut
diperkuat dengan ditemukannya keragaman tanaman nenas di lapang yang diperbanyak
secara konvensional. Menariknya, sebagian varian bersifat stabil selama proses
aklimatisasi di rumah kaca sehingga diduga telah terjadi keragaman somaklonal. Hasil
analisis RAPD memperkuat dugaan terjadinya keragaman somaklonal atau keragaman
yang terjadi pada sekuens DNA. Berdasarkan analisis RAPD tersebut, populasi lama
(umur 4 tahun) mempunyai koefisien kemiripan sebesar 0.32-0.91.
Untuk mengekstraksi khimera maka jaringan tanaman perlu dipisah-pisahkan.
Hasil organogenesis langsung memperlihatkan bahwa regeneran yang dihasilkan
memiliki fenotipe normal dengan penampilan yang seragam sehingga diduga teknik ini
dapat mengekstraksi khimera. Namun demikian, keragaman morfologi (sebesar 1.6%)
terdeteksi dari regeneran yang diinkubasikan dalam waktu yang lebih lama (hingga 4
bulan) sehingga timbul 2 dugaan. Dugaan pertama adalah khimera tidak berhasil
dipisahkan dengan metode organogenesis langsung. Dugaan kedua adalah periode in
vitro yang lebih lama menjadi penyebab terjadinya keragaman. Selanjutnya, pemisahan
khimera diupayakan melalui teknik organogenesis dan embriogenesis somatik tidak
langsung. Namun demikian, tipe dan tingkat keragaman justru meningkat (masing-
masing sebesar 2% dan 31.1%) sehingga diduga bahwa khimera bukan menjadi
penyebab utama terjadinya keragaman pada kultur in vitro nenas. Hasil evaluasi
keragaman fenotipe bibit yang diaklimatisasi di rumah kaca menunjukkan tidak
konsistennya karakter duri pada daun. Planlet yang berfenotipe normal (tepi daun halus)
ada yang berubah menjadi tanaman berdaun berduri dan sebaliknya planlet yang

berfenotipe berduri ada yang berubah menjadi normal. Terdapat laporan bahwa suhu
tinggi pada malam hari (Chan et al 2003) menyebabkan timbulnya duri pada daun
sehingga diduga bahwa interaksi genotipe dan lingkungan (G X E) sangat tinggi pada
tanaman nenas. Pengamatan lebih lanjut di rumah kaca menunjukkan adanya pola yang
sama sekali tidak beraturan untuk karakter duri pada daun karena duri dapat muncul
pada daun manapun secara silih berganti, baik pada salah satu sisi ataupun pada kedua
sisinya sehingga diduga terdapat gen yang secara aktif berinteraksi dengan stres
lingkungan sehingga menimbulkan ekspresi yang berbeda (daun halus menjadi berduri).
Telah diketahui bahwa embriogenesis somatik pada tanaman nenas kultivar Smooth
Cayenne bersifat unicellular origin, sehingga bibit yang dihasilkan seharusnya memiliki
morfologi yang konsisten (berupa fenotipe normal atau varian tertentu yang bersifat
stabil) karena masing-masing tanaman tersebut berasal dari satu sel. Perubahan fenotipe
pada bibit tersebut mengindikasikan adanya faktor lain yang mendasari perubahan
tersebut. Fakta-fakta yang ada sudah cukup kuat memunculkan dugaan adanya elemen
loncat (transposable element) dalam tanaman nenas. Berdasarkan penelusuran pustaka
diketahui bahwa retotransposon-like sequence telah terintegrasi ke dalam genom
tanaman nenas, dua sekuens di antaranya bahkan tidak mempunyai stop kodon sehingga
memungkinkan loncatnya elemen tersebut ke daerah genom lainnya (Thomson et al.
1998). Dilaporkan oleh Kumar dan Benetzen (1999) bahwa retrotransposon diaktivasi
oleh stres biotik dan abiotik karena retrotransposon memiliki sekuens cis-acting element
yang terdapat di dalam daerah U3 pada bagian 5’ long terminal repeat (LTR) yang
bersebelahan dengan daerah yang tidak ditranslasikan (untranslated region).
Untuk melihat seberapa besar pengaruh metode regenerasi dalam menimbulkan
keragaman duri pada daun maka evaluasi keragaman dilakukan juga pada bibit yang
diaklimatisasi di rumah kaca. Hasil evaluasi tersebut menunjukkan bahwa
organogenesis langsung menghasilkan bibit berfenotipe tepi daun halus dengan proporsi
tertinggi (48.3%) dibandingkan dengan organogenesis tidak langsung (41.4%) dan
embriogenesis somatik (27.1%) sehingga disimpulkan bahwa metode organogenesis
langsung merupakan metode yang paling efektif. Setelah diketahui bahwa periode in
vitro merupakan faktor yang dominan dalam menimbulkan keragaman somaklonal
maka dibentuklah populasi biakan in vitro yang baru dengan metode organogenesis
langsung untuk mereduksi keragaman somaklonal. Secara molekuler, populasi baru

tersebut memiliki koefisien kemiripan yang lebih tinggi dibandingkan dengan tanaman
kontrolnya (0.65-0.85) daripada populasi yang lama yang telah diekstraksi (0.33-0.63)
dan populasi asalnya atau yang belum diekstraksi (0.32-0.61) melalui metode
organogenesis langsung serta orgaogenesis dan embriogenesis somatik tidak langsung.
Akhirnya, diusulkan bahwa metode karakterisasi morfologi biakan in vitro yang
dihasilkan dalam penelitian ini dapat dimanfaatkan sebagai pedoman dalam pemilihan
varian-varian yang tak dikehendaki selama dalam kultur in vitro secara lebih dini.
Metode organogenesis langsung juga dapat diterapkan untuk propagasi tanaman nenas
secara masal. Varian yang dihasilkan dari penelitian ini dimanfaatkan untuk studi
forward genetic untuk mempelajari karakter yang menarik pada varian sebagaimana
disarankan oleh Wessler (1996). Metode regenerasi secara in vitro dan metode seleksi
dini dapat diterapkan untuk pengembangan komoditi nenas secara komersial, baik nenas
hasil persilangan maupun perbanyakan klonal. Di antara berbagai negara penghasil
nenas, Cuba berhasil memproduksi bibit nenas hibrida melalui teknik mikropropagasi.
Malaysia bahkan berhasil mengembangkan nenas Masmerah yang merupakan varian
hasil perbanyakan klonal dari kultivar Singapore Spanish. Demikian pula, Filipina
berhasil mengembangkan nenas tipe Queen tak berduri secara mikropropagasi (Chan et
al. 2003), sementara diketahui bahwa Filipina merupakan salah satu negara yang
mendominasi perdagangan nenas di dunia setelah Thailand (Rohrbach et al. 2003).
Di antara varian yang dihasilkan dalam penelitian ini, varian erect posture, wider
leaf, dan narrow leaf merupakan varian yang berpotensi besar untuk dikembangkan.
Varian erect posture dan narrow leaf mempunyai perawakan yang ramping dengan
daun yang tegak sehingga memungkinkan untuk ditanam dengan jarak tanam yang lebih
rapat (densitas tanaman dapat ditingkatkan per satuan luas). Selain itu, dengan tegaknya
daun maka fotosintesis dapat lebih efektif sehingga diharapkan fotosintat yang
terakumulasi dalam buah akan lebih banyak. Varian wider leaf juga diduga akan
menghasilkan buah dengan ukuran yang lebih besar sebagaimana dikatakan oleh
Coppens d’Eeckenbrugge dan Leal (2003) bahwa peningkatan ukuran buah pada nenas
sejalan dengan peningkatan ukuran organ lainnya, seperti daun lebih lebar, batang lebih
besar, tangkai bunga lebih pendek dan lebar. Sebaliknya beberapa varian diduga akan
berpengaruh negatif terhadap produksi buah. Varian branched shoot diduga akan gagal

membentuk bunga dan buah karena pertumbuhan vegetatif lebih dominan daripada
perkembangan generatifnya. Varian albino stripe leaf juga diduga akan menghasilkan
buah yang berukuran kecil atau abnormal karena fotosintat yang dihasilkan akan lebih
sedikit sehingga kurang mendukung perkembangan buah. Varian spiness spot leaf
sebaliknya berpotensi menghasilkan buah dengan mahkota yang berduri sehingga
kurang tepat dijadikan sebagai bahan tanam atau bibit. Varian powdery leaf juga akan
memberikan pengaruh yang negatif terhadap proses fotosintesis karena akan banyak
cahaya yang dipantulkan sehingga efisiensi fotosintesis menjadi menurun dan
selanjutnya berpengaruh terhadap perkembangan buah.
Selain varian yang berpotensi untuk dikembangkan, varian spiness spot leaf yang
100% berduri (complete spiness) juga bibit yang berfenotipe smooth 100% dapat
dimanfaatkan sebagai bahan studi untuk mengetahui mekanisme perubahan sifat
tersebut yang dikontrol oleh retrotransposon. Dilaporkan bahwa dari berbagai genotipe
tanaman yang dipertimbangkan bersifat stabil, duri pada daun merupakan karakter yang
bersifat tidak stabil dan mempunyai frekuensi yang tinggi (Collins dan Kerns 1938
dalam Chan et al. 2003). Hal ini akan menjadi bahan kajian yang sangat menarik dalam
mempelajari mekanisme aktifnya retrotransposon yang semakin dipicu oleh stres dalam
kultur in vitro dan upaya penanggulangannya untuk mempertahankan komposisi genetik
tanaman nenas dalam perbanyakan secara masal.

SIMPULAN DAN SARAN
Pemberian ZPT (berupa sitokinin atau auksin) mutlak diperlukan untuk
menginduksi proses diferensiasi eksplan basal daun nenas kultivar Smooth Cayenne.
Morfogenesis eksplan nenas pada media padat lebih baik daripada dalam media cair.
Penggunaan pikloram lebih baik daripada NAA dan 2,4-D dalam menginduksi
embriogenesis somatik. Penggunaan embrio somatik lebih baik daripada tunas in vitro
dan manitol (4%) lebih baik daripada paklobutrazol dan penurunan suhu dalam
menyimpan kapsul nenas selama 4 bulan. Terdapat 21 varian yang terdeteksi dari
populasi induk (umur 4 tahun), 8 varian bersifat epigenetik, 8 varian bersifat khimera,
dan 7 varian bersifat stabil. Karakterisasi morfologi menghasilkan dendogram dengan
koefisien kemiripan 0.49-0.91 (r=0.83). Karakterisasi molekuler membuktikan
terjadinya keragaman somaklonal (perubahan pada sekuens DNA). Evaluasi keragaman
biakan in vitro menunjukkan bahwa metode organogenesis langsung memiliki tingkat
keragaman yang paling rendah (1.6%) daripada metode organogenesis tidak langsung
(2%) dan embriogenesis somatik tidak langsung (31.1%), demikian pula dengan hasil
evaluasi keragaman bibit aklimatisasi dari populasi tersebut menunjukkan hasil yang
serupa. Populasi baru memiliki koefisien kemiripan genetik yang lebih tinggi (0.65-
0.85) daripada populasi induk, baik sebelum (0.32-0.61) maupun sesudah diekstraksi
(0.34-0.63) dengan 3 macam metode regenerasi. Secara umum, metode organogenesis
langsung merupakan metode yang paling efektif untuk perbanyakan bibit nenas kultivar
Smooth Cayenne secara masal. Penggunaan populasi baru yang dikombinasikan dengan
metode organogenesis langsung dapat mereduksi keragaman somaklonal.
Disarankan untuk melakukan evaluasi karakter agronomi dan biokimia terhadap
varian-varian yang dihasilkan dalam penelitian ini, melakukan rekayasa seluler untuk
peningkatan sifat toleransi tanaman nenas terhadap cekaman kekeringan dan
kemasaman serta peningkatan kualitas buah (peningkatan kadar asam askorbat dan
penurunan kadar Ca-oksalat). Selain itu, disarankan untuk melakukan studi forward
genetic terhadap varian-varian yang mempunyai karakter yang menarik.

DAFTAR PUSTAKA
Abrash, E.B. and D.C. Bergmann. 2009. Asymmetric cell divisions: A view from plant development. Developmental Cell 16: 783-796.
Adaniya S, Minemoto K, Moromizato Z, Molomura K. 2004. The use of CPPU for efficient propagation of pineapple. Hort Sci 100: 7–14.
Al-Zahim MA, Ford-Lloyd BV, Newbury HJ. 1999. Detection of somaclonal variation in garlic (Allium sativum L.) using RAPD and cytological analysis. Plant CellRep18: 473-477.
Arnold S, Sabala I, Bozhkov P, Dyachok J, Filonova L. 2002. Developmental pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult 69: 233-249.
Arteca RN. 1996. Plant Growth Substances. Chapman and Hall. New York.332p.
Ashmore SE. 1997. Status Report on The Development and Application of In Vitro Techniques for The Conservation and Use of Plant Genetic Resources. IPGRI (International Plant Genetic Resources Institute). Rome. Italy. 67 p.
Baez R, Lopes MTP, Salas CE, Hernandez M. 2007. In vitro antitumoral activity of stem pineapple (Ananas comosus) bromelain. Planta Med 73:1377-1383.
Bairu MW, Fennell CW, Staden J van. 2006. The effect of plant growth regulators on somaclonal variation in Cavendish banana (Musa AAA cv. ‘Zelig’). Sci Hort 108:347–351.
Bessembinder JJE, Staritsky G, Zandvoort EA. 1993. Longterm in vitro storage of Colocasia esculenta under minimal growth conditions. Plant Cell Tiss Org Cult 33:121-127.
Bhojwani SS, Razdan MK. 1983. Plant Tissue Culture: Theory and Practise. Elsevier. Amsterdam. New York. 502p.
[BPS] Badan Pusat Statistik Republik Indonesia. 2009. Produksi buah-buahan di Indonesia. Jakarta
Buffard-Morel JL et al. 1995. Initiation of somatic embryogenesis in coconut (Cocos nucifera L.). Pp. 217-223. In: Oropeza C et al., editor. Lethal Yellowing: Research and Practical Aspects. Kluwer Academic Publishers, Netherlands.
Cathey HM. 1975. Comparative plant growth-retarding activities of ancymidol with ACPC, phosfon, clhormequat, and SADH on ornamental plant species. Hort Science 10(3):204-215.

Chan YK, Coppens d’Eeckenbrugge G, Sanewski GM. 2003. Breeding and variety improvement. Pp. 33-56. In: Bartholomew DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany: Production and Uses. CABI Publishing.Wallingford.
Chen WH, Chen TM, Fu YM, Hsieh RM, Chen WS. 1998. Studies on somaclonal variation in Phalenopsis. Plant Cell Rep18: 7-13.
Collins JL. 1951. Notes on the origin, history, and genetic nature of the Cayenne pineapple. Pacific Science 5:3-17
Collins JL. 1960. The Pineapple Botany, Cultivation, and Utilization. Leonard Hill, London
Coppens d’Eeckenbrugge G, Leal F. 2003. Morphology, anatomy, and taxonomy. Pp. 13-32. In: Bartholomew DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany: Production and Uses. CABI Publishing.Wallingford.
Corredoira E, Valladares S, Vieitez AM. Morphohistological analysis of the origin and development of somatic embryos from leaves of mature Quercus robur. In Vitro Cell Dev Biol_Plant 42:525-533.
Cunha A, Fernandez-Ferreira M. 2012. Influence of medium parameters on somatic embryogenesis from hypocotyl explants of flax (Linum usitatissimum L.): Effect of carbon source, total inorganic nitrogen and balance between ionic forms and interaction between calcium and zeatin. http://dx.doi.org/10.1016/S0176-1617(99)80059-5.
Desbrunais AB, Noirot M, Charrier A. 1992. Slow growth in vitro conservation of coffee (Coffea spp.). Plant Cell Tissue Organ Cult 31:105-110.
Dewi IS, Jawak G, Roostika I, Sabda M, Purwoko BS, Adil WH. 2010. Konservasi in vitro tanaman jeruk besar (Citrus maxima (Burn.) Merr.) kultivar Srinyonya menggunakan osmotikum dan retardan. Jurnal Agrobiogen 6(2):84-90
Eeuwens CJ et al. 2002. Effects of tissue culture conditions during embryoid multiplication on the incidence of “mantled” flowering in clonally propagated oil palm. Plant Cell Tissue Organ Cult 70:311-323.
Feuser S, Meler K, Daquinta M, Guerra MP. 2003. Genotype fidelity of micropropagated pineapple (Ananas comosus) plantlets assessed by isozymes and RAPD markers. Plant Cell Tissue Organ Cult 72:221-227
Firoozabady E, Gutterson N. 2003. Cost-effective in vitro propagation methods for pineapple. Plant Cell Rep 21:844-850.
Firoozabady E, Moy Y. 2004. Regeneration of pineapple via somatic embryogenesis and organogenesis. In vitro Cell Dev Biol_Plant 40:67-74.

Gamez-Pastrana R, Martinez-Ocampo Y, Beristain CI, Gozales-Arnao MT. 2004. An improved cryopreservation protocol for pineapple apices using encapsulation-vitrification. CryoLetters 25(6): 405-414.
Gangopadhyay G, Bandyopadhyay T, Poddar R, Gangopadhyay SB, Mukherjee KK. 2005. Encapsulation of micro shoots in alginate beads for temporary storage. Curr Res 88(6): 972-977.
Gatica AM, Arrieta G, Espinoza AM. 2008. Direct somatic embryogenesis in Coffea arabica L.. cvs. Caturra and Catuaí: effect of triacontanol, light condition, and medium consistency. Agronomía Costarricense 32(1): 139-147.
Grout BWW. 1995. Introduction to the in vitro preservation of plant cells, tissues and organs. In: Grout B, editor. Genetic Preservation of Plant Cells In Vitro. Springer Lab Manual. Berlin-Heidelberg. Pp 1-17.
Hadiati S, Sukmadjaja D. 2004. Keragaman pola pita beberapa aksesi nenas berdasarkan analisis isozim. J Biotek Pert 7(2): 62-70.
Hagen SR, Muneta P, Augustin J, Tourneau D Le. 1991. Stability and utilization of picloram, vitamins, and sucrose in a tissue culture medium. Plant Cell Tissue and Organ Cult 25(1):45-48.DOI: 10.1007/BF00033911.(Abstract)
Hu CY, Wang PJ. 1983. Meristem, shoot tip, and bud culture. Pp. 177-227. In. Evans DA, Sharp WR, Amiroto PV, Yamada Y, editor. Handbook of Plant Cell Culture Vol. I. Techniques for Propagation and Breeding. McMilan Publishing, New York.
[IBPGR] International Board for Plant Genetic Resources. 1991. Descriptors for Pineapple. IBPGR Headquarters, Rome
Jimenez VM. 2005. Involvement of plant hormone and plant growth regulators on in vitro somatic embryogenesis. Plant Growth Regulation 47: 91-110.
Joachimiak A, Ilnicki T. 2003. Nuclear morphology, polyploidy, and chromatin elimination in tissue culture of Allium fistulosum L. Acta Societatis Botanicorum Poloniae 77(1):11-17.
Kaeppler SM, Kaeppler HF, Rhee Y. 2000. Epigenetic aspects of somaclonal variation in plants. Plant Mol Biol 43:179-188.
Keller ERJ, Senula A, Leunufna S, Grube M. 2006. Slow growth storage and cryopreservation-tools to facilitate germplasm maintenance of vegetatively propagated crops in living plant collections. Int J of Refr 29:411-417.
Kelley KB, Riechers DE. 2007. Recent development in auxin biology and new properties for auxinic herbicide research. Pesticide Biochemistry and Physiology 89: 1-11.

Khan S, Nasib A, Saeed BA. 2004. Employment of in vitro technology for large scale multiplication of pineapples (Ananas comosos). Pak J Bot 36(3): 611-615.
Kiernan JA. 1990. Histological and Histochemical Methods: Theory and Practice. 2nd edition. Pergamon Press.
Koornneef M. 1991. Variation and mutant selection in plant cell and tissue culture. In: Jones L, editor. Biotechnological Innovation in Crop Improvement. London: Butterworth-Heinemann Ltd. pp 100-115.
Kordestani GK, Karami O. 2008. Picloram-induced somatic embryogenesis in leaves of strawberry (Fragaria ananassa L.). Acta Biologica Cracoviensia Series Botanica 50(1): 69–72.
Kumar A, Bennetzen JZ. 1999. Plant retrotransposon. Annu. Rev. Genet 33: 479-532.
Laukkanen, H., L. Rautiainen, E. Taulavuori, and A. Hohtola. 2000. Changes in cellular structures and enzymatic activities during browning of Scots pine callus derived from mature buds. Tree Physiology 20: 467–475.
Lestari EG, Mariska I. 1997. Kultur in vitro sebagai metode pelestarian tumbuhan obat langka. Buletin Plasma Nutfah 2(1): 1-8.
Leunufna S. 2004. Improvement of the in vitro maintenance and cryopreservation of yams (Dioscorea spp.). Dissertation Martin-Luther–Universitat Halle-Wittenberg. Berlin. 12p. Unpublished.
Lincy AK, Remashree AB, Sasikumar B. 2009. Indirect and direct somatic embryogenesis from aerial stem explants of ginger (Zingiber officinale Rosc.). Acta Bot Croat 68 (1): 93–103.
Mariska I, Suwarno, Damardjati DS. 1996. Pengembangan konservasi in vitro sebagai salah satu bentuk pelestarian plasma nutfah di dalam bank gen. Dalam: Seminar Penyusunan Konsep Pelestarian Ex Situ Plasma Nutfah Pertanian. Bogor. 18 Desember 1996. Balitbio Bogor.
Masoud S, Hamta A. 2008. Cytogenetic analysis of somaclonal variation in regenerated plants of berseem clover (Trifolium alexandrium L.). Cryologia 61(4):392-396.
Meneses A, Flores D, Muñoz M, Arriet G, Espinoza AM. 2005. Effect of 2,4-D, hydric stress and light on indica rice (Oryza sativa) somatic embryogenesis. Int J Trop Biol 53 (3-4): 361-368.
Moubayidin L, Mambro RD, Sabatini S. 2009. Cytokinin–auxin crosstalk. Trends in Plant Science 14 (10): 557-562.
Murashige T, Skoog F. 1962. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15:473-497.

Noor NM, Clyde MM, Rao VR, Jeevamoney J. 2009. Radiosensitivity and in vitro studies of Citrus suhuiensis. Pp. 17-32. In: IAEA. Induced Mutation in Tropical Fruit Trees. Vienna.
Nursandi F, Poerwanto R, Sobir, Sujiprihati S, Purwito A. 2005a. Studi pertumbuhan tanaman nenas (Ananas comosus (L.) Merr.) kultivar Queen hasil in vitro dan analisis kestabilan genetik berdasarkan karakter morfologi dan RAPD. Dalam: Konggres Peripi 2005.
Nursandi F, Poerwanto R, Sobir, Sujiprihati S. 2003. Perbanyakan in vitro nenas (Ananas comosus (L.) Merr.) dengan menggunakan BAP dan TDZ. Dalam: Simposium Nasional dan Konggres VIII. Bandar Lampung.
Nursandi F, Sobir, Murtini. 2005b. Perbanyakan tanaman nenas (Ananas comosus (L.) Merr.) cv. Smooth Cayenne dengan teknik etiolasi secara in vitro. Tropika 13(1):15-24.
Nursandi F. 2006. Studi perbanyakan in vitro tanaman nenas (Ananas comosus L. Merr.) dan analisis kestabilan genetic berdasarkan karakter morofologi isozim, dan RAPD [disertasi]. Sekolah Pascasarjana, Institut Pertanian Bogor.117 hal.
Ochse JJ, Soule Jr MJ, Dijkman MJ, Wehlburg C. 1961. Tropical and Subtropical Agriculture. Vol. I. The Macmillan Company. New York.
Okamoto H, Hirochika H. 2001. Silencing of transposable elements in plants. Trends in Plant Sci 6(11):527-534.
Oktavia F, Siswanto, Budiani A, Sudarsono. 2003. Direct somatic embryogenesis and regeneration of arabica coffee plantlets (Coffea arabica) from different explant. Menara Perkebunan 71(2): 44-55.
Paciorek T, Bergmann DC. 2010. The secret to life is being different: asymmetric divisions in plant development. Current Opinion in Plant Biology 13:661–669.
Perez G, Yanes E, Isidron M, Lorenzo JC. 2009. Phenotypic and AFLP characterization of two new pineapple somaclones derived from in vitro culture. Plant Cell Tissue Organ Cult 96:113-116.
Perez G et al. 2011. Morphological and physiological characterization of two new pineapple somaclones derived from in vitro culture. In Vitro Cell Dev Biol_Plant. DOI 10.1007/s11627-011-9342-y.
Phillips GC, Hubstenberger JF, Hansen EE. 1995. Plant regeneration from callus and cell suspension cultures by somatic embryogenesis. Pp. 81 - 90. In: Gamborg OL, Phillips GC, editor. Plant Cell Tissue Organ Cult: Fundamental Methods. Springer. Berlin.

Podwyszyńska M. 2005. Somaclonal variationin micropropagated tulips based on phenotype observation. Journal of Fruit and Ornamental Plant Research 13:109-122.
Popluechai S, Onto S, Eungwanichayapant PD. 2007. Relationships between some Thai cultivars of pineapple (Ananas comosus) revealed by RAPD analysis. Songklanakarin J Sci Technol 29(6): 1491-1497.
Purnamaningsih R, Mariska I, Supriati Y. 2009. Penggunaan ABA dan paklobutrazol dalam perbanyakan nenas Simadu melalui kultur in vitro. Ber Bio 9(6): 751-758.
Rai MK, Asthana P, Singh SK, Jaiswal VS, Jaiswal U. 2009. The encapsulation technology in fruit plants – A review. Biotech Adv 27: 671-679.
Rani V, Raina SN. 2000. Genetic fidelity of organized meristem-derived micropropagated plants: a critical Reappraisal. In Vitro Cell Dev Biol_Plant 36:319-330.
Redenbaugh K. 1992. Synseeds: Application of synthetic seeds to crop improvement. CRC Press. 481 p.
[Ristek] Kementrian Riset dan Teknologi. 2009. Laporan Riset Unggulan Strategis Nasional dalam realistas kurun waktu 2000 – 2009. Jakarta.
Roostika I, Purnamaningsih R, Arief VN. 2008. Pengaruh sumber karbon dan kondisi inkubasi terhadap pertumbuhan kultur in vitro purwoceng (Pimpinella pruatjan Molk.). AgroBiogen 4(2): 65-69
Roostika I, Purnamaningsih R, Darwati I. 2009. Penyimpanan in vitro tanaman purwoceng (Pimpinella pruatjan Molk.) melalui aplikasi pengenceran media dan paklobutrazol. Jurnal Littri 15(2): 84-90
Roostika I, Sunarlim N, Arief VN. 2005. Penyimpanan kentang hitam (Coleus tuberosus) secara kultur in vitro. PP 24 (1):46-52
Roostika I, Sunarlim N. 2001. Penyimpanan in vitro tunas ubi jalar dengan penggunaan paclobutrazol dan ancymidol. PP 20 (3):48-56.
Sagiman S. 2007. Pemanfaatan lahan gambut dengan perspektif pertanian berkelanjutan. Orasi Ilmiah Guru Besar Fakultas Pertanian Universitas Tanjungpura. Pontianak.
Sakakibara. H. 2004. Cytokinin biosynthesis and metabolism. Pp. 95-114. In. Davies, P.J. (Ed.). Plant Hormones: Biosynthesis, Signal, Action. 3rd Edition. Kluwer Academic Publishers. London.
Santos MDM, Buso GCS, Torres AC. 2008. Evaluation of genetic variability in micropropagated propagules of ornamental pineapple (Ananas comosus var. bracteatus (Lindley) Coppens and Leal) using RAPD markers. Gen Mol Res 7(4):1097-1105.

Sidha M, Suprasanna P, Bapat VA, Kulkarni UG, Shinde BN. 2007. Developing somatic embryogenic culture system and plant regeneration in banana. Founder’s Day 285: 153-161.
Silva AE De, Kadir MA, Aziz MA, Kadzimin S. 2008. Callus induction in pineapple (Ananas comosus L.) cv. Moris and Jasapine. Int J of Agric Res 3(4): 261-267.
Smith MK, Ko H-L, Hamil SD, Sanewski GM, Graham MW. 2003. Biotechnology. Pp. 57-68. In: Bartholomew DP, Paull RE, Rohrbach KG, editor. The Pineapple Botany: Production and Uses. CABI Publishing.Wallingford.
Somsri S, et al. 2009. Development of seedless fruits mutants in citrus including tangerine (C. reticulate) and pummelo (C. grandis) through induced mutations and biotechnology. Pp. 33-40. In: IAEA. Induced Mutation in Tropical Fruit Trees. Vienna.
Soneji J., Rao PS, Mhatre M. 2002a. Somaclonal variation in micropropagated dormant axillary buds of pineapple(Ananas comosus L., Merr.). J Hort SciBiotech 77: 28-32.
Soneji JR, Rao PS, Mhatre M. 2002b. Suitability of RAPD for analyzing spined and spineless variant regenerants of pineapple (Ananas comosus L., Merr.). Plant Mol Biol Rep 20:307a-307i.
Soniya EV, Banerjee NS. Das MR. 2001. Genetic analysis of somaclonal variation among callus-derived plants of tomato. Current Science 80(9):1213-1215.
Sripaoraya S, Marchamt R, Power JB, Davey MR. 2003. Plant regeneration by somatic embryogenesis and organogenesis in commercial pineapple (Ananas comosus L.). In Vitro Cell Dev Biol_Plant 39:450-454.
Srivastava LM. 2002. Plant Growth and Development. Academic Press. New York. 772p.
Sudarmonowati E, Henshaw GG. 1996. The use of picloram and dicamba to induce somatic embryogenesis in cassava. Annales Bogoriensis 4 (1):27-34.
Suminar E. 2010. Induksi keragaman genetik dengan mutagen sinar gamma pada nenas (Ananas comosus (L.) Merr.) secara in vitro [seminar]. Sekolah Pascasarjana, Institut Pertanian Bogor. Bogor.
Sunarlim N, Roostika I, Arief VN. 2004. Penyimpanan in vitro gembili melalui pertumbuhan minimal. Dalam: Prosiding Seminar Kinerja Penelitian Mendukung Agribisnis Kacang-kacangan dan Umbi-umbian. BALITKABI. Malang.
Sunarlim N, Roostika I. 2003. Penggunaan zat penghambat tumbuh dan regulator osmotik manitol dalam penyimpanan ubi-ubian secara kultur jaringan. Dalam: Prosiding Seminar Teknologi Inovatif Agribisnis Kacang-kacangan dan Umbi-umbian untuk Mendukung Ketahan Pangan. BALITKABI. Malang.

Suryowinoto M. 1996. Pemuliaan Tanaman secara In Vitro. Pusat Antar Universitas-Bioteknologi UNiversitas Gadjah Mada. Yogyakarta. 252 hal.
Taiz L, Zeiger E. 2003. Plant Physiology-on line 3rd Ed. Sunderland: Sinauer Associates. doi:10.1093/aob/mcg079.
Tawar PN et al. 2008. An assessment of somaclonal variation in micropropagated plants of sugarcane by RAPD markers. Sugar Tech 10(2):124-127.
Thomson KG, Thomas JE, Dietzgen RG. 1998. Retrotransposon-like sequence intregated into genome of pineapple (Ananas comosus). Plant Mol Biol 38:461-465.
Uozumi N, Kohketsu K, Okamoto A, Kobayashi T. 1993. Light dependency in celery somatic embryogenesis and plantlet development in suspension culture. Plant Tissue Culture Letter 10(1): 25-32.
Vanneste S, Frim J. 2009. Auxin: A trigger for change in plant development. Cell 136: 1005-1016.
Wakasa K. 1979. Variation in the plants differentiated from the tissue culture of pineapple. Japan J Breed 29(1):13-22.
Wattimena, GA. 2012. Sumber pangan baru. Pp.126-147. Dalam: Poerwanto R, Siregar IZ, Suryani A, editor. Merevolusi Revolusi Hijau. IPB Press. Bogor.
Weerasinghe SS, Siriwardana AU. 2006. Fast propagation of pineapple (Ananas comosus) with stem cuttings. The J of Agric Sci 2(2): 55-59.
Williams JG, Kubelik AR, Livak KJ, Rafalski JA. 1990. DNA polymorphism amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res 18:6531-6535
Withers LA. 1985. Cryopreservation and storage of germplasm. In: Dixon DA, editor. Plant Cell Culture. IRL Press, Washington. Pp.169-190.
Zhao Y, Grout BWW, Crisp P. 2005. Variation in morphology and disease susceptibility of micropropagated rhubarb (Rheum rhaponticum) PC49, compared to conventional plants. Plant Cell Tissue Organ Cult 82:357-361.
Zucchi MI, Arizono H, Morais VA, Fungaro MHP, Vieira MLC. 2002. Genetic instability of sugarcane plants derived from meristem cultures. Gen Mol Biol 25(1):91-96.




















