Jawaban No.3
28
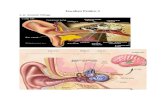
Radang (bahasa Inggris : inflammation) adalah rangkaian reaksi yang terjadi pada tempat jaringan yang mengalami cedera , seperti karena terbakar, atau terinfeksi. Radang atau inflamasi adalah satu dari respon utama sistem kekebalan terhadap infeksi dan iritasi . Inflamasi distimulasi oleh faktor kimia (histamin , bradikinin , serotonin , leukotrien , dan prostaglandin ) yang dilepaskan oleh sel yang berperan sebagai mediator radang di dalam sistem kekebalan untuk melindungi jaringan sekitar dari penyebaran infeksi. Radang mempunyai tiga peran penting dalam perlawanan terhadap infeksi: [1] memungkinkan penambahan molekul dan sel efektor ke lokasi infeksi untuk meningkatkan performa makrofaga menyediakan rintangan untuk mencegah penyebaran infeksi mencetuskan proses perbaikan untuk jaringan yang rusak. Respon peradangan dapat dikenali dari rasa sakit, kulit lebam, demam dll, yang disebabkan karena terjadi perubahan pada pembuluh darah di area infeksi: pembesaran diameter pembuluh darah, disertai peningkatan aliran darah di daerah infeksi. Hal ini dapat menyebabkan kulit tampak lebam kemerahan dan penurunan tekanan darah terutama pada pembuluh kecil. aktivasi molekul adhesi untuk merekatkan endotelia dengan pembuluh darah. kombinasi dari turunnya tekanan darah dan aktivasi molekul adhesi, akan memungkinkan sel darah putih bermigrasi ke endotelium dan masuk ke dalam jaringan . Proses ini dikenal sebagai ekstravasasi . Bagian tubuh yang mengalami peradangan memiliki tanda-tanda sebagai berikut: tumor atau membengkak calor atau menghangat
-
Upload
ratih-dwiputri-pradana -
Category
Documents
-
view
74 -
download
5
Transcript of Jawaban No.3

Radang (bahasa Inggris: inflammation) adalah rangkaian reaksi yang terjadi pada tempat jaringan yang mengalami cedera, seperti karena terbakar, atau terinfeksi. Radang atau inflamasi adalah satu dari respon utama sistem kekebalan terhadap infeksi dan iritasi. Inflamasi distimulasi oleh faktor kimia (histamin, bradikinin, serotonin, leukotrien, dan prostaglandin) yang dilepaskan oleh sel yang berperan sebagai mediator radang di dalam sistem kekebalan untuk melindungi jaringan sekitar dari penyebaran infeksi.
Radang mempunyai tiga peran penting dalam perlawanan terhadap infeksi:[1]
memungkinkan penambahan molekul dan sel efektor ke lokasi infeksi untuk meningkatkan performa makrofaga
menyediakan rintangan untuk mencegah penyebaran infeksi
mencetuskan proses perbaikan untuk jaringan yang rusak.
Respon peradangan dapat dikenali dari rasa sakit, kulit lebam, demam dll, yang disebabkan karena terjadi perubahan pada pembuluh darah di area infeksi:
pembesaran diameter pembuluh darah, disertai peningkatan aliran darah di daerah infeksi. Hal ini dapat menyebabkan kulit tampak lebam kemerahan dan penurunan tekanan darah terutama pada pembuluh kecil.
aktivasi molekul adhesi untuk merekatkan endotelia dengan pembuluh darah.
kombinasi dari turunnya tekanan darah dan aktivasi molekul adhesi, akan memungkinkan sel darah putih bermigrasi ke endotelium dan masuk ke dalam jaringan. Proses ini dikenal sebagai ekstravasasi.
Bagian tubuh yang mengalami peradangan memiliki tanda-tanda sebagai berikut:
tumor atau membengkak calor atau menghangat
dolor atau nyeri
rubor atau memerah
functio laesa atau daya pergerakan menurun
dan kemungkinan disfungsi organ atau jaringan.

Definisi
Inflamasi merupakan respons protektif setempat yang ditimbulkan oleh cedera atau kerusakan
jaringan, yang berfungsi menghancurkan, mengurangi, atau mengurung (sekuestrasi) baik agen
pencedera maupun jaringan yang cedera itu (Dorland, 2002).
Apabila jaringan cedera misalnya karena terbakar, teriris atau karena infeksi kuman, maka pada
jaringan ini akan terjadi rangkaian reaksi yang memusnahkan agen yang membahayakan jaringan atau
yang mencegah agen menyebar lebih luas. Reaksi-reaksi ini kemudian juga menyebabkan jaringan yang
cedera diperbaiki atau diganti dengan jaringan baru. Rangkaian reaksi ini disebut radang (Rukmono,
1973).
Agen yang dapat menyebabkan cedera pada jaringan, yang kemudian diikuti oleh radang adalah
kuman (mikroorganisme), benda (pisau, peluru, dsb.), suhu (panas atau dingin), berbagai jenis sinar
(sinar X atau sinar ultraviolet), listrik, zat-zat kimia, dan lain-lain. Cedera radang yang ditimbulkan oleh
berbagai agen ini menunjukkan proses yang mempunyai pokok-pokok yang sama, yaitu terjadi cedera
jaringan berupa degenerasi (kemunduran) atau nekrosis (kematian) jaringan, pelebaran kapiler yang
disertai oleh cedera dinding kapiler, terkumpulnya cairan dan sel (cairan plasma, sel darah, dan sel
jaringan) pada tempat radang yang disertai oleh proliferasi sel jaringan makrofag dan fibroblas,
terjadinya proses fagositosis, dan terjadinya perubahan-perubahan imunologik (Rukmono, 1973).
Secara garis besar, peradangan ditandai dengan vasodilatasi pembuluh darah lokal yang
mengakibatkan terjadinya aliran darah setempat yang berlebihan, kenaikan permeabilitas kapiler
disertai dengan kebocoran cairan dalam jumlah besar ke dalam ruang interstisial, pembekuan cairan
dalam ruang interstisial yang disebabkan oleh fibrinogen dan protein lainnya yang bocor dari kapiler
dalam jumlah berlebihan, migrasi sejumlah besar granulosit dan monosit ke dalam jaringan, dan
pembengkakan sel jaringan. Beberapa produk jaringan yang menimbulkan reaksi ini adalah histamin,
bradikinin, serotonin, prostaglandin, beberapa macam produk reaksi sistem komplemen, produk reaksi
sistem pembekuan darah, dan berbagai substansi hormonal yang disebut limfokin yang dilepaskan oleh
sel T yang tersensitisasi (Guyton & Hall, 1997).
Tanda-tanda radang (makroskopis)

Gambaran makroskopik peradangan sudah diuraikan 2000 tahun yang lampau. Tanda-tanda
radang ini oleh Celsus, seorang sarjana Roma yang hidup pada abad pertama sesudah Masehi, sudah
dikenal dan disebut tanda-tanda radang utama. Tanda-tanda radang ini masih digunakan hingga saat ini.
Tanda-tanda radang mencakup rubor (kemerahan), kalor (panas), dolor (rasa sakit), dan tumor
(pembengkakan). Tanda pokok yang kelima ditambahkan pada abad terakhir yaitu functio laesa
(perubahan fungsi) (Abrams, 1995; Rukmono, 1973; Mitchell & Cotran, 2003).
Umumnya, rubor atau kemerahan merupakan hal pertama yang terlihat di daerah yang
mengalami peradangan. Saat reaksi peradangan timbul, terjadi pelebaran arteriola yang mensuplai
darah ke daerah peradangan. Sehingga lebih banyak darah mengalir ke mikrosirkulasi lokal dan kapiler
meregang dengan cepat terisi penuh dengan darah. Keadaan ini disebut hiperemia atau kongesti,
menyebabkan warna merah lokal karena peradangan akut (Abrams, 1995; Rukmono, 1973).
Kalor terjadi bersamaan dengan kemerahan dari reaksi peradangan akut. Kalor disebabkan pula
oleh sirkulasi darah yang meningkat. Sebab darah yang memiliki suhu 37oC disalurkan ke permukaan
tubuh yang mengalami radang lebih banyak daripada ke daerah normal (Abrams, 1995; Rukmono,
1973).
Perubahan pH lokal atau konsentrasi lokal ion-ion tertentu dapat merangsang ujung-ujung saraf.
Pengeluaran zat seperti histamin atau zat bioaktif lainnya dapat merangsang saraf. Rasa sakit
disebabkan pula oleh tekanan yang meninggi akibat pembengkakan jaringan yang meradang (Abrams,
1995; Rukmono, 1973).
Pembengkakan sebagian disebabkan hiperemi dan sebagian besar ditimbulkan oleh pengiriman
cairan dan sel-sel dari sirkulasi darah ke jaringan-jaringan interstitial. Campuran dari cairan dan sel yang
tertimbun di daerah peradangan disebut eksudat meradang (Abrams, 1995; Rukmono, 1973).
Berdasarkan asal katanya, functio laesa adalah fungsi yang hilang (Dorland, 2002). Functio laesa
merupakan reaksi peradangan yang telah dikenal. Akan tetapi belum diketahui secara mendalam
mekanisme terganggunya fungsi jaringan yang meradang (Abrams, 1995).
Mekanisme radang
1. Radang akut

Radang akut adalah respon yang cepat dan segera terhadap cedera yang didesain untuk
mengirimkan leukosit ke daerah cedera. Leukosit membersihkan berbagai mikroba yang menginvasi
dan memulai proses pembongkaran jaringan nekrotik. Terdapat 2 komponen utama dalam proses
radang akut, yaitu perubahan penampang dan struktural dari pembuluh darah serta emigrasi dari
leukosit. Perubahan penampang pembuluh darah akan mengakibatkan meningkatnya aliran darah
dan terjadinya perubahan struktural pada pembuluh darah mikro akan memungkinkan protein
plasma dan leukosit meninggalkan sirkulasi darah. Leukosit yang berasal dari mikrosirkulasi akan
melakukan emigrasi dan selanjutnya berakumulasi di lokasi cedera (Mitchell & Cotran, 2003).
Segera setelah jejas, terjadi dilatasi arteriol lokal yang mungkin didahului oleh
vasokonstriksi singkat. Sfingter prakapiler membuka dengan akibat aliran darah dalam kapiler yang
telah berfungsi meningkat dan juga dibukanya anyaman kapiler yang sebelumnya inaktif. Akibatnya
anyaman venular pasca kapiler melebar dan diisi darah yang mengalir deras. Dengan demikian,
mikrovaskular pada lokasi jejas melebar dan berisi darah terbendung. Kecuali pada jejas yang
sangat ringan, bertambahnya aliran darah (hiperemia) pada tahap awal akan disusul oleh
perlambatan aliran darah, perubahan tekanan intravaskular dan perubahan pada orientasi unsur-
unsur berbentuk darah terhadap dinding pembuluhnya. Perubahan pembuluh darah dilihat dari
segi waktu, sedikit banyak tergantung dari parahnya jejas. Dilatasi arteriol timbul dalam beberapa
menit setelah jejas. Perlambatan dan bendungan tampak setelah 10-30 menit (Robbins & Kumar,
1995).
Peningkatan permeabilitas vaskuler disertai keluarnya protein plasma dan sel-sel darah
putih ke dalam jaringan disebut eksudasi dan merupakan gambaran utama reaksi radang akut.
Vaskulatur-mikro pada dasarnya terdiri dari saluran-saluran yang berkesinambungan berlapis
endotel yang bercabang-cabang dan mengadakan anastomosis. Sel endotel dilapisi oleh selaput
basalis yang berkesinambungan (Robbins & Kumar, 1995).
Pada ujung arteriol kapiler, tekanan hidrostatik yang tinggi mendesak cairan keluar ke
dalam ruang jaringan interstisial dengan cara ultrafiltrasi. Hal ini berakibat meningkatnya
konsentrasi protein plasma dan menyebabkan tekanan osmotik koloid bertambah besar, dengan
menarik kembali cairan pada pangkal kapiler venula. Pertukaran normal tersebut akan menyisakan
sedikit cairan dalam jaringan interstisial yang mengalir dari ruang jaringan melalui saluran limfatik.

Umumnya, dinding kapiler dapat dilalui air, garam, dan larutan sampai berat jenis 10.000 dalton
(Robbins & Kumar, 1995).
Eksudat adalah cairan radang ekstravaskuler dengan berat jenis tinggi (di atas 1.020) dan
seringkali mengandung protein 2-4 mg% serta sel-sel darah putih yang melakukan emigrasi. Cairan
ini tertimbun sebagai akibat peningkatan permeabilitas vaskuler (yang memungkinkan protein
plasma dengan molekul besar dapat terlepas), bertambahnya tekanan hidrostatik intravaskular
sebagai akibat aliran darah lokal yang meningkat pula dan serentetan peristiwa rumit leukosit yang
menyebabkan emigrasinya (Robbins & Kumar, 1995).
Penimbunan sel-sel darah putih, terutama neutrofil dan monosit pada lokasi jejas,
merupakan aspek terpenting reaksi radang. Sel-sel darah putih mampu memfagosit bahan yang
bersifat asing, termasuk bakteri dan debris sel-sel nekrosis, dan enzim lisosom yang terdapat di
dalamnya membantu pertahanan tubuh dengan beberapa cara. Beberapa produk sel darah putih
merupakan penggerak reaksi radang, dan pada hal-hal tertentu menimbulkan kerusakan jaringan
yang berarti (Robbins & Kumar, 1995).
Dalam fokus radang, awal bendungan sirkulasi mikro akan menyebabkan sel-sel darah
merah menggumpal dan membentuk agregat-agregat yang lebih besar daripada leukosit sendiri.
Menurut hukum fisika aliran, massa sel darah merah akan terdapat di bagian tengah dalam aliran
aksial, dan sel-sel darah putih pindah ke bagian tepi (marginasi). Mula-mula sel darah putih
bergerak dan menggulung pelan-pelan sepanjang permukaan endotel pada aliran yang tersendat
tetapi kemudian sel-sel tersebut akan melekat dan melapisi permukaan endotel (Robbins & Kumar,
1995).
Emigrasi adalah proses perpindahan sel darah putih yang bergerak keluar dari pembuluh
darah. Tempat utama emigrasi leukosit adalah pertemuan antar-sel endotel. Walaupun pelebaran
pertemuan antar-sel memudahkan emigrasi leukosit, tetapi leukosit mampu menyusup sendiri
melalui pertemuan antar-sel endotel yang tampak tertutup tanpa perubahan nyata (Robbins &
Kumar, 1995).
Setelah meninggalkan pembuluh darah, leukosit bergerak menuju ke arah utama lokasi
jejas. Migrasi sel darah putih yang terarah ini disebabkan oleh pengaruh-pengaruh kimia yang dapat
berdifusi disebut kemotaksis. Hampir semua jenis sel darah putih dipengaruhi oleh faktor-faktor

kemotaksis dalam derajat yang berbeda-beda. Neutrofil dan monosit paling reaktif terhadap
rangsang kemotaksis. Sebaliknya limfosit bereaksi lemah. Beberapa faktor kemotaksis dapat
mempengaruhi neutrofil maupun monosit, yang lainnya bekerja secara selektif terhadap beberapa
jenis sel darah putih. Faktor-faktor kemotaksis dapat endogen berasal dari protein plasma atau
eksogen, misalnya produk bakteri (Robbins & Kumar, 1995).
Setelah leukosit sampai di lokasi radang, terjadilah proses fagositosis. Meskipun sel-sel
fagosit dapat melekat pada partikel dan bakteri tanpa didahului oleh suatu proses pengenalan yang
khas, tetapi fagositosis akan sangat ditunjang apabila mikroorganisme diliputi oleh opsonin, yang
terdapat dalam serum (misalnya IgG, C3). Setelah bakteri yang mengalami opsonisasi melekat pada
permukaan, selanjutnya sel fagosit sebagian besar akan meliputi partikel, berdampak pada
pembentukan kantung yang dalam. Partikel ini terletak pada vesikel sitoplasma yang masih terikat
pada selaput sel, disebut fagosom. Meskipun pada waktu pembentukan fagosom, sebelum
menutup lengkap, granula-granula sitoplasma neutrofil menyatu dengan fagosom dan melepaskan
isinya ke dalamnya, suatu proses yang disebut degranulasi. Sebagian besar mikroorganisme yang
telah mengalami pelahapan mudah dihancurkan oleh fagosit yang berakibat pada kematian
mikroorganisme. Walaupun beberapa organisme yang virulen dapat menghancurkan leukosit
(Robbins & Kumar, 1995).
2. Radang kronis
Radang kronis dapat diartikan sebagai inflamasi yang berdurasi panjang (berminggu-minggu
hingga bertahun-tahun) dan terjadi proses secara simultan dari inflamasi aktif, cedera jaringan, dan
penyembuhan. Perbedaannya dengan radang akut, radang akut ditandai dengan perubahan
vaskuler, edema, dan infiltrasi neutrofil dalam jumlah besar. Sedangkan radang kronik ditandai oleh
infiltrasi sel mononuklir (seperti makrofag, limfosit, dan sel plasma), destruksi jaringan, dan
perbaikan (meliputi proliferasi pembuluh darah baru/angiogenesis dan fibrosis) (Mitchell & Cotran,
2003).
Radang kronik dapat timbul melalui satu atau dua jalan. Dapat timbul menyusul radang
akut, atau responnya sejak awal bersifat kronik. Perubahan radang akut menjadi radang kronik
berlangsung bila respon radang akut tidak dapat reda, disebabkan agen penyebab jejas yang
menetap atau terdapat gangguan pada proses penyembuhan normal. Ada kalanya radang kronik

sejak awal merupakan proses primer. Sering penyebab jejas memiliki toksisitas rendah
dibandingkan dengan penyebab yang menimbulkan radang akut. Terdapat 3 kelompok besar yang
menjadi penyebabnya, yaitu infeksi persisten oleh mikroorganisme intrasel tertentu (seperti basil
tuberkel, Treponema palidum, dan jamur-jamur tertentu), kontak lama dengan bahan yang tidak
dapat hancur (misalnya silika), penyakit autoimun. Bila suatu radang berlangsung lebih lama dari 4
atau 6 minggu disebut kronik. Tetapi karena banyak kebergantungan respon efektif tuan rumah dan
sifat alami jejas, maka batasan waktu tidak banyak artinya. Pembedaan antara radang akut dan
kronik sebaiknya berdasarkan pola morfologi reaksi (Robbins & Kumar, 1995).
Mediator kimia peradangan
Bahan kimia yang berasal dari plasma maupun jaringan merupakan rantai penting antara
terjadinya jejas dengan fenomena radang. Meskipun beberapa cedera langsung merusak endotelium
pembuluh darah yang menimbulkan kebocoran protein dan cairan di daerah cedera, pada banyak kasus
cedera mencetuskan pembentukan dan/atau pengeluaran zat-zat kimia di dalam tubuh. Banyak jenis
cedera yang dapat mengaktifkan mediator endogen yang sama, yang dapat menerangkan sifat stereotip
dari respon peradangan terhadap berbagai macam rangsang. Karena pola dasar radang akut stereotip,
tidak tergantung jenis jaringan maupun agen penyebab pada hakekatnya menyertai mediator-mediator
kimia yang sama yang tersebar luas dalam tubuh. Beberapa mediator dapat bekerja bersama, sehingga
memberi mekanisme biologi yang memperkuat kerja mediator. Radang juga memiliki mekanisme
kontrol yaitu inaktivasi mediator kimia lokal yang cepat oleh sistem enzim atau antagonis (Abrams,
1995; Robbins & Kumar, 1995).
Cukup banyak substansi yang dikeluarkan secara endogen telah dikenal sebagai mediator dari
respon peradangan. Identifikasinya saat ini sulit dilakukan. Walaupun daftar mediator yang diusulkan
panjang dan kompleks, tetapi mediator yang lebih dikenal dapat digolongkan menjadi golongan amina
vasoaktif (histamin dan serotonin), protease plasma (sistem kinin, komplemen, dan koagulasi
fibrinolitik), metabolit asam arakidonat (leukotrien dan prostaglandin), produk leukosit (enzim lisosom
dan limfokin), dan berbagai macam mediator lainnya (misal, radikal bebas yang berasal dari oksigen dan
faktor yang mengaktifkan trombosit) (Abrams, 1995; Robbins & Kumar, 1995).
1. Amina vasoaktif

Amina vasoaktif yang paling penting adalah histamin. Sejumlah besar histamin disimpan
dalam granula sel jaringan penyambung yang disebut sel mast. Histamin tersebar luas dalam tubuh.
Histamin juga terdapat dalam sel basofil dan trombosit. Histamin yang tersimpan merupakan
histamin yang tidak aktif dan baru menampilkan efek vaskularnya bila dilepaskan. Stimulus yang
dapat menyebabkan dilepaskannya histamin adalah jejas fisik (misal trauma atau panas), reaksi
imunologi (meliputi pengikatan antibodi IgE terhadap reseptor Fc pada sel mast), fragment
komplemen C3a dan C5a (disebut anafilaktosin), protein derivat leukosit yang melepaskan histamin,
neuropeptida (misal, substansi P), dan sitokin tertentu (misal, IL-1 dan IL-8) (Mitchell & Cotran,
2003; Robbins & Kumar, 1995; Abrams, 1995).
Pada manusia, histamin menyebabkan dilatasi arteriola, meningkatkan permeabilitas
venula, dan pelebaran pertemuan antar-sel endotel. Histamin bekerja dengan mengikatkan diri
pada reseptor-reseptor histamin jenis H-1 yang ada pada endotel pembuluh darah. Pada perannya
dalam fenomena vaskular, histamin juga dilaporkan merupakan bahan kemotaksis khas untuk
eosinofil. Segera setelah dilepaskan oleh sel mast, histamin dibuat menjadi inaktif oleh histaminase.
Antihistamin merupakan obat yang dibuat untuk menghambat efek mediator dari histamin. Perlu
diketahui bahwa obat antihistamin hanya dapat menghambat tahap dini peningkatan permeabilitas
vaskular dan histamin tidak berperan pada tahap tertunda yang dipertahankan pada peningkatan
permeabilitas (Mitchell & Cotran, 2003; Robbins & Kumar, 1995; Abrams, 1995).
Serotonin (5-hidroksitriptamin) juga merupakan suatu bentuk mediator vaasoaktif.
Serotonin ditemukan terutama di dalam trombosit yang padat granula (bersama dengan histamin,
adenosin difosfat, dan kalsium). Serotonin dilepaskan selama agregasi trombosit. Serotonin pada
binatang pengerat memiliki efek yang sama seperti halnya histamin, tetapi perannya sebagai
mediator pada manusia tidak terbukti (Mitchell & Cotran, 2003; Robbins & Kumar, 1995).
2. Protease plasma
Berbagai macam fenomena dalam respon radang diperantarai oleh tiga faktor plasma yang
saling berkaitan yaitu sistem kinin, pembekuan, dan komplemen. Seluruh proses dihubungkan oleh
aktivasi awal oleh faktor Hageman (disebut juga faktor XII dalam sistem koagulasi intrinsik). Faktor
XII adalah suatu protein yang disintesis oleh hati yang bersirkulasi dalam bentuk inaktif hingga
bertemu kolagen, membrana basalis, atau trombosit teraktivasi di lokasi jejas endotelium. Dengan

bantuan kofaktor high-molecular-weight kininogen (HMWK)/kininogen berat molekul tinggi, faktor
XII kemudian mengalami perubahan bentuk menjadi faktor XIIa. Faktor XIIa dapat membongkar
pusat serin aktif yang dapat memecah sejumlah substrat protein (Mitchell & Cotran, 2003).
Aktivasi sistem kinin pada akhirnya menyebabkan pembentukan bradikinin. Bradikinin
merupakan polipeptida yang berasal dari plasma sebagai prekursor yang disebut HMWK. Prekursor
glikoprotein ini diuraikan oleh enzim proteolitik kalikrein. Kalikrein sendiri berasal dari prekursornya
yaitu prekalikrein yang diaktifkan oleh faktor XIIa. Seperti halnya histamin, bradikinin menyebabkan
dilatasi arteriola, meningkatkan permeabilitas venula dan kontraksi otot polos bronkial. Bradikinin
tidak menyebabkan kemotaksis untuk leukosit, tetapi menyebabkan rasa nyeri bila disuntikkan ke
dalam kulit. Bradikinin dapat bertindak dalam sel-sel endotel dengan meningkatkan celah antar sel.
Kinin akan dibuat inaktif secara cepat oleh kininase yang terdapat dalam plasma dan jaringan, dan
perannya dibatasi pada tahap dini peningkatan permeabilitas pembuluh darah (Mitchell & Cotran,
2003; Robbins & Kumar, 1995).
Pada sistem pembekuan, rangsangan sistem proteolitik mengakibatkan aktivasi trombin
yang kemudian memecah fibrinogen yang dapat larut dalam sirkulasi menjadi gumpalan fibrin.
Faktor Xa menyebabkan peningkatan permeabilitas vaskular dan emigrasi leukosit. Trombin
memperkuat perlekatan leukosit pada endotel dan dengan cara menghasilkan fibrinopeptida
(selama pembelahan fibrinogen) dapat meningkatkan permeabilitas vaskular dan sebagai
kemotaksis leukosit (Mitchell & Cotran, 2003).
Ketika faktor XIIa menginduksi pembekuan, di sisi lain terjadi aktivasi sistem fibrinolitik.
Mekanisme ini terjadi sebagai umpan balik pembekuan dengan cara memecah fibrin kemudian
melarutkan gumpalan fibrin. Tanpa adanya fibrinolisis ini, akan terus menerus terjadi sistem
pembekuan dan mengakibatkan penggumpalan pada keseluruhan vaskular. Plasminogen activator
(dilepaskan oleh endotel, leukosit, dan jaringan lain) dan kalikrein adalah protein plasma yang
terikat dalam perkembangan gumpalan fibrin. Produk hasil dari keduanya yaitu plasmin, merupakan
protease multifungsi yang memecah fibrin (Mitchell & Cotran, 2003).
Sistem komplemen terdiri dari satu seri protein plasma yang berperan penting dalam
imunitas maupun radang. Tahap penting pembentukan fungsi biologi komplemen ialah aktivasi
komponen ketiga (C3). Pembelahan C3 dapat terjadi oleh apa yang disebut ”jalur klasik” yang

tercetus oleh pengikatan C1 pada kompleks antigen-antibodi (IgG atau IgM) atau melalui jalur
alternatif yang dicetuskan oleh polisakarida bakteri (misal, endotoksin), polisakarida kompleks, atau
IgA teragregasi, dan melibatkan serangkaian komponen serum (termasuk properdin dan faktor B
dan D). Jalur manapun yang terlibat, pada akhirnya sistem komplemen akan memakai urutan
efektor akhir bersama yang menyangkut C5 sampai C9 yang mengakibatkan pembentukan
beberapa faktor yang secara biologi aktif serta lisis sel-sel yang dilapisi antibodi (Mitchell & Cotran,
2003; Robbins & Kumar, 1995).
Faktor yang berasal dari komplemen, mempengaruhi berbagai fenomena radang akut, yaitu
pada fenomena vaskular, kemotaksis, dan fagositosis. C3a dan C5a (disebut juga anafilaktosin)
meningkatkan permeabilitas vaskular dan menyebabkan vasodilatasi dengan cara menginduksi sel
mast untuk mengeluarkan histamin. C5a mengaktifkan jalur lipoksigenase dari metabolisme asam
arakidonat dalam netrofil dan monosit. C5a juga menyebabkan adhesi neutrofil pada endotel dan
kemotaksis untuk monosit, eosinofil, basofil dan neutrofil. Komplemen yang lainnya, C3b, apabila
melekat pada dinding sel bakteri akan bekerja sebagai opsonin dan memudahkan fagositosis
neutrofil dan makrofag yang mengandung reseptor C3b pada permukaannya (Mitchell & Cotran,
2003).
a. Metabolit asam arakidonat
Asam arakidonat merupakan asam lemak tidak jenuh (20-carbon polyunsaturated fatty
acid) yang utamanya berasal dari asupan asam linoleat dan berada dalam tubuh dalam bentuk
esterifikasi sebagai komponen fosfolipid membran sel. Asam arakidonat dilepaskan dari
fosfolipid melalui fosfolipase seluler yang diaktifkan oleh stimulasi mekanik, kimia, atau fisik,
atau oleh mediator inflamasi lainnya seperti C5a. Metabolisme asam arakidonat berlangsung
melalui salah satu dari dua jalur utama, sesuai dengan enzim yang mencetuskan, yaitu jalur
siklooksigenase dan lipoksigenase. Metabolit asam arakidonat (disebut juga eikosanoid) dapat
memperantarai setiap langkah inflamasi. (Mitchell & Cotran, 2003).
Jalur siklooksigenase menghasilkan prostaglandin (PG) E2 (PGE2), PGD2, PGF2?, PGI2
(prostasiklin), dan tromboksan A2 (TXA2). Setiap produk tersebut berasal dari PGH2 oleh
pengaruh kerja enzim yang spesifik. PGH2 sangat tidak stabil, merupakan prekursor hasil akhir
biologi aktif jalur siklooksigenase. Beberapa enzim mempunyai distribusi jaringan tertentu.

Misalnya, trombosit mengandung enzim tromboksan sintetase sehingga produk utamanya
adalah TXA2. TXA2 merupakan agen agregasi trombosit yang kuat dan vasokonstriktor. Di sisi
lain, endotelium kekurangan dalam hal tromboksan sintetase, tetapi banyak memiliki
prostasiklin sintetase yang membentuk PGI2. PGI2 merupakan vasodilator dan penghambat
kuat agregasi trombosit. PGD2 merupakan metabolit utama dari jalur siklooksigenase pada sel
mast. Bersama dengan PGE2 dan PGF2?, PGD2 menyebabkan vasodilatasi dan pembentukan
edema. Prostaglandin terlibat dalam patogenesis nyeri dan demam pada inflamasi (Mitchell &
Cotran, 2003).
Jalur lipoksigenase merupakan jalur yang penting untuk membentuk bahan-bahan
proinflamasi yang kuat. 5-lipoksigenase merupakan enzim metabolit asam arakidonat utama
pada neutrofil. Produk dari aksinya memiliki karakteristik yang terbaik. 5-HPETE (asam 5-
hidroperoksieikosatetranoik) merupakan derivat 5-hidroperoksi asam arakidonat yang tidak
stabil dan direduksi menjadi 5-HETE (asam 5-hidroksieikosatetraenoik) (sebagai kemotaksis
untuk neutrofil) atau diubah menjadi golongan senyawa yang disebut leukotrien. Produk dari
5-HPETE adalah leukotrien (LT) A4 (LTA4), LTB4, LTC4, LTD4, dan LTE5. LTB4 merupakan agen
kemotaksis kuat dan menyebabkan agregasi dari neutrofil. LTC4, LTD4, dan LTE4 menyebabkan
vasokonstriksi, bronkospasme, dan meningkatkan permeabilitas vaskular (Mitchell & Cotran,
2003).
Lipoksin juga termasuk hasil dari jalur lipoksigenase yang disintesis menggunakan jalur
transeluler. Trombosit sendiri tidak dapat membentuk lipoksin A4 dan B4 (LXA4 dan LXB4), tetapi
dapat membentuk metabolit dari intermediat LTA4 yang berasal dari neutrofil. Lipoksin
mempunyai aksi baik pro- dan anti- inflamasi. Misal, LXA4 menyebabkan vasodilatasi dan
antagonis vasokonstriksi yang distimulasi LTC4. Aktivitas lainnya menghambat kemotaksis
neutrofil dan perlekatan ketika menstimulasi perlekatan monosit (Mitchell & Cotran, 2003).
b. Produk leukosit
Granula lisosom yang terdapat dalam neutrofil dan monosit mengandung molekul
mediator inflamasi. Mediator ini dilepaskan setelah kematian sel oleh karena peluruhan
selama pembentukan vakuola fagosit atau oleh fagositosis yang terhalang karena ukurannya
besar dan permukaan yang tidak dapat dicerna. Kalikrein yang dilepaskan dari lisosom

menyebabkan pembentukan bradikinin. Neutrofil juga merupakan sumber fosfolipase yang
diperlukan untuk sintesis asam arakidonat (Robbins & Kumar, 1995).
Di dalam lisosom monosit dan makrofag juga banyak mengandung bahan yang aktif
untuk proses radang. Pelepasannya penting pada radang akut dan radang kronik. Limfosit yang
telah peka terhadap antigen melepaskan limfokin. Limfokin merupakan faktor yang
menyebabkan penimbunan dan pengaktifan makrofag pada lokasi radang. Limfokin penting
pada radang kronik (Robbins & Kumar).
c. Mediator lainnya
Metabolit oksigen reaktif yang dibentuk dalam sel fagosit saat fagositosis dapat luruh
memasuki lingkungan ekstrasel. Diduga bahwa radikal-radikal bebas yang sangat toksik
meningkatkan permeabilitas vaskular dengan cara merusak endotel kapiler. Selain itu, ion-ion
superoksida dan hidroksil juga dapat menyebabkan peroksidase asam arakidonat tanpa enzim.
Akibatnya, akan dapat terbentuk lipid-lipid kemotaksis (Robbins & Kumar, 1995).
Aseter-PAF merupakan mediator lipid yang menggiatkan trombosit. Hal ini karena
menyebabkan agregasi trombosit ketika dilepaskan oleh sel mast. Selain sel mast, neutrofil dan
makrofag juga dapat mensintesis aseter-PAF. Aseter-PAF meningkatkan permeabilitas vaskular,
adhesi leukosit dan merangsang neutrofil dan makrofag (Robbins & Kumar, 1995).
Daftar Pustaka
1. Dorland, W.A.N. (2002). Kamus Kedokteran Dorland (Setiawan, A., Banni, A.P., Widjaja, A.C., Adji, A.S., Soegiarto, B., Kurniawan, D., dkk , penerjemah). Jakarta: EGC. (Buku asli diterbitkan 2000).
2. Rukmono (1973). Kumpulan kuliah patologi. Jakarta: Bagian patologi anatomik FK UI.3. Guyton, A.C. & Hall, J.E. (1997). Buku ajar fisiologi kedokteran (9th ed.) (Setiawan, I., Tengadi, K.A., Santoso,
A., penerjemah). Jakarta: EGC (Buku asli diterbitkan 1996).4. Abrams, G.D. (1995). Respon tubuh terhadap cedera. Dalam S. A. Price & L. M. Wilson, Patofisiologi: Konsep
klinis proses-proses penyakit (4th ed.)(pp.35-61)(Anugerah, P., penerjemah). Jakarta: EGC (Buku asli diterbitkan 1992).
5. Mitchell, R.N. & Cotran, R.S. (2003). Acute and chronic inflammation. Dalam S. L. Robbins & V. Kumar, Robbins Basic Pathology (7th ed.)(pp33-59). Philadelphia: Elsevier Saunders.
6. Robbins, S.L. & Kumar, V. (1995). Buku ajar patologi I (4th ed.)(Staf pengajar laboratorium patologi anatomik FK UI, penerjemah). Jakarta: EGC (Buku asli diterbitkan 1987).

Infeksi adalah kolonalisasi yang dilakukan oleh spesies asing terhadap organisme inang, dan bersifat pilang membahayakan inang. Organisme penginfeksi, atau patogen, menggunakan sarana yang dimiliki inang untuk dapat memperbanyak diri, yang pada akhirnya merugikan inang. Patogen mengganggu fungsi normal inang dan dapat berakibat pada luka kronik, gangrene, kehilangan organ tubuh, dan bahkan kematian. Respons inang terhadap infeksi disebut peradangan. Secara umum, patogen umumnya dikategorikan sebagai organisme mikroskopik, walaupun sebenarnya definisinya lebih luas, mencakup bakteri, parasit, fungi, virus, prion, dan viroid.
Simbiosis antara parasit dan inang, di mana satu pihak diuntungkan dan satu pihak dirugikan, digolongkan sebagai parasitisme. Cabang kedokteran yang menitikberatkan infeksi dan patogen adalah cabang penyakit infeksi.
Secara umum infeksi terbagi menjadi dua golongan besar:[1]
Infeksi yang terjadi karena terpapar oleh antigen dari luar tubuh Infeksi yang terjadi karena difusi cairan tubuh atau jaringan, seperti virus HIV, karena
virus tersebut tidak dapat hidup di luar tubuh.
Infeksi awal
Setelah menembus jaringan, patogen dapat berkembang pada di luar sel tubuh (ekstraselular) atau menggunakan sel tubuh sebagai inangnya (intraselular). Patogen intraselular lebih lanjut dapat diklasifikasikan lebih lanjut:
patogen yang berkembang biak dengan bebas di dalam sel, seperti : virus dan beberapa bakteri (Chlamydia, Rickettsia, Listeria).
patogen yang berkembang biak di dalam vesikel, seperti Mycobacteria.
Jaringan yang tertembus dapat mengalami kerusakan oleh karena infeksi patogen, misalnya oleh eksotoksin yang disekresi pada permukaan sel, atau sekresi endotoksin yang memicu sekresi sitokina oleh makrofaga, dan mengakibatkan gejala-gejala lokal maupun sistemik.[2]
terpuruknya mekanisme sistem kekebalan
Pada tahapan umum sebuah infeksi, antigen selalu akan memicu sistem kekebalan turunan, dan kemudian sistem kekebalan tiruan pada saat akut. Tetapi lintasan infeksi tidak selalu demikian, sistem kekebalan dapat gagal memadamkan infeksi, karena terjadi fokus infeksi berupa:[3]
subversi sistem kekebalan oleh patogen kelainan bawaan yang disebabkan gen

tidak terkendalinya mekanisme sistem kekebalan
Perambatan perkembangan patogen bergantung pada kemampuan replikasi di dalam inangnya dan kemudian menyebar ke dalam inang yang baru dengan proses infeksi. Untuk itu, patogen diharuskan untuk berkembangbiak tanpa memicu sistem kekebalan, atau dengan kata lain, patogen diharuskan untuk tidak menggerogoti inangnya terlalu cepat.
Patogen yang dapat bertahan hanya patogen yang telah mengembangkan mekanisme untuk menghindari terpicunya sistem kekebalan.
Variasi serotipe
Salah satu cara yang digunakan patogen untuk menghindari sistem kekebalan adalah dengan mengubah struktur permukaan selnya. Banyak patogen ekstraselular mempunyai tipe antigenik yang sangat beragam. Salah satu contoh adalah streptococcus pneumoniae, penyebab pneumonia, yang mempunyai banyak tipe antigenik dan baru diketahui 84 macam. Setiap macam mempunyai stuktur pelapis polisakarida yang berbeda. Tipe-tipe tersebut dibedakan berdasarkan uji serologi, sehingga disebut juga serotipe. Infeksi yang dilakukan oleh satu serotipe tertentu dapat memicu sistem kekebalan tiruan terhadapnya, tetapi tidak terhadap infeksi ulang yang dilakukan oleh serotipe yang berbeda, oleh karena sistem kekebalan tiruan melihat satu serotipe sebagai satu jenis organisme yang berbeda. Infeksi akut berulang dari antigen yang sama dapat terjadi karena hal ini.
Penggunaan kapsul pelindung yang mencegah lisis oleh sistem komplemen dan fagosit juga dilakukan Mycobacterium tuberculosis. Spesies bacterioides umumnya bakteri komensal yang berdiam di usus buntu mamalia. Beberapa spesies seperti Bacterioides fragilis adalah patogen oportunistik penyebab infeksi pada lapisan peritoneum. Spesies ini menghindari sistem kekebalan dengan mempengaruhi pencerap yang digunakan fagosit untuk menelan bakteri atau dengan menyamar sebagai sel organisme tersebut sehingga sistem kekebalan tidak mengenali mereka sebagai patogen.
Bakteri dan jamur mungkin juga membentuk lapisan bio kompleks, menyediakan perlindungan dari sel dan protein dari sistem kekebalan. Penelitian terbaru menunjukkan bahwa lapisan bio muncul di infeksi yang berhasil, termasuk infeksi kronis Pseudomonas aeruginosa dan Burkholderia cenocepacia, ciri utama dari cystic fibrosis.
Mutasi genetik

Deteksi trypanosome oleh antibodi akan memicu pergantian gen VSG pada DNA, sehingga dihasilkan protein VSG yang berbeda pula. Tubuh kemudian akan membuat antibodi baru dengan cara yang sama, tetapi setiap antibodi yang baru dibuat mengenali trypanosome, gen VSG akan berubah lagi sebelum sistem kekebalan terpicu. Dengan demikian trypanosome berada satu langkah lebih cepat dari sistem kekebalan, sehingga meskipun berupa protozoa yang berkembangbiak ekstraselular, fokus infeksinya bersifat kronik dan membentuk kompleks imun dan peradangan, hingga berakhir pada kerusakan saraf dan koma. Hal ini yang menyebabkan African trypanosomiasis mendapatkan julukan penyakit "tidur". Malaria adalah contoh lain penyakit yang disebabkan parasit protozoa dengan kemampuan tata-ulang DNA, yang sangat sulit diatasi oleh sistem kekebalan.
Metode kedua yang lebih dinamis ditunjukkan oleh virus influensa. Virus influensa dikenali oleh sistem kekebalan melalui hemaglutinin yang terdapat pada permukaan virus.
Mutasi genetik yang pertama disebut antigenic drift yang mengubah notasi gen ekspresi dari hemaglutinin, sebagai respon dari protein yang berada pada permukaan, neuraminidase. Mutasi yang lain mengubah epitop agar tidak dikenali oleh sel T, khususnya yang mempunyai pencerap CD8.[4]
Mutasi genetik yang kedua disebut antigenic shift yang terjadi karena tertukarnya RNA antara virus baru dengan virus yang telah lama berada dalam tubuh inang.
Mekanisme ketiga melibatkan tata-ulang DNA terprogram. African trypanosome mempunyai kemampuan untuk mengubah major surface antigen berkali-kali dengan satu kali infeksi. Trypanosome terbalut sebuah tipe glikoprotein yang disebut variant-specific glycoprotein (VSG), yang dengan mudah dapat dikenali oleh sistem kekebalan. Meskipun demikian, DNA trypanosome mengandung lebih dari 1000 gen VSG dengan ekspresi antigenik yang berlainan.
o Pada tingkat bakteri, kemampuan tata-ulang DNA juga dijumpai pada Salmonella typhimurium dan Neisseria gonorrhoeae.

Fokus infeksi laten
Dalam fisiologi, laten didefinisikan sebagai jedah waktu antara stimulus dan respon yang terpicu di dalam suatu organisme. Virus umumnya segera akan mengkoordinir sintesis protein viral yang dibutuhkan untuk proliferasi, setelah berhasil melakukan infeksi terhadap sebuah sel. Mekanisme semacam ini akan mengakibatkan kondisi akut yang akan segera direspon oleh sistem kekebalan tiruan. Sel T akan dengan mudah memindai fragmen dari protein viral yang tertera pada permukaan molekul MHC dan memadamkan infeksi.
Meskipun demikian, masih terdapat jenis virus yang lain yang mampu menunda proses sintesis protein viral di dalam sel. Kondisi ini disebut kondisi laten, saat tidak terjadi replikasi virus di dalam sel. Infeksi laten tidak menimbulkan penyakit dan keberadaan virus tidak terdeteksi oleh karena tidak terdapat fragmen viral pada molekul MHC. Salah satu contoh adalah virus Herpes Simplex, yang melakukan infeksi epitelia dengan fokus berupa sel saraf di daerah tersebut.
Setelah sistem kekebalan mengatasi infeksi pada epitelia, virus HS tetap berada dalam kondisi laten di dalam neuron saraf. Beberapa faktor seperti sinar matahari, infeksi bakteri dan perubahan hormonal akan mengaktivasi virus ini untuk bermigrasi melalui akson dan melakukan infeksi ulang pada jaringan epitelial. Fokus infeksi berupa neuron memiliki dua keunggulan:
peptida viral yang dihasilkan sangat sedikit, menghasilkan fragmen yang tidak menyolok neuron mempunyai molekul MHC kelas I, yang kecil, sehingga sulit dideteksi sel T CD8.
Contoh lain adalah virus Epstein-Barr (EBV), sebuah tipe virus herpes yang lain, memiliki kondisi laten di dalam sel B. Proliferasi sel B akan menghasilkan sel baru dengan EBV di dalamnya.
Evolusi fitur
Beberapa bakteri yang biasanya dicerna oleh makrofaga dengan proses fagositosis, telah berevolusi dan berhasil membuat makrofaga sebagai fokus infeksi. Salah satu contoh adalah Mycobacterium tuberculosis yang tertelan oleh makrofaga, akan menghalangi pencairan lisosom ke dalam fagosom dan melindunginya dari sitokina di dalam lisosom.

Listeria monocytogenes, bahkan dapat keluar dari fagosom dan masuk ke dalam sitoplasma dan membuat replikasi di dalamnya. Kemudian menginfeksi sel yang berdekatan, tanpa keluar dari ruang intraselularnya.
Sebuah parasit protozoa toxoplasma gondii, dapat membuat vesikel sendiri yang memisahkannya dari bagian sel yang lain. Hal ini memungkinkan T. gondii untuk membuat peptida dengan fragmen yang tidak termuat pada molekul MHC, sehingga keberadaannya tidak terdeteksi sistem kekebalan.
[sunting] Perlawanan patogen
Staphylococci aureus, salah satu penyebab mastitis pada ternak sapi. Kapsul yang besar melindung organisme ini dari sistem kekebalan sapi, sebagai inangnya. Citra ini diambil dengan 50.000x pembesaran dari substrat replikasi yang kering dan beku.
Respon patogen dalam menghadapi sistem kekebalan juga berlainan. Selain dengan berbagai cara untuk menghindar, beberapa patogen melakukan perlawanan. Staphylococci aureus melepaskan dua macam toksin yaitu staphylococcal enterotoxin dan toxic shock syndrome toxin-1 yang berperan sebagai superantigen.
“Superantigen adalah protein yang mengikat sejumlah pencerap antigen dari sel T. Ikatan ini menyebabkan sel T mengalamai apoptosis dengan sangat cepat. ”

Organisme lain seperti Streptococcus pyogenes, dan Bacillus anthracis memiliki mekanisme untuk membunuh langsung fagosit.
Banyak patogen melakukan perlawanan dalam rentang waktu infeksi akut. Hal merupakan tekanan terhadap sistem kekebalan (bahasa Inggris: immunosuppression) dan menyebabkan tubuh inang menjadi rentan terhadap infeksi susulan oleh patogen jenis lain. Contoh-contoh penting meliputi trauma, luka bakar dan operasi bedah besar. Pasien dengan luka bakar tidak dapat merespon infeksi, sehingga infeksi ringan pun dapat menyebabkan kematian.
Infeksi virus measles juga merupakan salah satu contoh tekanan terhadap sistem kekebalan. Banyak anak-anak yang menderita malnutrisi menjadi korban, hingga meninggal dunia, karena infeksi susulan pada saat sistem kekebalan tertekan oleh infeksi virus measles. Infeksi susulan biasanya berupa bakteri penyebab pneumonia. Virus measles mempunyai fokus infeksi pada sel dendritik sehingga mempengaruhi kinerja sel T dan sel B dalam sistem kekebalan, dan aktivasi makrofaga oleh sel TH1.
[sunting] Fokus infeksi
Salah satu contoh terbaik dari topik ini adalah fokus infeksi yang dimiliki oleh virus HIV, berupa putusnya mata rantai sistem kekebalan selular [5] karena padamnya kemampuan sel T CD4 untuk teraktivasi dan terdiferensiasi menjadi sel T pembantu. Terputusnya mata rantai tersebut terjadi perlahan tanpa memantik sistem kekebalan oleh sebab sifat laten retrovirus. Sejumlah kecil PSK Gambia dan Kenya yang selalu terpapar infeksi HIV selama 5 tahun melalui fluida reproduksi[6][7]
justru menunjukkan respon kekebalan tiruan sel T CD8 dan sel TH1[8] yang merespon berbagai macam epitop HIV tanpa disertai respon antibodi.
Selain itu, modus yang digunakan oleh virus HIV adalah pemotongan jalur informasi selular dengan menempel pada pencerap kemokina CCR5 dan CXCR4, selain pada CD4.[9] Pencerap CCR5 merupakan ekspresi dari sel dendritik, makrofaga dan sel T CD4. Ekspresi CXCR4 adalah pencerap pada sel T CD4 setelah teraktivasi.
Kompetisi pada area pencerap CCR5 oleh sekresi kemokina RANTES, MIP-1α, and MIP-1β menunjukkan respon kekebalan terhadap infeksi HIV.[10]