
PENGARUH FUNGI MIKORIZA ARBUSKULA (FMA) DAN … · menghasilkan gubal gaharu di alam. Namun...
106
PENGARUH FUNGI MIKORIZA ARBUSKULA (FMA) DAN MEDIA TUMBUH TERHADAP PERTUMBUHAN PLANTLING GAHARU (Gyrinops versteegii (Gilg.) Domke) HASIL MULTIPLIKASI IN-VITRO KUSRINER F. MBAUBEDARI SEKOLAH PASCASARJANA PROGRAM STUDI ILMU PENGETAHUAN KEHUTANAN INSTITUT PERTANIAN BOGOR BOGOR 2011
Transcript of PENGARUH FUNGI MIKORIZA ARBUSKULA (FMA) DAN … · menghasilkan gubal gaharu di alam. Namun...

i
PENGARUH FUNGI MIKORIZA ARBUSKULA (FMA)
DAN MEDIA TUMBUH TERHADAP PERTUMBUHAN PLANTLING GAHARU
(Gyrinops versteegii (Gilg.) Domke) HASIL MULTIPLIKASI IN-VITRO
KUSRINER F. MBAUBEDARI
SEKOLAH PASCASARJANA
PROGRAM STUDI ILMU PENGETAHUAN KEHUTANAN
INSTITUT PERTANIAN BOGOR
BOGOR
2011

ii
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Pengaruh Fungi Mikoriza Arbuskula
(FMA) dan Media Tumbuh terhadap Pertumbuhan Plantling Gaharu (Gyrinops versteegii
(Gilg) Domke) Hasil Multiplikasi In-vitro adalah karya saya dengan arahan dari komisi
pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana
pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak
diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar
Pustaka di bagian akhir tesis ini.
Bogor, Agustus 2008
Kusriner Fernando Mbaubedari
NRP. E051060201

iii
ABSTRACT
KUSRINER F. MBAUBEDARI. The Effect of Arbuscular Mycorrhizal Fungi
(AMF) Indigenous Papua and Plantling Media to The Growth of Agarwood
(Gyrinops versteegii (gilg) Domke) Plantling from In-vitro Multiplication. Under
academic supervision of IRDIKA MANSUR and SUPRIYANTO
These research works were begun with field observation in Asai Natural Forest in
July 2010, and experiments were conducted at Silviculture Laboratory, SEAMEO
BIOTROP, Bogor, from November 2010 to June 2011. Acclimatization method was
designed using completely randomized factorial design with four level planting media and
three level AMF factors, and using three replicates. The objectives were to identify the
species of AMF associated with Gyrinops versteegii collected from Asai Natural Forest,
West Papua, to observe the compatibility of indigenous AMF Papua to G. versteegii
plantling from in-vitro multiplication, on some media combination. The results showed
that there were 7 species of AMF found from 5 sampling plots from Asai Natural Forest
which are associated with G. versteghii; there were Glomus mossae, G. fasciculatum, G.
aggregatum, Glomus sp.1, Glomus sp.2, Glomus sp.3, and Acaulospora longula.
Application of AMF significantly affected to some growth parameters such as the survival
percentage of plantling, height and diameter of stem, fresh weight of top and root, roots
geometry, vigor, and plantling quality index as well as the presence of stomata. AMF
consortium inoculums from Asai Natural Forest gave a better response to the growth of
Gyrinops plantlings compared to Gigaspora margarita.
Keywords : Plantling, Gyrinops versteegii, AMF Consortium, symbiose

iv
RINGKASAN
KUSRINER F. MBAUBEDARI. Pengaruh Fungi Mikoriza Arbuskula (FMA)
Indigenous Papua dan Media Tumbuh terhadap Pertumbuhan Plantling Gaharu
(Gyrinops versteegii (Gilg) Domke) Hasil Multiplikasi In-vitro.
Untuk mendapatkan pohon penghasil gubal gaharu yang baik, harus menggunakan
bibit dari pohon gaharu potensial, yaitu bibit unggul dari pohon inang yang telah terbukti
menghasilkan gubal gaharu di alam. Namun demikian produktifitas benih yang rendah
menyebabkan kesulitan memperoleh anakan pohon gaharu dalam jumlah banyak di alam,
padahal untuk tujuan budidaya yang luas sangat diperlukan bibit berkualitas dalam jumlah
cukup dan tersedia tepat waktu. Di sisi lain, selama ini bibit yang digunakan berasal dari
biji atau semai hutan alam dimana jumlah bibit terbatas, kualitas bibit rendah dan peluang
memperoleh gubal gaharu setelah penanaman relatif kecil karena bibit yang digunakan
belum tentu berasal dari induk yang berpontesi menghasilkan gubal gaharu.
Kendala lain yang umumnya dihadapi adalah tidak semua pohon gaharu
menghasilkan buah setiap tahun, belum adanya kebun bibit unggul dan kebun benih serta
biji gaharu bersifat rekalsitran, selain itu adanya penebangan pohon induk dewasa di alam
oleh pencari gaharu menyebabkan hilangnya sumber benih. Rendahnya daya berbunga
dan produktifitas berbuah menyebabkan masalah regenerasi secara generatif, sementara
itu pembiakan secara vegetatif menggunakan stek dan cangkok membutuhkan bahan
induk yang banyak, maka kultur in vitro gaharu menjadi alternatif teknologi perbanyakan
gaharu unggul secara masal dan cepat.
Selain dapat menghasilkan bibit berkualitas dalam jumlah yang memadai, teknik
ini berpotensi mempertahankan sifat genetis dari pohon induk penghasil gaharu. Namun
demikian, ditemukan kendala dalam kultur jaringan gaharu yaitu memerlukan waktu yang
lama untuk menginduksi akar plantlet baik secara in vitro maupun ex vitro di green
house, sementara akar yang terbentuk umumnya relatif kecil, jumlahnya sedikit serta sulit
berkembang. Problem lain adalah tahapan aklimatisasi plantling gaharu hasil in vitro ke
media aklimatisasi merupakan tahapan kritis yang masih menjadi masalah karena
plantling telah terbiasa tumbuh pada kondisi lingkungan dengan kelembaban tinggi
sementara apabila diadaptasikan secara ex vitro dengan temperatur tinggi plantling akan
mengalami dehidrasi, layu dan mati dikarenakan mekanisme buka tutup stomata yang
rendah dan stomata tidak berfungsi optimal. Lingkungan tumbuh in vitro bersifat steril
mengakibatkan imunitas plantling rendah dan lebih rentan terhadap serangan hama dan

v
penyakit busuk akar oleh jamur pembusuk akar (lodoh). Akibatnya tingkat kematian
plantling gaharu sangat tinggi, yaitu 75 % dari plantlet yang diadaptasikan ke media
pertumbuhan plantling di green house. Kondisi ini menyebabkan kegagalan produksi
bibit gaharu potensial asal kultur in vitro. Penyebab lainnya yaitu lapisan kutikula
plantling tipis dan terbiasa menerima cahaya konstan, sedangkan pada ex vitro plantling
mendapatkan cahaya tidak konstan dan intensitas cahaya tinggi. Faktor lainnya ialah
dalam media in vitro nutrisi tersedia optimal sehingga plantlet bersifat heterotrop,
sedangkan diluar media kultur plantling dituntut bersifat autotrop. Selain itu, plantling
asal in vitro memiliki lignifikasi batang rendah sehingga cenderung bersifat vitrious
(sukulen).
Aplikasi FMA pada plantling gaharu akan memacu ramifikasi akar dan batang
plantling karena mikoriza memproduksi auksin IAA untuk menstimulir akar plantling
serta menghasilkan enzim phospatase untuk pembentukan lignin. Hal tersebut dapat
tercermin dari geometri akar yang terbentuk, dengan demikian diharapkan plantling
menjadi lebih tahan dan vigor bibit meningkat, sedangkan bio-charcoal akan berperan dalam
menyerap zat-zat toksis, pembenah media dalam menejemen unsur hara tanah dan memberikan
efek gelap di sekitar akar plantling sehingga auksin IAA di akar tidak rusak oleh cahaya dan IAA
akan berperan optimal mendukung perkembangan akar adventif plantling. Sementara itu dengan
adanya lignit maka asam humat akan berperan menyediakan nutrisi hara secara autotropik bagi
plantling di media pertumbuhan.
Sejauh ini pemanfaatan FMA, kompos, bio-charcoal, dan lignit untuk perbaikan
dan peningkatkan pertumbuhan bibit gaharu G. versteegii hasil multiplikasi in vitro yang
mampu beradaptasi di greenhouse dan di lapangan belum dilakukan. Kerjasama sinergis
beberapa perlakuan diharapkan dapat membuat terobosan baru dalam penyediaan bibit
gaharu yang berkualitas (genetik, fisik dan fisiologis). Tuiuan dari penelitian ini adalah
untuk mengetahui jenis-jenis Fungi Mikoriza Arbuskula (FMA) indigenous yang
bersimbiosis dengan G. versteegii asal Papua dan uji kompatibilitas FMA indigenous
Papua terhadap plantling Gyrinops versteegii hasil multiplikasi in-vitro pada berbagai
kombinasi media. Hasil penelitian ini diharapkan dapat memberikan sejumlah data dan
informasi keragaman FMA indigenous yang bersimbiosis dengan pohon G versteegii dan
mendapatkan kombinasi FMA indigenous dengan kompos, bio-charcoal dan lignit yang
efektif untuk produksi bibit gaharu bermutu serta peningkatan kualitas pertumbuhannya.

vi
Hasil isolasi dari trapping menunjukkan bahwa jumlah spora FMA alami
bervariasi dari 2 hingga 5 spora /10 gram sampel tanah. Ditemukan 7 spesies FMA dari 5
plot pengambilan sampel di hutan alam Asai yang bersimbiosis dengan G. Versteegii,
yaitu Glomus mossae, Glomus fasciculatum, Glomus aggregatum, Glomus sp1, Glomus
sp2, Glomus sp3. dan Acaulospora longula. Tingkat kolonisasi FMA alam pada akar
semai G. Versteegii sebesar 30 % hingga 80 % atau tingkat kolonisasi rendah hingga
tinggi. Struktur kolonisasi FMA yang ditemui berupa hifa internal, vesikula, dan spora
dalam akar. Inokulum FMA konsorsium asal Hutan Alam Asai memberikan respon positif
terhadap parameter pertumbuhan plantling Gyrinops lebih baik dari pada FMA
Gigasspora margarita. FMA berpengaruh nyata pada semua parameter diamati yaitu
kolonisasi, persentase hidup, tinggi, diameter, jumlah akar primer dan sekunder, panjang
akar primer dan sekunder, berat basah pucuk, berat basah akar, dan kekokohan bibit.
Media berpengaruh nyata terhadap kolonisasi, tinggi, diameter, jumlah akar primer,
panjang akar primer dan kekokohan bibit. Interaksi antara media dan FMA tidak
berpengaruh terhadap semua parameter yang diamati. Namun demikian FMA konsorsium
memberikan kontribusi relatif lebih baik dibanding FMA G. margarita maupun tanpa
pemberian FMA (kontrol). Besarnya kontribusi FMA konsorsium memberikan pengaruh
nyata terhadap berbagai parameter yang diamati diduga disebabkan karena secara alami
FMA konsorsium telah membangun simbiosis dengan tanaman inang G. versteegii.
Dalam penelitian ini keeratan hubungan antara FMA konsorsium dengan inang G.
versteegii dapat dilihat dari besarnya nilai persentase kolonisasi FMA konsorsium (32,8
%) sedangkan G. margarita (18,7 %), PGR FMA konsorsium (7,1 %) sedangkan G.
Margarita (2,1 %) dan DPU untuk FMA konsorsium (5,4 %) sedangkan DPU G.
Margarita (1,5 %). Indikator lain yang dapat dipakai untuk untuk melihat hubungan
simbiosis ini adalah nilai serapan hara P, dimana FMA konsorsium meningkatkan serapan
hara P pada jaringan tanaman plantling G. versteegii lebih baik dari G. margarita..
Penggunaan inokulum konsorsium alami lebih menjanjikan untuk pertumbuhan suatu jenis
yang diperbanyak secara in-vitro. Geometri akar menunjukan bahwa FMA berpengaruh
terhadap panjang dan jumlah akar primer. FMA konsorsium menghasilkan panjang akar
primer (7 cm) sedangkan G. Margarita (3.1 cm). FMA konsorsium menghasilkan jumlah
akar primer (16,2 akar) sedangkan G. Margarita (11, 1 cm). Berdasarkan nilai kekokohan
dan indeks mutu benih maka media tanah, pasir, kompos, bio-charcoal dan batu bara
muda menghasilkan bibit G. Versteegii lebih siap di lapangan.

vii
@Hak cipta milik IPB, tahun 2011
Hak cipta dilindungi undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumbernya.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan
karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu
masalah.
b. Pengutipan tidak merugikan kepentingan yang wajar IPB.
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
tulis dalam bentuk apapun tanpa izin IPB.

viii
PENGARUH FUNGI MIKORIZA ARBUSKULA (FMA)
DAN MEDIA TUMBUH TERHADAP PERTUMBUHAN PLANTLING GAHARU
(Gyrinops versteegii (Gilg) Domke) HASIL MULTIPLIKASI IN-VITRO
KUSRINER F. MBAUBEDARI
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Pascasarjana Institut Pertanian Bogor
SEKOLAH PASCASARJANA
PROGRAM STUDI ILMU PENGETAHUAN KEHUTANAN
INSTITUT PERTANIAN BOGOR
BOGOR
2011

ix
Judul Penelitian : Pengaruh Fungi Mikoriza Arbuskula (FMA) dan Media Tumbuh
terhadap Pertumbuhan Plantling Gaharu (Gyrinops versteegii
(Gilg) Domke) Hasil Multiplikasi In-Vitro
Nama : Kusriner F. Mbaubedari
NRP : E051060201
Program Studi : Ilmu Pengetahuan Kehutanan
Disetujui
Komisi Pembimbing
Dr. Ir. Irdika Mansur, M. For. Sc Dr. Ir. Supriyanto, DEA
Ketua Anggota
Diketahui
Ketua Program Studi Dekan Sekolah Pascasarjana IPB
Ilmu Pengetahuan Kehutanan
Prof. Dr. Ir. Fauzi Febrianto, MS Dr. Ir. Dahrul Syah, M. Sc. Agr
Tanggal Ujian: 1 Agustus 2011 Tanggal Lulus : 1 Agustus 2011

x
Penguji Luar Komisi pada Ujian Tesis: Dr. Ir. Sri Wilarso Budi R, M.S.

xi
PRAKATA
Puji syukur kepada Tuhan Yang Maha Kuasa atas Rahmat-Nya sehingga saya dapat
menghasilkan tesis berjudul Pengaruh Fungi Mikoriza Arbuskula (FMA) dan Media
Tumbuh terhadap Pertumbuhan Plantling Gaharu (Gyrinops versteegii (Gilg) Domke)
Hasil Multiplikasi In-vitro) ini dapat diselesaikan.
Pada kesempatan ini penulis menyampaikan terima kasih kepada Rektor Universitas
Negeri Papua atas kesempatan yang diberikan untuk melanjutkan studi ke Program Studi
Ilmu Pengetahuan Kehutanan Sekolah Pascasarjana Institut Pertanian Bogor. Terima kasih
juga disampaikan kepada Direktor Jenderal Pendidikan Tinggi (Dirjen DIKTI) atas
program pendidikan Pascasarjana BPPS T.A. 2006-2008 dan Pemerintah Propinsi Papua
atas Program Beasiswa Otonomi Khusus T. A. 2009-2010.
Ucapkan terima kasih serta penghargaan yang tulus disampaikan kepada Dr. Ir.
Irdika Mansur, M.For.Sc dan Dr. Ir. Supriyanto, DEA selaku komisi pembimbing yang
telah membantu mengarahkan, memberikan semangat dan perhatian khusus sejak awal
penentuan topik penelitian tesis hingga penyelesaian karya ilmiah ini. Pada kesempatan
ini, penulis juga menyampaikan terima kasih yang tulus kepada Dr. Ir. Sri Wilarso Budi R,
M.S selaku penguji luar komisi yang karena kesediaannya sebagai penguji sehingga ujian
tesis saya dapat terselenggarakan dengan baik.
Semoga Karya Ilmiah ini dapat bermanfaat untuk perkembangan teknik silvikultur
gaharu dan mikoriza di Indonesia.
Bogor, 1 Agustus 2011
Kusriner Fernando Mbaubedari

xii
RIWAYAT HIDUP
Penulis dilahirkan di Serui, Kecamatan Yapen Selatan, Kabupaten Yapen
Waropen, Propinsi Papua pada tanggal 17 Februari 1975. Penulis merupakan anak
pertama dari tujuh bersaudara dari ayah Fredrik Mbaubedari dan ibu Benselina Reba .
Tahun 1995 penulis lulus dari SMUN 417 Serui, pada tahun yang sama lulus seleksi
masuk Universitas Cenderawasih melalui jalur seleksi lokal siswa berpotensi. Penulis
memilih Program Studi Budidaya Hutan, Jurusan Kehutanan, Fakultas Pertanian
Universitas Cenderawasi (UNCEN). Penulis menamatkan pendidikan Sarjana tahun 2001.
Selama menjadi mahasiswa S1, penulis bekerja sebagai asisten pada mata kuliah Silvika
dan Ilmu Tanah Hutan, dan aktif dalam berbagai kegiatan kemahasiswaan. Pada tahun
2003, penulis diterima sebagai pengajar pada Universitas Papua di Manokwari, Papua
Barat,.

xiii
DAFTAR ISI
Halaman
PRAKATA ------------------------------------------------------------------------------- xi
DAFTAR ISI ---------------------------------------------------------------------------- xiii
DAFTAR TABEL ------------------------------------------------------------------------ xv
DAFTAR GAMBAR ------------------------------------------------------------------ xvi
DAFTAR LAMPIRAN ----------------------------------------------------------------- xvi
PENDAHULUAN ----------------------------------------------------------------------- 1
Latar Belakang -------------------------------------------------------------------- 1
Perumusan Masalah --------------------------------------------------------------- 3
Tujuan ------------------------------------------------------------------------------ 5
Manfaat ---------------------------------------------------------------------------- 5
Hipotesis --------------------------------------------------------------------------- 5
Kerangka Pemikiran -------------------------------------------------------------- 6
TINJAUAN PUSTAKA ---------------------------------------------------------------- 7
Gaharu ------------------------------------------------------------------------------- 7
Proses Pembentukan Gaharu ------------------------------------------------- 8
Gaharu Unggul ---------------------------------------------------------------- 10
Perbanyakan Pohon Penghasil Gaharu -------------------------------------- 10
Manfaat dan Nilai Ekonomi Gaharu ---------------------------------------- 11
Mikoriza ----------------------------------------------------------------------------- 12
Fungi Mikoriza Arbuskula --------------------------------------------------------- 14
Manfaat Mikoriza ------------------------------------------------------------------- 16
Mikoriza dan Gaharu G. versteegii ----------------------------------------------- 17
Bio-charcoal ------------------------------------------------------------------------ 17
Lignit (Batu Bara Muda) ----------------------------------------------------------- 19
METODE PENELITIAN ------------------------------------------------------------- 21
Tempat dan Waktu ---------------------------------------------------------------- 21
Bahan dan Alat --------------------------------------------------------------------- 21
Metode ------------------------------------------------------------------------------ 21
Prosedur Pelaksanaan --------------------------------------------------------- 22
Isolasi dan Identifikasi FMA ------------------------------------------------- 22
Produksi Inokulum FMA ----------------------------------------------------- 25
Aplikasi FMA, Kompos, Bio-charcoal dan Lignit pada Plantling -------- 25
Pengamatan dan Pengukuran ------------------------------------------------- 25
Analisa Data ------------------------------------------------------------------------ 27
HASIL ------------------------------------------------------------------------------------ 28
Kondisi Lokasi Pengambilan Sampel ------------------------------------------ 28
Vegetasi ------------------------------------------------------------------------- 28
Keadaan Tanah ----------------------------------------------------------------- 30
Keadaan Iklim ------------------------------------------------------------------ 30

xiv
Potensi Fungi Mikoriza Arbuskular Alami di Semai G. versteegii di Hutan
Alam Asai --------------------------------------------------------------------------- 31
Keberadaan jenis FMA -------------------------------------------------------- 31
Kolonisasi FMA alami pada semai G. versteegii --------------------------- 34
Aplikasi FMA pada Plantling Gaharu G. versteegii --------------------------- 36
Kondisi stomata plantlet dan plantling G. verstegii ----------------------- 36
Rekapitulasi hasil uji F terhadap berbagai parameter pengamatan ------- 37
Kolonisasi akar pada plantling G. versteegii ------------------------------- 39
Respon plantling G. versteegii terhadap mikoriza (percentage growth
respon, PGR) -------------------------------------------------------------------- 43
Ketergantungan plantling G. versteegii bermikoriza terhadap fosfor
(dependency of P uptake, DPU) ---------------------------------------------- 44
Persentase hidup plantling G. versteegii ------------------------------------- 46
Pertambahan tinggi plantling G. versteegii ---------------------------------- 48
Pertambahan diameter plantling G. versteegii ------------------------------ 51
Geometri akar ------------------------------------------------------------------- 53
Berat basah pucuk dan akar --------------------------------------------------- 57
Kekokohan plantling G. versteegii ------------------------------------------- 59
Hubungan antara kolonisasi FMA dengan parameter pertumbuhan
plantling G. versteegii ---------------------------------------------------------- 61
Searapan hara makro N, P dan K --------------------------------------------- 61
Indeks mutu bibit -------------------------------------------------------------- 63
PEMBAHASAN ------------------------------------------------------------------------- 64
Pengaruh FMA --------------------------------------------------------------------- 64
Pengaruh Media ------------------------------------------------------------------- 68
Pengaruh interaksi media tumbuh dengan FMA -------------------------- ------ 69
SIMPULAN DAN SARAN ------------------------------------------------------------- 70
DAFTAR PUSTAKA -------------------------------------------------------------------- 71
LAMPIRAN ------------------------------------------------------------------------------- 75

xv
DAFTAR TABEL
NO. Halaman
Teks
1. Keberadaan G. versteegii dan vegetasi lain dan pada lokasi pengambilan
sampel ------------------------------------------------------------------------------ 28
2. Hasil analisis tanah di lokasi penelitian ----------------------------------------- 30
3. Dokumentasi jenis spora FMA yang bersimbiosis dengan semai G.
versteegii dari hutan alam Asai, Manokwari Papua Barat ------------------ 32
4. Rekapitulasi sidik ragam pengaruh FMA terhadap pertumbuhan plantling
G. versteegii ------------------------------------------------------------------------ 38
5. Hasil análisis korelasi antara kolonisasi FMA dengan beberapa parameter
pertumbuhan plantling G. versteegii -------------------------------------------- 61

xvi
DAFTAR GAMBAR
NO. Halaman
Teks
1. Kerangka pemikiran pengembangan bibit gaharu potensial ----------------- 6
2. Distribusi pohon penghasil gaharu Aquilaria spp. dan Gyrinops spp. di
Papua, Indonesia dan dunia ------------------------------------------------------- 8
3. Empat faktor yang mempengaruhi terbentuknya gubal gaharu -------------- 9
4. Skema hirarki yang terdiri dari tipe-tipe mikoriza dan kategori interaksi
tumbuhan dengan cendawan lainnya ------------------------------------------- 14
5. Pohon gaharu G. versteegii dan vegetasi lain yang tumbuh disekitar G.
versteegii (a) dan keberadaan semai G. Versteegii --------------------------- 29
6. Sebaran jumlah spora FMA per plot pengambilan sampel --------------------- 34
7. Struktur kolonisasi FMA dengan akar semai G. versteegii --------------------- 35
8. Stomata di daun muda (a) dan di daun tua (b) pada planlet G. versteegii asal
kultur in-vitro dalam keadaan membuka dan sedikit ---------------------------- 37
9. Stomata daun muda (a) dan daun tua (b) pada plantling G. versteegii
dalam keadaan menutup dan jumlahnya lebih banyak ------------------------ 37
10. Visualisasi persentase kolonisasi FMA dengan plantling G. versteegii ----- 39
11.` Hasil Uji Duncan persentase kolonisasi FMA pada akar G. versteegii
(angka diikuti huruf yang sama berarti tidak ada perbedaan antar FMA yang
dicobakan pada α=0,05) ---------------------------------------------------------- 40
12. Hasil uji Duncan pengaruh media terhadap persentase kolonisasi pada akar
plantling G. versteegii (angka diikuti huruf yang sama berarti tidak ada
perbedaan antar FMA yang dicobakan pada α=0,05) ------------------------- 40
13. Visualisasi struktur kolonisasi FMA pada akar plantling G. versteegii ---- 41
14. Visualisasi respon plantling G. versteegii terhadap mikoriza --------------- 43
15. Hasil Uji Duncan PGR pada plantling G. versteegii (angka diikuti huruf
yang sama berarti tidak ada perbedaan antar FMA yang dicobakan pada
α=0,05). ------------------------------------------------------------------------------ 44
16. Visualisasi ketergantungan plantling G. versteegii bermikoriza terhadap
fosfor ------------------------------------------------------------------------------- 45
17. Hasil Uji Duncan DPU pada plantling G. versteegii (angka diikuti huruf
yang sama berarti tidak ada perbedaan antar FMA yang dicobakan pada
α=0,05) ------------------------------------------------------------------------------- 45

xvii
18. Visualisasi persentase hidup plantling G. versteegii sampai minggu ke-8 -- 46
19. Visualisasi kematian plantling G. versteegii akibat serangan busuk akar - 47
20. Hasil Uji Duncan hidup plantling G. versteegii (angka diikuti huruf yang
sama berarti tidak ada perbedaan antar FMA yang dicobakan pada α=0,05) 47
21. Visualisasi keadaan tinggi plantling G. versteegii pada berbagai kombinasi
media yang dicobakan ------------------------------------------------------------ 48
22. Hasil Uji Duncan terhadap tinggi plantling G. versteegii (angka diikuti
huruf yang sama berarti tidak ada perbedaan antar FMA yang dicobakan
pada α=0,05) ----------------------------------------------------------------------- 49
23. Hasil uji Duncan pengaruh media tumbuh terhadap tinggi plantling G.
versteegii. (angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05) --------------------------------------------- 49
24. Pengaruh kombinasi media tumbuh terhadap pertambahan tinggi plantling
G. Versteegii ----------------------------------------------------------------------- 50
25. Keragaan pertumbuhan tinggi plantling G. versteegii ------------------------ 50
26. Keragaan pertumbuhan plantling G. versteegii umur 8 MST dari semua
kombinasi perlakuan -------------------------------------------------------------- 51
27. Hasil Uji Duncan pengaruh FMA terhadap pertumbuhan diameter plantling
G. versteegii (Angka diikuti huruf yang sama berarti tidak ada perbedaan
antar FMA yang dicobakan pada α=0,05) -------------------------------------- 51
28. Hasil uji Duncan pengaruh media tumbuh terhadap diameter plantling G.
versteegii --------------------------------------------------------------------------- 52
29. Visualisasi pertambahan diameter plantling G. versteegii pada berbagai
kombinasi media tumbuh --------------------------------------------------------- 53
30. Hasil Uji Duncan pengaruh FMA terhadap jumlah akar primer dan
sekunder plantling G. versteegii (Angka diikuti huruf yang sama berarti
tidak ada perbedaan ----------------------------------------------------------------- 53
31. Hasil Uji Duncan pengaruh media terhadap jumlah akar primer plantling G.
versteegii (Angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05) --------------------------------------------- 54 53
32. Visualisasi pengaruh kombinasi FMA dan media tumbuh terhadap panjang
akar primer dan sekunder plantling G. Versteegii ----------------------------- 55

xviii
33. Hasil Uji Duncan pengaruh FMA terhadap panjang akar primer dan
sekunder plantling G. versteegii (Angka diikuti huruf yang sama berarti
tidak ada perbedaan antar FMA yang dicobakan pada α=0,05) ------------- 55
34. Hasil Uji Duncan pengaruh media terhadap panjang akar primer plantling
G. versteegii (Angka diikuti huruf yang sama berarti tidak ada perbedaan
antar FMA yang dicobakan pada α=0,05) -------------------------------------- 56
35. Visualisasi pengaruh kombinasi FMA dan media tumbuh terhadap panjang
akar primer dan sekunder plantling G. Versteegii ----------------------------- 56
36. Keragaan panjang akar primer (A) dan akar sekunder (B) plantling G.
Versteegii --------------------------------------------------------------------------- 57
37. Hasil Uji Duncan terhadap berat basah akar dan pucuk plantling G.
versteegii (angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05) --------------------------------------------- 58
38. Visualisasi pengaruh kombinasi FMA dan media tumbuh terhadap berat
basah akar dan pucuk plantling G. Versteegii ---------------------------------- 58
39. Hasil Uji Duncan pengaruh FMA terhadap kekokohan plantling G.
versteegii (angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05) --------------------------------------------- 59
40. Hasil Uji Duncan pengaruh media terhadap kekokohan plantling G.
versteegii --------------------------------------------------------------------------- 59
41. Visualisasi pengaruh FMA dan media terhadap kekokohan plantling G.
versteegii --------------------------------------------------------------------------- ` 60
42. Hasil analisis serapan hara makro N pada jaringan tanaman plantling G.
Versteegii --------------------------------------------------------------------------- 62
43. Hasil analisis serapan hara makro P pada jaringan tanaman plantling G.
Versteegii -------------------------------------------------------------------------- 62
44. Hasil analisis serapan hara makro K pada jaringan tanaman plantling G.
Versteegii --------------------------------------------------------------------------- 62

xix
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Persentase kolonisasi FMA hasil isolasi dari sampel tanah
komposit semai pohon penghasil gaharu G. versteegii dari hutan
alam Asai Kabupaten Manokwari, Propinsi Papua Barat ------------ 75
Lampiran 2. Rekapitulasi Uji Signifikasi terhadap PGR dan DPU plantling G.
Versteegii ----------------------------------------------------------------- 76
Lampiran 3. Uji Signifikasi terhadap PGR dan DPU plantling G. Versteegii --- 78
Lampiran 4. Hasil Uji Duncan persentase kolonisasi FMA pada plantling G.
versteegii ----------------------------------------------------------------- 78
Lampiran 5. Hasil Uji Duncan terhadap Nilai PGR plantling G. versteegii -- 78
Lampiran 6. Hasil Uji Duncan terhadap Nilai DPU plantling G. versteegii --- 79
Lampiran 7. Hasil Uji Duncan Pengaruh FMA terhadap persentase hidup
plantling G. versteegii --------------------------------------------------- 79
Lampiran 8. Hasil Uji Duncan Pengaruh FMA terhadap Tinggi plantling G.
Versteegii ------------------------------------------------------------------ 79
Lampiran 9. Hasil Uji Duncan Pengaruh FMA terhadap Diameter plantling G.
versteegii ----------------------------------------------------------------- 80
Lampiran 10. Hasil Uji Duncan t Pengaruh FMA terhadap Jumlah Akar Primer
plantling G. Versteegii ------------------------------------------------- 80
Lampiran 11. Hasil Uji Duncan Pengaruh FMA terhadap Jumlah Akar
Sekunder plantling G. versteegii -------------------------------------- 80
Lampiran 12. Hasil Uji Duncan Pengaruh media terhadap Jumlah Akar Primer
plantling G. versteegii -------------------------------------------------- 80
Lampiran 13. Hasil Uji Duncan pengaruh media terhadap Panjang Akar Primer
plantling G. Versteegii -------------------------------------------------- 81
Lampiran 14. Hasil Uji Duncan Pengaruh FMA terhadap Panjang Akar primer
plantling G. Versteegii -------------------------------------------------- 82
Lampiran 15. Hasil Uji Duncan Pengaruh FMA terhadap Panjang Akar
Sekunder plantling G. versteegii --------------------------------------- 83
Lampiran 16. Hasil Uji Duncan Pengaruh FMA terhadap Berat Basah Akar
plantling G. Versteegii --------------------------------------------------- 83

xx
Lampiran 17. Hasil Uji Duncan Pengaruh FMA terhadap Berat Basah Pucuk
plantling G. versteegii ---------------------------------------------------- 83
Lampiran 18. Hasil Uji Duncan Pengaruh FMA terhadap Kekokohan Bibit
plantling G. versteegii ---------------------------------------------------- 84
Lampiran 19. Indeks Mutu Bibit berdasarkan rata-rata diameter plantling G.
versteegii menggunakan sitem skoring ------------------------------- 84
Lampiran 20. Indeks Mutu Bibit berdasarkan rata-rata tinggi plantling G.
versteegii menggunakan sistem skoring ----------------------------- 85
Lampiran 21. Indeks Mutu Bibit berdasarkan rata-rata tingkat kekokohan
plantling G. versteegii menggunakan sitem skoring ---------------- 85
Lampiran 23. Nilai serapan hara N, P dan K pada plantling G. Versteegii -------- 86
Lampiran 24. Hasil Analisis Media Tumbuh Plantling G. versteegii -------------- 87
Lampiran 25. Hasil analisis jaringan plantling G. versteegii pada kombinasi
perlakuan media tumbuh ----------------------------------------------- 87

21
PENDAHULUAN
Latar Belakang
Gaharu merupakan Hasil Hutan Bukan Kayu (HHBK) bernilai ekonomi tinggi,
berwarna khas, mengandung aroma resin wangi jika dibakar dan dapat digunakan untuk
bahan parfum, dupa, obat-obatan, sabun mandi, kosmetik, dan pengharum ruangan.
Dengan demikian gaharu merupakan Hasil Hutan Bukan Kayu memiliki nilai ekonomi
yang tinggi.
Permintaan gaharu dunia saat ini diperkirakan 4000 ton pertahun namun Indonesia
hanya mampu menyediakan 200 ton pertahun dari potensi gaharu Indonesia yang
diperkirakan sebesar 600 ton pertahun. Potensi sebesar ini diambil dari hutan alam Papua,
Kalimantan, Sulawesi, Nusa Tenggara dan Sumatera meliputi 98% dan sisanya 2%
berasal dari perkebunan gaharu. Harga gaharu di Indonesia berkisar Rp 100.000-
150.000.000 per kg tergantung kualitasnya. Sementara itu harga gaharu super di China
dapat mencapai Rp 400 juta per kilogram dan 300 juta per kilogram di Timur Tengah
(Mashur, 2011). Total ekspor gaharu Indonesia dalam lima tahun terakhir berkisar 170-
573 ton/tahun dengan perkiraan perolehan devisa negara pada tahun 2006 sebesar
26.086.350 USD meningkat menjadi 85.987.500 USD pada tahun 2010. Menurut Menteri
Kehutanan RI, untuk memenuhi permintaan pasar dunia, maka eksport gaharu Indonesia
akan ditingkatkan menjadi 1000 ton/tahun yang didukung oleh pembangunan hutan
tanaman gaharu (Zulkifli, 2011).
Indonesia telah dikenal sebagai salah satu negara penghasil gaharu di dunia, karena
mempunyai lebih dari 25 jenis pohon penghasil gaharu yang tersebar di Sumatera,
Kalimantan, Sulawesi, Nusa Tenggara, Maluku dan Papua. G versteegii, Gyrinops
ladermanii dan Aquilaria filaria merupakan spesies penghasil gaharu yang dapat
ditemukan di New Guinea (The State Papua New Guinea and West Papua in Indonesian)
dan telah dikenal di dunia (Gunn et al., 2003).
Beragamnya permintaan produk berbahan baku gaharu dan tingginya harga gaharu
diperdagangan internasional menyebabkan perburuan gaharu (Aquilaria filaria dan G.
versteegii) menyebar sampai hutan-hutan alam Papua yang masih tersisa. Penebangan
pohon secara tidak selektif menyebabkan pohon yang tidak mengandung gaharu pun
ditebang. Hal tersebut disebabkan karena sulitnya mendekteksi keberadaan gubal gaharu
pada pohon gaharu yang masih hidup. Untuk memproleh satu pohon penghasil gubal
gaharu, pemburu gaharu dapat menebang 10 pohon gaharu. Sisa pohon gaharu di daerah-

22
daerah penghasil utama gaharu semakin lama semakin menipis antara lain Sumatera
(26%), Kalimantan (27%), Nusa Tenggara (5%), Sulawesi (4%), Maluku (6%), dan Papua
(37%) (Asgarin 2011). Semakin sulitnya mendapatkan gaharu di hutan alam
mengakibatkan Aquilaria spp. dan Gyrinops spp. dimasukkan dalam Apendix II (produksi
gaharu harus berasal dari perkebunan gaharu) pada konvensi CITES (Convention on
International Trade in Endangered Species of Wild Fauna and Flora) di Bangkok (Cites,
2004). Kekuatiran akan punahnya spesies gaharu di Indonesia, menyebabkan Kementerian
Kehutanan Indonesia sejak tahun 2005 menurunkan kuota ekspor gaharu menjadi hanya
125 ton/tahun.
Eksploitasi gaharu tanpa upaya budidaya menyebabkan potensi gaharu di hutan
alam semakin berkurang. Pola ini dapat diperbaiki melalui upaya konservasi,
pembangunan hutan tanaman gaharu yang didukung dengan tersedianya bibit unggul dan
teknologi bioproses gaharu yang efektif serta aplikasi bioteknologi pupuk hayati dan
bahan organik dalam meningkatkan pertumbuhan tanaman di lapangan melalui produksi
bibit berkualitas.
Pohon berpotensi menghasilkan gaharu dapat diperbanyak melalui kultur jaringan
dan dikembangkan dengan mikoriza untuk memacu pertumbuhannya pada media yang
tepat. Media tersebut dapat berupa, kompos, bio-charcoal dan lignit (batubara muda).
FMA diketahui berperan mengurangi stress yang disebabkan karena minimnya hara, aerasi
tanah kurang baik, struktur tanah padat, pH rendah, salinitas tinggi dan logam beracun
(Sieverding, 1991). Meningkatkan penyerapan unsur hara makro P, N dan beberapa hara
mikro (Baghel et al., 2009). Hifa mikoriza mampu memfilter logam berat dan tidak
meneruskannya ke tanaman (Smith and Read, 1997), melindungi perakaran tanaman dari
patogen berbahaya (Liu, 1991), perlindungan dari senyawa-senyawa radio nuklir (Pfleger
and Linderman, 1996) dan salinitas tanah (Delvian et al, 2001) serta memproduksi hormon
pertumbuhan seperti auksin, sitokinin dan gibberelin bagi tanaman. Dengan adanya auksin
proses penuaan akar menjadi lambat.
Bio-charcoal adalah arang yang dibuat dari material biologis dan diolah melalui
proses pirolisis. Teknologi ini telah lama digunakan oleh suku Indian Maya ribuan tahun
silam. Manfaat bio-charcoal adalah untuk menurunkan emisi rumah kaca, meningkatkan
simpanan karbon dalam tanah, soil conditioner dan soil managment, meningkatkan
porositas tanah dan aktifitas mikroba tanah, menyerap kontaminan toksis, menetralkan
keasaman tanah serta mencegah penyakit busuk akar yang disebabkan oleh serangan
patogen jamur (Steiner, 2007 ; Lehman, 2007 ; Supriyanto, 2010), sedangkan lignit

23
(batubara muda) adalah batubara tidak bernilai ekonomis yang ditimbun di lokasi tambang
sebagai limbah. Diduga lignit mengandung zat-zat humat (asam humat) sebagai unsur
organik dari proses dekomposisi tumbuhan dalam tanah, sedimen rawa dan gambut
sehingga dapat memperkaya media pertumbuhan bibit. Adanya peran potensial dari
mikoriza, kompos, bio-charcoal dan lignit, maka berpotensi dapat dimanfaatkan untuk
memperkaya media pertumbuhan plantling gaharu G. versteegii.
Perumusan Masalah
Untuk mendapatkan pohon penghasil gubal gaharu yang baik, harus menggunakan
bibit dari pohon gaharu potensial, yaitu bibit unggul dari pohon inang yang telah terbukti
menghasilkan gubal gaharu di alam, artinya sangat dipengaruhi oleh sifat genetik,
kemudian diperbanyak sebagai bibit unggul (clone) untuk kepentingan budidaya. Namun
demikian produktifitas benih yang rendah menyebabkan kesulitan memperoleh anakan
pohon gaharu dalam jumlah banyak di alam, padahal untuk tujuan budidaya yang luas
sangat diperlukan bibit berkualitas dalam jumlah cukup dan tersedia tepat waktu. Di sisi
lain, selama ini bibit yang digunakan berasal dari biji atau semai hutan alam dimana
jumlah bibit terbatas, kualitas bibit rendah dan peluang memperoleh gubal gaharu setelah
penanaman relatif kecil karena bibit yang dipakai belum tentu berasal dari induk yang
berpontesi menghasilkan gubal gaharu.
Kendala lain yang umumnya dihadapi adalah tidak semua pohon gaharu
menghasilkan buah setiap tahun, belum adanya kebun bibit unggul dan kebun benih serta
biji gaharu bersifat rekalsitran, selain itu adanya penebangan pohon induk dewasa di alam
oleh pencari gaharu menyebabkan hilangnya sumber benih. Rendahnya daya berbunga dan
produktifitas berbuah menyebabkan masalah regenerasi secara generatif, sementara itu
pembiakan secara vegetatif menggunakan stek dan cangkok membutuhkan bahan induk
yang banyak, maka kultur in vitro gaharu menjadi alternatif teknologi perbanyakan gaharu
unggul secara masal dan cepat.
Selain dapat menghasilkan bibit berkualitas dalam jumlah yang memadai, teknik
ini berpotensi mempertahankan sifat genetis dari pohon induk penghasil gaharu. Namun
demikian, ditemukan kendala dalam kultur jaringan gaharu yaitu memerlukan waktu yang
lama untuk menginduksi akar plantlet baik secara in vitro maupun ex vitro di green house,
sementara akar yang terbentuk umumnya relatif kecil, jumlahnya sedikit serta sulit
berkembang. Problem lain adalah tahapan aklimatisasi plantling gaharu hasil in vitro ke
media aklimatisasi merupakan tahapan kritis yang masih menjadi masalah karena

24
plantling telah terbiasa tumbuh pada kondisi lingkungan dengan kelembaban tinggi
sementara apabila diadaptasikan secara ex vitro dengan temperatur tinggi plantling akan
mengalami dehidrasi, layu dan mati dikarenakan mekanisme buka tutup stomata yang
rendah dan stomata tidak berfungsi optimal. Lingkungan tumbuh in vitro bersifat steril
mengakibatkan imunitas plantling rendah dan lebih rentan terhadap serangan hama dan
penyakit busuk akar oleh jamur pembusuk akar (lodoh). Akibatnya tingkat kematian
plantling gaharu sangat tinggi, yaitu 75 % dari plantlet yang diadaptasikan ke media
pertumbuhan plantling di green house. Kondisi ini menyebabkan kegagalan produksi bibit
gaharu potensial asal kultur in vitro. Penyebab lainnya yaitu lapisan kutikula plantling
tipis dan terbiasa menerima cahaya konstan, sedangkan pada ex vitro plantling
mendapatkan cahaya tidak konstan dan intensitas cahaya tinggi. Faktor lainnya ialah
dalam media in vitro nutrisi tersedia optimal sehingga plantlet bersifat heterotrop,
sedangkan diluar media kultur plantling dituntut bersifat autotrop. Selain itu, plantling
asal in vitro memiliki lignifikasi batang rendah sehingga cenderung bersifat vitrious
(sukulen).
Aplikasi FMA pada plantling gaharu akan memacu ramifikasi akar dan batang
plantling karena mikorhia memproduksi auksin IAA untuk menstimulir akar plantling
serta menghasilkan enzim phospatase untuk pembentukan lignin. Dengan demikian
diharapkan plantling menjadi lebih tahan dan vigor bibit meningkat, sedangkan bio-
charcoal akan berperan dalam menyerap zat-zat toksis, pembenah media dalam menejemen unsur
hara tanah dan memberikan efek gelap disekitar akar plantling sehingga auksin IAA di akar tidak
rusak oleh cahaya dan IAA akan berperan optimal mendukung perkembangan akar adventif
plantling. Sementara itu dengan adanya lignit maka asam humat akan berperan menyediakan
nutrisi hara secara ototropik bagi plantling di media pertumbuhan.
Sejauh ini pemanfaatan FMA, kompos, bio-charcoal, dan lignit untuk perbaikan
dan peningkatkan pertumbuhan bibit gaharu G. versteegii hasil multiplikasi in vitro yang
mampu adaptasi di green house dan lapangan belum dilakukan. Kerjasama sinergis
beberapa komponen perlakuan diharapkan dapat membuat terobosan baru dalam
penyediaan bibit gaharu yang berkualitas (genetik, fisik dan fisiologis). Dari uraian di atas,
dirumuskan permasalahan penelitian sebagai berikut :

25
1. Bagaimana keragaman jenis Fungi Mikoriza Arbuskula (FMA) indigenous yang
bersimbiosis dengan gaharu G. versteegii asal Papua?
2. Bagaimana kompatibilitas FMA indigenous Papua terhadap plantling G. versteegii
hasil multiplikasi in-vitro.
Tujuan
Penelitian ini bertujuan untuk :
(1) Mengetahui jenis-jenis Fungi Mikoriza Arbuskula (FMA) indigenous yang
bersimbiosis dengan G. versteegii asal Papua.
(2) Uji kompatibilitas FMA indigenous Papua terhadap plantling G. versteegii hasil
multiplikasi in-vitro pada berbagai kombinasi media.
Manfaat Penelitian
Hasil penelitian ini diharapkan dapat memberikan sejumlah data dan informasi
keragaman FMA indigenous yang bersimbiosis dengan pohon G. versteegii dan
mendapatkan kombinasi FMA indigenous dengan kompos, bio-charcoal dan lignit yang
efektif untuk produksi bibit gaharu bermutu serta peningkatan kualitas pertumbuhannya.
Hipotesis
Berdasarkan rumusan masalah dan tujuan penelitian, ditetapkan hipotesis
sebagai berikut :
a. Ditemukan lebih dari satu jenis FMA indigenous yang bersimbiosis dengan gaharu
alami G. versteegii di Papua.
b. Kompatibilitas FMA indigenous Papua dapat meningkatkan daya hidup dan
pertumbuhan plantling gaharu Gyrinops versteegii hasil multiplikasi in vitro pada
berbagai kombinasi media.
26
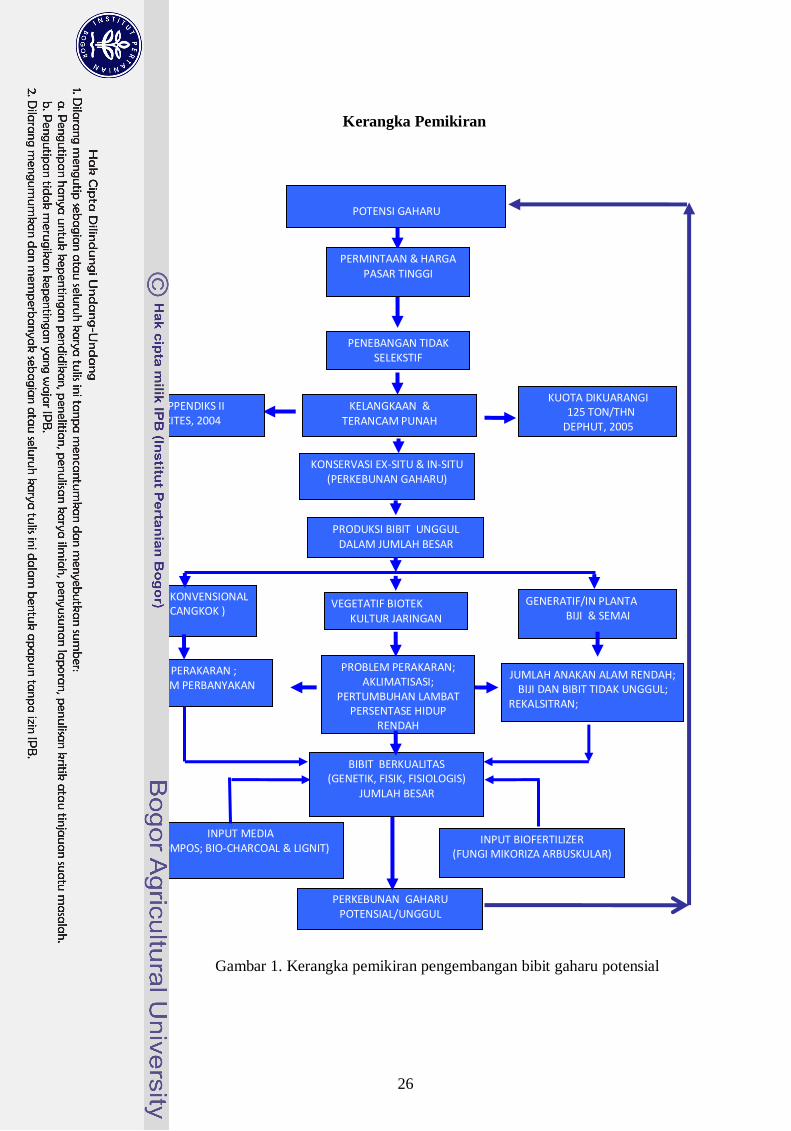
Kerangka Pemikiran
POTENSI GAHARU
KELANGKAAN & TERANCAM PUNAH
KONSERVASI EX-SITU & IN-SITU (PERKEBUNAN GAHARU)
VEGETATIF KONVENSIONAL (STEK, CANGKOK )
GENERATIF/IN PLANTA BIJI & SEMAI
PROBLEM PERAKARAN; AKLIMATISASI;
PERTUMBUHAN LAMBAT PERSENTASE HIDUP
RENDAH
PERKEBUNAN GAHARU POTENSIAL/UNGGUL
BIBIT BERKUALITAS (GENETIK, FISIK, FISIOLOGIS)
JUMLAH BESAR
PERMINTAAN & HARGA PASAR TINGGI
PENEBANGAN TIDAK SELEKSTIF
KUOTA DIKUARANGI 125 TON/THN DEPHUT, 2005
APPENDIKS II CITES, 2004
PROBLEM PERAKARAN ; PROBLEM PERBANYAKAN
INPUT MEDIA (KOMPOS; BIO-CHARCOAL & LIGNIT)
VEGETATIF BIOTEK KULTUR JARINGAN
INPUT BIOFERTILIZER (FUNGI MIKORIZA ARBUSKULAR)
PRODUKSI BIBIT UNGGUL DALAM JUMLAH BESAR
JUMLAH ANAKAN ALAM RENDAH; BIJI DAN BIBIT TIDAK UNGGUL;
REKALSITRAN;
Gambar 1. Kerangka pemikiran pengembangan bibit gaharu potensial

27
TINJAUAN PUSTAKA
Gaharu
Gaharu berasal dari bahasa Sangsekerta disebut “aguru” yang artinya
“tenggelam” atau bahasa Melayu yang artinya “harum” (Sumarna, 2007). Dalam
perdagangan internasional dikenal dengan nama ”agarwood, eaglewood, aloeswood”
(Gunn et al., 2003), yaitu sebutan untuk hasil hutan non kayu yang berupa damar wangi
(aromatic resin) dari genus Aquilaria dan genus lain pada famili Thymelaeaceae,
sedangkan gubal gaharu adalah substansi aromatik berupa gumpalan kayu berwarna coklat
muda, coklat kehitaman sampai hitam yang terbentuk pada pohon gaharu akibat infeksi
mikroorganisme penyebab penyakit.
Family Thymelaceae ini terdiri dari Gonystyloideae, Aquilariodeae,
Thymelaeiodeae, Gilgiodaphniodeae. Beberapa genus anggota Thymeleaceae penghasil
gaharu adalah Aetoxylon sp, Aquilaria spp., Enkleia sp, Gonystilus sp dan Wilkstromeia sp
(Hou, 1960). Diantara genus pohon tersebut, Aquilaria spp. diketahui sebagai penghasil
gubal gaharu yang terbaik, demikian juga 2 jenis Gyrinops spp., yaitu G. versteegii dan
G. ledermanii (Rahayu & Sitomorang, 2004).
Telah diketahui ada 24 spesies Aquilaria spp. tersebar di hutan tropis Asia mulai
dari Banglades, India, Pakistan, Myanmar, Laos, Thailand, Singapura, Kamboja, Cina
Selatan, Malaysia, Philipina dan Indonesia. Enam spesies diantaranya yaitu Aquilaria
beccariana, A. cumingiana, A. filaria, G. versteegii, A. malaccensis, A. microcarpa dan A.
hirta ditemukan di wilayah Indonesia Asia (Hou, 1960; Cites, 2004), sedangkan 7 spesies
Gyrinops spp., 6 diantaranya terdapat di Indonesia timur dan satu spesies terdapat di
Srilangka, 20 spesies Gonystilus tersebar di Asia Tenggara mulai dari Malaysia, Serawak,
Sabah, Philipina, Indonesia, Kepulauan Salomon dan Kepulauan Nikobar (Parman dan
Mulyaningsih, 1996).
Pohon penghasil gaharu Gyrinops spp. memiliki distribusi yang cukup luas pada ras
geografi. G. moluccana ditemukan di Maluku dan Halmahera, G. Versteegii ditemukan di
Nusa Tenggara Timur, Nusa Tengara Barat dan Propinsi Papua, sedangkan G. ladermanii,
G. salicifolia, G. audate dan G. podocarpus ditemukan di Propinsi Papua dan Papua New
Guinea. Di Pulau Papua sendiri distribusi populasi G. ladermanii lebih banyak ditemukan
di Papua New Guinea, sedangkan G. versteegii populasi terbanyak di propinsi Papua
Indonesia (Hou, 1960) (Gambar 1).

28
Gambar 2. Distribusi pohon penghasil gaharu Aquilaria spp. dan Gyrinops spp. di Papua,
Indonesia dan dunia.
Proses Pembentukan Gubal Gaharu
Gubal gaharu sering disebut dengan istilah “gaharu” adalah substansi aromatik
berupa gumpalan berwarna coklat muda, coklat kehitaman sampai hitam (Hou, 1960).
Proses terbentuknya gaharu pada pohon penghasil gaharu diduga sebagai akibat proses
patogenesis yang diawali dengan rangsangan luka pada batang pohon, cabang atau ranting
dan pengaruh fisik lainnya. Menurut Goodman et al. (1986) gubal gaharu diduga terkait
dengan mekanisme ketahanan inang terhadap rangkaian proses yang terjadi pada
patogenis tumbuhan berupa metabolit sekunder dari golongan senyawa aromatik.
Metabolit beraroma harum ini umumnya dari golongan sesquiterpenoid dapat terbentuk
pada pohon gaharu akibat adanya luka atau induksi dari mokroorganisme yang
menginfeksinya atau interaksi faktor biotik dan abiotik.
Patogenesis tumbuhan adalah pertarungan antara inang dengan pathogen yang
kompatibel dimana menurut Agrios (1997) keberhasilan proses infeksi oleh suatu pathogen
sehingga dapat menginduksi gaharu dipengaruhi oleh 4 (empat) faktor : 1). Inang yang
rentan, yaitu jenis pohon gaharu; 2) Patogen yang virulen, artinya organisme pathogen
yang potensial menyebabkan penyakit pada pohon inang gaharu; 3) Lingkungan yang
mendukung dan 4) Peranan manusia dalam memodifikasi lingkungan, patogen dan pohon
inang.

29
Seperti layaknya flora lain yang dapat mempertahankan diri, pohon gaharupun akan
mempertahankan diri dengan cara memperbaiki luka yang ada dan menyembuhkan
jaringan yang sakit atau rusak dengan memproduksi resin atau menangkal cendawan.
Proses ke arah tersebut merupakan upaya mempertahankan diri sehingga akan
menyebabkan terjadinya gubal gaharu yang mengandung damar wangi (Soehartono, 2001).
Gubal gaharu sebenarnya adalah resin pohon yang tidak dieksudasikan keluar
melainkan terdeposit dalam jaringan kayu. Resin yang terdeposit ini mengakibatkan kayu
yang seratnya lepas dan berwarna putih berubah menjadi kompak dan hitam serta wangi.
Resin ini dari golongan sesquiterpen yang mudah menguap (Ishihara et al., 1991) dan
merupakan senyawa fitoaleksin, yaitu senyawa yang dibentuk oleh tumbuhan sebagai
respon terhadap gangguan misalnya infeksi mikroorganisme (Kunoh, 1990).
Gambar 3. Empat faktor yang mempengaruhi terbentuknya gubal gaharu (Agrios, 1997)
Umumnya di alam pohon akan menghasilkan gubal gaharu setelah berumur lebih
dari 20 tahun, karena pada umur ini diameter batang pohon sudah cukup besar dan struktur
batangpun sudah cukup permanen dan baik untuk produksi gubal gaharu. Menurut Giano
(1986) dalam Zinch and Compton (2001), pada umur ini hanya 10 persen dari batang
tanaman atau pohon yang dapat menghasilkan gaharu, sedangkan menurut Sumarna
(2007), gaharu potensial yang ditanam sudah dapat diinokulasi mikroba untuk
pembentukan gubal pada umur 5 tahun atau pada saat telah terbentuk organ reproduktif
(berbunga dan berbuah).
MANUSIA
(pohon gaharu)
INANG
MANUSIA
WAKTU

30
Gaharu Unggul
Gaharu unggul adalah pohon gaharu yang terbukti di alam memiliki gubal gaharu
yang kemudian akan diperbanyak sebagai bibit unggul (clone) untuk kepentingan budidaya
(Sitorus, 2006). Kegiatan ini meliputi seleksi, baik seleksi in planta (pada pohon) dan
seleksi in vitro di laboratorium. Seleksi in planta pada pohon gaharu di alam telah terbukti
menghasilkan gubal gaharu yang dapat digunakan untuk keperluan breeding dan
perbanyakan bibit unggul secara in vitro, stek atau cangkok.
Hal ini penting karena berdasarkan hasil studi yang telah dilakukan oleh
PUSLITBANG Kehutanan dan SEAMEO BIOTROP di hutan alam pada beberapa daerah
penghasil gaharu di Indonesia seperti Sumatera, Kalimantan dan Sulawesi, Maluku, Nusa
Tenggara dan Papua menunjukan bahwa tidak semua pohon penghasil gaharu di hutan
alam dapat menginduksi gaharu. Pada umumnya dari pohon-pohon gaharu dewasa
berumur 25 – 50 tahun di hutan-hutan alam hanya 10 % saja yang dapat memproduksi
gubal gaharu (Isnaini, 2003).
Sementara itu, jumlah ekspor gaharu Indonesia pada tahun 1985 sekitar 1487 ton,
namun pada tahun 1995 ekspor gaharu Indonesia kurang menjadi 300 ton sehingga
CITES telah memasukan A. malaccensis ke dalam daftar appendix II (Barden, 2000)
akibatnya kuota ekspor gaharu dibatasi 250 ton/tahun. Namun demikian sejak tahun 2000
Indonesia hanya mampu memasok gaharu 15 % dari kuota CITES.
Semakin sulitnya mendapatkan gaharu di hutan alam mengakibatkan Aquilaria spp.
dan Gyrinops spp dimasukkan dalam Apendix II pada konvensi CITES tahun 2004 di
Bangkok (CITES, 2004). Kekuatiran akan punahnya spesies gaharu di Indonesia,
menyebabkan Departemen Kehutanan Indonesia sejak tahun 2005 menurunkan kuota
ekspor gaharu menjadi hanya 125 ton/tahun.
Perbanyakan Pohon Penghasil Gaharu
Bibit gaharu untuk kepentingan budidaya dapat diperoleh dari semai di alam,
perkecambahan biji, stek, cangkokan dan kultur jaringan (Sumarna, 2007). Kendala utama
yang dihadapi dalam upaya pengembangan gaharu melalui upaya budidaya di Indonesia
adalah produktifitas benih di alam rendah. Produktifitas benih yang rendah menyebabkan
kesulitan memperoleh anakan pohon gaharu dalam jumlah banyak di alam padahal untuk
tujuan budidaya yang luas sangat diperlukan bibit berkualitas dalam jumlah cukup dan
tersedia tepat waktu, oleh karena itu model perkembangbiakan vegetatif melalui stek pucuk
dan kultur jaringan merupakan alternative yang baik untuk perbanyakan gaharu unggul.

31
Kendala lain yang umumnya terdapat hampir disebagian besar daerah di Indonesia dan
Papua adalah tidak semua pohon gaharu menghasilkan buah setiap tahun, belum adanya
kebun bibit unggul dan kebun benih (Sitomorang, 2000). Rendahnya daya berbunga dan
produktifitas berbuah menyebabkan masalah regenerasi secara generatif, sementara itu
pembiakan secara vegetatif dengan menggunakan stek dan cangkok membutuhkan bahan
induk yang banyak. Kultur jaringan merupakan salah satu teknik alternatif perbanyakan
gaharu secara masal dan cepat.
Manfaat dan Nilai Ekonomi Gaharu
Gaharu telah dimanfaatkan oleh manusia menjadi komoditi perdagangan sekitar
800 tahun yang lalu karena memiliki nilai ekonomi yang sangat tinggi. Pohon gaharu
memiliki nilai ekonomi karena produk kayunya, gubal gaharu yang mengandung resin
wangi, kemendangan, serpih/ ampas gaharu dan serbuk gaharu. Minyak gaharu merupakan
produk gaharu yang paling mahal di Amerika, Eropa, Timur Tengah, India, Tibet dan
China.
Gaharu memiliki berbagai ragam manfaat, namun pada dasarnya gaharu dapat
dikelompokan ke dalam 4 manfaat besar, yaitu :
1. Manfaat dibidang industri parfum dan kosmetik, seperti : gas parfum dan minyak
gaharu, sabun, sampo dan bedak serta pengharum ruangan (Hayne, 1987; Barden et al.,
2000; Boruah & Singh, 2000).
2. Manfaat dibidang kesehatan dan obat-obatan, seperti : anti asmatik, anti mikrobia,
stimulant kerja saraf dan pencernaan, liver, hepatitis, penghilang rasa sakit, kanker,
paru-paru, rematik, cacar, malaria, sakit perut, obat kuat pada masa kehamilan dan
bersalin serta perangsang birahi (Hayne, 1987; Barden et al., 2000; Boruah & Singh,
2000).
3. Manfaat di bidang agama dan kepercayaan, seperti : upacara-upacara religius pada
umat Hindu, Budha dan Islam, pengharum ruangan sembayang, abu dan batang
kemenyan dibakar ketika melakukan ritual kepercayaan, benda-benda rohani seperti
rosario dan tasbih (Barden et al., 2000).
4. Manfaat lain : kayu gaharu kualitas kemedangan untuk perabot rumah tangga seperti
meja, kursi, rak buku, hiasan dinding, ukiran serta kulit kayu untuk pembuatan pakaian
adat, noken dan cawat pada sebagian masyarakat di Papua (Sumarna, 2007).
Harga gubal gaharu bervariasi dari Rp 5.000.000 - 15.000.000,-/kg tergantung dari
kualtitas, bentuk dan corak, warna dan aroma. Gaharu kemedangan bervariasi dari Rp

32
50.000 – 500.000,-/kg ; bubuk gaharu atau ampas gaharu yang diperoleh dari sisa
pembersihan gubal gaharu bervariasi dari Rp 10.000,- sampai Rp 50.000,-/kg.
Bervariasinya harga juga ditentukan berdasarkan rantai perdagangan dan negara pemesan
(Sumarna, 2007). Menurut Mashur (2011) harga gaharu di Indonesia berkisar Rp 100.000-
150.000.000 per kg tergantung kualitasnya. Sementara itu harga gaharu super di China
dapat mencapai Rp 400 juta per kilogram dan 300 juta per kilogram di Timur Tengah.
Total ekspor gaharu Indonesia dalam lima tahun terakhir berkisar 170-573 ton/tahun
dengan perkiraan perolehan devisa negara pada tahun 2006 sebesar 26.086.350 USD
meningkat menjadi 85. 987.500 USD pada tahun 2010 (Zulkifli, 2011).
Mikoriza
Mikoriza berasal dari kata Myces artinya cendawan dan Rhiza artinya akar
(Sieverding, 1991). Brundrett et. al (2004) menyarankan definisi baru mikoriza sebagai
suatu asosiasi simbiotik yang esensial bagi kedua partner, antara suatu cendawan
(terspesialisasi untuk hidup dalam tanah dan tumbuhan) dan akar (atau organ yang
mengadakan kontak-substrat lainnya) dari suatu tumbuhan hidup, yang terutama
bertanggungjawab untuk transfer hara. Mikoriza terjadi dalam suatu organ tumbuhan yang
terspesialisasi dimana hubungan kontak yang dekat berasal dari perkembangan cendawan-
tumbuhan yang tersinkronisasi.
Menurut Sylvia (2004) mikoriza merujuk pada suatu asosiasi atau simbiosis antara
tumbuhan dan cendawan yang mengkoloni korteks akar selama periode pertumbuhan aktif.
Simbiosis ini dicirikan oleh pergerakan hara dua arah (bi-directional movement) yaitu
karbon mengalir dari tumbuhan ke cendawan dan hara inorganik dari cendawan ke
tumbuhan dalam suatu linkage antara akar dan tanah yang mengindikasikan adanya peran
kritis bagi tanaman terutama pada kondisi lingkungan yang tidak mendukung pertumbuhan
yang optimal.
Mikoriza memberikan manfaat berupa bahan materi dan jasa. Materi yang
diberikan yaitu unsur hara, sedangkan jasa yang diberikan yaitu perlindungan terhadap
patogen, perlindungan terhadap kekeringan, perlindungan dari logam berat dan senyawa-
senyawa radio nuklir serta dalam membantu pembentukkan struktur tanah (Pfleger and
Linderman, 1996).
Langkah awal dalam pengembangan cendawan mikoriza adalah melakukan seleksi
untuk mendapatkan isolat yang infektif yaitu mampu menembus dan menyebar ke dalam
akar dan efektif yaitu mampu mempertinggi pertumbuhan atau toleransi tanaman terhadap

33
tekanan lingkungan. Hal ini disebabkan karena individu isolat FMA secara genetik sangat
bervariasi dalam sifatnya sehingga perlu screening isolat untuk keefektifan dengan
mengombinasi inang dan media tumbuh yang kompatibel dengan sistem produksi
pembibitan. Sumber isolat FMA tersebar seluruh kondisi tanah yang bervegetasi.
Pengambilan tanah-akar di bawah tegakan tanaman/pohon (daerah rhizosfer) yang diduga
terkoloni oleh FMA merupakan cara yang sering dilakukan untuk mendapatkan inokulum
FMA. Dalam suatu campuran tanah-akar ini belum dapat diketahui secara pasti baik jenis,
jumlah, potensi kualitas maupun pengaruhnya terhadap tanaman, sehingga perlu
identifikasi, pemurnian, dan uji efektifitasnya. Selanjut menurutnya untuk membedakan
antara spesifiksitas (specificity) yaitu kemampuan bawaan (innate ability) untuk
mengkoloni, keinfektifan (infectiveness) yaitu jumlah kolonisasi, dan keefektifan
(effectiveness) yaitu respon tumbuhan terhadap kolonsiasi. Dengan demikian penilaian
haruslah didasarkan atas faktor-faktor ini (Sylvia, 2004).
Menurut Brundrett et al. (2004), mikoriza harus merupakan asosiasi mutualistik
“balanced”, dimana cendawan dan tumbuhan mengubah bahan pokok yang diperlukan
untuk pertumbuhan dan daya hidupnya. Selanjutnya Brundrett (2004) mengusulkan
skema klasifikasi hirarkis simbiosis mikoriza sebagaimana pada Gambar 3.
Asosiasi mutualistik menempati kuadran saling menguntungkan (++), dan berbeda
dengan kuadran relatif menguntungkan (+) dan berbahaya (-). Pada kuadran atas (++)
BERMANFAAT BAGI
CENDAWAN
BERMANFAAT BAGI
TUMBUHAN
BERBAHAYA BAGI
TUMBUHAN
BERBAHAYA BAGI
CENDAWAN
+
+ + –
En
dop
hy
tism
e
Fakultatif obligat
Cen
daw
an
An
tag
on
is
+ –
– –
Ek
splo
itat
if
B
alan
ced
Antagonisme
tumbuhan
Parasitisme
Mikoriza
Gambar 4. Skema hirarki yang terdiri dari tipe-tipe mikoriza dan kategori interaksi
tumbuhan dengan cendawan lainnya (Brundrett et al., 2004).

34
merupakan ketergantungan tanaman yang meningkat, dimulai dari mikoriza fakultatif dan
tumbuhan non-mikoriza dengan titik kulminasinya pada tumbuhan berasosiasi mikoriza
obligat, sedangkan garis kedua searah sumbu vertikal mewakili sifat berbahaya atau
menguntungkan terhadap cendawan. Parasitik dan asosiasi antagonistik menempati dua
kuadran lainnya dengan tumbuhan-cendawan.
Fungi Mikoriza Arbuskula (FMA)
Fungi Mikoriza Arbuskula (FMA) sering disebut juga endomikoriza, ada juga yang
menggunakan istilah Vesicular-Arbuskula Mikoriza (V-AM). Istilah FMA digunakan
untuk membedakan tipe mikoriza ini dari ektomikoriza, karena disini biasanya tidak ada
sarung (sheath) miselium cendawan di sekitar akar seperti yang terdapat pada sekeliling
akar ektomikoriza. Fungi Mikoriza Arbuskula merupakan cendawan yang penyebarannya
sangat luas di dunia mulai dari daerah padang pasir, temperit, tropika dan dapat berasosiasi
lebih dari 90% tanaman yang ada di bumi. VAM telah diketahui di dalam akar tanaman
lebih dari 100 tahun yang lalu, tetapi struktur reproduktifnya baru diketahui 30 tahun
terakhir. Cendawan VA mikoriza telah berhasil diekstraksi dan ditumbuhkan pada tanaman
hidup di pot kultur (Gardeman, 1963). Diversitas FMA tidak mengikuti
keanekaragaman tanaman namun keanekaragaman spesies tanaman mungkin diatur oleh
tipe FMA. Alasan mengapa FMA dapat meningkatkan penyerapan hara dalam tanah
(Abbott & Robson, 1992), karena hifa dari FMA dapat menjangkau dan mengambil hara
dari dalam tanah dan selanjutnya mensuplai hara ke tanaman melalui akarnya,
meningkatkan penyerapan hara dan konsentrasi hara pada permukaan penyerapan,
merubah secara kimia sifat-sifat hara sehingga memudahkan penyerapannya ke dalam akar
tanaman. Dengan demikian menyebabkan terjadinya peningkatan penyerapan unsur hara,
kondisi ini banyak terjadi pada tanaman yang mempunyai akar yang kasar, tersebar tipis
dan sedikit rambut akarnya. Kapasitas pengambilan hara dapat ditingkatkan jika terjadi
kolonisasi mikoriza pada akar karena akar yang dikolonisasi diperpanjang, ukuran
percabangan serta diameter akar diperbesar dan luas permukaan absorpsi akan diperluas.
Hormon pertumbuhan seperti auksin, sitokinin dan gibberelin bagi tanaman dapat juga
meningkat. Akar sebagai penyerap unsur hara dan air akan bertahan lebih lama karena
dengan meningkatnya auksin, maka proses penuaan akar menjadi lambat (Karagiannidis et
al., 1995).
Resistensi terhadap kekeringan lebih baik pada tanaman yang bermikoriza daripada
yang tidak bermikoriza. Tanaman bermikoriza akan cepat kembali pulih setelah periode

35
kekeringan berakhir karena hifa FMA masih mampu menyerap air pada pori-pori tanah
pada saat akar tanaman sudah tidak mampu. Hifa dapat mengambil air relatif lebih banyak
karena penyebarannya di dalam tanah sangat luas.
Menurut Daniels dan Menge (1981) dan Abbott & Robson (1992), kemampuan
meningkatkan penyerapan hara dan pertumbuhan tanaman berbeda antara spesies dan
strain FMA, setiap spesies FMA mempunyai innate effectiveness atau kemampuan
spesifik. Keefektivan diartikan sebagai kemampuan FMA dalam meningkatkan
pertumbuhan tanaman pada kondisi tanah yang kurang menguntungkan. Beberapa faktor
yang berhubungan dengan keefektivan dari suatu spesies FMA, yaitu FMA mampu untuk
membentuk hifa yang ekstensif dan penyebaran hifa yang baik di dalam tanah, FMA
mampu untuk membentuk infeksi yang ekstensif pada seluruh sistem perakaran yang
berkembang dari suatu tanaman, FMA mempunyai hifa yang mampu menyerap fosfor dari
larutan tanah.
Manfaat Mikoriza
Manfaat mikoriza di alam ini sudah tidak dapat disangkal lagi karena telah banyak
ditulis para pakar. Menurut Brundrett et al. (1996), manfaat dari mikoriza dapat
dikelompokan menjadi tiga, yaitu manfaat bagi tanaman, manfaat bagi ekosistem dan
manfaat bagi manusia
a. Manfaat mikoriza bagi tanaman
Meningkatkan pasokan unsur hara tanaman dengan memperbesar volume tanah yang
dapat dikelolah oleh akar tanaman, meningkatkan pasokan hara dengan jalan menyerap
bentuk-bentuk hara yang secara normal tidak tersedia bagi tanaman, beberapa jenis
cendawan ektomikoriza dan ericoid memiliki kapasitas untuk membongkar senyawa-
senyawa fenolik yang ada dalam tanah (Smith and Read, 1997), senyawa fenolik diyakini
dapat mengganggu berlangsungnya serapan hara oleh tanaman, kolonisasi akar oleh
ektomikoriza dan FMA dapat memberikan perlindungan terhadap serangan cendawan
parasitik dan nematoda memungkinkan terjadinya perpindahan hara dari tanaman yang
sudah mati ke tanaman yang masih hidup.
b. Manfaat mikoriza bagi ekosistem
Hifa-hifa yang ada di dalam tanah berperan penting dalam pendauran hara dengan cara
membantu mencegah kehilangan hara dari sistem, khususnya ketika akar tanaman sedang
tidak aktif, hifa merupakan saluran yang digunakan untuk memindahkan karbon dari akar
tanaman ke jazad renik lainnya yang terlibat dalam proses daur hara. Dengan kata lain

36
bekerjasama dengan anggota lain dari rantai pangan dekomposisi tanah, cendawan
mikoriza dapat menyumbangkan simpanan karbon dalam tanah dengan jalan mengubah
kualitas dan kuantitas bahan organik tanah (hifa mikoriza berperan penting dalam
memperbaiki struktur tanah (Grifftiths et. al., 1992), FMA juga dapat bersinergis dengan
mikroba potensial lainnya, seperti bakteri penambat N bebas dan bakteri pelarut fosfat
(Barea et al. 1992). Serta sinergis dengan jasad – jasad renik selulotik seperti Trichoderma
sp. Berdasarkan kemampuan tersebut, maka FMA dapat berfungsi untuk meningkatkan
biodiversitas mikroba potensial di sekitar perakaran tanaman.
c. Manfaat mikoriza bagi umat manusia
Cendawan ektomikoriza secara ekonomis dan nutrisi merupakan sumber pangan
yang penting artinya bagi umat manusia. Tubuh Buah dari fungi ektomikorhiza
Scleroderma sinnamariense yang bersimbiosis dengan tanaman melinjo dapat dijadikan
sebagai bahan pangan. Jamur ektomikoriza juga telah digunakan sebagai obat-obatan dan
pewarna alami. Cendawan juga memiliki nilai keindahan/estetika dan merupakan bagian
penting dalam budaya, tradisi dan cara menghargai alam oleh umat manusia (Mansur,
2010). Keragaman cendawan merupakan bio-indikator kualitas lingkungan, cendawan
yang telah beradaptasi dengan kondisi tanah lokal diperlukan untuk pertanian, hortikultura
dan kehutanan.
Mikoriza dan Gaharu G. versteegii
Sejauh ini kajian mengenai keberadaan Fungi Mikoriza Arbuskula pada G.
versteegii asal Papua baru dilaporkan oleh Worabai (2009) yang telah berhasil
mengidentifikasikan 3 jenis spora indigenous yakni Glomus sp., Acaulospora sp., dan
Scutellospora sp. dengan persen kolonisasi akar tertinggi 16,84% dan terendah adalah
8,77%. Sementara itu, uji pemanfaatan FMA untuk memacu pertumbuhan tanaman
penghasil gaharu ini belum pernah dilakukan.
Pemanfaatan FMA pada gaharu oleh Sumarna, 2007 melaporkan bahwa aplikasi
mikorhiza ternyata menghasilkan bibit tanaman gaharu lebih cepat mencapai kondisi siap
tanam dalam kisaran waktu 2–3 bulan sejak proses perkecambahan benih. Selain itu bibit
yang diperoleh relatif lebih sehat, tahan gangguan penyakit dan hama akar dibanding
tanpa perlakuan mikorhiza. Bibit bermikorhiza memiliki kemampuan hidup di lapangan
jauh lebih tinggi dibandingkan bibit tak bermikorhiza. Hal ini didukung hasil penelitian
Karyaningsih (2009), dimana pemberian FMA jenis G. margarita mampu membentuk

37
kolonisasi, sporalisasi dan berpengaruh sangat nyata terhadap pertumbuhan semai gaharu
A. crassna.
Sampai saat ini aplikasi FMA untuk pertumbuhan plantling gaharu Papua G
versteghii belum pernah dilakukan. Untuk itu perlu dilakukan uji pemanfaatan FMA untuk
memacu dan meningkatkan pertumbuhan bibit gaharu ini.
Bio-Charcoal
Bio-charcoal adalah arang yang dibuat dari material biologis atau bahan organik
apapun yang diolah melalui proses pirolisis (pembakaran dengan suplai oksigen yang
minimal). Bio-charcoal telah digunakan untuk kegiatan pertanian dalam rangka
meningkatkan produktifitas lahan dalam budidaya pertanian yang berkelanjutan sejak suku
Indian Maya menggunakan teknik tebang, cincang dan bakar (Steiner et al., 2004). Baru-
baru ini, telah diteliti oleh banyak peneliti bahwa bio-charcoal mampu menurunkan emisi
gas rumah kaca dari tanah dan dapat meningkatkan produksi padi (IRRI), jagung
(University of Georgia di Atlanta), Singkong (Terra Pretta, Brazil), karena bio-charcoal
dapat berfungsi sebagai manajer tanah dan pembenah tanah (Steiner, 2007).
Fungsi dari penambahan bio-charcoal tersebut di dalam tanah adalah untuk
meningkatkan kesuburan tanah (pembenah tanah), meningkatkan aktifitas mikroba tanah
dan menurunkan emisi karbon ke udara. Lehman (2007) telah menunjukkan hasil
penelitiannya bahwa bio-charcoal dapat digunakan untuk meningkatkan struktur dan
kesuburan tanah yang pada gilirannya meningkatkan produksi biomassa.
Peranan bio-charcoal sebagai soil conditioner meliputi perbaikan sifat fisika, kimia
dan biologi. Pengaruh bio-charcoal terhadap sifat fisik tanah antara lain meningkatkan
stabilitas struktur dan peningkatan kandungan air tanah, peningkatan porositas dan aerase
tanah, menjadikan media lebih porous serta meningkatkan kapasitas luasan permukaan.
Peranan pada sifat kimia tanah akibat penambahan bio-charcoal ke dalam tanah yaitu
peningkatan pH tanah, Kapasitas Tukar Kation, Kejenuhan Basa dan penurunan ion Al+
dan H+. Adapun pengaruh bio-charcoal terhadap aktifitas biologi tanah antara lain sebagai
aktifator mikoriza dan rhizobium. Pemberian arang sekam terhadap bakteri penambat
nitrogen dan cendawan mikoriza mampu meningkatkan jumlah bintil akar pada tanaman
leugum, meningkatkan jumlah spora dan tingkat kolonisasi Glomus etunicatum (Ogawa,
1994; Glaser at al., 2002 ; Braida et al., 2003 diacu Ahmad, 2006).
Di Indonesia teknologi bio-charcoal juga telah digunakan pada system perladangan
berpindah, namun teknik tersebut hanya menghasilkan 2% arang karena dibakar dengan

38
suplai oksigen yang berlebihan (tempat terbuka). Pemanfaatan bio-charcoal dibidang
kehutanan telah dilakukan oleh Badan Litbang Kehutanan Indonesia pada tanamaan Acacia
mangium, Shorea leprosulla dan Pinus merkusii berumur 2,5-3 tahun menunjukan
pengaruh sangat nyata terhadap indikator pertumbuhan (tinggi, diameter, biomasa dan akar
tanaman) (Siregar, 2010). Sementara itu hasil uji coba pemanfaatan bio-charcoal untuk
produksi FMA menggunakan inang Shorgum bicolour menunjukan pengaruh yang
signifikan terhadap jumlah spora (Supriyanto, 2010).
Lignit (Batubara Muda)
Lignit disebut juga batubara muda atau batubara cokelat, tidak ekonomis digunakan
sebagai bahan bakar. Lignit memiliki kelembaban yang tinggi dan kandungan karbon yang
rendah. Dengan adanya masalah tersebut, apabila terdapat lapisan batubara lignit dalam
penambangan batubara, maka penambang hanya mengambil lapisan yang berkualitas
tinggi, sedangkan lignit akan disingkirkan atau ditimbun kembali di lokasi tambang.
Batubara dengan mutu yang rendah, seperti lignit dan sub-bitumen biasanya lebih lembut
dengan materi yang rapuh, berwarna suram seperti tanah dan memiliki kelembaban tinggi
dengan demikian energinya rendah.
Lignit (batu bara muda) mengandung zat-zat humat (asam humat) sebagai unsur
organik utama yang banyak terdapat dalam air alami, bahan-bahan yang tahan degradasi
yang dihasilkan selama dekomposisi dari tumbuhan yang terjadi sebagai endapan dalam
tanah, sedimen rawa. Asam humat adalah salah satu senyawa yang terkandung dalam
humat substance yang merupakan hasil dekomposisi bahan organik, utamanya bahan
nabati yang terdapat dalam batu bara muda, tanah gambut, kompos atau humus (Senn dan
Kigman, 1973).
Asam humat merupakan hasil akhir proses dekomposisi bahan organik, berwarna
hitam, kecoklatan, relatif tahan terhadap degradasi serta mengandung muatan negatif yang
dapat dipengaruhi pH (Stevenson, 1994). Proses pembentukan batubara muda dapat
melalui dekomposisi flora dan fauna yang merupakan bagian dari transformasi biokimia
organik sebagai titik awal. Dalam pertumbuhan gambut, sisa tumbuhan akan mengalami
perubahan baik secara fisik maupun kimiawi. Setelah tumbuhan mati, proses degradasi
biokimia lebih berperan. Proses pembusukan akan terjadi oleh kerja mikrobiologi (bakteri
anaerob). Bakteri ini bekerja dalam suasana tanpa oksigen menghancurkan bagian yang
lunak dari tumbuhan seperti selulosa, protoplasma dan pati. Melalui proses itu terjadi
perubahan dari kayu menjadi lignit dan batubara berbitumen. Dalam suasana kekurangan

39
oksigen terjadi proses biokimia yang berakibat keluarnya air (H2O) dan sebagian unsur
karbon akan hilang dalam bentuk karbon dioksida (CO2), karbon monoksida (CO) dan
methan (CH4).
Indonesia termasuk negara dengan sumber tambang batubara terbesar di dunia.
Cadangannya diperkirakan 36 ton. Hanya saja 50 – 85 persennya berkualitas rendah. Ini
dilihat dari nilai kalori pembakarannya yang rendah, dan kadar sulfur serta airnya yang
tergolong tinggi ( Sukandarrumudi, 2006).
Asam humat merupakan komponen organik, yang dapat berasosiasi menjadi ion
yang aktif serta bersiaft kolodial dan relatif stabil. Berperan besar dalam memperbaiki
kesuburan tanah, baik secara kimia, fisika dan biologi tanah. Memperbaiki strukjtur tanah,
meningkatkan kapasitas memegang air dan Kapasits Tukar Kation (KTK) tanah,
menurunkan kalarutan unsur beracun seperti Fe dan Al karena muatan negatif dan gugus
fungsional karboksil (- COOH) dan hidroksil (-OH).
Pemanfaatan batubara muda sebagai pembenah tanah telah dilakukan oleh
Karyanigsih, 2009 dan hasil penelitiannya menunjukkan bahwa pemanfaatan batubara
muda dan Glomus margarita sebagai pembenah tanah dalam media pertumbuhan
berpengaruh sangat nyata terhadap peningkatan pertumbuhan semai gaharu A. crassna,
kolonisasi dan jumlah spora G. margarita.

40
METODE PENELITIAN
Tempat dan Waktu
Survey pohon penghasil gaharu G. versteegii untuk kajian keragaman FMA
indigenous, dilaksanakan dalam bulan Juli 2010 pada hutan alam di daerah Asai
Kabupaten Manokwari Papua Barat. Survey dilakukan bersama-sama dengan
masyarakat pencari gaharu dengan cara jelajah lokasi penelitian sejauh 1 km. Setiap
pohon induk gaharu yang ditemukan dalam jarak pandang 10 meter kiri dan kanan jalan
dijadikan sebagai plot pengambilan contoh semai dan tanah untuk isolasi dan
identifikasi keberadaan mikorizanya. Dalam survey ini juga dicatat vegetasi lain yang
tubuh pada radius 2 meter dari pohon induk gaharu.
Kegiatan isolasi FMA, identifikasi, multiplikasi FMA dilakukan pada
Laboratorium Silvikultur Fakultas Kehutanan IPB. Aplikasi FMA, bio-charcoal,
kompos dan lignit pada plantling gaharu dilaksanakan di Laboratorium Mikoriza dan
greenhouse SEAMEO BIOTROP. Analisis tanah dilakukan di Laboratorium Penelitian
dan Uji Tanah, Balai Besar Litbang Sumberdaya Lahan Pertanian, Badan Penelitian dan
Pengembangan Pertanian Bogor. Penelitian ini berlangsung dari awal Nopember 2010
sampai dengan bulan Juni 2011.
Bahan dan Alat
Bahan-bahan yang digunakan adalah inokulum FMA komposit hasil trapping
asal Manokwari, Papua Barat. Plantlet G. versteegii asal Papua yang telah
dikembangkan di SEAMEO BIOTROP melalui teknik kultur jaringan, Pueraria
(Pueraria javanica), kompos, lignit (batubara muda) asal Desa Sukajaya Kabupaten
Sukabumi, zeolit, tanah, pasir, KOH 1%, KOH 10%, HCl 2%, H2O2 2%, Glyserin 50 %,
Acid Fushin 0,02 %, larutan PVLG, larutan Melzer’s reagen, aquades, pot plastik hitam,
polybag 10 x 15 cm, kertas nitroselulosa, sungkup plastik, paranet dan label.
Alat-alat yang digunakan meliputi: camera digital, pH-fertility meter,
mikroskop binokuler/dissecting microscope, sentrifus, termohydrometer, saringan
berukuran 425 μm, 125 μm, 60 μm dan 45 μm, pinset spora, cawan petri, gelas becker,
kaca obyek dan cover glass, pipet, jarum ose, timbangan analitik, oven, autoclave,
magnetic stirrer, pacul, ayakan tanah 1 mm, dan alat tulis.

41
Metode
Penelitian ini dirancang menggunakan Rancangan Acak Lengkap (RAL) pola
faktorial 2 faktor (4x3). Perlakuan dan taraf perlakuan yang diberikan sebagai berikut.
I. Faktor Perlakuan 1 ; Media (M)
Mo Tanah : Pasir (control) (v/v) 1 : 1
M1 Tanah : Pasir : Kompos (v/v) : Bio-Charcoal (v/v) 1 : 1 : 0,5 : 0,2
M2 Tanah : Pasir : Lignit (v/v) : Bio-Charcoal (v/v) 1 : 1 : 0,4 : 0,2
M3 Tanah : Pasir : Kompos:Lignit (v/v) : Bio-Charcoal 1 : 1 : 0,5 : 0,4 : 0,2
II. Faktor Perlakuan 2 ; FMA (F)
F0 Tanpa Inokulum FMA 0
F1 Inokulum FMA indigenous (konsorsium) 10 g
F2 Inokulum FMA spora tunggal(Gigaspora sp) 10 g
Dari taraf perlakuan yang diberikan, diperoleh 12 kombinasi perlakuan. Masing-
masing perlakuan diulang 3 kali sehingga diperlukan 36 bibit hasil kultur jaringan.
Kombinasi media merupakan hasil uji pendahuluan untuk mengukur parameter tingkat
kesuburan dan tingkat keasaman kombinasi lignit, bio-charcoal, kompos dan tanah
menggunakan alat ukur pH-fertility meter. Media disterilkan menggunakan autoclave
dengan tekanan 15 atm pada suhu 120 0C selama 2 jam. Lignit yang digunakan, terlebih
dahulu digerus lalu diayak menjadi tepung menggunakan ayakan berukuran 10 mm.
Sebelum ditanam, media disiram dengan air hingga kapasitas lapang lalu diperam selama
satu minggu. Khusus untuk kombinasi media yang menggunakan lignit, media disiram
dengan KOH 1% sebanyak 10 ml untuk mempercepat pelarutan asam humat dari lignit
Prosedur Pelaksanaan
Isolasi dan Identifikasi FMA
a. Pengambilan Sampel Tanah dari Semai G. versteegii
Pengambilan sampel tanah komposit dari bawah tegakan G. verstegii sebanyak 5
(lima) plot. Dari setiap plot diambil satu sampel rizosfer tanah untuk ekstraksi FMA dan
satu sampel tanah untuk analisis sifat tanah, masing-masing sebanyak 1 kg dengan
kedalaman 0 – 20 cm. Semai G. veerstegii yang diambil berukuran tinggi 75 – 100 cm.
b. Ekstraksi Spora FMA dengan Teknik penyaringan basah (Metode Gardeman and
Nicholson, 1963 ) dan Metoda Sentrifugasi (Brundrett et al., 1996)

42
Tanah yang telah diambil dari lapangan ditimbang sebanyak 10 gr, lalu dimasukkan ke
dalam gelas piala 500 atau 1000 ml atau bekas botol air mineral 1 L dan ditambahkan
500 ml air, agregat tanah dipecahkan dan tanah tersebut diaduk dengan tangan agar
spora terbebas dari tanah.
Larutan dibiarkan selama kurang lebih 15-30 detik tergantung tekstur tanahnya sampai
partikel-partikel tanah mengendap, selanjutnya supernatant dituang ke dalam penyaring
bersusun berturut-turut 425 μm, 125 μm, 69 μm, 45 μm. Tanah pada penyaring
disemprot dengan air mengalir. Penyemprotan dilakukan dengan hati-hati agar tidak
merusak struktur spora.
Akar-akar dipisahkan dari tanah jika ada, untuk kemudian diamati kemungkinan
adanya spora pada akar atau kolonisasi infeksi perakaran melalui pewarnaan (staining)
akar.
Supernatant keruh hasil penyaringan dari penyaring dengan ukuran 45 dan 69 µm
masing-masing dimasukkan ke dalam tabung sentrifusi. Kemudian larutan sukrosa 60
% ditambahkan ke dalam tabung sentrifusi dengan perbandingan supernatant keruh dan
sukrosa adalah 3 : 1 (v/v), kemudian disentrifusikan pada kecepatan putar 1200-1300
rpm selama 5 menit.
Hasil sentrifusi dituangkan ke penyaring yang sesuai, lalu dibilas dengan air perlahan-
lahan agar tidak terjadi plasmolisis spora. Selanjutnya supernatant dituang pada kertas
saring, lalu dikeringanginkan dan diamati di bawah mikroskop. Jumlah spora dihitung
dan diidentifikasi jenis FMA-nya.
c. Pewarnaan akar (staining) (Metode Phillips dan Hayman, 1970 hasil modifikasi
Supriyanto, 2009).
Sampel akar diambil dan dipotong akar kecil diameter ≤ 2 mm yaitu akar serabut/akar
rambut sebanyak kurang lebih 2 gram
Akar dicuci hingga bersih untuk menghilangkan kotoran berupa tanah, serasah,
kompos, pasir dan lain-lain.
Fiksasi akar dilakukan dengan cara merendam potongan akar dalam larutan FAA
Bilas larutan fiksator (FAA) dalam air mengalir hingga larutan fiksator hilang.
Hilangkan komponen tanin dan fenol dalam akar dengan merendam dalam larutan
KOH 10% (v/v) dengan cara dipanaskan selama 5-10 dihitung sejak mendidih.
Pindahkan akar-akar tersebut dalam saringan plastik dan bilas dengan air mengalir
untuk menghilangkan larutan KOH 10 %, tanin dan fenol.

43
Rendam akar-akar yang sudah dihilangkan tanin dan fenolnya dalam hydrogen
peroksida (H2O2 2% w/v) selama 5-10 menit.
Bilas akar-akar tersebut dengan air mengalir untuk menghilangkan larutan hydrogen
peroksida dengan menggunakan saringan plastik.
Rendam akar-akar tersebut dalam larutan HCl 2% (v/v) selama 10-30 menit atau
sampai akar berwarna putih. Larutan HCl digunakan untuk penjernih (clearing)
komponen sitoplasma.
Staining akar-akar tersebut dalam larutan asam fushin 0,2 % dan larutan lacto-glycerol,
lalu dipanaskan selama 5-10 menit dihitung sejak mendidih.
Akar-akar tersebut dibilas dengan air mengalir hingga larutan asam fushin hilang.
Kemudian akar direndam dalam larutan destaining glyserin 50 % untuk menghilangkan
kelebihan larutan staining asam fushin 0,2 %
Pengamatan dilakukan di bawah mikroskop binukuler untuk mengetahui keberadaan
hifa, vesikula, spora intra radikal dan arbuskula
Untuk pembuatan preparat awetan, maka akar-akar tersebut dipotong dengan ukuran 1
cm dan letakan di atas kaca preparat yang sudah ditetesi larutan PVLG kemudian
ditutup dengan cover glass. Spesimen dibiarkan hingga kering lalu diamati dan
dilakukan pemotretan dengan dissecting microscope/stereo
Kolonisasi perakaran dilihat berdasarkan kolonisasi oleh struktur FMA (hifa eksternal,
hifa internal, spora intradikal, vesikula dan arbuskula).
d. Identifikasi Fungi Mikoriza Arbuskula (FMA)
Identifikasi dilakukan berdasarkan karakter morfologi spora hasil ekstraksi. Peubah
yang diamati dalam mendeskripsikan spora meliputi bentuk spora, ukuran spora, warna
spora, ornamen/asesoris permukaan spora, substanding hifa dan Bulbous suspensor;
Pembuatan preparat spora untuk identifikasi menggunakan glass slide, cover glass,
polyvinyl alcohol-lactic acid-glycerol (PVLG) dan larutan pewarna Melzer’s. Pengamatan
dilakukan menggunakan mikroskop yang dihubungkan dengan kamera CCTV dan USB
converter untuk dapat diamati di komputer, dan dapat dilakukan pengambilan gambar
(capturing).
Produksi Inokulum FMA
Teknik penangkaran/trapping yang digunakan mengikuti metoda Brundrett et al.,
(1996), menggunakan pot plastik dengan media tanah contoh dari bawah tegakan gaharu
sebanyak 100 gram dan zeolit steril. Pueraria javanica digunakan sebagai tanaman inang.

44
Masing-masing sampel tanah diulang 2 kali. Propagul diamati setelah penangkaran
berumur 3 bulan lalu penyiraman dihentikan selama 2 minggu (stressing agent) kemudian
propagul FMA diisolasi, diidentifikasi dan pendugaan potensi inokulum untuk aplikasi
perlakuan.
Aplikasi FMA, Kompos, Bio-Charcoal dan Lignit pada Plantling Gaharu
Penanaman
1) Media tanah, FMA dengan bio-charcoal, kompos dan batu bara muda dicampur sesuai
taraf perlakuan dan dimasukan ke dalam polybag berukuran 10 x 15 cm.
2) Polybag berisi media tanam dibuat lobang tanam sedalam ± 5 cm, kemudian
diinokulasi dengan spora FMA konsorsium (bentuk granular) hasil trapping dan FMA
spora tunggal hasil trapping ke dalam polybag sesuai perlakuan yang dicobakan.
3) Bibit ditanam tepat di atas granular, lalu ditutup kembali dengan media.
4) Pada setiap polybag ditutup dengan sungkup plastik transparan untuk mencegah
terjadinya transpirasi berlebihan dan mempertahankan kelembaban udara di sekitar
bibit.
5) Selanjutnya ditempatkan di dalam rumah kaca secara random menurut kaidah RAL.
Pemeliharaan
Pemeliharaan meliputi penyiraman secara teratur sesuai kebutuhan dan kondisi
tanaman, pembersihan dari gulma serta pemeliharaan terhadap serangan hama dan
penyakit.
Pengamatan dan Pengukuran
Parameter utama yang dipakai dalam menjawab tujuan penelitian :
1. Parameter Keragaman Jenis (bentuk spora, ukuran spora, warna spora,
ornamen/asesoris permukaan spora, substanding hifa dan bulbous suspensor).
2. Parameter Kompatibilitas (Persentase kolonisasi akar, Percentage growth respon
(PGR), Dependency of P uptake (DPU).
3. Parameter Pertumbuhan (Persentase plantling hidup, pertambahan tinggi dan diameter,
berat kering tanaman dan Indeks Mutu Bibit).
Peubah yang diamati dan diukur dalam penelitian ini meliputi :
1) Kolonisasi akar. Menggunakan Metode Slide berdasarkan bidang pandang terinfeksi
(+) dan tidak terinfeksi (-) terhadap 10 potong akar dalam 3 kali ulangan dengan
rumus:

45
Jumlah potongan akar yang terinfeksi
% Kolonisasi = x 100 %
Jumlah seluruh potongan akar yang diamati
2) Respon tanaman terhadap mikoriza ditentukan berdasarkan percentage growth respon
(PGR) menurut Hetrik dan Wilson (1993), sebagai berikut :
BK tanaman terinokulasi – BK tanaman tidak terinokulasi
PGR = x 100 %
BK tanaman tidak terinokulasi
3) Ketergantungan tanaman bermikoriza terhadap fosfor atau dependency of P uptake
(DPU) ditentukan dengan rumus Tawaraya, Tokarin dan Wagatsuma (2001) yang
diacu Asmarahman (2008) sebagai berikut :
Kandungan P Tan bermikoriza – Kandungan P Tan Tidak bermikoriza
DPU = x 100 %
Kandungan P Tanaman bermikorhiza
4) Plantling hidup pada umur 8 MST; dihitung dengan rumus :
Jumlah plantling hidup
% bibit hidup = x 100 %
Jumlah plantling yang ditanam
5) Pertambahan Tinggi Bibit (cm). Pengukuran tinggi bibit dilakukan dengan
menggunakan mistar mulai dari pangkal batang (diberi tanda batas) hingga titik
tumbuh tunas pucuk semai setelah tinggi awal ditentukan. Pengukuran dilakukan satu
minggu sekali selama 3 bulan.
6) Pertambahan Diameter Bibit (mm). Pengukuran diameter bibit dilakukan
menggunakan mini kaliper, di atas pangkal batang. Pengukuran dilakukan satu
minggu sekali selama 3 bulan.
7) Biomasa Kering Tanaman. Dilakukan setelah pengamatan tinggi dan diameter selesai.
Sampel tanaman dipotong, bagian pucuk dan akarnya dibungkus kertas secara
terpisah, kemudian dioven pada suhu 80 0C selama 2 x 24 jam. Penimbangan
dilakukan setelah mencapai berat kering konstan.
8) Indeks Mutu Bibit (IMB) diukur pada umur 3 bulan. Nilai IMB ditentukan melalui
parameter pertumbuhan yang sesuai menggunakan sistem skoring.
9) Analisa serapan hara tanaman meliputi serapan nitrogen, fosfor dan kalium yang
dilakukan di Laboratorium Penelitian dan Uji Tanah, Balai Besar Litbang
Sumberdaya Lahan Pertanian, Badan Penelitian dan Pengembangan Pertanian Bogor.
Setelah diperoleh N dan P pada setiap tanaman maka serapan hara tanaman dihitung

46
dengan mengalikan berat kering tanaman terhadap kadar haranya (Harjono dan
Warsito 1992).
P serapan = P kandungan x BK
N serapan = N kandungan x BK
11). Analisis histologi stomata plantlet. Analisis dilakukan untuk mengetahui keberadaan
stomata plantlet dan plantling.
12) Geometri akar, dihitung berdasarkan jumlah dan panjang baik akar primer maupun
akar sekunder.
13) Parameter pendukung: analisa awal keadaan tanah alam dari plot-plot pengambilan
sampel FMA, keadaan tanah pada media tanam meliputi kandungan N, P, K, pH, KB
dan KTK; suhu (oC) dan kelembaban (% RH) harian ditempat percobaan.
Analisis Data
Data hasil pengamatan pada setiap peubah dianalisis dengan uji F, melalui program
SPSS. Perbedaan antar perlakuan diuji dengan menggunakan Duncan’s Multiple Range
Test (DMRT) pada slang kepercayaan 95 %. Untuk mengetahui hubungan persentase
kolonisasi FMA terhadap parameter pertumbuhan dilakukan uji korelasi Pearson pada
selang kepercayaan 99%.
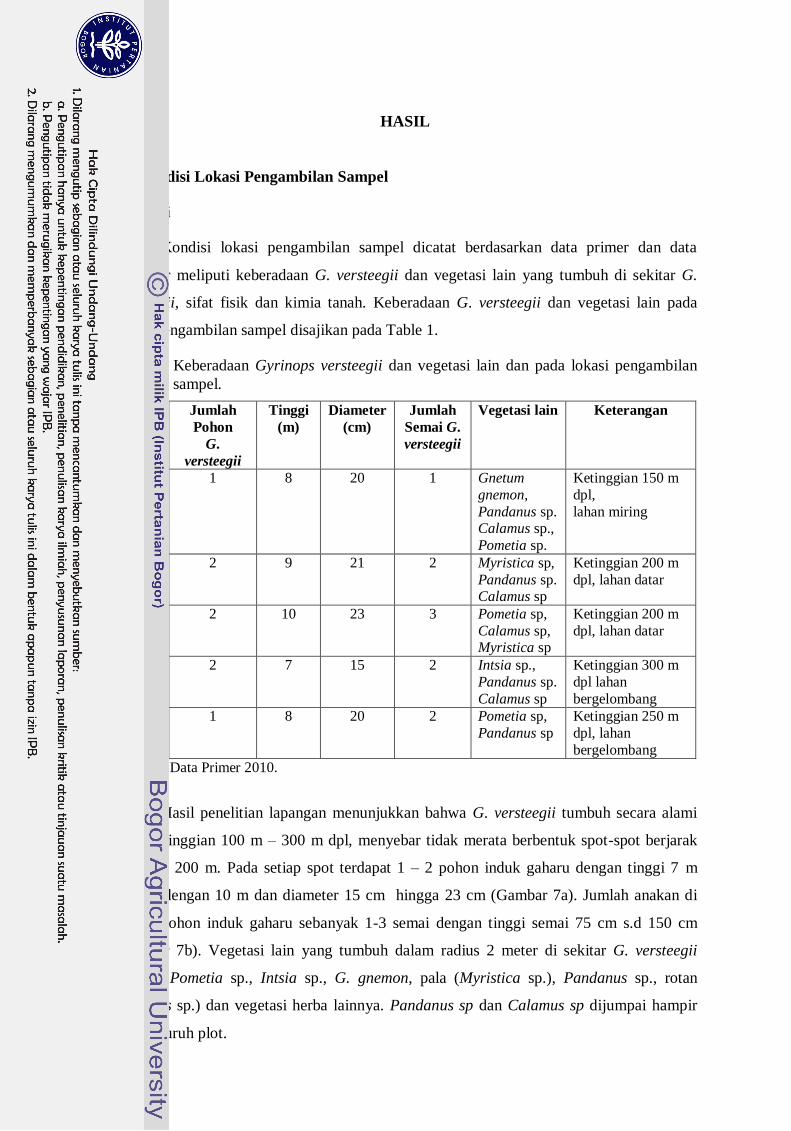
47
HASIL
1. Kondisi Lokasi Pengambilan Sampel
Vegetasi
Kondisi lokasi pengambilan sampel dicatat berdasarkan data primer dan data
sekunder meliputi keberadaan G. versteegii dan vegetasi lain yang tumbuh di sekitar G.
versteegii, sifat fisik dan kimia tanah. Keberadaan G. versteegii dan vegetasi lain pada
lokasi pengambilan sampel disajikan pada Table 1.
Tabel 1. Keberadaan Gyrinops versteegii dan vegetasi lain dan pada lokasi pengambilan
sampel.
Nomer
Plot
Jumlah
Pohon
G.
versteegii
Tinggi
(m)
Diameter
(cm)
Jumlah
Semai G.
versteegii
Vegetasi lain Keterangan
1 1 8 20 1 Gnetum
gnemon,
Pandanus sp. Calamus sp.,
Pometia sp.
Ketinggian 150 m
dpl,
lahan miring
2 2 9 21 2 Myristica sp,
Pandanus sp. Calamus sp
Ketinggian 200 m
dpl, lahan datar
3 2 10 23 3 Pometia sp,
Calamus sp, Myristica sp
Ketinggian 200 m
dpl, lahan datar
4 2 7 15 2 Intsia sp.,
Pandanus sp.
Calamus sp
Ketinggian 300 m
dpl lahan
bergelombang
5 1 8 20 2 Pometia sp,
Pandanus sp
Ketinggian 250 m
dpl, lahan
bergelombang
Sumber : Data Primer 2010.
Hasil penelitian lapangan menunjukkan bahwa G. versteegii tumbuh secara alami
pada ketinggian 100 m – 300 m dpl, menyebar tidak merata berbentuk spot-spot berjarak
150 m – 200 m. Pada setiap spot terdapat 1 – 2 pohon induk gaharu dengan tinggi 7 m
sampai dengan 10 m dan diameter 15 cm hingga 23 cm (Gambar 7a). Jumlah anakan di
bawah pohon induk gaharu sebanyak 1-3 semai dengan tinggi semai 75 cm s.d 150 cm
(Gambar 7b). Vegetasi lain yang tumbuh dalam radius 2 meter di sekitar G. versteegii
adalah Pometia sp., Intsia sp., G. gnemon, pala (Myristica sp.), Pandanus sp., rotan
(calamus sp.) dan vegetasi herba lainnya. Pandanus sp dan Calamus sp dijumpai hampir
pada seluruh plot.

48
Jika dilihat dari jumlah anakan yang tumbuh maka jumlahnya tidak banyak
mengingat diameter pohonnya sudah cukup besar. Hal ini karena benih G. versteegii
termasuk jenis rekalsitran atau cepat kehilangan daya kecambah. Disisi lain kadang-
kadang ditemukan juga semai yang tumbuh menggerombol karena benih yang jatuh
mengelompok dalam satu tempat yang berdekatan. Gyrinops versteegii juga mampu
menghasilkan trubusan yang tumbuh seperti semai normal. Hal ini penting jika ditebang
diharapkan akan menghasilkan individu baru yang berasal dari trubusan (Gambar 7.a).
Ketika dilakukan survey pada bulan Juli 2010 tidak ditemukan bunga dan benih G.
versteegii atau kulit buahnya di lantai hutan, dengan demikian musim berbunga dan
berbuah G. versteegii terjadi pada bulan-bulan yang lain.
Kondisi lantai hutan lembab dan ditutup oleh serasah setebal 5-10 cm. Dalam
kondisi seperti ini buah G. versteegii akan cepat membusuk sehingga jumlah semai yang
dijumpai juga sangat sedikit.
Pada Gambar 5a nampak ada bekas luka pengecekan terjadinya gaharu oleh pencari
gaharu. Luka tersebut dibiarkan dengan harapan akan terbentuk gaharu dikemudian hari.
Hal ini berarti pencari gaharu telah mengenal proses pembentukan gaharu dengan cara
melukai pohon G. versteegii yang tumbuh di hutan.
a b
Gambar 5. Pohon gaharu Gyrionops versteegii dan vegetasi lain yang tumbuh
disekitar Gyrionops versteegii (a) dan keberadaan semai Gyrionops
versteegii
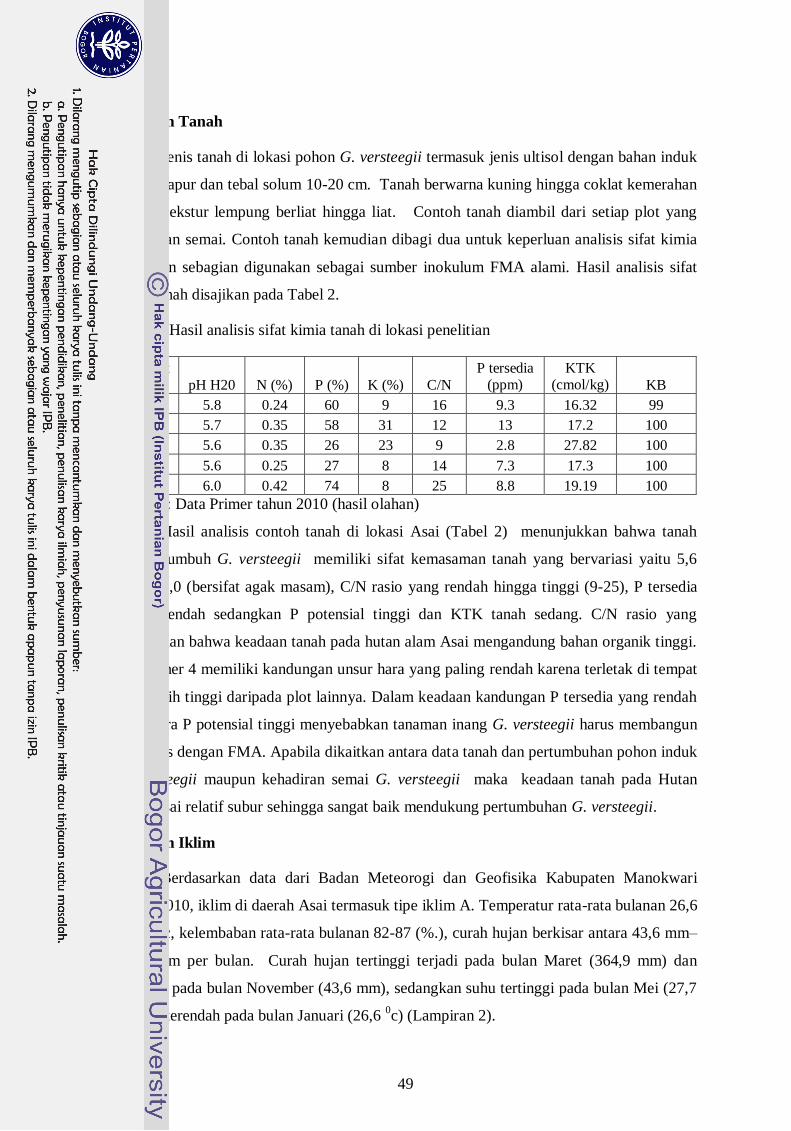
49
Keadaan Tanah
Jenis tanah di lokasi pohon G. versteegii termasuk jenis ultisol dengan bahan induk
batuan kapur dan tebal solum 10-20 cm. Tanah berwarna kuning hingga coklat kemerahan
dengan tekstur lempung berliat hingga liat. Contoh tanah diambil dari setiap plot yang
ditemukan semai. Contoh tanah kemudian dibagi dua untuk keperluan analisis sifat kimia
tanah dan sebagian digunakan sebagai sumber inokulum FMA alami. Hasil analisis sifat
kimia tanah disajikan pada Tabel 2.
Tabel 2. Hasil analisis sifat kimia tanah di lokasi penelitian
No. Plot Contoh pH H20 N (%) P (%) K (%) C/N
P tersedia (ppm)
KTK (cmol/kg) KB
Plot 1 5.8 0.24 60 9 16 9.3 16.32 99
Plot2 5.7 0.35 58 31 12 13 17.2 100
Plot3 5.6 0.35 26 23 9 2.8 27.82 100
Plot4 5.6 0.25 27 8 14 7.3 17.3 100
Plot5 6.0 0.42 74 8 25 8.8 19.19 100
Sumber : Data Primer tahun 2010 (hasil olahan)
Hasil analisis contoh tanah di lokasi Asai (Tabel 2) menunjukkan bahwa tanah
tempat tumbuh G. versteegii memiliki sifat kemasaman tanah yang bervariasi yaitu 5,6
hingga 6,0 (bersifat agak masam), C/N rasio yang rendah hingga tinggi (9-25), P tersedia
sangat rendah sedangkan P potensial tinggi dan KTK tanah sedang. C/N rasio yang
mencirikan bahwa keadaan tanah pada hutan alam Asai mengandung bahan organik tinggi.
Plot nomer 4 memiliki kandungan unsur hara yang paling rendah karena terletak di tempat
yang lebih tinggi daripada plot lainnya. Dalam keadaan kandungan P tersedia yang rendah
sementara P potensial tinggi menyebabkan tanaman inang G. versteegii harus membangun
simbiosis dengan FMA. Apabila dikaitkan antara data tanah dan pertumbuhan pohon induk
G. versteegii maupun kehadiran semai G. versteegii maka keadaan tanah pada Hutan
Alam Asai relatif subur sehingga sangat baik mendukung pertumbuhan G. versteegii.
Keadaan Iklim
Berdasarkan data dari Badan Meteorogi dan Geofisika Kabupaten Manokwari
Tahun 2010, iklim di daerah Asai termasuk tipe iklim A. Temperatur rata-rata bulanan 26,6
– 27,7 0c, kelembaban rata-rata bulanan 82-87 (%.), curah hujan berkisar antara 43,6 mm–
364,9 mm per bulan. Curah hujan tertinggi terjadi pada bulan Maret (364,9 mm) dan
terendah pada bulan November (43,6 mm), sedangkan suhu tertinggi pada bulan Mei (27,7
0C) dan terendah pada bulan Januari (26,6
0c) (Lampiran 2).

50
Berdasarkan data iklim tersebut maka lokasi pengambilan contoh termasuk
memiliki curah hujan yang tinggi dan sering tergenang oleh air hujan. Hal ini juga terkait
erat dengan kemampuan regenerasi G. versteegii. Dalam kondisi seperti ini maka benih
G. versteeigii akan mudah busuk sehingga tingkat regenerasi alaminya rendah.
2. Potensi Fungi Mikoriza Arbuskular Alami di semai G. versteegii di Hutan Alam
Asai
Keberadaan Jenis FMA
Untuk mengetahui potensi FMA alami, dilakukan trapping untuk memicu
sporulisasi FMA di green house menggunakan inang Pueraria javanica dan media zeolit
selama 3 bulan. Informasi potensi FMA alami diperlukan untuk pembibitan G. versteegii di
greenhouse atau untuk penanaman di lapangan. Jika potensinya rendah maka perlu
dilakukan inokulasi.
Hasil isolasi dari trapping menunjukkan bahwa jumlah spora FMA alami
bervariasi dari 2 hingga 5 spora /10 gram sampel tanah. Identifikasi berdasarkan karakter
morfologi FMA antara lain ukuran diameter spora, warna spora, ornamen/asesoris
permukaan spora, substanding hifa dan bulbous suspensor sehingga di lokasi penelitian
ditemukan 7 spesies yang berasosiasi dengan semai G. versteegii, yaitu Glomus mossae,
Glomus fasciculatum, Glomus aggregatum, Glomus sp1, Glomus sp2, Glomus sp3 dan
Acaulospora sp1 (Tabel 3).
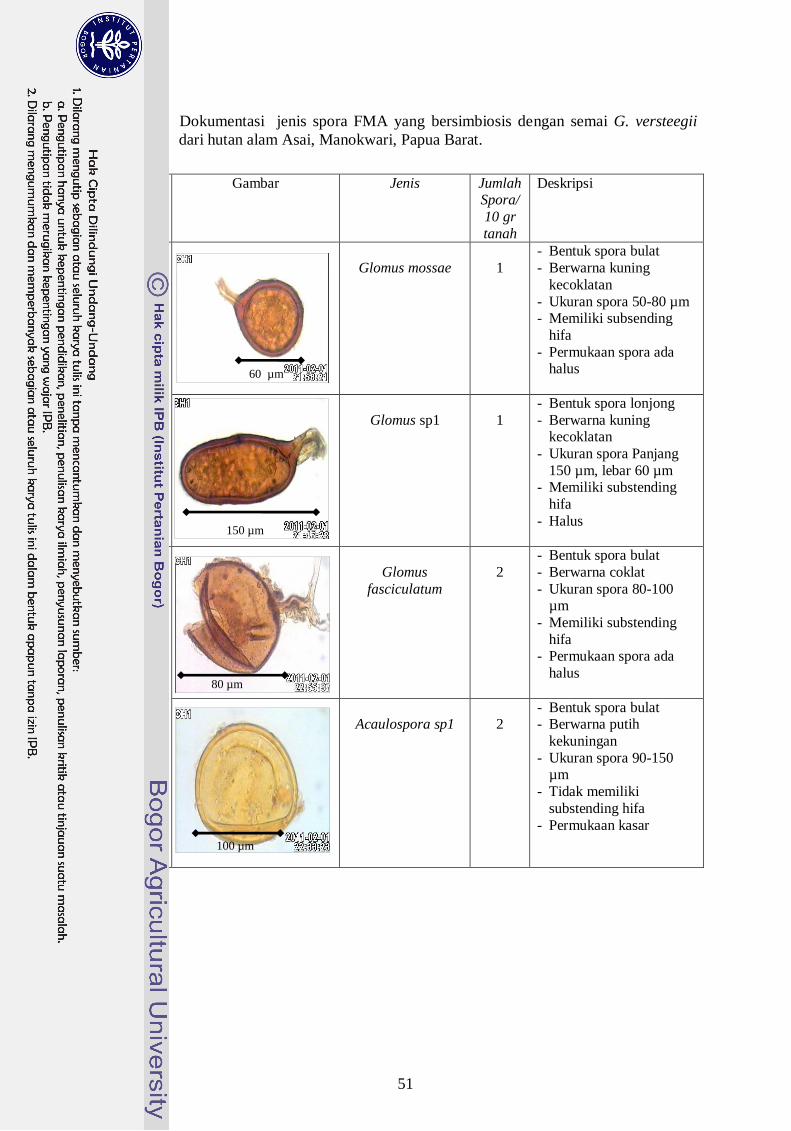
51
Tabel 3. Dokumentasi jenis spora FMA yang bersimbiosis dengan semai G. versteegii
dari hutan alam Asai, Manokwari, Papua Barat.
Plot Contoh
Gambar Jenis Jumlah Spora/
10 gr
tanah
Deskripsi
Plot 1
Glomus mossae
1
- Bentuk spora bulat
- Berwarna kuning
kecoklatan
- Ukuran spora 50-80 µm - Memiliki subsending
hifa
- Permukaan spora ada halus
Glomus sp1
1
- Bentuk spora lonjong
- Berwarna kuning kecoklatan
- Ukuran spora Panjang
150 µm, lebar 60 µm - Memiliki substending
hifa
- Halus
Plot 2
Glomus
fasciculatum
2
- Bentuk spora bulat
- Berwarna coklat
- Ukuran spora 80-100 µm
- Memiliki substending
hifa - Permukaan spora ada
halus
Acaulospora sp1
2
- Bentuk spora bulat - Berwarna putih
kekuningan
- Ukuran spora 90-150 µm
- Tidak memiliki
substending hifa
- Permukaan kasar
60 µm µm µm
150 µm
80 µm mmµm
100 µm
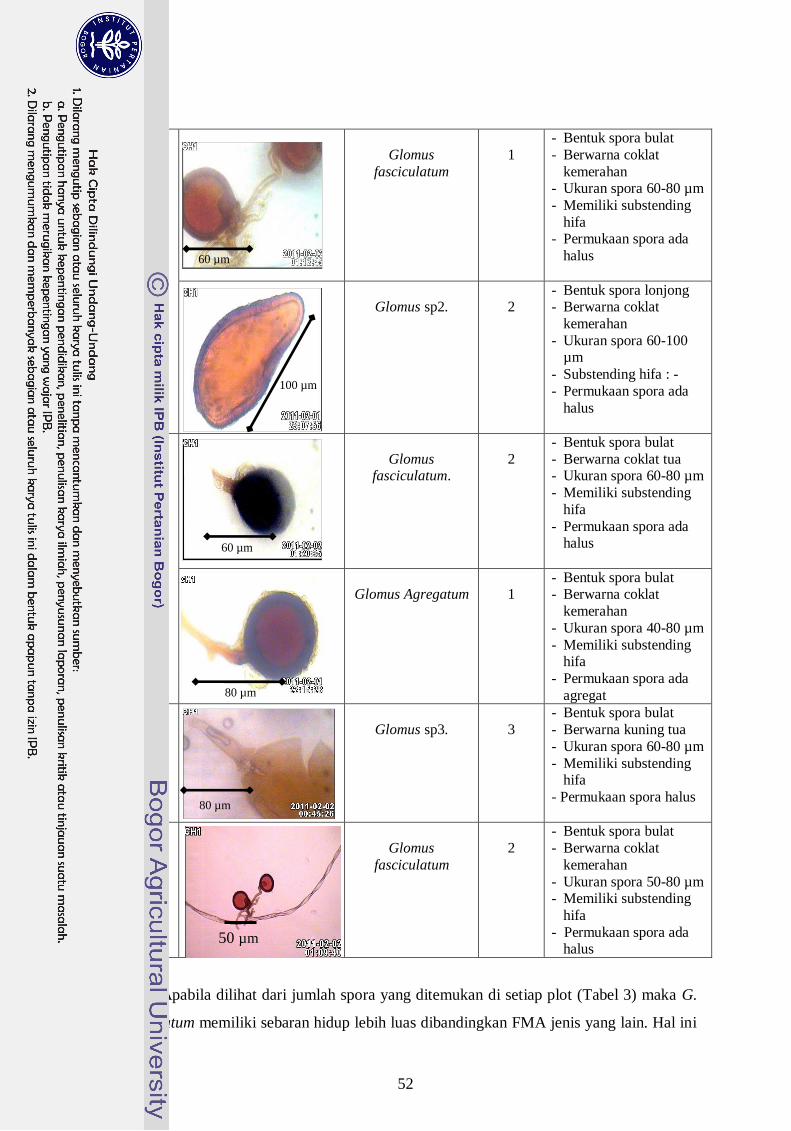
52
Plot 3
Glomus
fasciculatum
1
- Bentuk spora bulat
- Berwarna coklat
kemerahan - Ukuran spora 60-80 µm
- Memiliki substending
hifa - Permukaan spora ada
halus
Glomus sp2.
2
- Bentuk spora lonjong - Berwarna coklat
kemerahan
- Ukuran spora 60-100 µm
- Substending hifa : -
- Permukaan spora ada
halus
Plot 4
Glomus fasciculatum.
2
- Bentuk spora bulat
- Berwarna coklat tua - Ukuran spora 60-80 µm
- Memiliki substending
hifa
- Permukaan spora ada halus
Glomus Agregatum
1
- Bentuk spora bulat - Berwarna coklat
kemerahan
- Ukuran spora 40-80 µm
- Memiliki substending hifa
- Permukaan spora ada
agregat
Plot 5
Glomus sp3.
3
- Bentuk spora bulat
- Berwarna kuning tua
- Ukuran spora 60-80 µm
- Memiliki substending hifa
- Permukaan spora halus
Glomus
fasciculatum
2
- Bentuk spora bulat
- Berwarna coklat
kemerahan
- Ukuran spora 50-80 µm - Memiliki substending
hifa
- Permukaan spora ada halus
Apabila dilihat dari jumlah spora yang ditemukan di setiap plot (Tabel 3) maka G.
fasciculatum memiliki sebaran hidup lebih luas dibandingkan FMA jenis yang lain. Hal ini
60 µm
100 µm
100 µm
80 µm
60 µm
80 µm
50 µm 50 µm
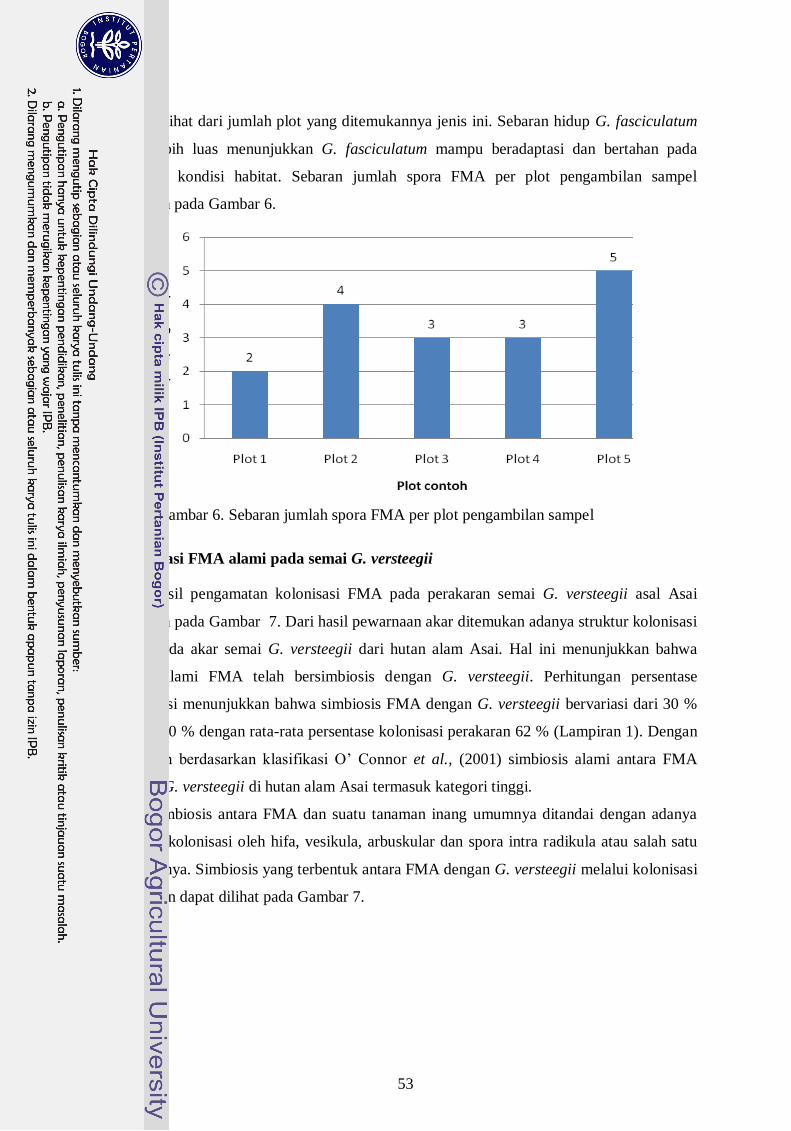
53
dapat dilihat dari jumlah plot yang ditemukannya jenis ini. Sebaran hidup G. fasciculatum
yang lebih luas menunjukkan G. fasciculatum mampu beradaptasi dan bertahan pada
berbagai kondisi habitat. Sebaran jumlah spora FMA per plot pengambilan sampel
disajikan pada Gambar 6.
Gambar 6. Sebaran jumlah spora FMA per plot pengambilan sampel
Kolonisasi FMA alami pada semai G. versteegii
Hasil pengamatan kolonisasi FMA pada perakaran semai G. versteegii asal Asai
disajikan pada Gambar 7. Dari hasil pewarnaan akar ditemukan adanya struktur kolonisasi
FMA pada akar semai G. versteegii dari hutan alam Asai. Hal ini menunjukkan bahwa
secara alami FMA telah bersimbiosis dengan G. versteegii. Perhitungan persentase
kolonisasi menunjukkan bahwa simbiosis FMA dengan G. versteegii bervariasi dari 30 %
hingga 80 % dengan rata-rata persentase kolonisasi perakaran 62 % (Lampiran 1). Dengan
demikian berdasarkan klasifikasi O’ Connor et al., (2001) simbiosis alami antara FMA
dengan G. versteegii di hutan alam Asai termasuk kategori tinggi.
Simbiosis antara FMA dan suatu tanaman inang umumnya ditandai dengan adanya
struktur kolonisasi oleh hifa, vesikula, arbuskular dan spora intra radikula atau salah satu
diantaranya. Simbiosis yang terbentuk antara FMA dengan G. versteegii melalui kolonisasi
perakaran dapat dilihat pada Gambar 7.
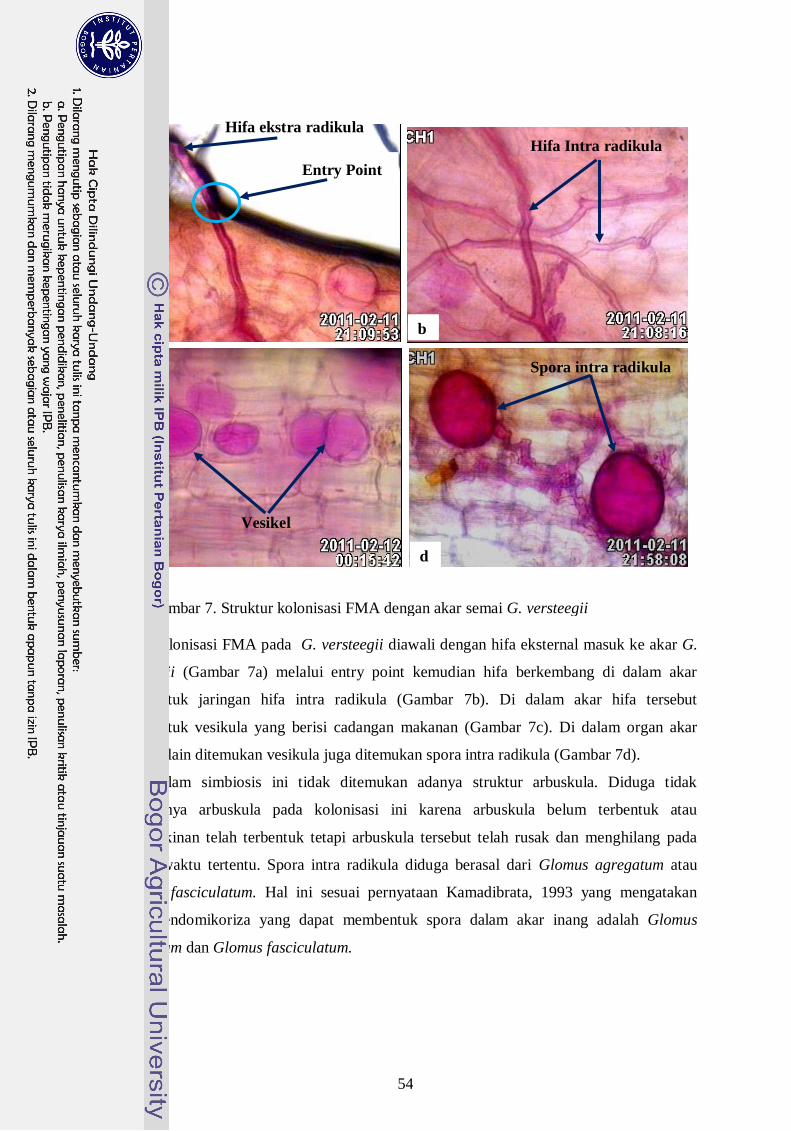
54
Kolonisasi FMA pada G. versteegii diawali dengan hifa eksternal masuk ke akar G.
versteegii (Gambar 7a) melalui entry point kemudian hifa berkembang di dalam akar
membentuk jaringan hifa intra radikula (Gambar 7b). Di dalam akar hifa tersebut
membentuk vesikula yang berisi cadangan makanan (Gambar 7c). Di dalam organ akar
selain selain ditemukan vesikula juga ditemukan spora intra radikula (Gambar 7d).
Dalam simbiosis ini tidak ditemukan adanya struktur arbuskula. Diduga tidak
terdapatnya arbuskula pada kolonisasi ini karena arbuskula belum terbentuk atau
kemungkinan telah terbentuk tetapi arbuskula tersebut telah rusak dan menghilang pada
jangka waktu tertentu. Spora intra radikula diduga berasal dari Glomus agregatum atau
Glomus fasciculatum. Hal ini sesuai pernyataan Kamadibrata, 1993 yang mengatakan
bahwa endomikoriza yang dapat membentuk spora dalam akar inang adalah Glomus
agregatum dan Glomus fasciculatum.
Hifa Ekstra radikula Entry Point
Hifa Intra radikula
Vesikel
Spora intra radikula
Gambar 7. Struktur kolonisasi FMA dengan akar semai G. versteegii
a
c
b a
d
Hifa ekstra radikula

55
3. Aplikasi FMA pada plantling gaharu G. versteegii
Kondisi Stomata plantlet dan plantling G. versteegii
Stomata merupakan bagian dari organ daun yang sangat penting dalam serapan C02
ke dalam tanaman. Mekanisme buka tutupnya stomata dipengaruhi faktor lingkungan dan
fisiologis tanaman. Mekanisme buka tutup stomata berpengaruh terhadap kinerja
fotosintesis. Bibit yang dikembangkan melalui kultur jaringan mempunyai beberapa
masalah ketika dipindahkan ke kondisi autotropik yaitu mudah layu karena transpirasi
sangat besar yang tidak diimbang serapan air yang memadai. Stomata pada hasil kultur
jaringan pada umumya dalam keadaan selalu membuka karena hidup dalam kondisi
kelembaban tinggi, nutrisi selalu terpenuhi, kondisi media yang aseptik, intensitas cahaya
yang selalu terkontrol dan konstan.
Secara visual terdapat perbedaan keadaan bentuk, kerapatan dan letak stomata pada
permukaan daun plantlet gaharu G. versteegii asal kultur in-vitro dengan plantling gaharu
G. versteegii pada percobaan di greenhouse. Plantling adalah bibit hasil in-vitro yang
sudah diaklimatisasi. Hasil pemotretan menunjukkan bahwa stomata plantlet asal kultur in-
vitro berjumlah sedikit, letak stomata tenggelam pada permukaan daun dan stomata pada
plantlet dalam keadaan terbuka sempurna (Gambar 8 a dan b), sedangkan di daun plantling
G. versteegii pada percobaan di greenhouse menunjukkan bahwa stomata berjumlah
banyak dan muncul di atas permukaan daun dalam keadaan tertutup atau tidak terbuka
penuh (Gambar 9 a & b). Diduga perbedaan ini disebabkan karena adaptasi fisiologis dan
morfologi dari tanaman untuk meningkatkan kinerja fotosintesis dengan keadaan
lingkungan.
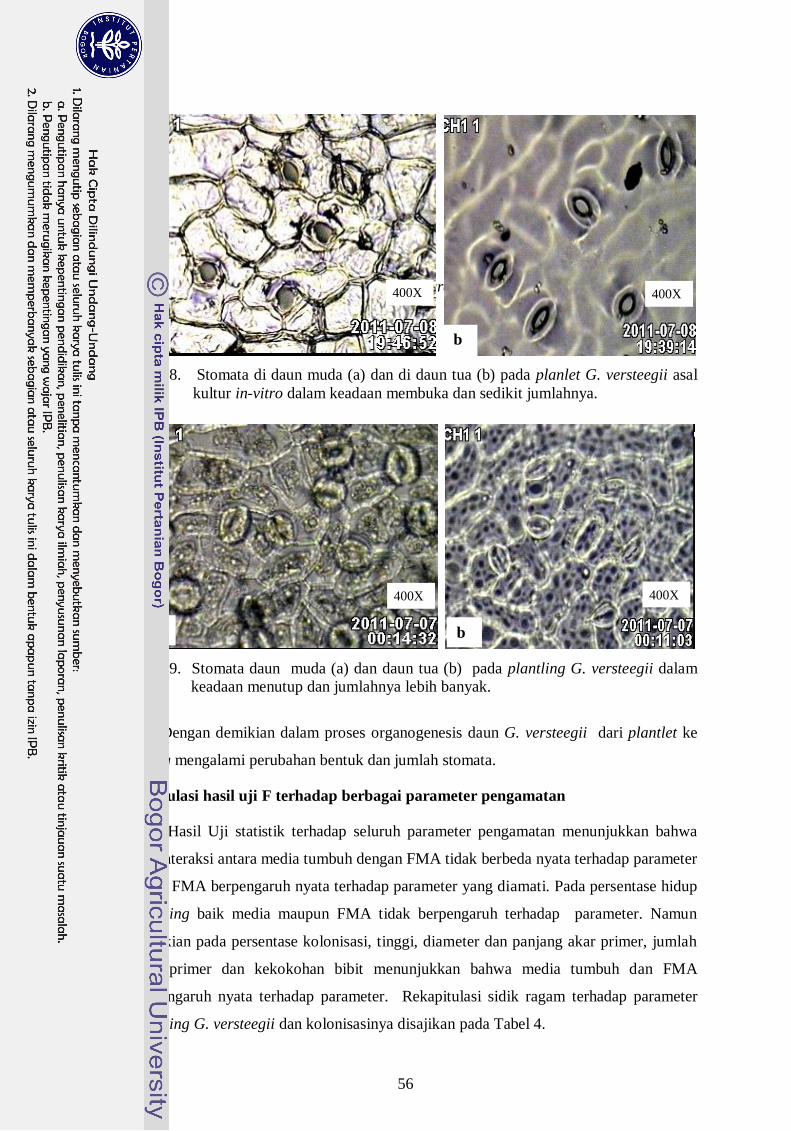
56
Pembesarx Pembesaran 400 x
Gambar 8. Stomata di daun muda (a) dan di daun tua (b) pada planlet G. versteegii asal
kultur in-vitro dalam keadaan membuka dan sedikit jumlahnya.
Gambar 9. Stomata daun muda (a) dan daun tua (b) pada plantling G. versteegii dalam
keadaan menutup dan jumlahnya lebih banyak.
Dengan demikian dalam proses organogenesis daun G. versteegii dari plantlet ke
plantling mengalami perubahan bentuk dan jumlah stomata.
Rekapitulasi hasil uji F terhadap berbagai parameter pengamatan
Hasil Uji statistik terhadap seluruh parameter pengamatan menunjukkan bahwa
dan interaksi antara media tumbuh dengan FMA tidak berbeda nyata terhadap parameter
tetapi FMA berpengaruh nyata terhadap parameter yang diamati. Pada persentase hidup
plantling baik media maupun FMA tidak berpengaruh terhadap parameter. Namun
demikian pada persentase kolonisasi, tinggi, diameter dan panjang akar primer, jumlah
akar primer dan kekokohan bibit menunjukkan bahwa media tumbuh dan FMA
berpengaruh nyata terhadap parameter. Rekapitulasi sidik ragam terhadap parameter
plantling G. versteegii dan kolonisasinya disajikan pada Tabel 4.
a b
a b
400X 400X
400X 400X
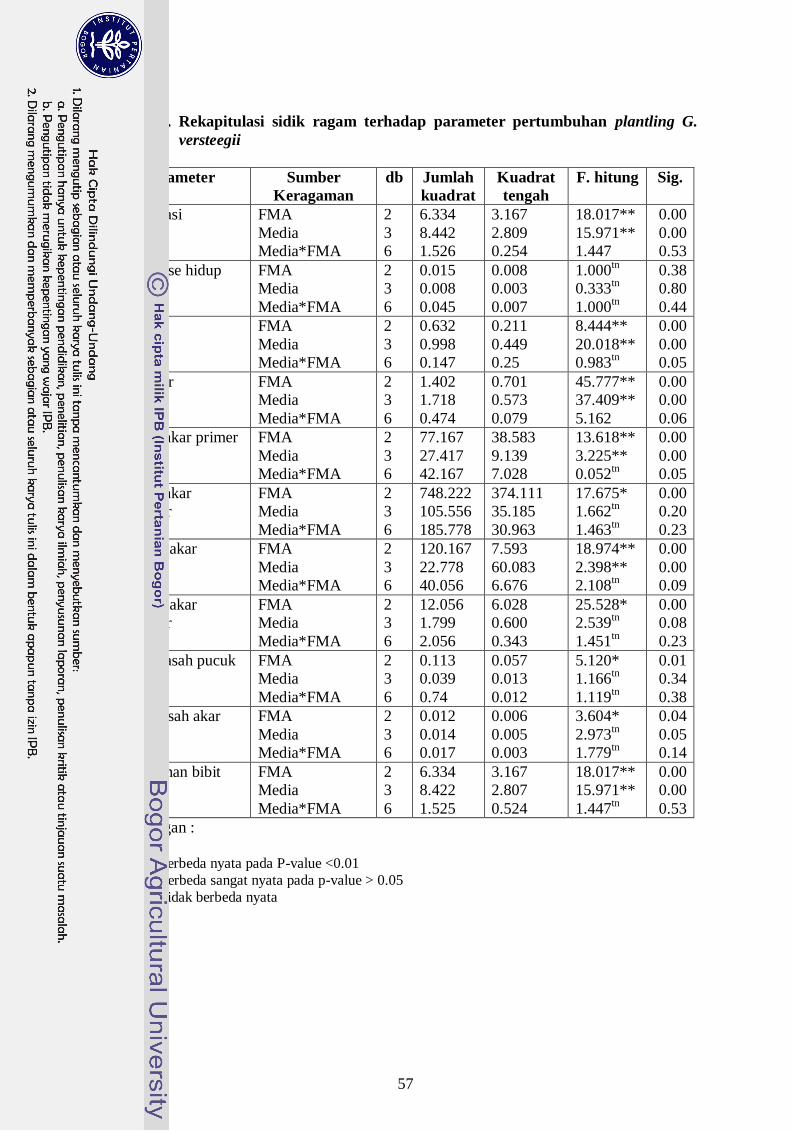
57
Tabel 4. Rekapitulasi sidik ragam terhadap parameter pertumbuhan plantling G.
versteegii
Parameter Sumber
Keragaman
db Jumlah
kuadrat
Kuadrat
tengah
F. hitung Sig.
Kolonisasi FMA
Media
Media*FMA
2
3
6
6.334
8.442
1.526
3.167
2.809
0.254
18.017**
15.971**
1.447
0.00
0.00
0.53
Persentase hidup FMA
Media
Media*FMA
2
3
6
0.015
0.008
0.045
0.008
0.003
0.007
1.000tn
0.333tn
1.000tn
0.38
0.80
0.44
Tinggi FMA
Media
Media*FMA
2
3
6
0.632
0.998
0.147
0.211
0.449
0.25
8.444**
20.018**
0.983tn
0.00
0.00
0.05
Diameter FMA
Media
Media*FMA
2
3
6
1.402
1.718
0.474
0.701
0.573
0.079
45.777**
37.409**
5.162
0.00
0.00
0.06
Jumlah akar primer FMA
Media
Media*FMA
2
3
6
77.167
27.417
42.167
38.583
9.139
7.028
13.618**
3.225**
0.052tn
0.00
0.00
0.05
Jumlah akar
sekunder
FMA
Media
Media*FMA
2
3
6
748.222
105.556
185.778
374.111
35.185
30.963
17.675*
1.662tn
1.463tn
0.00
0.20
0.23
Panjang akar
primer
FMA
Media
Media*FMA
2
3
6
120.167
22.778
40.056
7.593
60.083
6.676
18.974**
2.398**
2.108tn
0.00
0.00
0.09
Panjang akar
sekunder
FMA
Media
Media*FMA
2
3
6
12.056
1.799
2.056
6.028
0.600
0.343
25.528*
2.539tn
1.451tn
0.00
0.08
0.23
Berat basah pucuk FMA
Media
Media*FMA
2
3
6
0.113
0.039
0.74
0.057
0.013
0.012
5.120*
1.166tn
1.119tn
0.01
0.34
0.38
Berat basah akar FMA
Media
Media*FMA
2
3
6
0.012
0.014
0.017
0.006
0.005
0.003
3.604*
2.973tn
1.779tn
0.04
0.05
0.14
Kekokohan bibit FMA
Media
Media*FMA
2
3
6
6.334
8.422
1.525
3.167
2.807
0.524
18.017**
15.971**
1.447tn
0.00
0.00
0.53
Keterangan :
* = Berbeda nyata pada P-value <0.01
** = Berbeda sangat nyata pada p-value > 0.05
tn = Tidak berbeda nyata
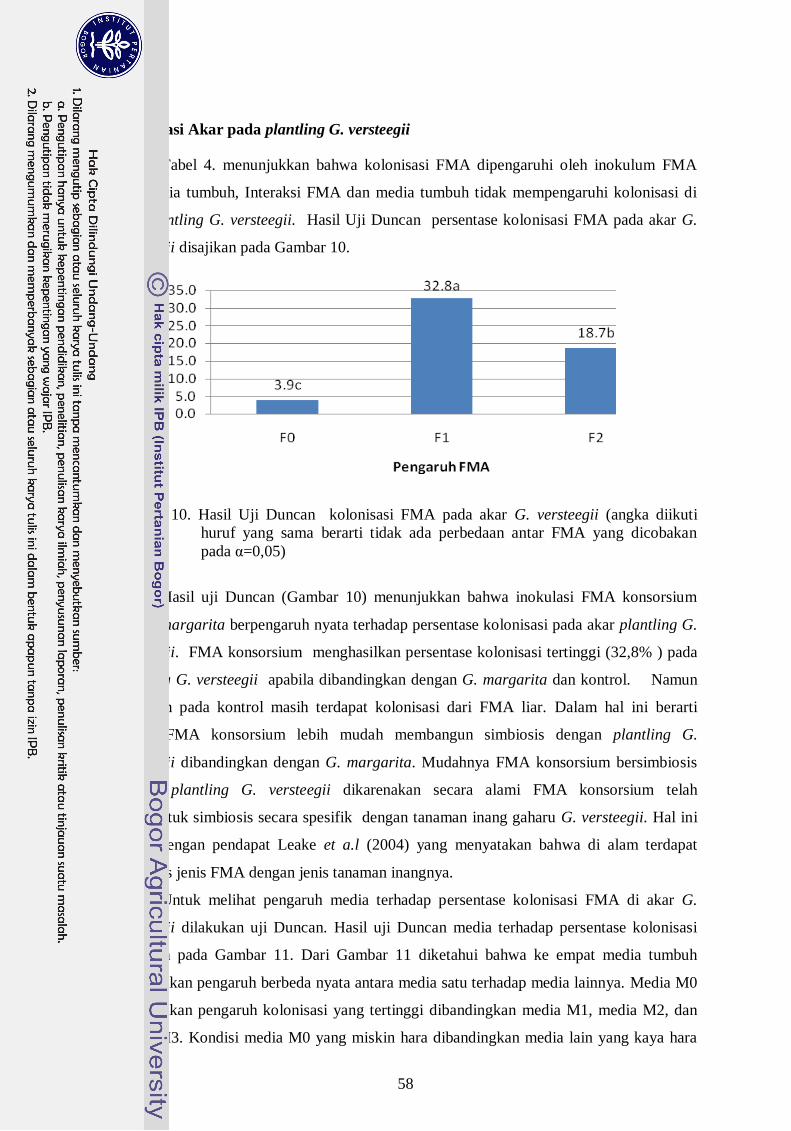
58
Kolonisasi Akar pada plantling G. versteegii
Tabel 4. menunjukkan bahwa kolonisasi FMA dipengaruhi oleh inokulum FMA
dan media tumbuh, Interaksi FMA dan media tumbuh tidak mempengaruhi kolonisasi di
akar plantling G. versteegii. Hasil Uji Duncan persentase kolonisasi FMA pada akar G.
versteegii disajikan pada Gambar 10.
Gambar 10. Hasil Uji Duncan kolonisasi FMA pada akar G. versteegii (angka diikuti
huruf yang sama berarti tidak ada perbedaan antar FMA yang dicobakan
pada α=0,05)
Hasil uji Duncan (Gambar 10) menunjukkan bahwa inokulasi FMA konsorsium
dan G. margarita berpengaruh nyata terhadap persentase kolonisasi pada akar plantling G.
versteegii. FMA konsorsium menghasilkan persentase kolonisasi tertinggi (32,8% ) pada
plantling G. versteegii apabila dibandingkan dengan G. margarita dan kontrol. Namun
demikian pada kontrol masih terdapat kolonisasi dari FMA liar. Dalam hal ini berarti
bahwa FMA konsorsium lebih mudah membangun simbiosis dengan plantling G.
versteegii dibandingkan dengan G. margarita. Mudahnya FMA konsorsium bersimbiosis
dengan plantling G. versteegii dikarenakan secara alami FMA konsorsium telah
membentuk simbiosis secara spesifik dengan tanaman inang gaharu G. versteegii. Hal ini
sesuai dengan pendapat Leake et a.l (2004) yang menyatakan bahwa di alam terdapat
spesifitas jenis FMA dengan jenis tanaman inangnya.
Untuk melihat pengaruh media terhadap persentase kolonisasi FMA di akar G.
versteegii dilakukan uji Duncan. Hasil uji Duncan media terhadap persentase kolonisasi
disajikan pada Gambar 11. Dari Gambar 11 diketahui bahwa ke empat media tumbuh
memberikan pengaruh berbeda nyata antara media satu terhadap media lainnya. Media M0
memberikan pengaruh kolonisasi yang tertinggi dibandingkan media M1, media M2, dan
media M3. Kondisi media M0 yang miskin hara dibandingkan media lain yang kaya hara
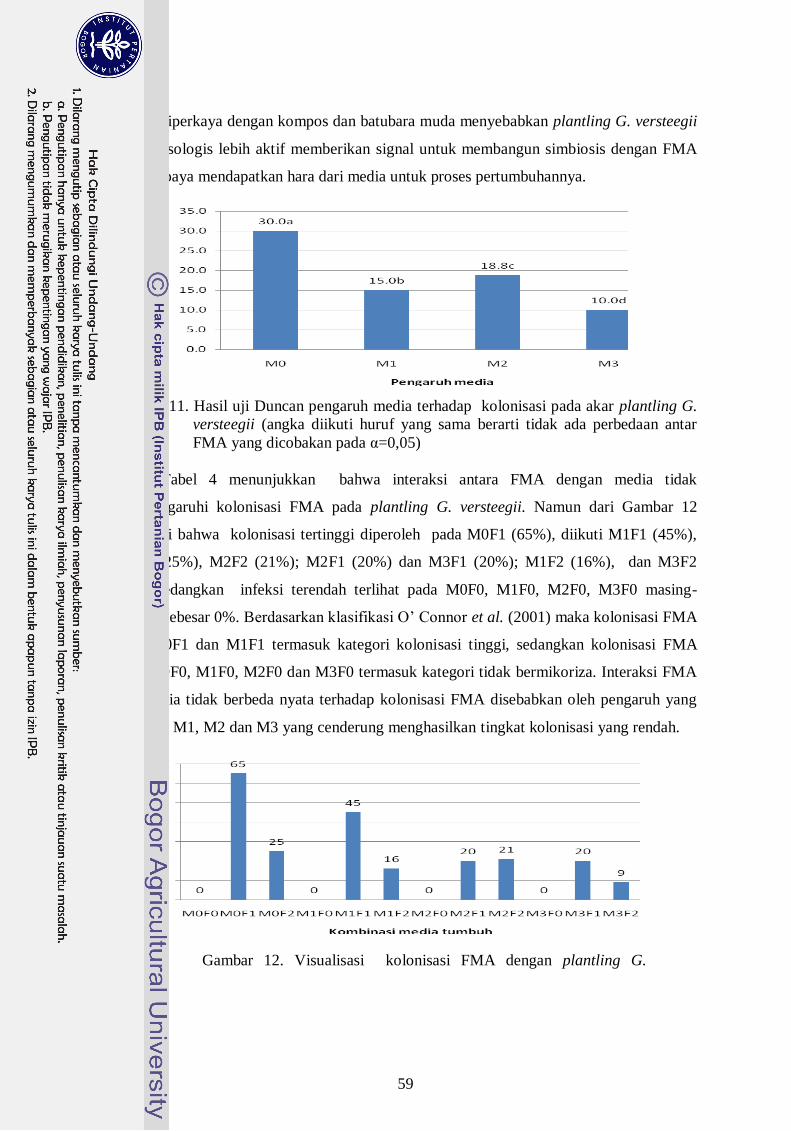
59
karena diperkaya dengan kompos dan batubara muda menyebabkan plantling G. versteegii
secara fisologis lebih aktif memberikan signal untuk membangun simbiosis dengan FMA
dalam upaya mendapatkan hara dari media untuk proses pertumbuhannya.
Gambar 11. Hasil uji Duncan pengaruh media terhadap kolonisasi pada akar plantling G.
versteegii (angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05)
Tabel 4 menunjukkan bahwa interaksi antara FMA dengan media tidak
mempengaruhi kolonisasi FMA pada plantling G. versteegii. Namun dari Gambar 12
diketahui bahwa kolonisasi tertinggi diperoleh pada M0F1 (65%), diikuti M1F1 (45%),
M0F2 (25%), M2F2 (21%); M2F1 (20%) dan M3F1 (20%); M1F2 (16%), dan M3F2
(9%), sedangkan infeksi terendah terlihat pada M0F0, M1F0, M2F0, M3F0 masing-
masing sebesar 0%. Berdasarkan klasifikasi O’ Connor et al. (2001) maka kolonisasi FMA
pada M0F1 dan M1F1 termasuk kategori kolonisasi tinggi, sedangkan kolonisasi FMA
pada M0F0, M1F0, M2F0 dan M3F0 termasuk kategori tidak bermikoriza. Interaksi FMA
dan media tidak berbeda nyata terhadap kolonisasi FMA disebabkan oleh pengaruh yang
kuat dari M1, M2 dan M3 yang cenderung menghasilkan tingkat kolonisasi yang rendah.
Gambar 12. Visualisasi kolonisasi FMA dengan plantling G.
versteegii
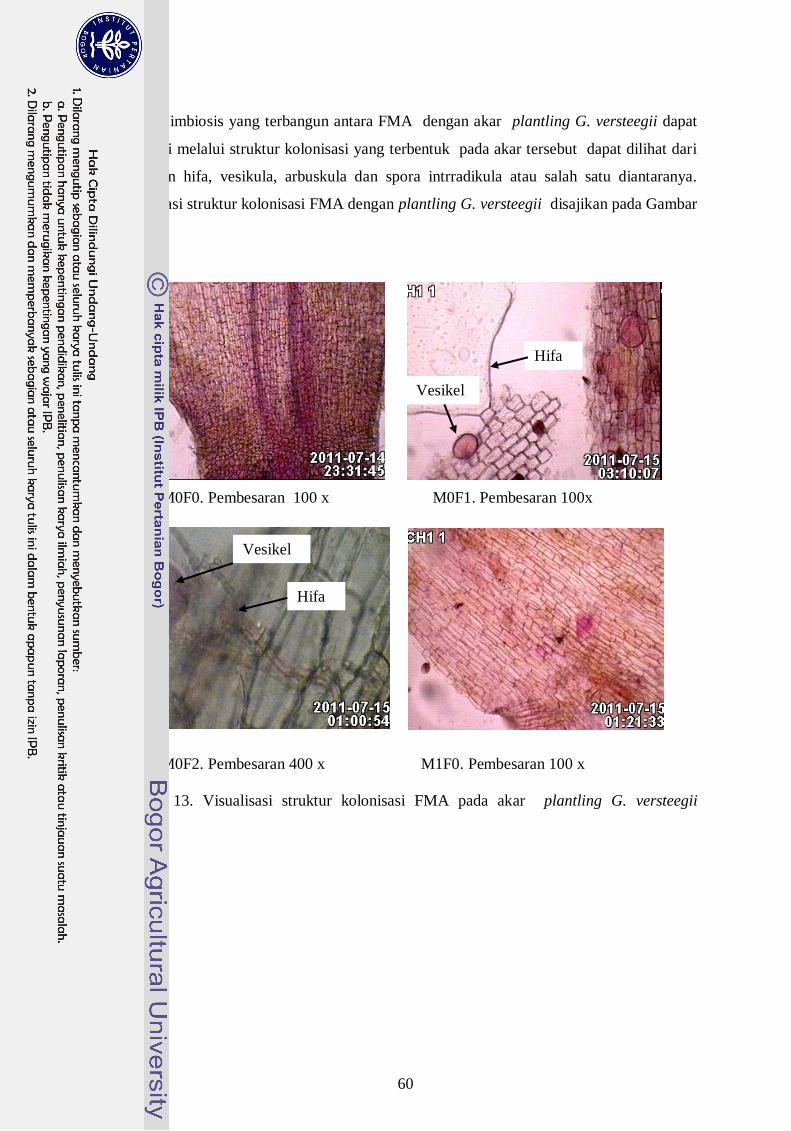
60
Simbiosis yang terbangun antara FMA dengan akar plantling G. versteegii dapat
diketahui melalui struktur kolonisasi yang terbentuk pada akar tersebut dapat dilihat dari
kehadiran hifa, vesikula, arbuskula dan spora intrradikula atau salah satu diantaranya.
Visualisasi struktur kolonisasi FMA dengan plantling G. versteegii disajikan pada Gambar
13.
M0F0. Pembesaran 100 x M0F1. Pembesaran 100x
M0F2. Pembesaran 400 x M1F0. Pembesaran 100 x
Gambar 13. Visualisasi struktur kolonisasi FMA pada akar plantling G. versteegii
Vesikel
Hifa
Vesikel
Hifa
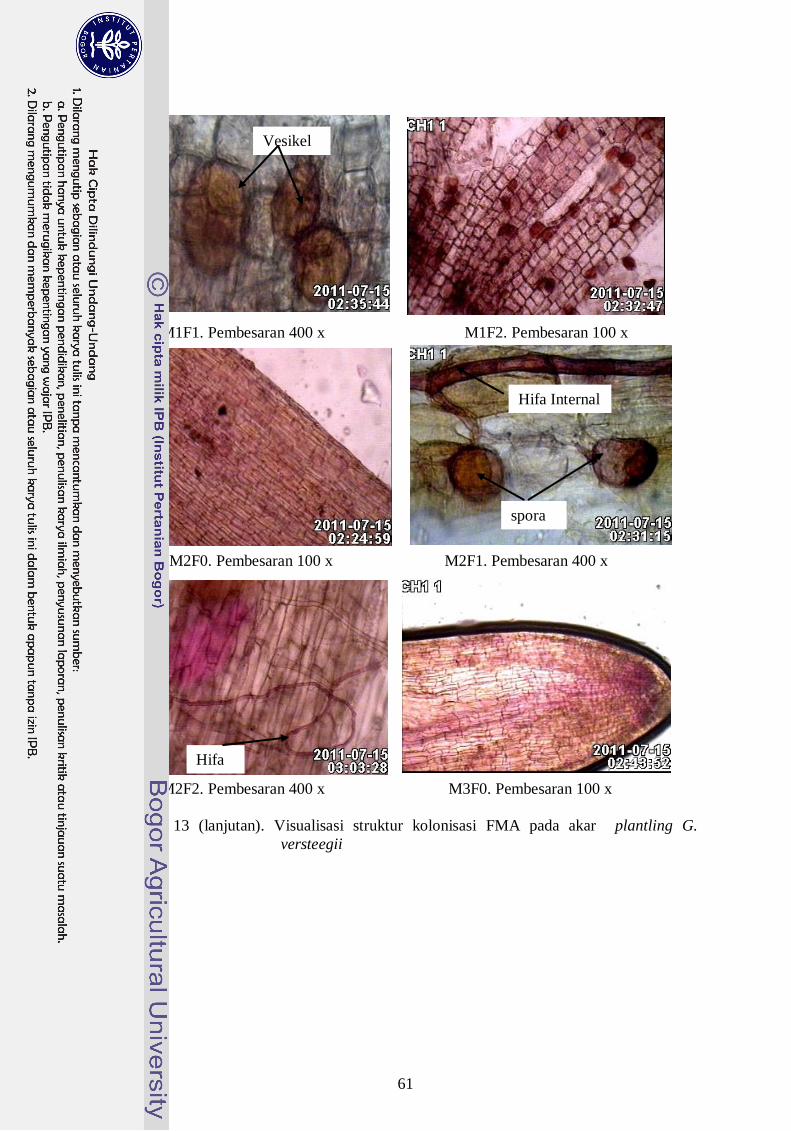
61
M1F1. Pembesaran 400 x M1F2. Pembesaran 100 x
M2F0. Pembesaran 100 x M2F1. Pembesaran 400 x
M2F2. Pembesaran 400 x M3F0. Pembesaran 100 x
Gambar 13 (lanjutan). Visualisasi struktur kolonisasi FMA pada akar plantling G.
versteegii
Vesikel
spora
Hifa Internal
Hifa
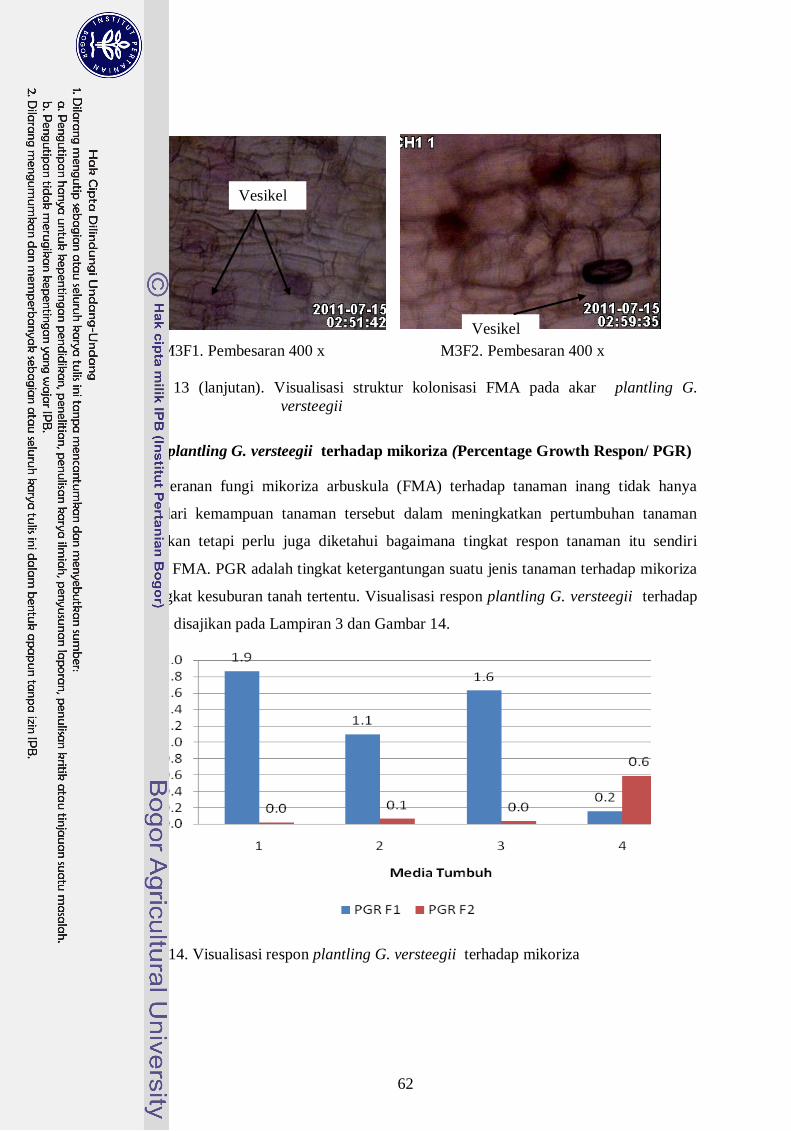
62
M2F2. Pembesaran 100 x M3F0. Pembesaran 100 x
M3F1. Pembesaran 400 x M3F2. Pembesaran 400 x
Gambar 13 (lanjutan). Visualisasi struktur kolonisasi FMA pada akar plantling G.
versteegii
Respon plantling G. versteegii terhadap mikoriza (Percentage Growth Respon/ PGR)
Peranan fungi mikoriza arbuskula (FMA) terhadap tanaman inang tidak hanya
dilihat dari kemampuan tanaman tersebut dalam meningkatkan pertumbuhan tanaman
inang, akan tetapi perlu juga diketahui bagaimana tingkat respon tanaman itu sendiri
terhadap FMA. PGR adalah tingkat ketergantungan suatu jenis tanaman terhadap mikoriza
pada tingkat kesuburan tanah tertentu. Visualisasi respon plantling G. versteegii terhadap
mikoriza disajikan pada Lampiran 3 dan Gambar 14.
Gambar 14. Visualisasi respon plantling G. versteegii terhadap mikoriza
Vesikel
Vesikel
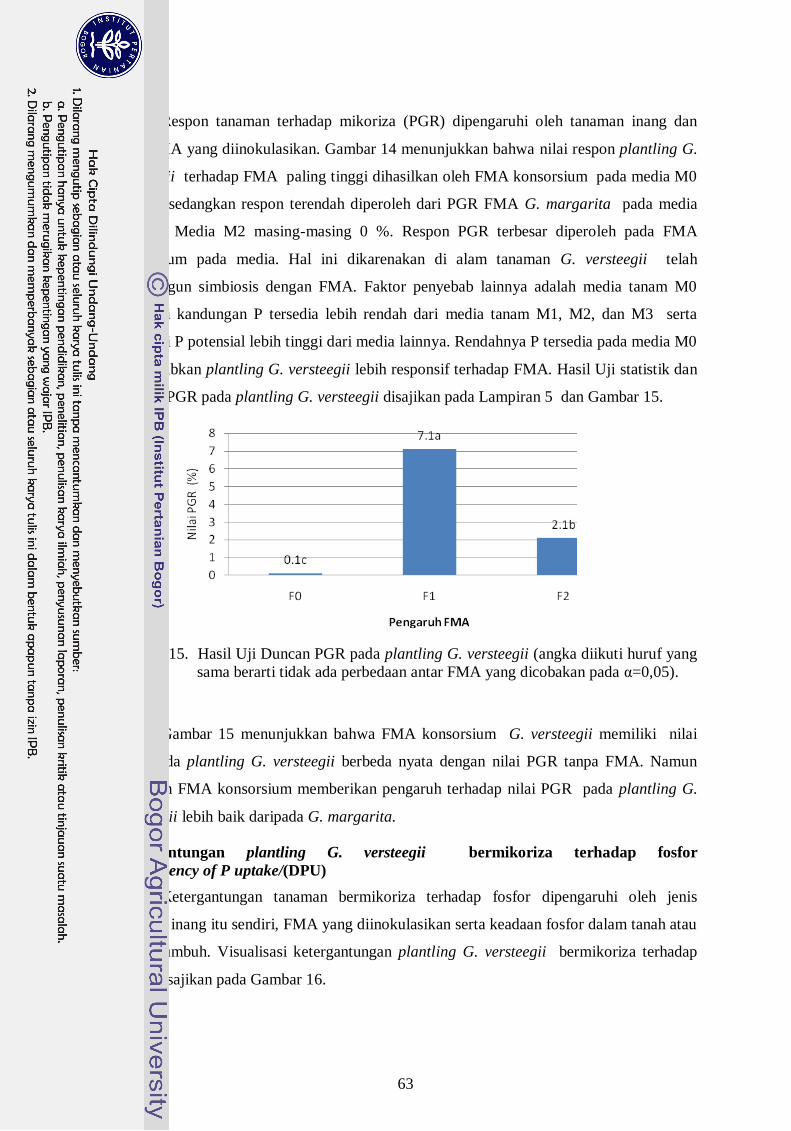
63
Respon tanaman terhadap mikoriza (PGR) dipengaruhi oleh tanaman inang dan
jenis FMA yang diinokulasikan. Gambar 14 menunjukkan bahwa nilai respon plantling G.
versteegii terhadap FMA paling tinggi dihasilkan oleh FMA konsorsium pada media M0
(19 %), sedangkan respon terendah diperoleh dari PGR FMA G. margarita pada media
M0 dan Media M2 masing-masing 0 %. Respon PGR terbesar diperoleh pada FMA
konsorsium pada media. Hal ini dikarenakan di alam tanaman G. versteegii telah
membangun simbiosis dengan FMA. Faktor penyebab lainnya adalah media tanam M0
memiliki kandungan P tersedia lebih rendah dari media tanam M1, M2, dan M3 serta
memiliki P potensial lebih tinggi dari media lainnya. Rendahnya P tersedia pada media M0
menyebabkan plantling G. versteegii lebih responsif terhadap FMA. Hasil Uji statistik dan
Duncan PGR pada plantling G. versteegii disajikan pada Lampiran 5 dan Gambar 15.
Gambar 15. Hasil Uji Duncan PGR pada plantling G. versteegii (angka diikuti huruf yang
sama berarti tidak ada perbedaan antar FMA yang dicobakan pada α=0,05).
Gambar 15 menunjukkan bahwa FMA konsorsium G. versteegii memiliki nilai
PGR pada plantling G. versteegii berbeda nyata dengan nilai PGR tanpa FMA. Namun
demikian FMA konsorsium memberikan pengaruh terhadap nilai PGR pada plantling G.
Versteegii lebih baik daripada G. margarita.
Ketergantungan plantling G. versteegii bermikoriza terhadap fosfor
(Dependency of P uptake/(DPU)
Ketergantungan tanaman bermikoriza terhadap fosfor dipengaruhi oleh jenis
tanaman inang itu sendiri, FMA yang diinokulasikan serta keadaan fosfor dalam tanah atau
media tumbuh. Visualisasi ketergantungan plantling G. versteegii bermikoriza terhadap
fosfor disajikan pada Gambar 16.
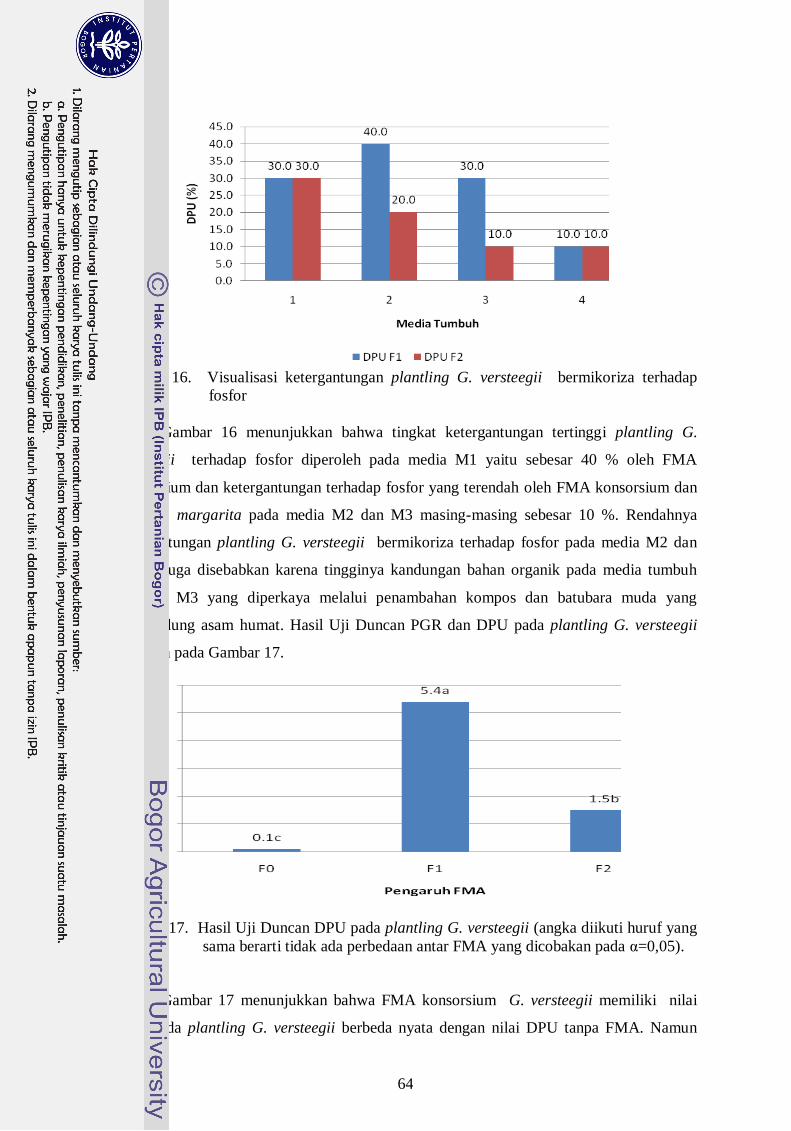
64
Gambar 16. Visualisasi ketergantungan plantling G. versteegii bermikoriza terhadap
fosfor
Gambar 16 menunjukkan bahwa tingkat ketergantungan tertinggi plantling G.
versteegii terhadap fosfor diperoleh pada media M1 yaitu sebesar 40 % oleh FMA
Konsorsium dan ketergantungan terhadap fosfor yang terendah oleh FMA konsorsium dan
FMA G. margarita pada media M2 dan M3 masing-masing sebesar 10 %. Rendahnya
ketergantungan plantling G. versteegii bermikoriza terhadap fosfor pada media M2 dan
M3 diduga disebabkan karena tingginya kandungan bahan organik pada media tumbuh
M2 dan M3 yang diperkaya melalui penambahan kompos dan batubara muda yang
mengandung asam humat. Hasil Uji Duncan PGR dan DPU pada plantling G. versteegii
disajikan pada Gambar 17.
Gambar 17. Hasil Uji Duncan DPU pada plantling G. versteegii (angka diikuti huruf yang
sama berarti tidak ada perbedaan antar FMA yang dicobakan pada α=0,05).
Gambar 17 menunjukkan bahwa FMA konsorsium G. versteegii memiliki nilai
DPU pada plantling G. versteegii berbeda nyata dengan nilai DPU tanpa FMA. Namun
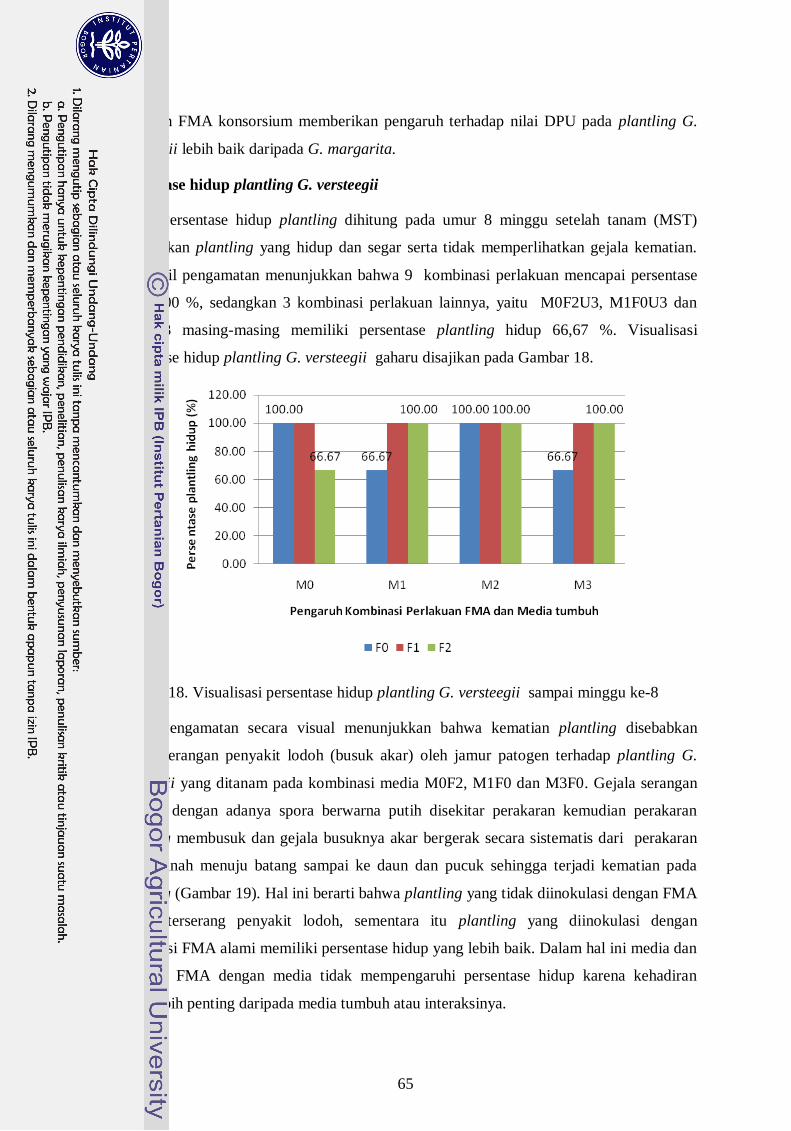
65
demikian FMA konsorsium memberikan pengaruh terhadap nilai DPU pada plantling G.
Versteegii lebih baik daripada G. margarita.
Persentase hidup plantling G. versteegii
Persentase hidup plantling dihitung pada umur 8 minggu setelah tanam (MST)
berdasarkan plantling yang hidup dan segar serta tidak memperlihatkan gejala kematian.
Dari hasil pengamatan menunjukkan bahwa 9 kombinasi perlakuan mencapai persentase
hidup 100 %, sedangkan 3 kombinasi perlakuan lainnya, yaitu M0F2U3, M1F0U3 dan
M3F0U3 masing-masing memiliki persentase plantling hidup 66,67 %. Visualisasi
persentase hidup plantling G. versteegii gaharu disajikan pada Gambar 18.
Gambar 18. Visualisasi persentase hidup plantling G. versteegii sampai minggu ke-8
Pengamatan secara visual menunjukkan bahwa kematian plantling disebabkan
karena serangan penyakit lodoh (busuk akar) oleh jamur patogen terhadap plantling G.
versteegii yang ditanam pada kombinasi media M0F2, M1F0 dan M3F0. Gejala serangan
ditandai dengan adanya spora berwarna putih disekitar perakaran kemudian perakaran
plantling membusuk dan gejala busuknya akar bergerak secara sistematis dari perakaran
dalam tanah menuju batang sampai ke daun dan pucuk sehingga terjadi kematian pada
plantling (Gambar 19). Hal ini berarti bahwa plantling yang tidak diinokulasi dengan FMA
mudah terserang penyakit lodoh, sementara itu plantling yang diinokulasi dengan
kolonisasi FMA alami memiliki persentase hidup yang lebih baik. Dalam hal ini media dan
interaksi FMA dengan media tidak mempengaruhi persentase hidup karena kehadiran
FMA lebih penting daripada media tumbuh atau interaksinya.
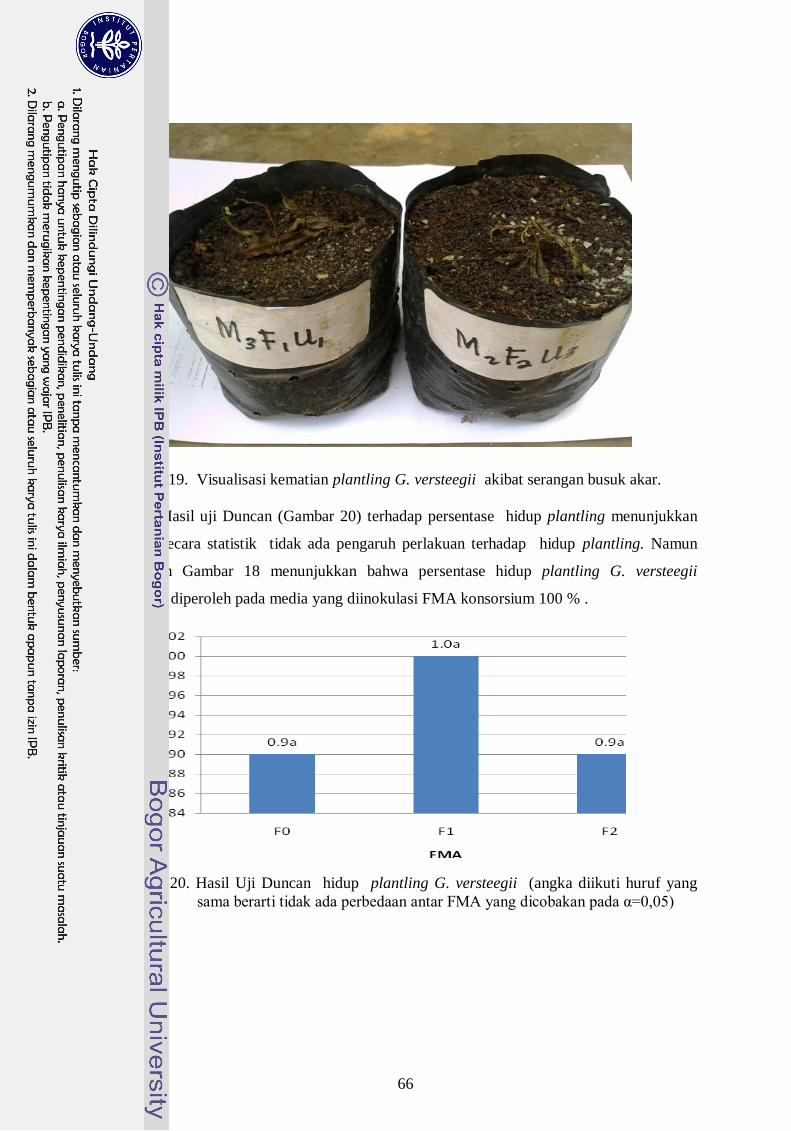
66
Gambar 19. Visualisasi kematian plantling G. versteegii akibat serangan busuk akar.
Hasil uji Duncan (Gambar 20) terhadap persentase hidup plantling menunjukkan
bahwa secara statistik tidak ada pengaruh perlakuan terhadap hidup plantling. Namun
demikian Gambar 18 menunjukkan bahwa persentase hidup plantling G. versteegii
tertinggi diperoleh pada media yang diinokulasi FMA konsorsium 100 % .
Gambar 20. Hasil Uji Duncan hidup plantling G. versteegii (angka diikuti huruf yang
sama berarti tidak ada perbedaan antar FMA yang dicobakan pada α=0,05)

67
Pertambahan tinggi plantling G. versteegii
Parameter tinggi dan diameter digunakan sebagai indikator parameter pertumbuhan
plantling G. versteegii untuk menduga pengaruh lingkungan atau perlakuan yang
dicobakan. Hasil sidik ragam menunjukkan bahwa FMA dan media berpengaruh terhadap
pertambahan tinggi plantling G. Versteegii (Tabel 4). Visualisasi dinamika pertumbuhan
tinggi plantling G. versteegii sampai akhir pengamatan disajikan pada Gambar 21.
Gambar 21. Visualisasi keadaan tinggi plantling G. versteegii pada berbagai kombinasi
media yang dicobakan
Hasil uji Duncan pengaruh FMA (Gambar 22) menunjukkan bahwa secara statistik
ada perbedaan pengaruh FMA yang dicobakan terhadap pertumbuhan tinggi plantling,
yaitu perlakuan dengan FMA konsorsium memberikan pengaruh lebih baik daripada
dengan G. margarita dan tanpa FMA begitu pula perlakuan dengan G. margarita
memberikan pengaruh lebih tinggi dengan tanpa FMA. Hal ini berarti FMA alami lebih
baik daripada FMA G. margarita.
C
B A
D
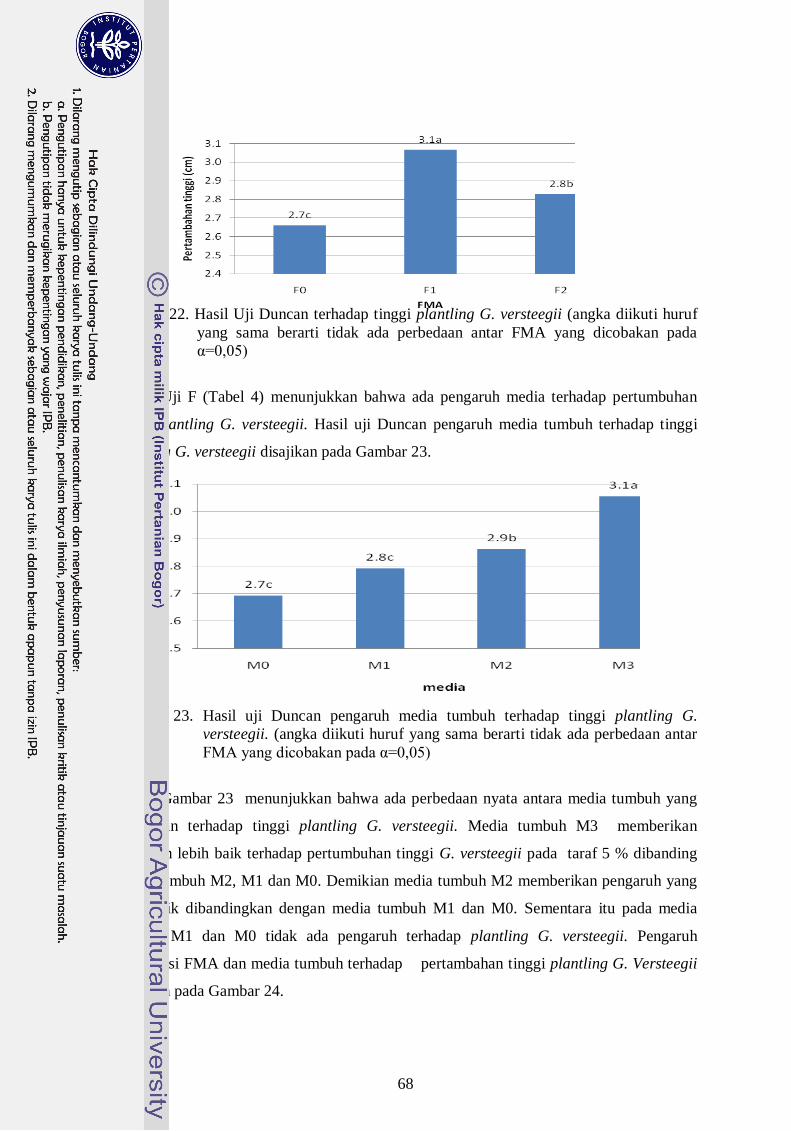
68
Gambar 22. Hasil Uji Duncan terhadap tinggi plantling G. versteegii (angka diikuti huruf
yang sama berarti tidak ada perbedaan antar FMA yang dicobakan pada
α=0,05)
Uji F (Tabel 4) menunjukkan bahwa ada pengaruh media terhadap pertumbuhan
tinggi plantling G. versteegii. Hasil uji Duncan pengaruh media tumbuh terhadap tinggi
plantling G. versteegii disajikan pada Gambar 23.
Gambar 23. Hasil uji Duncan pengaruh media tumbuh terhadap tinggi plantling G.
versteegii. (angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05)
Gambar 23 menunjukkan bahwa ada perbedaan nyata antara media tumbuh yang
dicobakan terhadap tinggi plantling G. versteegii. Media tumbuh M3 memberikan
pengaruh lebih baik terhadap pertumbuhan tinggi G. versteegii pada taraf 5 % dibanding
media tumbuh M2, M1 dan M0. Demikian media tumbuh M2 memberikan pengaruh yang
lebih baik dibandingkan dengan media tumbuh M1 dan M0. Sementara itu pada media
tumbuh M1 dan M0 tidak ada pengaruh terhadap plantling G. versteegii. Pengaruh
kombinasi FMA dan media tumbuh terhadap pertambahan tinggi plantling G. Versteegii
disajikan pada Gambar 24.
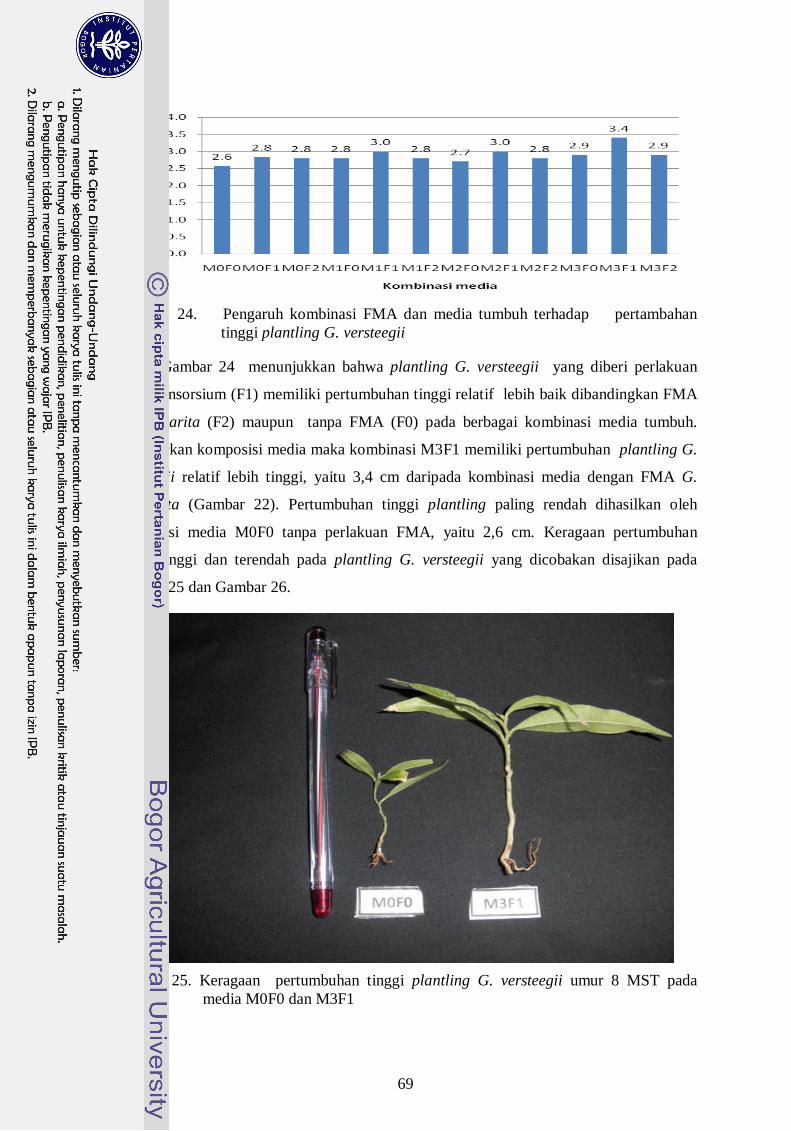
69
Gambar 24. Pengaruh kombinasi FMA dan media tumbuh terhadap pertambahan
tinggi plantling G. versteegii
Gambar 24 menunjukkan bahwa plantling G. versteegii yang diberi perlakuan
FMA konsorsium (F1) memiliki pertumbuhan tinggi relatif lebih baik dibandingkan FMA
G. margarita (F2) maupun tanpa FMA (F0) pada berbagai kombinasi media tumbuh.
Berdasarkan komposisi media maka kombinasi M3F1 memiliki pertumbuhan plantling G.
versteegii relatif lebih tinggi, yaitu 3,4 cm daripada kombinasi media dengan FMA G.
Margarita (Gambar 22). Pertumbuhan tinggi plantling paling rendah dihasilkan oleh
kombinasi media M0F0 tanpa perlakuan FMA, yaitu 2,6 cm. Keragaan pertumbuhan
paling tinggi dan terendah pada plantling G. versteegii yang dicobakan disajikan pada
Gambar 25 dan Gambar 26.
Gambar 25. Keragaan pertumbuhan tinggi plantling G. versteegii umur 8 MST pada
media M0F0 dan M3F1
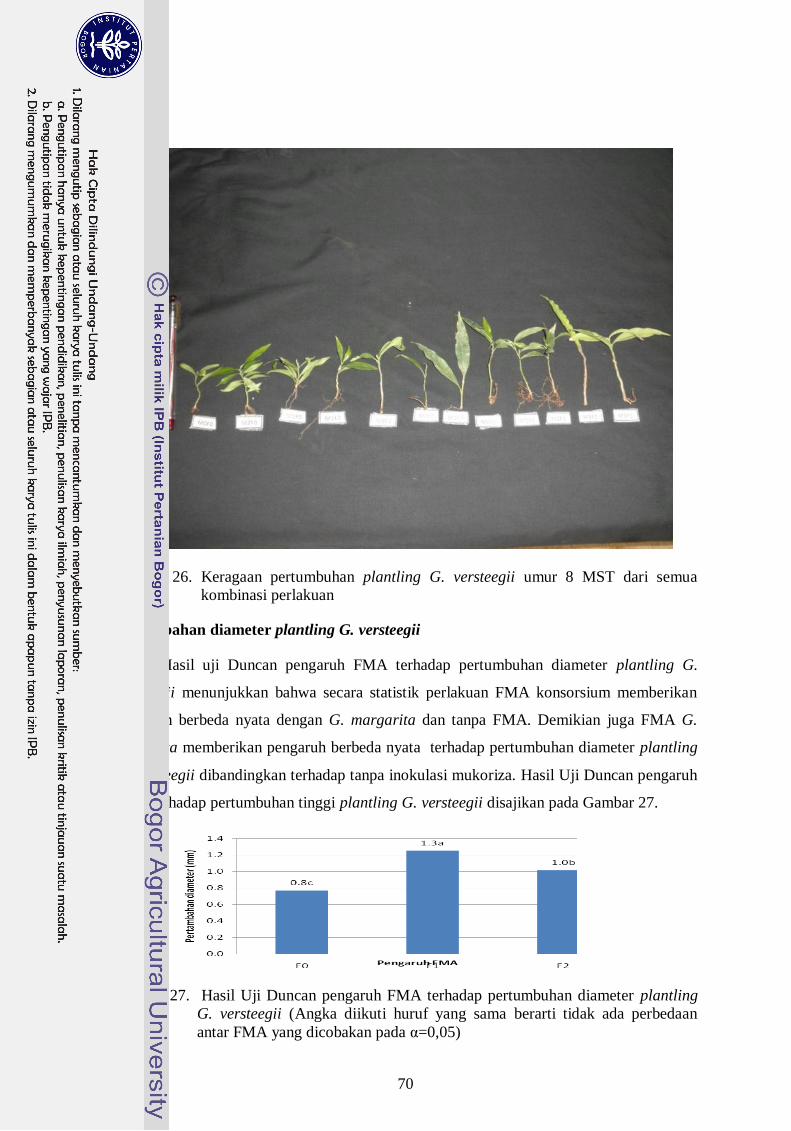
70
Gambar 26. Keragaan pertumbuhan plantling G. versteegii umur 8 MST dari semua
kombinasi perlakuan
Pertambahan diameter plantling G. versteegii
Hasil uji Duncan pengaruh FMA terhadap pertumbuhan diameter plantling G.
versteegii menunjukkan bahwa secara statistik perlakuan FMA konsorsium memberikan
pengaruh berbeda nyata dengan G. margarita dan tanpa FMA. Demikian juga FMA G.
margarita memberikan pengaruh berbeda nyata terhadap pertumbuhan diameter plantling
G. versteegii dibandingkan terhadap tanpa inokulasi mukoriza. Hasil Uji Duncan pengaruh
FMA terhadap pertumbuhan tinggi plantling G. versteegii disajikan pada Gambar 27.
Gambar 27. Hasil Uji Duncan pengaruh FMA terhadap pertumbuhan diameter plantling
G. versteegii (Angka diikuti huruf yang sama berarti tidak ada perbedaan
antar FMA yang dicobakan pada α=0,05)
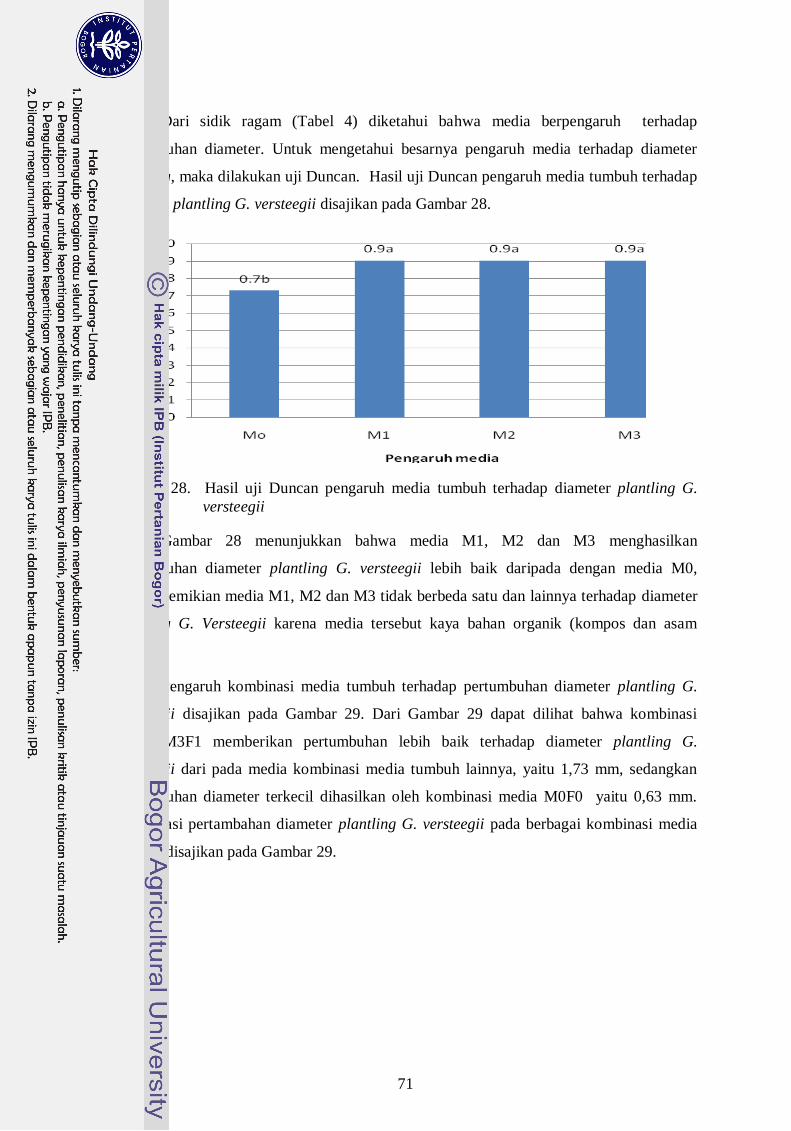
71
Dari sidik ragam (Tabel 4) diketahui bahwa media berpengaruh terhadap
pertumbuhan diameter. Untuk mengetahui besarnya pengaruh media terhadap diameter
plantling, maka dilakukan uji Duncan. Hasil uji Duncan pengaruh media tumbuh terhadap
diameter plantling G. versteegii disajikan pada Gambar 28.
Gambar 28. Hasil uji Duncan pengaruh media tumbuh terhadap diameter plantling G.
versteegii
Gambar 28 menunjukkan bahwa media M1, M2 dan M3 menghasilkan
pertumbuhan diameter plantling G. versteegii lebih baik daripada dengan media M0,
namun demikian media M1, M2 dan M3 tidak berbeda satu dan lainnya terhadap diameter
plantling G. Versteegii karena media tersebut kaya bahan organik (kompos dan asam
humat).
Pengaruh kombinasi media tumbuh terhadap pertumbuhan diameter plantling G.
versteegii disajikan pada Gambar 29. Dari Gambar 29 dapat dilihat bahwa kombinasi
media M3F1 memberikan pertumbuhan lebih baik terhadap diameter plantling G.
versteegii dari pada media kombinasi media tumbuh lainnya, yaitu 1,73 mm, sedangkan
pertumbuhan diameter terkecil dihasilkan oleh kombinasi media M0F0 yaitu 0,63 mm.
Visualisasi pertambahan diameter plantling G. versteegii pada berbagai kombinasi media
tumbuh disajikan pada Gambar 29.
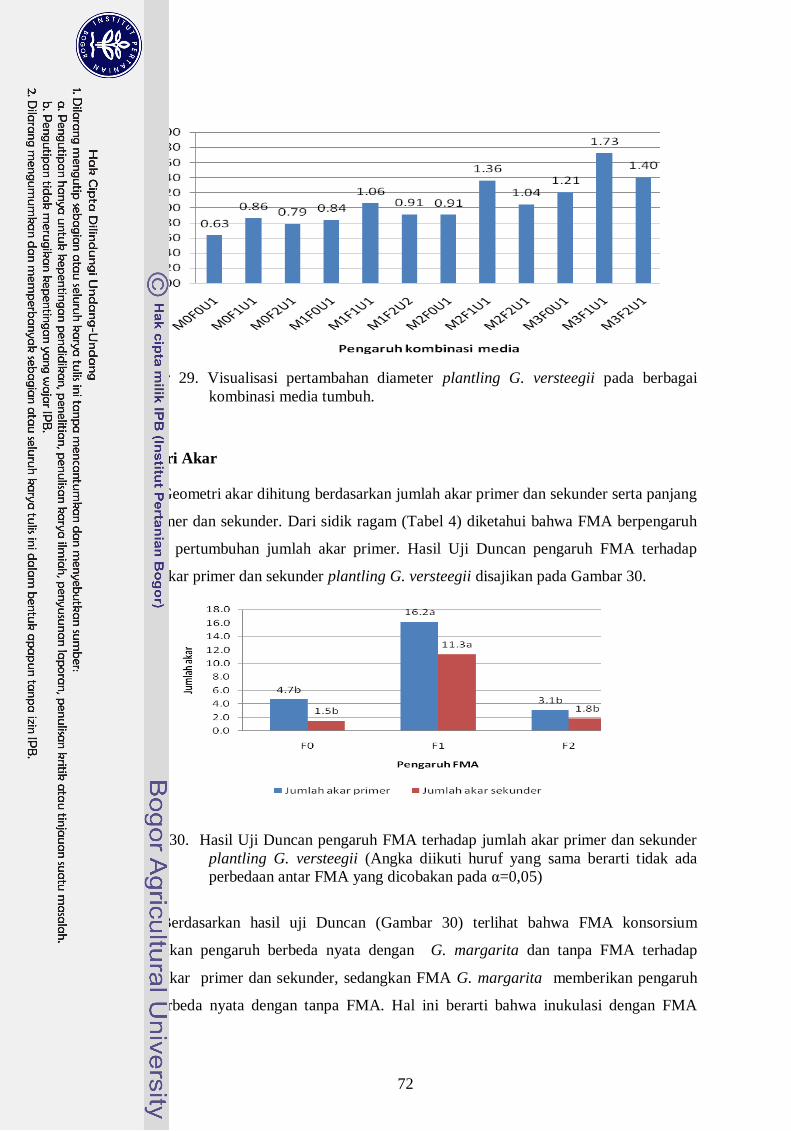
72
Gambar 29. Visualisasi pertambahan diameter plantling G. versteegii pada berbagai
kombinasi media tumbuh.
Geometri Akar
Geometri akar dihitung berdasarkan jumlah akar primer dan sekunder serta panjang
akar primer dan sekunder. Dari sidik ragam (Tabel 4) diketahui bahwa FMA berpengaruh
terhadap pertumbuhan jumlah akar primer. Hasil Uji Duncan pengaruh FMA terhadap
jumlah akar primer dan sekunder plantling G. versteegii disajikan pada Gambar 30.
Gambar 30. Hasil Uji Duncan pengaruh FMA terhadap jumlah akar primer dan sekunder
plantling G. versteegii (Angka diikuti huruf yang sama berarti tidak ada
perbedaan antar FMA yang dicobakan pada α=0,05)
Berdasarkan hasil uji Duncan (Gambar 30) terlihat bahwa FMA konsorsium
memberikan pengaruh berbeda nyata dengan G. margarita dan tanpa FMA terhadap
jumlah akar primer dan sekunder, sedangkan FMA G. margarita memberikan pengaruh
tidak berbeda nyata dengan tanpa FMA. Hal ini berarti bahwa inukulasi dengan FMA
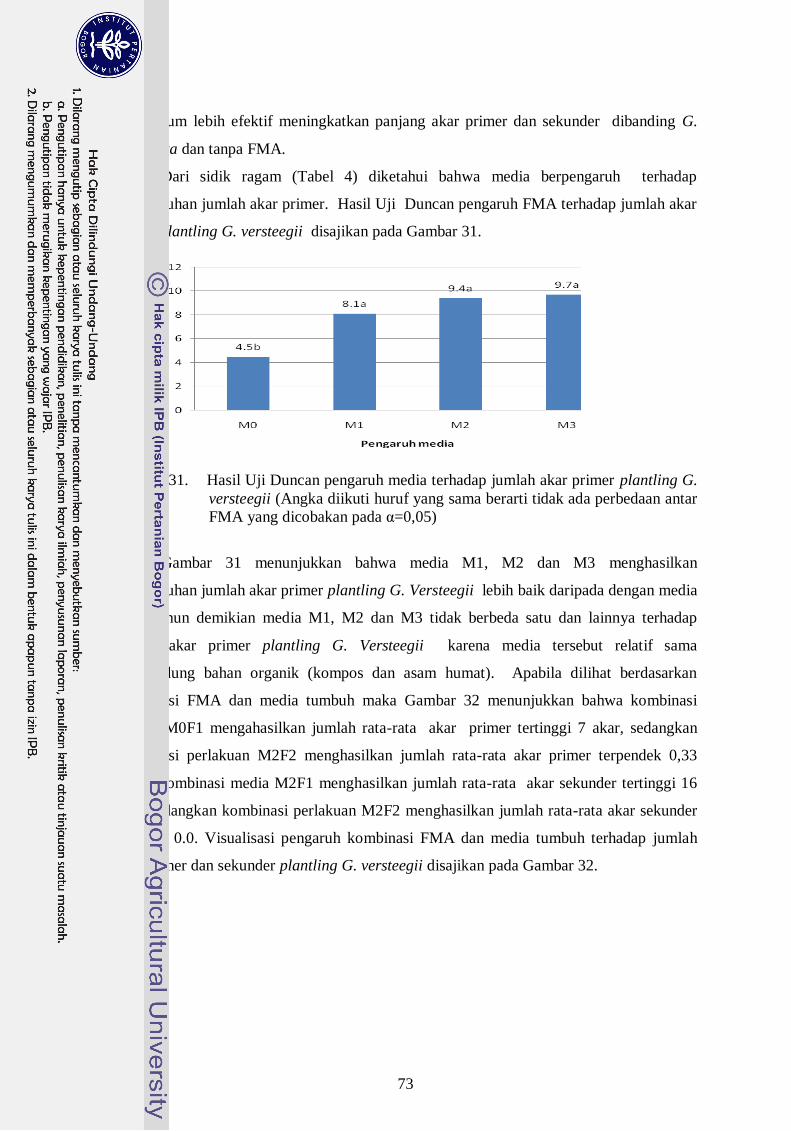
73
konsorsium lebih efektif meningkatkan panjang akar primer dan sekunder dibanding G.
margarita dan tanpa FMA.
Dari sidik ragam (Tabel 4) diketahui bahwa media berpengaruh terhadap
pertumbuhan jumlah akar primer. Hasil Uji Duncan pengaruh FMA terhadap jumlah akar
primer plantling G. versteegii disajikan pada Gambar 31.
Gambar 31. Hasil Uji Duncan pengaruh media terhadap jumlah akar primer plantling G.
versteegii (Angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05)
Gambar 31 menunjukkan bahwa media M1, M2 dan M3 menghasilkan
pertumbuhan jumlah akar primer plantling G. Versteegii lebih baik daripada dengan media
M0, namun demikian media M1, M2 dan M3 tidak berbeda satu dan lainnya terhadap
jumlah akar primer plantling G. Versteegii karena media tersebut relatif sama
mengandung bahan organik (kompos dan asam humat). Apabila dilihat berdasarkan
kombinasi FMA dan media tumbuh maka Gambar 32 menunjukkan bahwa kombinasi
media M0F1 mengahasilkan jumlah rata-rata akar primer tertinggi 7 akar, sedangkan
kombinasi perlakuan M2F2 menghasilkan jumlah rata-rata akar primer terpendek 0,33
akar. Kombinasi media M2F1 menghasilkan jumlah rata-rata akar sekunder tertinggi 16
akar, sedangkan kombinasi perlakuan M2F2 menghasilkan jumlah rata-rata akar sekunder
terendah 0.0. Visualisasi pengaruh kombinasi FMA dan media tumbuh terhadap jumlah
akar primer dan sekunder plantling G. versteegii disajikan pada Gambar 32.
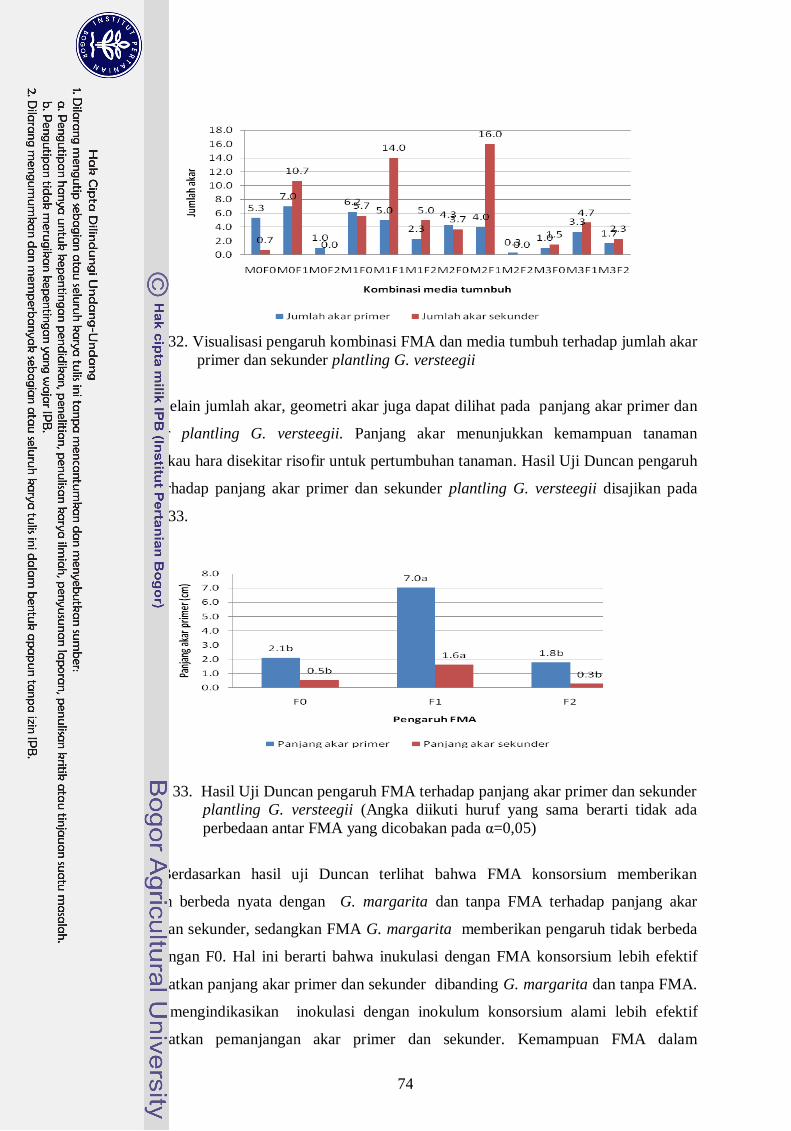
74
Gambar 32. Visualisasi pengaruh kombinasi FMA dan media tumbuh terhadap jumlah akar
primer dan sekunder plantling G. versteegii
Selain jumlah akar, geometri akar juga dapat dilihat pada panjang akar primer dan
sekunder plantling G. versteegii. Panjang akar menunjukkan kemampuan tanaman
menjangkau hara disekitar risofir untuk pertumbuhan tanaman. Hasil Uji Duncan pengaruh
FMA terhadap panjang akar primer dan sekunder plantling G. versteegii disajikan pada
Gambar 33.
Gambar 33. Hasil Uji Duncan pengaruh FMA terhadap panjang akar primer dan sekunder
plantling G. versteegii (Angka diikuti huruf yang sama berarti tidak ada
perbedaan antar FMA yang dicobakan pada α=0,05)
Berdasarkan hasil uji Duncan terlihat bahwa FMA konsorsium memberikan
pengaruh berbeda nyata dengan G. margarita dan tanpa FMA terhadap panjang akar
primer dan sekunder, sedangkan FMA G. margarita memberikan pengaruh tidak berbeda
nyata dengan F0. Hal ini berarti bahwa inukulasi dengan FMA konsorsium lebih efektif
meningkatkan panjang akar primer dan sekunder dibanding G. margarita dan tanpa FMA.
Hal ini mengindikasikan inokulasi dengan inokulum konsorsium alami lebih efektif
meningkatkan pemanjangan akar primer dan sekunder. Kemampuan FMA dalam
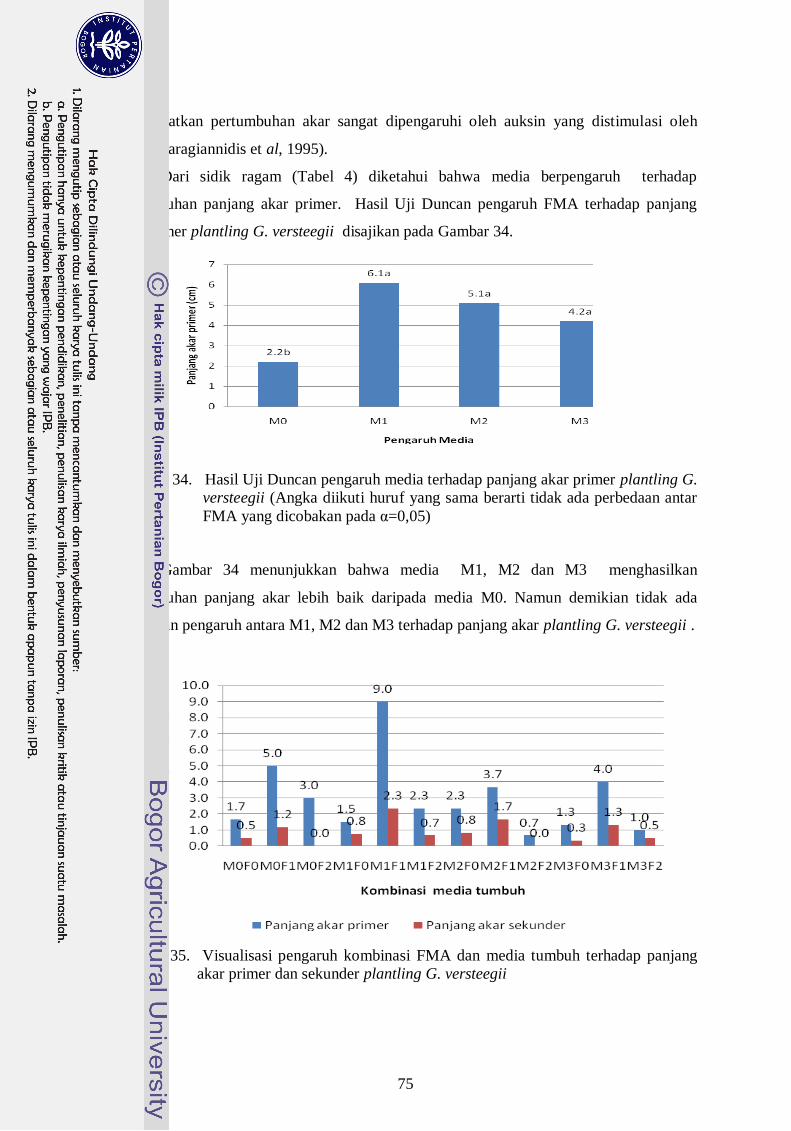
75
meningkatkan pertumbuhan akar sangat dipengaruhi oleh auksin yang distimulasi oleh
FMA (Karagiannidis et al, 1995).
Dari sidik ragam (Tabel 4) diketahui bahwa media berpengaruh terhadap
pertumbuhan panjang akar primer. Hasil Uji Duncan pengaruh FMA terhadap panjang
akar primer plantling G. versteegii disajikan pada Gambar 34.
Gambar 34. Hasil Uji Duncan pengaruh media terhadap panjang akar primer plantling G.
versteegii (Angka diikuti huruf yang sama berarti tidak ada perbedaan antar
FMA yang dicobakan pada α=0,05)
Gambar 34 menunjukkan bahwa media M1, M2 dan M3 menghasilkan
pertumbuhan panjang akar lebih baik daripada media M0. Namun demikian tidak ada
perbedaan pengaruh antara M1, M2 dan M3 terhadap panjang akar plantling G. versteegii .
Gambar 35. Visualisasi pengaruh kombinasi FMA dan media tumbuh terhadap panjang
akar primer dan sekunder plantling G. versteegii

76
Apabila diilihat dari kombinasi perlakuan maka panjang akar primer dan akar
sekunder terpanjang dihasilkan oleh kombinasi M2F1 akar primer terpanjang (9 cm),
sedangkan kombinasi perlakuan M2F2 memiliki akar primer terpendek (0 %). Akar
sekunder terpanjang dihasilkan oleh M1F1 (2,3 cm) dan akar sekunder terpendek
dihasilkan oleh kombinasi M0F2 dan M2F2 masing-masing 0 cm. Visualisasi panjang akar
primer dan sekunder disajikan pada Gambar 35. Secara umum FMA konsorsium
memberikan pengaruh berbeda nyata terhadap panjang akar primer nyata dengan tanpa
mikoriza. Berdasarkan hasil uji Duncan (Gambar 33) terlihat bahwa FMA konsorsium
memberikan pengaruh berbeda nyata dengan G. margarita dan tanpa FMA terhadap
perpanjangan akar primer dan sekunder, sedangkan FMA G. margarita memberikan
pengaruh tidak berbeda nyata dengan F0. Hal ini berarti bahwa inukulasi dengan FMA
konsorsium lebih efektif meningkatkan pemanjangan akar primer dan sekunder dibanding
G. margarita dan tanpa FMA. Keragaan panjang akar primer dan akar sekunder disajikan
pada Gamabr 36.
Gambar 36. Keragaan panjang akar primer (A) dan akar sekunder (B) plantling G.
versteegii
Berat Basah Pucuk dan Akar
Berat basah pucuk adalah berat segar pada saat tanaman uji dipanen caranya
dengan memisahkan antara pangkal batang dengan akar tanaman. Hasil sidik ragam
menunjukan bahwa FMA berpengaruh terhadap berat basah pucuk akar. Hasil Uji Duncan
terhadap berat basah akar dan pucuk plantling G. versteegii disajikan pada Gambar 37.
A B
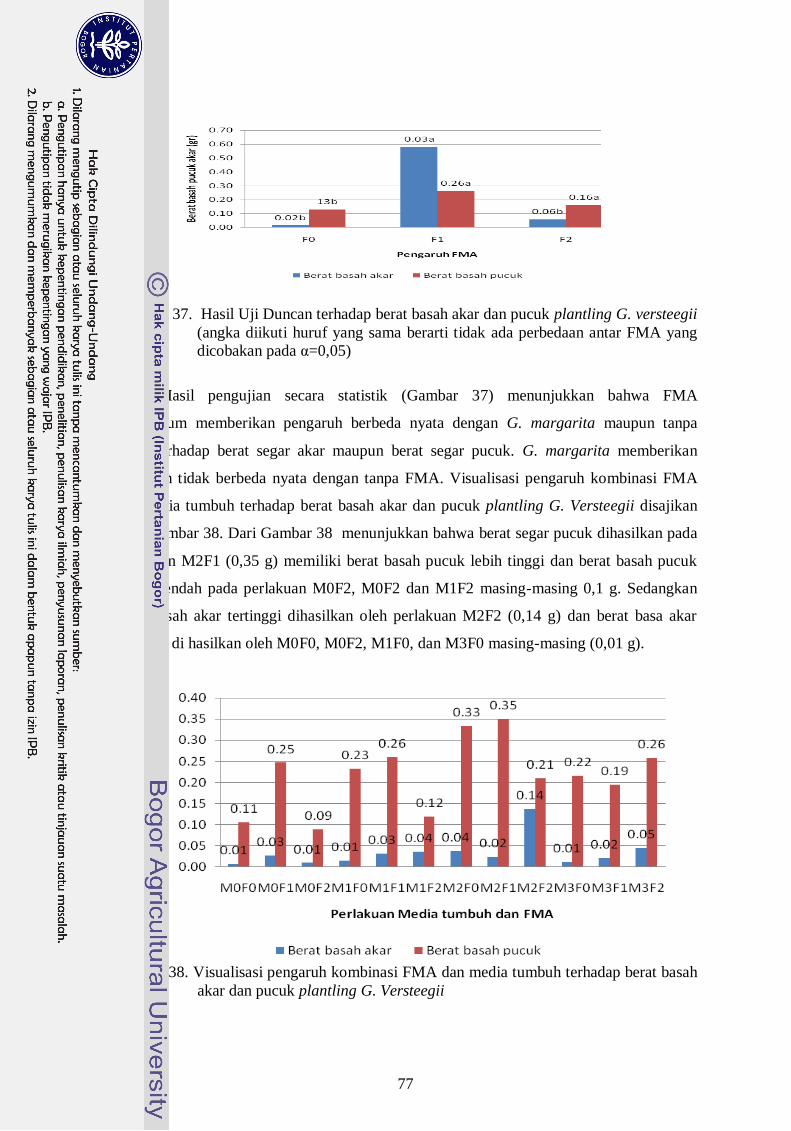
77
Gambar 37. Hasil Uji Duncan terhadap berat basah akar dan pucuk plantling G. versteegii
(angka diikuti huruf yang sama berarti tidak ada perbedaan antar FMA yang
dicobakan pada α=0,05)
Hasil pengujian secara statistik (Gambar 37) menunjukkan bahwa FMA
konsorsium memberikan pengaruh berbeda nyata dengan G. margarita maupun tanpa
FMA terhadap berat segar akar maupun berat segar pucuk. G. margarita memberikan
pengaruh tidak berbeda nyata dengan tanpa FMA. Visualisasi pengaruh kombinasi FMA
dan media tumbuh terhadap berat basah akar dan pucuk plantling G. Versteegii disajikan
pada Gambar 38. Dari Gambar 38 menunjukkan bahwa berat segar pucuk dihasilkan pada
perlakuan M2F1 (0,35 g) memiliki berat basah pucuk lebih tinggi dan berat basah pucuk
paling rendah pada perlakuan M0F2, M0F2 dan M1F2 masing-masing 0,1 g. Sedangkan
berat basah akar tertinggi dihasilkan oleh perlakuan M2F2 (0,14 g) dan berat basa akar
terendah di hasilkan oleh M0F0, M0F2, M1F0, dan M3F0 masing-masing (0,01 g).
Gambar 38. Visualisasi pengaruh kombinasi FMA dan media tumbuh terhadap berat basah
akar dan pucuk plantling G. Versteegii
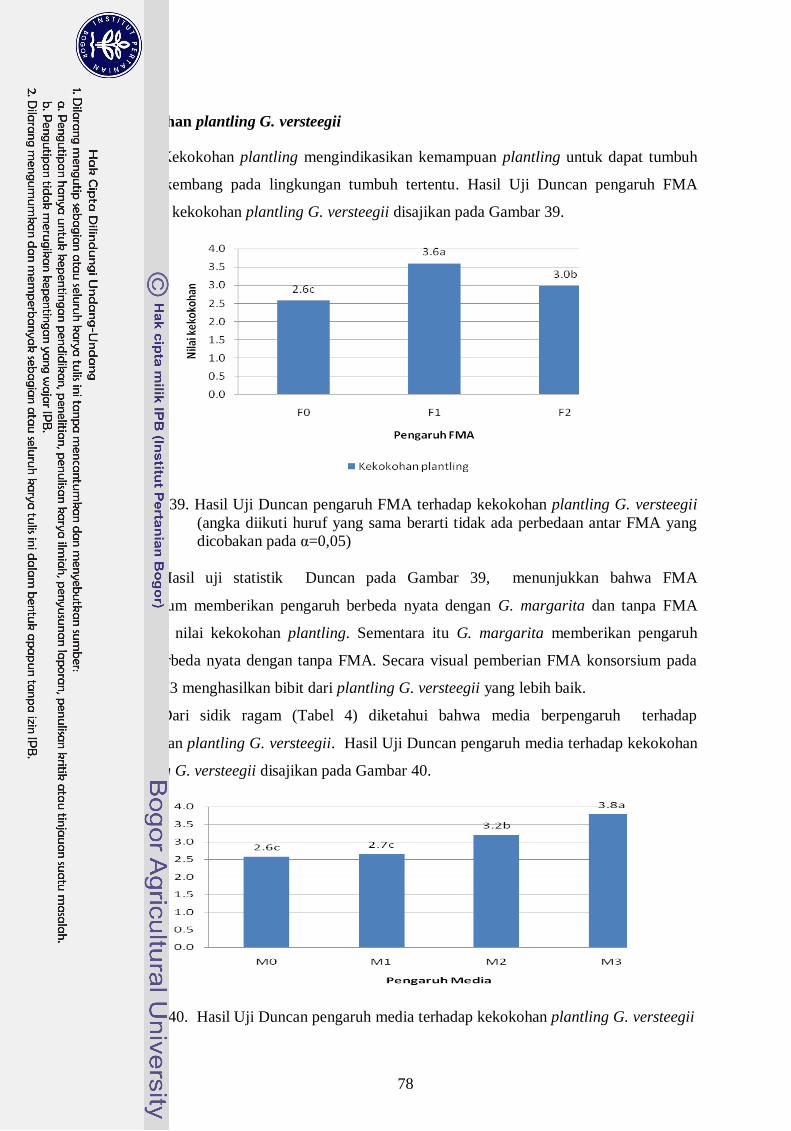
78
Kekokohan plantling G. versteegii
Kekokohan plantling mengindikasikan kemampuan plantling untuk dapat tumbuh
dan berkembang pada lingkungan tumbuh tertentu. Hasil Uji Duncan pengaruh FMA
terhadap kekokohan plantling G. versteegii disajikan pada Gambar 39.
Gambar 39. Hasil Uji Duncan pengaruh FMA terhadap kekokohan plantling G. versteegii
(angka diikuti huruf yang sama berarti tidak ada perbedaan antar FMA yang
dicobakan pada α=0,05)
Hasil uji statistik Duncan pada Gambar 39, menunjukkan bahwa FMA
konsorsium memberikan pengaruh berbeda nyata dengan G. margarita dan tanpa FMA
terhadap nilai kekokohan plantling. Sementara itu G. margarita memberikan pengaruh
tidak berbeda nyata dengan tanpa FMA. Secara visual pemberian FMA konsorsium pada
media M3 menghasilkan bibit dari plantling G. versteegii yang lebih baik.
Dari sidik ragam (Tabel 4) diketahui bahwa media berpengaruh terhadap
kekokohan plantling G. versteegii. Hasil Uji Duncan pengaruh media terhadap kekokohan
plantling G. versteegii disajikan pada Gambar 40.
Gambar 40. Hasil Uji Duncan pengaruh media terhadap kekokohan plantling G. versteegii
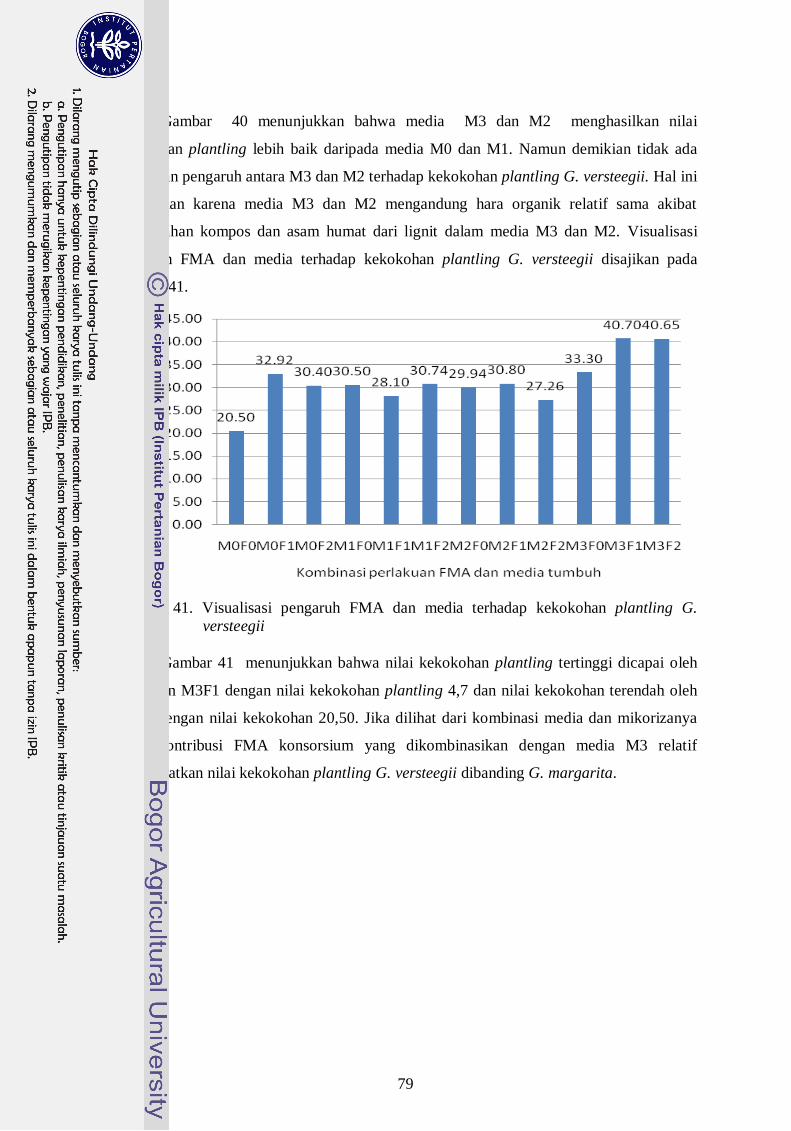
79
Gambar 40 menunjukkan bahwa media M3 dan M2 menghasilkan nilai
kekokohan plantling lebih baik daripada media M0 dan M1. Namun demikian tidak ada
perbedaan pengaruh antara M3 dan M2 terhadap kekokohan plantling G. versteegii. Hal ini
disebabkan karena media M3 dan M2 mengandung hara organik relatif sama akibat
penambahan kompos dan asam humat dari lignit dalam media M3 dan M2. Visualisasi
pengaruh FMA dan media terhadap kekokohan plantling G. versteegii disajikan pada
Gambar 41.
Gambar 41. Visualisasi pengaruh FMA dan media terhadap kekokohan plantling G.
versteegii
Gambar 41 menunjukkan bahwa nilai kekokohan plantling tertinggi dicapai oleh
perlakuan M3F1 dengan nilai kekokohan plantling 4,7 dan nilai kekokohan terendah oleh
M0F0 dengan nilai kekokohan 20,50. Jika dilihat dari kombinasi media dan mikorizanya
maka kontribusi FMA konsorsium yang dikombinasikan dengan media M3 relatif
meningkatkan nilai kekokohan plantling G. versteegii dibanding G. margarita.
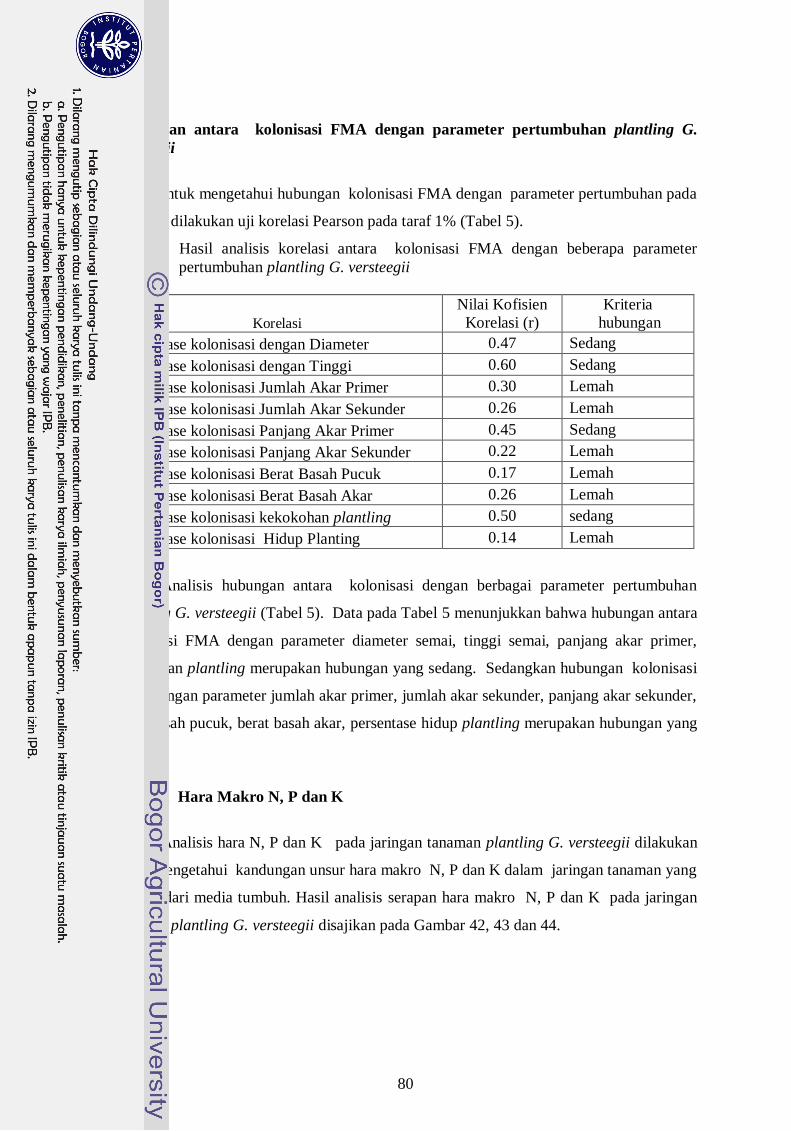
80
Hubungan antara kolonisasi FMA dengan parameter pertumbuhan plantling G.
versteegii
Untuk mengetahui hubungan kolonisasi FMA dengan parameter pertumbuhan pada
tanaman dilakukan uji korelasi Pearson pada taraf 1% (Tabel 5).
Tabel 5. Hasil analisis korelasi antara kolonisasi FMA dengan beberapa parameter
pertumbuhan plantling G. versteegii
Korelasi
Nilai Kofisien
Korelasi (r)
Kriteria
hubungan
Persentase kolonisasi dengan Diameter 0.47 Sedang
Persentase kolonisasi dengan Tinggi 0.60 Sedang
Persentase kolonisasi Jumlah Akar Primer 0.30 Lemah
Persentase kolonisasi Jumlah Akar Sekunder 0.26 Lemah
Persentase kolonisasi Panjang Akar Primer 0.45 Sedang
Persentase kolonisasi Panjang Akar Sekunder 0.22 Lemah
Persentase kolonisasi Berat Basah Pucuk 0.17 Lemah
Persentase kolonisasi Berat Basah Akar 0.26 Lemah
Persentase kolonisasi kekokohan plantling 0.50 sedang
Persentase kolonisasi Hidup Planting 0.14 Lemah
Analisis hubungan antara kolonisasi dengan berbagai parameter pertumbuhan
plantling G. versteegii (Tabel 5). Data pada Tabel 5 menunjukkan bahwa hubungan antara
kolonisasi FMA dengan parameter diameter semai, tinggi semai, panjang akar primer,
kekokohan plantling merupakan hubungan yang sedang. Sedangkan hubungan kolonisasi
FMA dengan parameter jumlah akar primer, jumlah akar sekunder, panjang akar sekunder,
berat basah pucuk, berat basah akar, persentase hidup plantling merupakan hubungan yang
lemah.
Serapan Hara Makro N, P dan K
Analisis hara N, P dan K pada jaringan tanaman plantling G. versteegii dilakukan
untuk mengetahui kandungan unsur hara makro N, P dan K dalam jaringan tanaman yang
diserap dari media tumbuh. Hasil analisis serapan hara makro N, P dan K pada jaringan
tanaman plantling G. versteegii disajikan pada Gambar 42, 43 dan 44.
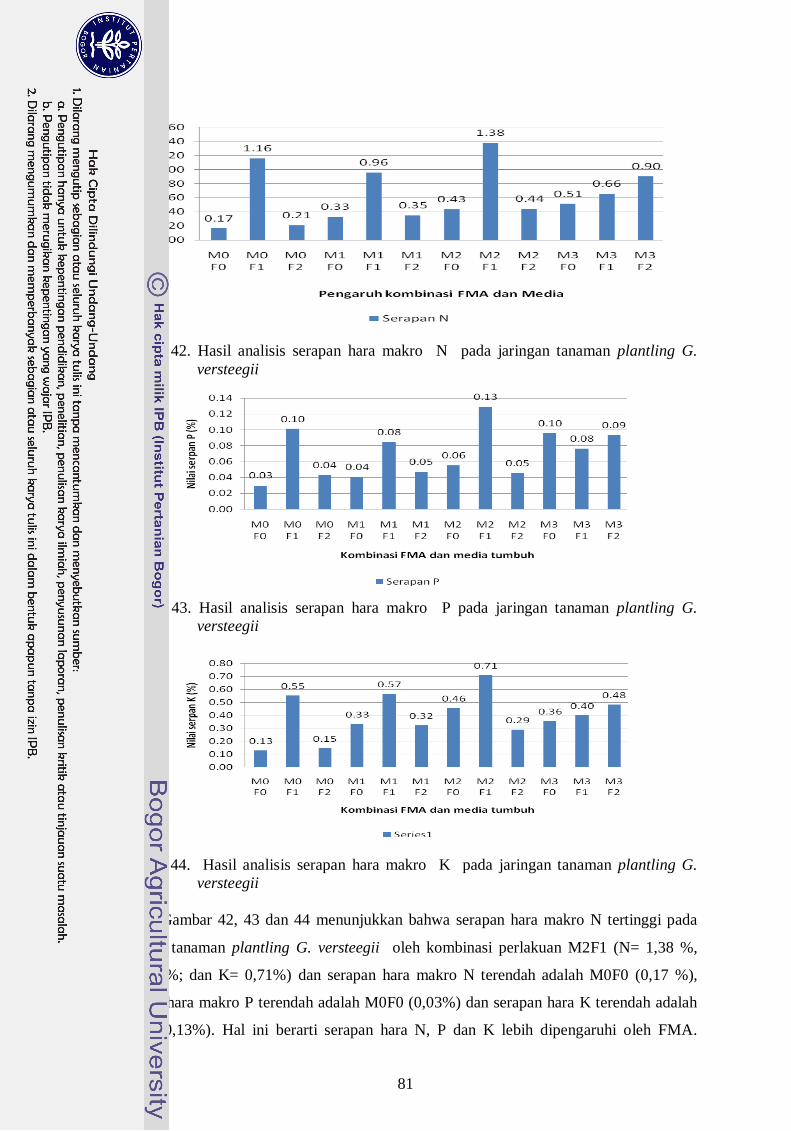
81
Gambar 42. Hasil analisis serapan hara makro N pada jaringan tanaman plantling G.
versteegii
Gambar 43. Hasil analisis serapan hara makro P pada jaringan tanaman plantling G.
versteegii
Gambar 44. Hasil analisis serapan hara makro K pada jaringan tanaman plantling G.
versteegii
Gambar 42, 43 dan 44 menunjukkan bahwa serapan hara makro N tertinggi pada
jaringan tanaman plantling G. versteegii oleh kombinasi perlakuan M2F1 (N= 1,38 %,
P=0,13 %; dan K= 0,71%) dan serapan hara makro N terendah adalah M0F0 (0,17 %),
serapan hara makro P terendah adalah M0F0 (0,03%) dan serapan hara K terendah adalah
M0F0 (0,13%). Hal ini berarti serapan hara N, P dan K lebih dipengaruhi oleh FMA.

82
Secara umum pemberian inokulasi FMA meningkatkan penyerapan hara makro terutama
N, P dan K pada jaringan tanaman plantling G. versteegii.
Indeks mutu bibit
Kualitas bibit tanaman dicirikan dengan perakaran dan pertumbuhan yang baik
sehingga apabila bibit-bibit tersebut dipindahkan dari persemaian ke lapangan dapat
tumbuh dan berkembang menjadi bibit yang mampu bertahan hidup pada kondisi lapang.
Kemampuan daya tahan hidup ini dapat diukur dengan indek mutu bibit (IMB). Indeks
mutu bibit dihitung menggunakan teknik skoring terhadap parameter tinggi, diameter dan
kekekohan plantling G. versteegii. Nilai hasil kualitas indeks mutu bibit plantling G.
versteegii menggunakan system skoring disajikan pada Lampiran 20, 21 dan 22. Dari
Lampiran 20 dan 21 menunjukkan bahwa berdasarkan nilai rata-rata tinggi dan diameter
maka indeks mutu bibit tertinggi adalah M2F1, M3F2 dan M3F1, sedangkan indeks mutu
bibit terendah adalah M0F0. Tingginya kualitas bibit plantling G. versteegii pada
kombinasi media M2F1, M3F2 dan M3F1 disebabkan karena adanya penyerapan hara
yang lebih baik dari FMA konsorsium maupun G. margarita serta tingginya kandungan
bahan organik pada media ini akibat diperkaya dengan bahan organik menggunakan
kompos dan batubara. Apabila dilihat berdasarkan nilai kekokohan bibit (Lampiran 22)
menunjukkan bahwa indeks mutu bibit tertinggi adalah M3F1 sedangkan indeks mutu bibit
terendah adalah M0F0, M0F2 dan M1F0. Secara visual tingginya kualitas bibit plantling
G. versteegii pada kombinasi media M3F1 disebabkan karena adanya pertambahan
pertumbuhan tinggi dan diameter plantling G. versteegiii oleh FMA konsorsium pada
media yang diperkaya kompos, batubara muda dan arang sekam.

83
PEMBAHASAN
Pengaruh FMA
Secara statistik aplikasi FMA memberikan pengaruh nyata terhadap semua
parameter yang diamati (Tabel 4). FMA konsorsium memberikan kontribusi relatif lebih
baik dari FMA G. margarita maupun tanpa pemberian FMA (kontrol). Besarnya kontribusi
FMA konsorsium memberikan pengaruh nyata terhadap berbagai parameter yang diamati
disebabkan karena secara alami FMA konsorsium telah membangun simbiosis dengan
tanaman inang G. versteegii. Dalam penelitian ini keeratan hubungan antara FMA
konsorsium dengan inang G. versteegii dapat dilihat dari besarnya nilai persentase
kolonisasi FMA, PGR dan DPU. Indikator lain yang dapat dipakai untuk untuk melihat
hubungan simbiosis ini adalah nilai serapan hara P, dimana FMA konsorsium
meningkatkan serapan hara P pada jaringan tanaman plantling G. versteegii lebih baik dari
G. margarita.
Pertumbuhan primer yang terjadi disebabkan aktifitas meristem pada ujung batang
tanaman. Aktifitas meristem yang selalu melakukan pembelahan sangat dipengaruhi oleh
fosfor. Fosfor merupakan bagian esensial dari banyak gula fosfat yang berperan dalam
nukleotida, seperti RNA dan DNA, serta bagian dari fosfolipid pada membran. Fosfor
berperan penting pula dalam metabolisme energi, karena keberadaannya dalam ATP, ADP,
AMP dan pirofosfat (Ppi) (Salisbury dan Ross 1995). Dengan demikian apabila kadar P
meningkat maka aktifitas meristem pada ujung batang pun meningkat sehingga batang
bertambah tinggi.
Keuntungan FMA pada tanaman akan meningkatkan penyerapan P karena
keberadaan hifa eksternalnya yang menyebar luas ke dalam tanah. Selain P, keberadaan
FMA juga dapat membantu meningkatkan penyerapan kalsium (Widiastuti dan Tahardi
1993) yang berperan dalam pembelahan dan pemanjangan sel (Gardner et al.1991).
Pertambahan tinggi tanaman juga dipengaruhi oleh aktifitas beberapa hormon
seperti giberalin dan sitokinin (Goldsworthy dan Fisher 1992). Menurut Anas dan Santosa
(1993), FMA pada akar tanaman dapat menghasilkan sejumlah hormon pengatur tumbuh
seperti giberalin dan sitokinin sehingga keberadaan FMA pada akar tanaman dapat
berperan dalam proses pemanjangan batang. Klein (2000) juga melaporkan bahwa banyak
mikroba di dalam rizosfer yang dapat menghasilkan campuran organik komplek seperti
giberalin yang dapat mengubah morfologi dan fisiologi tumbuhan. Dengan demikian
keberadaan FMA dapat meningkatkan rata-rata pertambahan tinggi tanaman secara lebih
baik dari pada kontrol (tanpa perlakuan).

84
Pertambahan diameter sangat dipengaruhi oleh fotosintat sebagai penyusun
kerangka karbon untuk dan suplai H2O untuk memperlancar semua proses fisiologis dan
metabolisme di dalam jaringan tanaman (Kramer dan Kozlowski 1960). Keberadaan FMA
membantu meningkatkan suplai H2O dengan bantuan hifa eksternal yang dapat masuk
kedalam rongga tanah yang diameternya lebih kecil daripada diameter akar (Fakuara
1994). Pertambahan diameter merupakan pertumbuhan sekunder yang juga dipengaruhi
oleh nitrogen untuk pembentukan sel-sel baru (Goldsworthy dan Fisher 1992). Dengan
adanya FMA maka kadar N pun meningkat dan akhirnya berperan dalam pertambahan
diameter. Serapan N lebih besar pada tanaman bermikoriza dari pada tanaman tanpa
mikoriza. Hal ini terbukti dari hasil analisis jaringan yang dilakukan pada plantling G.
versteegii.
Kolonisasi FMA diakar juga mempengaruhi geometri akar yang disebabkan oleh
peran auksin yang dihasilkan oleh FMA. Geometri akar divisualisasi berdasarkan jumlah
akar primer dan sekunder serta panjang akar primer dan sekunder. Aplikasi FMA
memberikan pengaruh terhadap panjang akar primer dan sekunder dan jumlah akar primer
dann sekunder plantling G. versteegii dibandingkan tanpa aplikasi FMA. Namun FMA
konsorsium lebih efektif meningkatkan pertumbuhan akar primer dan sekunder termasuk
jumlah akar primer dan jumlah akar sekunder plantling G. versteegii.
Garcia-Garrido dan Ocampo (2002) menyatakan bahwa interaksi FMA dan
tanaman inang melibatkan pengenalan sinyal molekul spesifik agar tercipta hubungan
simbiosis mutualisme. Keberadaan FMA dapat berpengaruh terhadap pengenalan akar
tanaman inang serta berpengaruh pula terhadap daya penerimaan akar melalui signal
eksudat yang dikeluarkan oleh plantling G. versteegii untuk membangun simbiosis.
Perbedaan kemampuan FMA dalam meningkatkan pertumbuhan akar sangat dipenagruhi
oleh auksin yang dihasilkan oleh FMA.
Secara statistik FMA konsorsium lebih efektif menginduksi terbentuknya akar
primer dan akar sekunder pada media tanah dan pasir (kontrol). Diduga hal ini disebabkan
karena media tersebut miskin akan hara sehingga plantling G. versteegii pada media ini
lebih aktif memproduksi eksudat sebagai signal untuk membangun simbiosis dengan FMA.
Dengan adanya simbiosis yang terbentuk maka FMA membantu inang plantling G.
versteegii menginduksi akar dengan cara mengeluarkan auksin IAA yang mempengaruhi
pembelahan dan pemanjangan sel akar tanaman pada daerah merismatik.
Berdasarkan hasil sidik ragam pada taraf 5% menunjukkan bahwa berat tajuk akar
dipengaruhi oleh inokulasi FMA konsorsium. Penyerapan H2O lebih optimal dilakukan

85
oleh FMA yang memiliki hifa yang panjang. Dari hasil penelitian terlihat bahwa tanaman
yang mendapat perlakuan FMA mempunyai nilai rata-rata berat segar yang lebih tinggi
bila dibandingkan dengan perlakuan tanpa FMA (kontrol).
Nitrogen sangat dibutuhkan oleh tanaman, sebagai penyusun asam amino, protein
dan komponen lainnya. Nitrogen juga sangat penting dalam respirasi, meningkatkan reaksi
enzimatik, dan meningkatkan metabolisme sel. Nitrogen diserap akar dalam bentuk
amonium atau ion nitrat. Nitrogen yang dapat terikat oleh tanaman akan selalu dibutuhkan,
sedangkan mengenai jumlahnya tergantung pada kebutuhan tanaman itu sendiri.
Berdasarkan hasil sidik ragam pada taraf 5% untuk parameter serapan hara N, serapan hara
P inokulasi FMA konsorsium menunjukkan pengaruh nyata pada jaringan plantling G.
versteegii. Jumlah unsur hara yang dapat diserap tanaman inang merupakan jumlah unsur
hara yang terlarut dan tersedia dalam larutan tanah. FMA mampu memanfaatkan unsur
hara yang relatif tidak tersedia untuk tanaman dan kemudian mengalirkannya ke tanaman
inang (Bornner dan Galston 1952).
Inokulasi FMA meningkatkan pertumbuhan tanaman dan serapan hara seperti N, P
dan K (Johansson et al. 2004). Setiap jenis FMA mempunyai kemampuan yang berbeda
dalam meningkatkan serapan P. Hal tersebut dapat terjadi karena adanya perbedaan
gerakan P dalam hifa sehingga menyebabkan terjadinya perbedaan efisiensi antara jenis
FMA dalam meningkatkan pertumbuhan tanaman (Anas dan Santosa 1993). Keberhasilan
semai dicirikan memiliki perakaran dan pertumbuhan yang baik, yang apabila bibit-bibit
tersebut dipindahkan dari persemaian ke lapangan dapat tumbuh dan berkembang menjadi
bibit, yang mampu bertahan hidup pada kondisi lapang. Kemampuan daya tahan hidup ini
dapat diukur dengan indek mutu bibit (IMB).
Pada plantling G. versteegii inokulan FMA konsorsium dan G. Margarita dapat
meningkatkan nilai indeks mutu bibit plantling G. versteegii pada parameter tinggi,
diameter dan kekokohan bibit dibandingkan dengan tanpa FMA pada kombinasi media
yang diberikan kompos, arang sekam dan batubara muda. Dengan demikian pemanfaatan
FMA dan kombinasi media tanah, pasir, kompos, arang sekam dan batubara muda pada
plantling G. versteegii akan menghasilkan bibit yang lebih siap dipindahkan ke lapangan
akan mampu bertahan hidup pada kondisi lapang .
Kolonisasi berhubungan erat dengan jenis FMA dan jenis tanaman inang. Dengan
demikian, translokasi nutrisi dan pemanfaatan karbon oleh tanaman inang pun berbeda-
beda. Kolonisasi mikoriza di dalam akar dan panjang miselium di dalam tanah merupakan
parameter yang menentukan peningkatan serapan fosfor. Dari hasil penelitian diketahui

86
bahwa FMA dapat berasosiasi dengan plantling G. versteegii simbiosis dimulai pada saat
propagul FMA merespon keberadaan akar di dalam tanah. Penetrasi terjadi pada saat hifa
FMA membentuk apresorium yang berlekatan dengan sel epidermis akar. Hifa kemudian
menyebar didalam ruang antar sel dan masuk ke dalam sel (Brundrett et al. 1996).
Hasil sidik ragam pada taraf 1% menunjukkan bahwa inokulasi FMA konsorsium
dan G. margarita berpengaruh nyata terhadap kolonisai FMA pada plantling G. versteegii.
Hal ini menunjukkan bahwa kedua FMA memiliki kemampuan dalam mengkolonisasi akar
tanaman inang. Menurut Johansson et al. (2004) fenomena kecocokan interaksi antara
tanaman inang dengan FMA terdapat pada tingkat biokimia dan genetik. Dengan demikian
interaksi antara FMA dan inang plantling G. versteegii yang cocok akan menghasilkan
pertumbuhan tanaman inang secara lebih baik.
Kolonisasi FMA pada akar tanaman dapat diamati dari keberadaan beberapa
struktur FMA, seperti hifa internal, arbuskula, vesikula dan spora. Menurut Smith dan
Read (1997), bahwa hifa internal berfungsi sebagai alat translokasi unsur hara, eksternal
berfungsi menyerap unsur hara dan air, vesikula berfungsi sebagai tempat cadangan
makanan terutama lipid, sedangkan arbuskula merupakan struktur infeksi yang sangat
penting dalam simbiosis FMA, karena arbuskula berfungsi dalam proses transfer unsur
hara antara kedua simbion (fungi dengan akar tanaman).
Perhitungan kolonisasi FMA di dalam akar hanya menentukan ada atau tidaknya
FMA yang mengkoloni akar dan melewatkan hifa-hifa eksternal yang tersebar di dalam
tanah (Hart dan Reader 2002). Besar kecilnya kolonisasi FMA di dalam akar hanya
mencerminkan sebagian kemampuannya dalam membantu penyerapan nutrisi bagi
tanaman (Hart dan Reader 2002). Hifa internal FMA di dalam akar berperan dalam proses
transfer nutrisi dari hifa ekternal ke dalam sel - sel akar tanaman inang. Namun demikian
pertumbuhan hifa dan akar inang sangat dipengaruhi oleh media tumbuh yang digunakan.
Pengaruh Media
Penggunaan media berupa tanah, kompos, bio-charcoal dan lignit (batubara muda)
digunakan sebagai upaya mendapatkan kombinasi media yang cocok untuk produksi
bibit gaharu yang diperbanyak menggukan teknik kultur jaringan. Penambahan kompos
menguntungkan karena mengandung bahan organik terdekomposisi sehingga mudah
diserap oleh tanaman, disamping itu dengan penambahan kompos akan meningkatkan
porositas dan menjadikan media tidak kompak sehingga memudahkan perkembangan akar
plantling.

87
Dalam penelitian ini penambahan bio-charcoal diharapkan dapat fungsi sebagai
soil conditioner dan soil managment, meningkatkan porositas tanah dan aktifitas mikroba
tanah, menyerap kontaminan toksis, menetralkan keasaman tanah serta mencegah penyakit
busuk akar yang disebabkan oleh serangan patogen jamur (Steiner, 2007 ; Lehman, 2007 ;
Supriyanto, 2010), memberikan efek gelap di sekitar akar plantling sehingga auksin IAA
di akar tidak rusak oleh cahaya dan IAA akan berperan optimal mendukung
perkembangan akar adventif plantling. Peran lain yang diberikan oleh bio-charcoal adalah
berperan dalam menyerap zat-zat toksis dalam media tumbuh dan memberikan efek gelap
disekitar akar plantling sehingga auksin IAA di akar tidak rusak oleh cahaya dan IAA
akan berperan optimal mendukung perkembangan akar adventif plantling. Penggunaan
lignit (batubara muda) dalam media dimanfaatkan sebagai pembenah media karena
batubara muda mengandung asam humat sehingga dapat berperan menyediakan nutrisi
hara secara ototropik bagi plantling di media pertumbuhan.
Secara statistik media memberikan pengaruh tidak berbeda nyata terhadap
parameter lainnya namun pada parameter persentase kolonisasi media tumbuh
berpengaruh nyata terhadap parameter. Meskipun media memberikan pengaruh tidak
nyata terhadap beberapa parameter yang diuji secara statistik menggunakan uji Duncan,
namun bila dilihat berdasarkan nilai rata-rata maka media dengan penambahan kompos,
arang sekam dan batubara muda ternyata memberikan keragaan plantling yang baik.
Kombinasi media kompos, arang sekam dan batubara muda memberikan nilai kokokohan
bibit lebih baik untuk dapat dipindahkan ke lapangan dibandingkan dengan media
kontrol dari tanah dan pasir.
Pengaruh Interaksi media tumbuh dengan FMA
Hasil sidik ragam (Tabel 4) menunjukkan bahwa tidak ada interaksi antara media
tumbuh dan FMA yang diberikan terhadap parameter diamati, namun jika dilihat dari nilai
indeks mutu bibit dan kekokohan bibit maka kombinasi media tanah, pasir, kompos bio-
charcoal dan lignit menghasilkan bibit tanaman dari plantling G. versteegii yang relatif
lebih siap untuk disapih ataupun ditanam di lapangan. Penambahan kompos dan lignit
berati menambahkan unsur hara organik dalam media tumbuh untuk meningkatkan
ketersediaanya dalam media dengan harapan tersedia cukup hara yang dapat diserap oleh
akar plantling.
Inokulasi FMA diharapkan memicu terbentuknya akar serta membantu akar dalam
menyerap hara dari media tubuh. Sementara itu dengan adanya kompos dan lignit maka

88
akan menambah hara organik yang dapat diserap akar plantling yang dibantu oleh adanya
FMA. Penambahan bio-charcoal diharapkan memberikan efek gelap sehingga auksin pada
akar plantling dapat berfungsi optimal dalam ramifikasi akar. Selain itu dengan adanya
bio-charcoal maka media tumbuh akan menjadi lebih poros sehingga aerasi dalam media
akan lebih baik untuk perkembangan perakaran plantling. Adanya pori-pori pada bio-
biocharcoal menjadi tempat berkembang dan berlindung organisme tanah. Bio-charcoal
juga meningkatkan kelembaban di sekitar akar plantling sehingga mendukung kehidupan
organisma tanah dalam hal ini adalah FMA.
Analisis statistik yang dipakai menggunaka pada taraf α0.05 ternyata tidak dapat
mendeteksi pengaruh interaksi antara FMA dan media tumbuh, namun demikian apabila
taraf α dinaikan menjadi 0.1. ternyata dapat mendeksi pengaruh interaksi antar FMA dan
media. Diduga interaksi ini tidak nyata disebabkan karena waktu pengamatan yang relatif
singkat. Hal ini berarti bahwa jika umur pertumbuhan plantling G. versteegii pada media
dilanjutkan diatas waktu 3 bulan, maka interakasi antara FMA dan media tumbuh akan
memberikan pengaruh nyata pada parameter yang diamati.

89
SIMPULAN DAN SARAN
Simpulan
1. Secara alami FMA telah membangun simbiosis dengan gaharu alam yang dibuktikan
dengan adanya persentase kolonisasi yang terbangun dengan kategori tinggi
2. Dalam penelitian ini ditemukan 7 spesies FMA dari 5 plot pengambilan sampel di
hutan alam Asai yang bersimbiosis dengan G. versteegii, yaitu G. mossae, G.
fasciculatum, G. aggregatum, Glomus sp1, Glomus sp2, Glomus sp3. dan Acaulospora
sp.1
3. Inokulum FMA konsorsium asal Hutan Alam Asai secara signifikan meningkatkan
pertumbuhan plantling G. versteegii lebih baik dari pada FMA Gigasspora margarita
dan plantling yang tidak diinokulasi mikoriza. Hal ini berarti kompatibilitas FMA
alami lebih tinggi daripada FMA G. Margarita.
4. Berdasarkan nilai kekokohan dan indeks mutu benih maka media tanah dengan
perbandingan 1 : 1 (v/v) yang dicampur dengan, kompos, bio-charcoal dan batu bara
muda masing-masing 0,5:0,4:0,2 (v/v) menghasilkan bibit G. versteegii lebih baik
dibandingkan kombinasi media lainnya.
Saran
Dalam upaya meningkatkan kualitas bibit gaharu asal kultur perlu disesuaikan
kondisi plantlet yang tepat saat aplikasi FMA. Penggunaan inokulum konsorsium alami
yang dikombinasikan dengan tanah, pasir, kompos, bio-charcoal dan lignit lebih
menjanjikan untuk pertumbuhan suatu jenis yang diperbanyak secara in-vitro.

90
DAFTAR PUSTAKA
Abbott L.K. & Robson A.D, Jasper D. A, and Gezey C., 1992. What is the role of VA
mycorrhizal Soil? Hal : 37 – 41.
Agrios GN. 1988. Plant Pathology. Third edition. San diego, California: Academic Press,
Inc.
Ahmad A. G, 2006. Pengaruh Arang Kayu dan Cendawan Endomikoriza terhadap Proses
Biokimia karbon Pada Semai Manglit (Michellia Montana Blue). Tesis Pascasarjana
IPB.
Anas I, Santosa DA. 1993. Mikoriza Vesikular-Arbuskular. Di dalam: Anas I. Pupuk
Hayati (Biofertilizer). Bogor: Laboratorium Biologi Tanah, Jurusan Tanah Fakultas
Pertanian Institut Pertanian Bogor.
ASGARIN (Asosiasi Pengusaha Ekspotir Gaharu Indonesia). 2001. Masalah/Kendala
Pengusahaan Kayu Gaharu. Direktorat Bina Usaha Perhutanan Rakyat. Ditjen RLPS
Departemen Kehutanan, Jakarta
Asmarahman C, 2008. Pemanfaatan Mikoriza dan Rhizobium Untuk Meningkatkan
Pertumbuhan Semai Kayu Energi Pada Media Tanah Bekas Tambang Semen. (Tesis).
Sekolah Pascasarjana IPB. Bogor.
Baghel RK, Sharma R, Pandey AK. 2009. Actifity of Phosphatase in the ectomycorrhizal
fungus Cantarellus tropicialis under controlled conditions. Journal of Tropical Forest
Science, 21 : 218-222.
Barden A, Awang N. A, Mulliken T, and Song M, 2000. Hearth of the Matter: Agarwood
Use and Trade and Cites Implementation for Aquilaria malaccensis. A Traffic
Network Report. Traffic Internasional. Cambridge, UK.
Barea JM, Azcon-Aguilar C. 1992. Interactions between Mycorrhizal Fungi and Other
Rhizosphere Microorganisms. In: Allen, M.F. (Ed). Micorrhizal Functioning. An
Integrative Plant-Funggal Process. New York: Chamman & Hall, Inc.
Bickelhaupt DH. 1980. Nursery Soil and Seedling Analysis Methodology. Proc. North
American Forest Tree Nursery Soil. Workshop. New york: July 28 August 1980: 237-
260.
Boruah P. and Singh R.S., 2000. Unraveling the secret of agar formation in agarwood. Di
dalam : Anonim, editor. Souvenir on a Seminar on Agar Scope and Dimension of Agar
(Aquilaria spp). Plantation in The NE Region ; Guwahati, India 22-23 November
2000. Assam : AATMA. Hal 1-2.
Boyce JS. 1948. Forest Pathology. Second Edition. New York: McGrraw - Hill Book
Company, Inc.
Brundrett M, Bougher N, Dell B, Grove T, Malajczuk N. 1996. Working with Mycorrhizas
in Forestry and Agriculture. 1996. Australian Centre for International Agricultural
Research (ACIAR). Diambil dari
http://www.ffp.csiro.au/research/mycorrhiza/index.html.[15 Desember 2006].
Brundrett, M. 2004. Diversity and classification of mycorrhizal associations.
Biol.Rev.2004.79.pp.473-495.©Cambridge Philosophical Society. (Edisi Revisi 20
Juni 2003).
CITES, 2004. Convention on International Trade in endangered Species of Wild Fauna
and flora : Amandements to Appendices I and II of CITES.
http://www.cites.org/common/cop/13/raw/ props/ID-Aguilaria-Gyrinops.pdf.
Delvian, Setiadi Y, Soedarmadi. 2001. Korelasi antara Salinitas Tanah dan Distribusi,
Populasi dan Dinamika Musiman Fungi Mikoriza Arbuskula di Hutan Pantai. Makalah
disampaikan pada Seminar Nasional dan Workshop Pemanfataan Mikoriza pada
Pertanian dan Rehabilitasi Lahan. Bengkulu 11- 13 Juni 2001. Bengkulu. Universitas
Bengkulu.

91
Fakuara Y. 1994. Peranan Mikoriza dalam Peredaran Hara dan Peningkatan Kualitas
Bibit. Di dalam: Laporan Program Penelitian Biologi dan Bioteknologi mikoriza.
Bogor: SEMEO BIOTROP. hlm 174-179.
Garcia-Garrido JM, Ocampo JA. 2002. Regulation of the Plant Defence Response in
Arbuscular Mycorrhizal Symbiosis. Journal of Experimental Botany 53 (373): 1377-
1386.
Gardeman J. W. And T. H. Nicholson, 1963. Spores of Mycorrhizal Endogene Spcies
Extraced from Soil by West Sieving and Decanting. The British Mycological Society
46 : 235-224.
Gardner FP, Pearce RB, Mitchell RL. 1991. Fisiologi Tanaman Budidaya. Jakarta: UI
Press.
Goodman R. N, Kiraly Z. Wood K. R., 1986. The Biochemistry and Physiology of Plant
Disease. Colombia: University of Missouri. Press.
Gunn and P. Stevens; M. Singadan; l. Sunnari and P. Chatterton, 2003. Eaglewood in
Papua New Guinea Tropical Rein Forest Project Vietnam. Working Paper No. 51.
Hetrick BAD, Wilson GWT.1993. Mycorrhizal Dependences of Modern Wheat Cultivar
and Ancestors; a Synthesis Can.J. Bot. 71: 512-518.
Hou D., 1960. Thymelaeaceae. Dalam Van Steenis, C. G. G. J (ed.). Flora Melasiana.
Gronigen: Walters-Noordhoff Publishing (1): 1-48.
Ishihara M, T. Tsuneya, M. Shiga and K. Uneyama, 1991. Three Sesquiterpenes from
Agarwood. Phytochemistry 30 (2) : 563-566.
Isnaini, 2003. Respon Tunas Gaharu (Aquilaria cressna dan A. filaria) terhadap Inokulasi
Acremonium pada Tiga Konsentrasi Medium Kultur. Makalah disampikan pada
Kongres XVII Seminar Ilmiah Perhimpunan Fitopatologi Indonesia. Bandung 6 – 8
Agustus 2003.
Johansson JF, Paul LR and Finlay RD. 2004. Microbial Interactions in the
Mycorrhizosphere and Their Significance For Sustainable Agriculture. FEMS
Microbial Ecology. hlm 1-10.
Karagiannidis N, Nikolaou N and Mattheou A., 1995. Influence of vesicular-arbuscular
mycorrhizal species on the growth and nutrient uptake of 3 grappevine rootstocks and
one table grape cultivar. Vitis. 34 (2) : 85-89.
Karyaningsih E., 2009. Pembenah Tanah dan Fungi Mikorhiza Arbuskula (FMA) untuk
Peningkatan Kualitas Bibit Tanaman Kehutanan pada Areal Bekas Tambang Batubara.
[Tesis Pascasarjana IPB].
Klein DA. 2000. The Rhizosphere. In: Lederberg J. (ed) Encyclopedia of Microbiology.
Second Edition. Vol. IV. USA: Academic Press. hlm 177.
Kramer PJ, Kozlowski TT. 1960. Physiology of Tree. USA: McGraww Hill. Book
Company.
Kunoh H, 1990. Ultrastucture and Mobilization of ions near infection sites. In Ann. Rev.
Phythopathol 28:93-111.
Leake JR, Jhonson D, Donnelly DP, Muckle GE, Boddy L, Read DJ. 2004. Networks of
power and influence : the role of mycorrhizal mycelium in controlling plant
communities and agroecosystem functioning. Can J Bot. 82: 1016-1045.
Lehman, J. 2007. A handful of carbon, Nature Vol. 447.
Linderman RG, Pfleger EL. 1994. Mycorrhizae and Plant Health. St Paul: APS Press. The
American Phytopathologycal Society.
Liu R. J, 1995. Effect of vesicular-arbuscular mycorrhizal fungi on Verticullium wilt of
cotton. Mycorrhiza 5 : 293-297.
Mashur, 2011. Indonesia Ekspor Gaharu ke China. Ekonomi Suara Karya, 15 Maret 2011.
http://www.bataviase.co.id/node/602572. [20 April 2011]

92
Parman T, Mulyaningsih dan M. Y. A Rahman, 1996. Studi Ekologi Gubal Gaharu pada
Pohon Ketimunan (A. Filaria). Temu Pakar Gaharu. Kanwil Dephut. Propinsi NTB,
11-12 April 1996.
Salisbury FB, Ross CW. 1995. Fisiologi Tumbuhan. Jilid I. Bandung: Institut Teknologi
Bandung.
Samingan T. 1983. Dendrologi. Jakarta: PT Gramedia. 90 hal.
Senn, T.L., and A.R. Kingman, 1973. A Review of Humus and Humic Acids. Research
Series Report No. 145. South Carolina Agricultural Experiment Station, Clemson.
Setiadi Y. 1990. Cross Inoculant Studies and Screening Rhizobial Isolates for Acacia
mangium Willd. IUC-BIOTEK-IPB.
Setiadi Y. 1992. Petunjuk Laboratorium Mikrobiologi Tanah Hutan. Bogor: Pusat Antar
Universitsas. Institut Pertanian Bogor.
Sieverding, E. 1991. Vesicular Arbuscular Mycorrihzal Management in Tropical
Agrosystem. Technical Cooperation Federal Republic of Germany. Eschborn. p. 37-76.
Siregar C. A, 2010. Pemanfaatan Arang Sekam Sebagai Soil Conditioner untuk
Memperbaikai Kualitas Tanah. Makalah Pelatihan Teknologi Bio-Charcoal Untuk
Budidaya Tanaman Kehutanan dan Pertanian Yang Berkelanjutan. SEAMEO
BIOTROP, 29 November 2010.
Smith, S.E., Read, D.J. 1997. Mycorrhizal Symbiosis. Second Eds. San Diego California.
Academic Press. Harcourt Brace and Company Publ.. p.34-160.
Steiner C. 2007. Charcoal as soil amandement : carbon negative energy and soil
restoration, UNFCC, Bali.
Stevonson, F.J. 1994. Humus Chemictry, Genesic, Composition, Reactions. A Wiley-
Interscience & Sons. New York. 496 pp.
Suhartono T. R, dan Newton, 2001. Status and Disribution of Aquilaria spp in Indonesia
and the Sustainebility of the Gaharu [Phd], Edinburg: Institut of Ecology and resource
Management.
Sumarna, Y. 2007. Budidaya Gaharu. Seri Agribusines. Penebar Swadaya. Jakarta.
Sumiasri N, Harmastini. 1990. Prospek Sengon (Paraserianthes falcataria(L) Fosberg) di
Jawa Barat. Bogor. Jurnal Penelitian dan Pengembangan Kehutanan. Departemen
Kehutanan. hlm 7–9.
Supriyanto, 2009. Modifikasi Teknik Pewarnaan Akar Terkolonisasi Cendawan
Endomikorhiza. Makalah Pelatihan Dasar Isolasi dan Inokulasi Mikoriza untuk
Tanaman Pertanian dan Kehutanan. SEAMEO-BIOTROP, 24-25 Juni 2009.
Supriyanto, 2010. Pemanfatan Bio-Charcoal untuk Kultur Endomikoriza. Makalah
Pelatihan Teknologi Bio-Charcoal Untuk Budidaya Tanaman Kehutanan dan Pertanian
Yang Berkelanjutan. SEAMEO BIOTROP, 29 November 2010.
Sutedjo MM, Kartasapoetra AG, Sastroatmodjo RDS. 1996. Mikrobiologi Tanah. Jakarta.
PT. Rineka Cipta.
Sylvia, D.M. 2006. Overview of mycorrhizal symbiosis : Based on a chapter in Prinsples
and Application of Soil Microbiology. http://Cropsoil.psu.edu/sylvia/mycorrhiza.htm.
Tawaraya K, Tokarin K, Wagatsuma T. 2001. Dependence of Allium fistulosum Cultivar
on the Arbuscular Mycorrhizal Fungu, Glomus fasciculatum. Applied Soil Ecology 17:
119-134.
Tawaraya K, Tokarin K, Wagatsuma T. 2001. Dependence of Allium fistulosum Cultivar
on the Arbuscular Mycorrhizal Fungu, Glomus fasciculatum. Applied Soil Ecology 17:
119-134.
Turnbull JW, Martenz PN, Hall N. 1986. Notes on Lesser-Known Australian Trees and
Shrubs with Potential for Wood and Agroforestry. In: Multipurpose Australian Trees
and Shrubs, Lesser Known Spezies for Fuelwood and Agroforestry. (Turnbull, J.W.ed).
ACIAR Canbera. hlm.81-90.

93
Umboh M.I.J, Rahayu G, Affandi A, 2000. Upaya Peningkatan Produksi Gubal Gaharu:
Mikropropagasi Aquilaria Species dan Upaya Peningkatan Bioproses Gubal Gaharunya
[Laporan Penelitian RUT V] Jakarta. Menristek-Dewan Riset Nasional.
Worabay, D. 2009. Identifikasi Fungi Mikoriza Arbuskula (FMA) Pada Pohon Penghasil
Gaharu Gyrinops versteghii Gilg (domke). Di Kampung Asai Kabupaten Manokwari
(Skripsi Sarjana Kehutanan UNIPA).
Wulan T. 2006. Pengaruh Inokulasi Cendawan Mikoriza Arbuskula (CMA) dan Beberapa
Isolat Rhizobakteri terhadap Pertumbuhan Bibit Aquilaria malacensis Lamk. Dan A.
microcarpa Baill. Dipersemaian. [Skripsi]. Jatinangor. Jurusan Biologi Fakultas
FMIPA. Universitas Padjajaran.
Yudhy HB. 2006. Pemanfaatan CMA dan Bradirhizobium dalam Meningkatkan
Produktifitas Kedelai pada Sistem Agroforestri Kayu Bawang (Scorodocarpus
bornensis, Burm. F) Di Ultisol [Disertasi]. Bogor. Sekolah Pascasarjana, Institut
Pertanian Bogor.
Zich F, Compton I. 2001. The Final Frontier: Toward Sustainable Management of Papua
New Guinea’s Agarwood Resource. Sydney: TRAFFIC Oceania and the WWF South
Pacific Programme.
Zulkifli, 2011. Indonesia Kini Ekspor Gaharu langsung ke Tiongkok. Investor Daily
Indonesia. Senin, 14 Maret 2011.http://www.investor.co.id/agribusiness/ indonesia-
kini-ekspor-gaharu-langsung-ke-tiongkok/7628. [ 20 April 2011].

94
Lampiran
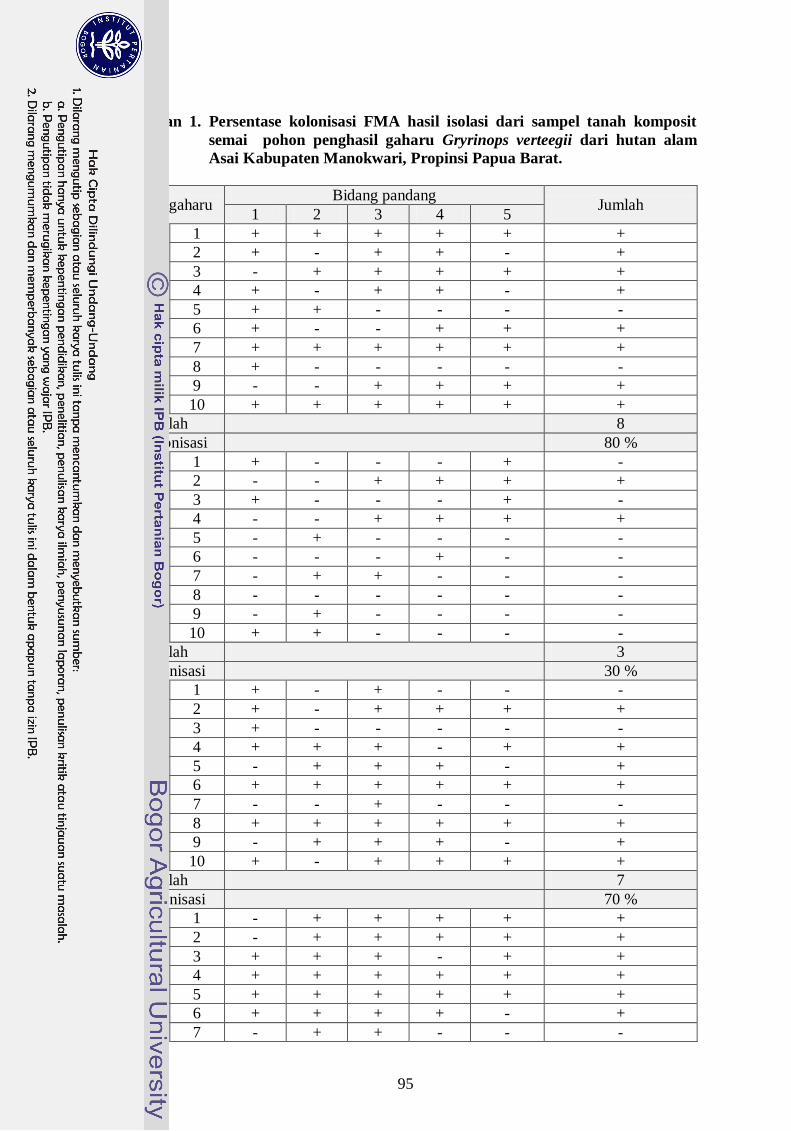
95
Lampiran 1. Persentase kolonisasi FMA hasil isolasi dari sampel tanah komposit
semai pohon penghasil gaharu Gryrinops verteegii dari hutan alam
Asai Kabupaten Manokwari, Propinsi Papua Barat.
Sampel gaharu Bidang pandang
Jumlah 1 2 3 4 5
G1 1 + + + + + +
2 + - + + - +
3 - + + + + +
4 + - + + - +
5 + + - - - -
6 + - - + + +
7 + + + + + +
8 + - - - - -
9 - - + + + +
10 + + + + + +
Jumlah 8
% Kolonisasi 80 %
G2 1 + - - - + -
2 - - + + + +
3 + - - - + -
4 - - + + + +
5 - + - - - -
6 - - - + - -
7 - + + - - -
8 - - - - - -
9 - + - - - -
10 + + - - - -
Jumlah 3
% kolonisasi 30 %
G3 1 + - + - - -
2 + - + + + +
3 + - - - - -
4 + + + - + +
5 - + + + - +
6 + + + + + +
7 - - + - - -
8 + + + + + +
9 - + + + - +
10 + - + + + +
Jumlah 7
% kolonisasi 70 %
G4 1 - + + + + +
2 - + + + + +
3 + + + - + +
4 + + + + + +
5 + + + + + +
6 + + + + - +
7 - + + - - -
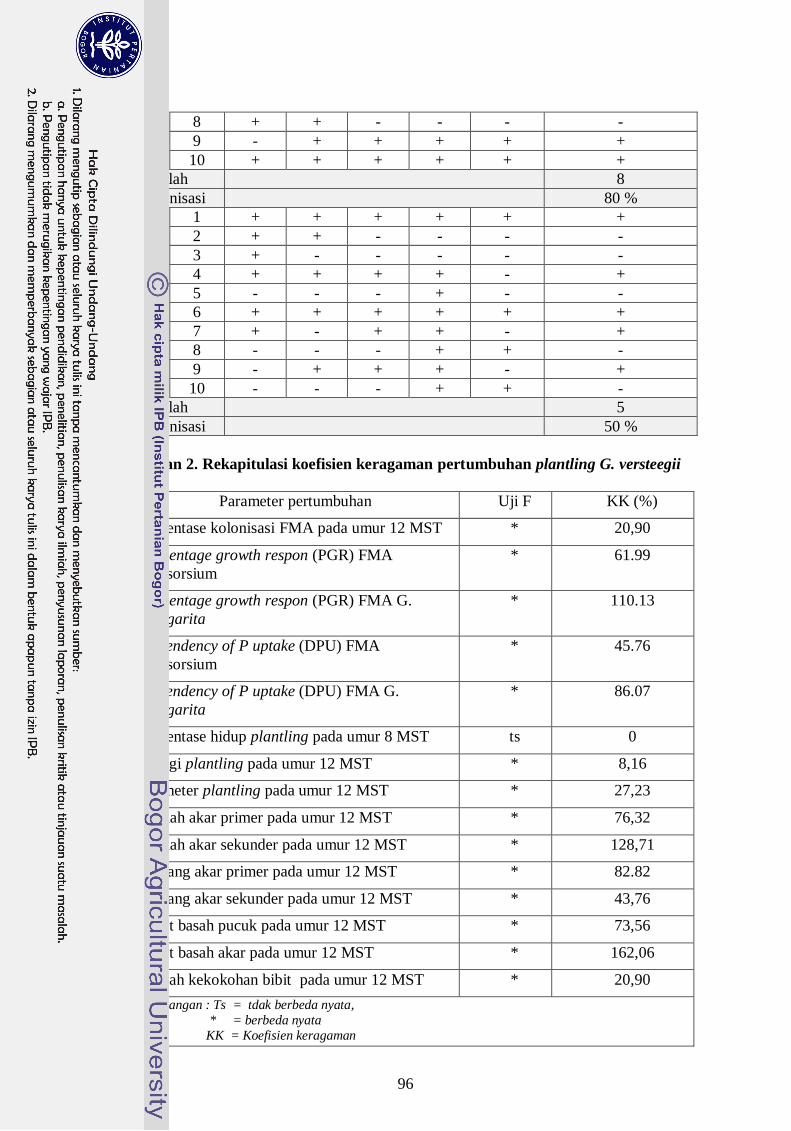
96
8 + + - - - -
9 - + + + + +
10 + + + + + +
Jumlah 8
% kolonisasi 80 %
G5 1 + + + + + +
2 + + - - - -
3 + - - - - -
4 + + + + - +
5 - - - + - -
6 + + + + + +
7 + - + + - +
8 - - - + + -
9 - + + + - +
10 - - - + + -
Jumlah 5
% kolonisasi 50 %
Lampiran 2. Rekapitulasi koefisien keragaman pertumbuhan plantling G. versteegii
Parameter pertumbuhan Uji F KK (%)
Persentase kolonisasi FMA pada umur 12 MST * 20,90
Percentage growth respon (PGR) FMA
Konsorsium
* 61.99
Percentage growth respon (PGR) FMA G.
Margarita
* 110.13
Dependency of P uptake (DPU) FMA
Konsorsium
* 45.76
Dependency of P uptake (DPU) FMA G.
Margarita
* 86.07
Persentase hidup plantling pada umur 8 MST ts 0
Tinggi plantling pada umur 12 MST * 8,16
Diameter plantling pada umur 12 MST * 27,23
Jumlah akar primer pada umur 12 MST * 76,32
Jumlah akar sekunder pada umur 12 MST * 128,71
Panjang akar primer pada umur 12 MST * 82.82
Panjang akar sekunder pada umur 12 MST * 43,76
Berat basah pucuk pada umur 12 MST * 73,56
Berat basah akar pada umur 12 MST * 162,06
Nisbah kekokohan bibit pada umur 12 MST * 20,90
Keterangan : Ts = tdak berbeda nyata,
* = berbeda nyata KK = Koefisien keragaman
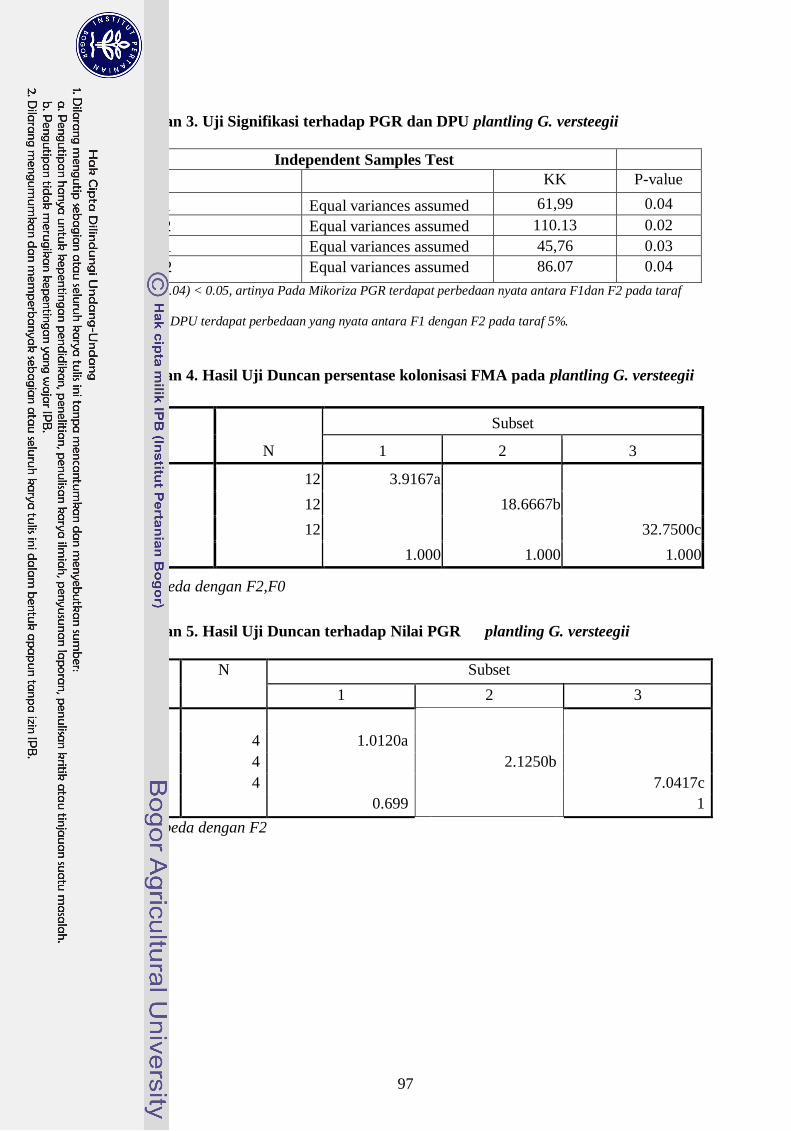
97
Lampiran 3. Uji Signifikasi terhadap PGR dan DPU plantling G. versteegii
Independent Samples Test
KK P-value
PGR F1 Equal variances assumed 61,99 0.04
PGR F2 Equal variances assumed 110.13 0.02
DPU F1 Equal variances assumed 45,76 0.03
DPU F2 Equal variances assumed 86.07 0.04
P-value (0.04) < 0.05, artinya Pada Mikoriza PGR terdapat perbedaan nyata antara F1dan F2 pada taraf
5%.
P-value < DPU terdapat perbedaan yang nyata antara F1 dengan F2 pada taraf 5%.
Lampiran 4. Hasil Uji Duncan persentase kolonisasi FMA pada plantling G. versteegii
FMA N
Subset
1 2 3
F0 12 3.9167a
F2 12 18.6667b
F1 12 32.7500c
Sig. 1.000 1.000 1.000
F1 berbeda dengan F2,F0
Lampiran 5. Hasil Uji Duncan terhadap Nilai PGR plantling G. versteegii
PGR N Subset
1 2 3
PGR
F0 4 1.0120a
F2 4 2.1250b
F1 4 7.0417c
Sig. 0.699 1
F1 berbeda dengan F2
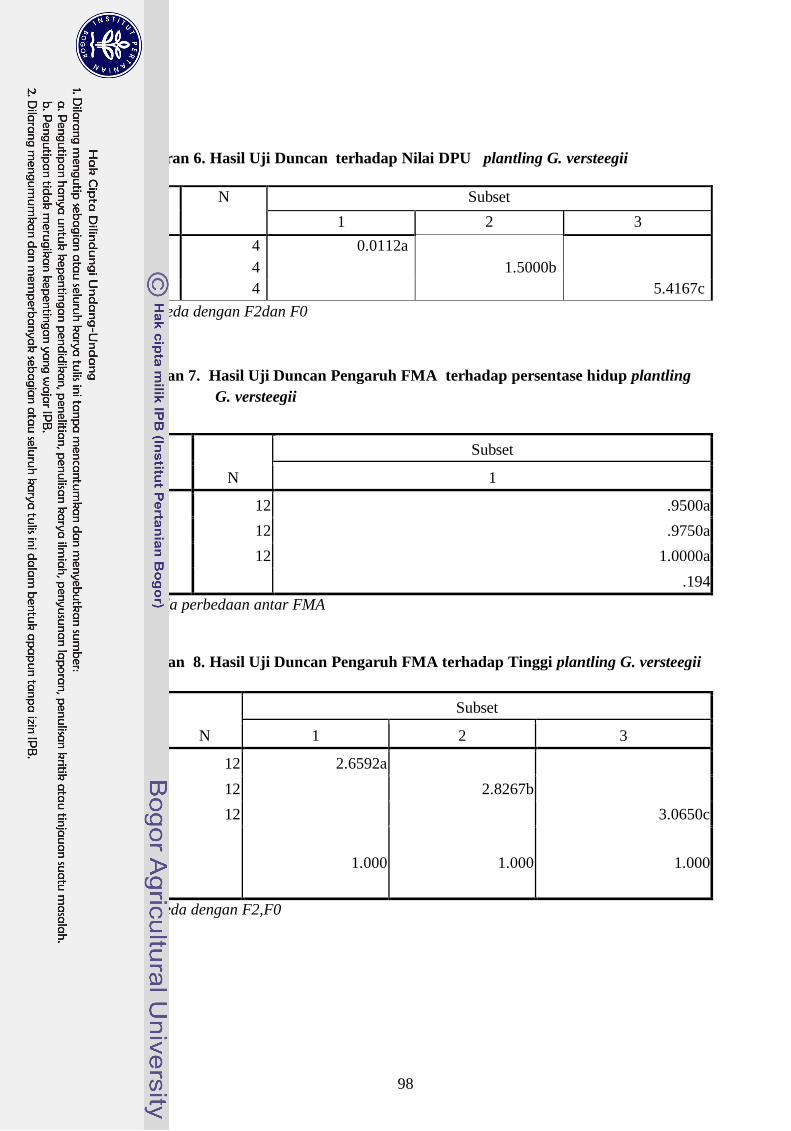
98
Lampiran 6. Hasil Uji Duncan terhadap Nilai DPU plantling G. versteegii
DPU N Subset
1 2 3
F0 4 0.0112a
F2 4 1.5000b
F1 4 5.4167c
F1 berbeda dengan F2dan F0
Lampiran 7. Hasil Uji Duncan Pengaruh FMA terhadap persentase hidup plantling
G. versteegii
FMA N
Subset
1
F0 12 .9500a
F2 12 .9750a
F1 12 1.0000a
Sig. .194
Tidak ada perbedaan antar FMA
Lampiran 8. Hasil Uji Duncan Pengaruh FMA terhadap Tinggi plantling G. versteegii
FMA N
Subset
1 2 3
F0 12 2.6592a
F2 12 2.8267b
F1 12 3.0650c
Sig.
1.000 1.000 1.000
F1 berbeda dengan F2,F0
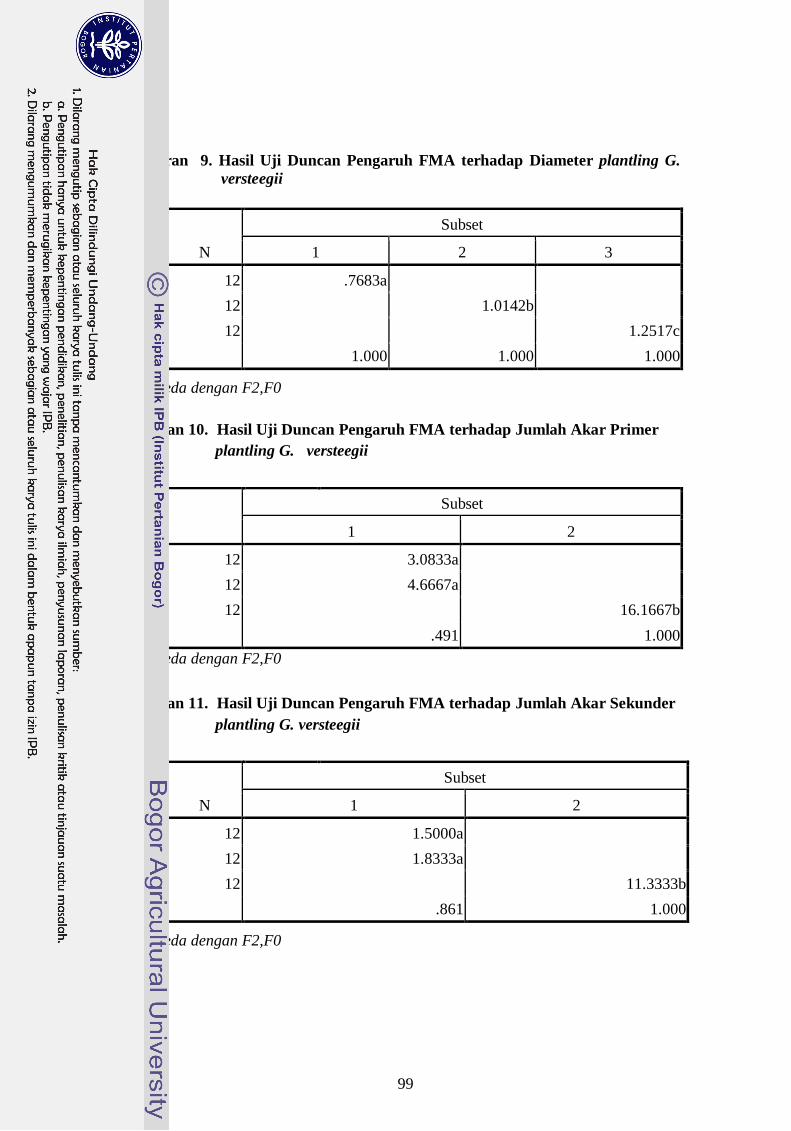
99
Lampiran 9. Hasil Uji Duncan Pengaruh FMA terhadap Diameter plantling G.
versteegii
FMA N
Subset
1 2 3
F0 12 .7683a
F2 12 1.0142b
F1 12 1.2517c
Sig. 1.000 1.000 1.000
F1 berbeda dengan F2,F0
Lampiran 10. Hasil Uji Duncan Pengaruh FMA terhadap Jumlah Akar Primer
plantling G. versteegii
FMA
Subset
1 2
F2 12 3.0833a
F0 12 4.6667a
F1 12 16.1667b
Sig. .491 1.000
F1 berbeda dengan F2,F0
Lampiran 11. Hasil Uji Duncan Pengaruh FMA terhadap Jumlah Akar Sekunder
plantling G. versteegii
FMA N
Subset
1 2
F0 12 1.5000a
F2 12 1.8333a
F1 12 11.3333b
Sig. .861 1.000
F1 berbeda dengan F2,F0

100
Lampiran 12. Hasil Uji Duncan Pengaruh media terhadap Jumlah Akar Primer
plantling G. versteegii
Media N
Subset
1 2
M0 9 4.5556a
M1 9 8.1111b
M2 9 9.4444b
M3 9 9.7778b
Sig. .078
M1, M2 dan M3 berbeda dengan M0

101
Lampiran 13. Hasil Uji Duncan Pengaruh media terhadap Panjang Akar primerer
plantling G. versteegii
Media N
Subset
1 2
M0 9 2.2889a
M3 9 4.2222b
M2 9 5.1111b
M1 9 6.1111b
Sig. .391 .158
M1, M2 dan M3 berbeda dengan M0
Lampiran 14. Hasil Uji Duncan Pengaruh FMA terhadap Panjang Akar Primer
plantling G. Versteegii
FMA N
Subset
1 2
F2 12 1.7917a
F0 12 2.1250a
F1 12 7.0417b
Sig. .699 1.000
F1 berbeda dengan F2,F0
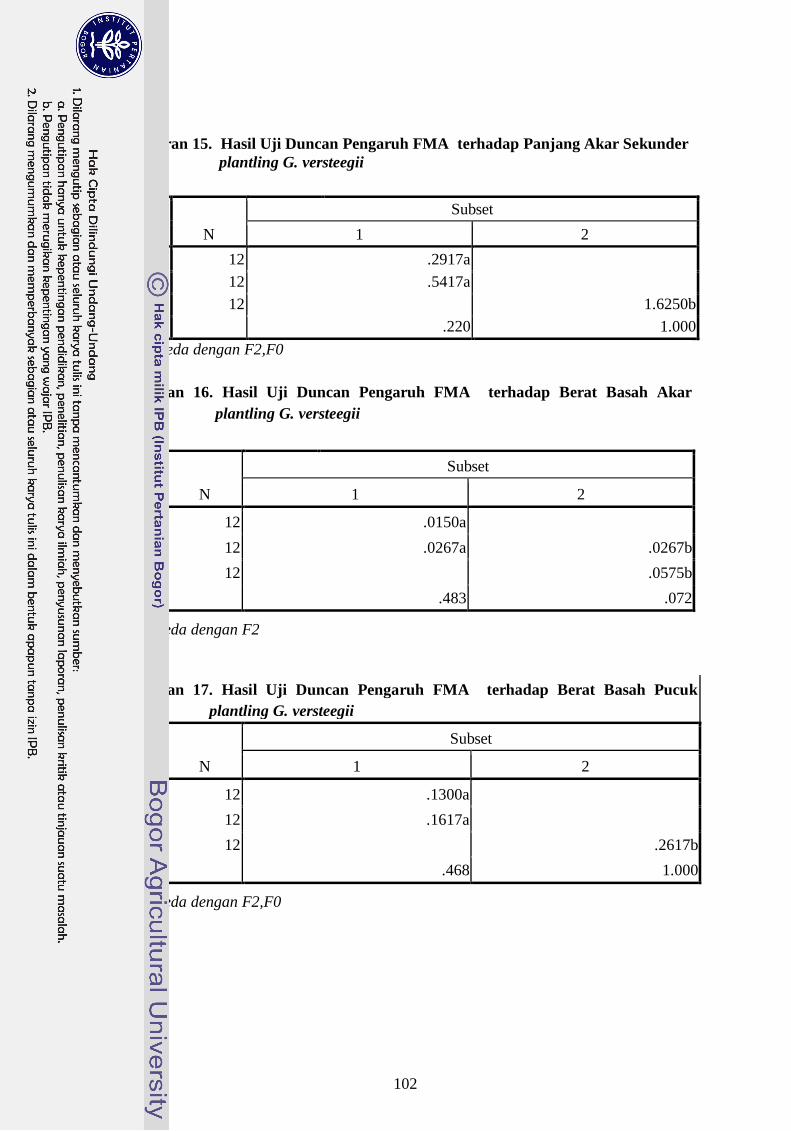
102
Lampiran 15. Hasil Uji Duncan Pengaruh FMA terhadap Panjang Akar Sekunder
plantling G. versteegii
FMA N
Subset
1 2
F2 12 .2917a
F0 12 .5417a
F1 12 1.6250b
Sig. .220 1.000
F1 berbeda dengan F2,F0
Lampiran 16. Hasil Uji Duncan Pengaruh FMA terhadap Berat Basah Akar
plantling G. versteegii
FMA N
Subset
1 2
F0 12 .0150a
F1 12 .0267a .0267b
F2 12 .0575b
Sig. .483 .072
F0 berbeda dengan F2
Lampiran 17. Hasil Uji Duncan Pengaruh FMA terhadap Berat Basah Pucuk
plantling G. versteegii
FMA N
Subset
1 2
F0 12 .1300a
F2 12 .1617a
F1 12 .2617b
Sig. .468 1.000
F1 berbeda dengan F2,F0
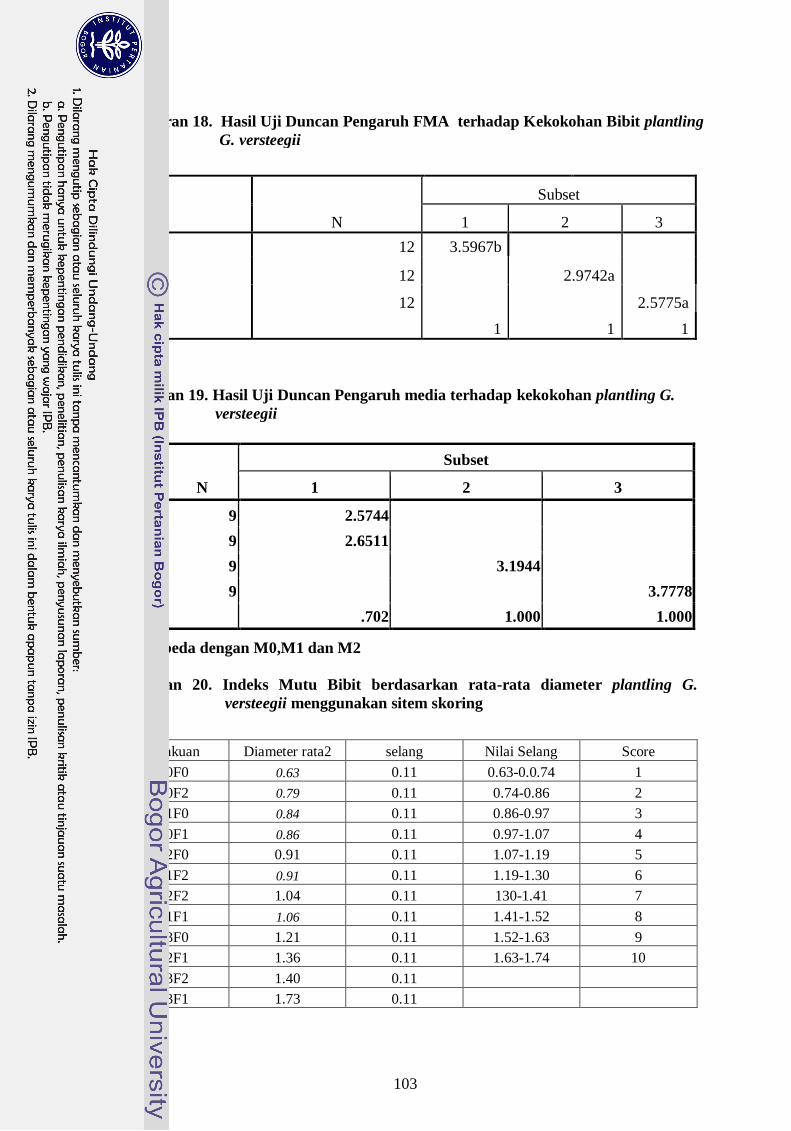
103
Lampiran 18. Hasil Uji Duncan Pengaruh FMA terhadap Kekokohan Bibit plantling
G. versteegii
FMA N
Subset
1 2 3
F1 12 3.5967b
F2 12
2.9742a
F0 12
2.5775a
Sig. 1 1 1
Lampiran 19. Hasil Uji Duncan Pengaruh media terhadap kekokohan plantling G.
versteegii
Media N
Subset
1 2 3
M0 9 2.5744
M1 9 2.6511
M2 9 3.1944
M3 9 3.7778
Sig. .702 1.000 1.000
M3 berbeda dengan M0,M1 dan M2
Lampiran 20. Indeks Mutu Bibit berdasarkan rata-rata diameter plantling G.
versteegii menggunakan sitem skoring
Perlakuan Diameter rata2 selang Nilai Selang Score
M0F0 0.63 0.11 0.63-0.0.74 1
M0F2 0.79 0.11 0.74-0.86 2
M1F0 0.84 0.11 0.86-0.97 3
M0F1 0.86 0.11 0.97-1.07 4
M2F0 0.91 0.11 1.07-1.19 5
M1F2 0.91 0.11 1.19-1.30 6
M2F2 1.04 0.11 130-1.41 7
M1F1 1.06 0.11 1.41-1.52 8
M3F0 1.21 0.11 1.52-1.63 9
M2F1 1.36 0.11 1.63-1.74 10
M3F2 1.40 0.11
M3F1 1.73 0.11
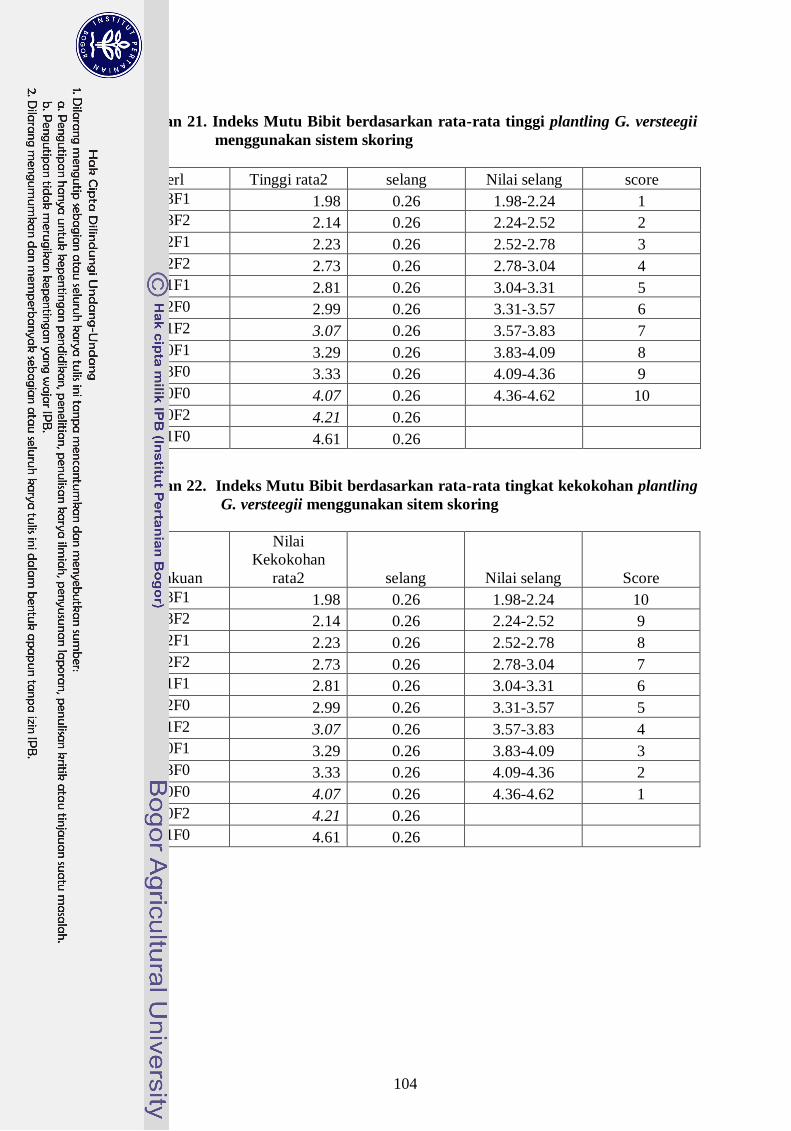
104
Lampiran 21. Indeks Mutu Bibit berdasarkan rata-rata tinggi plantling G. versteegii
menggunakan sistem skoring
Perl Tinggi rata2 selang Nilai selang score
M3F1 1.98 0.26 1.98-2.24 1
M3F2 2.14 0.26 2.24-2.52 2
M2F1 2.23 0.26 2.52-2.78 3
M2F2 2.73 0.26 2.78-3.04 4
M1F1 2.81 0.26 3.04-3.31 5
M2F0 2.99 0.26 3.31-3.57 6
M1F2 3.07 0.26 3.57-3.83 7
M0F1 3.29 0.26 3.83-4.09 8
M3F0 3.33 0.26 4.09-4.36 9
M0F0 4.07 0.26 4.36-4.62 10
M0F2 4.21 0.26
M1F0 4.61 0.26
Lampiran 22. Indeks Mutu Bibit berdasarkan rata-rata tingkat kekokohan plantling
G. versteegii menggunakan sitem skoring
Perlakuan
Nilai
Kekokohan
rata2 selang Nilai selang Score
M3F1 1.98 0.26 1.98-2.24 10
M3F2 2.14 0.26 2.24-2.52 9
M2F1 2.23 0.26 2.52-2.78 8
M2F2 2.73 0.26 2.78-3.04 7
M1F1 2.81 0.26 3.04-3.31 6
M2F0 2.99 0.26 3.31-3.57 5
M1F2 3.07 0.26 3.57-3.83 4
M0F1 3.29 0.26 3.83-4.09 3
M3F0 3.33 0.26 4.09-4.36 2
M0F0 4.07 0.26 4.36-4.62 1
M0F2 4.21 0.26
M1F0 4.61 0.26
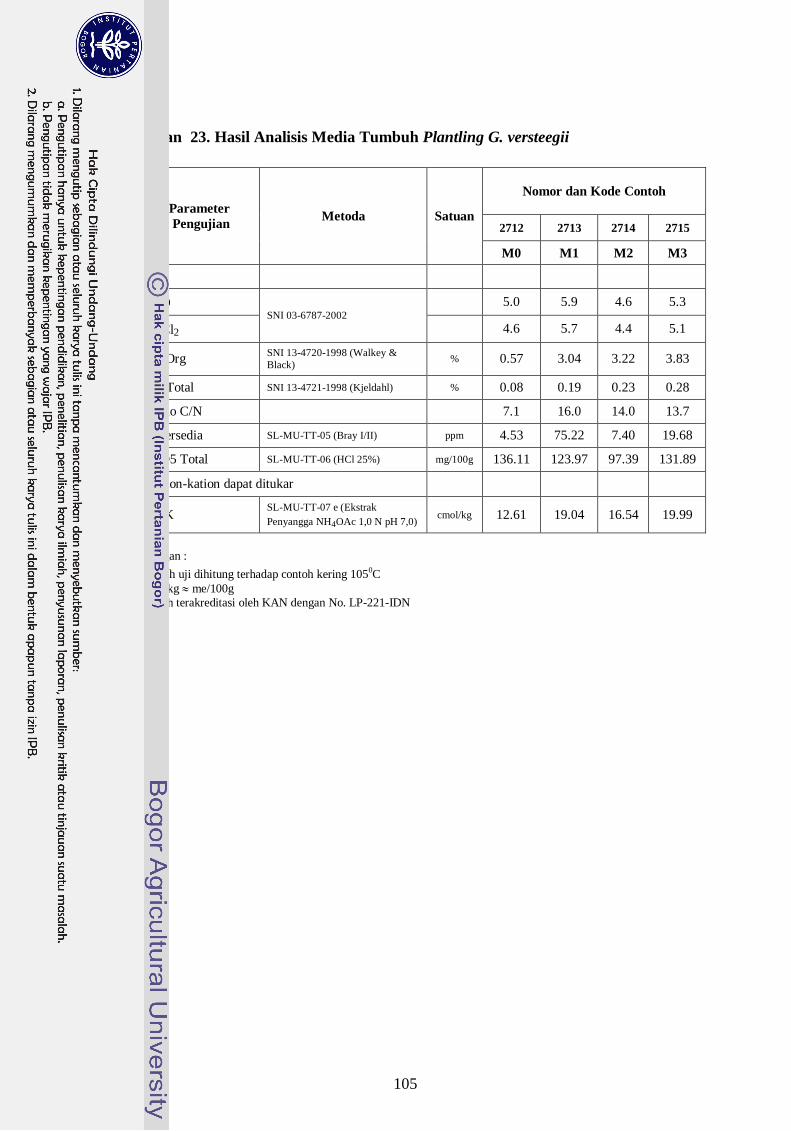
105
Lampiran 23. Hasil Analisis Media Tumbuh Plantling G. versteegii
No. Parameter
Pengujian Metoda Satuan
Nomor dan Kode Contoh
2712 2713 2714 2715
M0 M1 M2 M3
1 *pH
H2O SNI 03-6787-2002
5.0 5.9 4.6 5.3
CaCl2 4.6 5.7 4.4 5.1
2 *C Org SNI 13-4720-1998 (Walkey &
Black) % 0.57 3.04 3.22 3.83
3 *N Total SNI 13-4721-1998 (Kjeldahl) % 0.08 0.19 0.23 0.28
4 Rasio C/N 7.1 16.0 14.0 13.7
5 P Tersedia SL-MU-TT-05 (Bray I/II) ppm 4.53 75.22 7.40 19.68
6 P2O5 Total SL-MU-TT-06 (HCl 25%) mg/100g 136.11 123.97 97.39 131.89
Kation-kation dapat ditukar
7 KTK SL-MU-TT-07 e (Ekstrak
Penyangga NH4OAc 1,0 N pH 7,0) cmol/kg 12.61 19.04 16.54 19.99
Keterangan :
- Contoh uji dihitung terhadap contoh kering 1050C
- cmol/kg me/100g
- * Telah terakreditasi oleh KAN dengan No. LP-221-IDN
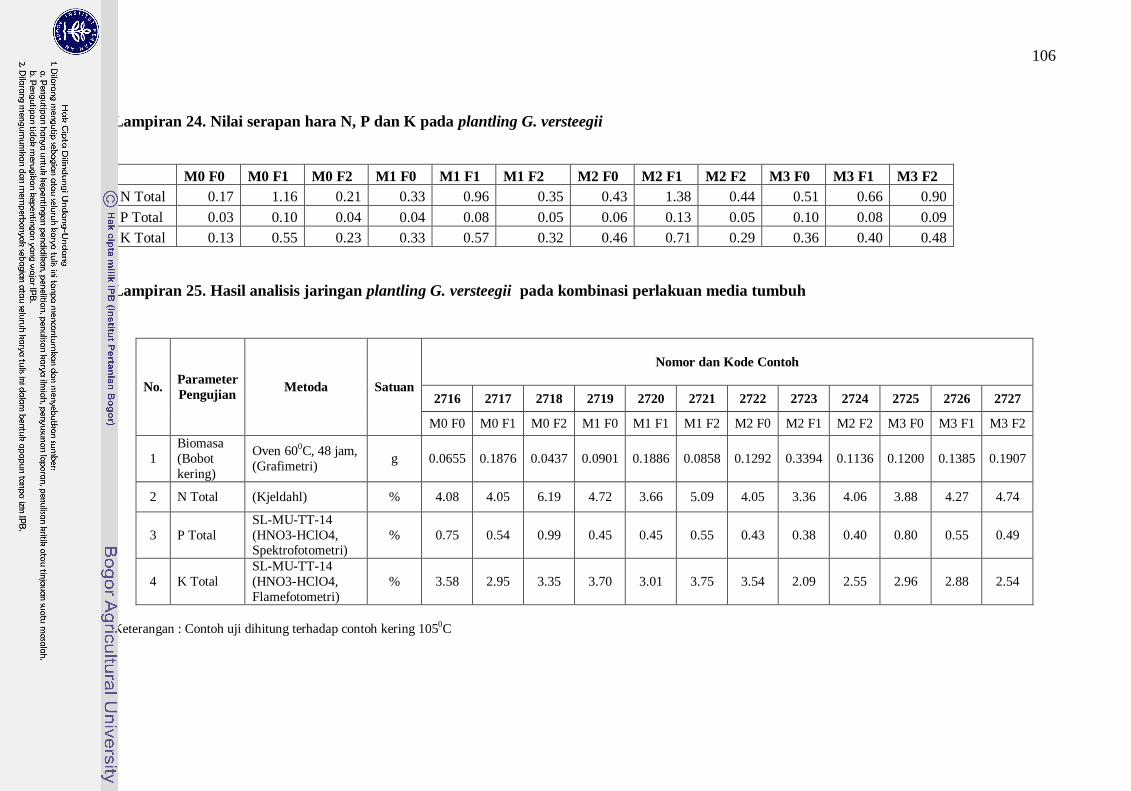
106
Lampiran 24. Nilai serapan hara N, P dan K pada plantling G. versteegii
M0 F0 M0 F1 M0 F2 M1 F0 M1 F1 M1 F2 M2 F0 M2 F1 M2 F2 M3 F0 M3 F1 M3 F2
N Total 0.17 1.16 0.21 0.33 0.96 0.35 0.43 1.38 0.44 0.51 0.66 0.90
P Total 0.03 0.10 0.04 0.04 0.08 0.05 0.06 0.13 0.05 0.10 0.08 0.09
K Total 0.13 0.55 0.23 0.33 0.57 0.32 0.46 0.71 0.29 0.36 0.40 0.48
Lampiran 25. Hasil analisis jaringan plantling G. versteegii pada kombinasi perlakuan media tumbuh
No. Parameter
Pengujian Metoda Satuan
Nomor dan Kode Contoh
2716 2717 2718 2719 2720 2721 2722 2723 2724 2725 2726 2727
M0 F0 M0 F1 M0 F2 M1 F0 M1 F1 M1 F2 M2 F0 M2 F1 M2 F2 M3 F0 M3 F1 M3 F2
1
Biomasa
(Bobot
kering)
Oven 600C, 48 jam,
(Grafimetri) g 0.0655 0.1876 0.0437 0.0901 0.1886 0.0858 0.1292 0.3394 0.1136 0.1200 0.1385 0.1907
2 N Total (Kjeldahl) % 4.08 4.05 6.19 4.72 3.66 5.09 4.05 3.36 4.06 3.88 4.27 4.74
3 P Total
SL-MU-TT-14
(HNO3-HClO4, Spektrofotometri)
% 0.75 0.54 0.99 0.45 0.45 0.55 0.43 0.38 0.40 0.80 0.55 0.49
4 K Total
SL-MU-TT-14
(HNO3-HClO4,
Flamefotometri)
% 3.58 2.95 3.35 3.70 3.01 3.75 3.54 2.09 2.55 2.96 2.88 2.54
Keterangan : Contoh uji dihitung terhadap contoh kering 1050C




















