







Bahasa
Halaman
Hukum


Physiological and Molecular Plant Pathology (1996) 48, 131–144
Variation in sensitivity to tomatine and rishitin among
isolates of Fusarium oxysporum f.sp. lycopersici, and
strains not pathogenic on tomato
P. S", A. M. T#, A. A. S#, M. A. M#
and D C. S"*
"Department of Plant Biology, and the Maryland Agriculture Experiment Station, Uni�ersity of Maryland, CollegePark, MD 20742, U.S.A. and #Agricultural Genetic Engineering Research Institute, Agricultural Research Center,9 Gamaa st, Giza, Egypt
(Accepted for publication September 1995)
Studies of several fungal plant pathogens have found an association between greater virulence andincreased tolerance of the host’s defense compounds among different isolates of that pathogenspecies. This study examined 17 Fusarium oxysporum isolates to determine if tolerance of either oftwo fungitoxic compounds produced by tomato, rishitin and tomatine, would correlate withvirulence or pathogenicity on tomato. Among the 12 pathogenic isolates (forma specialislycopersici), quantitative levels of virulence were significantly correlated with rishitin tolerance and,in more limited circumstances, with tomatine tolerance. A group of four highly virulent isolatesdisplayed a relatively high tolerance of both tomatine and rishitin compared to the other isolates.When these pathogenic isolates were compared to five F. oxysporum isolates nonpathogenic ontomato, the nonpathogens generally displayed the highest sensitivity to tomatine, but not torishitin. Although these results do not prove a role for rishitin or tomatine tolerance in virulenceor pathogenicity, they do indicate that sufficient natural variation in these traits exists for themto contribute to an isolate’s disease potential on tomato. # 1996 Academic Press Limited
INTRODUCTION
Fusarium oxysporum (Sacc.) Snyder & Hans. causes wilt disease in a number of crops.
Individual strains normally show a high degree of host specificity and are grouped into
formae speciales according to the ability to cause disease on certain hosts.
Tomato (Lycopersicon esculentum Mill.) is a host to two formae speciales, F. oxysporum f.sp.
lycopersici, which causes a vascular wilt, and F. oxysporum f.sp. radicis-lycopersici, which
causes a crown rot [11]. F. oxysporum isolates which are not pathogenic to tomato have
also been isolated from tomato roots [8]. It is not known what traits are unique to the
pathogenic F. oxysporum which allow infection of tomato, or which traits produce more
virulent pathogens within a forma specialis.
One trait that may potentially determine either pathogenicity or virulence on a
specific host is the ability of the fungus to tolerate fungistatic or toxic compounds
produced by the host, such as the constitutively produced phytoanticipins [21] or the
*To whom correspondence should be addressed.Abbreviations used in text : CR, Castle Rock II-PVP; IPa-VF, Improved Pearson-VF; PDA, potato
dextrose agar ; PDB, potato dextose broth; PI, Pearson A-1 Improved; SM, Super Marmand.
0885–5765}96}02013114 $12.00}0 # 1996 Academic Press Limited

132 P. Suleman et al.
induced phytoalexins [22]. Such tolerance may result from either degradation of the
compound or nondegradative mechanisms [22]. Studies of other pathogen-plant
systems have identified variation in host phytoalexin tolerance among different isolates
of a pathogen. In these studies, higher tolerance to the host phytoalexin or
phytoanticipin was correlated with higher virulence on that host [4, 6, 14, 16, 18, 20].
Two fungitoxic compounds produced in tomato are tomatine, a saponin, and rishitin,
a terpenoid. Previous studies have shown that levels of tomatine and rishitin produced
in tomato stems and roots approach or exceed inhibitory levels (ED&!
) for F. oxysporum
f.sp. lycopersici [15]. This suggests that these compounds have the potential to limit
growth of this fungus within the plant and so allow faster growth of isolates possessing
greater tolerance of these compounds.
Tolerance of rishitin or tomatine has been found to correlate with increased
virulence in two other plant-pathogen systems. Rishitin tolerance has been associated
with high virulence of Gibberella pulicaris (Fries) Sacc. (anamorph: Fusarium sambucinum
Fuckel) on potato among both field isolates and progeny of a sexual cross [6].
Tomatine tolerance has been associated with the ability of Fusarium solani mutants to
cause rot on green tomato fruits [5]. F. oxysporum f.sp. lycopersici has been shown to
enzymatically detoxify tomatine [9], and, like a number of other tomato pathogens, is
more tolerant of tomatine than other fungi which are not pathogenic on tomato [1].
Thus, by analogy to the other pathogen systems, a variation in virulence among
different isolates of F. oxysporum f.sp. lycopersici could possibly result from differences
among the isolates in their tolerance of either rishitin or tomatine. Differences in host
specificity among F. oxysporum, in particular the ability to be pathogenic on tomato,
could also arise in part from an isolate’s tolerance of these tomato compounds. Since
the relative tolerance of different F. oxysporum isolates, pathogenic or nonpathogenic,
have not been directly compared, evidence for or against such relationships is lacking.
This study evaluates the variation of rishitin and tomatine tolerance in 17 isolates of
F. oxysporum in order to determine if differences correlate with pathogenicity or virulence
on race 1-susceptible tomato. Recently collected isolates were the focus of the study in
order to be certain that the virulence of these strains was not altered by long-term
storage and subculture. Additional F. oxysporum isolates which were collected from
tomato or associated soils and found not to be pathogenic on tomato were also
evaluated. Tolerance of rishitin and tomatine was tested for three states of growth
(spore germination, germ tube elongation, and growth of mature mycelia) so as to
detect differential expression of tolerance if it were developmentally regulated. Wilt
pathogens would be expected to pass through all three stages during spread in the
xylem. Understanding the variation intolerance of host chemicals among different
isolates of this asexually-reproducing pathogen could explain the underlying
determinants of virulence.
MATERIALS AND METHODS
Plant material
The tomato cultivars used in the studies included Pearson A-1 Improved (PI),
Improved Pearson VF (IPa-VF), (Peto Seed Co., Woodland, CA, U.S.A.) ; Super
Marmand (SM) and Castle Rock II-PVP (CR), (Sun Seed Co., Hollster, CA, U.S.A.).
The cultivars SM, and PI are susceptible to races 1, 2 and 3. The cultivar IPa-vf is

Virulence and chemical tolerance of Fusarium oxysporum 133
resistant to race 1 and susceptible to races 2 and 3. The cultivar CR is resistant to races
1 and 2 but susceptible to race 3. All plants were grown in sterilized sand}top soil mix
(1:1) in the greenhouse. Soil temperature was maintained with bottom heat set to
27³3 °C.
Isolates of Fusarium oxysporum
Isolates from Egypt and Maryland were collected from either suspected infected plants
or from soil in fields with a history of continuous tomato cultivation. Isolates were
obtained from surface-disinfested stem sections and plated on Komada’s Fusarium-
selective medium [12] or from soil by dilution-plating soil suspensions on Komada’s
medium and selected from the plates by characteristic growth and morphology after
7–10 days. Single-spore cultures of each isolate were sub-cultured on potato dextrose
Agar (PDA) slants for further use. All isolates described here were identified as F.
oxysporum from morphological features [3]. Reference strains of F. oxysporum f.sp.
lycopersici were obtained from the American Type Culture Collection: race 1 g16417
(abbreviated to ATCC 417) and race 2 g16605 (ATCC 605). Fusarium oxysporum f.sp.
pisi, strain 247, was kindly provided by Dr Hans D. VanEtten of the University of
Arizona.
Plant inoculation
Microconidia of all isolates were harvested from seven-day old PDA cultures grown at
26 °C. Aqueous suspensions of microconidia were filtered through two layers of
miracloth to remove mycelium before dilution to a final concentration of
1¬10' microconidia ml−" with additional sterile water. Five-leaf stage seedlings were
used. Plants were uprooted from flats and the root system gently washed to remove soil
particles. Three millimetres of tissue was excised from the tip of the tap root and the
root system was dipped into the spore suspension for 30 s and immediately transplanted.
In order to determine pathogenicity, four plants of cultivar SM were tested for each
isolate with two plants per 4 inch pot. Isolates were considered nonpathogenic if no wilt
symptoms occurred after 45 days in all four plants inoculated with that isolate.
Characterization by race and �irulence
Tomato cultivars susceptible and resistant to race 1 or to race 2 of F. oxysporum f.sp.
lycopersici (SM, PI, IPa-VF, CR) were used to characterize isolates by race and
virulence. Plants at the five-leaf stage were inoculated as described above. Plants were
evaluated at 45 days after inoculation by a disease index rating of 0–10, a modification
of Wellman’s rating [23] of 1–15. Evaluations were based on number of plants infected,
number of internodes from the bottom of the plant at which leaves display symptoms
and the extent of vascular discoloration (to the nearest internode from the bottom of
the plant). Inoculated plants which displayed neither wilt symptoms nor vascular
discoloration were considered to be uninfected and excluded from the disease rating so
as to evaluate virulence independent from the efficiency of establishing infection.
Generally, a plant receiving a score of 7–8 was defoliated in all but the topmost 3–4
leaves, which showed some chlorosis. Plants with scores of 4–5 generally displayed little
defoliation but half the leaves were chlorotic. Plants with scores of 2–3 displayed
chlorosis on only the lowest two to three leaves and displayed vascular discoloration
between the roots and the soil line.
Plants were also evaluated by culturing 5 mm pieces of surface disinfested stem tissue

134 P. Suleman et al.
at each internode to determine the extent of vertical spread of the pathogen. Plants at
45 days post-inoculation generally had 10–11 internodes. One experiment tested 36
plants of each cultivar for each isolate while a replicate experiment tested 10 plants of
each cultivar for each isolate. Results of both experiments were combined. Data on
disease index and stem colonization were analyzed separately by analysis of variance
(ANOVA) and mean separation by Duncan’s multiple range test (DMRT) at the level
of P¯ 0±05.
Preparation of rishitin and tomatine
Rishitin was elicited in slices of potato with arachidonic acid and prepared by a
modified protocol described by Desjardins and Gardner [6]. Partial purification of
rishitin was achieved by column chromatography of the residue obtained from ethyl
ether extract of the treated potato slices. Rishitin was eluted between 600–900 ml and
the fractions were reduced by vacuum evaporation to about 1±0 ml. The residue was
resuspended in hexane:ethyl acetate (7:3) and the rishitin was detected by TLC (silica
gel 60A, 20¬20 cm) using 20 µl aliquots of fractions and plates developed in
hexane:acetone (4:1). Rishitin was visualized, after spraying the plates with 0±5%
(w}v) vanillin in sulphuric acid, as red spots which turn blue within minutes.
Preparative plates were sprayed only on the edges and the adjacent unsprayed material
collected by scraping the silica from the plate. Rishitin was eluted from the silica with
hexane:ethyl acetate (7:3) and the TLC was repeated. Chemical purity, as determined
by gas chromatography and mass spectrum analysis indicated a minimum of 95±6%
rishitin. Mass spectroscopy was provided by the University of Maryland spectroscopy
facility. Tomatine was obtained from Sigma Chemical Co. (St. Louis, MO, U.S.A.).
Phytoalexin tolerance
Isolates were tested in potato dextrose broth (PDB) media amended with rishitin or
tomatine to determine the sensitivity of spore germination and germ tube elongation.
Both compounds were used at concentrations of 25, 50, and 100 µg ml−" in media.
Tomatine was dissolved in 0±5% acetic acid and then citrate-phosphate buffer pH 6
was added. Rishitin was dissolved in ethanol. Controls were amended with the volume
of solvent alone equivalent to a 50 µg ml−" treatment. The PDB medium had a pH of
5±0³0±1 with or without amendments and is close to that of tomato xylem fluid (5±3to 6±4, [15]). Approximately 400 microconidia ml−" were pipetted into each of three
replicate tubes of 2±0 ml of amended PDB. Spore germination counts were taken every
3 h for 18 h, while germ tube growth measurements were taken 12 and 24 h after
addition of spores.
Hyphal growth was evaluated from radial growth rates on PDA amended with
tomatine or rishitin, as described above. For each isolate, a 2 mm plug taken from
5-day-old cultures was placed upside down on PDA medium. All treatments were
maintained at 26}21 °C³1 (light and dark temp. in a growth chamber). Hyphal
growth measurements were taken daily for 6 days and the growth rate of each isolate
determined by regression of three replicates together. This rate was compared to the
radial growth rate on unamended PDA (three replicates) to express the percentage
reduction in growth rate. Data for spore germination, germ tube length and radial
growth were analyzed individually by ANOVA and by a means separation by
Virulence and chemical tolerance of Fusarium oxysporum 135
Duncan’s multiple range test (DMRT), at P¯ 0±05, using MSTATC (Michigan State
University). Correlative analyses were performed with MSTATC between data and
using replicate virulence measurements.
RESULTS
Characterization of pathogenicity and race
A total of 25 isolates of F. oxysporum were collected in Egypt and 41 isolates were
collected from Maryland. Ten of these isolates were able to cause wilt symptoms on
tomato plants susceptible to all three races of F. oxysporum f.sp. lycopersici and so were
identified as pathogens of this forma specialis. The race of each isolate, as determined
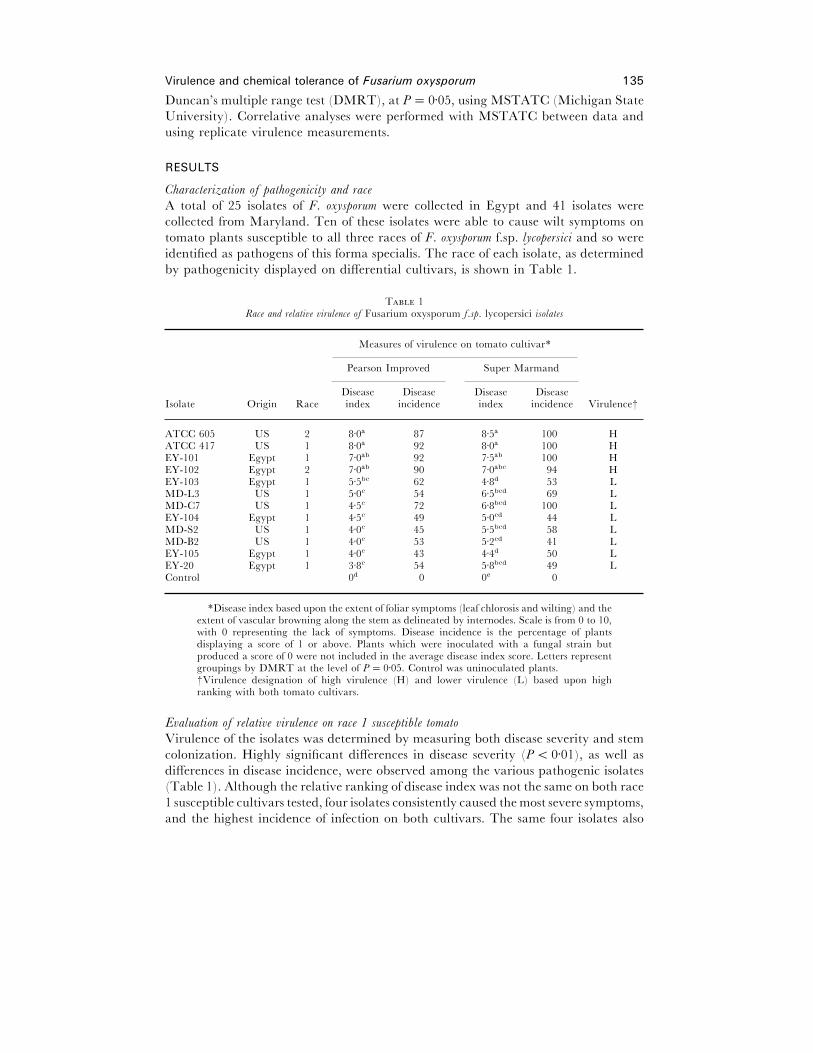
by pathogenicity displayed on differential cultivars, is shown in Table 1.
T 1Race and relati�e �irulence of Fusarium oxysporum f.sp. lycopersici isolates
Measures of virulence on tomato cultivar*
Pearson Improved Super Marmand
Isolate Origin RaceDiseaseindex
Diseaseincidence
Diseaseindex
Diseaseincidence Virulence†
ATCC 605 US 2 8±0a 87 8±5a 100 HATCC 417 US 1 8±0a 92 8±0a 100 HEY-101 Egypt 1 7±0ab 92 7±5ab 100 HEY-102 Egypt 2 7±0ab 90 7±0abc 94 HEY-103 Egypt 1 5±5bc 62 4±8d 53 LMD-L3 US 1 5±0c 54 6±5bcd 69 LMD-C7 US 1 4±5c 72 6±8bcd 100 LEY-104 Egypt 1 4±5c 49 5±0cd 44 LMD-S2 US 1 4±0c 45 5±5bcd 58 LMD-B2 US 1 4±0c 53 5±2cd 41 LEY-105 Egypt 1 4±0c 43 4±4d 50 LEY-20 Egypt 1 3±8c 54 5±8bcd 49 LControl 0d 0 0e 0
*Disease index based upon the extent of foliar symptoms (leaf chlorosis and wilting) and theextent of vascular browning along the stem as delineated by internodes. Scale is from 0 to 10,with 0 representing the lack of symptoms. Disease incidence is the percentage of plantsdisplaying a score of 1 or above. Plants which were inoculated with a fungal strain butproduced a score of 0 were not included in the average disease index score. Letters representgroupings by DMRT at the level of P¯ 0±05. Control was uninoculated plants.†Virulence designation of high virulence (H) and lower virulence (L) based upon highranking with both tomato cultivars.
E�aluation of relati�e �irulence on race 1 susceptible tomato
Virulence of the isolates was determined by measuring both disease severity and stem
colonization. Highly significant differences in disease severity (P! 0±01), as well as
differences in disease incidence, were observed among the various pathogenic isolates
(Table 1). Although the relative ranking of disease index was not the same on both race
1 susceptible cultivars tested, four isolates consistently caused the most severe symptoms,
and the highest incidence of infection on both cultivars. The same four isolates also
136 P. Suleman et al.
displayed the highest levels of colonization of tomato stem on SM (not shown). Thus,
for comparative purposes, we designated a group of four highly virulent F. oxysporum
f.sp. lycopersici isolates, and a group of eight lower virulence isolates (Table 1), although
there was continuous variation among isolates in the scores.
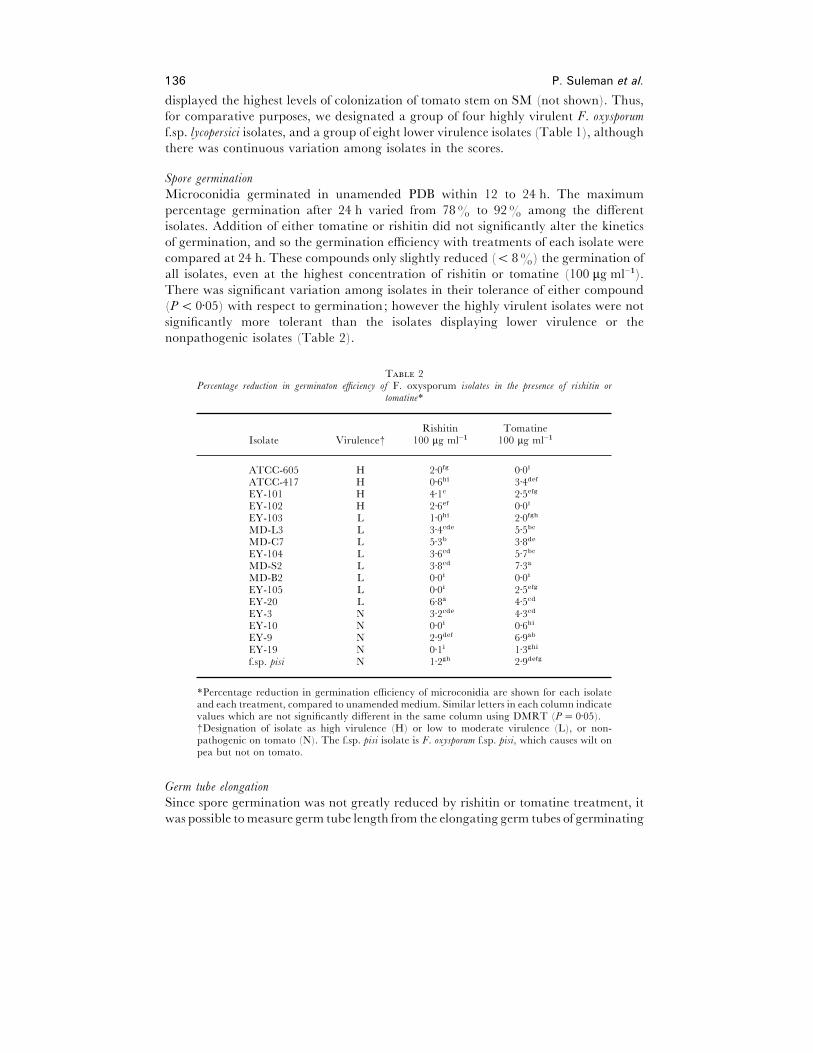
Spore germination
Microconidia germinated in unamended PDB within 12 to 24 h. The maximum
percentage germination after 24 h varied from 78% to 92% among the different
isolates. Addition of either tomatine or rishitin did not significantly alter the kinetics
of germination, and so the germination efficiency with treatments of each isolate were
compared at 24 h. These compounds only slightly reduced (! 8%) the germination of
all isolates, even at the highest concentration of rishitin or tomatine (100 µg ml−").
There was significant variation among isolates in their tolerance of either compound
(P! 0±05) with respect to germination; however the highly virulent isolates were not
significantly more tolerant than the isolates displaying lower virulence or the
nonpathogenic isolates (Table 2).
T 2Percentage reduction in germinaton efficiency of F. oxysporum isolates in the presence of rishitin or
tomatine*
Rishitin TomatineIsolate Virulence† 100 µg ml−" 100 µg ml−"
ATCC-605 H 2±0fg 0±0i
ATCC-417 H 0±6hi 3±4def
EY-101 H 4±1c 2±5efg
EY-102 H 2±6ef 0±0i
EY-103 L 1±0hi 2±0fgh
MD-L3 L 3±4cde 5±5bc
MD-C7 L 5±3b 3±8de
EY-104 L 3±6cd 5±7bc
MD-S2 L 3±8cd 7±3a
MD-B2 L 0±0i 0±0i
EY-105 L 0±0i 2±5efg
EY-20 L 6±8a 4±5cd
EY-3 N 3±2cde 4±3cd
EY-10 N 0±0i 0±6hi
EY-9 N 2±9def 6±9ab
EY-19 N 0±1i 1±3ghi
f.sp. pisi N 1±2gh 2±9defg
*Percentage reduction in germination efficiency of microconidia are shown for each isolateand each treatment, compared to unamended medium. Similar letters in each column indicatevalues which are not significantly different in the same column using DMRT (P¯ 0±05).†Designation of isolate as high virulence (H) or low to moderate virulence (L), or non-pathogenic on tomato (N). The f.sp. pisi isolate is F. oxysporum f.sp. pisi, which causes wilt onpea but not on tomato.
Germ tube elongation
Since spore germination was not greatly reduced by rishitin or tomatine treatment, it
was possible to measure germ tube length from the elongating germ tubes of germinating
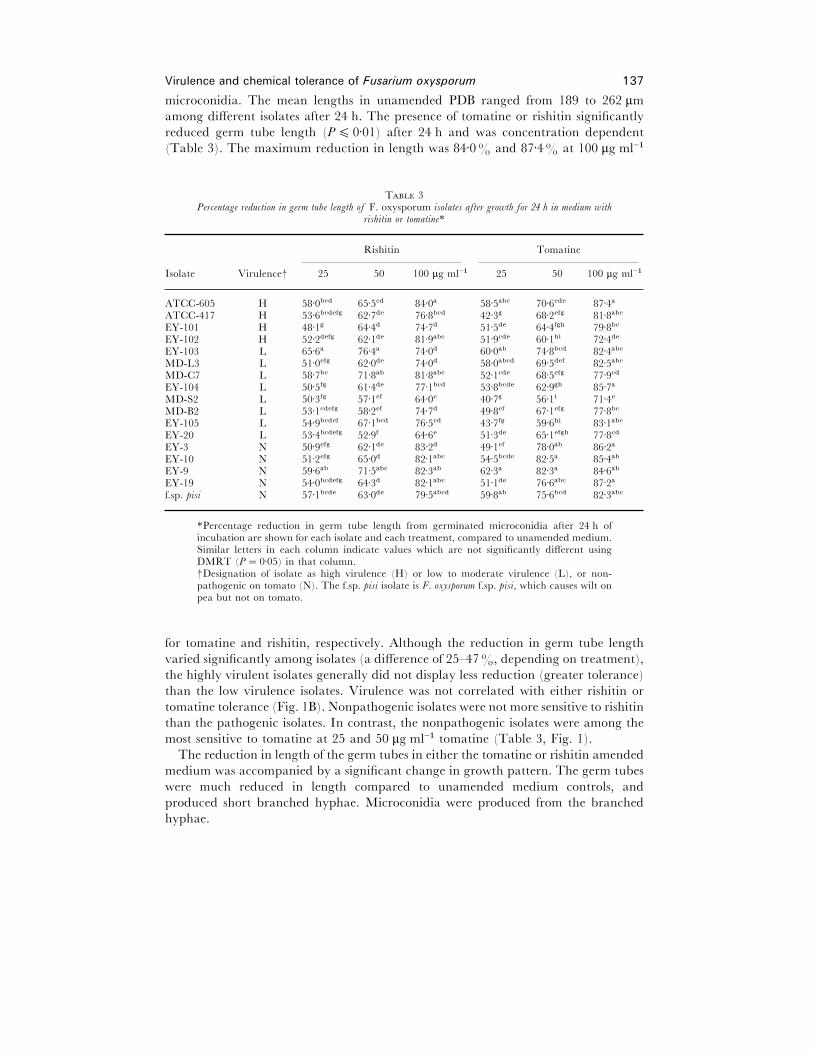
Virulence and chemical tolerance of Fusarium oxysporum 137
microconidia. The mean lengths in unamended PDB ranged from 189 to 262 µm
among different isolates after 24 h. The presence of tomatine or rishitin significantly
reduced germ tube length (P% 0±01) after 24 h and was concentration dependent
(Table 3). The maximum reduction in length was 84±0% and 87±4% at 100 µg ml−"
T 3Percentage reduction in germ tube length of F. oxysporum isolates after growth for 24 h in medium with
rishitin or tomatine*
Rishitin Tomatine
Isolate Virulence† 25 50 100 µg ml−" 25 50 100 µg ml−"
ATCC-605 H 58±0bcd 65±5cd 84±0a 58±5abc 70±6cde 87±4a
ATCC-417 H 53±6bcdefg 62±7de 76±8bcd 42±3g 68±2efg 81±8abc
EY-101 H 48±1g 64±4d 74±7d 51±5de 64±4fgh 79±8bc
EY-102 H 52±2defg 62±1de 81±9abc 51±9cde 60±1hi 72±4de
EY-103 L 65±6a 76±4a 74±0d 60±0ab 74±8bcd 82±4abc
MD-L3 L 51±0efg 62±0de 74±0d 58±0abcd 69±5def 82±5abc
MD-C7 L 58±7bc 71±8ab 81±8abc 52±1cde 68±5efg 77±9cd
EY-104 L 50±5fg 61±4de 77±1bcd 53±8bcde 62±9gh 85±7a
MD-S2 L 50±3fg 57±1ef 64±0e 40±7g 56±1i 71±4e
MD-B2 L 53±1cdefg 58±2ef 74±7d 49±8ef 67±1efg 77±8bc
EY-105 L 54±9bcdef 67±1bcd 76±5cd 43±7fg 59±6hi 83±1abc
EY-20 L 53±4bcdefg 52±9f 64±6e 51±3de 65±1efgh 77±8cd
EY-3 N 50±9efg 62±1de 83±2d 49±1ef 78±0ab 86±2a
EY-10 N 51±2efg 65±0d 82±1abc 54±5bcde 82±5a 85±4ab
EY-9 N 59±6ab 71±5abc 82±3ab 62±3a 82±3a 84±6ab
EY-19 N 54±0bcdefg 64±3d 82±1abc 51±1de 76±6abc 87±2a
f.sp. pisi N 57±1bcde 63±0de 79±5abcd 59±8ab 75±6bcd 82±3abc
*Percentage reduction in germ tube length from germinated microconidia after 24 h ofincubation are shown for each isolate and each treatment, compared to unamended medium.Similar letters in each column indicate values which are not significantly different usingDMRT (P¯ 0±05) in that column.†Designation of isolate as high virulence (H) or low to moderate virulence (L), or non-pathogenic on tomato (N). The f.sp. pisi isolate is F. oxysporum f.sp. pisi, which causes wilt onpea but not on tomato.
for tomatine and rishitin, respectively. Although the reduction in germ tube length
varied significantly among isolates (a difference of 25–47%, depending on treatment),
the highly virulent isolates generally did not display less reduction (greater tolerance)
than the low virulence isolates. Virulence was not correlated with either rishitin or
tomatine tolerance (Fig. 1B). Nonpathogenic isolates were not more sensitive to rishitin
than the pathogenic isolates. In contrast, the nonpathogenic isolates were among the
most sensitive to tomatine at 25 and 50 µg ml−" tomatine (Table 3, Fig. 1).
The reduction in length of the germ tubes in either the tomatine or rishitin amended
medium was accompanied by a significant change in growth pattern. The germ tubes
were much reduced in length compared to unamended medium controls, and
produced short branched hyphae. Microconidia were produced from the branched
hyphae.
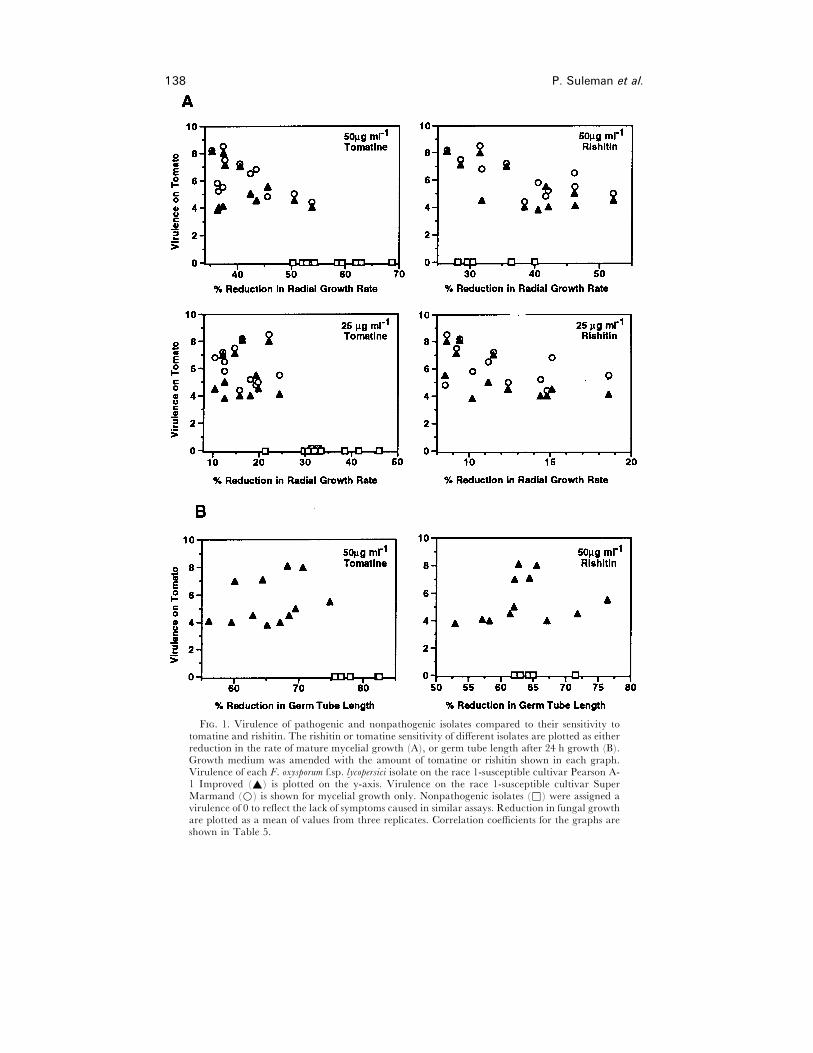
138 P. Suleman et al.
F. 1. Virulence of pathogenic and nonpathogenic isolates compared to their sensitivity totomatine and rishitin. The rishitin or tomatine sensitivity of different isolates are plotted as eitherreduction in the rate of mature mycelial growth (A), or germ tube length after 24 h growth (B).Growth medium was amended with the amount of tomatine or rishitin shown in each graph.Virulence of each F. oxysporum f.sp. lycopersici isolate on the race 1-susceptible cultivar Pearson A-1 Improved (_) is plotted on the y-axis. Virulence on the race 1-susceptible cultivar SuperMarmand (D) is shown for mycelial growth only. Nonpathogenic isolates (*) were assigned avirulence of 0 to reflect the lack of symptoms caused in similar assays. Reduction in fungal growthare plotted as a mean of values from three replicates. Correlation coefficients for the graphs areshown in Table 5.
Virulence and chemical tolerance of Fusarium oxysporum 139
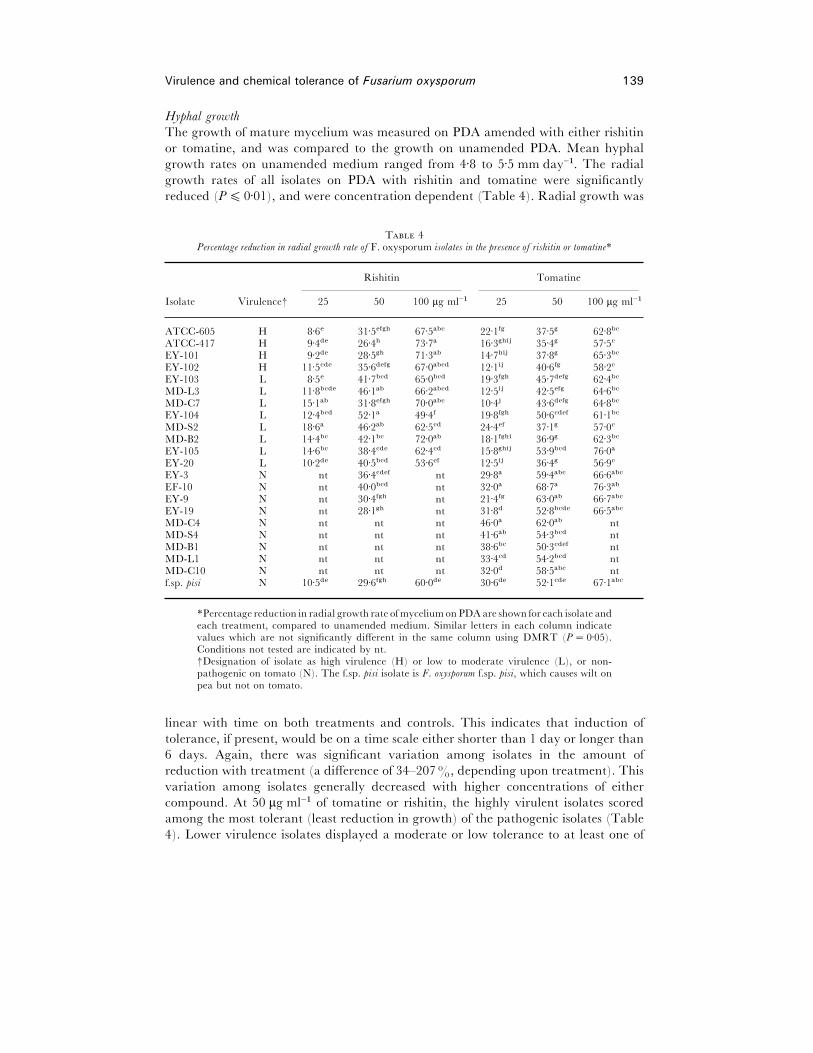
Hyphal growth
The growth of mature mycelium was measured on PDA amended with either rishitin
or tomatine, and was compared to the growth on unamended PDA. Mean hyphal
growth rates on unamended medium ranged from 4±8 to 5±5 mm day−". The radial
growth rates of all isolates on PDA with rishitin and tomatine were significantly
reduced (P% 0±01), and were concentration dependent (Table 4). Radial growth was
T 4Percentage reduction in radial growth rate of F. oxysporum isolates in the presence of rishitin or tomatine*
Rishitin Tomatine
Isolate Virulence† 25 50 100 µg ml−" 25 50 100 µg ml−"
ATCC-605 H 8±6e 31±5efgh 67±5abc 22±1fg 37±5g 62±8bc
ATCC-417 H 9±4de 26±4h 73±7a 16±3ghij 35±4g 57±5c
EY-101 H 9±2de 28±5gh 71±3ab 14±7hij 37±8g 65±3bc
EY-102 H 11±5cde 35±6defg 67±0abcd 12±1ij 40±6fg 58±2c
EY-103 L 8±5e 41±7bcd 65±0bcd 19±3fgh 45±7defg 62±4bc
MD-L3 L 11±8bcde 46±1ab 66±2abcd 12±5ij 42±5efg 64±6bc
MD-C7 L 15±1ab 31±8efgh 70±0abc 10±4j 43±6defg 64±8bc
EY-104 L 12±4bcd 52±1a 49±4f 19±8fgh 50±6cdef 61±1bc
MD-S2 L 18±6a 46±2ab 62±5cd 24±4ef 37±1g 57±0c
MD-B2 L 14±4bc 42±1bc 72±0ab 18±1fghi 36±9g 62±3bc
EY-105 L 14±6bc 38±4cde 62±4cd 15±8ghij 53±9bcd 76±0a
EY-20 L 10±2de 40±5bcd 53±6ef 12±5ij 36±4g 56±9c
EY-3 N nt 36±4cdef nt 29±8a 59±4abc 66±6abc
EF-10 N nt 40±0bcd nt 32±0a 68±7a 76±3ab
EY-9 N nt 30±4fgh nt 21±4fg 63±0ab 66±7abc
EY-19 N nt 28±1gh nt 31±8d 52±8bcde 66±5abc
MD-C4 N nt nt nt 46±0a 62±0ab ntMD-S4 N nt nt nt 41±6ab 54±3bcd ntMD-B1 N nt nt nt 38±6bc 50±3cdef ntMD-L1 N nt nt nt 33±4cd 54±2bcd ntMD-C10 N nt nt nt 32±0d 58±5abc ntf.sp. pisi N 10±5de 29±6fgh 60±0de 30±6de 52±1cde 67±1abc
*Percentage reduction in radial growth rate of myceliumon PDA are shown for each isolate andeach treatment, compared to unamended medium. Similar letters in each column indicatevalues which are not significantly different in the same column using DMRT (P¯ 0±05).Conditions not tested are indicated by nt.†Designation of isolate as high virulence (H) or low to moderate virulence (L), or non-pathogenic on tomato (N). The f.sp. pisi isolate is F. oxysporum f.sp. pisi, which causes wilt onpea but not on tomato.
linear with time on both treatments and controls. This indicates that induction of
tolerance, if present, would be on a time scale either shorter than 1 day or longer than
6 days. Again, there was significant variation among isolates in the amount of
reduction with treatment (a difference of 34–207%, depending upon treatment). This
variation among isolates generally decreased with higher concentrations of either
compound. At 50 µg ml−" of tomatine or rishitin, the highly virulent isolates scored
among the most tolerant (least reduction in growth) of the pathogenic isolates (Table
4). Lower virulence isolates displayed a moderate or low tolerance to at least one of

140 P. Suleman et al.
these compounds. For example, at 50 µg ml−" of either compound, the lower virulence
isolate EY-104 was very sensitive to both tomatine and rishitin. Three other lower
virulence isolates, MD-S2, MD-B2 and EY-20, were as tolerant to tomatine as the high
virulence isolates, but displayed only low to moderate tolerance for rishitin. Conversely,
the lower virulence isolate EY-105 was relatively tolerant of rishitin but sensitive to
tomatine. Thus, at this concentration, there appeared to be an association of relatively
high tolerance to both compounds with only the group of highly virulent pathogens.
However, such a relationship was not strictly observed at lower (25 µg ml−") or higher
(100 µg ml−") concentrations of either compound.
In order to extend the above analysis, the correlation of tolerance for each
compound with quantitative virulence measurements (disease index) was tested.
Virulence of the different isolates was significantly (P% 0±05) correlated with their
tolerance of rishitin, at 25 or 50 µg ml−" rishitin for cultivar PI and at 50 µg ml−"
rishitin for cultivar SM (Table 5). Rishitin tolerance was also significantly correlated
T 5Correlation analyses between �irulence of Fusarium oxysporum f.sp. lycopersici isolates on two tomato
culti�ars and their tolerance of rishitin and tomatine
Conc.Rishitin Tomatine
Cultivar µg ml−" r* P† r P
Pearson Improved 25 ®0±666 ** ®0±032 NS50 ®0±701 ** ®0±378 NS
100 ®0±534 NS ®0±193 NSSuper Marmand 25 ®0±464 NS ®0±164 NS
50 ®0±701 ** ®0±610 *100 ®0±515 NS ®0±287 NS
*Coefficient of correlation.†Significance of correlation coefficient. * and ** indicate significance at P¯ 0±05 and 0±01respectively. NS indicates not significant at P¯ 0±05.
to the disease incidence under similar circumstances (not shown). There was no
significant correlation of rishitin tolerance at 100 µg ml−" rishitin to either virulence or
disease incidence, apparently due to the more uniform sensitivity of the isolates at this
concentration. Virulence on the cultivar PI was not significantly correlated to tomatine
tolerance at any concentration (Fig. 1A, Table 5). However, virulence on another
cultivar, SM, was significantly correlated with tomatine tolerance at the 50 µg ml−"
concentration (P! 0±05).
Comparison of pathogenic and nonpathogenic isolates indicated that all non-
pathogenic isolates were among the most sensitive to tomatine. In order to further test
this result, five additional nonpathogenic F. oxysporum strains were evaluated for
tomatine tolerance in radial growth assays (Table 4). At 25 and 50 µg ml−" tomatine,
growth of most nonpathogenic isolates was significantly more reduced (less tolerant)
than was any pathogenic strain (Table 4, Fig. 1A). For example, at 25 µg ml−"
tomatine, eight out of 10 nonpathogenic isolates were significantly more sensitive than

Virulence and chemical tolerance of Fusarium oxysporum 141
any pathogenic isolates (P! 0±05), displaying a 1±5 to 4-fold greater reduction in
growth rate in response to tomatine. In contrast, the nonpathogenic isolates showed no
such association with rishitin sensitivity. All nonpathogenic isolates were as tolerant to
rishitin as the pathogenic isolates, including, in some cases, those in the high virulence
group.
DISCUSSION
Variation in virulence among isolates of a pathogen can provide the basis for further
genetic and biochemical studies to identify virulence determinants. Variation in the
virulence of F. oxysporum f.sp. lycopersici has not been studied much beyond the original
observations by Wellman [23]. The 12 isolates of F. oxysporum f.sp. lycopersici used in the
present study varied significantly with respect to both their ability to incite disease and
the severity of symptoms that they caused on infected plants. Here, we focused upon
symptom severity (vertical extent of foliar symptoms and vascular discoloration) as a
measure of virulence. Although the variation in virulence among isolates was
continuous, with the relative ranking dependent upon the tomato cultivar used, one
group of four pathogenic isolates colonized tomato stems more aggressively than the
other isolates tested, causing more severe symptoms and higher disease incidence, on
both cultivars of race 1-susceptible tomato. This group of high virulence isolates
included examples of both race 1 and race 2 genotypes, indicating that the virulence
of these isolates is independent of their race. Relative ranking among the lower
virulence isolates was more dependent upon the cultivar of tomato used.
Evaluation of the growth of these F. oxysporum isolates in the presence of rishitin and
tomatine demonstrated levels of tolerance which are consistent with a previous report
using a single F. oxysporum f.sp. lycopersici isolate (e.g. ED&!
of 8 µg ml−" for germ tube
growth with either compound [15]), considering the pH dependence of toxicity with
these compounds and the different media used in each study [1]. Different stages of
growth of these F. oxysporum isolates displayed different levels of sensitivity to these
compounds and differences in correlation of sensitivity to virulence. Spore germination
was largely unaffected by either compound, even at 100 µg ml−", and so the variation
measured between isolates would presumably have little consequence in secondary
germination within the plant. Germ tube elongation was the most sensitive stage to
both compounds but the degree of sensitivity among isolates did not correlate with
virulence. Radial growth rate, however, displayed the widest variation among isolates
and was correlated with virulence and pathogenicity under certain conditions.
Evaluation of a possible association of tolerance with virulence focused upon the
pathogenic isolates. Mycelial growth in the presence of rishitin was strongly and
significantly correlated with virulence under several of the experimental conditions
evaluated: at 25 and 50 µg ml−" rishitin in the tolerance assays, and when virulence on
either tomato cultivar was considered. Since rishitin has been found to be present at
concentrations ranging from 8 to 44 µg g−" fresh wt. in vascular and stem tissue
respectively [7, 15], there may be circumstances where this could lead to selective
growth of more tolerant isolates within the host. The lack of correlation of virulence
with rishitin tolerance at 100 µg ml−" rishitin does not detract from this conclusion
since rishitin has not been reported to accumulate to such high concentrations in

142 P. Suleman et al.
tomato. Correlation of virulence with tomatine tolerance was only found to be
significant under limited experimental conditions : only when virulence on the SM
cultivar was considered, and only at one tomatine concentration (50 µg ml−") tested in
the tolerance assay. Although this concentration is similar to that found in tomato
(50 µg g−" fresh wt. in root and stem tissue [13], 100 µg g−" fresh wt. in stem vascular
tissue [15]), tomatine tolerance may affect F. oxysporum virulence in more narrow
circumstances. The cultivar-dependence of the correlations could be attributed to the
higher virulence of five moderately virulent isolates on SM than on PI. Four of these
isolates were found to be highly tolerant of tomatine, but relatively more sensitive to
rishitin. Since tomatine or rishitin levels have been reported to vary by 2 to 6-fold
among different tomato cultivars [7, 17], the relative contributions of tolerance of each
compound to virulence could depend upon the levels of each compound in the cultivar
tested.
Although the analyses show separate correlations of virulence with rishitin tolerance,
and to a lesser extent with tomatine tolerance, an isolate’s tolerance of both compounds
together may provide a more accurate prediction of virulence since both compounds
occur in challenged tissues. Indicative of this possibility, the highly virulent isolates
generally displayed high tolerance of both tomatine and rishitin at 50 µg ml−". In
contrast, the lower virulence isolates displayed a similar high level of tolerance of only
one, or neither, of these compounds. Thus the correlations of virulence with tolerance
of one compound alone may underestimate the significance of its potential contribution
to virulence. Desjardins and Gardner [6] reported a similar relationship in G. pulicaris,
where tolerance of both potato phytoalexins rishitin and lubimin were necessary, but
not sufficient, for high virulence.
Despite the apparent association between the level of rishitin tolerance with
virulence, this association is only correlative and does not prove a role in virulence.
Two concerns might minimize such a role. The first concern is that the tolerance
displayed during spore germination and germ tube elongation did not correlate with
virulence. Any effect of differential tolerance on virulence could therefore function only
during the stage of growth of mature mycelia. However, the lack of perforation plates
in tomato xylem vessel elements has been used to argue that continued sporulation and
germination may not be necessary for spread in tomato xylem; fungal growth in the
xylem is mostly in the form of non-sporulating hyphae [19]. Therefore, limitation of
hyphal growth could indeed be expected to affect virulence more than a limitation of
germination or germ tube elongation. A second concern is that the variation in rishitin
tolerance among strains of F. oxysporium f.sp. lycopersici is much narrower than that
displayed by G. pulicaris, a pathogen of potato, to the same compound (18–88%
reduction in hyphal growth rate on 200 µg ml−" rishitin [6]). This suggests that
differences in rishitin tolerance may have a more limited role in the variation in its
virulence than in G. pulicaris. Part of this difference between pathogens may, however,
be related to the generally higher concentrations of rishitin in potato tubers (90 µg g−"
fresh wt. or more [10]) than in tomato.
When the pathogenic and nonpathogenic F. oxysporum isolates were compared, the
nonpathogens appeared to possess greater sensitivity to tomatine during germ tube
elongation and growth of mature mycelia at 25 and 50 µg ml−". Similar concentrations
of tomatine are generally thought to exist in healthy tomato tissue (50 µg g−" fresh wt.

Virulence and chemical tolerance of Fusarium oxysporum 143
in root and stem tissue [13], 100 µg g−" fresh wt. in stem vascular tissue [14]), and thus
may provide some selection against growth of more tomatine-sensitive strains. A
smaller growth differential between pathogens and nonpathogens would be expected
in the xylem fluids where concentrations of tomatine are higher (100–400 µg ml−"
[17]), but growth of nonpathogens is generally halted prior to this stage of infection [2].
Similar tolerance studies with F. oxysporum f.sp. radicis-lycopersici, which causes crown
and root rot in tomato, also indicate a greater tolerance of tomatine among isolates
pathogenic on tomato than the nonpathogenic isolates tested here. Sixteen isolates of
this forma specialis displayed only a 6–43% reduction in hyphal growth rate at
100 µg ml−" tomatine (Suleman, Straney and McGovern, unpublished). In the light of
the differences in tomatine sensitivity between the two tomato pathogenic formae
speciales and the nonpathogens, we could speculate that a certain level of tolerance of
tomatine may be required, but not in itself be sufficient, for certain types of pathogenic
growth on tomato. Although this study can not rule out a similar role for rishitin,
similar extremes in rishitin sensitivity do not appear to occur frequently in
nonpathogenic F. oxysporum associated with tomato.
Collectively, these results indicate that differences in virulence among pathogenic
isolates correlate most strongly to quantitative differences in tolerance of rishitin and
only more narrowly to differences in tolerance of tomatine. However, a certain
threshold level of tomatine tolerance is associated with pathogenicity on tomato since
growth of nonpathogens was generally more sensitive to tomatine than was that of
pathogenic isolates. Such different effects with the two compounds are consistent with
the different timing of rishitin and tomatine production in tomato. The preformed
nature of tomatine would allow inhibition of the establishment of an infection, while
the induced nature of rishitin would affect fungal growth only later during infection.
Although such differences in tolerance can not fully account for all differences in
virulence or pathogenicity of the different isolates, there are general trends which are
consistent with such relationships contributing to disease potential. An analogous
situation was observed with G. pulicaris, where tolerance to two different phytoalexins
in potato were necessary, but not sufficient, for high virulence in potato [6]. In this
case, sexual genetic analysis clarified the role of rishitin tolerance in G. pulicaris
virulence [6]. Similar manipulation of F. oxysporum, through parasexual genetics or
molecular cloning will be necessary to prove such role for tomatine or rishitin tolerance
in F. oxysporum f.sp. lycopersici. This study has characterized the variation among a set
of field isolates, and suggests possible roles for tomatine and rishitin tolerance, for
further detailed analysis.
The authors wish to thank Ann Desjardins and Susan McCormick of the Mycotoxin
Research Unit, USDA}ARS, Peoria, IL for their assistance and kindly providing
rishitin standards. Peto Seed Co. kindly supplied tomato seed which is not distributed
in U.S. market. Research was supported by a collaborative grant from the National
Agricultural Research Program (NARP) and the U.S.AID}Cairo, and coordinated in
the U.S. by USDA}OICD. Further support was provided through a NARP fellowship
to A. A. Saleh and a Peace Fellowship from the Egyptian Embassy (U.S.) to A. M.
Tohamy for work at the University of Maryland. Scientific article number A7783 of
the Maryland Agricultural Experiment Station.

144 P. Suleman et al.
REFERENCES
1. Arneson PA, Durbin RD. 1968. The sensitivity of fungi to α-tomatine. Phytopathology 58 : 536–537.2. Beswetherick JT, Bishop CD. 1993. An ultrastructural study of tomato roots inoculated with
pathogenic and nonpathogenic necrotrophic fungi and a saprophytic fungus. Plant Pathology 42 :577–588.
3. Booth C. 1971. The genus Fusarium. Kew, England: Commonwealth Mycol. Inst.4. Bowyer P, Clarke BR, Lunness P, Daniels MJ, Osbourn AE. 1995. Host range of a plant
pathogenic fungus determined by a saponin detoxifying enzyme. Science 267 : 371–374.5. Defago G, Kern H. 1983. Induction of Fusarium solani mutants insensitive to tomatine, their
pathogenicity and aggressiveness to tomato fruits and pea plants. Physiological and Molecular PlantPathology 22 : 29–37.
6. Desjardins AE, Gardner HW. 1989. Genetic analysis in Gibberella pulicaris : Rishitin tolerance, rishitinmetabolism and virulence on potato tubers. Molecular Plant Microbe Interactions 2 : 26–34.
7. Elgersma DM, Liem JI. 1989. Accumulation of phytoalexins in susceptible and resistant near-isogeniclines of tomato infected with Verticillium albo-atrum or Fusarium oxysporum f.sp. lycopersici. Physiological andMolecular Plant Pathology 34 : 545–555.
8. Elias KS, Schneider RW, Lear MM. 1991. Analysis of vegetative compatibility groups innonpathogenic populations of Fusarium oxysporum isolated from symptomless tomato roots. CanadianJournal of Botany 69 : 2089–2094.
9. Ford, JE, McCance DJ, Drysdale RB. 1979. The hydrolysis of tomatine, by an inducible extracellularenzyme from Fusarium oxysporum f.sp. lycopersici. In: Hawkes JG, Lester RN, Skelding AD, eds. Linn.Soc. Symp. Ser. �ol. 7; The Biology and Taxonomy of the Solanaceae. London: Academic Press.
10. Henfling JWDM, Kuc’ J. 1979. A semi-micro method for the quantitation of sesquiterpene stressmetabolites in potato tuber. Phytopathology 69 : 609–612.
11. Jarvis WR. 1988. Fusarium crown and root rot of tomatoes. Phytoprotection 69 : 49–69.12. Komada H. 1975. Development of a selective medium for quantitative isolation of Fusarium oxysporum
from natural soil. Re�iews of Plant Protection Research 8 : 114–125.13. Langcake P, Drysdale RB, Smith H. 1972. Post-infectional production of an inhibitor of Fusarium
oxysporum f.sp. lycopersici by tomato plants. Physiological Plant Pathology 2 : 17–25.14. Lucy MC, Matthews PS, VanEtten HD. 1988. Metabolic detoxification of the phytoalexins
maackiain and medicarpan by Nectria haematococca field isolates : relationship to virulence on chickpea.Physiological and Molecular Plant Pathology 33 : 187–199.
15. McChance DJ, Drysdale RB. 1975. Production of tomatine and rishitin in tomato plants inoculatedwith Fusarium oxysporum f.sp. lycopersici. Physiologial Plant Pathology 7 : 221–230.
16. Miao VPW, VanEtten HD. 1992. Genetic analysis of the role of phytoalexin detoxification in virulenceof fungus Nectria haematococca on chickpea (Cicer arietinum). Applied En�ironmental Microbiology 58 :809–814.
17. Smith CA, MacHardy WE. 1982. The significance of tomatine in the host response of susceptible andresistant tomato isolines infected with two races of Fusarium oxysporum f.sp. lycopersici Lycopersiconesculentum. Phytopathology 72 : 415–419.
18. Soledade M, Pedras C, Taylor J. 1993. Metabolism of the phytoalexin brassinin by the ‘‘Blackleg’’fungus. Journal of Natural Products 56 : 731–738.
19. Stromberg EL, Corden ME. 1980. Scanning electron microscopy of Fusarium oxysporum f.sp. lycopersiciin xylem vessels of wilt-resistant and susceptible tomato plants. Canadian Journal of Botany 58 :2360–2366.
20. Tegtmeier KJ, VanEtten HD. 1982. The role of pisatin tolerance and degradation in the virulence ofNectria haematococca on peas : A genetic analysis. Phytopathology 72 : 608–612.
21. VanEtten HD, Mansfield JW, Bailey JA, Farmer EE. 1994. Two classes of plant antibiotics :Phytoalexins versus ‘‘phytoanticipins ’’. Plant Cell 6 : 1191–1192.
22. VanEtten HD, Matthews DE, Matthews PS. 1989. Phytoalexin detoxification: Importance forpathogenicity and practical considerations. Annual Re�iew of Phytopathology 27 : 143–164.
23. Wellman FL. 1940. Differences in Growth Characters and Pathogenicity of Fusarium Wilt Isolations tested on ThreeTomato Varieties. Technical Bull. no. 705. Washington, DC; U.S. Department of Agriculture.
Top Related
Copyright © 2022 FDOKUMEN

