







Bahasa
Halaman
Hukum


Special Issue Article
Received: 27 February 2014 Revised: 16 April 2014 Accepted: 18 April 2014 Published online in Wiley Online Library: 20 May 2014
(wileyonlinelibrary.com) DOI 10.1002/psc.2651
554
J. Pept. Sci. 2014; 20:
The role of water and structure on thegeneration of reactive oxygen species inpeptide/hypericin complexes‡
Márcia I. Souza,a Emerson R. Silva,a Ygor M. Jaques,a Fabio F. Ferreira,a
Eudes E. Filetib and Wendel A. Alvesa*
Hybrid associates formed between peptide assemblies and fluorophores are attractive mainly because of their uniqueproperties for biomedical applications. Recently, we demonstrated that the production of reactive oxygen species (ROS) byhypericin and their stability in excited states are enhanced upon conjugation with L,L-diphenylalanine microtubes (FF-MNTs).Although the detailed mechanisms responsible for improving the photophysical properties of ROS remain unclear, tentativehypotheses have suggested that the driving force is the growth of overall dipolar moments ascribed either to couplingbetween aligned H2O dipoles within the ordered structures or to the organization of hypericin molecules on peptideinterfaces. To provide new insights on ROS activity in hypericin/FF-MNTs hybrids and further explore the role of water in thisrespect, we present results obtained from investigations on the behavior of these complexes organized into differentcrystalline arrangements. Specifically, we monitored and compared the photophysical performance of hypericin bound toFF-MNTs with peptides organized in both hexagonal (water-rich) and orthorhombic (water-free) symmetries. From a theoret-ical perspective, we present the results of new molecular dynamics simulations that highlight the distinct hypericin/peptideinteraction at the interface of FF-MNTs for the different symmetries. As a conclusion, we propose that although waterenhances photophysical properties, the organization induced by peptide structures and the availability of a hydrophobicenvironment surrounding the hypericin/peptide interface are paramount to optimizing ROS generation. The findingspresented here provide useful basic research insights for designing peptide/fluorophore complexes with outstanding techno-logical potential. Copyright © 2014 European Peptide Society and John Wiley & Sons, Ltd.
Additional supporting information may be found in the online version of this article at the publisher’s web site.
Keywords: peptide nanostructures; hypericin; photodynamic therapy; supramolecular chemistry; L,L-diphenylalanine
* Correspondence to: Wendel A. Alves: Centro de Ciências Naturais e Humanas,Universidade Federal do ABC, Santo André, São Paulo, SP, Brazil. E-mail:[email protected]
‡ This article is published in Journal of Peptide Science as part of the SpecialIssue devoted to contributions presented at the 1st International Conferenceon Peptide Materials for Biomedicine and Nanotechnology, Sorrento, October28–31, 2013, edited by Professor Giancarlo Morelli, Professor Claudio Tonioloand Professor Mariano Venanzi.
a Centro de Ciências Naturais e Humanas, Universidade Federal do ABC,09210-170, Santo André, SP, Brazil
b Instituto de Ciência e Tecnologia, Universidade Federal de São Paulo, 12231-280,São José dos Campos, SP, Brazil
Introduction
The homo-dipeptide L,L-diphenylalanine (FF) [1] is consideredone of the simplest peptides able to generate ordered arrays insolution [2–5]. FF typically forms highly anisotropic structuresfollowing a multi-level ordering into fibrils and tubes whosedimensions range from a few tens of nanometers up to themicrometer scale. On the one hand, for self-assembly occurringin an aqueous environment, the crystallographic arrangementhas been appropriately described according to the hexagonalsymmetry group P61. In this configuration, FF units form cyclicheaders stack into a columnar phase parallel to the long axis ofthe structure with carboxyl and amine groups forming hydro-philic tunnels able to host H2O molecules and other hydrophilicspecies [6–8]. It has been shown that the water content withinthese galleries plays a major role in determining stability,morphology [9] and electronic properties [10]. As an example ofthis water-dependent behavior, two of us recently reported onthe splitting of the conduction and valence bands in FFnanotubes, likely due to the alignment of individual dipolemoments leading to a larger dipole moment of the overallstructure [10]. Moreover, when FF-based assemblies are producedunder anhydrous conditions, they are frequently described interms of orthorhombic groups [11–13]. Similarly, hydrated FFnanotubes exhibit a hexagonal→ orthorhombic phase transition
554–562
when incubated at temperatures above 150 °C. This phenomenonhas been ascribed to the release of water and the change inpeptide backbones from a linear conformation to a cyclic one(2,5-dioxopiperazine), followed by a re-organization of the crystal-line structure [11]. The observed transition is irreversible andinduces strong modifications on FF-MNTs (L,L-diphenylalanine mi-cro/nanotubes) properties, ranging from the nanoscopic to themacroscopic level [14,15]. Clearly, this realignment at themolecular level affects the balance of interactions. Specifically, forFF-MNTs, it is well-known that hydrogen bonds are extensively
Copyright © 2014 European Peptide Society and John Wiley & Sons, Ltd.

PEPTIDE NANOSTRUCTURES FOR GENERATION OF REACTIVE OXYGEN SPECIES
formed between peptide–peptide and peptide–water moleculeswhereas aromatic interactions are abundant between aromatic sidechains, which can be directly altered by phase organization [16,17].In a recent study [18], we demonstrated that the generation of
reactive oxygen species (ROS) by hypericin is reinforced uponcomplexation with FF micro/nanotubes (FF-MNTs). Hypericin isa natural photosensitizing compound found in vegetables ofthe genus Hypericium, among which St. John’s Wort is the mostcommon [19,20]. Among the several applications of hypericin inbiomedicine, this fluorophore has lately been explored inphotodynamic therapy because of its high yield for the produc-tion of ROS upon illumination with visible light (550–600 nm)[21,22]. The ROS output has been observed to be improved by>50% in the presence of FF-MNTs relative to that of barehypericin, as indirectly measured from the absorbance profile of1,3-diphenylisobenzofuran (DPBF). In addition, the characteristicfluorescence lifetimes have been observed to be significantlyboosted, demonstrating an enhancement in the photophysicalproperties and stabilization of ROS in excited states uponassociation with the peptide assemblies [23]. These results havebeen obtained for FF-MNTs assembled in a hexagonal spacegroup (P61), with evidence of the presence of water moleculesin the interstices of the peptide matrix [6,7]. Unfortunately, thedetailed mechanisms leading to improvements in ROS activityin hypericin+FF-MNTs complexes remain unclear, but tentativehypotheses have ascribed the aforementioned observations to(i) an increase in the overall dipolar moment due to thealignment of individual water dipoles within the structure and(ii) the organization of hypericin molecules on the surface ofFF-MNTs, which would lead to a coupling of hypericin dipolesand an increase in the resulting dipolar moment [18,24].To provide new insights into ROS activity in hypericin/FF-MNTs
hybrids and further explore the role of water in this system, herein,we present results obtained from investigations on the behaviorof these systems for different crystalline arrangements in solution.In particular, we monitored and compared the production of ROSby hypercin bound to FF-MNTs in both hexagonal (water-rich) andorthorhombic (water-free) structures. We also compared thesedata with ROS production by hypericin in the presence of freeH2O molecules in DMSO+water solutions. From a theoretical per-spective, we provide new molecular dynamics (MD) simulationsthat illustrate the distinct behavior of the hypericin/peptideinteraction at the interface of FF-MNTs for the different crystallo-graphic symmetries. As a conclusion, we propose that althoughthe presence of water dipoles appears to be an importantparameter able to enhance the photophysical properties of ROS,the organization induced by peptide structures and the creationof a hydrophobic environment surrounding hypericin/peptideinterface is paramount to optimizing ROS generation.
555
Materials and Methods
Chemicals
The linear form of L,L-diphenylalanine and 1,1,1,3,3,3 hexafluor-2-propanol (HFIP) were purchased from Sigma-Aldrich (USA). 1,3-Diphenylisobenzofuran, dimethyl sulfoxide (DMSO), methyl alcoholand chloroform were purchased from Synth (Brazil). Hypericin wassynthesized as reported elsewhere [25,26]. Ultrapure water (R> 18mol l�1Ω cm�1) was obtained from a Milli-Q system. All reagentswere of analytical grade and were used without further processing.
J. Pept. Sci. 2014; 20: 554–562 Copyright © 2014 European Peptide Society a
Preparation of Hypericin/FF-MNTs
The preparation and functionalization of FF-MNTs with hypericinwere performed in two steps: first, we diluted 3mg of FF in 30μlof HFIP and added aliquots of a stock solution of hypericin inmethanol (1.3 × 10�3mol l�1) up to the desired concentration ofthe fluorophore. Eleven samples were prepared and vortexedfor several minutes; the corresponding concentrations ofhypericin in the final solutions, ranging from 0.3 to 5.7μmol l�1,are indicated in Figure S1. In the second step, 150μl of ultrapurewater was added to the solutions, and the solutions were againvortexed. After approximately 20min, the precipitation of the firstfunctionalized hypericin/FF-MNTs was observed. Samples werekept in the dark, at room temperature, and left to dry in air fora few days. We assume that all of the peptides were convertedto FF-MNTs and all hypericin was incorporated into the struc-tures. As discussed in the Introduction, and shown in whatfollows, the tubes obtained following this protocol have P61crystallographic symmetry. To obtain tubes with orthorhombiccrystal ordering, the hypericin/FF-MNTs described previouslywere incubated at 150 °C for 60min. The orthorhombic symmetrywas inspected by X-ray diffraction (see results in what follows) toconfirm the phase transition. Absorbance (500–700 nm) andfluorescence (550–700 nm) data revealed identical the spectro-scopic signature of hypericin before and after heating (FiguresS2 and S3).
Structural, Morphological and Physical Characterization
X-ray diffraction (XRD): dry powdered samples of hypericin/FF-MTNs were loaded into 0.3mm glass capillaries and mountedhorizontally in a rotating holder at the center of a 2θ goniometerat the D12A-XRD1 beamline of LNLS (Brazilian Synchrotron LightLaboratory, Campinas, Brazil). Samples containing the highesthypericin concentration were chosen for high-resolution XRDmeasurements. X-rays with a wavelength λ=1.24044(2) Å wereselected by a double-bounce Si(111) monochromator with watercooling for the first crystal, whereas the second crystal was bentfor sagittal focusing. Experiments were conducted at roomtemperature, and slits were adjusted to provide a beam size of0.2 × 2mm2 on the samples. The wavelength and the zero pointwere determined from a Rietveld refinement [27,28] of theSRM626a alumina standard. The diffracted beam was detectedusing a Mythen 1K strip detector (Dectris). The incoming beamwas also monitored by a scintillation counter to normalize thedecay of the primary beam. Scanning was performed over a 2θrange of 4–30°, in step sizes of 0.001° and 2× 106 counts per step.The software Topas-Academic v.5 (Brisbane, Australia) [29] wasused to refine the orthorhombic model of the hypericin/FF-MNTs.Microscopy assays: polarizing and fluorescence imaging wereperformed using a confocal laser scanning microscope (LeicaTCS SP2, Germany) at the Centre de Recherche Paul Pascal(Pessac, France). Excitation wavelengths were chosen near theabsorbance peaks of hypericin (Figure S2), provided by the 514and 543 nm lines of an argon laser. Fluorescence images wererecorded across the hypericin emission range, between 575 and700 nm (Figure S3). Samples were prepared by sandwiching dropsof the hypericin/FF-MNTs solutions between glass slides andcoverslips. SEM images were obtained using a FEI Quanta 650FEG microscope at LNNANO (Brazilian Nanotechnology NationalLab, Campinas, Brazil). Droplets from solutions of the complexeswere deposited onto copper plaques, dried overnight in
nd John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jpepsci
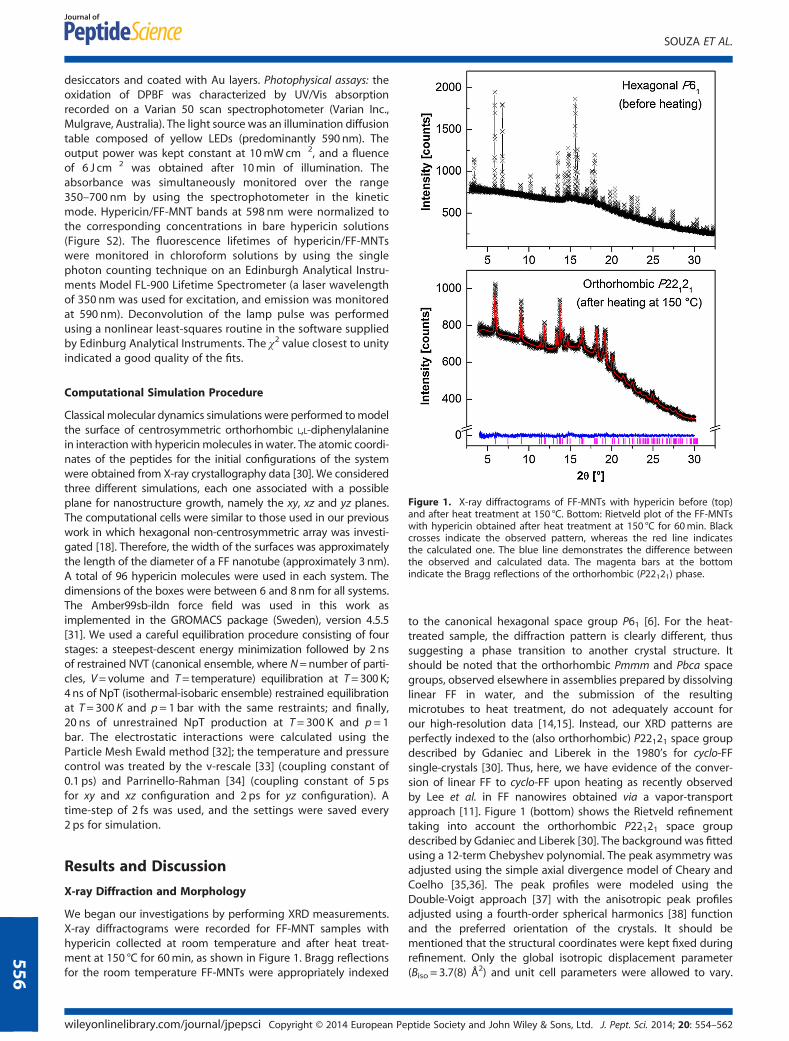
Figure 1. X-ray diffractograms of FF-MNTs with hypericin before (top)and after heat treatment at 150 °C. Bottom: Rietveld plot of the FF-MNTswith hypericin obtained after heat treatment at 150 °C for 60min. Blackcrosses indicate the observed pattern, whereas the red line indicatesthe calculated one. The blue line demonstrates the difference betweenthe observed and calculated data. The magenta bars at the bottomindicate the Bragg reflections of the orthorhombic (P22121) phase.
SOUZA ET AL.
556
desiccators and coated with Au layers. Photophysical assays: theoxidation of DPBF was characterized by UV/Vis absorptionrecorded on a Varian 50 scan spectrophotometer (Varian Inc.,Mulgrave, Australia). The light source was an illumination diffusiontable composed of yellow LEDs (predominantly 590 nm). Theoutput power was kept constant at 10mWcm�2, and a fluenceof 6 J cm�2 was obtained after 10min of illumination. Theabsorbance was simultaneously monitored over the range350–700 nm by using the spectrophotometer in the kineticmode. Hypericin/FF-MNT bands at 598 nm were normalized tothe corresponding concentrations in bare hypericin solutions(Figure S2). The fluorescence lifetimes of hypericin/FF-MNTswere monitored in chloroform solutions by using the singlephoton counting technique on an Edinburgh Analytical Instru-ments Model FL-900 Lifetime Spectrometer (a laser wavelengthof 350 nm was used for excitation, and emission was monitoredat 590 nm). Deconvolution of the lamp pulse was performedusing a nonlinear least-squares routine in the software suppliedby Edinburg Analytical Instruments. The χ2 value closest to unityindicated a good quality of the fits.
Computational Simulation Procedure
Classical molecular dynamics simulations were performed tomodelthe surface of centrosymmetric orthorhombic L,L-diphenylalaninein interaction with hypericin molecules in water. The atomic coordi-nates of the peptides for the initial configurations of the systemwere obtained from X-ray crystallography data [30]. We consideredthree different simulations, each one associated with a possibleplane for nanostructure growth, namely the xy, xz and yz planes.The computational cells were similar to those used in our previouswork in which hexagonal non-centrosymmetric array was investi-gated [18]. Therefore, the width of the surfaces was approximatelythe length of the diameter of a FF nanotube (approximately 3 nm).A total of 96 hypericin molecules were used in each system. Thedimensions of the boxes were between 6 and 8nm for all systems.The Amber99sb-ildn force field was used in this work asimplemented in the GROMACS package (Sweden), version 4.5.5[31]. We used a careful equilibration procedure consisting of fourstages: a steepest-descent energy minimization followed by 2nsof restrained NVT (canonical ensemble, where N=number of parti-cles, V=volume and T= temperature) equilibration at T=300K;4 ns of NpT (isothermal-isobaric ensemble) restrained equilibrationat T= 300 K and p= 1 bar with the same restraints; and finally,20 ns of unrestrained NpT production at T= 300 K and p = 1bar. The electrostatic interactions were calculated using theParticle Mesh Ewald method [32]; the temperature and pressurecontrol was treated by the v-rescale [33] (coupling constant of0.1 ps) and Parrinello-Rahman [34] (coupling constant of 5 psfor xy and xz configuration and 2 ps for yz configuration). Atime-step of 2 fs was used, and the settings were saved every2 ps for simulation.
Results and Discussion
X-ray Diffraction and Morphology
We began our investigations by performing XRD measurements.X-ray diffractograms were recorded for FF-MNT samples withhypericin collected at room temperature and after heat treat-ment at 150 °C for 60min, as shown in Figure 1. Bragg reflectionsfor the room temperature FF-MNTs were appropriately indexed
wileyonlinelibrary.com/journal/jpepsci Copyright © 2014 European Pe
to the canonical hexagonal space group P61 [6]. For the heat-treated sample, the diffraction pattern is clearly different, thussuggesting a phase transition to another crystal structure. Itshould be noted that the orthorhombic Pmmm and Pbca spacegroups, observed elsewhere in assemblies prepared by dissolvinglinear FF in water, and the submission of the resultingmicrotubes to heat treatment, do not adequately account forour high-resolution data [14,15]. Instead, our XRD patterns areperfectly indexed to the (also orthorhombic) P22121 space groupdescribed by Gdaniec and Liberek in the 1980’s for cyclo-FFsingle-crystals [30]. Thus, here, we have evidence of the conver-sion of linear FF to cyclo-FF upon heating as recently observedby Lee et al. in FF nanowires obtained via a vapor-transportapproach [11]. Figure 1 (bottom) shows the Rietveld refinementtaking into account the orthorhombic P22121 space groupdescribed by Gdaniec and Liberek [30]. The background was fittedusing a 12-term Chebyshev polynomial. The peak asymmetry wasadjusted using the simple axial divergence model of Cheary andCoelho [35,36]. The peak profiles were modeled using theDouble-Voigt approach [37] with the anisotropic peak profilesadjusted using a fourth-order spherical harmonics [38] functionand the preferred orientation of the crystals. It should bementioned that the structural coordinates were kept fixed duringrefinement. Only the global isotropic displacement parameter(Biso = 3.7(8) Å2) and unit cell parameters were allowed to vary.
ptide Society and John Wiley & Sons, Ltd. J. Pept. Sci. 2014; 20: 554–562
PEPTIDE NANOSTRUCTURES FOR GENERATION OF REACTIVE OXYGEN SPECIES
The hydrogen atoms were constrained to have Biso values 1.2times greater than the non-hydrogen ones. A good visual fit aswell as suitable statistical R-factors and goodness-of-fit indicatorwere obtained. The refined unit cell parameters and statistical fac-tors were as follows: a=6.185(1) Å, b=10.397(2) Å, c= 23.786(3) Å,V=1529.6(5) Å3, Rwp = 1.272% Rp = 1.023%, RBragg = 0.171% andχ2 = 0.866. The unit cells of hypericin/FF-MNT before and afterheat treatment are exhibited in Figure S4.We also characterized our hypericin/FF-MNTs hybrids by
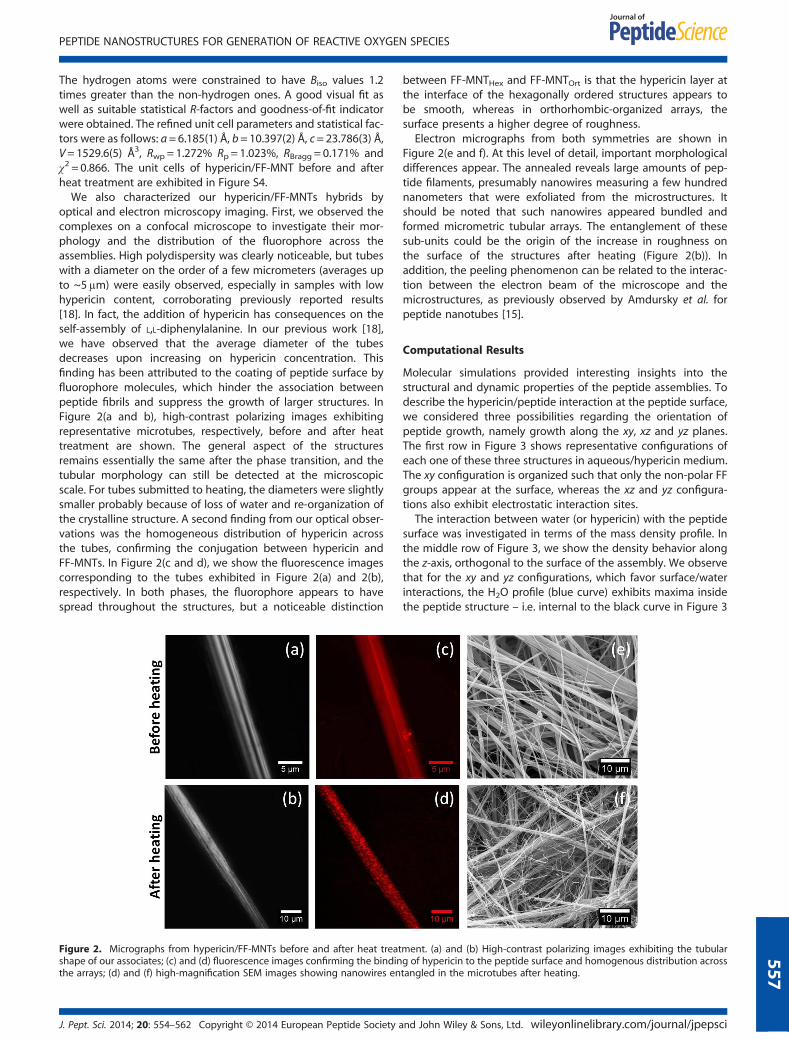
optical and electron microscopy imaging. First, we observed thecomplexes on a confocal microscope to investigate their mor-phology and the distribution of the fluorophore across theassemblies. High polydispersity was clearly noticeable, but tubeswith a diameter on the order of a few micrometers (averages upto ~5μm) were easily observed, especially in samples with lowhypericin content, corroborating previously reported results[18]. In fact, the addition of hypericin has consequences on theself-assembly of L,L-diphenylalanine. In our previous work [18],we have observed that the average diameter of the tubesdecreases upon increasing on hypericin concentration. Thisfinding has been attributed to the coating of peptide surface byfluorophore molecules, which hinder the association betweenpeptide fibrils and suppress the growth of larger structures. InFigure 2(a and b), high-contrast polarizing images exhibitingrepresentative microtubes, respectively, before and after heattreatment are shown. The general aspect of the structuresremains essentially the same after the phase transition, and thetubular morphology can still be detected at the microscopicscale. For tubes submitted to heating, the diameters were slightlysmaller probably because of loss of water and re-organization ofthe crystalline structure. A second finding from our optical obser-vations was the homogeneous distribution of hypericin acrossthe tubes, confirming the conjugation between hypericin andFF-MNTs. In Figure 2(c and d), we show the fluorescence imagescorresponding to the tubes exhibited in Figure 2(a) and 2(b),respectively. In both phases, the fluorophore appears to havespread throughout the structures, but a noticeable distinction
Figure 2. Micrographs from hypericin/FF-MNTs before and after heat treatshape of our associates; (c) and (d) fluorescence images confirming the bindinthe arrays; (d) and (f) high-magnification SEM images showing nanowires en
J. Pept. Sci. 2014; 20: 554–562 Copyright © 2014 European Peptide Society a
between FF-MNTHex and FF-MNTOrt is that the hypericin layer atthe interface of the hexagonally ordered structures appears tobe smooth, whereas in orthorhombic-organized arrays, thesurface presents a higher degree of roughness.
Electron micrographs from both symmetries are shown inFigure 2(e and f). At this level of detail, important morphologicaldifferences appear. The annealed reveals large amounts of pep-tide filaments, presumably nanowires measuring a few hundrednanometers that were exfoliated from the microstructures. Itshould be noted that such nanowires appeared bundled andformed micrometric tubular arrays. The entanglement of thesesub-units could be the origin of the increase in roughness onthe surface of the structures after heating (Figure 2(b)). Inaddition, the peeling phenomenon can be related to the interac-tion between the electron beam of the microscope and themicrostructures, as previously observed by Amdursky et al. forpeptide nanotubes [15].
Computational Results
Molecular simulations provided interesting insights into thestructural and dynamic properties of the peptide assemblies. Todescribe the hypericin/peptide interaction at the peptide surface,we considered three possibilities regarding the orientation ofpeptide growth, namely growth along the xy, xz and yz planes.The first row in Figure 3 shows representative configurations ofeach one of these three structures in aqueous/hypericin medium.The xy configuration is organized such that only the non-polar FFgroups appear at the surface, whereas the xz and yz configura-tions also exhibit electrostatic interaction sites.
The interaction between water (or hypericin) with the peptidesurface was investigated in terms of the mass density profile. Inthe middle row of Figure 3, we show the density behavior alongthe z-axis, orthogonal to the surface of the assembly. We observethat for the xy and yz configurations, which favor surface/waterinteractions, the H2O profile (blue curve) exhibits maxima insidethe peptide structure – i.e. internal to the black curve in Figure 3
ment. (a) and (b) High-contrast polarizing images exhibiting the tubularg of hypericin to the peptide surface and homogenous distribution acrosstangled in the microtubes after heating.
nd John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jpepsci
557
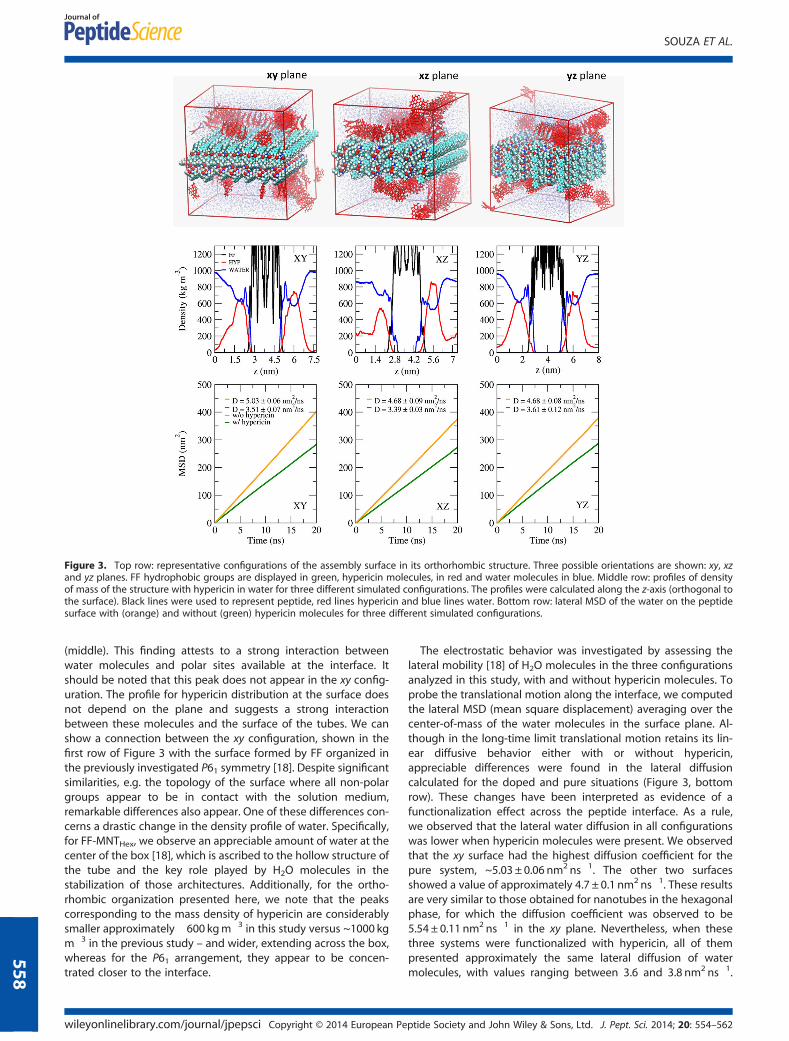
Figure 3. Top row: representative configurations of the assembly surface in its orthorhombic structure. Three possible orientations are shown: xy, xzand yz planes. FF hydrophobic groups are displayed in green, hypericin molecules, in red and water molecules in blue. Middle row: profiles of densityof mass of the structure with hypericin in water for three different simulated configurations. The profiles were calculated along the z-axis (orthogonal tothe surface). Black lines were used to represent peptide, red lines hypericin and blue lines water. Bottom row: lateral MSD of the water on the peptidesurface with (orange) and without (green) hypericin molecules for three different simulated configurations.
SOUZA ET AL.
558
(middle). This finding attests to a strong interaction betweenwater molecules and polar sites available at the interface. Itshould be noted that this peak does not appear in the xy config-uration. The profile for hypericin distribution at the surface doesnot depend on the plane and suggests a strong interactionbetween these molecules and the surface of the tubes. We canshow a connection between the xy configuration, shown in thefirst row of Figure 3 with the surface formed by FF organized inthe previously investigated P61 symmetry [18]. Despite significantsimilarities, e.g. the topology of the surface where all non-polargroups appear to be in contact with the solution medium,remarkable differences also appear. One of these differences con-cerns a drastic change in the density profile of water. Specifically,for FF-MNTHex, we observe an appreciable amount of water at thecenter of the box [18], which is ascribed to the hollow structure ofthe tube and the key role played by H2O molecules in thestabilization of those architectures. Additionally, for the ortho-rhombic organization presented here, we note that the peakscorresponding to the mass density of hypericin are considerablysmaller approximately �600 kgm�3 in this study versus ~1000 kgm�3 in the previous study – and wider, extending across the box,whereas for the P61 arrangement, they appear to be concen-trated closer to the interface.
wileyonlinelibrary.com/journal/jpepsci Copyright © 2014 European Pe
The electrostatic behavior was investigated by assessing thelateral mobility [18] of H2O molecules in the three configurationsanalyzed in this study, with and without hypericin molecules. Toprobe the translational motion along the interface, we computedthe lateral MSD (mean square displacement) averaging over thecenter-of-mass of the water molecules in the surface plane. Al-though in the long-time limit translational motion retains its lin-ear diffusive behavior either with or without hypericin,appreciable differences were found in the lateral diffusioncalculated for the doped and pure situations (Figure 3, bottomrow). These changes have been interpreted as evidence of afunctionalization effect across the peptide interface. As a rule,we observed that the lateral water diffusion in all configurationswas lower when hypericin molecules were present. We observedthat the xy surface had the highest diffusion coefficient for thepure system, ~5.03 ± 0.06 nm2 ns�1. The other two surfacesshowed a value of approximately 4.7 ± 0.1 nm2 ns�1. These resultsare very similar to those obtained for nanotubes in the hexagonalphase, for which the diffusion coefficient was observed to be5.54 ± 0.11 nm2 ns�1 in the xy plane. Nevertheless, when thesethree systems were functionalized with hypericin, all of thempresented approximately the same lateral diffusion of watermolecules, with values ranging between 3.6 and 3.8 nm2 ns�1.
ptide Society and John Wiley & Sons, Ltd. J. Pept. Sci. 2014; 20: 554–562
PEPTIDE NANOSTRUCTURES FOR GENERATION OF REACTIVE OXYGEN SPECIES
These values are substantially lower than the value observed forthe P61 symmetry, 4.33 ± 0.04 nm2 ns�1 [18]. These lower valuescan be ascribed to a greater interaction between water moleculesand hypericin at the interface for the orthorhombic configura-tion, decreasing the movement of the fluorophore parallel tothe peptide surface.
Photophysical Characterization
Upon excitation with visible light, hypericin is able to transferenergy to oxygen and give rise to ROS, including 1O2 [22,39]. To eval-uate the photophysical properties of ROS in our hypericin-containingcomplexes, we used an indirect method based on the oxygen-accepting properties of DPBF, which produces o-dibenzoylbenzenein the presence of 1O2. Therefore, by monitoring the disappearanceof DPBF, we were able to follow the kinetics of ROS production[40]. We focused on three types of samples dispersed in DMSO: barehypericin, hypericin/FF-MNThex and hypericin/FF-MNTort. In addition,we also evaluated bare hypericin in DMSO+water solutions toprovide a framework for comparison between the scenario in whichH2Omolecules are ‘free and disordered’ in solutionwith one inwhichthey appear ‘trapped and ordered’ within P61 structures.According to the procedures detailed in the Materials and
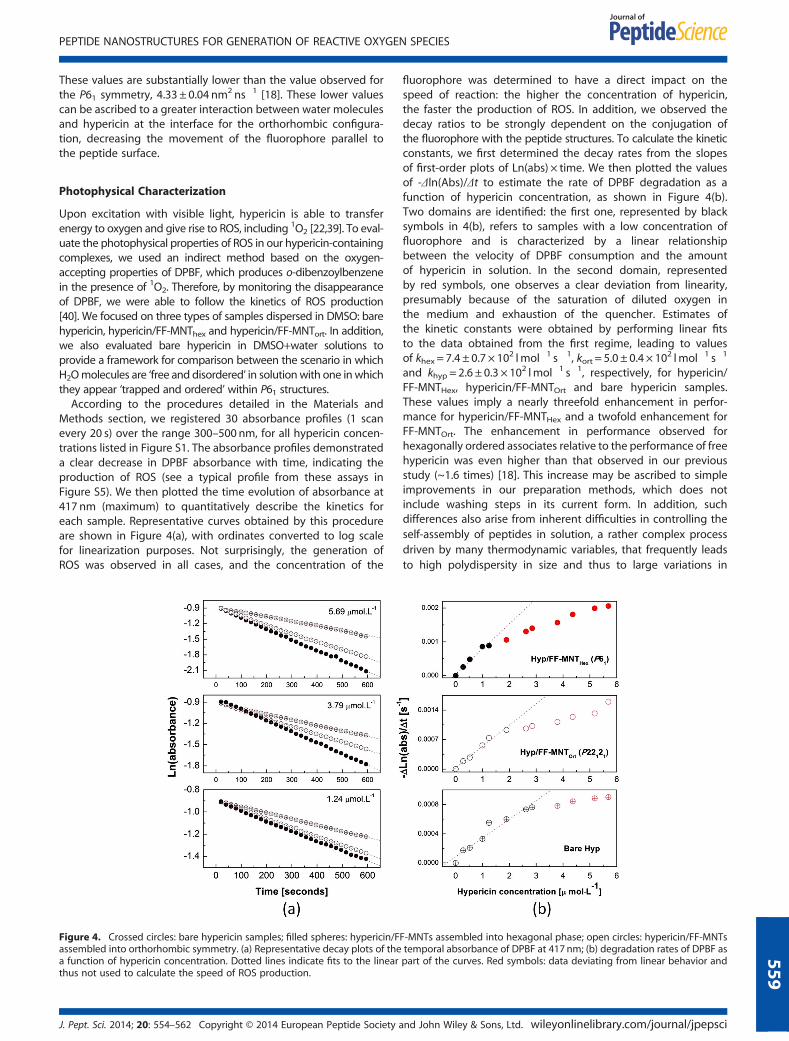
Methods section, we registered 30 absorbance profiles (1 scanevery 20 s) over the range 300–500 nm, for all hypericin concen-trations listed in Figure S1. The absorbance profiles demonstrateda clear decrease in DPBF absorbance with time, indicating theproduction of ROS (see a typical profile from these assays inFigure S5). We then plotted the time evolution of absorbance at417 nm (maximum) to quantitatively describe the kinetics foreach sample. Representative curves obtained by this procedureare shown in Figure 4(a), with ordinates converted to log scalefor linearization purposes. Not surprisingly, the generation ofROS was observed in all cases, and the concentration of the
Figure 4. Crossed circles: bare hypericin samples; filled spheres: hypericin/Fassembled into orthorhombic symmetry. (a) Representative decay plots of thea function of hypericin concentration. Dotted lines indicate fits to the linearthus not used to calculate the speed of ROS production.
J. Pept. Sci. 2014; 20: 554–562 Copyright © 2014 European Peptide Society a
fluorophore was determined to have a direct impact on thespeed of reaction: the higher the concentration of hypericin,the faster the production of ROS. In addition, we observed thedecay ratios to be strongly dependent on the conjugation ofthe fluorophore with the peptide structures. To calculate the kineticconstants, we first determined the decay rates from the slopesof first-order plots of Ln(abs) × time. We then plotted the valuesof -Δln(Abs)/Δt to estimate the rate of DPBF degradation as afunction of hypericin concentration, as shown in Figure 4(b).Two domains are identified: the first one, represented by blacksymbols in 4(b), refers to samples with a low concentration offluorophore and is characterized by a linear relationshipbetween the velocity of DPBF consumption and the amountof hypericin in solution. In the second domain, representedby red symbols, one observes a clear deviation from linearity,presumably because of the saturation of diluted oxygen inthe medium and exhaustion of the quencher. Estimates ofthe kinetic constants were obtained by performing linear fitsto the data obtained from the first regime, leading to valuesof khex = 7.4 ± 0.7 × 102 lmol�1 s�1, kort = 5.0 ± 0.4 × 102 lmol�1 s�1
and khyp = 2.6 ± 0.3 × 102 lmol�1 s�1, respectively, for hypericin/FF-MNTHex, hypericin/FF-MNTOrt and bare hypericin samples.These values imply a nearly threefold enhancement in perfor-mance for hypericin/FF-MNTHex and a twofold enhancement forFF-MNTOrt. The enhancement in performance observed forhexagonally ordered associates relative to the performance of freehypericin was even higher than that observed in our previousstudy (~1.6 times) [18]. This increase may be ascribed to simpleimprovements in our preparation methods, which does notinclude washing steps in its current form. In addition, suchdifferences also arise from inherent difficulties in controlling theself-assembly of peptides in solution, a rather complex processdriven by many thermodynamic variables, that frequently leadsto high polydispersity in size and thus to large variations in
F-MNTs assembled into hexagonal phase; open circles: hypericin/FF-MNTstemporal absorbance of DPBF at 417 nm; (b) degradation rates of DPBF aspart of the curves. Red symbols: data deviating from linear behavior and
nd John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jpepsci
559
SOUZA ET AL.
560
interfacial area between hypericin and FF-MNTs [3]. Similarly, ki-netics are also strongly dependent onmultiple quantum and ther-modynamic phenomena cooperating in a complex frameworkand small changes in factors such as incubation time, oxidationdegree and environmental conditions often result distinct behav-iors [41]. Using identical protocols through which absorbance pro-files were followed under normalized conditions, we systemicallyobserved a substantial increase in the estimated kinetic constantsfor hypericin/FF-MNTs when compared with those of freehypericin in solution.
It should be noted that the enhancement in ROS output occurseven for micro/nanotubes exhibiting P22121 symmetry. In otherwords, a remarkable improvement in ROS generation is foundeven in the absence of water within the particles’ structure.Nevertheless, it should be noted that the increase in kineticconstants was higher for hypericin associated with FF-MNTsorganized into the hexagonal phase, likely because of the spatialorganization of hypericin on the hydrophobic surface of theassemblies and the facilitation of charge transfer [10]. The strongnon-polar nature of the interface allows for the creation of apacking pattern of hypericin on the borders of the tubes, wherethe fluorophore appears to spread across the surface [18]. Onthe other hand, assemblies organized into orthorhombic symme-try have more hydrophilic groups available at the interface (seediscussions on MD results previously) and presumably have ahigher propensity for hypericin aggregation, thus losing thecapacity to generate ROS compared with FF-MNTs in thehexagonal phase.
Fluorescence lifetimes in chloroform were also measured forsamples containing hypericin at 2.9μmol l�1, and the respectiveresults are shown in Table 1 together with the correspondingχ2red parameters. The proximity of this parameter to unity is astrong indicator of the goodness of fits performed to the fluores-cence decay data and attests reliability of the measured lifetimes.Similar to the behavior observed for the kinetic constants, thelifetimes increase appreciably when the fluorophore appears tobe conjugated to peptides, with respective improvements of~35% and ~20% for FF-MNTHex and FF-MNTOrt. These resultsdemonstrate that the peptide assemblies stabilize the excitedstate of the photosensitizer and indicate an enhancement inphotophysical properties because longer fluorescence lifetimesimply that more energy is available to be transferred to 3O2 andpotentially boost ROS generation [22].
Overall, the results presented previously indicate that the bestperformances are always obtained for structures organized intothe hexagonal phase. On the one hand, it has been reported thatthe photophysical behavior of hypericin can be strongly affectedby guest H2O molecules in DMSO, and fluorescence lifetimeshave been observed to decrease with an increase in the volumepercent of water [42]. On the other hand, the reaction rate
Table 1. Fluorescence lifetimes in chloroform for hypericin andhypericin/FF-MNTs complexes (hypericin concentration at 2.9μmol l�1)
Sample τ ±σ (ns) χ2red
Bare hypericin 3.904 ± 0.002 0.949
Hypericin/FF-MNTOrt 4.663 ± 0.003 1.378
Hypericin/FF-MNTHex 5.274 ± 0.003 1.367
The proximity of χ2red parameter to unity attests reliability ofmeasurements.
wileyonlinelibrary.com/journal/jpepsci Copyright © 2014 European Pe
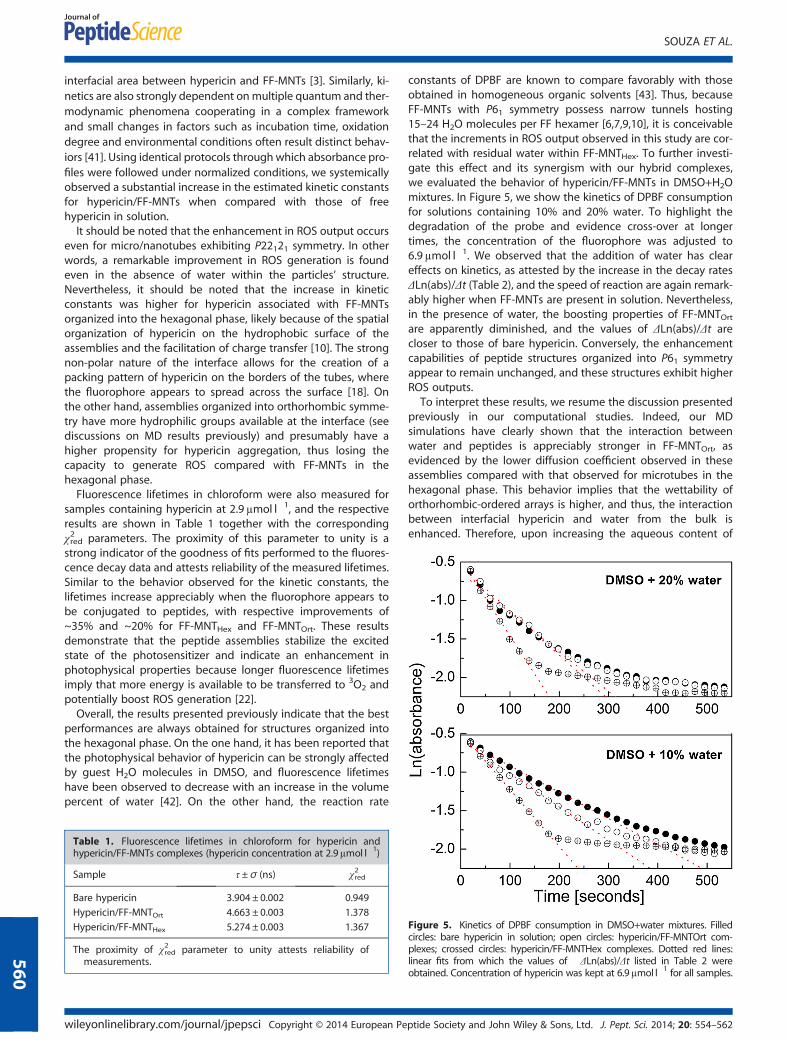
constants of DPBF are known to compare favorably with thoseobtained in homogeneous organic solvents [43]. Thus, becauseFF-MNTs with P61 symmetry possess narrow tunnels hosting15–24 H2O molecules per FF hexamer [6,7,9,10], it is conceivablethat the increments in ROS output observed in this study are cor-related with residual water within FF-MNTHex. To further investi-gate this effect and its synergism with our hybrid complexes,we evaluated the behavior of hypericin/FF-MNTs in DMSO+H2Omixtures. In Figure 5, we show the kinetics of DPBF consumptionfor solutions containing 10% and 20% water. To highlight thedegradation of the probe and evidence cross-over at longertimes, the concentration of the fluorophore was adjusted to6.9μmol l�1. We observed that the addition of water has cleareffects on kinetics, as attested by the increase in the decay ratesΔLn(abs)/Δt (Table 2), and the speed of reaction are again remark-ably higher when FF-MNTs are present in solution. Nevertheless,in the presence of water, the boosting properties of FF-MNTOrtare apparently diminished, and the values of ΔLn(abs)/Δt arecloser to those of bare hypericin. Conversely, the enhancementcapabilities of peptide structures organized into P61 symmetryappear to remain unchanged, and these structures exhibit higherROS outputs.To interpret these results, we resume the discussion presented
previously in our computational studies. Indeed, our MDsimulations have clearly shown that the interaction betweenwater and peptides is appreciably stronger in FF-MNTOrt, asevidenced by the lower diffusion coefficient observed in theseassemblies compared with that observed for microtubes in thehexagonal phase. This behavior implies that the wettability oforthorhombic-ordered arrays is higher, and thus, the interactionbetween interfacial hypericin and water from the bulk isenhanced. Therefore, upon increasing the aqueous content of
Figure 5. Kinetics of DPBF consumption in DMSO+water mixtures. Filledcircles: bare hypericin in solution; open circles: hypericin/FF-MNTOrt com-plexes; crossed circles: hypericin/FF-MNTHex complexes. Dotted red lines:linear fits from which the values of �ΔLn(abs)/Δt listed in Table 2 wereobtained. Concentration of hypericin was kept at 6.9μmol l�1 for all samples.
ptide Society and John Wiley & Sons, Ltd. J. Pept. Sci. 2014; 20: 554–562

Table 2. Decay rates, ΔLn(abs)/Δt, of DBPF consumption in DMSO+water mixtures (hypericin concentration at 6.9μmol l�1).
�ΔLn(abs)/Δt [s�1]
Water 10% Water 20%
Bare hypericin 0.0034± 0.0001 0.0051± 0.0003
Hypericin/FF-MNTOrt 0.0043± 0.0001 0.0058± 0.0003
Hypericin/FF-MNTHex 0.0074± 0.0001 0.0099± 0.0004
PEPTIDE NANOSTRUCTURES FOR GENERATION OF REACTIVE OXYGEN SPECIES
the medium, H2O molecules permeate the hypericin layer, andthe photophysical properties of the complexes approach thoseobserved for free hypericin in bulk solution. On the other hand,lower wettability in hexagonally ordered structures likely favorsthe appearance of a confined hydrophobic environment sur-rounding assemblies in which non-polar species – namely,hypericin, 1O2 and DPBF – are brought together to create ananoreactor, in line with observations previously made forhypericin/liposome complexes [39,44]. In addition, the formationof such a hydrophobic layer close to the interface potentiallyleads to tighter packing of water in proximity to hypericin onthe surface and likely to the alignment of H2O dipoles in thehydration shell surrounding the associates. A similar packingeffect could also occur with the guest H2O molecules hosted inthe hydrophilic core of FF hexamers, giving rise to the best per-formance of ROS production observed for hypericin/FF-MNTHexunder anhydrous conditions.The role of unbound hypericin in solution also merits to be
discussed in our interpretation of the previously described results.In principle, interactions between fluorophore and peptides arenot specific, and the generation of ROS in our samples presumablyarises from either bound or unbound hypericin. Thus, a fraction offluorophore molecules would be at dynamic equilibrium, driven byassociation and dissociation constants at interface. In this case again,the strong hydrophobic behavior found in hexagonal assembliesplays a central role because the higher the hydrophobicity, thehigher the residence time of the fluorophore (and other non-polarspecies) in the complex. Thus, the presence of a highly hydrophobicmicro-environment surrounding peptide interfaces also hasconsequences on the dynamic equilibrium of the system.
561
Conclusions
We investigated the behavior of complexes formed betweenhypericin and FF-MNTs either in a water-containing hexagonalphase or in a water-free phase exhibiting orthorhombic symme-try. Significant improvements in the lifetimes of excited statesand higher decay rates of DPBF consumption were observed inboth the hexagonal and orthorhombic arrangements. These find-ings indicate a higher efficiency of ROS generation in these com-plexes, even in the absence of residual water in the peptidematrix. Thus, our results demonstrate that spatial arrangementplays a paramount role in determining photophysical behavior,where the abundance of hydrophobic sites at the hypericin/peptide interface favors the creation of a nanoreactor in whichnon-polar species such hypericin, DPBF and singlet oxygen arebrought together in a confined geometry. In addition, the self-assembled structures can induce template-assisted ordering ofhypericin at the interface and lead to potential coupling betweenhypericin dipolar moments. Moreover, the best performanceswere systematically observed in hypericin/FF-MNTHex hosting
J. Pept. Sci. 2014; 20: 554–562 Copyright © 2014 European Peptide Society a
highly packed H2O molecules within hydrophilic narrow tunnelsin the peptide matrix. Therefore, we speculate that a synergisticcooperation between organized and aligned water dipoles couldbe give rise to the enhancement of photophysical properties.Molecular dynamics simulations indicated stronger interactionsbetween water and associates assembled in the orthorhombicphase, allowing for the permeation of the hypericin layer byH2O molecules. For this reason, the enhancement capabilities ofFF-MNTOrt are strongly diminished in an aqueous environment,as observed in our DMSO+water mixtures. In addition, thepresence of hydrophilic sites on the peptides’ surface is anobstacle to the creation of a hydrophobic shell able to confinenon-polar species, explaining why the worst performance wasobserved for the highly hydrophobic structures exhibitinghexagonal symmetry. These findings provide useful fundamentalinsights for designing peptide/fluorophore complexes with out-standing technological potential.
Acknowledgements
Prof. Anderson O. Ribeiro and MSc Gislaine P. de Andrade arekindly acknowledged for providing the hypericin compoundsused in this work. Financial support was provided by FAPESP(grants no. 2013/12997-0, 2013/17193-6, 2008/53576-9 and2008/10537-3) and INCT in Bioanalytics (FAPESP, grant no. 08/57805-2 and CNPq, grant no. 573672/2008-3) and CNPq (grantno. 472197/2012-6). M. I. S. is grateful to CAPES for a PhD fellow-ship. E. R. S. acknowledges FAPESP (proc. no. 2013/12674-6) andPNPD-CAPES (proc. no. 23038007044201108) for post-doctoralgrants. The LNNano and LNLS staffs are kindly recognized forinvaluable help and access to SEM and synchrotron facilities(proposals SEM-LV-14754 and XRD1 – 14395). Dr. LaurenceNavailles deserves very special thanks for receiving M. I. S. and E.R. S. at CRPP (Pessac, France) during the confocal experiments.
References1 Jorgensen WL, Chandrasekhar J, Madura JD, Impey RW, Klein ML.
Comparison of simple potential functions for simulating liquid water.J. Chem. Phys. 1983; 79: 926–935.
2 Reches M, Gazit E. Casting metal nanowires within discrete self-assembled peptide nanotubes. Science 2003; 300: 625–627.
3 Reches M, Gazit E. Molecular self-assembly of peptide nanostructures:mechanism of association and potential uses. Curr. Nanosci. 2006; 2:105–111.
4 Demirel G, Malvadkar N, Demirel MC. Control of protein adsorptiononto core-shell tubular and vesicular structures of diphenylalanine/parylene. Langmuir 2010; 26: 1460–1463.
5 Maude S, Tai LR, Davies RP, Liu B, Harris SA, Kocienski PJ, Aggeli A. Pep-tide synthesis and self-assembly. Top. Curr. Chem. 2012; 310: 27–69.
6 Görbitz CH. Nanotube formation by hydrophobic dipeptides. Chem.Eur. J. 2001; 7: 5153–5159.
7 Gorbitz CH. The structure of nanotubes formed by diphenylalanine,the core recognition motif of Alzheimer’s [small beta]-amyloid poly-peptide. Chem. Commun. 2006; 2332–2334.
8 Silva RF, Araújo DR, Silva ER, Ando RA, Alves WA. L-diphenylalaninemicrotubes as a potential drug-delivery system: characterization,release kinetics, and cytotoxicity. Langmuir 2013; 29: 10205–10212.
9 Kim J, Han TH, Kim Y-I, Park JS, Choi J, Churchill DG, Kim SO, Ihee H.Role of water in directing diphenylalanine assembly into nanotubesand nanowires. Adv. Mater. 2010; 22: 583–587.
10 Andrade-Filho T, Ferreira FF, Alves WA, Rocha AR. The effects of watermolecules on the electronic and structural properties of peptidenanotubes. Phys. Chem. Chem. Phys. 2013; 15: 7555–7559.
11 Lee JS, Yoon I, Kim J, Ihee H, Kim B, Park CB. Self-assembly of semicon-ducting photoluminescent peptide nanowires in the vapor phase.Angew. Chem. Int. Ed. 2011; 50: 1164–1167.
nd John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jpepsci

SOUZA ET AL.
562
12 Mason TO, Chirgadze DY, Levin A, Adler-Abramovich L, Gazit E,Knowles TPJ, Buell AK. Expanding the solvent chemical space forself-assembly of dipeptide nanostructures. ACS Nano 2014; 8: 1243–1253.
13 Amaral HR, Kogikoski S, Jr, Silva ER, Souza JA, Alves WA. Micro- andnano-sized peptidic assemblies prepared via solid-vapor approach:morphological and spectroscopic aspects. Mater. Chem. Phys. 2012;137: 628–636.
14 Heredia A, Bdikin I, Kopyl S, Mishina E, Semin S, Sigov A, German K,Bystrov V, Gracio J, Kholkin AL. Temperature-driven phase transforma-tion in self-assembled diphenylalanine peptide nanotubes. J. Phys. DAppl. Phys. 2010; 43: 462001.
15 Amdursky N, Beker P, Koren I, Bank-Srour B, Mishina E, Semin S,Rasing T, Rosenberg Y, Barkay Z, Gazit E, Rosenman G. Structuraltransition in peptide nanotubes. Biomacromolecules 2011; 12:1349–1354.
16 Sunde M, Serpell LC, Bartlam M, Fraser PE, Pepys MB, Blake CCF. Com-mon core structure of amyloid fibrils by synchrotron X-ray diffraction.J. Mol. Biol. 1997; 273: 729–739.
17 Blake C, Serpell L. Synchrotron X-ray studies suggest that the core ofthe transthyretin amyloid fibril is a continuous β-sheet helix. Structure1996; 4: 989–998.
18 Souza MI, Jaques YM, de Andrade GP, Ribeiro AO, da Silva ER, Fileti EE,Ávilla ÉS, Pinheiro MVB, Krambrock K, Alves WA. Structural andphotophysical properties of peptide micro/nanotubes functionalizedwith hypericin. J. Phys. Chem. B 2013; 117: 2605–2614.
19 Karioti A Bilia AR Hypericins as potential leads for new therapeutics.Int. J. Mol. Sci. 2010; 11: 562–594.
20 Gbur P, Dedic R, Chorvat D, Miskovsky P, Hala J, Jancura D. Time-resolved luminescence and singlet oxygen formation after illumina-tion of the hypericin-low-density lipoprotein complex. Photochem.Photobiol. 2009; 85: 816–823.
21 SanchezCortes S, Miskovsky P, Jancura D, Bertoluzza A. Specific inter-actions of antiretroviraly active drug hypericin with DNA as studied bysurface-enhanced resonance Raman spectroscopy. J. Phys. Chem.1996; 100: 1938–1944.
22 Guedes RC, Eriksson LA. Theoretical study of hypericin. J. Photochem.Photobiol. A 2005; 172: 293–299.
23 Martins TD, de Souza MI, Cunha BB, Takahashi PM, Ferreira FF, SouzaJA, Fileti EE, Alves WA. Influence of pH and pyrenyl on the structuraland morphological control of peptide nanotubes. J. Phys. Chem. C2011; 115: 7906–7913.
24 Zhang HY, Chen DZ. Mechanism for intramolecular proton transferinvolving hypericin. Dyes Pigm. 2000; 46: 17–21.
25 Falk H, Meyer J, Oberreiter M. A convenient semisynthetic route tohypericin. Monatsh. Chem. 1993; 124: 339–341.
26 Huygens A, Kamuhabwa AR, de Witte PAM. Stability of different for-mulations and ion pairs of hypericin. Eur. J. Pharm. Biopharm. 2005;59: 461–468.
27 Rietveld HM. Line profiles of neutron powder-diffraction peaks forstructure refinement. Acta Crystallogr. 1967; 22: 151–152.
28 Rietveld HM A profile refinement method for nuclear and magneticstructures. J. Appl. Crystallogr. 1969; 2: 65–71.
wileyonlinelibrary.com/journal/jpepsci Copyright © 2014 European Pe
29 Coelho AA, Evans J, Evans I, Kern A, Parsons S. The TOPAS symboliccomputation system. Powder Diffr. 2011; 26: s22–s25.
30 Gdaniec M, Liberek B. Structure of cyclo(-l-phenylalanyl-l-phenylalanyl-).Acta Crystallogr. Sect. C Cryst. Struct. Commun. 1986; 42: 1343–1345.
31 Hess B, Kutzner C, van der Spoel D, Lindahl E. GROMACS 4: algorithmsfor highly efficient, load-balanced, and scalable molecular simulation.J. Chem. Theory Comput. 2008; 4: 435–447.
32 Darden T, York D, Pedersen L. Particle mesh Ewald: an N�log(N)methodfor Ewald sums in large systems. J. Chem. Phys. 1993; 98: 10089–10092.
33 Bussi G, Donadio D, Parrinello M. Canonical sampling through velocityrescaling. J. Chem. Phys. 2007; 126: 014101.
34 Martoňák R, Laio A, Parrinello M. Predicting crystal structures: theParrinello-Rahman method revisited. Phys. Rev. Lett. 2003; 90: 075503.
35 Cheary RW, Coelho AA. Axial divergence in a conventional X-raypowder diffractometer. II. Realization and evaluation in aFundamental-Parameter profile fitting procedure. J. Appl.Crystallogr. 1998; 31: 862–868.
36 Cheary RW, Coelho AA. Axial divergence in a conventional X-raypowder diffractometer. I. Theoretical foundations. J. Appl. Crystallogr.1998; 31: 851–861.
37 Balzar D X-Ray-diffraction line broadening – modeling and applica-tions to high-Tc superconductors. J. Res. Natl. Inst. Stand. Technol.1993; 98: 321–353.
38 Jarvinen M. Application of symmetrized harmonics expansion to cor-rection of the preferred orientation effect. J. Appl. Crystallogr. 1993;26: 525–531.
39 Hadjur C, Jeunet A, Jardon P. Photosensitization by hypericin-electron-spin-resonance (esr) evidence for the formation of singletoxygen and superoxide anion-radicals in an in-vitro model.J. Photochem. Photobiol. B 1994; 26: 67–74.
40 AmatGuerri F, Lempe E, Lissi EA, Rodriguez FJ, Trull FR. Water-soluble1,3-diphenylisobenzofuran derivatives. Synthesis and evaluation assinglet molecular oxygen acceptors for biological systems.J. Photochem. Photobiol. A 1996; 93: 49–56.
41 Lakowicz JR. Principles of Fluorescence Spectroscopy, Springer: NewYork,2006.
42 Falk H, Meyer J. On the homo-association and heteroassociation ofhypericin. Monatsh. Chem. 1994; 125: 753–762.
43 Lindig BA, Rodgers MAJ, Schaap AP. Determination of the lifetime ofsinglet oxygen in D2O using 9,10-anthracenedipropionic acid, awater-soluble probe. J. Am. Chem. Soc. 1980; 102: 5590–5593.
44 Bouirig H, Eloy D, Jardon P. Generation and reactions of singlet oxy-gen photosensitized by hypericin in dipalmitoylphosphatidylcholinevesicles – experimental evidence of a delayed photooxidation.J. Chim. Phys. Phys. Chim. Biol. 1992; 89: 1391–1411.
Supporting Information
Additional supporting information may be found in the onlineversion of this article at the publisher's web site.
ptide Society and John Wiley & Sons, Ltd. J. Pept. Sci. 2014; 20: 554–562
Top Related
Copyright © 2022 FDOKUMEN

