
BAB II TINJAUAN PUSTAKA 2.1. Tinjauan Tentang Kacang Gude
31
8 BAB II TINJAUAN PUSTAKA 2.1. Tinjauan Tentang Kacang Gude Tanaman kacang gude merupakan perdu dengan tinggi mencapai 3 m dan merupakan kacang tahunan dengan umur yang tidak terlalu panjang yaitu hanya 1- 5 tahun. Batang kacang gude berbulu halus dan bercabang banyak, daunnya ganda, beranak daun berjumlah tiga. Bunganya berbentuk kupu-kupu, berwarna jingga, ataupun kecoklatan berjumlah majemuk. Buahnya polong, dapat mencapai 7,5 cm, lurus atau membengkok seperti sabit, membulat, menjorong atau agak persegi. Biji kacang gude berwarna putih, krim, coklat, dan ungu kehitaman (Valenzuela dan Smith, 2002). Gambaran fisik kecambah kacang gude dapat dilihat pada Gambar 2.1., sedangkan klasifikasi kacang gude dapat dijelaskan sebagai berikut : Kingdom : Plantae Sub Kingdom : Viridiplantae Infa Kingdom : Streptophyta Super Divisi : Embryophyta Divisi : Tracheophyta Sub Divisi : Spermatophyta Kelas Divisi : Magnoliopsida Super Ordo : Rosanae Ordo : Fabales Famili : Fabaceae Genus : Cajanus adans Spesies : Cajanus cajan L. Mil. Sp.
Transcript of BAB II TINJAUAN PUSTAKA 2.1. Tinjauan Tentang Kacang Gude

8
BAB II
TINJAUAN PUSTAKA
2.1. Tinjauan Tentang Kacang Gude
Tanaman kacang gude merupakan perdu dengan tinggi mencapai 3 m dan
merupakan kacang tahunan dengan umur yang tidak terlalu panjang yaitu hanya 1-
5 tahun. Batang kacang gude berbulu halus dan bercabang banyak, daunnya ganda,
beranak daun berjumlah tiga. Bunganya berbentuk kupu-kupu, berwarna jingga,
ataupun kecoklatan berjumlah majemuk. Buahnya polong, dapat mencapai 7,5 cm,
lurus atau membengkok seperti sabit, membulat, menjorong atau agak persegi. Biji
kacang gude berwarna putih, krim, coklat, dan ungu kehitaman (Valenzuela dan
Smith, 2002). Gambaran fisik kecambah kacang gude dapat dilihat pada Gambar
2.1., sedangkan klasifikasi kacang gude dapat dijelaskan sebagai berikut :
Kingdom : Plantae
Sub Kingdom : Viridiplantae
Infa Kingdom : Streptophyta
Super Divisi : Embryophyta
Divisi : Tracheophyta
Sub Divisi : Spermatophyta
Kelas Divisi : Magnoliopsida
Super Ordo : Rosanae
Ordo : Fabales
Famili : Fabaceae
Genus : Cajanus adans
Spesies : Cajanus cajan L. Mil. Sp.

9
Terdapat dua macam varietas kacang gude yang dikenal petani di Indonesia,
yaitu:
1. Varietas Flavus (jenis Genjah) : umumnya lebih genjah(lekas berbuah dan
lebat), tanaman lebih pendek batangnya kekuningan, polongnya hijau
kalau telah masak berwarna terang, biasanya tiap polong berisi tiga biji.
2. Varietas Bicolor (jenis Umur panjang) : bersifat tahunan, tajuk lebih besar,
batangnya merah keunguan, polongnya berwarna gelap dan berbulu berisi
4-5 biji, kalau masak berwarna gelap.
.
Gambar 2.1. Kecambah kacang gude (Sumber : Koleksi Pribadi)
Kacang gude jenis genjah berbuah pada umur 12-14 minggu dan buah akan
masak dalam waktu 5-6 bulan. Kacang gude jenis umur panjang akan berbuah
lebih lambat dan memerlukan waktu 9-12 bulan untuk menghasilkan buah masak.

10
Dari segi kesehatan, kacang gude memiliki banyak kegunaan sebagai tanaman
obat tradisional yang merakyat, dan telah digunakan selama bertahun-tahun dalam
mengobati diabetes, mengeluarkan batu ginjal, menstabilkan periode menstruasi,
iritasi kulit, hepatitis, campak, sakit kuning, disentri dan penyakit lainya. Biji
kacang gude mengandung inhibitor protease (inhibitor terhadap tripsin,
kimotripsin), fitolektin, serta metabolit sekunder lainnya, yang berperan dalam
sistem pertahanan diri (Muangman et al., 2011).
Akhir-akhir ini, tanaman legum dipertimbangkan tidak hanya sebagai sumber
protein bernilai penting untuk fungsi tertentu dan pertumbuhan badan saja, tetapi
juga sebagai sumber senyawa bioaktif seperti peptida bioaktif, yang dapat
bermanfaat bagi kesehatan manusia dan melindungi pangan dari kerusakan.
Protein biji kacang gude telah difraksinasi sebagai protein albumin (larut
dalam air), globulin (larut dalam alkohol), glutelin (larut dalam asam atau basa),
protein sisa dan nitrogen non protein (Pande dan Pant tahun 1978 dalam Salunkhe
et al., 2009). Globulin berperan sebagai protein cadangan dalam kacang-kacangan.
Protein cadangan merupakan protein-protein yang terakumulasi selama fase
tertentu dalam perkembangan biji dan selanjutnya didegradasi untuk menyediakan
nitrogen untuk sintesa protein baru setelah perkecambahan. Untuk menjaga peran
tersebut, globulin ditemukan kaya dengan asam amino yang banyak mengandung
atom nitrogen seperti arginin, lisin, asparagin dan glutamin (Potter & Hotchkiss,
1998)
Albumin dan globulin dikenal merupakan komponen utama dalam protein
kacang-kacangan. Osborne dan Campbell adalah peneliti pertama yang menyatakan

11
dua kelompok utama yang terdapat dalam globulin yang diberinama vicilin dan
legumin. Kedua protein globulin ini memiliki perbedaan kelarutan, legumin
memiliki kelarutan lebih rendah dalam larutan garam dibandingkan dengan vicilin.
Selain itu, vicilin lebih mudah terkoagulasi oleh panas dalam larutan. Danielsson
(1949) mempelajari kecenderungan sedimentasi dari legumin dan vicilin dengan
metode ultrasentrifugasi, dan menemukan bahwa legumin dan vicilin memiliki
perbedaan koefisien sentrifugasi. Berdasarkan studi ini, legumin dan vicilin
selanjutnya lebih dikenal sebagai 11S globulin dan 7S globulin. Legumin tersusun
atas 6 sub unit masing-masing dengan berat molekul sekitar 60 kDa, sedangkan
berat molekul totalnya antara 300 sampai 400 kDa.
Albumin merupakan protein non cadangan dalam biji legum, tetapi beberapa
studi juga menunjukkan bahwa albumin mungkin berperan sebagai protein
cadangan. Berat molekul albumin pada biji kacang buncis berada dalam kisaran 11
sampai 105 kDa dengan 24 pita pada pola elektroforetik. Albumin dianggap sebagai
fraksi yang paling bergizi berdasarkan kandungan asam amino esensialnya yang
tinggi (Sathe and Venkatachalam, 2007).
Bagian kotiledon dari biji kacang gude merupakan sumber nutrien utama yang
dimanfaatkan oleh konsumen. Tabel 2.1. menunjukkan komposisi asam amino dari
beberapa protein yang berhasil difraksinasi dari kotiledon kacang gude. Seperti
protein legum pada umumnya, protein kacang gude rendah dalam kandungan asam
amino metionin, sistein, triptofan dan kaya dengan lisin (Salunkhe et al., 2009).

12
Tabel.2.1.
Komposisi asam amino (g/16g N) dari fraksi-fraksi protein dalam kotiledon
kacang gude
Jenis Asam Amino Albumin Globulin Glutelin Prolamin
Lisin 10,0 6,9 7,1 1,0
Histidin 3,5 3,4 4,3 1,0
Arginin 6,4 7,0 7,6 1,3
Asam Aspartat 13,9 10,8 11,8 3,9
Treonin 6,0 3,6 5,2 0,6
Serin 6,1 4,7 6,0 1,0
Asam Glutamat 24,3 22 25,1 15,9
Prolin 4,8 3,3 7,1 2,3
Glisin 5,9 4,1 4,8 1,3
Alanin 7,2 4,1 5,7 1,3
Sistein 3,2 0,9 1,0 0,4
Valin 6,2 5,5 5,8 1,8
Metionin 1,7 0,8 1,3 0,3
Isoleusin 4,1 3,4 5,1 0,7
Leusin 7,7 6,7 9,2 0,8
Tirosin 4,2 3,9 3,9 0,9
Fenilalanin 4,7 10,9 8,0 6,5
Sumber : Singh and Jambunathan, 1982.
2.2. Asam Amino dan Peptida
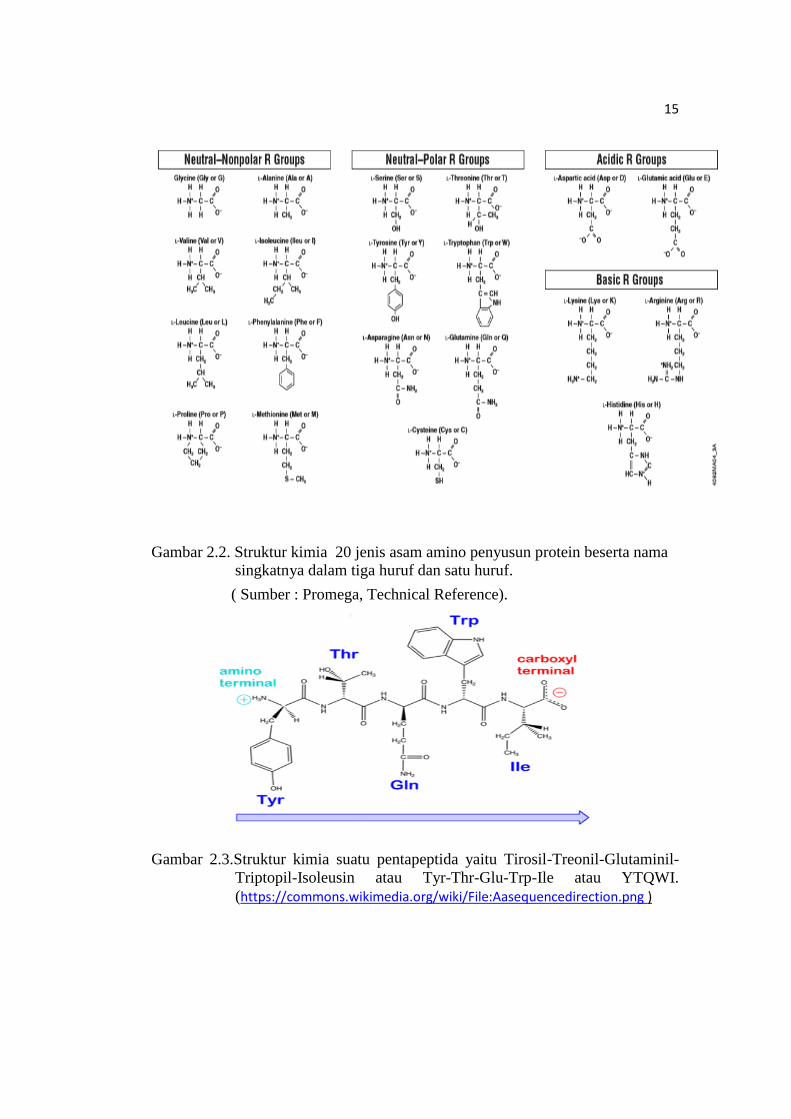
Terdapat 20 jenis asam amino yang berfungsi sebagai unit pembangun
(monomer) molekul protein, yang dibedakan satu sama lain oleh struktur kimia dari
rantai sampingnya (jenis gugus R yang terikat pada atom Cα), seperti dapat dilihat
pada Gambar 2.2. Berdasarkan sifat kepolaran gugus R (rantai sampingnya)
tersebut, ke 20 jenis asam amino dapat diklasifikasikan menjadi 4 kelompok
yaitu :
1. Asam amino netral dengan gugus R non polar (hidrofobik) yaitu alanin, leusin,
isoleusin, valin, prolin, fenilalanin dan triptofan dan metionin.

13
2. Asam amino netral dengan gugus R polar (hidrofilik) yaitu : serin, treonin,
tirosin, asparagin, glutamin, sistein.
3. Asam amino asam (gugus R bermuatan negatif) karena memiliki gugus R
karboksil yang bermuatan negatif pada pH 6,0 –7,0 yaitu asam glutamat dan
asam aspartat.
4. Asam amino basa (gugus R bermuatan positif) karena memiliki gugus R amina
yang bermuatan positif pada pH 6,0 – 7,0 yaitu lisin dan arginin.
Peptida merupakan produk dari hasil kondensasi beberapa asam amino atau
juga merupakan hasil hidrolisis parsial dari protein. Peptida memiliki berat molekul
dan ukuran lebih kecil daripada protein induknya. Peptida – peptida yang dihasilkan
dari hidrolisis protein dapat memiliki sifat kimia yang bervariasi karena peptida-
peptida tersebut memiliki variasi dalam hal panjang rantainya, komposisi asam
aminonya, dan fungsionalitasnya. Sifat fisikokimia dari peptida–peptida tersebut
meliputi kelarutan, ukuran, hidrofobisitas permukaan, muatan, sifat asam basa, dan
kemampuannya dalam mengkelat logam. Sifat fisikokimia ini sangat tergantung
pada keberadaan dan sifat kimia dari gugus fungsi yang membentuk rantai samping
asam-asam amino penyusunnya. Terdapat asam amino yang memiliki rantai
samping berupa gugus fungsi yang dapat membentuk ion (seperti gugus karboksil,
gugus amino, gugus tiol), gugus fungsi hidrofobik seperti gugus alkil, dan gugus
netral hidrofilik (seperti gugus amida). Suatu peptida memang mengandung gugus
karboksil-α dan gugus amino-α bebas pada ujung kanan dan ujung kirinya, akan
tetapi gugus-gugus ini hampir tidak berperan tergantung pada konformasi peptida
dan bagaimana posisi dari gugus-gugus ini, apalagi jika peptidanya berbentuk

14
siklik. Beberapa ion logam transisi seperti tembaga (Cu2+) dan nikel (Ni2+), dapat
membentuk kompleks dengan beberapa gugus fungsi dalam peptida dan asam
amino seperti asam glutamat dan histidin, dan membatasi fungsionalitas dari
gugus-gugus fungsi tersebut (Barrett dan Elmore, 1998).
Pada Gambar 2.3, ditampilkan contoh struktur kimia dari suatu pentapeptida
dengan urutan asam amino Tyr-Thr-Glu-Trp-Ile atau YTQWI. Tampak bahwa
dalam struktur peptida tersebut, gugus α-amino dan gugus α-karboksil dari asam-
asam amino yang menyusun bagian dalam rantai peptida tidak akan bebas lagi
membentuk ion karena sudah digunakan untuk membentuk ikatan peptida, yang
masih bebas membentuk ion adalah gugus α-amino dari asam amino yang terletak
di ujung kiri (Amino-terminal end) dan gugus α-karboksil dari asam amino di ujung
kanan (Carboxyl-terminal end) dalam rantai peptida.
2.3. Manfaat Kesehatan Peptida Bioaktif
Peptida bioaktif telah didiskusikan secara luas dalam komunitas ilmiah sebagai
salah satu kelompok nutrasetikal yang menjadi perhatian dalam bidang pangan dan
gizi. Banyak literatur yang melaporkan tentang bioaktivitas peptida secara in vitro
dan berbagai jenis aktivitas telah dilaporkan, meliputi sifat antimikroba,
antikarsinogenik, antiinflamasi, antihipertensi, kemampuan menurunkan
kolesterol, antitrombotik, aktivitas antioksidan, meningkatkan absorpsi/
bioavailabilitas mineral, dan aktivitas opioid (Malaguti et al., 2014). Beberapa dari
peptida-peptida tersebut ditemukan bersifat bifungsional, misalnya satu jenis
peptida bersifat aktif antioksidan dan sekaligus aktif antihipertensi
15
Gambar 2.2. Struktur kimia 20 jenis asam amino penyusun protein beserta nama
singkatnya dalam tiga huruf dan satu huruf.
( Sumber : Promega, Technical Reference).
Gambar 2.3.Struktur kimia suatu pentapeptida yaitu Tirosil-Treonil-Glutaminil-
Triptopil-Isoleusin atau Tyr-Thr-Glu-Trp-Ile atau YTQWI.
(https://commons.wikimedia.org/wiki/File:Aasequencedirection.png )

16
Mellander (1950) dalam Wang & Gonzales (2005) merupakan ahli yang
bertanggung jawab pertama kali mempelajari hubungan antara peptida bioaktif
yang dihasilkan dari hidrolisis protein kasein dengan peningkatan kalsifikasi tulang
pada bayi yang menderita rakhitis. Berdasarkan database dari Bipep dan BioPD
(Bioactive Peptide Database), menyatakan bahwa lebih dari 1200 peptida bioaktif
berbeda telah tercatat ditemukan. Data beberapa jenis peptida bioaktif (dengan
berbagai aktivitas biologis) yang diturunkan dari makanan beserta urutan asam
aminonya dapat dilihat pada Tabel 2.2.
Dalam pengajaran ilmu gizi dan metabolisme zat gizi, diberikan pengertian
bahwa protein yang dikonsumsi akan dihidrolisis oleh enzim-enzim pencernaan dan
akhirnya menjadi asam-asam amino; dan zat gizi inilah yang kemudian akan diserap
oleh usus halus lalu masuk ke dalam peredaran darah. Hasil-hasil penelitian
kinetika mengenai pencernaan susu yang dilakukan belakangan ini menunjukkan
bahwa peptida bioaktif masih terdapat dalam usus halus walaupun protein susu
telah dicerna oleh enzim-enzim pankreatik. Chabance et al. (1998) menemukan
bahwa dua macam peptida yang berasal dari protein susu dan yoghurt yang
dikonsumsi, yaitu κ-kasein-glikopeptida dan peptida N-terminal dari αS1-kasein
dapat dideteksi dalam plasma darah. Hasil penelitian ini memberikan bukti bahwa
peptida bioaktif dari protein susu dapat diserap oleh usus halus (tidak harus diubah
terlebih dahulu menjadi asam amino), sehingga kemudian dapat menunjukkan
aktivitas fisiologis dalam bermacam-macam organ tubuh.
Untuk dapat melaksanakan fungsinya, sesungguhnya peptida bioaktif tidak
selalu harus diserap oleh usus dan masuk ke dalam peredaran darah, karena

17
senyawa tersebut dapat pula bekerja di dalam usus. Sebagai contoh peptida
”penekan nafsu makan” (anorectic, appetite suppressant) bekerja di dalam usus, di
mana mereka menstimulir opioid dan reseptor hormon serta menginduksi perasaan
kenyang, sedangkan peptida bioaktif lain yang mempunyai aktivitas hipotensif atau
anti kanker harus diserap oleh usus halus dan masuk ke dalam peredaran darah,
untuk selanjutnya ditransportasikan ke organ target.
Dalam beberapa penelitian tentang pendekatan diet untuk mengontrol atau
mencegah penyakit-penyakit degeneratif kronis, peptida bioaktif yang diturunkan
dari protein biji-bijian dapat menjadi salah satu sumber komponen yang dapat
meningkatkan kesehatan. Proses fermentasi biji-bijian yang menghasilkan peptida
bioaktif ditunjukkan pada produk fermentasi dari kedelai seperti natto, tempe, dan
douchi yang ditemukan mengandung peptida bioaktif antioksidan, antihipertensi
dan antijamur. Hasil penelitian menunjukkan bahwa teknik pengolahan makanan
mempengaruhi aktivitas antihipertensi dari produk-produk berbasis kedelai.
Peptida bioaktif merupakan unsur penting pada berbagai produk maupun
ingredien yang dipasarkan sebagai makanan fungsional maupun nutrasetikal. Pada
produk-produk ini, peptida bioaktif ditambahkan atau diperkaya dengan cara
modifikasi proses manufaktur yang sudah umum (misalnya dengan mengubah
parameter proses atau kultur starter yang digunakan). Namun beberapa dari produk
ini merupakan makanan tradisional yang sekarang ditawarkan dengan strategi
pemasaran yang berbeda. Food-derived bioactive peptides juga memiliki potensi
yang tinggi digunakan sebagai ingredien pada farmasetikal (obat). Sebagai
contohnya yaitu : kapsul penurun tekanan darah telah dikomersialisasi yang

18
mengandung oligopeptida Katsuobushi LKPNM yang diproduksi dari ikan bonito
kering menggunakan enzim termolisin, yang dikonversi menjadi bentuk aktifnya
yaitu peptida LKP oleh enzim pencernaan.
Beberapa jenis peptida penghambat ACE telah sukses dikomersialisasi sebagai
suplemen bioaktif yang ditambahkan pada matrik makanan seperti susu asam (sour
milk) dengan merk Calpis, yang diproduksi oleh Calpis Food Industry co, Ltd.
Tokyo, Jepang, diketahui mengandung peptida IPP (15 mg/L) dan VPP (11 mg/L)
(Hata et al., 1996). IPP and VPP juga diformulasi ke dalam susu asam merk
Evolus® pada konsentrasi berturut-turut 15 mg/L and 20 mg/L, dan
dikomersialisasi oleh Valio Ltd, Valio, Finlandia (Seppo et al., 2003).
2.4. Peptida Bioaktif Penghambat ACE dan Antihipertensi
Beberapa jenis peptida juga memiliki aktivitas sebagai antihipertensi.
Hipertensi mempengaruhi hampir seperempat populasi dunia, dan merupakan
faktor resiko yang utama dan belum terkendali dalam penyakit kardiovaskular dan
komplikasinya. Tekanan darah merupakan suatu keseimbangan antara output
jantung (cardiac output) dan resistensi vascular peripelar, yang dikontrol oleh
suatu sistem yang kompleks yang meliputi Renin Angiotensin System (RAS),
sistem syaraf simpatetik, ginjal dan mekanisme keseimbangan cairan (Hernández-
Ledesma et al., 2011).

19
Tabel 2.2.
Manfaat kesehatan dari berbagai peptida bioaktif yang diturunkan dari makanan
(Hernandez-Ledesma et al., 2011)
Aktivitas
Biologis
Sumber
Makanan
Protein Induk Nama Peptida/
Urutan Asam Amino
(singkatan satu huruf)
Antihipertensi Kedelai Protein Kedelai NWGPLV
Ikan Protein Otot Ikan LKP, IKP, LRP
Daging Protein otot
daging
IKW, LKP
Susu α-Laktalbumin,
β-Laktoglobulin
Laktokinin (misalnya
WLAHK, LRP, LKP)
Brokoli Protein Tanaman YPK
Antimikroba Telur Ovotransferin OTAP-92(fraksi109-200)
Susu Laktoferin Laktofericin
Kacang buncis Protein Legum Vulgarinin (Antijamur)
Pengikatan
Mineral
Susu α-, β-kasein Caseinophosphopeptide
disingkat CPP.
Anti Kolesterol Kedelai Glisinin LPYPR
Antioksidan Ikan Otot Sardin MY
Susu α-Laktalbumin,
β-Laktoglobulin
MHIRL, YVEEL,
WYSLAMAAASDI
Antitrombotik Susu κ-kasein κ-kasein (fraksi 106-116)
RAS merupakan sistem vasokontriksi yang kuat dalam sistem mamalia yang
tidak hanya bertanggung jawab untuk mempertahankan level tekanan darah, namun
juga mempengaruhi keseimbangan cairan dan garam sebagai produk akhir dari
Angiotensin II. Jalur utama dimulai dari degradasi Angiotensinogen, suatu peptida
precursor dari hati, oleh renin, dan melepaskan dekapeptida Angiotensin I.
Angiotensin I selanjutnya dipotong pada dipeptida ujung karboksilnya oleh aksi
Angiotensin I - Converting Enzyme (ACE) pada bagian luminal dari endothelium
vascular, untuk membentuk oktapeptida Angiotensin II. Fungsi fisiologis dari
Angiotensin II ditentukan karena kemampuannya untuk terikat pada dua reseptor

20
yaitu, reseptor Angiotensin II tipe 1 (AT1) dan reseptor Angiotensin II tipe 2 (AT2)
(Eriksson et al., 2002). Melalui ikatan langsung dengan reseptor AT1, maka
angiotensin II mampu menstimulasi sekresi aldosteron dan menginduksi
vasokontriksi (pengkerutan pembuluh darah), dan juga mempromosi reabsorpsi
natrium dan air dalam ginjal, dan bahkan meningkatkan tekanan darah. Namun,
stimulasi yang berlebihan pada reseptor AT1 dapat menginduksi suatu mekanisme
penyeimbang yang dikirimkan oleh reseptor AT2 yang menyebabkan vasodilatasi
dan penghambatan pertumbuhan sel. ACE tidak hanya mengkatalisa konversi
Angiotensin I menjadi vasokontsriktor Angiotensin II, ACE juga mampu
mengkatalisa degradasi bradikinin menjadi peptida yang tidak aktif. Bradikinin
menstimulasi reseptor Bradikini B2 untuk menginduksi produksi nitrat oksida
(NO) dan prostasiklin, yang merupakan vasodilator yang kuat. Dengan demikian,
penghambatan ACE memiliki peran ganda yaitu mengurangi pembentukan
Angiotensin II dan juga mencegah degradasi vasodilator, yang secara keseluruhan
menyebabkan penurunan tekanan darah.
Stimulasi terhadap sistem RAS menghasilkan suatu peningkatan tekanan darah
yang akut pada level sirkulasi. Selain itu, jika sistem RAS diaktivasi, dapat
mengakibatkan efek merusak jangka panjang pada organ-organ akhir dengan
menginduksi proses-proses yang mendorong proses pertumbuhan dan fibrotik pada
tingkat jaringan. Untuk mencapai penghambatan yang efektif terhadap sistem RAS
pada tingkat jaringan, maka dibutuhkan penghambat ACE dengan dosis yang lebih
tinggi, dan kombinasi dengan terapi lain yang direkomendasikan selama treatmen
ACE inhibitor, seperti Angiotensin II receptor blockers (Weinberg et al., 2000).

21
Terdapat beberapa jenis ACE inhibitor sintetis yang telah sukses
dikomersialisasi, sebagai contohnya adalah captopril yang telah sukses digunakan
dalam praktik klinis. Namun captopril dan inhibitor sintetis ini, memiliki efek
samping yang signifikan seperti batuk, eksantema, perubahan rasa, ruam kulit,
gangguan lambung dan edema pada bibir. Akibat efek samping ini, maka
dibutuhkan ACE inhibitor yang dapat diperoleh secara alami dari makanan sebagai
pengganti ACE inhibitor sintetis. Peptida penghambat ACE alami dari makanan
tidak memiliki efek samping seperti obat sintetis. Peptida alami memiliki daya
hambat yang lebih rendah daripada ACE inhibitor sintetis (yang dapat memiliki
nilai IC50 pada daerah nM). Mengingat daya hambat yang lebih rendah ini, hal ini
Karena peptida tersebut menghambat ACE pada tingkat yang lebih rendah, ini
berpotensi memungkinkan tingkat bradykinin yang lebih aman dalam tubuh.
Dengan demikian, untuk alasan ini, peptida penghambat ACE telah menarik
perhatian sebagai agen pencegahan potensial untuk kontrol hipertensi (Hernandez-
Ledesma, et al., 2011).
2.4.1. Tinjauan Tentang Enzim ACE
Angiotensin converting enzyme (ACE) termasuk Zn-metaloproteinase. Atom
Zn berikatan dengan tiga gugus asam amino dan satu molekul air. Aktivitas
pengikatan inhibitor ACE terhadap ACE berdasarkan interaksi yang kuat antara
atom Zn dan gugus pengkelat inhibitor ACE (Ghassem et al., 2012). Struktur tiga
dimensi ACE ditunjukkan pada Gambar 2.4.

22
Gambar 2.4. Struktur tiga dimensi ACE (Sumber: PDB 1086).
Sisi aktif ACE memiliki tiga sub sisi yakni S1 (antepenultimate), S1’
(penultimate) dan S2 (ultimate) yang memiliki karakter berbeda dalam mengikat
tiga asam amino bagian C-terminal dari substrat atau inhibitor, berada pada dua sisi
aktif yang homolog. ACE lebih menyukai substrat atau inhibitor kompetitif yang
mengandung asam amino hidrofobik pada ketiga posisi C-terminal. Untuk interaksi
antara enzim dan inhibitor, tiga sub sisi dari sisi aktif enzim dengan sekuen asam
amino yang berbeda harus berikatan dengan substrat. Pengikatan inhibitor atau
substrat terhadap enzim secara umum terjadi pada tripeptida bagian C-terminal.
ACE (Angiotensin I-converting enzyme) (EC 3.4.15.1) merupakan suatu enzim
kunci yang mengatur tekanan darah melalui sistem RAS (Renin-Angiotensin
Sistem). ACE merupakan suatu enzim dipeptidil karboksidase, yang mengkatalisa
konversi Angiotensin I (dekapeptida) menjadi Angiotensin II (oktapeptida) yang
merupakan suatu vasokonstriktor yang ampuh dan memiliki peran fisiologis

23
penting dalam pengaturan tekanan darah dan cairan serta keseimbangan garam pada
mamalia. Selain itu ACE juga berperan dalam mengkatalisa degradasi bradikinin
(suatu vasodilator) menjadi peptida in aktif. Kedua peran ACE dalam sistem RAS
inilah yang menjadi alasan bahwa aktivasi terhadap ACE secara keseluruhan dapat
meningkatkan tekanan darah. Angiotensin converting enzyme (ACE) bersifat non
spesifik dan memotong unit-unit dipeptida pada substrat yang mengandung banyak
asam amino. Konversi angiotensin I menjadi angiotensin II yang cepat secara in
vivo terjadi melalui aksi ACE yang terdapat pada lumen permukaan sel endotelial
menembus ke sistem vaskuler. Sistem renin angiotensin terdiri atas tiga komponen,
yakni renin, angiotensinogen dan ACE. Renin merupakan protease utama, mampu
menentukan laju produksi angiotensin II. Gambar 2.5. menunjukkan peran ACE
dalam sistem RAS.
2.4.2. Peptida Bioaktif Antihipertensi.
Sejumlah besar peptida-peptida penghambat ACE telah diisolasi dari hasil
pencernaan berbagai protein makanan, terutama pada susu, ikan dan daging.
Peptida penghambat ACE pada umumnya memiliki rantai pendek, seringkali
membawa asam amino non polar seperti prolin. Peptida antihipertensi (hipotensif)
dan immunomodulator yaitu Val-Pro-Pro dan Ile-Pro-Pro sebagai contohnya, dapat
dilepaskan dari prekursor protein (β-kasein dan κ-kasein) oleh enzim dari
Lactobacillus helveticus.

24
Gambar 2.5. Pengaturan tekanan darah oleh enzim ACE dalam sistem RAS
(Li et al., 2004)
Cushman & Cheung (1971) telah membagi senyawa penghambat (inhibitor)
ACE ke dalam empat kelompok yaitu, reagen pengkelat, reagen sulfidril, ion logam
berat dan peptida. Reagen pengkelat seperti EDTA bekerja sebagai inhibitor
melalui interaksinya dengan ion Zn (yang merupakan kofaktor penting bagi ACE).
Reagen pengkelat dan reagen sulfidril mampu mengkelat ion Zn, sedangkan
inhibitor ion logam berat (misalnya CdBr2) bekerja melalui mekanisme penukar ion
dengan cara menggantikan ion Zn. Inhibitor kelompok terakhir yaitu peptida
bekerja dengan cara dengan mekanisme sebagai inhibitor kompetitif karena
memiliki struktur yang mirip dengan substrat alami dari ACE yang juga merupakan
peptida yaitu Angiotensin I (suatu dekapeptida).

25
Studi aktivitas penghambatan ACE secara in vitro dilakukan dengan cara
memonitor berlangsungnya reaksi konversi oleh enzim ACE menggunakan substrat
spesifik (HHL) akibat keberadaan dan ketidakberadaaan inhibitor (Ledesma et al.,
2011). Cushman & Cheung (1971) mengembangkan metode uji penghambatan
ACE menggunakan hippuryl-L-histidyl-L-leucine (HHL) sebagai substrat. Metode
ini melibatkan tahap ekstraksi menggunakan pelarut etil asetat terhadap produk
asam hipurat (HA) yang dilepaskan oleh aktivitas ACE pada waktu tertentu. Jumlah
asam hipurat yang terbentuk akibat aktivitas ACE ini ditentukan secara
spektrofotometri dengan pembacaan absorbansi pada 228 nm. Satu unit aktivitas
ACE dapat didefinisikan sebagai produksi satu μmol asam hipurat selama 1 menit
pada 37 °C.
Peptida penghambat ACE dapat bekerja dalam tiga mekanisme dan
diklasifikasikan sebagai peptida type inhibitor, type substrat dan type prodrug
(berdasarkan perubahan aktivititas penghambatan ACE-nya setelah hidrolisis
peptida oleh enzim ACE itu sendiri mengingat enzim ACE sendiri merupakan
enzim peptidase yang dapat menghidrolisis ikatan peptida tertentu). Peptida type
inhibitor merupakan peptida yang aktivitas penghambatan ACEnya tidak signifikan
mengalami perubahan karena peptida tersebut resistan terhadap pemecahan oleh
ACE. Peptida type substrat menunjukkan sedikit penurunan aktivitas
penghambatan ACE karena peptida ini telah mengalami degradasi oleh ACE.
Peptida type prodrug mengacu pada peptida tertentu yang akan menghasilkan
peptida lebih pendek (akibat pemotongan oleh ACE itu sendiri) yang memiliki

26
aktivitas penghambatan ACE kuat. Peptida jenis prodrug cenderung menghasilkan
peptida yang memberi efek hipotensif yang tahan lama secara in vivo.
2.5. Tinjauan Tentang Perkecambahan
Perkecambahan merupakan suatu proses dimana radikula (akar embrionik)
memanjang ke luar menembus kulit biji. Di balik gejala morfologi dengan
pemunculan radikula tersebut, terjadi proses fisiologi-biokimia yang kompleks,
dikenal sebagai proses perkecambahan fisiologis (Salisbury, 1985).
Perkecambahan dimulai dengan proses penyerapan air ke dalam sel-sel. Proses ini
merupakan proses fisika. Masuknya air pada biji menyebabkan enzim aktif bekerja.
Bekerjanya enzim merupakan proses kimia. Enzim amilase bekerja memecah
tepung menjadi maltosa, selanjutnya maltosa dihidrolisis oleh maltase menjadi
glukosa. Protein juga dipecah menjadi asam-asam amino. Perkecambahan biji
berhubungan dengan aspek kimiawi. Proses tersebut meliputi beberapa tahapan,
antara lain imbibisi, sekresi hormon dan enzim, hidrolisis cadangan makanan,
pengiriman bahan makanan terlarut dan hormon ke daerah titik tumbuh atau daerah
lainnya, serta fotosintesis. Proses penyerapan cairan pada biji (imbibisi) terjadi
melalui mikropil. Air yang masuk ke dalam kotiledon menyebabkan volumenya
bertambah, akibatnya kotiledon membengkak. Pembengkakan tersebut pada
akhirnya menyebabkan pecahnya testa. Bagian-bagian penting dari kecambah dapat
dilihat pada Gambar 2.6.

27
Gambar 2.6. Bagian-bagian dari kecambah
Sumber : Bewley, J. D. and M. Black., 1978.
Untuk meningkatkan nilai nutrisi, daya cerna dan juga untuk mengurangi faktor
antinutrisi dari kacang-kacangan, maka sejumlah metoda dapat diaplikasikan
meliputi perendaman, pengupasan, pemanasan, perkecambahan (germinasi) dan
fermentasi. Perkecambahan merupakan proses biologis di mana tanaman
memulihkan kembali aktivitasnya untuk mencapai potensinya sehingga enzim-
enzimnya menjadi sangat aktif. Perkecambahan dapat menyebabkan perubahan
yang signifikan dalam karakteristik biokimia biji-bijian dan selama proses tersebut,
protein-protein cadangan dapat didegradasi oleh protease-protease endogen yang
aktif dalam fase perkecambahan. Teknik ini merupakan proses yang tidak mahal
dan efektif yang dapat digunakan untuk meningkatkan nilai nutrisi dari biji-bijian
melalui peningkatan daya cerna protein dan penurunan senyawa-senyawa
antinutrisi (Rocha et al., 2015).

28
2.6. Produksi Peptida Bioaktif dengan Enzim Protease.
Peptida bioaktif dapat dilepaskan dari protein induknya melalui hidrolisis
enzimatik selama pencernaaan gastrointestinal, fermentasi maupun pematangan
selama pengolahan pangan atau proteolisis oleh enzim-enzim food grade yang
berasal dari mikroba, hewan maupun tanaman. Jika sekuen asam amino dalam
peptidanya telah diketahui, kemungkinan juga peptida tersebut dapat disintesis
secara kimia maupun enzimatis atau melalui teknologi DNA rekombinan.
Produk hasil hidrolisis protein disebut dengan hidrolisat protein yang
mengandung suatu campuran komponen berbagai jenis peptida rantai pendek
maupun asam amino bebas, tergantung pada derajat hidrolisisnya apakah secara
parsial maupun sampai sempurna. Dari berbagai jenis rantai peptida yang terdapat
dalam hidrolisat protein ini, beberapa di antaranya memiliki aktivitas biologis
sehingga disebut peptida bioaktif. Sumber kandidat protein yang potensial sebagai
protein induk yang mempunyai aktivitas biologis laten untuk menghasilkan peptida
bioaktif dapat bersumber dari jaringan atau bagian organisme yang kaya akan
protein seperti susu, telur, daging, ikan dan juga dari protein nabati dari tanaman.
Telah dilaporkan bahwa keuntungan tambahan dari hidrolisis protein adalah
peningkatan hidrofobisitas karena proteolisis membuka lipatan dalam rantai.
Pemutusan ikatan peptida meningkatkan jumlah gugus amino dan karboksil bebas
sehingga meningkatkan kelarutan protein. Selain itu, hidrolisis yang memproduksi
peptida pendek dengan berat molekul di bawah 1000 Da memiliki rasa yang kurang
pahit. Akan tetapi, telah dilaporkan bahwa hidrolisis ekstensif dapat menyebabkan
efek yang sebaliknya yang mempengaruhi sifat fungsional dari peptida-peptida.

29
Beberapa faktor yang mempengaruhi produksi peptida bioaktif meliputi, waktu
hidrolisis, derajat hidrolisis protein, rasio enzim-substrat, dan pretreatmen protein
sebelum hidrolisis. Sebagai contohnya, perlakuan termal protein dapat
meningkatkan efektitivitas hidrolisis enzimatis, kemungkinan disebabkan karena
terjadi peningkatan interaksi enzim-substrat karena pemanasan memicu pembukaan
lipatan protein. Enzim-enzim protease industri seperti Alkalase, Flavourenzyme,
dan Protamex dari mikroba, dan juga enzim-enzim dari tanaman (seperti papain)
dan hewan (misalnya pepsin dan tripsin), telah digunakan secara luas untuk
memproduksi peptida antihipertensi dan antioksidan. Berdasarkan posisi
pemotongan ikatan peptidanya, enzim protease dan peptidase dibedakan menjadi
endoprotease yang memotong ikatan peptida di dalam rantai protein, dan
eksoprotease yang memotong ikatan peptida dari ujung rantai protein. Sebagian
besar jenis protease merupakan endoprotease, sedangkan yang tergolong
eksoprotease yaitu jenis karboksipeptidase dan aminopeptidase. Masing-masing
jenis protease memiliki spesifitas yang berbeda, yaitu jenis ikatan peptida tertentu
yang lebih disukai untuk dihidrolisis.
Dalam Tabel 2.3 ditampilkan beberapa jenis protease yang dilengkapi dengan
jenis residu asam amino yang disukai (yang mengapit ikatan peptida yang akan
dipotong) (Mathew dan Van Holde, 1996).
2.7. Pencernaan Gastrointestinal in vitro
Hidrolisis protein pangan oleh kombinasi enzim-enzim pencernaan merupakan
strategi umum untuk meniru proses pencernaan in vivo. Dengan menggunakan
pepsin, tripsin, α-kimotripsin, atau pankreatin, protein-protein mengalami

30
degradasi yang mirip seperti pada sistem pencernaan manusia, yaitu melepaskan
peptida-peptida yang diharapkan akan resistan terhadap pencernaan in vivo
(Vermeirssen et al., 2005). Contoh penelitian sejenis yang menggunakan enzim-
enzim gastrointestinal di antaranya adalah, pepsin dan pankreatin digunakan untuk
menghidrolisis protein biji matahari (Megías et al., 2004).
Enzim pepsin merupakan protease yang terdapat di lambung (gastro), memiliki
pH optimum 1,0 -3,0 dan memiliki berat molekul 34,6 kDa. Pepsin lebih menyukai
memotong ujung C protein dari fenilalanin (Phe), leusin (Leu), tirosin (Tyr) dan
triptofan (Trp) (Promega, 2016). Pankreatin merupakan suatu campuran enzim
yang dihasilkan oleh pankreas mamalia. Enzim-enzim ini meliputi enzim-enzim
proteolitik (tripsin, kimotripsin, karboksipeptidase dan pankropeptidase), amilase,
dan lipase (Frey and Hegeman, 2010). Aktivitas enzim-enzim pankreatin terbaik
diamati pada pH 6,5 – 9,0 dan suhu 37 – 40 oC. Untuk aktivitas proteolitik maka
yang dikenal peranannya adalah tripsin dan kimotripsin. Diklasifikasikan sebagai
serin protease karena memiliki residu serin pada sisi aktifnya, tripsin merupakan
enzim pankreatik yang paling terkenal untuk aktivitas proteolitiknya. Tripsin
terutama memotong peptida dari asam amino basa, khususnya lisin dan arginin.
Pemotongan ini biasanya terjadi pada ujung karboksil dari asam-asam amino
tersebut. Jika suatu residu prolin mengikuti asam amino basa, maka tidak akan
terjadi hidrolisis oleh tripsin. Kimotripsin juga merupakan suatu serin protease.
Proteolisis terutama oleh kimotripsin teramati terutama pada ujung karboksil dari
residu tirosin, triptofan, dan fenilalanin yang semuanya merupakan asam amino
aromatik. Rantai samping aromatik ini (dengan struktur cincinnya dapat secara

31
tepat menempati kantung hidrofobik (sisi aktif) dari kimotripsin. Sisi pemotongan
lainnya meliputi ikatan amida dekat ujung karboksil dari leusin, meskipun katalisis
ini relatif bertahap.
Tabel 2.3.
Spesifitas beberapa jenis enzim protease (jenis asam amino yang disukai)
No Nama Enzim Posisi ikatan yang disukai (Preferred Site)
/Jenis asam amino yang mengapit ikatan peptida
1 Tripsin R1 = Lys, Arg. R2 ≠ Pro
2 Kimotripsin R1 = Tyr, Phe, Leu, Ile, Val, Trp, dan His. R2 ≠ Pro
3 Pepsin R1 = Phe, Leu, dan banyak lainnya. R2 ≠ Pro
4 Karboksipeptidase A R 2 = asam amino C terminal (ujung kiri dari rantai).
R1 ≠ Pro
5 Trombin
(pembekuan darah)
R1 = Arg. R2 ≠ Pro
6 Papain R1 = Arg, Lys, Phe-X (bagian CO dari asam amino
setelah Phe)
7 Bromelain R1 = Lys, Ala, Tyr, Gly.
8 Termolisin R2 = sama dengan kimotripsin, R1 ≠ Pro
Sumber: Mathew dan Van Holde, 1996.
2.8. Fraksinasi Peptida Bioaktif
Umumnya, kandungan peptida dalam makanan tidak melimpah seperti yang
diharapkan. Selain itu, kehadiran senyawa lain nonpeptida (yaitu, lipid, gula) juga
dapat mengganggu dalam analisis peptida. Oleh karena itu, dalam prakteknya sulit
untuk menganalisis peptida makanan dengan akurasi yang baik tanpa melakukan
langkah persiapan sampel. Persiapan sampel ini dapat terdiri dari beragam prosedur
untuk isolasi, pemurnian, dan prakonsentrasi analit, lebih dari satu langkah yang

32
diperlukan dalam banyak kasus. RP-LC (Reverse Phase-Liquid Chromatography)
dan CE (Capillary Electrophoresis) merupakan metode analitik dasar yang
digunakan untuk analisis peptidom makana secara kemometrik (Minkiewicz et al.,
2008 dalam Martínez-Maqueda et al., 2013).
Secara umum, sampel makanan pertama-tama diberikan tahap pembersihan
sampel awal untuk menghilangkan zat yang mengganggu dan kemudian, langkah
fraksinasi yang berbeda diterapkan. Analisis peptida langsung pada sampel
makanan tanpa tahap persiapan apapun jarang dilaporkan dalam literatur. Ekstraksi
peptida biasanya diikuti dengan pembersihan sampel awal untuk menghilangkan
komponen makanan lainnya (yaitu, protein dan lipid).
Deproteinisasi, merupakan prosedur pembersihan awal yang paling penting
dalam analisis peptida, dilakukan oleh pengendapan protein menggunakan
beberapa agen. Deproteinisasi juga bisa bertindak sebagai prosedur fraksinasi untuk
peptida karena kelarutannya tergantung pada agen pengendap dan proporsinya.
Setelah pengendapan, metode sentrifugasi dan filtrasi digunakan untuk
memisahkan protein dari peptida terlarut. Selain itu, penerapan perlakuan panas
atau langkah ultrasentrifugasi dengan kecepatan tinggi untuk menghilangkan
protein telah dilaporkan. Selektifitas pengendapan secara langsung tergantung pada
jenis bahan pengendap yang digunakan. Selain penggunaan pelarut organik seperti
etanol, metanol, atau aseton, larutan yang mengandung asam seperti TCA atau asam
fluoroasetat (TFA) merupakan pengendap protein yang klasik. Pengendapan
saltingout, berdasarkan polaritas, dengan konsentrasi garam atau pengendapan yang
tinggi dengan menyesuaikan pH ke pI (titik isoelektrik) protein merupakan opsi

33
lainnya. Contoh representatif ditemukan dalam isolasi dan identifikasi suatu peptida
penghambat (ACE) dari biji soba (buckwheat) utuh setelah langkah-langkah
pencucian dengan ekstraksi dietil eter untuk menghilangkan sebagian besar
kandungan lemak dan deproteinisasi dengan menyesuaikan pH ke pI protein soba
(Ma et al. 2006 dalam Martínez-Maqueda et al., 2013). Pada beberapa kasus,
penerapan homogenisasi dan atau deproteinisasi cukup untuk analisis peptida lebih
lanjut. Sayangnya, sebagian besar sampel memerlukan langkah tambahan untuk
mencapai isolasi dan konsentrasi peptida yang sesuai sebelum analisis.
Metode filtrasi membran merupakan suatu proses fisik untuk memisahkan
komponen-komponen dengan menggunakan bahan membran yang memungkinkan
lewatnya komponen terpilih tergantung pada sifat bahan dari membrannya.
Kelompok filtrasi membran yang paling umum meliputi mikrofiltrasi (MF),
ultrafiltrasi (UF), nanofiltrasi (NF) dan Reverse Osmosis (RO). Faktor yang
membedakan kelompok filtrasi membran ini yaitu mekanisme pemisahan secara
keseluruhan (yaitu ukuran, muatan atau keduanya), sifat material membran, gaya
dorong untuk pemisahan (yaitu vakum atau tekanan), tujuan utama filtrasinya, dan
sifat sampel yang akan dipisahkan. Metode UF merupakan salah satu metode filtrasi
membran yang paling popular, yang memisahkan atau memekatkan komponen
target yang terdapat dalam suatu larutan melalui efek saringan molekular.
Efek saringan molekular terjadi karena perbedaan ukuran antara pori membran
dengan komponen pada larutan. Membran UF dapat diidentifikasi berdasarkan
ukuran nominal porinya (khususnya di antara 0,01 - 0,05 µm) atau oleh nilai
MWCO (Molecular Weight Cut Off) yang umumnya antara 2-500 kDa (Pirnie,

34
2005). Tabel 2.4. memberikan petunjuk untuk membedakan satu sama lain antara
beberapa metode filtrasi membran aliran tangensial berdasarkan ukuran pori
membran, MWCO dari senyawanya, campuran dan permeasinya.
Tabel 2.4.
Karakteristik beberapa metode filtrasi membran aliran tangensial (Pirnie, 2005).
Metoda Ukuran Pori
Membran(nm)
MWCO
(kDa)
Tekanan
(Pa)
Permeasi
Reverse
Osmosis (RO)
<0,6 <0,5 435-1015 Air.
Nanofiltrasi
(NF)
0,6 – 5 0,3 – 2 145 -580 Air, Zat terlarut
dengan BM
rendah.
Ultrafiltrasi
(UF)
5-50 2-500 7-145 Zat terlarut
dengan BM lebih
tinggi dan
makromolekul.
Mikrofiltrasi
(MF)
50-5000 >500 7-29 BM lebih tinggi
dan koloid.
Dalam filtrasi membran, larutan yang akan difiltrasi (feed) diberikan kondisi
agar kontak dengan suatu membran. Tekanan trans membran (TMP) diberikan ke
dalam sistem untuk mendorong larutan melewati membran. Suatu bagian larutan
tertentu akan berdifusi melalui membran (permeate) sedangkan beberapa bagian
akan tertahan atau ditolak oleh membran (retentate atau concentrate).
Ultrafiltrasi merupakan metode yang paling umum diterapkan sebagai tahap
awal purifikasi. Mengingat peptida penghambat ACE pada umumnya merupakan
peptida pendek, maka kualifikasi membran MWCO 5 kDa, 3 kDa, dan 1 kDa
paling sering digunakan pada berbagai studi. Ultrafiltrasi merupakan metode yang
sederhana, cepat dan reliable untuk pengkayaan peptida bioaktif, dan telah diakui
sebagai teknik yang sesuai untuk aplikasi skala industri (Zhu et al., 2010).

35
2.9. Tinjauan Tentang Metode LC-MS
Dewasa ini metode LC-MS (Liquid Chromatography - Mass Spectrofotometry)
semakin meningkat penerapannya dalam bidang yang lebih luas seperti, bidang
farmasi, lingkungan, pangan dan material industri. Pada umumnya, kromatografi
cair memisahkan komponen dalam sampel berdasarkan perbedaan afinitasnya (atau
kekuatan retensi) terhadap fase diam atau fase gerak, selanjutnya mendeteksi
komponen-komponen yang terpisah menggunakan UV, fluoresen, atau
konduktivitas listrik tergantung pada sifat-sifatnya.
Beberapa detektor terutama mengidentifikasi senyawa berdasarkan waktu
retensi dan mengkuantitasi senyawa berdasarkan intensitas dan luas puncak.
Kromatografi menawarkan resolusi yang baik, tetapi untuk mengkualifikasi dan
mengkuantitasi senyawa akan menjadi sulit jika banyak komponen terelusi pada
waktu yang hampir bersamaan, misalnya selama analisis banyak analit secara
simultan. Sebaliknya, mass spectrometry (MS) menawarkan teknik deteksi yang
sangat sensitif yang mengionisasi komponen sampel menggunakan berbagai,
selanjutnya memisahkan ion-ion yang dihasilkan dalam kondisi vakum berdasarkan
rasio massa terhadap muatan (m/z) dan mengukur intensitas dari masing-masing
ion. Mengingat spektra masssa yang disediakan oleh MS dapat menunjukkan
tingkat konsentrasi ion dari senyawa dengan massa tertentu, maka hal ini akan
sangat membantu dalam analisis kualitatif. Hal ini mengingat masa merupakan
informasi spesifik pada molekul tertentu dan MS mampu menyediakan informasi
tersebut secara langsung. Akan tetapi hal ini hanya bisa diterapkan jika kita
mengukur satu komponen saja. Jika banyak komponen (multikomponen)

36
diinjeksikan secara simultan, maka akan menjadi sangat sulit untuk menganalisa
spektranya.
Dengan demikian, maka sistem LC-MS menggabungkan metode kromatografi
cair (LC) yang dikenal unggul dari segi resolusi pemisahannya dengan metode MS
yang dikenal kapabilitasnya dalam analisis kualitatif. Spektra massa yang diperoleh
melalui pengukuran ini akan memberikan informasi tentang berat molekul dan
informasi struktur komponen-komponen yang dielusi, yang melengkapi informasi
kualitatif berdasarkan waktu retensi yang diperoleh menggunakan detector LC
lainnya.
Selain itu, pengukuran SIM (selected ion monitoring) mendeteksi senyawa
berdasarkan pada massa, yang merupakan parameter yang sangat selektif. Hal ini
memungkinkan pada analisis kuantitatif yang menghindari pengaruh senyawa lain
(kontaminan) bahkan hal ini tidak cukup dilakukan oleh pemisahan oleh LC saja.
Dalam hal penyediaan keluasan aplikasi pada berbagai jenis senyawa dan tingginya
selektivitas, spektrometer massa menawarkan karakteristik yang memuaskan
sebagai sebuah detektor LC.
Spektrometer massa telah digunakan sejak lama sebagai detektor pada sistem
kromatografi gas (GC) dalam metode GCMS, yang telah dikenal keunggulannya.
GCMS menyedikan makna yang sangat efektif untuk memisahkan dan identifikasi
senyawa, namun aplikasinya terbatas pada senyawa yang volatil dengan berat
molekul rendah, dan sampelnya juga harus memiliki stabilitas termal yang tinggi.
Sebaliknya, jika senyawa –senyawa tersebut terlarut pada fase gerak yang berupa
cairan (non volatil), maka LC mampu menganalisis bahkan senyawa-senyawa yang

37
sangat tidak volatile maupun senyawa yang tidak stabil yang sulit dianalisa
menggunakan GCMS. Dengan kata lain, salah satu keunggulan LC-MS adalah
memiliki aplikasi yang luas.
Sistem spektrometer massa terdiri atas suatu perangkat untuk memasukkan
sampel (seperti unit HPLC atau GC), antarmuka untuk menghubungkan perangkat
tersebut, sumber ion yang mengionisasi sampel, lensa elektrostatik yang secara
efisien mengenali ion yang dihasilkan, unit analisa massa yang memisahkan ion
berdasarkan rasio massa-muatan (m/z), dan unit detektor yang mendeteksi ion-ion
yang terpisah. Terdapat banyak jenis sistem MS yang tersedia tergantung pada
metode yang digunakan untuk memisahkan ion. Contoh berikut menunjukkan
komponen dari sistem MS quadrupole ionisasi atmosfer khas, yang umum
digunakan sebagai detektor LCMS (Gambar 2.7.). Unit ionisasi atmosfer
didasarkan pada ionisasi elektrospray (ESI = Electrospray Ionization), ionisasi
kimia atmosfer (APCI = Atmospheric Chemical Ionization), atau metode ionisasi
lainnya, dan berfungsi sebagai sumber ion dan antarmuka dengan sistem HPLC.
Ion yang dihasilkan dalam unit ini dilepaskan dari pelarut, kemudian difokuskan ke
balok menggunakan octupole atau cara lain, kemudian dikirim ke quadrupole. Arus
langsung dan arus bolak-balik frekuensi tinggi diterapkan ke quadrupole, sehingga
hanya ion dengan target rasio m/z tertentu yang berhasil melewati quadrupole.
Jumlah ion yang mencapai detektor diubah menjadi sinyal dan output ke computer.

38
Gambar 2.7. Komponen peralatan penyusun sistem LC-MS
(https://www.shimadzu.com/an/lcms/support/intro/lib/lctalk/46/46intro.html)




















