
KEANEKARAGAMAN DAN PARASITISASI PARASITOID PADA … · keanekaragaman dan parasitisasi parasitoid...
35
KEANEKARAGAMAN DAN PARASITISASI PARASITOID PADA PERTANAMAN SAYURAN DI BOGOR MUHAMAD NURHUDA NUGRAHA DEPARTEMEN PROTEKSI TANAMAN FAKULTAS PERTANIAN INSTITUT PERTANIAN BOGOR BOGOR 2013
-
Upload
truongcong -
Category
Documents
-
view
240 -
download
0
Transcript of KEANEKARAGAMAN DAN PARASITISASI PARASITOID PADA … · keanekaragaman dan parasitisasi parasitoid...

KEANEKARAGAMAN DAN PARASITISASI PARASITOID
PADA PERTANAMAN SAYURAN DI BOGOR
MUHAMAD NURHUDA NUGRAHA
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
2013

PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Keanekaragaman dan
Parasitisasi Parasitoid pada Pertanaman Sayuran di Bogor adalah benar karya saya
dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun
kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip
dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah
disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir
skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut
Pertanian Bogor.
Bogor, September 2013
Muhamad Nurhuda Nugraha
NIM A34070052

ABSTRAK
MUHAMAD NURHUDA NUGRAHA. Keanekaragaman dan Parasitisasi
Parasitoid pada Pertanaman Sayuran di Bogor. Dibimbing oleh DAMAYANTI
BUCHORI.
Pada suatu pertanaman, lingkungan agroekosistem dapat memengaruhi
keanekaragaman serta keefektifan komunitas parasitoid sebagai musuh alami
serangga hama. Sebuah survei untuk mempelajari keanekaragaman serangga
parasitoid dan parasitisasinya dilakukan di beberapa lahan sayuran di Bogor pada
bulan Agustus sampai Oktober 2012 berdasarkan 3 kriteria pertanaman berbeda,
yaitu: pola budidaya (organik dan konvensional), keanekaragaman tanaman
sayuran di sekitar (5-8 sp. dan 9-15 sp.) dan perbedaan umur pertanaman (3, 4 dan
5 bulan). Dengan membuat garis transek sepanjang 50 langkah di setiap lahan
pertanaman yang telah dipilih, telur larva dan pupa serangga hama dikumpulkan,
lalu dihitung kejadian parasitisasi dan keanekaragaman parasitoidnya. Hasil
penelitian ini menunjukkan tidak ada perbedaan yang signifikan terhadap tiga
kriteria pertanaman yang digunakan. Walaupun begitu, berdasarkan
keanekaragaman tanaman sekitar, tingkat parasitisasi pada lahan dengan
keanekaragaman tanaman sayuran yang lebih sedikit (5-8 sp.), lebih tinggi
dibandingkan lahan dengan keanekaragaman tanaman sayuran yang lebih banyak
(9-15 sp.). Hal ini dapat disebabkan karena keterbatasan unit pengambilan contoh
dan faktor lain yang memengaruhi seperti habitat sekitar, cuaca dan interaksi yang
kompleks antar spesies dalam suatu agroekosistem.
Kata kunci: keanekaragaman, parasitisasi, parasitoid, praktek pertanian, umur
tanaman

ABSTRACT
MUHAMAD NURHUDA NUGRAHA. Diversity and Parasitism of Parasitoids in
Vegetable Plants in Bogor. Supervised by DAMAYANTI BUCHORI.
In an agricultural area, agroecosystem could influence diversity and
parasitism of parasitoids as natural enemies of insect pests. A survey to learn
diversity and rate of parasitism of insect pests by parasitoids had been conducted
in several agricultural fields with different criteria, such as farming practice
(organic and convensional), species richness of vegetable crops (5-8 sp. and 9-15
sp.) and different crop ages (3, 4 and 5 months), in Bogor from August to October
2012. By making a 50 steps line transect for every field, eggs, larvae and pupa of
insect pests were collected monthly, reared in laboratory and incidence of
parasitism were calculated. The results showed that there were no significant
differences on diversity and rate of parasitism based on three criterias used.
However, based on the diversity of vegetable plant species around, the rate of
parasitism in field with less plant species diversity (5-8 sp.) was higher than the
field with more vegetable plant species diversity (9-15 sp.). These results might be
caused by the limited number of samplings conducted and other potential factors
such as surrounding habitat, seasonal fluctuation, and complex interactions
between species in agroecosystem.
Key words: biodiversity, parasitism, parasitoids, farming practice, different plant
age

© Hak Cipta Milik IPB, tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
yang wajar IPB.
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
dalam bentuk apa pun tanpa izin IPB.

KEANEKARAGAMAN DAN PARASITISASI PARASITOID
PADA PERTANAMAN SAYURAN DI BOGOR
MUHAMAD NURHUDA NUGRAHA
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Pertanian
pada
Program Studi Proteksi Tanaman
DEPARTEMEN PROTEKSI TANAMAN
FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR
BOGOR
2013

Judul Skripsi : Keanekaragaman dan Parasitisasi Parasitoid pada
Pertanaman Sayuran di Bogor Nama Mahasiswa : Muhamad Nurhuda Nugraha
NIM : A34070052
Disetujui oleh
Dr. Ir. Damayanti Buchori, MSc.
Dosen Pembimbing
Diketahui oleh
Dr. Ir. Abdjad Asih Nawangsih, M.Si
Ketua Departemen
Tanggal lulus :

PRAKATA
Syukur alhamdulillah penulis panjatkan kehadirat Allah SWT karena berkat
rahmat dan karuniaNya, penelitian dan penulisan tugas akhir sarjana ini dapat
diselesaikan. Tugas akhir sarjana ini berjudul: ”Keanekaragaman dan Parasitisasi
Parasitoid pada Pertanaman Sayuran di Bogor” dan berlangsung sejak Maret 2012
hingga September 2013. Penulis sangat bersyukur atas ilmu dan pengalaman yang
didapat selama proses meraih gelar Sarjana Pertanian di Institut Pertanian Bogor.
Penghargaan dan ucapan terima kasih penulis sampaikan kepada Dr. Ir.
Damayanti Buchori, M.Sc atas bimbingan yang diberikan hingga penelitian dan
penulisan tugas akhir sarjana ini diselesaikan, dan juga kepada Dr. Akhmad Rizali
atas arahan dan pelatihan yang diberikan. Penulis yakin ilmu baru yang didapat
seperti cara penggunaan perangkat keras GPS Map dan perangkap lunak Quantum
GIS dan R, akan lebih bermanfaat suatu hari nanti. Ucapan terima kasih juga
penulis sampaikan kepada Mba Adha Sari dan Mba Nita selaku laboran di
Laboratorium Pengendalian Hayati yang telah banyak membantu dan juga kepada
Pa Sudarsono serta Yasin Farid sebagai teman dan rekan kerja yang baik dalam
penelitian ini. Penulis juga ucapkan terima kasih kepada pihak-pihak yang telah
membantu dalam tahap identifikasi dan analisis statistik yaitu Pa Puji, Mba Laras,
Rizki, Tuti, Pa Uyung serta kepada teman-teman di Goettingen yang telah banyak
membantu memudahkan akses terhadap jurnal-jurnal internasional. Kepada Bu
Tri, Pa Giyanto dan Pa Kikin atas diskusi dan arahan selama penulis menempuh
studi di departemen ini. Kepada seluruh teman-teman di Departemen Proteksi
Tanaman dan Laboratorium Pengendalian Hayati Faperta IPB secara khusus dan
teman-teman dari berbagai departemen dan fakultas di Institut Pertanian Bogor
baik junior maupun senior serta teman seperjuangan atas dukungan dan
kebersamaannya. Akhirnya, ucapan terima kasih penulis sampaikan sedalam-
dalamnya kepada Ibu dan Bapak yang sangat Penulis cintai atas perhatian, kasih
sayang, kepercayaan, doa serta kesabaran yang diberikan. Juga atas diskusi dan
cerita-cerita masa lalu yang menginspirasi dan memotivasi penulis dalam
menjalani hidup ini.
Akhir kata, tidak ada yang sempurna di dunia ini, karena kesempurnaan
hanya milik Tuhan Sang Pencipta seluruh alam. Semoga tulisan ini bermanfaat.
Aamiin.
Bogor, September 2013
Muhamad Nurhuda Nugraha

i
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 3
Manfaat Penelitian 3
BAHAN DAN METODE 4
Waktu dan Lokasi Penelitian 4
Metode Penelitian 5
Survei dan Penentuan Lokasi 5
Pengambilan Contoh Serangga 9
Identifikasi Serangga 9
Analisis Data 9
HASIL DAN PEMBAHASAN 10
Keanekaragaman, Serangan Hama dan Parasitisasi Parasitoid pada
Pertanaman Sayuran di Bogor 10
Pengaruh Sistem Budidaya terhadap Keanekaragaman dan Tingkat
Parasitisasi Parasitoid: Studi Kasus pada Pertanaman Caisin 13
Pengaruh Keanekaragaman Tanaman Sayuran di Sekitar terhadap
Keanekaragaman dan Tingkat Parasitisasi Parasitoid: Studi Kasus pada
Pertanaman Polikultur Kubis 15
Pengaruh Perbedaan Umur Tanaman terhadap Keanekaragaman
Parasitoid: Studi Kasus pada Pertanaman Terung 18
SIMPULAN DAN SARAN 20
DAFTAR PUSTAKA 21
RIWAYAT HIDUP 25

vi
DAFTAR TABEL
1. Deskripsi lokasi penelitian di daerah Bogor 4
2. Keanekaragaman spesies sayuran di sekitar pertanaman polikultur kubis
di Desa Tugu Selatan, Cisarua 7
3. Keanekaragaman, serangan hama dan parasitisasi parasitoid pada
pertanaman sayuran di Bogor 10
4. Frekuensi serangan hama pada pertanaman sayuran di Bogor 11
5. Jumlah parasitoid (per individu inang) pada pertanaman sayuran di Bogor 12
6. Keanekaragaman, tingkat serangan hama dan parasitisasi parasitoid pada
pertanaman caisin organik dan konvensional 13
7. Komposisi spesies parasitoid antar lahan dengan sistem pertanian berbeda
berdasarkan indeks kemiripan Sorensen 14
8. Kelimpahan hama dan parasitoid pada pertanaman terung dengan umur
3, 4 dan 5 bulan 19
DAFTAR GAMBAR
1. Peta lokasi penelitian di daerah Bogor 5
2. Gambar pertanaman caisin (a) organik di Desa Tugu Selatan dan (b)
konvensional di Desa Situ Daun 6
3. Pertanaman kubis dengan keanekaragaman (a) 5-8 spesies tanaman
sayuran dan (b) 9-15 spesies tanaman sayuran di sekitar 7
4. Pertanaman terung berumur (a) 3 bulan di Desa Bojong dan (b) terung
berumur 4 bulan di Desa Bantarsari 8
5. Rataan tingkat serangan hama dan parasitisasinya pada pertanaman
sayuran di Bogor 13
6. Hubungan tropik herbivor (hama) dan parasitoidnya pada (a) pertanaman
caisin organik dan (b) pertanaman caisin konvensional 14
7. Keanekaragaman, tingkat serangan hama dan parasitisasi parasitoid pada
pertanaman kubis polikultur dengan 5-8 spesies dan 9-15 spesies
tanaman sayuran di sekitarnya 15
8. Hubungan tropik herbivor (hama) dan parasitoidnya pada pertanaman
kubis polikultur dengan (a) 5-8 spesies dan (b) 9-15 spesies tanaman
sayuran di sekitarnya 16
9. Keanekaragaman parasitoid pada pertanaman kubis polikultur dengan
(a) 5-8 dan (b) 9-15 spesies tanaman sayuran di sekitar serta (c) irisan
diantara keduanya 17
10. Keanekaragaman, tingkat serangan hama dan parasitisasi parasitoid pada
pertanaman terung dengan umur 3 bulan, 4 bulan dan 5 bulan 18
11. Hubungan tropik herbivor (hama) dan parasitoidnya pada pertanaman
terung dengan umur (a) 3 bulan, (b) 4 bulan dan (c) 5 bulan 19

1
PENDAHULUAN
Latar Belakang
Sayuran memiliki peranan penting dalam kehidupan manusia sebagai salah
satu sumber asupan vitamin dan mineral. Pada tahun 2011, produktivitas rata-rata
sayuran segar di Indonesia mencapai 9,5 ton/ha (FAO 2013). Nilai ini tidak
sebesar tahun sebelumnya yang mencapai 10 ton/ha. Penurunan ini tidak lepas
dari permasalahan hama dalam praktek budidayanya. Di sisi lain, praktek
budidaya dengan penggunaan pestisida sintetik yang berlebihan pada sayuran
segar dalam mengendalikan permasalahan hama dinilai lebih banyak
menimbulkan efek negatif di kalangan produsen maupun konsumen. Efek negatif
ini dapat berupa timbulnya resistensi hama terhadap pestisida, kontaminasi pada
bahan pangan serta pencemaran lingkungan.
Pengendalian hama dan penyakit terpadu (PHT) merupakan suatu konsep
yang dikembangkan sebagai salah satu bentuk resolusi dari efek negatif
intensifikasi dalam pertanian. Dalam konsep ini, serangga musuh alami memiliki
peran yang sangat penting dalam menekan serangan organisme pengganggu
tanaman (Losey dan Vaugan 2006; Buchori et al. 2008). Salah satu bentuk jasa
ekosistem (ecosystem services) dalam suatu areal pertanian yang banyak
menguntungkan petani adalah parasitisme oleh serangga parasitoid terhadap
serangga hama.
Secara sederhana, parasitoid dapat diartikan sebagai serangga yang stadia
larvanya mampu memarasit serangga lainnya (Godfray 1994). Walaupun
demikian, parasitoid merupakan kelompok yang dipisahkan dari kelompok
organisme parasit pada umumnya. Hal ini menurut Gordh et al. (1999)
dikarenakan parasitoid memiliki beberapa karakteristik yang unik, diantaranya
adalah: 1) sifat parasitisasi parasitoid hanya diekspresikan pada stadia larva, 2)
stadia imago hidup bebas di alam, 3) larva parasitoid biasanya membunuh dan
memakan inangnya, 4) parasitoid memiliki ukuran tubuh yang kurang lebih sama
dengan inangnya, 5) parasitoid dan inangnya berada pada grup taksonomi yang
berdekatan (serangga dengan serangga). Berdasarkan keanekaragaman, biologi
dan perkembangan/jenis parasitisasinya, serangga parasitoid telah diperhatikan
sejak lama sebagai salah satu agen pengendalian hayati yang efektif. Sejak awal
1970, sebanyak 907 spesies parasitoid yang didominasi oleh ordo Hymenoptera
(84,4%) dan Diptera (1,4%) telah digunakan dalam program pengendalian hayati
di berbagai tempat di penjuru dunia (Clausen 1978).
Dalam upaya pelestarian serangga parasitoid, terdapat beberapa hal yang
dapat memengaruhi keanekaragaman dan keefektifan komunitas parasitoid
sebagai musuh alami pada suatu pertanaman, diantaranya adalah pengelolaan
lingkungan pada suatu agroekosistem (Letourneau dan Altieri 1999). Setiap
wilayah akan memiliki karakter agroekosistem pertanaman yang berbeda sebagai
hasil dari iklim, topografi, tanah, kondisi ekonomi dan sejarah penggunan lahan
yang memengaruhinya. Perbedaan karakter agroekosistem pertanaman juga dapat
ditentukan oleh tujuan dan skala produksinya. Hal ini berkaitan dengan teknologi,
sumberdaya dan cara budidaya yang digunakan (Mattson et al. 1984; Altieri
1989). Walaupun demikian, pertanaman dapat dikategorikan berdasarkan tipe dari

2
pertanian atau agroekosistemnya (Norman 1979). Pertanaman organik dan
konvensional atau pertanaman polikultur dan monokultur merupakan contoh
kategori pertanaman yang dapat dibedakan berdasarkan pola budidayanya.
Pada pertanaman yang dikelola secara organik, keanekaragaman spesies
pada tiga level tropik (tanaman-herbivor-parasitoid) lebih tinggi secara signifikan
dibandingkan dengan lahan yang dikelola secara konvensional (Macfadyen 2009).
Sebenarnya hubungan keanekaragaman musuh alami pada suatu pertanaman
dengan keefektifan dalam pengendalian hayati itu sendiri masih belum begitu
jelas. Hal ini karena adanya pola interaksi yang sangat kompleks antar spesies
yang terjadi dalam suatu ekosistem. Menurut Stireman et al. (2005),
keanekaragaman musuh alami dapat meningkatkan keefektifan pengendalian
hayati apabila musuh alami yang berbeda dapat melengkapi satu sama lain. Model
ekologi seperti ini dapat terjadi apabila setiap spesies musuh alami
memangsa/memarasit kelompok hama, baik dari jenis atau waktu yang berbeda.
Keanekaragaman musuh alami juga berpotensi meningkatkan keefektifan
pengendalian hayati, karena dapat meningkatkan kemungkinan hadirnya agen
musuh alami yang superior. Namun, interaksi negatif antar spesies musuh alami
dapat juga terjadi dan mengakibatkan pengandalian hayati menjadi kurang efektif.
Keanekaragaman musuh alami seperti parasitoid dapat meningkatkan keefektifan
pengendalian hayati apabila interaksi positif yang terjadi lebih kuat daripada
interaksi negatif yang dihasilkan diantara spesies musuh alami dalam suatu
agroekosistem (Letourneau dan Bothwell 2008).
Di sisi lain, pertanaman yang dikelola secara konvensional dengan
intensifikasi berupa peningkatan penggunaan pupuk kimia dan pestisida, dapat
menyebabkan degradasi habitat dan kehilangan keanekaragaman beberapa grup
taksonomi (Lohaus et al. 2012). Penggunaan pupuk kimia dapat mempengaruhi
pertumbuhan, perkembangan, respon biokimia dan fenologi tanaman secara
langsung. Hal ini secara tidak langsung juga akan mempengaruhi interaksi antara
tanaman, herbivor dan musuh alaminya. Kualitas nutrisi herbivor dipengaruhi oleh
kualitas nutrisi tanaman, sedangkan tingkat parasitisasi dipengaruhi juga oleh
kualitas inangnya (herbivor). Benrey dan Denmo (1997) menunjukkan bahwa
larva Pieris rapae yang pertumbuhannya lambat sebagai akibat dari kurangnya
nutrisi yang didapat dari tanaman, memiliki tingkat parasitisasi oleh Cotesia
glomerata yang lebih tinggi dibandingkan dengan larva yang pertumbuhannya
lebih cepat. Aplikasi pestisida lewat udara (semprot) juga dapat mempengaruhi
interaksi antara tanaman, herbivor dan parasitoidnya dalam suatu ekosistem secara
lebih luas. Hal ini karena dampak pesitisida terdapat organisme non-target
menjadi tidak terhindarkan. Keefektifan pengandalian hayati menjadi terganggu
sebagai akibat dari toksisitas semprotan yang diaplikasikan secara intensif dan
berulang-ulang (Ridgway et al. 1976; Riehl et al. 1980).
Keanekaragaman tanaman sekitar sebagai salah satu bagian dari struktur
jaring-jaring makanan juga dapat memengaruhi keberadaan dan tingkat
parasitisasi oleh serangga parasitoid. Hal ini berkaitan dengan ketersediaan inang
alternatif, nektar sebagai sumber makanan parasitoid dewasa dan selter sebagai
tempat berlindung yang disediakan oleh tanaman lain di sekitar pertanaman
(Menalled et al. 1999). Semakin beragamnya suatu pertanaman, semakin
melimpah pula parasitoidnya dan diharapkan memberikan pengendalian hama
yang lebih baik. Moreira dan Mooney (2013) menunjukkan bahwa pertanaman

3
sayuran Baccharis salicifolia polikultur dengan empat genotipe berbeda,
menghasilkan populasi aphid, semut dan parasitoid yang lebih melimpah
dibandingkan pertanaman Baccharis salicifolia monokultur. Hal ini tentunya
mempengaruhi perubahan pola interaksi tropik yang terjadi dalam agroekosistem
pertanaman tersebut. Sebaliknya, penurunan keanekaragaman inang dapat
memengaruhi keanekaragaman organism parasitoid secara negatif. Hal ini
tergambarkan dalam model hubungan ekologi antara inang dan
parasit/parasitoidnya (Lafferty 2012).
Umur tanaman sebagai bagian dari fenologi tanaman dapat memberikan
informasi kejadian parasitisasi dan keanekaragaman spesies parasitoid pada
tanaman tersebut (Barron et al. 2004). Hal ini menjadi penting karena jenis hama
yang menyerang dan parasitoid yang berperan dalam pengendalian hayati dapat
berbeda tergantung pada kondisi pertumbuhan dan perkembangan tanaman yang
berbeda.
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari tingkat parasitisasi serta
keanekaragaman serangga parasitoid pada beberapa pertanaman sayuran di daerah
Bogor berdasarkan tiga kriteria pertanaman, yaitu pola budidaya, keanekaragaman
tanaman sayuran di sekitar lahan dan perbedaan umur tanaman.
Manfaat Penelitian
Manfaat penelitian ini adalah untuk memberikan informasi dasar mengenai
keanekaragaman, persebaran serta parasitisasi parasitoid pada beberapa lahan
pertanaman sayuran di wilayah Bogor serta mengetahui faktor-faktor yang dapat
memengaruhinya. Informasi ini diharapkan dapat menjadi bahan masukan dalam
pembuatan model pertanian berkelanjutan yang diiringi dengan sistem PHT.
4
BAHAN DAN METODE
Waktu dan Lokasi Penelitian
Penelitian ini dilaksanakan dari bulan Maret 2012 sampai Mei 2013.
Pengambilan contoh serangga dilakukan selama tiga bulan berturut-turut dari
bulan Agustus sampai Oktober 2012 pada beberapa lahan sayuran di empat desa
di daerah Bogor, yaitu: Tugu Selatan (Cisarua), Situ Daun (Tenjolaya), Bojong
(Kemang) dan Bantarsari (Ranca Bungur) (Tabel 1). Curah hujan pada bulan
Agustus, September and Oktober 2012 masing-masing adalah 110 mm, 370 mm
dan 374 mm. Proses identifikasi dilakukan di Laboratorium Pengendalian Hayati,
Departemen Proteksi Tanaman, Institut Pertanian dari bulan November 2012
sampai April 2013.
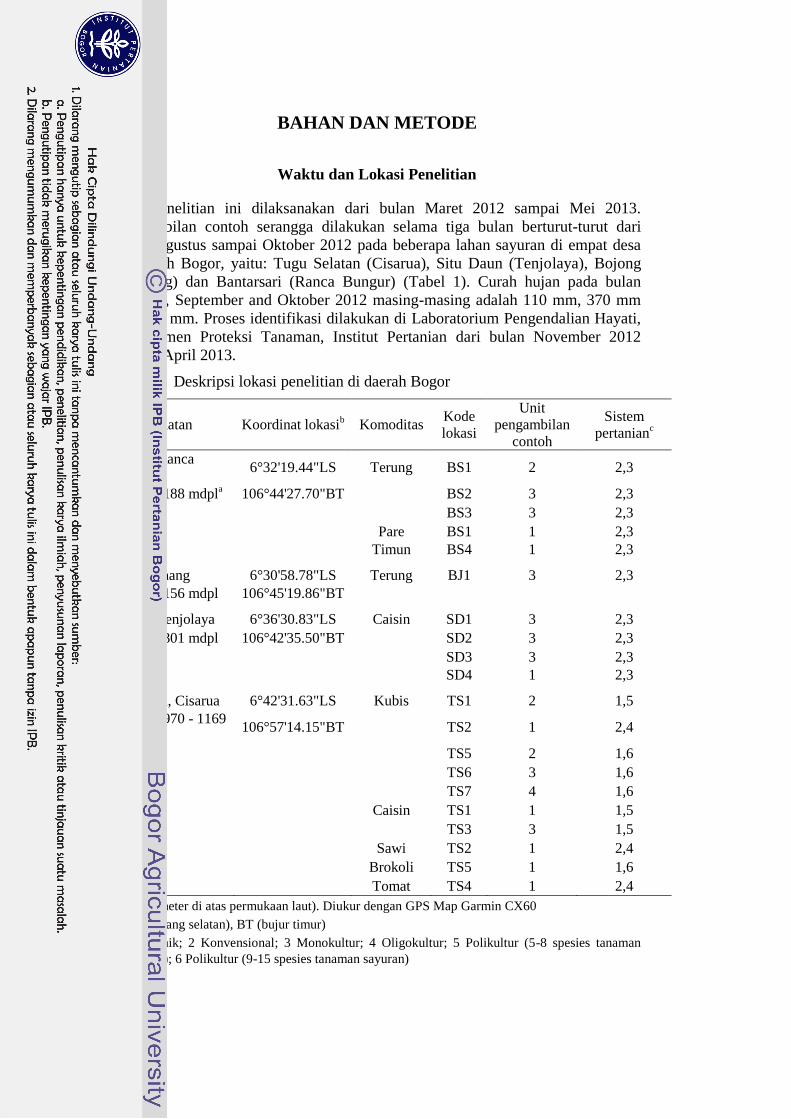
Tabel 1 Deskripsi lokasi penelitian di daerah Bogor
Desa, Kecamatan Koordinat lokasib Komoditas
Kode
lokasi
Unit
pengambilan
contoh
Sistem
pertanianc
Bantarsari, Ranca
Bungur 6°32'19.44"LS Terung BS1 2 2,3
Ketinggian: 188 mdpla 106°44'27.70"BT
BS2 3 2,3
BS3 3 2,3
Pare BS1 1 2,3
Timun BS4 1 2,3
Bojong, Kemang 6°30'58.78"LS Terung BJ1 3 2,3
Ketinggian: 156 mdpl 106°45'19.86"BT
Situ Daun, Tenjolaya 6°36'30.83"LS Caisin SD1 3 2,3
Ketinggian: 301 mdpl 106°42'35.50"BT
SD2 3 2,3
SD3 3 2,3
SD4 1 2,3
Tugu Selatan, Cisarua 6°42'31.63"LS Kubis TS1 2 1,5
Ketinggian: 970 - 1169
mdpl 106°57'14.15"BT
TS2 1 2,4
TS5 2 1,6
TS6 3 1,6
TS7 4 1,6
Caisin TS1 1 1,5
TS3 3 1,5
Sawi TS2 1 2,4
Brokoli TS5 1 1,6
Tomat TS4 1 2,4
a mdpl (meter di atas permukaan laut). Diukur dengan GPS Map Garmin CX60
b LS (lintang selatan), BT (bujur timur)
c 1 Organik; 2 Konvensional; 3 Monokultur; 4 Oligokultur; 5 Polikultur (5-8 spesies tanaman
sayuran); 6 Polikultur (9-15 spesies tanaman sayuran)

5
Metode Penelitian
Survei dan Penentuan Lokasi Penelitian ini dimulai dengan melakukan pencarian lahan pertanaman
sayuran di wilayah Bogor. Sebanyak 31 calon wilayah diseleksi menjadi 16
wilayah yang tersebar di 15 desa dan 11 kecamatan berdasarkan kemudahan akses
dan kriteria yang ditentukan. Lokasi pengambilan contoh kemudian dipilih
menjadi 5 wilayah di 4 desa berdasarkan 3 kriteria pertanaman, yaitu sistem
budidaya, keanekaragaman tanaman sekitar dan perbedaan umur tanaman
(Gambar 1). Berdasarkan lokasi yang dipilih, ditemukan delapan jenis komoditas
sayuran yang dapat diamati, yaitu caisin, kubis, brokoli, sawi, terung, tomat,
timun dan pare (Tabel 1).
Gambar 1 Peta lokasi penelitian di daerah Bogor
Caisin merupakan komoditas yang mewakili kriteria pertanaman
berdasarkan sistem budidayanya, yaitu organik dan konvensional (Tabel 1).
Pertananaman caisin yang ada di Desa Tugu Selatan, Cisarua dan Situ Daun,
Tenjolaya memiliki sistem pertanaman yang cukup berbeda. Caisin di Desa Tugu
Selatan dikelola secara polikultur-organik dengan mengandalkan pupuk organik
dan pestisida nabati dan kultur teknis sebagai cara untuk mengendalikan serangan
hama dan penyakitnya. Sedangkan caisin di Desa Situ Daun dikelola secara
monokultur-konvensional yang sangat bergantung pada penggunaan pupuk dan
pestisida kimia sintetik dalam praktek budidayanya (Gambar 2).
Bogor, Jawa Barat

6
Gambar 2 Gambar pertanaman caisin (a) organik di Desa Tugu Selatan dan (b)
konvensional di Desa Situ Daun
Pada pertanaman sayuran di Desa Tugu Selatan, terdapat dua pertanaman
polikultur organik yang dikelola secara terpisah (Gambar 3). Kedua pertanaman
ini dapat dibedakan berdasarkan keanekaragaman spesies tanaman sayuran yang
ada di sekitarnya. Satu pertanaman memiliki keanekaragaman 5-8 spesies
tanaman sayuran, sedangkan pertanaman lain memiliki keanekaragaman 9-15
spesies tanaman sayuran (Tabel 2). Berdasarkan ketersediaan unit pengambilan
contoh selama tiga bulan pengamatan, kubis merupakan komoditas sayuran yang
dapat mewakili kedua pertanaman berdasarkan keanekaragaman tanaman sayuran
di sekitar lahan.
a
b

7
Tabel 2 Keanekaragaman spesies sayuran di sekitar pertanaman polikultur kubis
di Desa Tugu Selatan, Cisarua
5-8 spesies tanaman sayurana 9-15 spesies tanaman sayuran
b
Caisin (Brassica rapa var. parachinensis
L.)
Caisin (Brassica apa var. parachinensis
L.)
Bayam Hijau (Amaranthus hybridus L.) Bayam Hijau (Amaranthus hybridus L.)
Timun (Cucumis sativus L.) Timun (Cucumis sativus L.)
Tomat (Solanum lycopersicum L.) Tomat (Solanum lycopersicum L.)
Buncis (Phaseolus vulgaris L.) Buncis (Phaseolus vulgaris L.)
Selada (Lactuca sativa L.) Selada (Lactuca sativa L.)
Kacang Panjang (Vigna unguiculata
subsp. sesquipedalis L.)
Kacang Panjang (Vigna unguiculata
subsp. sesquipedalis L.)
Wortel (Daucus carota L.) Wortel (Daucus carota L.)
Kemangi (Ocimum citriodorum Vis.)
Kapri (Pisum sativum L.)
Jagung (Zea mays L.)
Brokoli (Brassica oleracea L.)
Terung (Solanum melongena L.)
Kacang Hijau (Vigna radiata L.)
Kacang Merah (Vigna angularis Willd.) a terletak di lokasi TS1
b terletah di lokasi TS5, TS6, TS7
Gambar 3 Pertanaman kubis dengan keanekaragaman (a) 5-8 spesies tanaman
sayuran dan (b) 9-15 spesies tanaman sayuran di sekitar
a b

8
Terung digunakan untuk mewakili sayuran dengan perbedaan umur. Hal ini
karena pertanaman terung di Desa Bojong dan Bantarsari pada awal pengambilan
contoh (Agustus 2012) telah memasuki umur 3 bulan dan memulai fase
generatifnya. Sehingga empat lokasi yang ada di dua desa ini dapat digunakan
untuk melihat serangan hama dan parasitisasi parasitoid selama tiga bulan
pengambilan contoh berturut-turut. Selanjutnya, didapat tiga perbedaan umur
tanaman terung berbeda, yaitu, 3 bulan, 4 bulan dan 5 bulan (Gambar 4). Pada
penelitian ini, kontrol manajemen pengelolaan lahan seperti pemberian pupuk dan
manajemen terhadap hama sengaja tidak diperhatikan untuk melihat perbandingan
antara pertanaman yang dikelola secara organik dan konvensional.
Gambar 4 Pertanaman terung berumur (a) 3 bulan di Desa Bojong dan (b) 4 bulan
di Desa Bantarsari
a
b

9
Pengambilan Contoh Serangga Pengambilan contoh serangga hama dilakukan setiap bulannya dari bulan
Agustus sampai Oktober 2012. Pada setiap lahan dilakukan pengambilan contoh
pada transek sepanjang 50 langkah dengan waktu tiap transek 30 menit. Jenis
hama yang diambil pada penelitian ini meliputi serangga dari ordo Lepidoptera
dan Coleoptera yang merupakan jenis hama yang mendominasi pada pertanaman
sayuran. Telur, larva dan pupa serangga hama yang ditemukan di jalur transek
dikumpulkan dan dimasukan ke dalam wadah untuk kemudian diberi label.
Serangga kemudian dibawa ke laboratorium untuk dipelihara dengan diberi pakan
alami dan dihitung kejadian parasitisasinya.
Identifikasi Serangga
Identifikasi serangga hama dan parasitoid (ordo Hymenoptera dan Diptera)
yang muncul dilakukan di Laboratorium Pengendalian Hayati, hingga ke tingkat
morfospesies berdasarkan struktur morfologi, jenis inang dan parasitoid dengan
mengacu pada buku The Pest of Crops in Indonesia (Kalshoven 1981),
Hymenoptera of the World (Goulet dan Huber 1993) dan Manual of Nearctic
Diptera Volume 2 (McAlpine 1987). Khusus indentifikasi hingga spesies
dilakukan dengan bantuan taksonomis dari LIPI.
Analisis Data
Keanekaragaman, persentase serangan hama dan parasitisasi parasitoid
dihitung secara terpisah berdasarkan jenis komoditas dan kriteria pertanaman yang
dimilikinya. Persentase serangan hama dan parasitisasi parasitoid dihitung dengan
rumus yang dimodifikasi dari Hamid et al. (2003), yaitu:
Pengaruh sistem budidaya terhadap kekayaan spesies, tingkat serangan hama dan
parasitisasi parasitoid diuji dengan analisis nilai tengah (t-test) dan analisis ragam
(One way ANOVA) menggunakan program SPSS 17 untuk Windows (SPSS
2008). Kemiripan komposisi spesies parasitoid antar lokasi dihitung
menggunakan Indeks Sorensen yang memiliki persamaan:
IS : Indeks Kemiripan Spesies
A : Jumlah spesies parasitoid di lahan 1
B : Jumlah spesies parasitoid di lahan 2
C : Jumlah spesies parasitoid yang sama di kedua lahan yang
dibandingkan
Perbedaan komposisi spesies parasitoid antar lahan uji dengan Analysis of
Similarity (ANOSIM) menggunakan perangkat lunak R dengan paket vegan (R
Core Team 2013). Gambar interaksi tropik inang-parasitoid dibuat dengan
perangkat lunak R dengan paket bipartite.
10
HASIL DAN PEMBAHASAN
Keanekaragaman, Serangan Hama dan Parasitisasi Parasitoid pada
Pertanaman Sayuran di Bogor
Pengambilan contoh dengan metode transek garis yang dilakukan pada
penelitian ini menggambarkan kelimpahan relatif serangan hama sekaligus tingkat
kematiannya yang disebabkan oleh parasitoid (parasitism rate) di suatu
agroekosistem pertanian pada rentang waktu tiga bulan pengamatan. Penelitian ini
memperlihatkan bahwa tanaman sayuran famili Brassicaceae (kubis, caisin,
brokoli, sawi) menunjukkan kecenderungan tingkat serangan hama yang lebih
tinggi dibandingkan famili Solanaceae (terung dan tomat) dan Curcubitaceae (pare
dan timun) (Tabel 3). Rendahnya tingkat serangan hama pada famili Solanaceae
dan Curcubitaceae diduga karena adanya dengan gangguan intensif pada skala
agroekosistem yang bersifat letal, yaitu saat digunakannya pestisida sintetik. Hal
serupa juga diduga terjadi pada pertanaman caisin di enam lokasi yang
menunjukkan tingkat serangan hama yang berbeda-beda (Tabel 1). Selain hama,
penggunaan pestisida sintetik juga dapat mengganggu keberadaan musuh alami
serangga seperti parasitoid apabila diaplikasikan terus menerus walaupun dengan
dosis sub-letal (De Cock et al. 1996).
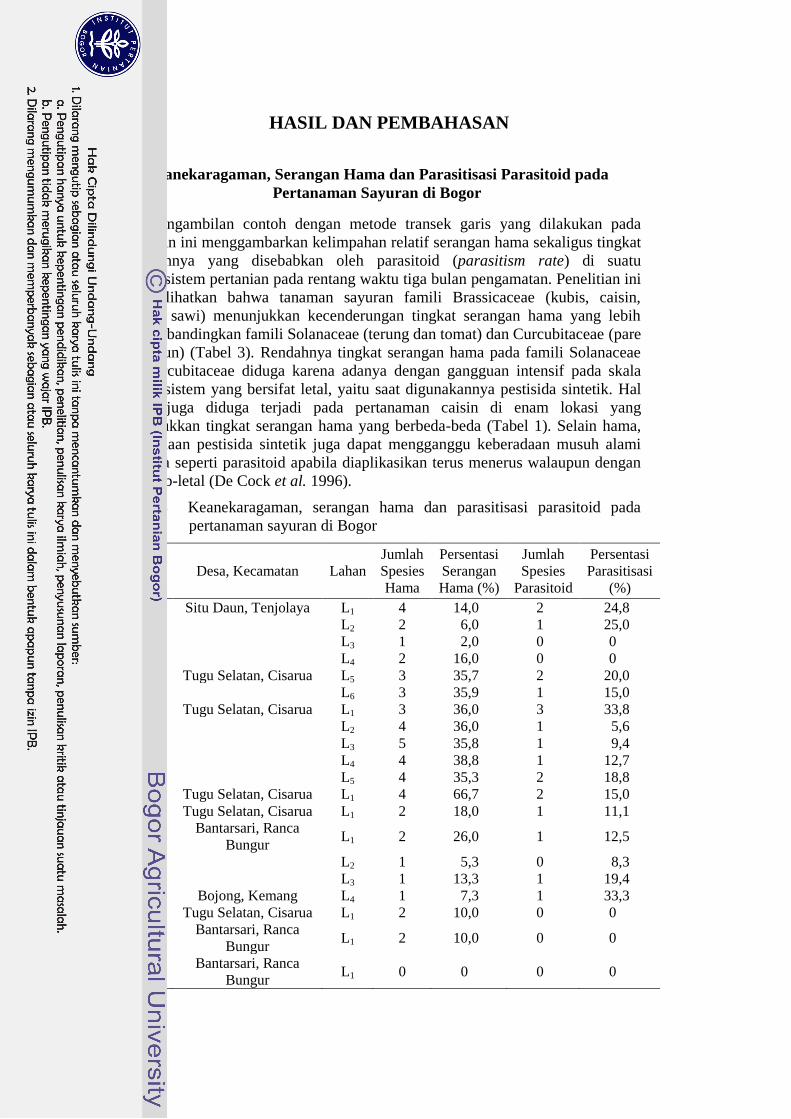
Tabel 3 Keanekaragaman, serangan hama dan parasitisasi parasitoid pada
pertanaman sayuran di Bogor
Komoditas Desa, Kecamatan Lahan
Jumlah
Spesies
Hama
Persentasi
Serangan
Hama (%)
Jumlah
Spesies
Parasitoid
Persentasi
Parasitisasi
(%)
Caisin Situ Daun, Tenjolaya L1 4 14,0 2 24,8
L2 2 6,0 1 25,0
L3 1 2,0 0 0
L4 2 16,0 0 0
Tugu Selatan, Cisarua L5 3 35,7 2 20,0
L6 3 35,9 1 15,0
Kubis Tugu Selatan, Cisarua L1 3 36,0 3 33,8
L2 4 36,0 1 5,6
L3 5 35,8 1 9,4
L4 4 38,8 1 12,7
L5 4 35,3 2 18,8
Brokoli Tugu Selatan, Cisarua L1 4 66,7 2 15,0
Sawi Tugu Selatan, Cisarua L1 2 18,0 1 11,1
Terung Bantarsari, Ranca
Bungur L1 2 26,0 1 12,5
L2 1 5,3 0 8,3
L3 1 13,3 1 19,4
Bojong, Kemang L4 1 7,3 1 33,3
Tomat Tugu Selatan, Cisarua L1 2 10,0 0 0
Pare Bantarsari, Ranca
Bungur L1 2 10,0 0 0
Timun Bantarsari, Ranca
Bungur L1 0 0 0 0
11
Delapan komoditas sayuran yang dipilih di empat desa di wilayah Bogor
menunjukkan tingkat parasitisasi parasitoid yang beragam (Tabel 3). Selain
karena faktor ketersediaan inang dan kompetisi di alam, hal ini diduga karena
dipengaruhi faktor alam lain yang memengaruhi, salah satunya adalah kondisi
habitat sekitar (Tylianakis et al. 2007). Tidak adanya kejadian parasitisasi pada
dua sayuran caisin di Desa Situ Daun, pare dan timun di Desa Bantarsari, serta
tomat di Desa Tugu Selatan diduga karena pengaruh intensifikasi dalam pertanian
(Lohaus et al. 2013).
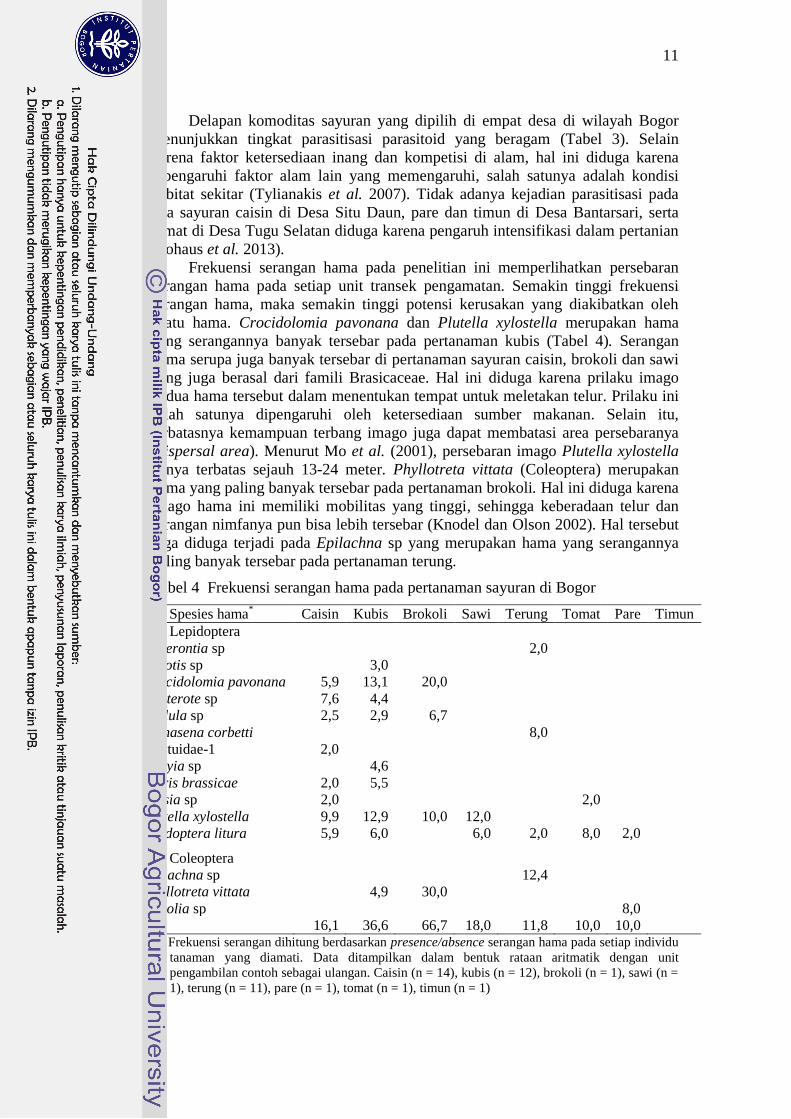
Frekuensi serangan hama pada penelitian ini memperlihatkan persebaran
serangan hama pada setiap unit transek pengamatan. Semakin tinggi frekuensi
serangan hama, maka semakin tinggi potensi kerusakan yang diakibatkan oleh
suatu hama. Crocidolomia pavonana dan Plutella xylostella merupakan hama
yang serangannya banyak tersebar pada pertanaman kubis (Tabel 4). Serangan
hama serupa juga banyak tersebar di pertanaman sayuran caisin, brokoli dan sawi
yang juga berasal dari famili Brasicaceae. Hal ini diduga karena prilaku imago
kedua hama tersebut dalam menentukan tempat untuk meletakan telur. Prilaku ini
salah satunya dipengaruhi oleh ketersediaan sumber makanan. Selain itu,
terbatasnya kemampuan terbang imago juga dapat membatasi area persebaranya
(dispersal area). Menurut Mo et al. (2001), persebaran imago Plutella xylostella
hanya terbatas sejauh 13-24 meter. Phyllotreta vittata (Coleoptera) merupakan
hama yang paling banyak tersebar pada pertanaman brokoli. Hal ini diduga karena
imago hama ini memiliki mobilitas yang tinggi, sehingga keberadaan telur dan
serangan nimfanya pun bisa lebih tersebar (Knodel dan Olson 2002). Hal tersebut
juga diduga terjadi pada Epilachna sp yang merupakan hama yang serangannya
paling banyak tersebar pada pertanaman terung.
Tabel 4 Frekuensi serangan hama pada pertanaman sayuran di Bogor
Spesies hama* Caisin Kubis Brokoli Sawi Terung Tomat Pare Timun
Ordo Lepidoptera
Acherontia sp
2,0
Agrotis sp
3,0
Crocidolomia pavonana 5,9 13,1 20,0
Eupterote sp 7,6 4,4
Hellula sp 2,5 2,9 6,7
Mahasena corbetti
8,0
Noctuidae-1 2,0
Orgyia sp
4,6
Pieris brassicae 2,0 5,5
Plusia sp 2,0
2,0
Plutella xylostella 9,9 12,9 10,0 12,0
Spodoptera litura 5,9 6,0
6,0 2,0 8,0 2,0
Ordo Coleoptera
Epilachna sp
12,4
Phyllotreta vittata
4,9 30,0
Rodolia sp
8,0
Total 16,1 36,6 66,7 18,0 11,8 10,0 10,0
* Frekuensi serangan dihitung berdasarkan presence/absence serangan hama pada setiap individu
tanaman yang diamati. Data ditampilkan dalam bentuk rataan aritmatik dengan unit
pengambilan contoh sebagai ulangan. Caisin (n = 14), kubis (n = 12), brokoli (n = 1), sawi (n =
1), terung (n = 11), pare (n = 1), tomat (n = 1), timun (n = 1)
12
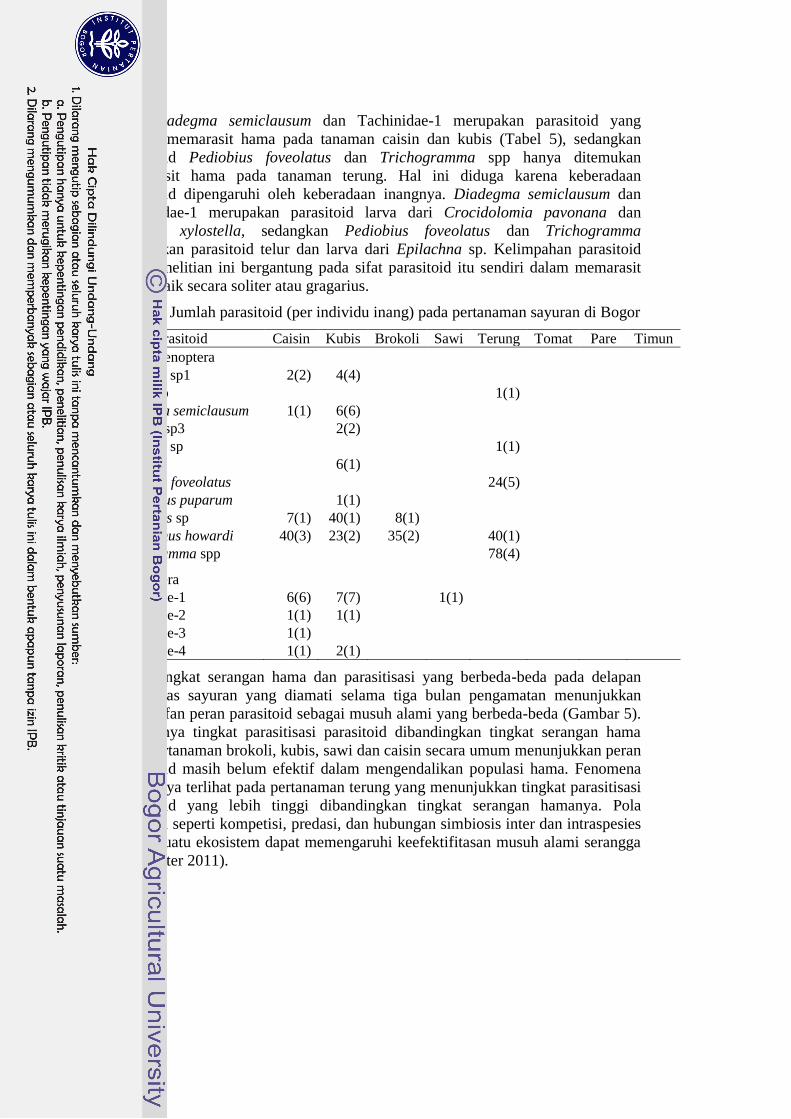
Diadegma semiclausum dan Tachinidae-1 merupakan parasitoid yang
banyak memarasit hama pada tanaman caisin dan kubis (Tabel 5), sedangkan
parasitoid Pediobius foveolatus dan Trichogramma spp hanya ditemukan
memarasit hama pada tanaman terung. Hal ini diduga karena keberadaan
parasitoid dipengaruhi oleh keberadaan inangnya. Diadegma semiclausum dan
Tachinidae-1 merupakan parasitoid larva dari Crocidolomia pavonana dan
Plutella xylostella, sedangkan Pediobius foveolatus dan Trichogramma
merupakan parasitoid telur dan larva dari Epilachna sp. Kelimpahan parasitoid
pada penelitian ini bergantung pada sifat parasitoid itu sendiri dalam memarasit
hama, baik secara soliter atau gragarius.
Tabel 5 Jumlah parasitoid (per individu inang) pada pertanaman sayuran di Bogor
Spesies Parasitoid Caisin Kubis Brokoli Sawi Terung Tomat Pare Timun
Ordo Hymenoptera
Apanteles sp1 2(2) 4(4)
Bracon sp
1(1)
Diadegma semiclausum 1(1) 6(6)
Eriborus sp3 2(2)
Goryphus sp
1(1)
Idris sp 6(1)
Pediobius foveolatus
24(5)
Pteromalus puparum 1(1)
Telenomus sp 7(1) 40(1) 8(1)
Tetrastichus howardi 40(3) 23(2) 35(2)
40(1)
Trichogramma spp
78(4)
Ordo Diptera
Tachinidae-1 6(6) 7(7)
1(1)
Tachinidae-2 1(1) 1(1)
Tachinidae-3 1(1)
Tachinidae-4 1(1) 2(1)
Tingkat serangan hama dan parasitisasi yang berbeda-beda pada delapan
komoditas sayuran yang diamati selama tiga bulan pengamatan menunjukkan
keefektifan peran parasitoid sebagai musuh alami yang berbeda-beda (Gambar 5).
Rendahnya tingkat parasitisasi parasitoid dibandingkan tingkat serangan hama
pada pertanaman brokoli, kubis, sawi dan caisin secara umum menunjukkan peran
parasitoid masih belum efektif dalam mengendalikan populasi hama. Fenomena
sebaliknya terlihat pada pertanaman terung yang menunjukkan tingkat parasitisasi
parasitoid yang lebih tinggi dibandingkan tingkat serangan hamanya. Pola
interaksi seperti kompetisi, predasi, dan hubungan simbiosis inter dan intraspesies
dalam suatu ekosistem dapat memengaruhi keefektifitasan musuh alami serangga
(Showalter 2011).
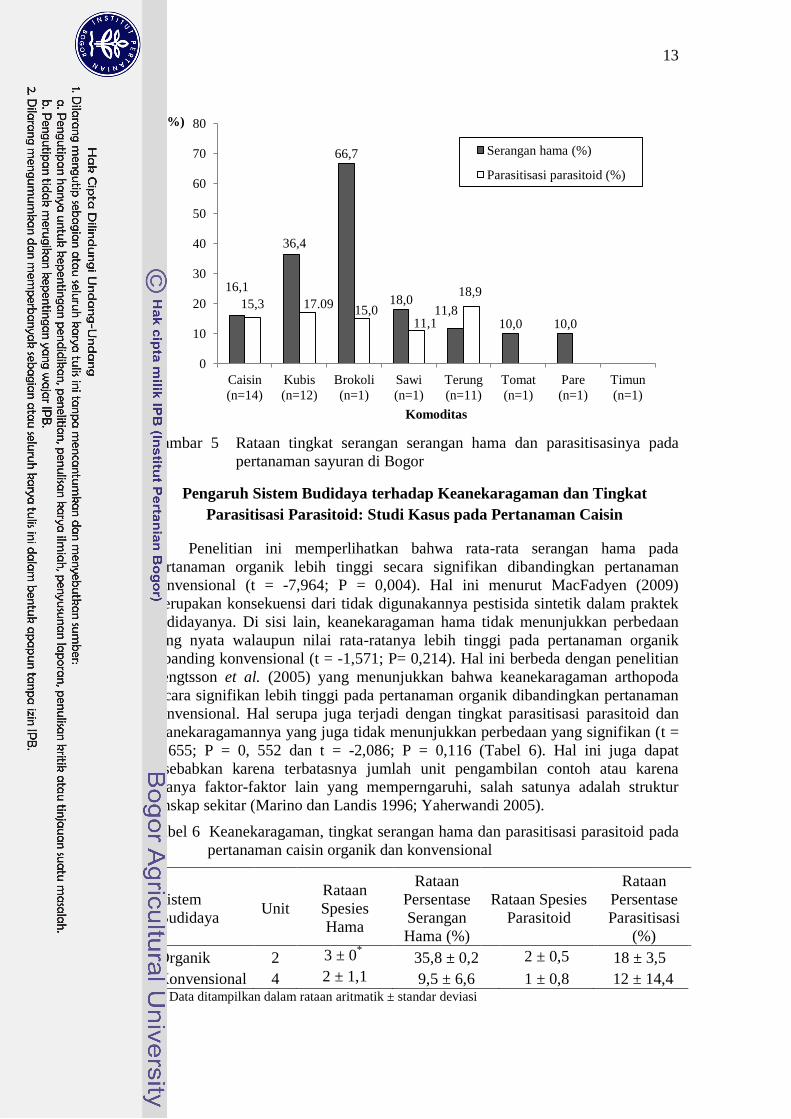
13
Gambar 5 Rataan tingkat serangan serangan hama dan parasitisasinya pada
pertanaman sayuran di Bogor
Pengaruh Sistem Budidaya terhadap Keanekaragaman dan Tingkat
Parasitisasi Parasitoid: Studi Kasus pada Pertanaman Caisin
Penelitian ini memperlihatkan bahwa rata-rata serangan hama pada
pertanaman organik lebih tinggi secara signifikan dibandingkan pertanaman
konvensional (t = -7,964; P = 0,004). Hal ini menurut MacFadyen (2009)
merupakan konsekuensi dari tidak digunakannya pestisida sintetik dalam praktek
budidayanya. Di sisi lain, keanekaragaman hama tidak menunjukkan perbedaan
yang nyata walaupun nilai rata-ratanya lebih tinggi pada pertanaman organik
dibanding konvensional (t = -1,571; P= 0,214). Hal ini berbeda dengan penelitian
Bengtsson et al. (2005) yang menunjukkan bahwa keanekaragaman arthopoda
secara signifikan lebih tinggi pada pertanaman organik dibandingkan pertanaman
konvensional. Hal serupa juga terjadi dengan tingkat parasitisasi parasitoid dan
keanekaragamannya yang juga tidak menunjukkan perbedaan yang signifikan (t =
-0,655; P = 0, 552 dan t = -2,086; P = 0,116 (Tabel 6). Hal ini juga dapat
disebabkan karena terbatasnya jumlah unit pengambilan contoh atau karena
adanya faktor-faktor lain yang memperngaruhi, salah satunya adalah struktur
lanskap sekitar (Marino dan Landis 1996; Yaherwandi 2005).
Tabel 6 Keanekaragaman, tingkat serangan hama dan parasitisasi parasitoid pada
pertanaman caisin organik dan konvensional
Sistem
Budidaya Unit
Rataan
Spesies
Hama
Rataan
Persentase
Serangan
Hama (%)
Rataan Spesies
Parasitoid
Rataan
Persentase
Parasitisasi
(%)
Organik 2 3 ± 0* 35,8 ± 0,2 2 ± 0,5 18 ± 3,5
Konvensional 4 2 ± 1,1 9,5 ± 6,6 1 ± 0,8 12 ± 14,4 *
Data ditampilkan dalam rataan aritmatik ± standar deviasi
16,1
36,4
66,7
18,0 11,8
10,0 10,0
15,3 17.09 15,0
11,1
18,9
0
10
20
30
40
50
60
70
80
Caisin
(n=14)
Kubis
(n=12)
Brokoli
(n=1)
Sawi
(n=1)
Terung
(n=11)
Tomat
(n=1)
Pare
(n=1)
Timun
(n=1)
(%)
Komoditas
Serangan hama (%)
Parasitisasi parasitoid (%)

14
Komposisi spesies parasitoid dengan sistem budidaya yang sama memiliki
nilai indeks kemiripan yang tinggi (Tabel 7). Sedangkan komposisi spesies
parasitoid antar lahan dengan sistem budidaya berbeda menunjukkan rentang nilai
yang cukup luas, yaitu dari 0,73 hingga 0,89. Namun, hasil analisis kemiripan
menggunakan ANOSIM, sistem budidaya juga tidak memiliki pengaruh yang
signifikan terhadap komposisi spesies antar lahan (R = 0,429; P = 0,186).
Tabel 7 Komposisi spesies parasitoid antar lahan dengan sistem pertanian
berbeda berdasarkan indeks kemiripan Sorensen
Lahan* LO1 LO2 LK1 LK2 LK3 LK4
LO1 1 LO2 0,82 1
LK1 0,76 0,73 1 LK2 0,80 0,76 0,82 1
LK3 0,89 0,84 0,84 0,89 1 LK4 0,89 0,84 0,84 0,84 1 1
* LO = lahan organik; LK = lahan konvensional
Interaksi tropik memperlihatkan hubungan makan-memakan suatu
organisme antar level tropik. Hubungan tropik inang-parasitoid pada kedua sistem
budidaya memperlihatkan kompleksitas struktur interaksi yang hampir sama,
walaupun memiliki komposisi parasitoid yang berbeda (Gambar 6). Pada pola
interaksi tersebut diketahui bahwa parasitoid Tachinidae-1 dan Tetrastichus
howardi mendominasi kompetisi dengan memarasit lebih satu inang. Menurut
Hawkin (1994) dalam suatu komunitas inang-parasitoid, hanya ada satu atau
beberapa spesies parasitoid saja yang memiliki pengaruh besar dalam perannya
sebagai agen pengendali hayati. Hal ini disebabkan karena adanya perbedaan
kapabilitas pencarian inang tertentu oleh parasitoid di setiap habitat berbeda pada
kondisi dan waktu tertentu. Walaupun begitu, interaksi inter dan intraspesies pada
masing-masing level tropik (inang-parasitoid) pada penelitian ini masih belum
begitu diketahui.
Gambar 6 Hubungan tropik herbivor (hama) dan parasitoidnya pada (a)
pertanaman caisin organik dan (b) pertanaman caisin konvensional
N = 9
N = 46
N = 7
N = 32
Parasitoid
Herbivor
1 6 7 1 6 7
8 19 20 21 18 16 10 17
1 C. pavonana, 6 Plutella xylostella, 7 Spodoptera litura
8 Apanteles sp1, 10 D. semiclausum, 16 Tachinidae-1, 17 Tachinidae-2, 18 Tachinidae-3
19 Tachinidae-4, 20 Telenomus sp, 21 Tetrastichus howardi
(a) Organik (b) Konvensional
15
Pengaruh Keanekaragaman Tanaman Sayuran di Sekitar terhadap
Keanekaragaman dan Tingkat Parasitisasi Parasitoid: Studi Kasus pada
Pertanaman Polikultur Kubis
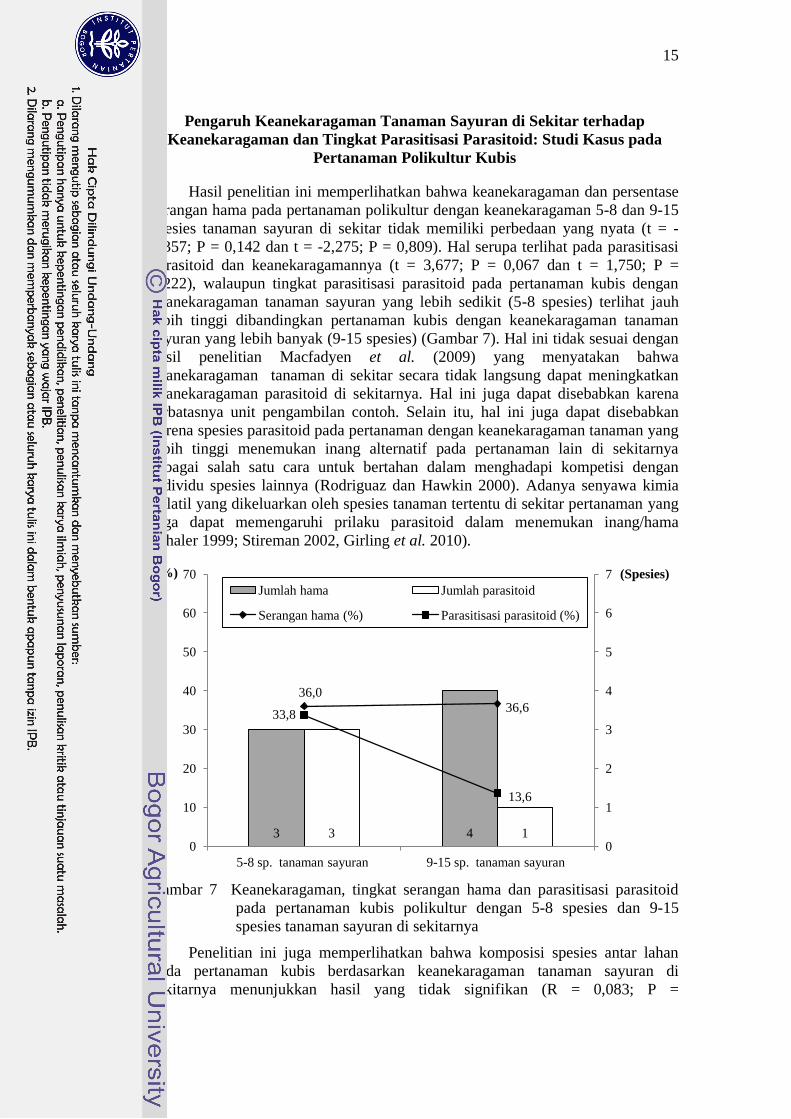
Hasil penelitian ini memperlihatkan bahwa keanekaragaman dan persentase
serangan hama pada pertanaman polikultur dengan keanekaragaman 5-8 dan 9-15
spesies tanaman sayuran di sekitar tidak memiliki perbedaan yang nyata (t = -
2,357; P = 0,142 dan t = -2,275; P = 0,809). Hal serupa terlihat pada parasitisasi
parasitoid dan keanekaragamannya (t = 3,677; P = 0,067 dan t = 1,750; P =
0,222), walaupun tingkat parasitisasi parasitoid pada pertanaman kubis dengan
keanekaragaman tanaman sayuran yang lebih sedikit (5-8 spesies) terlihat jauh
lebih tinggi dibandingkan pertanaman kubis dengan keanekaragaman tanaman
sayuran yang lebih banyak (9-15 spesies) (Gambar 7). Hal ini tidak sesuai dengan
hasil penelitian Macfadyen et al. (2009) yang menyatakan bahwa
keanekaragaman tanaman di sekitar secara tidak langsung dapat meningkatkan
keanekaragaman parasitoid di sekitarnya. Hal ini juga dapat disebabkan karena
terbatasnya unit pengambilan contoh. Selain itu, hal ini juga dapat disebabkan
karena spesies parasitoid pada pertanaman dengan keanekaragaman tanaman yang
lebih tinggi menemukan inang alternatif pada pertanaman lain di sekitarnya
sebagai salah satu cara untuk bertahan dalam menghadapi kompetisi dengan
individu spesies lainnya (Rodriguaz dan Hawkin 2000). Adanya senyawa kimia
volatil yang dikeluarkan oleh spesies tanaman tertentu di sekitar pertanaman yang
juga dapat memengaruhi prilaku parasitoid dalam menemukan inang/hama
(Thaler 1999; Stireman 2002, Girling et al. 2010).
Gambar 7 Keanekaragaman, tingkat serangan hama dan parasitisasi parasitoid
pada pertanaman kubis polikultur dengan 5-8 spesies dan 9-15
spesies tanaman sayuran di sekitarnya
Penelitian ini juga memperlihatkan bahwa komposisi spesies antar lahan
pada pertanaman kubis berdasarkan keanekaragaman tanaman sayuran di
sekitarnya menunjukkan hasil yang tidak signifikan (R = 0,083; P =
3 4 3 1
36,0
36,6 33,8
13,6
0
1
2
3
4
5
6
7
0
10
20
30
40
50
60
70
5-8 sp. tanaman sayuran 9-15 sp. tanaman sayuran
(Spesies) (%)
Jumlah hama Jumlah parasitoid
Serangan hama (%) Parasitisasi parasitoid (%)

16
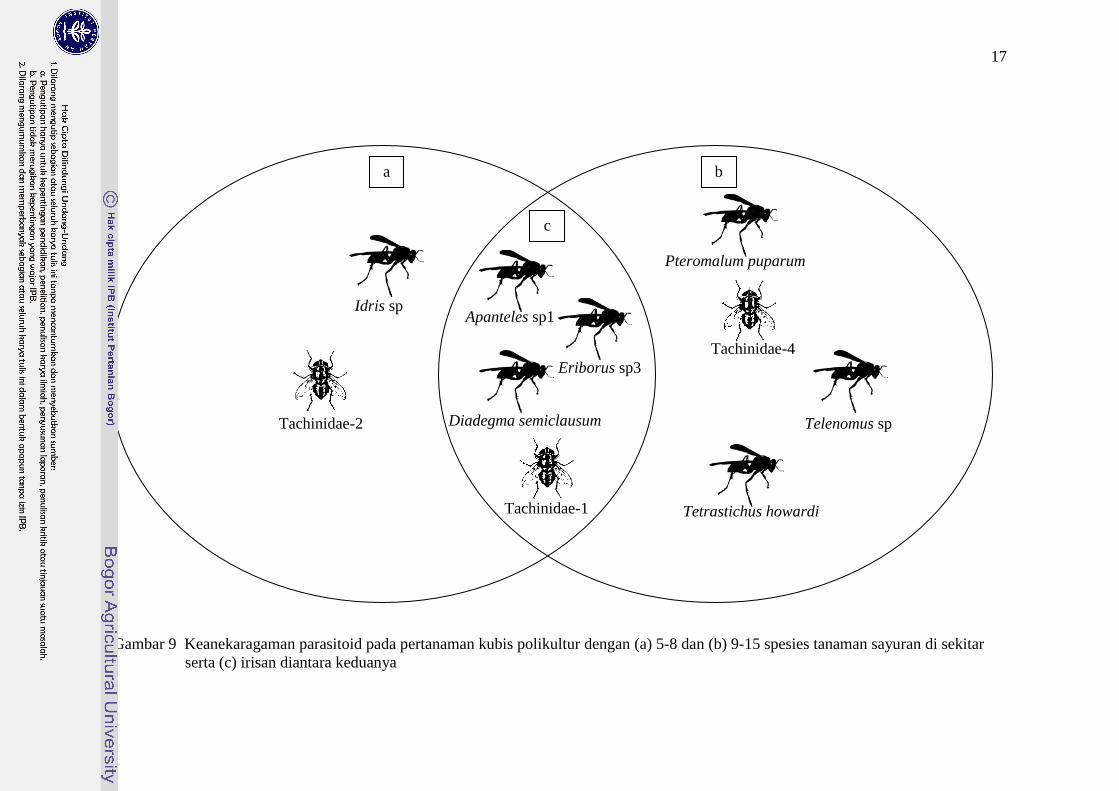
0,499). Terdapat beberapa speseis parasitoid yang ditemukan pada kedua
kelompok pertanaman kubis, yaitu: Apanteles sp1, Diadegma semiclausum,
Eriborus sp3 dan Tachinidae-1 (Gambar 9). Hal ini diduga karena adanya
persamaan jenis sayuran yang ada pada kedua pertanaman kubis, yaitu: caisin,
bayam hijau, timun, tomat, buncis, selada, kacang panjang dan wortel (Tabel 2).
Namun, ada juga spesies parasitoid yang hanya ditemukan pada pertanaman kubis
dengan keanekaragaman 5-8 spesies atau 9-15 spesies tanaman sayuran di
sekitarnya saja. Idris sp dan Tachinidae-2 merupakan speseis parasitoid yang
hanya ditemukan pada pertanaman kubis dengan keanekaragaman 5-8 spesies
tanaman sayuran di sekitarnya, sedangkan Pteromalum puparum, Tachinidae-4,
Telenomus sp dan Tetrastichus howardi merupakan speseis parasitoid yang hanya
ditemukan pada pertanaman kubis dengan keanekaragaman 9-15 speseis tanaman
sayuran di sekitarnya. Hal ini diduga karena adanya pengaruh tanaman atau
vegetasi yang ada di sekitar pertanaman yang memengaruhi perilaku pencarian
inang/hama serangga parasitoid.
Struktur interaksi tropik pada pertanaman dengan keanekaragaman tanaman
dengan 9-15 spesies tanaman sayuran disekitarnya terlihat lebih kompleks
dibandingkan dengan pertanaman dengan keanekaragaman tanaman dengan 5-8
spesies tanaman sayuran disekitarnya (Gambar 8). Hal ini dapat dilihat dari
jumlah spesies yang terlibat dan tingkat parasitisasi yang terjadi di kedua jaring-
jaring makanan. Pola interaksi ini memperlihatkan bahwa Diadegma semiclausum
lebih mendominasi persaingan dengan 3 spesies parasitoid lainnya dalam
memarasit Plutella xylostella. Sedangkan Tachinidae-1 bertahan dalam kompetisi
dengan memarasit dua spesies inang berbeda. Menurut Althof (2003), strategi
parasitisasi oleh suatu individu spesies parasitoid dapat memengaruhi spesifisitas
inang, di mana parasitoid koinobiont memiliki inang yang lebih spesifik dari pada
parasitoid idiobiont. Parasitoid koinobion biasanya membiarkan inang tumbuh
lebih lanjut setelah proses parasitisasi, sedangkan parasitoid idiobiont tidak. Hal
ini nantinya dapat memengaruhi tingkat parasitisasi suatu spesies parasitoid saat
berkompetisi dengan spesies lainnya dalam suatu komunitas.
Gambar 8 Hubungan tropik herbivor (hama) dan parasitoidnya pada pertanaman
kubis polikultur dengan (a) 5-8 spesies dan (b) 9-15 spesies tanaman
sayuran di sekitarnya
1 C. pavonana 4 Phyllotreta vittata, 5 Pieris brassicae, 6 Plutella xylostella, 7 Spodoptera litura
8 Apanteles sp1, 10 D. semiclausum, 11 Eriborus sp3, 13 Idris sp, 15 Pteromalum puparum
16 Tachinidae-1, 17 Tachinidae-2, 19 Tachinidae-4, 20 Telenomus sp, 21 Tetrastichus howardi
(b) 9-15 spesies tanaman sayuran (a) 5-8 spesies tanaman sayuran
8 10 11 15 16 19 20 21 8 10 1
1
13 16 17
Parasitoid
N = 16 N = 9
N = 93 N = 24
Herbivor
7 6 5 1 1 6 4
N = 9
N = 24
4
17
Gambar 9 Keanekaragaman parasitoid pada pertanaman kubis polikultur dengan (a) 5-8 dan (b) 9-15 spesies tanaman sayuran di sekitar
serta (c) irisan diantara keduanya
a
Apanteles sp1
Diadegma semiclausum
Eriborus sp3
Tachinidae-1
Idris sp
Tachinidae-2
Pteromalum puparum
Tachinidae-4
Telenomus sp
Tetrastichus howardi
b
c
18
Pengaruh Perbedaan Umur Tanaman terhadap Keanekaragaman dan
Tingkat Parasitisasi Parasitoid: Studi Kasus pada Pertanaman Terung
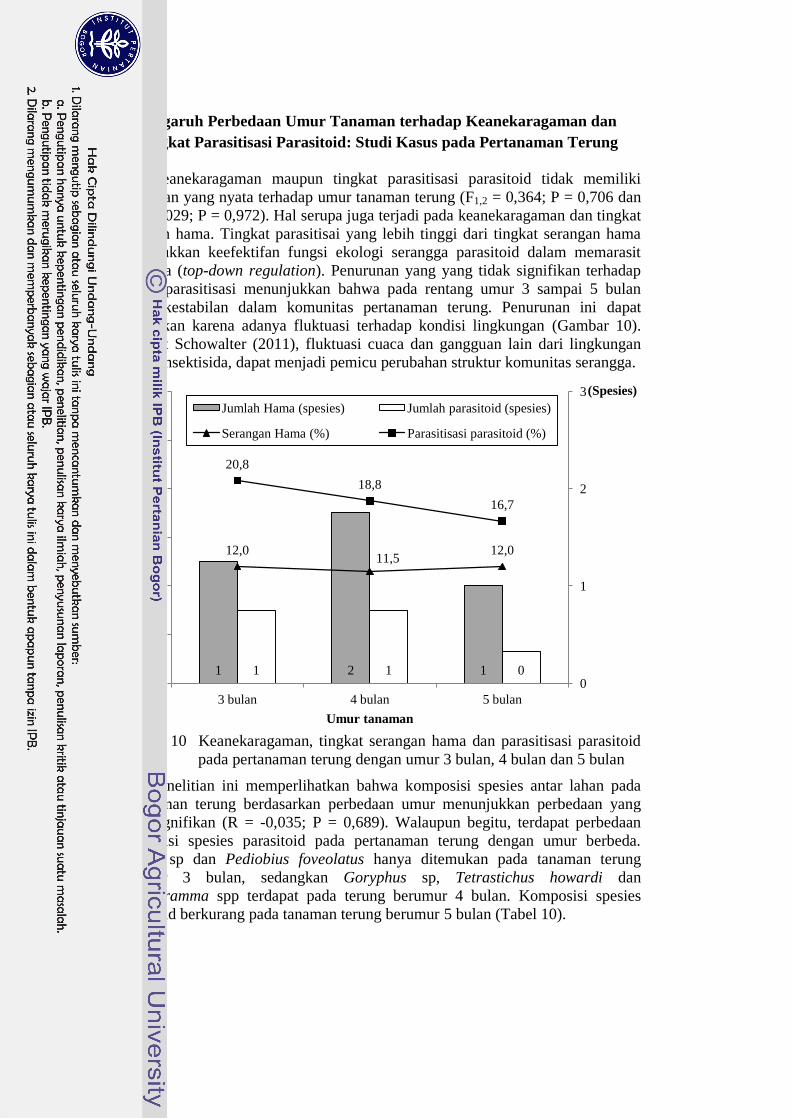
Keanekaragaman maupun tingkat parasitisasi parasitoid tidak memiliki
perbedaan yang nyata terhadap umur tanaman terung (F1,2 = 0,364; P = 0,706 dan
F1,2 = 0,029; P = 0,972). Hal serupa juga terjadi pada keanekaragaman dan tingkat
serangan hama. Tingkat parasitisai yang lebih tinggi dari tingkat serangan hama
menunjukkan keefektifan fungsi ekologi serangga parasitoid dalam memarasit
inangnya (top-down regulation). Penurunan yang yang tidak signifikan terhadap
tingkat parasitisasi menunjukkan bahwa pada rentang umur 3 sampai 5 bulan
terjadi kestabilan dalam komunitas pertanaman terung. Penurunan ini dapat
disebabkan karena adanya fluktuasi terhadap kondisi lingkungan (Gambar 10).
Menurut Schowalter (2011), fluktuasi cuaca dan gangguan lain dari lingkungan
seperti insektisida, dapat menjadi pemicu perubahan struktur komunitas serangga.
Gambar 10 Keanekaragaman, tingkat serangan hama dan parasitisasi parasitoid
pada pertanaman terung dengan umur 3 bulan, 4 bulan dan 5 bulan
Penelitian ini memperlihatkan bahwa komposisi spesies antar lahan pada
pertanaman terung berdasarkan perbedaan umur menunjukkan perbedaan yang
tidak signifikan (R = -0,035; P = 0,689). Walaupun begitu, terdapat perbedaan
komposisi spesies parasitoid pada pertanaman terung dengan umur berbeda.
Bracon sp dan Pediobius foveolatus hanya ditemukan pada tanaman terung
berumur 3 bulan, sedangkan Goryphus sp, Tetrastichus howardi dan
Trichogramma spp terdapat pada terung berumur 4 bulan. Komposisi spesies
parasitoid berkurang pada tanaman terung berumur 5 bulan (Tabel 10).
1 2 1 1 1 0
12,0 11,5
12,0
20,8
18,8
16,7
0
1
2
3
0
5
10
15
20
25
30
3 bulan 4 bulan 5 bulan
(Spesies) (%)
Umur tanaman
Jumlah Hama (spesies) Jumlah parasitoid (spesies)
Serangan Hama (%) Parasitisasi parasitoid (%)

19
Tabel 8 Kelimpahan hama dan parasitoid pada pertanaman terung dengan umur 3,
4 dan 5 bulan
Spesies Serangga Umur tanaman
3 bulan 4 bulan 5 bulan
Hama Acherontia sp
2
Epilachna sp 22 16 18
Mahasena corbetti
4
Spodoptera litura 2 1
Parasitoid
Bracon sp 1
Goryphus sp
1
Pediobius foveolatus 24
Tetrastichus howardi
40
Trichogramma spp
40 38
Hubungan tropik inang-parasitoid selama 3 bulan pengamatan pada
pertanaman terung memperlihatkan struktur interaksi tropik yang sederhana
(Gambar 11). Hal ini dilihat dari sedikitnya spesies yang terlibat dan pola interaksi
yang terbentuk. Terdapat perubahan terhadap komposisi spesies pada dua level
tropik. Walaupun begitu, belum diketahui bagaimana cuaca dan faktor alam
lainnya memengaruhi setiap individu spesies hama dan parasitoid dalam
penelitian ini.
Gambar 11 Hubungan tropik herbivor (hama) dan parasitoidnya pada pertanaman
terung dengan umur (a) 3 bulan, (b) 4 bulan dan (c) 5 bulan
2 Epilachna sp, 3 Mahasena corbetti, 7 Spodoptera litura
9 Bracon sp, 12 Goryphus sp, 14 Pediobius foveolatus, 21 Tetrastichus howardi, 22 Trichogramma sp
Parasitoid Parasitoid
Herbivor Herbivor
9 14 12 21 22 22 (a) 3 bulan (b) 4 bulan (c) 5 bulan
7 2 2 3 2
N = 6
N = 24
N = 5
N = 20 N = 18
N = 1

20
SIMPULAN DAN SARAN
Berdasarkan pola budidayanya, pertanaman organik dan konvensional
memiliki tingkat parasitisasi yang tidak jauh berbeda. Pada kedua jenis
pertanaman ini, perbedaan komposisi spesies parasitoid antar lahan tidak
menunjukkan perbedaan yang signifikan. Pertanaman polikultur dengan
keanekaragaman tanaman yang lebih rendah (5-8 sp.) memiliki tingkat parasitisasi
yang lebih tinggi dibanding pertanaman polikultur dengan keanekaragaman
tanaman yang lebih tinggi (9-15 sp.). Walaupun demikian, diantara keduanya
tidak menunjukkan perbedaan yang signifikan. Pertanaman terung berdasarkan
perbedaan umur juga tidak menunjukkan perbedaan yang signifikan. Pada rentang
umur 3-5 bulan, tingkat parasitisasi parasitoid menunjukkan nilai lebih tinggi
dibandingkan tingkat parasitisasi hamanya.
Untuk lebih memahami pengaruh agroekosistem terhadap keefektifan
pengendalian hayati, diperlukan penelitian lebih lanjut terkait faktor-faktor lain
yang dapat memengaruhi tingkat parasitisasi dan keanekaragaman parasitoid
dengan unit pengambilan contoh yang lebih banyak. Selain itu, diperlukan juga
studi lebih dalam terhadap struktur jaring-jaring makanan dalam suatu ekosistem
dan memahami interaksi yang terjadi di dalamnya untuk mengetahui peranan
setiap individu spesies dalam proses parasitisasi serangga hama.

21
DAFTAR PUSTAKA
[FAO] Food and Agriculture Organization of the United Nations. 2013.
FAOSTAT database. Tersedia pada: http://faostat3.fao.org/ home/ index.
Html #DOWNLOAD.
Althoff DM. 2003. Does parasitoid attack strategy influence host specificity? A
test with New World braconids. Ecological Entomology [Internet]. [diunduh
2013 Jul 3]; 28:500–502. Tersedia pada: http://althofflab .syr. edu/
_reprints/Althoff03.pdf.
Altieri MA. 1989. Agroecology: a new research and development paradigm for
word agriculture. Agriculture, Ecosystems, and Environment [Internet].
[diunduh 2013 Jul 17]; 27 (1989):37-46. Tersedia pada:
http://www.agroecologie.be/img/download/Altieri1989_agroecology_a_new
_research.pdf.
Barron MC, Wratten SD, Barlow ND. 2004. Phenology and parasitism of the red
admiral butterfly Bassaris gonerilla (Lepidoptera: Nymphalidae). New
Zealand Journal of Ecology [Internet]. [diunduh 2013 Apr 29]; 28(1): 105-
111. Tersedia pada: http://www.nzes.org.nz/nzje/free_issues/
NZJEcol28_1_105.pdf.
Bengtsson J, Anhstrom J, Weibull AC. 2005. The effects of organic agriculture on
biodiversity and abundance: a meta-analysis. Journal of Applied Ecology
[Internet]. [diunduh 2013 Jul 3]; 42:261–269. Tersedia pada:
http://www.bgu.ac.il/desert_agriculture/Agroecology/Reading/ Bengtsson05
.pdf. doi: 10.1111/j.1365-2664.2005.01005.x.
Benrey B, Denno RF. 1997. The slow-growth-high-mortality hypothesis: A test
using the cabbage butterfly. Ecology [Internet]. [diunduh 2013 Sep 5];
78(4), 987-999. Tersedia pada: http://www.jstor.org/
discover/10.2307/2265852?uid=3738224&uid=2129&uid=2&uid=70&uid=
4&sid=21102607923547.
Buchori D, Sahari B, Nurindah. 2008. Conservation of Agroecosystem through
Utilization of Parasitoid Diversity: Lesson for Promoting Sustainable
Agriculture and Ecosystem Health. Hayati [Internet]. [diunduh 2013 Apr
29]; 15:165–172. Tersedia pada: http://core.kmi. open.ac.uk/ display
/5516350.
Clausen CP. 1978. Introduced Parasites and Predators of Arthropod Pests and
Weeds: A World Review. Washington (US) : USDA.
De Cock A, DeClercq P, Tirry L, Degheele O. 1996. Toxicity of Diafenthiuron
and imidacloprid to the predatory bug Podisus maculiventris (Heteroptera:
pentatomidae). Environmental Entomology [Internet]. [diunduh 2013 Sep
3]; 25:476-480. Tersedia pada: http://link.springer.com/ article/
10.1007%2FBF02769828.
Girling RD, Stewart-Jones A, Dherbecourt J, Staley JT, Wright DJ, Poppy GM.
2010. Parasitoids select plants more heavily infested with their caterpillar
hosts: a new approach to aid interpretation of plant headspace volatiles.
Proceedings of Royal Society B [Internet]. [diunduh 2013 Jul 3]; Tersedia
pada: http://rspb.royalsocietypublishing.org/content/early/2011/ 01/21/rspb.
2725:1-8. full. doi:10.1098/rspb.2010.2725.

22
Godfray HCJ. 1994. Parasitoids. Behavioral and Evolutionary Ecology. New
Jersey (US): Princeton University Press.
Gordh G, Legner EF, Caltagirone LE. 1999. Biology of parasitic Hymenoptera. Di
dalam Bellows TS, Fisher TW, editor. Handbook of Biological Control:
Principles and Applications of Biological Control. San Diego (US):
Academic Press.
Goulet H, Huber JT. 1993. Hymenoptera of the World. An Identification Guide to
Families. Ottawa (CA): Agriculture Canada.
Hamid H, Buchori D, Triwidodo H. 2003. Keanekaragaman Parasitoid dan
Parasitisasinya pada Pertanaman Padi di Kawasan Taman Nasional Gunung
Halimun. Hayati [Internet]. [diunduh 2013 Jul 3]; 10:85–90. Tersedia pada:
http://repository.ipb.ac.id/bitstream/handle/123456789/9204/Hasmiandy_Ha
mid_keanekaragaman.pdf.
Hawkins BA. 1994. Pattern and Process in Host-Parasitoid Interaction.
Cambridge (UK): Cambridge University Press.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Jakarta (ID): PT Ichtiar
Baru Van Hoeve.
Knodel JJ, Olson DL. 2002. Crucifer Flea Beetle Biology and Integrated Pest
Management in Canola. North Dakota (US). North Dakota State University.
[Internet]. [diunduh 2013 Sep 3]. Tersedia pada: http://www.ag.ndsu.
edu/pubs/plantsci/pests/e1234.pdf.
Lafferty KD. 2012. Biodiversity loss decreases parasite diversity: theory and
patterns. Phil. Trans. Proceedings of Royal Society B [Internet]. [diunduh
2013 Jul 15]; 367: 2814–2827. Tersedia pada:
http://rstb.royalsocietypublishing .org /content/367/1604/2814.full. doi: 10.
1098 / rstb.2012. 0110.
Letourneau DK, Altieri MA. 1999. Biology of parasitic Hymenoptera. Di dalam
Bellows TS, Fisher TW, editor. Handbook of Biological Control: Principles
and Applications of Biological Control. San Diego (US): Academic Press.
Letourneau DK, Bothwell SG. 2002. Comparison of organic and conventional
farms: challenging ecologists to make biodiversity functional. The
Ecological Society of America [Internet]. [diunduh 2013 Apr 29]; 430-438.
Tersedia pada: http://www.esajournals.org/doi/abs/ 10.1890/ 070081?j
ournalCode=fron.
Lohaus K, Vidal S, Theis C. 2013. Farming practices change food web structures
in cereal aphid-parasitoid-hyperparasitoid communities. Oecologia
[Internet]. [diunduh 2013 Feb 28]; 171:249-259. Tersedia pada: http:/
link.springer.com /article/ 10.1007%2Fs00442-012-2387-8. doi:10. 1007/
s00442-012-2387-8.
Losey JE, Vaughan M. 2006. The economic value of ecological services provided
by insects. BioScience [Internet]. [diunduh 2013 Jan 28]; 56(4):311-323.
Tersedia pada: http://www.xerces.org/ wp-content/ uploads/ 2008/
09/economic_value_ insects.pdf.

23
Macfadyen S, Gibson R, Polaszek A, Morris RJ, Craze PG, Planque R,
Symondson WOC, Memmott J. 2009. Do differences in food web structure
between organic and conventional farms affect the ecosystem service of pest
control?. Ecology Letters [Internet]. [diunduh 2013 Apr 28]; 12:229-238.
Tersedia pada: http://www.insectecology.com/wordpress/wp-content/
uploads/2009/07/Macfad yenetal2009EcolLetters.pdf. doi: 10.1111/j.1461-
0248. 2008.01279.x.
Mattson PC, Altieri M A, Gagne WC. 1984. Modification of small farmer practice
for better pest management. Annual Review of Entomology [Internet].
[diunduh 2013 Jul 17]; 29:303-402. Tersedia pada:
http://www.annualreviews.org/doi/pdf/10.1146/annurev.en.29.010184.0021
23.
Marino PC, Landis DA. 1996. Effect of Landscape structure on parasitoid
diversity and parasitism in agroecosystems [Internet]. Ecology Application
[diunduh pada 2013 Apr 29]; 6(1):276-284. Tersedia pada:
http://www.landislab.ent.msu.edu/pdf/ Landis% 20PDF%20Collection/
63.Marino.Landis.1996.Effects%20of%20 Landscape%20Structure%20on%
20Parasitoid%20Diversity%20and%20Parasitism%20in%20Agroecosystem
s.pdf.
McAlpine JF. 1987. Manual of Nearctic Diptera Volume 2. Ottawa (CA):
Research Branch Agriculture Canada.
Menalled FD, Marino PC, Gage SH dan Landis DA. 1999. Does agricultural
landscape structure affect parasitism and parasitoid diversity?. Ecology
Application [Internet]. [diunduh 2012 Des 23]; 9(2):634-641. Tersedia pada:
http://www.landsat.org/publications/pdfs_ps.
Mo J, Baker G, Keller M, Roush R. 2001. Estimation of some characteristic
dispersal ranges of diamondback moth (Plutella xylostella) (Lepidoptera:
Plutellidae). Di dalam: Endersby NM, Ridland PM, editor. The management
of diamondback moth and other crucifer pests. Proceedings of the 4th
International Workshop; 2001 Nov; Melbourne, Australia. Melbourne
(AU): Department of Primary Industries. hlm. 107-114 [diunduh 2012 Des
13]; Tersedia pada: http://web.entomology.cornell. edu/Shelton
/diamondback-moth/pdf/2001papers/2001DBM14.pdf.
Moreira X, Mooney KA. 2013. Influence of plant genetic diversity on interactions
between higher trophic levels. Biology Letters [Internet]. [diunduh 2013
Aug 31]; 9 (3):1-4. Tersedia pada: http://www.ncbi.nlm.
nih.gov/pubmed/23485879. doi: 10.1098/rsbl.2013.0133.
Norman MJT. 1979. Annual Cropping Systems in The Tropics: An Introduction.
Gainesville (US): University Press.
R Development Core Team. 2013. R: A language and environment for statistical
computing. Vienna (AT). R Foundation for Statistical Computing. Tersedia
pada: http://www.R-project.org.
Ridgway RL, King EG, Carfillo JL. 1976. Augmentation of natural enemies for
control of plant pests in the western hemisphere. Di dalam: Ridgway RL,
Vinson SB, editor. Biological control by augmentation of natural enemies
New York (USA): Plenum Press. hlm. 379-416. [diunduh 2013 Sep 5].
Tersedia pada: http://link.springer.com/content/pdf/10.1007/978-1-4684-
2871-1_13.pdf

24
Riehl LA, Brooks RF, McCoy CW, Fisher TW, Dean HA. 1980.
Accomplishments toward improving integrated pest management for citrus.
Di dalam: Huffaker CB, editor. New technology of pest control. New York
(USA): John Wiley, & Sons. hlm. 319-363.
Rodriguez MA, Hawkins BA. 2000. Diversity, function and stability in parasitoid
communities. Ecology letters [Internet]. [diunduh 2013 Jul 2]; 3:35-40.
Tersedia pada: http://www2.uah.es/ marodriguez/ MARodriguez_Papers/
Rodriguez_&_ Hawkins_ 2000_EcoLetts.pdf.
Schowalter TD. 2011. Insect Ecology. An Ecosystem Approach. Edisi ke-3. San
Diego (US): Academic Press.
Stireman JO. 2002. Host location and selection cues in a generalist tachinid
parasitoid. Entomologia Experimentalis et Applicata [Internet]. [diunduh
2013 Jul 2]; 103: 23–34. Tersedia pada: http://www. wright.edu /~john.
stireman/ StiremanEntexpAppl.pdf.
Stireman JO, Nason JD, Heard S. 2005. Host-associated genetic differentiation in
phytophagous insects: General phenomenon or isolated exceptions?
evidence from a goldenrod-insect community. Evolution [Internet].
[diunduh 2013 Sep 5]; 59:2573-2587. Tersedia pada:
http://stiremanlab.files.wordpress.com/2012/07/stiremanetal2005evolution.p
df.
SPSS Inc. 2008. SPSS Base 17.0 for Windows User's Guide. Chicago (US). SPSS
Inc.
Thaler JS. 1999. Jasmonate-inducible plant defences cause increased parasitismof
herbivores. Nature [Internet]. [diunduh 2013 Jul 3]; 399:696-688. Tersedia
pada: http:// purple.niagara.edu/wje/Bio123/Thaler%201999%20Plant%20
defenses.pdf.
Tylianakis JM, Tscharntke T, dan Lewis OT. 2007. Habitat modification alters the
structure of tropical host-parasitoid food webs. Nature [Internet]. [diunduh
2012 Des 10]; 445:202-205. Tersedia pada: http://www.nature.com/
nature/journal/v445/n7124/abs/nature05429.html.
Yaherwandi. 2005. Keanekaragaman Hymenoptera parasitoid pada beberapa tipe
lanskap pertanian di Daerah Aliran Sungai (DAS) Cianjur, Kabupaten
Cianjur, Jawa Barat [disertasi]. Bogor (ID): Institut Pertanian Bogor.

25
RIWAYAT HIDUP
Penulis yang bernama Muhamad Nurhuda Nugraha, dilahirkan di kota
Rangkas Bitung pada tanggal 25 Oktober 1989 dan tumbuh besar di Pandeglang,
Banten sebagai anak ketiga dari tiga bersaudara dari pasangan Didi Atmaja dan
Enok Purnama. Penulis lulus dari MTsN I Pandeglang pada tahun 2004 dan lulus
dari SMAN I Pandeglang lalu diterima sebagai mahasiswa Departemen Proteksi
Tanaman, Institut Pertanian Bogor pada tahun 2007 melalui jalur USMI.
Selama menempuh studi di IPB, Penulis aktif di lembaga kerohanian FKRD
(Forum Komunikasi Rohis Departemen) dan beberapa kepanitiaan dan organisasi
lainnya di kampus. Pada tahun 2010 Penulis mengikuti mobility program dari
SEAMEO RIHED untuk belajar selama satu semester di departemen Tropical
Agriculture International Program, Kasetsart University, Thailand dalam skema
MIT Kredit Transfer Mahasiswa. Pada tahun 2011, Penulis juga mendapatkan
beasiswa dari Erasmus Mundus: EXPERTS untuk belajar di Fakultas Pertanian
selama satu semester di Goettingen University, Jerman. Selama menempuh
program sarjana di IPB, Penulis mengikuti beberapa pelatihan, diantaranya adalah
Pelatihan Penggunaan R Statistic, Pelatihan Penggunaan GPS & Quantum GIS
serta Training in Rapid Biodiversity Assessment 2013.




















