







Bahasa
Halaman
Hukum


Three-dimensional structure of a barley ββ-D-glucan exohydrolase,a family 3 glycosyl hydrolaseJoseph N Varghese1*, Maria Hrmova2 and Geoffrey B Fincher2*
Background: Cell walls of the starchy endosperm and young vegetative tissuesof barley (Hordeum vulgare) contain high levels of (1→3,1→4)-β-D-glucans.The (1→3,1→4)-β-D-glucans are hydrolysed during wall degradation ingerminated grain and during wall loosening in elongating coleoptiles. These keyprocesses of plant development are mediated by several polysaccharideendohydrolases and exohydrolases.
Results: The three-dimensional structure of barley β-D-glucan exohydrolaseisoenzyme ExoI has been determined by X-ray crystallography. This is the firstreported structure of a family 3 glycosyl hydrolase. The enzyme is a two-domain,globular protein of 605 amino acid residues and is N-glycosylated at three sites.The first 357 residues constitute an (α/β)8 TIM-barrel domain. The seconddomain consists of residues 374–559 arranged in a six-stranded β sandwich,which contains a β sheet of five parallel β strands and one antiparallel β strand,with three α helices on either side of the sheet. A glucose moiety is observed ina pocket at the interface of the two domains, where Asp285 and Glu491 arebelieved to be involved in catalysis.
Conclusions: The pocket at the interface of the two domains is probably theactive site of the enzyme. Because amino acid residues that line this active-sitepocket arise from both domains, activity could be regulated through the spatialdisposition of the domains. Furthermore, there are sites on the second domainthat may bind carbohydrate, as suggested by previously published kinetic dataindicating that, in addition to the catalytic site, the enzyme has a second bindingsite specific for (1→3,1→4)-β-D-glucans.
IntroductionTwo basic β-glucan exohydrolases with apparent molecu-lar masses of 69 kDa and 71 kDa have been purified fromextracts of eight day germinated barley seedlings and aredesignated isoenzymes ExoI and ExoII, respectively [1].β-Glucan exohydrolases have also been detected in elon-gating coleoptiles of barley [2,3] and maize [4], and insuspension-cultured soybean cells [5]. The barleyenzymes preferentially hydrolyse the (1→3)-β-glucan,laminarin, but also hydrolyse (1→3,1→4)-glucans,(1→3,1→6)-β-glucans and a range of glucooligosaccha-rides and aryl-β-glucosides [6]. Both isoforms releasesingle glucosyl residues from the non-reducing ends ofsubstrates, and proton nuclear magnetic resonance(NMR) shows that the anomeric configuration of releasedglucose is retained after hydrolysis [1]. Detailed kineticanalyses showed that resultant Hill plots had gradients of1.0 both for laminarin and for 4-nitrophenyl β-glucoside,but 2.0 for the cell wall (1→3,1→4)-β-glucan from barley[6]. The gradient of 2.0 indicated positive cooperativity ofbinding of (1→3,1→4)-β-glucan to the enzyme and led tothe conclusion that barley β-glucan exohydrolases haveanother binding site for cell wall (1→3,1→4)-β-glucans;
the second site does not appear to bind (1→3)-β-glucansor other, smaller, substrates [6].
Glycosyl hydrolases have been classified into a number offamilies on the basis of amino acid sequence alignmentsand hydrophobic cluster analyses [7]. It is becomingapparent that members of each family have similar three-dimensional (3D) conformations. For example, in previouswork from our laboratories, it was shown that barley(1→3)-β-glucan endohydrolases and (1→3,1→4)-β-glucanendohydrolases, both of which are classified in family 17,have (α/β)8 barrel folds that are very similar; the root meansquare deviation (rmsd) in Cα positions is 0.65 Å over 278amino acid residues [8]. Sequence alignments andhydrophobic-cluster analyses show that the barleyβ-glucan exohydrolases are members of the family 3 glyco-syl hydrolases [6,7], but no 3D structures are currentlyavailable for this group of enzymes. However, we recentlycrystallized β-glucan exohydrolase isoenzyme ExoI frombarley using vapour diffusion in the presence of ammo-nium sulphate and polyethylene glycol [9]. Crystal dimen-sions of up to 0.8 mm × 0.4 mm × 0.6 mm were observedand the crystals diffracted to at least 2.2 Å resolution [9].
Addresses: 1Biomolecular Research Institute, 343Royal Parade, Parkville, Victoria 3052, Australia and2Department of Plant Science, University ofAdelaide, Waite Campus, South Australia 5064,Australia.
*Corresponding authors.E-mail: [email protected]
Key words: barley β-D-glucan exohydrolaseisoenzyme ExoI, β-D-glucan, family 3 glycosylhydrolase, X-ray crystallography
Received: 11 September 1998Revisions requested: 13 October 1998Revisions received: 19 November 1998Accepted: 15 December 1998
Published: 29 January 1999
Structure February 1999, 7:179–190http://biomednet.com/elecref/0969212600700179
© Elsevier Science Ltd ISSN 0969-2126
Research Article 179

In the present work, the 3D structure of the barleyβ-glucan exohydrolase isoenzyme ExoI has been deter-mined by X-ray crystallography and this therefore repre-sents the first structure reported for a family 3 glycosylhydrolase and for a β-glucan glucohydrolase.
Results and discussionThe 3D structure of barley β-glucan exohydrolase isoen-zyme ExoI, a family 3 glycosyl hydrolase, has now beensolved at 2.2 Å resolution by X-ray crystallography, usingmultiple isomorphous replacement (MIR) techniques(Table 1). The heavy atom derived phases were accurateenough to build the entire molecule unambiguously andsimulated annealing was not necessary as a means of refin-ing the structure.
The complete amino acid sequence of the enzyme has beendeduced from the nucleotide sequence of a near full-lengthcDNA (AJ Harvey and GBF, unpublished results) and fromamino acid sequences of tryptic peptides generated from thepurified enzyme [1]. The mature enzyme contains 605amino acid residues, and its sequence has been lodged inthe GenBank and EMBL databases under accessionnumber AF102868. The X-ray data were collected from twocrystals [9] and electron density was continuous for theentire Cα polypeptide chain, except that the last three aminoacid residues at the C terminus could not be observed in theelectron-density map. The electron densities of most aminoacid sidechains were well-defined and identifiable residuescorresponded in all cases to those deduced from the cDNAsequence. In several instances, the electron density ofsidechains was used to confirm that the nucleotide sequencewas translated in the correct reading frame; the cDNA is par-ticularly GC-rich and sequencing compressions werecommon (AJ Harvey and GBF, unpublished results).
Two isoforms of the barley β-glucan exohydrolase familyhave been purified and characterized [1,2], and Southernhybridization analyses show that there are approximatelyeight members of the barley β-glucan exohydrolase genefamily (AJ Harvey and GBF, unpublished results). Isoen-zymes ExoI and ExoII exhibit 72% sequence identity at theamino acid level (AJ Harvey and GBF, unpublishedresults). We predict that the 3D structures of isoenzymeExoII and other barley isoforms (N Sakurai, personal com-munication), together with related β-glucan exohydrolasesfrom maize [4] and higher plants generally [5], will be verysimilar to that described here for barley β-glucan exohydro-lase isoenzyme ExoI. Furthermore, other family 3 glycosylhydrolases, such as the β-glucosidase from nasturtium(Tropaeolum majus) [10] and the β-glucan glucohydrolasefrom Nicotiana tabacum (N Koizumi, unpublished results,GenBank accession number G3582436) and Pseudomonasfluorescens [11], which have sequence identities relative tothe barley β-glucan exohydrolase of 71%, 72% and 44%,respectively, are likely to adopt similar protein folds.
However, the (1→3)-β-glucan exohydrolase from thehuman pathogenic fungus Candida albicans, which is amember of the family 5 group of glycosyl hydrolases [12,13],will probably exhibit significant structural differences.
Domain structureBarley β-glucan exohydrolase isoenzyme ExoI is a glyco-sylated protein consisting of two distinct domains, whichare connected by a helix-like strand of 16 residues(Figures 1 and 2). The first domain (residues 1–357) formsan (α/β)8 barrel and the second domain (residues 374–559)forms an (α/β)6 sheet, consisting of five parallel β strandsand one antiparallel strand, with three helices on eitherside of it. The C terminus forms a long antiparallel loop(Figure 1). Overall, the molecule is an elongated structureof about 82 Å in length. It is 55 Å in width and 35 Å thickat the (α/β)8-barrel domain, but it tapers to 30 Å in widthand 26 Å in thickness at the (α/β)6-sheet domain.
Domain 1 is a short cylindrical structure of approximately55 Å in diameter and 35 Å in thickness and, when vieweddown the cylinder axis of the disk with domain 2 below(Figure 1), the pseudo-eightfold (α/β)8 barrel axis pointsup and out at an angle of about 45° to the right of thecylinder axis. Domain 2 is wedge-shaped and is about 45 Ålong, 26 Å wide and 26 Å thick. Domain 2 lies in closecontact with the lower cylinder surface of domain 1 andprotrudes downwards at an angle of 30°, with the upperloops of domain 2 bordering the barrel axis of domain 1.The upper surface of the molecule is quite flat, apart fromthe protrusion of carbohydrate attached to Asn498.
180 Structure 1999, Vol 7 No 2
Table 1
Data collection and MIR phasing statistics.
Native PIP MHG
Data collectionResolution (Å) 2.2 2.2 2.6Completeness* 78 (32) 75 (33) 70 (38)Multiplicity 3.7 2.5 3.5⟨I/σ(I)⟩* 20.1 (2.8) 19.8 (2.4) 11.1 (2.1)Rmerge* 5.4 (14.2) 8.2 (20.5) 9.3 (17.2)Wilson B 26.7 41.8 39.7MIR phasingResolution (Å) 90–2.3 90–2.6Sites per molecule 4† 2Total occupation 1.84 1.7Phasing power‡§ 0.9 (1.2) [0.7] 1.0 (1.1) [1.1]Rcullis
¶§ 0.77 (0.79) [0.76] 0.87 (0.89) [0.61]
*Numbers in parentheses are the values in the high resolution shell,where Rmerge = Σ(Ii–⟨I⟩)ΣIi summed over all independent reflections.†Each site was split into pairs. §The three numbers represent summationover: centric (acentric) [acentric anomalous] reflections. ‡Root meansquare (rms) fh/residual = √(Σ( Fderi–FPH)2), where fh and FPH are thecalculated heavy-atom and derivative structure factors, respectively.¶Rcullis = Σ||fh|–(|Fderi|–|FNati|)|/Σ||Fderi|–|FNati||, where Fderi and FNati are theobserved derivative and native structure factors, respectively.

The two domains are very closely associated and a shallowgroove runs along the upper edge of the interface of thedomains. In the central region of this groove is a smallpocket, and there is a cleft between the barrel domain andthe C-terminal loop, toward the right-hand side (Figure 1).A glucose moiety is bound noncovalently, but stably, inthe pocket. The pocket sits above the pseudo-eightfoldaxis of the (α/β)8 barrel of domain 1.
There are disulphide bridges connecting Cys151 andCys159 in domain 1, and Cys513 and Cys518 in domain 2.
The four remaining cysteine residues in the enzyme(Cys81, Cys 146, Cys204, Cys427) are unpaired and two ofthese (Cys146 and Cys427) are bound to mercury ions inthe crystal. Six cis proline residues were observed at posi-tions 146, 295, 318, 405, 504 and 579 in the amino acidsequence, and cis peptide bonds were found beforeresidues His207 and Val209.
The (α/β)8-barrel domainThe N terminus begins at the top of the molecule whenviewed in the orientation shown in Figure 1 (see Figure 2
Research Article Barley ββ-glucan exohydrolase Varghese, Hrmova and Fincher 181
Figure 1
A MOLSCRIPT [50] stereo representation ofthe overall structure of barley β-glucanexohydrolase isoenzyme ExoI. (a) The (α/β)8barrel that comprises domain 1 is representedby red α helices and green β strands, withpurple coils connecting them. The (α/β)6sheet that constitutes domain 2 isrepresented by orange α helices and lightblue β strands, with yellow coils connectingthem. The C terminus is represented by lightgreen antiparallel sheets. The yellow andgreen balls represent the N terminus andC terminus, respectively, and the two lightblue balls and the three dark blue ballsrepresent the two putative catalytic aminoacid residues and the three N-glycosylationsites, respectively. Helices are labelled withthe uppercase letter of the β strands (lowercase) they follow. Positive and negativesubscripts on helices represent successivehelices following or preceding thecorresponding strands, respectively. Thebound glucose moiety is shown in the active-site region at the interface of the two domains.(b) A Cα trace of the protein in the sameorientation, with every tenth residuenumbered. The colour scheme is as in (a).Transparent GRASP [51] molecular surfacesof the two domains are overlayed on thefigure, with the (α/β)8 barrel in pink, and the(α/β)6 sheet in green. The asparagine-linkedcarbohydrate sites are represented by orangespheres, and blue, red and purple spheresrepresent N-acetyl-D-glucosaminyl, mannosyland fucosyl residues, respectively.

for residue assignments). The long axis of the enzyme isvertical in Figure 1 and the barrel axis points out of thepaper and 45° to the right. There are two helices, A–2 andA–1, near the N terminus before the first β strand a of the(α/β)8 barrel is entered about halfway up the disk. Thechain exits on the upper-middle surface of the disk andwinds itself into an (α/β) pair, with helix A on the outsideand β strand b towards the left side of the disk axis. Thismotif is repeated with helix B and β strand c, the β barrelwinding anti-clockwise. Exiting c is a long strand thatends with a small helix C1 and turns back under the barrelto enter β strand d via a long helix C2. This loop and thetwo helices are the main contacts with the second domain.The polypeptide exits β strand d as an extended strandand loop, before entering β strand e via helix D (now onthe right-hand side of the disk and above helix C2) and along loop under the disk. This is repeated for the strandexiting β strand e, except that there are two closely placedcis peptide bonds between Lys206 and His207 andPhe208 and Val209 in the extended strand that enters along helical segment (E1, E2) prior to entering β strand f(see Figure 3 for an electron-density map of this region).The typical (α/β) motif is repeated for helix F andβ strand g (Figure 1). Between β strands g and h are two
helices (G1, G2); the last β strand, h, completes the (α/β)8barrel. Exiting β strand h at the top of the barrel is a longhelix H1, which runs up along the top of the moleculebefore turning down at the top of the molecule as the longhelix H2. The chain continues along the underside of themolecule in a helix-like, extended conformation that con-nects the first domain to the second domain (Figure 1).
Most of the interactions between the domains involvehydrogen bonds or are ionic in nature. Interactionsbetween mainchain peptide atoms and sidechain groupspredominate and there are a number of charged groups(His98, His110, Arg526, Arg569) buried in the interfaceregion. Hydrophobic interactions between the domains arerestricted to the lower region of the interface (Figure 1).
The (α/β)6-sheet domainThe chain enters the second domain via a long helix H3,which lies underneath the molecule as it is orientated inFigure 1. It subsequently forms the sixth (antiparallel)outer β strand i (to the right of domain 2) of a six-strandedβ-sheet structure. The chain then loops across the bottomof the molecule and enters the second β strand j, afterwhich, via the two helices J1 and J2 that make up the left
182 Structure 1999, Vol 7 No 2
Figure 2
A-2 A-1 a A b B
D Y V L Y K D A T K P V E D R V A D L L G R M T L A E K I G Q M T Q I E R L V A T P D V L R D N F I G S L L S G G G S V P R K G A T A K E W Q D M V D G F Q K A1 10 20 30 40 50 60 70 80
cisc C1 C2 d
C M S T R L G I P M I Y G I D A V H G Q N N V Y G A T I F P H N V G L G A T R D P Y L V K R I G E A T A L E V R A T G I Q Y A F A P C I A V C R D P R W G R C Y90 100 110 120 130 140 150 160
cis cis CHOD e E1 E2
E S Y S E D R R I V Q S M T E L I P G L Q G D V P K D F T S G M P F V A G K N K V A A C A K H F V G D G G T V D G I N E N N T I I N R E G L M N I H M P A Y K N170 180 190 200 210 220 230 240
Catalytic cis cisf F g G1 G2 h
A M D K G V S T V M I S Y S S W N G V K M H A N Q D L V T G Y L K D T L K F K G F V I S D W E G I D R I T T P A G S D Y S Y S V K A S I L A G L D M I M V P N K250 260 270 280 290 300 310 320
H1 H2 H3 i
Y Q Q F I S I L T G H V N G G V I P M S R I D D A V T R I L R V K F T M G L F E N P Y A D P A M A E Q L G K Q E H R D L A R E A A R K S L V L L K N G K T S T D330 340 350 360 370 380 390 400
cisj J1 J2 k K
A P L L P L P K K A P K I L V A G S H A D N L G Y Q C G G W T I E W Q G D T G R T T V G T T I L E A V K A A V D P S T V V V F A E N P D A E F V K S G G F S Y A410 420 430 440 450 460 470 480
Catalytic CHO cisl L m M n N
I V A V G E H P Y T E T K G D N L N L T I P E P G L S T V Q A V C G G V R C A T V L I S G R P V V V Q P L L A A S D A L V A A W L P G S E G Q G V T D A L F G D490 500 510 520 530 540 550 560
cis CHO
F G F T G R L P R T W F K S V D Q L P M N V G D A H Y D P L F R L G Y G L T T N A T K K Y570 580 590 600
Structure
An ALSCRIPT [52] representation of the sequence of barley β-glucanexohydrolase isoenzyme ExoI, colour-coded and labelled as describedin Figure 1, to represent the secondary structural elements of thethree-dimensional fold of the molecule. The secondary structuralelements are displayed below the sequence as cylinders (α helices)
and arrows (β strands), and lines designate the connecting coils. Theputative catalytic acids (white up-arrows), N-glycosylation sites (blackup-arrows; CHO represents carbohydrate), cis-proline residues (whitedown-arrows) and non-proline cis peptide bonds (black down-arrows)are also shown.

side of domain 2, the chain enters β strand k at the bottomleft-hand side of the domain. The chain exits strand k,which forms the outermost strand of the sheet, and formsa helix K that runs down the bottom edge of the domainand on the other side of the sheet, into β strand l. Thechain then forms a long loop that approaches domain 1and, via helix L on the upper surface of the domain andpointing down the molecule, enters β strand m, whichnow runs parallel to the long axis of the molecule towardsdomain 2. This structure is repeated for helix M andβ strand n, except than the loop connecting β strand mand helix M is short. Strand n forms the final strand in asix-stranded β sheet consisting of strands k, j, l, m, n and i,where the first five strands are parallel and the sixth isantiparallel (Figure 1). Three helices K, L and M lie onthe underside of this sheet, and H3, J1, J2 and N are on thetopside of the sheet (Figure 1).
The C terminusThe final secondary structural element in the seconddomain is helix N, which runs into two antiparallel strands(residues 560–602) that proceed up the right-hand side ofthe interface between domains 1 and 2 and are perpendic-ular to the β-sheet structure (Figure 1). The antiparallelstrands form two pseudo β sheets that terminate at theunderside of the molecule (Figure 1). As mentionedearlier, the last three residues at the C terminus are notvisible in the electron-density map, presumably becauseof high thermal mobility in this region. It is also possiblethat the residues have been removed before or duringpurification by carboxypeptidases that are present in thegerminated barley grain [14]. The complete sequenceoverlayed by the secondary structure elements of ExoI isshown on Figure 2.
Carbohydrate structureFrom the electron-density map, carbohydrate has beenidentified at three sites. Amino acid residues Asn221,
Asn498 and Asn600 appear to have extra electron densityemanating from their sidechain amide nitrogen atoms.These are the only potential N-glycosylation sites on theenzyme and no O-linked carbohydrate was observed. Themolecular mass deduced from the cDNA sequence is65,390 Da. The overall molecular mass as determined bymatrix-assisted laser desorption time-of-flight (MALDI-TOF) mass spectrometry was 68,592 Da (± 46 Da; datanot shown). Thus, it can be calculated that N-linked car-bohydrate constitutes approximately 4.7% by weight ofthe glycosylated protein.
A single N-acetyl-D-glucosamine (GlcNAc) residueattached to Asn221 can be discerned in the electron-density map, but no further monosaccharide residues canbe seen at this position. Similarly, electron density atAsn600 reveals that this residue is glycosylated but theresolution is very poor and the structure of the carbohy-drate could not be modelled. However, the data on thecarbohydrate attached to Asn498 are sufficiently clear toenable the definition of several glycosyl residues(Figure 4). The oligosaccharide consists of a core structureof two (1→4)-β-linked GlcNAc residues, followed by abranching mannosyl residue. The (1→3)-β-linked branchof this mannosyl residue, which would project away fromthe enzyme’s surface, is not observed, but the C6 atom islinked to another mannosyl residue, which, in turn, is(1→2)-β-linked to a terminal GlcNAc residue. A (1→3)-α-linked fucosyl residue is also linked to the GlcNAc that isattached to Asn498 (Figure 4). There is a suggestion in theelectron density of a (1→2)-linked xylosyl residueattached to the branching mannosyl residue. The struc-ture is represented diagrammatically as follows:
GlcNAcβ1→2Manα1→6Manβ1→4GlcNAcβ1→4GlcNAcβ1→N-Asn498
2 3
↑ ↑Xylβ1(?) Fucα1
Research Article Barley ββ-glucan exohydrolase Varghese, Hrmova and Fincher 183
Figure 3
A MOLSCRIPT [50] stereo representation ofthe region of amino acid residues 206–211 ofdomain 1 of the barley β-glucan exohydrolaseisoenzyme ExoI. The cis peptide linkagesbetween Lys206 and His207 and betweenPhe208 and Val209 are clearly observed inthe electron density of the 2Fobs–Fcal map,and are represented by a green caged meshcontour (s = 2.1), where Fobs and Fcal are theobserved and refined structure factors.Carbon, oxygen, sulphur and nitrogen atomsare represented by black, red, yellow and bluespheres, respectively.
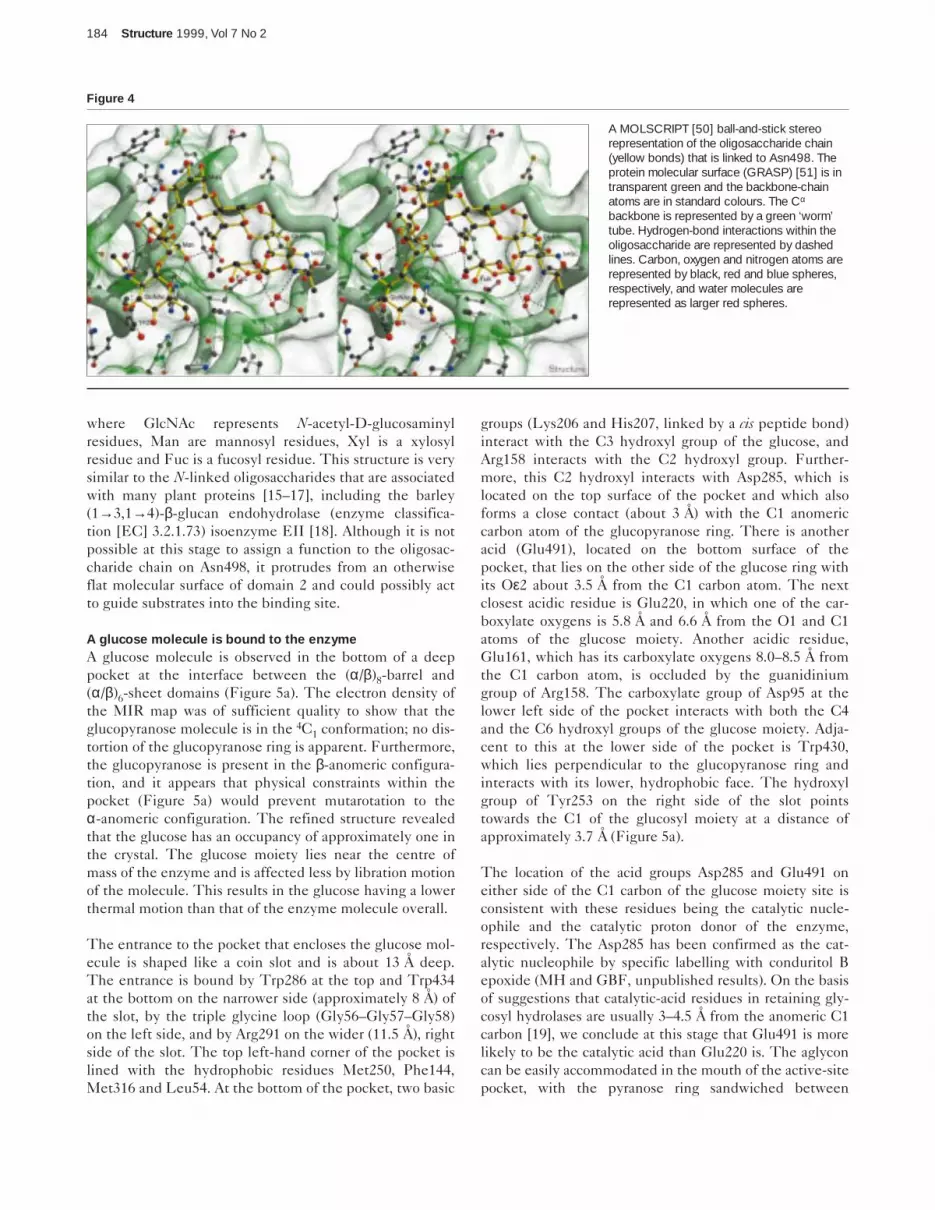
where GlcNAc represents N-acetyl-D-glucosaminylresidues, Man are mannosyl residues, Xyl is a xylosylresidue and Fuc is a fucosyl residue. This structure is verysimilar to the N-linked oligosaccharides that are associatedwith many plant proteins [15–17], including the barley(1→3,1→4)-β-glucan endohydrolase (enzyme classifica-tion [EC] 3.2.1.73) isoenzyme EII [18]. Although it is notpossible at this stage to assign a function to the oligosac-charide chain on Asn498, it protrudes from an otherwiseflat molecular surface of domain 2 and could possibly actto guide substrates into the binding site.
A glucose molecule is bound to the enzymeA glucose molecule is observed in the bottom of a deeppocket at the interface between the (α/β)8-barrel and(α/β)6-sheet domains (Figure 5a). The electron density ofthe MIR map was of sufficient quality to show that theglucopyranose molecule is in the 4C1 conformation; no dis-tortion of the glucopyranose ring is apparent. Furthermore,the glucopyranose is present in the β-anomeric configura-tion, and it appears that physical constraints within thepocket (Figure 5a) would prevent mutarotation to theα-anomeric configuration. The refined structure revealedthat the glucose has an occupancy of approximately one inthe crystal. The glucose moiety lies near the centre ofmass of the enzyme and is affected less by libration motionof the molecule. This results in the glucose having a lowerthermal motion than that of the enzyme molecule overall.
The entrance to the pocket that encloses the glucose mol-ecule is shaped like a coin slot and is about 13 Å deep.The entrance is bound by Trp286 at the top and Trp434at the bottom on the narrower side (approximately 8 Å) ofthe slot, by the triple glycine loop (Gly56–Gly57–Gly58)on the left side, and by Arg291 on the wider (11.5 Å), rightside of the slot. The top left-hand corner of the pocket islined with the hydrophobic residues Met250, Phe144,Met316 and Leu54. At the bottom of the pocket, two basic
groups (Lys206 and His207, linked by a cis peptide bond)interact with the C3 hydroxyl group of the glucose, andArg158 interacts with the C2 hydroxyl group. Further-more, this C2 hydroxyl interacts with Asp285, which islocated on the top surface of the pocket and which alsoforms a close contact (about 3 Å) with the C1 anomericcarbon atom of the glucopyranose ring. There is anotheracid (Glu491), located on the bottom surface of thepocket, that lies on the other side of the glucose ring withits Oε2 about 3.5 Å from the C1 carbon atom. The nextclosest acidic residue is Glu220, in which one of the car-boxylate oxygens is 5.8 Å and 6.6 Å from the O1 and C1atoms of the glucose moiety. Another acidic residue,Glu161, which has its carboxylate oxygens 8.0–8.5 Å fromthe C1 carbon atom, is occluded by the guanidiniumgroup of Arg158. The carboxylate group of Asp95 at thelower left side of the pocket interacts with both the C4and the C6 hydroxyl groups of the glucose moiety. Adja-cent to this at the lower side of the pocket is Trp430,which lies perpendicular to the glucopyranose ring andinteracts with its lower, hydrophobic face. The hydroxylgroup of Tyr253 on the right side of the slot pointstowards the C1 of the glucosyl moiety at a distance ofapproximately 3.7 Å (Figure 5a).
The location of the acid groups Asp285 and Glu491 oneither side of the C1 carbon of the glucose moiety site isconsistent with these residues being the catalytic nucle-ophile and the catalytic proton donor of the enzyme,respectively. The Asp285 has been confirmed as the cat-alytic nucleophile by specific labelling with conduritol Bepoxide (MH and GBF, unpublished results). On the basisof suggestions that catalytic-acid residues in retaining gly-cosyl hydrolases are usually 3–4.5 Å from the anomeric C1carbon [19], we conclude at this stage that Glu491 is morelikely to be the catalytic acid than Glu220 is. The aglyconcan be easily accommodated in the mouth of the active-sitepocket, with the pyranose ring sandwiched between
184 Structure 1999, Vol 7 No 2
Figure 4
A MOLSCRIPT [50] ball-and-stick stereorepresentation of the oligosaccharide chain(yellow bonds) that is linked to Asn498. Theprotein molecular surface (GRASP) [51] is intransparent green and the backbone-chainatoms are in standard colours. The Cα
backbone is represented by a green ‘worm’tube. Hydrogen-bond interactions within theoligosaccharide are represented by dashedlines. Carbon, oxygen and nitrogen atoms arerepresented by black, red and blue spheres,respectively, and water molecules arerepresented as larger red spheres.

Trp286 and Trp434, and some of the hydroxyl groups ofthe sugar could be stabilized by Arg291. It might be arguedthat the glucose-binding pocket of the barley β-glucan exo-hydrolase is not associated with the active site, because‘non-catalytic’ glucose-binding sites are found on a numberof other proteins. However, the glucose-binding site on thebarley enzyme (Figure 5a) is quite different from non-cat-alytic glucose-binding sites on proteins such as glycogenphosphorylase [20], glucose/galactose-binding protein [21]and xylose isomerase [22], where acidic amino acidresidues on either side of the C1 carbon are invariablyabsent. In addition, the glucose bound to the barleyenzyme interacts exclusively with charged and hydropho-bic residues, whereas the glucose bound to the non-cat-alytic sites of the other glucose-binding proteins interactswith neutral residues such as asparagine. Finally, the sug-gestion that the pocket represents the active site of theenzyme has been confirmed by the observation that a com-petitive inhibitor binds to the enzyme in exactly the sameregion, and simultaneously displaces the bound glucose(unpublished results). Deeper, funnel-shaped pockets arefound at the active sites of β-glucosidases and 6-phospho-β-galactosidases [23–25], but these other pockets are muchlonger, are narrower at the exit in comparison with thebarley β-glucan exohydrolase and can accommodate sub-strates with up to eight glucosyl residues [25,26].
The presence of the glucose moiety in the active site ofthe enzyme raises questions regarding its origin. One pos-sibility is that the glucose was present in tissue extractsand was loosely bound in the active-site region of theenzyme during its purification. The protein underwentseveral cycles of purification prior to crystallization,including fractional precipitation with ammonium sul-phate, anion and cation ion-exchange chromatography,chromatofocusing, hydrophobic-interaction chromatogra-phy and gel filtration [1]. Given the relatively low bindingconstant of glucose to the enzyme [1], one would expectthat any glucose in the original tissue extracts that mightbind weakly to the active site would have diffused awayby the time the enzyme was purified and crystallized. Noglucose was used in buffers during the purification proce-dure [1]. It would appear therefore that the glucose istightly bound to the barley β-glucan exohydrolase. Consis-tent with this conclusion is the observation from therefined structure that the glucose has thermal parameterssimilar to those of the surrounding amino acid residues(data not shown). This indicates a very high binding affin-ity for the glucose molecule at this site.
It might also be possible that loosely bound glucose origi-nally present in the active site has been chemically modi-fied to form a lactone or some other derivative that binds
Research Article Barley ββ-glucan exohydrolase Varghese, Hrmova and Fincher 185
Figure 5
(a) (b)
Structure
Glucose trapped in the active site of β-glucan exohydrolase isoenzymeExoI. (a) A MOLSCRIPT [50] stereo representation of the active-sitepocket of barley β-glucan exohydrolase isoenzyme ExoI. Nearest-neighbour hydrogen-bond interactions between the bound glucosemoiety (cyan) and the contact amino acid residues (standard colours)in the active site pocket are shown as dashed lines. The molecularsurfaces (GRASP) [51] of domains 1 and 2 are represented bytransparent red and green surfaces, respectively. Carbon, oxygen andnitrogen atoms are represented by black, red and blue spheresrespectively, and water molecules are represented as larger redspheres. (b) A schematic drawing of the bonding interactions of thebound glucose molecule, shown as 4C1 β-glucopyranose with atomicnumbering of C atoms, in the active site of the barley β-glucan
exohydrolase isoenzyme ExoI. The dotted lines indicate hydrogenbonds and ionic interactions between the glucose and acidic andbasic amino acid residues of the enzyme, and a water molecule(distances are in Å). Represented on an arc at the top left-hand side ofthe diagram are a group of four hydrophobic residues (Leu54, Met316,Phe144 and Met250), which interact with the C3, C5 and C6 atomson the hydrophobic face of the bound glucopyranose ring.Represented on an arc on the other, lower, side of the ring arehydrophobic interactions of Trp430 with carbon atoms C2 and C4.Note that the carboxylate oxygen of Asp285, which is not H-bonded tothe C2 hydroxyl of the sugar, has a close contact (less than 3 Å) withthe anomeric C1 carbon of the glucopyranose ring (not shown).

to the enzyme more strongly. In the case of viral neu-raminidase, it has been shown that neuraminic acid in theactive site is slowly converted to deoxydehydroneuraminicacid, which is a more potent inhibitor of neuraminidasethan is neuraminic acid [27]. Because the barley β-glucanexohydrolase was purified over a period of several weeksand crystals grew for 100 days or more [9], it is possiblethat any weakly bound glucose originally present couldhave been oxidised to gluconolactone during theprolonged purification and crystallization procedures.D-Glucono-1,5-lactone is a considerably stronger inhibitorof the enzyme than glucose [1]. If the bound moiety in theactive site were indeed a lactone, enzyme activity wouldbe measurably lowered. However, on redissolving thecrystals, the specific activity was the same as the native,freshly purified enzyme [9].
On the basis of these considerations and data, we con-clude that the glucose probably represents the product ofthe enzyme-catalysed reaction and that the glucoseproduct has not been released from the surface of thebinary enzyme–product complex after hydrolysis is com-pleted. This further implies that the glucose remainsbound to the enzyme until another, incoming substratemolecule displaces it. But what would be the structuralbasis for this release mechanism as the incoming substratemolecule binds to the binary enzyme–product complex?The two-domain structure of the active site could allow achange in the spatial orientation between domains whensubstrate binds to the enzyme or when the aglycone disso-ciates after hydrolysis. The long loop (residues 358–373)that connects the two domains (Figure 1) has a partiallyformed helical structure and could, when an incomingsubstrate molecule is bound, become fully helical. Thus,changes in the conformation of the loop could allow it toact as a hinge that would, in turn, allow the two domains toseparate in such a way that conformational changes in theregion of the pocket would release the bound glucose.
Substrate bindingThe barley β-glucan exohydrolase is inhibited by glucose,the product of hydrolysis that is released from nonreducingtermini of a range of substrates, and by glucose analoguessuch as conduritol B epoxide, D-glucono-1,5-lactone and2,4-dinitrophenyl 2-deoxy-2-fluoro-glucopyranoside [1](MH and GBF, unpublished results). On this basis, thepocket containing the noncovalently bound glucose moietyin the crystal structure is likely to represent the active siteof the enzyme. The site lies at the junction of the (α/β)8domain and the (α/β)6 domain, above the pseudo-eightfoldaxis of the (α/β)8 barrel. According to the structural classifi-cation of proteins (SCOP), a location for the active siteabove the pseudo-eightfold axis is typical of enzymes with(β/α)8 folds and, in particular, of members of the β-gly-canase family of the glycosyltransferase superfamily [28].On the other hand, the barley β-glucan exohydrolase does
not appear to belong to the 4/7 superfamily [29] of(β/α)8-barrel glycosyl hydrolases, where the catalyticacid/base and nucleophile active site residues are found atthe carboxy ends of the fourth and seventh β strands of thebarrel, respectively.
The putative active-site pocket is lined by 17 amino acidresidues, three of which (Trp430, Trp434 and Glu491) arepart of the second domain. The site is characterised by thepresence of several acidic and basic amino acid residues,and by two hydrophobic patches that interact with thebound glucose moiety (Figure 5). Among these residuesare the putative active-site residues Asp285 and Glu491;the latter arises from domain 2. Such an arrangement ofamino acid residues in the active-site region is reminiscentof the influenza virus neuraminidase [27], which is a glycanexohydrolase from family 34 [7,30]. In the neuraminidase,Glu276 forms short H bonds with hydroxyl groups of thetriol group of neuraminic acid, Asp151 is likely to be thecatalytic acid, and the nucleophile is probably Tyr406 [27].
In the active-site region of the barley β-glucan exohydro-lase, the hydroxyl groups of the bound glucose are allinvolved in multiple hydrogen-bonding and ionic interac-tions with seven active-site amino acid residues(Figure 5b). The C1 hydroxyl of the glucose forms twohydrogen bonds with Glu491 and one with Tyr253, the C2hydroxyl interacts with both Arg158 and Asp285, the C3hydroxyl forms three tetrahedrally directed interactionswith Arg158, Lys206 and His207, the C4 hydroxyl inter-acts with both Asp95 and Lys206, and the C6 hydroxylinteracts with Asp95 and a water molecule (Figure 5b).Hydrophobic interactions of Leu54, Met316, Phe144 andMet250 with the C3H, C5H and C6H regions on thehydrophobic face of the glucopyranose ring, together withinteractions between Trp430 and the C2H and C4H onthe other face, further stabilize the glucose molecule inthe active-site pocket of the enzyme.
The likely locations of the catalytic and binding sites onthe barley enzyme can also be gleaned from analysis ofresidues that are conserved among closely related family 3glycosyl hydrolases [1, 10, 11; AJ Harvey and GBF, unpub-lished results, GenBank accession number AF102868;N Koizumi, unpublished results, GenBank accessionnumber G3582436]. Apart from the substitution of Tyr253in the barley enzyme with a Phe residue in the corre-sponding position of the bacterial enzyme [11], thearrangement of amino acid residues in the active-sitepockets (Figure 5) is totally conserved.
The active site and the hydrolytic mechanismOn the basis of the bell-shaped pH dependence of barleyβ-glucan exohydrolase activity, Hrmova and Fincher [6]concluded that at least two ionizable amino acid residuesparticipate in catalysis, and ionization constants for the
186 Structure 1999, Vol 7 No 2

two catalytic amino acid residues were estimated atapproximately 4.7 and 5.9. Furthermore, proton NMRshowed that the anomeric configuration of releasedglucose is retained during hydrolysis [1]. Enzymic hydrol-ysis of glycosidic linkages in oligosaccharides and polysac-charides is widely believed to be initiated by protonationof the glycosidic O atom, via proton transfer from anactive-site-located catalytic acid [19,31]. Following proto-nation, the aglycone is released from the enzyme surfaceand the reducing terminal residue of the bound substrateis converted to a positively charged oxycarbonium ion.The oxycarbonium ion is stabilized, either electrostaticallyor by a covalent enzyme–substrate intermediate, by anucleophilic amino acid residue, which usually carries acarboxylate group. Hydration of the oxycarbonium orhydrolysis of the covalent intermediate results in theregeneration of the glycosyl residue and reprotonation ofthe catalytic acid.
In the barley β-glucan exohydrolase structure, the twoacidic amino acid residues Asp285 and Glu491, which arisefrom domains 1 and 2 of the enzyme, respectively, areappropriately positioned in relation to the C1 hydroxylgroup of the bound glucose molecule and have been iden-tified as likely catalytic residues (Figure 5). One of the car-boxylate oxygens of Asp285 has a close contact (less than3 Å) with the C1 carbon of the glucose moiety. This car-boxylate oxygen could stabilize the positive charge on theC1 carbon of the oxycarbonium ion during catalysis, iden-tifying it as the most likely candidate for the nucleophilein the catalytic mechanism. This situation is similar to theshort O…C=O contacts found in small organic compounds,in which the oxygen atom acts as the nucleophile in anincipient nucleophilic addition reaction with the carbonatom [32]. Similar short contacts have been observed inthe structures of neuraminidase/sialic acid complexes [27].In addition, in bovine trypsin complexed with the bovine-trypsin inhibitor, the oxygen atom of the putative serinenucleophile (Ser195) has a short contact (2.7 Å) with apeptide carbonyl (Lys15) of the inhibitor [33].
In the barley β-glucan exohydrolase, Glu491 is positionedon the other side of the glucopyranose ring (in comparisonwith the catalytic nucleophile Asp285), approximately 3.5 Åfrom the anomeric C1 carbon (Figure 5), and is a strongcandidate for the catalytic acid. Although Glu491 appearssolvated in the structure, it could facilitate the activation ofa water molecule that would subsequently act as the protondonor in the catalytic mechanism. The presence of thehydrophobic residues Trp430 and Trp434 in close proxim-ity to Glu491 (Figure 5a) might be expected to increase thepKa of Glu491, although interactions with Arg158 wouldtend to deprotonate the acid or decrease its pKa.
The location of Asp285 in the barley β-glucan exohydro-lase (Figure 5) above the hydrophobic face of the bound
glucopyranose ring is consistent with the enzyme having aretaining mechanism. Proton NMR has been used previ-ously to demonstrate that β-glucose is indeed releasedduring hydrolysis of a variety of substrates by the barleyenzyme [1]. Further support for a role for Asp285 andGlu491 in catalysis is provided by the observation thatthese residues are amongst the most highly conservedresidues in family 3 glycosyl hydrolases, although theneighbouring Glu220 is also highly conserved among theplant exohydrolases (data not shown). It should now bepossible to use active-site-directed inhibitors [25] and site-directed mutagenesis [34] to confirm the role of theseresidues in catalysis.
As mentioned earlier, the 16-residue helix-like loop thatconnects the two domains (Figure 1) could act as a hingethat would allow the two domains to move relative to eachother. Because three residues of domain 2, including theputative catalytic acid Glu491, are located close to the activesite and are likely to contribute to catalysis, movement ofthe second domain away from, or closer to, the active site ondomain 1 would almost certainly affect catalytic activity andcould be used to regulate enzyme activity.
A potential role for domain 2 in binding (1→3,1→4)-ββ-glucanKinetic data previously obtained for the barley β-glucanexohydrolase isoenzyme ExoI suggested that there ismore than one binding site on the enzyme for the(1→3,1→4)-β-glucan substrate [6]. Given that domain 1 islikely to contain the catalytic site, the additional(1→3,1→4)-β-glucan-binding site could be located on thesecond domain. Consistent with this possibility is theidentification of a potential binding site, based on thetopology of the open β-sheet domain (Figure 3). Brändén[35] has described carbohydrate-binding sites in a numberof proteins where two adjacent loops that emerge on oppo-site sides of a β sheet form a binding site between them[36]. The most likely region for (1→3,1→4)-β-glucanbinding would be on the loops that emerge from theC-terminal ends of β strands j and l (Figure 1). This wouldinvolve residues 417–440 (loop A) emerging from β strandj and residues 484–504 (loop B) emerging from β strand l.In addition, the 17-residue loop (residues 429–445) con-necting helices J1 and J2 has characteristics similar to cellu-lose-binding domains from bacterial cellulases [37], whererelatively large numbers of hydroxy amino acids, a lowabundance of charged amino acid residues and conservedTrp and Gly residues are observed (Figures 1 and 2). Itshould now be possible to experimentally confirm thelocation of the (1→3,1→4)-β-glucan-binding site by firstblocking enzyme activity by soaking crystals withinhibitors such as conduritol B epoxide [25], and subse-quently adding (1→3,1→4)-β-glucooligosaccharides to thecrystals. The (1→3,1→4)-β-glucooligosaccharides wouldbe expected to bind to the second binding site identifiedby Hrmova and Fincher [6], and enzyme inactivation
Research Article Barley ββ-glucan exohydrolase Varghese, Hrmova and Fincher 187

would ensure that the (1→3,1→4)-β-glucooligosaccharideswould not be hydrolyzed.
Comparison with other family 3 glycosyl hydrolasesCurrently, there are over 60 members of the family 3group of glycosyl hydrolases [7,30]. Multiple amino acidsequence alignments and hydrophobic-cluster analysesshow that these members fall into approximately six sub-groups that appear as clearly distinguishable branches onan unrooted phylogenetic radial tree (data not shown).The four plant members, two from barley [1] (AJ Harveyand GBF, unpublished results; GenBank accessionnumber AF102868), and one each from nasturtium [10]and tobacco (N Koizumi, unpublished results; GenBankaccession number G3582436), are closely clustered. Therelated (1,4)-β-glucan glucohydrolase from Pseudomonas[11] belongs to a different cluster. Our preliminary analy-ses indicate that each of the plant enzymes has a domainorganization that is similar to the barley β-glucan exo-hydrolase isoenzyme ExoI described here, and that aminoacid residues implicated in catalysis are highly conserved.The availability of the 3D structure for the barley enzyme(Figure 1) will now allow a detailed comparison of thestructural features and evolutionary relationships of othermembers of family 3 glycosyl hydrolases.
Biological implicationsThe presence of the β-glucan exohydrolases in youngseedlings, as well as their broad substrate specificities,raise important questions regarding the biological func-tions of these enzymes. One possible role for theβ-glucan exohydrolases could be in the auxin-inducedelongation of cells in growing coleoptiles. In this process,it has been suggested that matrix-phase polysaccharidesof cell walls, including (1→3,1→4)-β-glucans and het-eroxylans, could be partially hydrolysed, and that theresultant ‘loosening’ of noncovalent cross-links betweencellulose microfibrils could allow osmotically inducedturgor pressure to stretch the cell wall and cause the cellto expand. The β-glucan exohydrolases are certainlyassociated with cell walls in elongating coleoptiles ofboth barley and maize, and decreases in cell wall(1→3,1→4)-β-glucan levels are observed in both coleop-tiles and young leaves as development proceeds.
However, it is not clear how exohydrolases wouldmediate the ‘loosening’ process via partial exohydrolysisof cell wall (1→3,1→4)-β-glucans, unless an endohydro-lase first released a new nonreducing end. It should beemphasised that cell elongation in higher plants is notwell understood and a more thorough knowledge of theaction mechanisms of the enzymes involved in wall loos-ening could provide clues as to the mechanism of cellelongation and, hence, to the crucial and as yet unan-swered questions regarding the more general mecha-nisms of plant growth.
Of particular importance in describing the enzyme’smechanism of action will be the reconciliation of itsthree-dimensional structure with previous indicationsfrom kinetic analyses that a specific, non-catalytic(1→3,1→4)-β-glucan-binding site is present. The secondbinding site for (1→3,1→4)-β-glucans could serve toattach the enzyme to cell walls. It could simply anchorthe enzyme to (1→3,1→4)-β-glucans during hydrolysisor it could form a more integral component of thehydrolytic mechanism, for example, by feeding thenonreducing terminus of the polysaccharide into the cat-alytic site. If the specific (1→3,1→4)-β-glucan-bindingsite enables the enzyme to attach to fibrillar or morerefractory regions of the wall, then the enzyme mightdisrupt these regions physically to facilitate overallenzyme penetration and hydrolysis of the wall. Thiscould also provide opportunities for engineering theenzyme to allow it to bind to other insoluble substrates,or to express the (1→3,1→4)-β-glucan binding domainfor use as an ‘antibody’ to locate the polysaccharide inhistochemical studies.
Materials and methodsEnzyme purification and characterizationThe barley β-glucan exohydrolase isoenzyme ExoI was purified fromyoung seedlings as previously described [1,6]. The purity of thepreparation was confirmed by the presence of a single band duringsodium dodecyl sulphate (SDS) polyacrylamide gel electrophoresisand by N-terminal amino acid sequence analysis of the first 100residues, which demonstrated that the protein corresponded toβ-glucan exohydrolase isoenzyme ExoI. In addition, recoveries ofamino acid residues during sequencing were close to theoreticalvalues and no secondary sequences were observed. The molecularmass of the purified enzyme was estimated on a Finnigan Lasermat2000 MALDI-TOF mass spectrometer (Finnigan MAT, Hemel Hemp-stead, UK), using sinapinic acid as matrix.
Crystallization and data collectionCrystals were obtained as described by Hrmova et al. [9]. Data werecollected from a single large crystal (0.8 mm × 0.6 mm × 0.4 mm) in aglass capillary at 18°C on a Rigaku R-axis II Image Plate X-ray detector,mounted on a MAC Science SRA M18XH1 rotating anode X-ray gen-erator, which was operated at 47 kV and 60 mA with focussing mirrorsfor Cu Kα. Data-collection and heavy-atom refinement statistics aregiven in Table 1. A native data set of 148,526 observations (> 1σ)were collected and reduced to 40,310 unique reflections. The crystalhad a tetragonal space group P41212 or P43212, with cell edges102.09 Å × 102.09 Å × 184.50 Å. Data from a PIP [di-µ-iodo bis(ethyl-enediamine) di-platinum (II) nitrate] derivative were collected from thesame crystal by removing the seal at one end of the capillary and dis-solving a grain of PIP into the mother liquor. The capillary was resealedfor 18 h, which allowed diffusion of PIP into the crystal. The crystal wassubsequently pushed out of the mother liquor and data were collected.A total of 106,488 intensities (> 1σ) were collected, reducing to42,372 unique reflections. A third data set was collected on a secondsmaller crystal, obtained by micro-seeding techniques [38,39], soakedin 5 mM MHG (methyl mercury chloride) for 18 h. A total of 83,154observations, reducing to 23,992 unique observations, were collectedto 2.5 Å resolution. All data were processed using the HKL software[40] (see Table 1 for data collection statistics).
Heavy-atom location and refinementFor the PIP derivative, four platinum-binding sites were located by differ-ence Patterson methods and vector-verification methods using the
188 Structure 1999, Vol 7 No 2

PROTEIN software [41]. These sites were also located in the anomalousdifference Patterson and were indicative of a strong anomalous signal.Two mercury-binding sites were located in the MHG derivative by similarmethods and, using difference Fourier methods, each derivative gave aconsistent set of heavy-atom sites for the other derivative in the spacegroup P43212. Using anisotropic temperature factors, the platinum siteswere split into two sites along the direction of maximum anisotropy.
Heavy-atom refinement and phasing (see Table 1 for statistics) werecarried out by the maximum likelihood Bayesian methodology, using theprogram SHARP [42] with data from the PIP derivative to 2.3 Å andfrom the MHG derivative to 2.5 Å. A 2.3 Å map using the SHARP MIRphases was solvent-flattened by the Solomon method [43] using 55%solvent (calculated solvent content of 63%) and the phases extendedto 2.2 Å. A reliability index based on calculated structure factors fromthe back-transformed solvent-flattened map was 0.167.
Structure determination and refinementThe protein was clearly interpretable in the solvent-flattened map. Thehigh quality of the map was probably due to the lack of systematic errorsin the differences between the native and the PIP data (collected on thesame crystal), the low mosaic spread (< 0.1°) of the large crystal, andimprovements in heavy-atom refinement [42]. The map was skeletonised,and this was used as a guide for tracing the Cα chain of the protein,using the graphics program O version 6.1 [44]. All residues in the chainwere identified unambiguously, apart from the last three C-terminal aminoacids (residues 603–605). An atomic model was built using Lego frag-ments in the program O, including N-linked carbohydrates at Asn221and Asn498, and a glucose molecule bound in the putative active site ofthe molecule. Water molecules could be seen in this map, but were notbuilt. The atomic model was refined with the maximum-likelihood refine-ment program REFMAC [45], and SIGMAA-weighted maps [46] wereused for model building. One hexosyl residue attached to Asn221 andseven hexosyl residues attached to Asn498 were refined. A putativexylosyl residue attached to the C2 hydroxyl of the branching mannosylresidue in the carbohydrate attached to Asn498 was observed, but notmodelled. Two hundred and twenty water molecules were added to thestructure, using the program ARPP [47], by adding 20 water moleculesat a time to peaks (> 5σ) in the difference Fourier. Refinement cross vali-dation was carried out by selecting, at random, 5% of the data (2128reflections), which were used throughout the refinement to calculate freeR factors. Final refinement statistics are presented in Table 2. The esti-mated overall coordinate error based on R value, Free R value andmaximum likelihood is 0.195, 0.176 and 0.106 Å, respectively.
The stereochemical quality of the model was assessed with theprogram PROCHECK [48]. A Ramachandran plot indicated that90.3% of the amino acid residues are in the ‘most-favoured’ regions,9.1% are in ‘additionally allowed’ regions, 0.4% are in ‘generouslyallowed’ regions and 0.2% in ‘disallowed’ regions. The ‘disallowed’residue is Ile432, but this residue is well-defined by electron density.
Accession numbersThe atomic coordinates and structure factors of the refined proteinmodel of barley β-glucan exohydrolase isoenzyme ExoI determined here
have been deposited with the Brookhaven Protein Data Bank [49]under codes 1EX1 and R1EX1SF, respectively.
AcknowledgementsWe thank Peter M Colman for his continuing and high level support, BrianSmith for useful discussions and ideas relating to catalytic mechanisms,Andrew J Harvey for providing amino acid sequence data, Ross Degori forhelp with refinement of the structure and Bert van Donkelaar for skilled tech-nical assistance. We are grateful to Bruce Stone for his advice and ongoinginterest. The work was supported by grants (to GBF) from the AustralianResearch Council and the Grains Research and Development Corporationof Australia.
References1. Hrmova, M., et al., & Fincher, G.B. (1996). Barley β-D-glucan
exohydrolases with ß-D-glucosidase activity. J. Biol. Chem.271, 5277-5286.
2. Kotake, T., Nagawa, N., Takeda, K. & Sakurai, N. (1997). Purificationand characterization of wall bound exo-1,3-β-glucanase from barley(Hordeum vulgare L.) seedlings. Plant Cell Physiol. 38, 194-200.
3. Sakurai, N. & Masuda, Y. (1978). Auxin-induced changes in barleycoleoptile cell wall composition. Plant Cell Physiol. 19, 1217-1223.
4. Inouhe, M., McClellan, M. & Nevins, D.J. (1997). Developmentalregulation of polysaccharide metabolism and growth in the primary cellwalls of maize. Int. J. Biol. Macromol. 21, 21-28.
5. Cline, K. & Albersheim, P. (1981). Host-pathogen interactions. XVI.Purification and characterization of a glucosyl hydrolase/transferasepresent in the walls of soybean cells. Plant Physiol. 68, 207-220.
6. Hrmova, M. & Fincher, G.B. (1998). Barley β-D-glucan exohydrolases.Substrate specificity and kinetic properties. Carbohydr. Res.305, 209-221.
7. Henrissat, B. & Bairoch, A. (1993). New families in the classification ofglycosyl hydrolases based on amino acid sequence similarities.Biochem. J. 293, 781-788.
8. Varghese, J.N., Garrett, T.P.J., Colman P.M., Chen, L., Høj, P.B. &Fincher, G.B. (1994). Three-dimensional structures of two plant β-glucan endohydrolases with distinct substrate specificities. Proc.Natl Acad. Sci. USA 91, 2785-2789.
9. Hrmova, M., Varghese, J.N., Høj, P.B. & Fincher, G.B. (1998).Crystallization and preliminary X-ray analysis of β-D-glucanexohydrolase isoenzyme ExoI from barley (Hordeum vulgare). ActaCryst. D 54, 687-689.
10. Crombie, H.J., Chengappa, S., Hellyer, A. & Reid, J.S.G. (1998). Axyloglucan oligosaccharide-active, transglycosylating β-D-glucosidasefrom the cotyledons of nasturtium (Tropaeolum majus L.) seedlings —purification, properties and characterization of a cDNA clone. Plant J.15, 27-38.
11. Rixon, J.E., Ferreira, L.M.A., Durrant, A.J., Laurie, J.I., Hazlewood, G.P.& Gilbert, H.J. (1992). Characterization of the gene celD and itsencoded product 1,4-β-glucan glucohydrolase D from Pseudomonasfluorescens subsp. cellulosa. Biochem. J. 285, 947-955.
12. Cutfield, S., Brooke, G., Sullivan, P. & Cutfield, J. (1992).Crystallization of the exo-(1,3)-β-glucanase from Candida albicans. J. Mol. Biol. 225, 217-218.
13. MacKenzie, L.F., Brooke, G.S., Cutfield, J.F., Sullivan, P.A. & Withers,S.G. (1997). Identification of Glu330 as the catalytic nucleophile ofCandida albicans exo-β-(1,3)-glucanase. J. Biol. Chem.272, 3161-3167.
14. Mikola, J. (1983). Proteases, peptidases, and inhibitors ofendogenous proteinases in germinating seeds. In Seed Proteins.(Daussant, J., Mosse, J. & Vaughan, J., eds), pp. 35-52, AcademicPress, London.
15. Takahashi, N., et al., & Arata, Y. (1986). Xylose-containing commonstructural unit in N-linked oligosaccharides of laccase from sycamorecells. Biochemistry 25, 388-395.
16. Sturm, A., Johnson, K.D., Szumilo, T., Albein, A.D. & Chrispeels, M.J.(1987). Subcellular localization of glycosidases andglycosyltransferases involved in the processing of N-linkedoligosaccharides. Plant Physiol. 85, 741-745.
17. Imberty, A., Delage, M-M., Bourne, Y., Cambillau, C. & Perez, S. (1991).Data bank of three-dimensional structures of disaccharides: Part II,N-acetyllactosaminic type N-glycans. Comparison with the crystalstructure of a biantennary octasaccharide. Glycoconjugate J. 8, 456-483.
18. Harthill, J.E. & Thomsen, K.K. (1995). Analysis of glycan structures ofbarley (1,3;1,4)-β-D-glucan 4-glucanohydrolase isoenzyme EII. PlantPhysiol. Biochem. 33, 9-18.
Research Article Barley ββ-glucan exohydrolase Varghese, Hrmova and Fincher 189
Table 2
Final model statistics*.
Resolution (Å) 20–2.2Number of reflections 39,456 (1633)Completeness (%) 83.3 (37.8)Rwork 17.0 (20.2)Rfree 21.1 (28.3)Rms bonds (Å) 0.013Rms angles (°) 2.1
*The numbers in parentheses are the values in the high resolution shell.

19. White, A. & Rose, D.R. (1997). Mechanism of catalysis by retaining β-glycosyl hydrolases. Curr. Opin. Struct. Biol. 7, 645-651.
20. Martin, J.L., Johnson, L.N., & Withers, S.G. (1990) Comparison of thebinding of glucose and glucose-1-phosphate derivatives to T-stateglycogen phosphorylase. Biochemistry 29, 10745-10757.
21. Mowbray, S.L. (1992). Ribose and glucose-galactose receptors.Competitors in bacterial chemotaxis. J. Mol. Biol. 227, 418-440.
22. Carrell, H.L., Glusker, J.P., Burger, V., Manfre, F., Tritsch, D. &Biellmann, J.-F. (1989) X-ray analysis of D-xylose isomerase at 1.9 Å:native enzyme complexed with substrate and a mechanism designedinhibitor. Proc. Natl Acad. Sci. USA 86, 4440-4444.
23. Barrett, T., Suresh, S.G., Tolley, S.P., Dodson, E.J. & Hughes, M.A.(1995). The crystal structure of cyanogenic, β-glucosidase from whiteclover, a family 1 glycosyl hydrolase. Structure 3, 951-960.
24. Weismann, C., Beste, G., Hengstenberg, W. & Schultz, G.E. (1995).The three-dimensional structure of 6-phospho-β-galactosidase fromLactococcus lactis. Structure 3, 961-968.
25. Hrmova, M., MacGregor, E.A., Biely, P., Stewart, R.J. & Fincher, G.B.(1998). Substrate binding and catalytic mechanism of a barleyβ-D-glucosidase/(1,4)-β-D-glucan exohydrolase. J. Biol. Chem.273, 11134-11143.
26. Burmeister, W.P., Henrissat B., Bosso, C., Cusack, S. & Ruigrok,R.W.H. (1993) Influenza B virus neuraminidase can synthesize its owninhibitor. Structure 1, 19-26.
27. Varghese, J.N., McKimm-Breschkin, J., Caldwell, J.B., Kortt, A.A. &Colman, P.M. (1992). The structure of the complex between influenzavirus neuraminidase and sialic acid, the viral receptor. Proteins 14, 327-332.
28. Murzin, A.G., Brenner, S.E., Hubbard, T. & Chothia, C. (1995). SCOP:a structural classification of proteins database for the investigation ofsequences and structures. J. Mol. Biol. 247, 536-540.
29. Jenkins, J., Leggio, L.L., Harris, G. & Pickergill, R. (1995). β-glucosidase, β-galactosidase, family A cellulases, family F xylanasesand two barley glucanases form a superfamily of enzymes with 8-foldβ/α architecture and with two conserved glutamates near the carboxy-terminal ends of β-strands four and seven. FEBS Lett. 362, 281-285.
30. Henrissat, B. (1998). Glycosidase families. Biochem. Soc. Trans.26, 153-156.
31. McCarter, J.D. & Withers, S.G. (1994). Mechanism of enzymaticglycoside hydrolysis. Curr. Opin. Struct. Biol. 4, 885-892.
32. Burgi, H.B., Dunitz, J.D. & Shefter, E. (1974). Chemical reaction paths.IV. Aspects of O…C=O interactions in crystals. Acta Cryst. B 30, 1517-1527.
33. Marquart, M., Walter, J., Deisenhofer, J., Bode, W. & Huber, R. (1983).The geometry of the reactive site and the peptide groups in trypsin,trypsinogen and its complexes with inhibitors. Acta Cryst. B 39, 480-490.
34. Chen, L., Garrett, T.P.J., Fincher, G.B. & Høj, P.B. (1995). A tetrad ofionizable amino acids is important for catalysis in barley β-glucanases.J. Biol. Chem. 270, 8093-8101.
35. Brändén, C.I. (1980). Relation between structure and function of α/βproteins. Q. Rev. Biophys. 13, 317-338.
36. Gilliland, G.L. & Quicho, F.A. (1981). Structure of the L-arabinose-binding protein from Escherichia coli at 2.4 Å. J. Mol. Biol. 146, 341-362.
37. Gilkes, N.R., Henrissat, B., Kilburn, D.G., Miller, J.C. Jr. & Warren, R.A.J.(1991) Domains in microbial β-1,4-glycanases: sequence conservation,function, and enzyme families. Microbiol. Rev. 55, 303-315.
38. McPherson, A. (1990). Current approaches to macromoleculecrystallization. Eur. J. Biochem. 189, 1-23.
39. Chayen, N.E. (1998). Comparative studies of protein crystallization byvapour diffusion and microbatch techniques. Acta Cryst. D 54, 8-15.
40. Otwinowski, Z. & Minor, W. (1996). A processing of X-ray diffractiondata collected in oscillation mode. Methods Enzymol. 276, 307-326.
41. Stiegeman, W. (1974). Protein: Program System for the CrystalStructure Analysis of Proteins. Ph.D. Dissertation, Technical UniversityMunich, Germany.
42. De la Fortelle, E. & Bricogne, G. (1997). A maximum-likelihood heavy-atom parameter refinement for multiple isomorphous replacement andmultiwavelength anomalous diffraction methods. Methods Enzymol.276, 472-494.
43. Abrahams, J.P. & Leslie, A.G.W. (1996). Methods used in thestructure determination of bovine mitochondrial F1 ATPase. ActaCryst. D 52, 30-42.
44. Jones, T.A., Zou, J.Y., Cowan, S.W. & Kjeldgaard, M. (1991). Improvedmethods for building protein models in electron density maps and thelocation of errors in these models. Acta Cryst. A 47,110-119.
45. Murshudov, G.N., Vagin, A.A. & Dodson, E.J. (1997). Refinement ofmacromolecular structures by maximum-likelihood method. Acta Cryst.D 53, 240-255.
46. Read, R.J. (1986). Improved Fourier coefficients for maps usingphases from partial structures with errors. Acta Cryst. A 42, 140-149.
47. Lamzin, V.S. & Wilson, K.S. (1993). Automated refinement of proteinmodels. Acta Cryst. D 49, 129-147.
48. Laskowski, R.A., MacArthur, M.W., Moss, D.S. & Thorton, J.M. (1993).PROCHECK: a program to check the stereochemical quality ofprotein structures. J. Appl. Cryst. 26, 283-291.
49. Bernstein, F.C., et al., & Tasumi, M. (1977). The protein data bank: acomputer-based archival file for macromolecular structures. J. Mol.Biol. 112, 535-542.
50. Kraulis, P.J. (1991). MOLSCRIPT: a program to produce both detailedand schematic plots of protein structures. J. Appl. Cryst. 24, 946-950.
51. Nicholls, A., Sharp, K. & Honig B. (1991). Protein folding andassociation: insights from the interfacial and thermodynamicproperties of hydrocarbons. Proteins 4, 281-296.
52. Barton, G.J. (1993). ALSCRIPT: a tool to format multiple sequencealignments, Protein Eng. 6, 37-40.
190 Structure 1999, Vol 7 No 2
Because Structure with Folding & Design operates a‘Continuous Publication System’ for Research Papers, thispaper has been published on the internet before being printed(accessed from http://biomednet.com/cbiology/str). Forfurther information, see the explanation on the contents page.
Top Related
Copyright © 2022 FDOKUMEN

