







Bahasa
Halaman
Hukum


ABSTRACT
There is limited information on the impacts of present-daysolar ultraviolet-B radiation (UV-B) on biomass and grainyield of field crops and on the mechanisms that confer tol-erance to UV-B radiation under field conditions. We inves-tigated the effects of solar UV-B on aspects of thebiochemistry, growth and yield of barley crops using repli-cated field plots and two barley strains, a catalase (CAT)-deficient mutant (RPr 79/4) and its wild-type mother line(Maris Mink). Solar UV-B reduced biomass accumulationand grain yield in both strains. The effects on cropbiomass accumulation tended to be more severe in RPr79/4 (≈ 32% reduction) than in the mother line (≈ 20%reduction). Solar UV-B caused measurable DNA damagein leaf tissue, in spite of inducing a significant increase inUV-absorbing sunscreens in the two lines. Maris Minkresponded to solar UV-B with increased CAT and ascor-bate peroxidase (APx) activity. No effects of UV-B on totalsuperoxide dismutase (SOD) activity were detected.Compared with the wild type, RPr 79/4 had lower CATactivity, as expected, but higher APx activity. Neither ofthese activities increased in response to UV-B in RPr 79/4.These results suggest that growth inhibition by solar UV-Binvolves DNA damage and oxidative stress, and that con-stitutive and UV-B-induced antioxidant capacity may playan important role in UV-B tolerance.
Key-words:antioxidants; barley; catalase; DNA damage; UV-B.
INTRODUCTION
The effects of ultraviolet-B (UV-B) radiation on terrestrialplants have been studied in considerable detail over the lasttwo decades. Experiments carried out by physiologicalecologists have produced information on the morphologi-cal and growth responses to UV-B of several agronomi-cally important species under realistic field conditions
[references in Caldwell et al. (1995) and Rozema et al.(1997)]. In parallel with these efforts, laboratory studieshave addressed the effects of UV-B on damage to cellularcomponents (e.g. in the form of lipid peroxidation, proteindegradation, and DNA damage), and investigated adaptivebiochemical responses to UV-B, such as the accumulationof UV-absorbing pigments, the increase in antioxidantenzymes, and the effects of UV on photoenzymatic DNArepair [see reviews by Bornman & Teramura (1993) andStrid, Chow & Anderson (1994)].
Several generalizations have emerged from these stud-ies. From field experiments it has been established thatambient levels of solar UV-B received at low (Searles,Caldwell & Winter 1995), intermediate (Ballaré et al.1996), and high latitudes (Rousseaux et al. 1998) caninhibit stem elongation and cause reductions in leaf areaexpansion in various plant species. Photosynthesis per unitleaf area was found to be rather insensitive to solar UV-B(or to realistic supplementation treatments) under fieldconditions (Beyschlag et al. 1988; Searles et al. 1995).Nonetheless, plant growth (biomass accumulation) can beinhibited by ambient UV-B as a result of UV-B-inducedreductions in leaf area expansion (Ballaré et al. 1996).Growth inhibition appears to be transient in some systems,and limited to the early phases of development (Rozemaet al. 1997). However, the published information ongrowth responses of mature plants and on the impacts ofsolar UV-B on crop yield is still very limited. In the reportsavailable at present, morphological responses have beenobserved frequently, but conflicting data have been pub-lished regarding the effects of solar UV-B on crop yield[references in Bornman & Teramura (1993)].
Laboratory and greenhouse studies have shed somelight on the mechanisms of UV-B-induced growth inhibi-tion and UV-B protection. Evidence has been obtained forthe idea that some of the morphological responses to UV-B (e.g. inhibition of hypocotyl elongation) are mediatedby specific photoreceptors (Ensminger 1993; Ballaré,Barnes & Flint 1995a), and may play adaptive rolesreducing UV-B exposure (Ballaré et al. 1995b). The accu-mulation of phenylpropanoid derivatives is a common
Plant, Cell and Environment (1999) 22, 61–70
© 1999 Blackwell Science Ltd
The effects of solar ultraviolet-B radiation on the growth and
yield of barley are accompanied by increased DNA damage and
antioxidant responses
C. A. MAZZA,1 D. BATTISTA,1 A. M. ZIMA, 1 M. SZWARCBERG-BRACCHITTA,1 C. V. GIORDANO,1
A. ACEVEDO,2 A. L. SCOPEL1 & C. L. BALLARÉ1
1IFEVA, Departamento de Ecología, Facultad de Agronomía, Universidad de Buenos Aires, Av. San Martín 4453 (1417) BuenosAires, Argentina, and 2IGEAF CICA-INTA (1712) Castelar, and Departamento de Ciencia y Tecnología, Universidad Nacional deQuilmes (1876) Bernal, Argentina
ORIGINAL ARTICLE OA 220 EN
Correspondence: C. L. Ballaré. Fax: +54-1514-8730; e-mail:[email protected]
61

response to UV-B; biochemical studies have shown thatseveral key enzymes of the pathway are coordinatelyinduced in response to UV irradiation (Beggs &Wellmann 1994), and the importance of phenylpropanoidcompounds in UV screening and protection has been con-vincingly demonstrated using flavonoid-deficientmutants [see Landry, Chapple & Last (1995) and Reuber,Bornman & Weissenböck (1996)]. Laboratory studiesalso suggest that UV-B can cause oxidative damage tolipids and other cellular components (Kramer et al. 1991;Malanga & Puntarulo 1995; Takeuchi et al. 1995), andactivate antioxidant defences (Willekens et al. 1994;Malanga & Puntarulo 1995; Kim et al. 1996a; Rao,Paliyath & Ormrod 1996; Takeuchi et al. 1996). As anadditional damaging effect, UV-B radiation can inducelesions in DNA in the form of cyclobutane pyrimidinedimers (CPDs) and pyrimidine (6,4) pyrimidinone dimers(6,4 photoproducts) (Britt 1995).
There is a significant knowledge gap between field andlaboratory studies, which has two components. First, theoccurrence of certain effects of UV-B under field condi-tions has not yet been demonstrated. There are virtually nopublished studies on oxidative damage and antioxidantresponses induced by solar UV-B under field conditions.Kim et al. (1996a) have shown an increase in antioxidantenzymes in rice and cucumber in response to UV-B irradia-tion in laboratory experiments, but UV-B supplementationstudies with rice by the same group failed to detect signifi-cant antioxidant responses in the field (Kim et al. 1996b).Second, although some responses are known to occur inthe field, their functional implications are still unclear. Forexample, although it has been demonstrated that solar UV-B increases the steady-state level of CPDs in field-grownplants (Ballaré et al. 1996), the physiological conse-quences of variations in the total CPD load have not beenestablished (Taylor et al. 1996).
The field experiments that we report in this paper weredesigned to assess the impact of ambient UV-B on thegrowth and yield of barley and to narrow the gap betweenbiochemical and physiological studies. Barley is an inter-esting model system for two reasons. First, commercialvarieties have short stature, which facilitates studies thatextend until crop maturity. Second, it provides an oppor-tunity to study the effects of solar UV-B on a cool-seasoncrop. This is significant because most field studies ongrowth and biochemical responses to solar UV-Breported thus far have been carried out in the summer.Specifically, we addressed the following questions. (1)Does springtime solar UV-B affect the morphology,biomass accumulation and yield of barley crops? (2)Does springtime solar UV-B affect DNA integrity? (3) Dobarley plants respond to solar UV-B radiation withchanges in antioxidant enzyme levels? (4) Is antioxidantcapacity an important factor in UV-B tolerance? As a firstapproach to define the role of antioxidant capacity in UV-B tolerance we carried out comparative experiments thatincluded a catalase (CAT)-deficient mutant and its wild-type mother line.
MATERIALS AND METHODS
Plant material and experimental design
All the experiments were carried out in the experimentalfields of IFEVA (34°35′ S; 58°29′ W), Buenos Aires,Argentina. Seeds of the CAT-deficient barley (HordeumvulgareL.) mutant RPr 79/4 and its normal mother line cv.Maris Mink (Kendall et al. 1983) were sown in individual300 cm3 plastic cones filled with standard topsoil. Therewere two sowing dates: 10 June 1997 and 17 June 1997.Seedlings were allowed to emerge in the field under eitherclear polyester films (0·1 mm thick, Mylar-D, Dupont,UK), which virtually cut off all UV radiation below310 nm [– UV-B treatment, see spectrum in Ballaré et al.(1996)], or Aclar films (0·04 mm thick, Allied Signal,Pottsville, PA, USA), which have very high transmittanceover the whole UV waveband (+ UV-B treatment). Sowingwas carried out in pots (rather than directly in the soil) inorder to ensure uniform initial size; on 4 July 1997 [24 or17 d after sowing (DAS)] the seedlings were transplantedto their final field location. The seedlings were planted inrows in 1× 1 m plots; the space between the rows was18 cm with the two genotypes planted in alternate rows;planting density was ≈ 50 m–2. For each sowing date therewere three replicates of each UV-B treatment, following arandomized complete block design (total number of fieldplots = 12). On the north and south side of each plot therewas a border row of wheat; the east and west sides werecovered with curtains of clear polyester or Aclar, depend-ing on the UV-B treatment. The filters were raised periodi-cally to maintain them ≈ 5 cm above the barley canopy.The level of UV-B attenuation was checked with a broad-band UV-B detector (SUD/240/W attached to a IL-1700research radiometer; International Light, Newburyport,MA, USA; peak spectral response at 290 nm; half-bandwidth = 20 nm), and it was found to be consistently greaterthan 95% at the centre of the – UV-B plots. The plots wereirrigated as needed to maintain the soil water content nearfield capacity, and weeds were controlled manually. Thecrops were harvested on 15 December 1997, when theplants in both treatments were yellowish and ripe.
Growth and morphology
All morphological measurements were taken on plantslocated at the centre of each plot. Leaf length and tillernumbers were measured several times during the grow-ing season. On the final harvest the number of tillers,spikes, and seeds produced per plant were counted. Drymass was obtained after oven-drying the plants at 70 °Cfor at least 48 h.
DNA damage analysis
For DNA damage analysis we collected the middle third ofthe youngest fully expanded leaf available at the time ofsampling (three leaves per plot) at 1300 h. The samples wereimmediately frozen in liquid nitrogen. DNA extraction was
62 C. A. Mazza et al.
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
carried out under dim orange light, essentially as describedby Doyle & Doyle (1987) using a cetyltrimethylammo-nium bromide (CTAB)-based procedure modified by theuse of polyvinyl polypyrrolidone (PVP) to eliminatepolyphenols during DNA purification. DNA was quanti-tated with ethidium bromide (Gallagher 1994) using aPeltier-cooled CCD camera/imager system (Fluor-SMultiImager, Bio-Rad, Hercules, CA, USA) for fluores-cence detection. DNA damage was assayed by determina-tion of CPDs using a method adapted from Stapleton, Mori& Walbot (1993). In brief, DNA samples (≈ 3000 ng) in TEbuffer were denaturated and immobilized on a chargedNylon blotting membrane (Zeta-Probe, Bio-Rad); CPDswere detected using the TDM-2 monoclonal antibody (giftfrom Dr Toshio Mori, Nara Medical University, Japan).The method is based on the detection of primary-boundantibody by alkaline phosphatase-conjugated secondaryantibody (Bio-Rad) using a chemiluminescent substrate(CSPD®, Tropix, Bedford, MA, USA). Chemiluminescencewas detected with the CCD/imager system.
Antioxidant enzymes and pigments
Samples for enzyme and pigment determinations werealways collected at solar noon on sunny days. For enzymeanalysis we collected the middle third of the youngest fullyexpanded leaf available at the time of sampling (threeleaves per plot). The tissue was wrapped in aluminium foil,placed on ice and processed within 15 min of the samplingtime. For determinations of total CAT and ascorbate perox-idase (APx) activities, the tissue (100 mg) was homoge-nized in 1 cm3 of 50 mol m–3 HEPES (pH 7·5) containing0·1 mol m–3 ethylene diamine tetraacetic acid (EDTA),PVP, and centrifuged at 9500g for 20 min at 4 °C. Enzymeswere assayed in the supernatant. CAT activity was mea-sured spectrophotometrically by following the consump-tion of H2O2 at 240 nm in a reaction mixture containing50 mol m–3 potassium phosphate buffer (pH 7·0),24 mol m–3 H2O2 and 20 mm3 of the sample in a final vol-ume of 600 mm3 (Aebi 1984). The reaction was recordedfor 60 s after the addition of the sample. APx activity wasmeasured spectrophotometrically by the decrease inabsorbance of ascorbate at 290 nm in a reaction mixturecontaining 50 mol m–3 potassium phosphate buffer(pH 7·0), 0·1 mol m–3 EDTA, 0·5 mol m–3 ascorbate and20 mm3 of the sample in a final volume of 620 mm3. Thereaction was started by adding 3 mm3 of 9·8 mol m–3 H2O2
and the change in absorbance was followed for 30 s. Nocorrection for the oxidation of ascorbate in the absence ofsample was necessary (Nakano & Asada 1981). For deter-minations of superoxide dismutase (SOD) activity, the tis-sue (100 mg) was homogenized in 5 cm3 of 90% (v/v)acetone and centrifuged at 565g for 20 min at 4 °C. Thepellet was resuspended in 5 cm3 of 90% (v/v) acetone andcentrifuged at 4000g for 10 min at 4 °C. This step wasrepeated and the pellet was resuspended in 50 mol m–3
potassium phosphate buffer (pH 7·8), 0·5 mol m–3 EDTAand centrifuged at 10 000g for 15 min at 4 °C. SOD activity
was assayed spectrophotometrically (550 nm) in the super-natant based on its capacity to inhibit the reduction ofcytochrome c by superoxide radicals generated by xan-thine–xanthine oxidase (Beuchamp & Fridovich 1971). Thereaction mixture contained 50 mol m–3 potassium phos-phate buffer (pH 7·8), 0·1 mol m–3 EDTA, 20 mmol m–3
cytochrome c, 53 mmol m–3 xanthine, and the amount ofxanthine oxidase required to detect a change of 0·04absorbance units min–1. One unit of SOD activity wasdefined as the amount of sample necessary to produce 50%inhibition of the cytochrome c reduction rate. Protein deter-mination was carried out according to Bradford (1976).
For pigment analysis we collected four leaf discs per plot(0·45 cm diameter, youngest fully expanded leaf). Eachdisc was placed in 1·4 cm3 of 99:1 methanol:HCl andallowed to extract for 48 h at – 4 °C. Absorbance of theextracts was read at 305 nm or 546 nm for determinationsof total UV-absorbing compounds or anthocyanins, respec-tively. The dry mass of the discs was determined afteroven-drying at 70 °C for 4 h.
Statistical analyses were performed using PROC GLMin theSASv 6·12 package (SAS Institute, Cary, NC, USA); appro-priate transformations of the primary data were used whenneeded to meet the assumptions of the analysis of variance.
RESULTS
Solar UV-B affects morphological developmentand increases the concentration of UV-absorbingcompounds
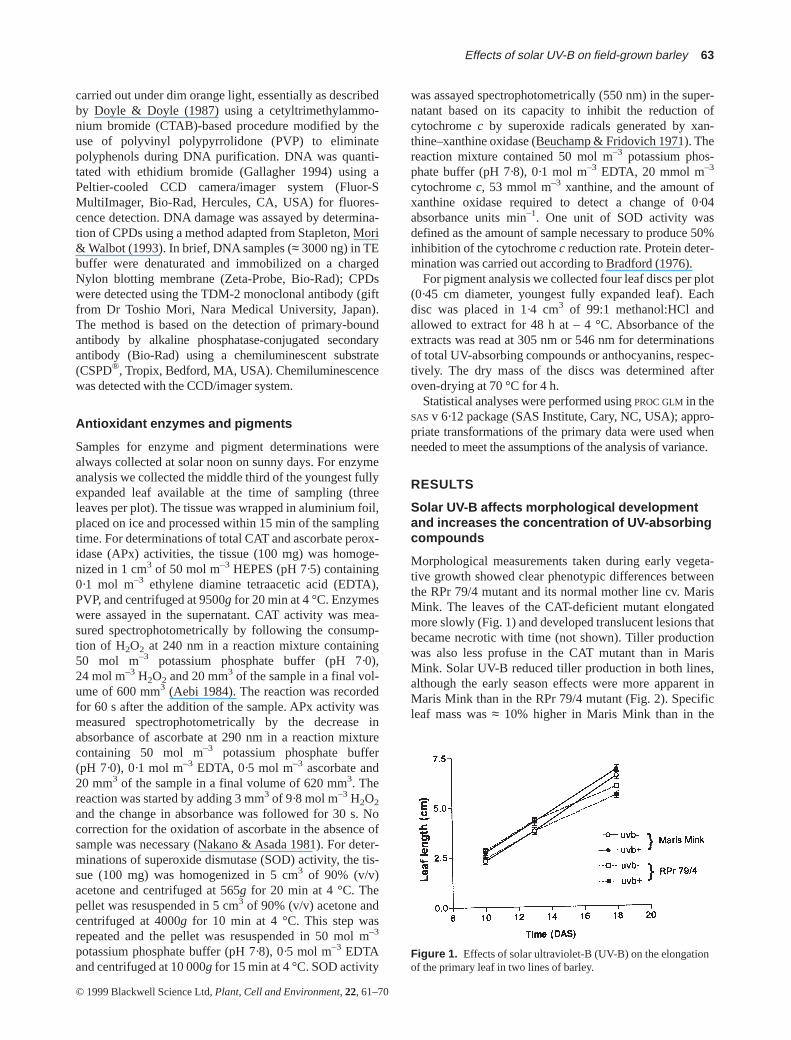
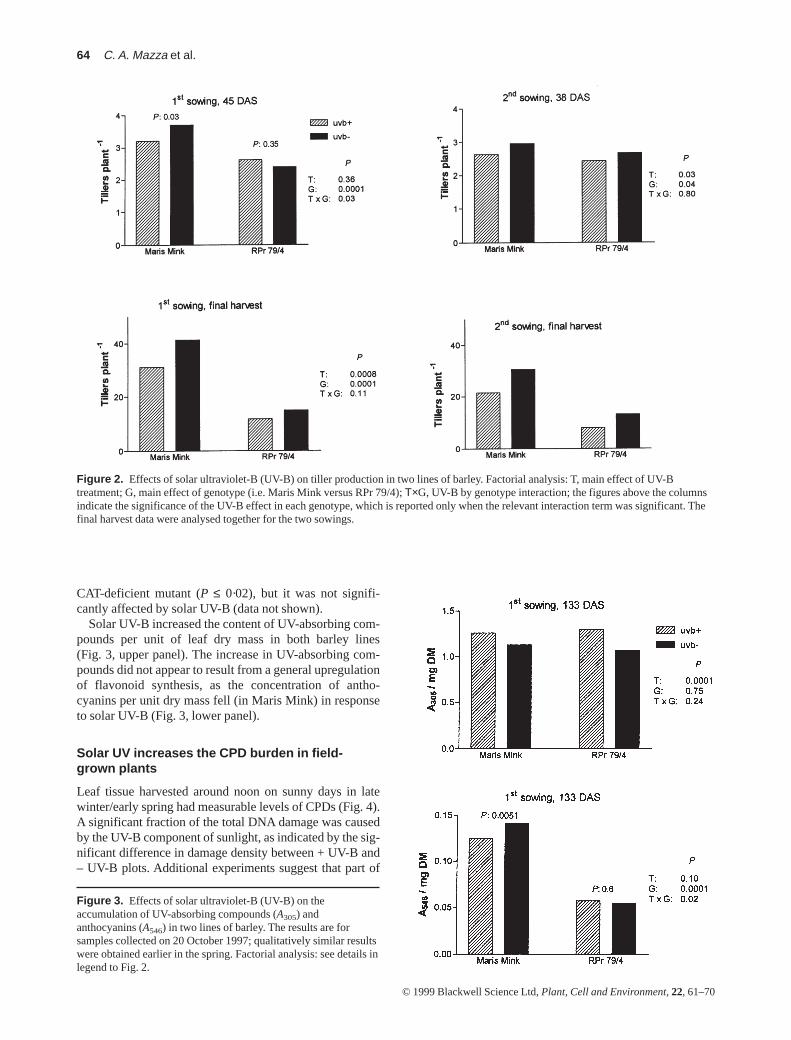
Morphological measurements taken during early vegeta-tive growth showed clear phenotypic differences betweenthe RPr 79/4 mutant and its normal mother line cv. MarisMink. The leaves of the CAT-deficient mutant elongatedmore slowly (Fig. 1) and developed translucent lesions thatbecame necrotic with time (not shown). Tiller productionwas also less profuse in the CAT mutant than in MarisMink. Solar UV-B reduced tiller production in both lines,although the early season effects were more apparent inMaris Mink than in the RPr 79/4 mutant (Fig. 2). Specificleaf mass was ≈ 10% higher in Maris Mink than in the
Effects of solar UV-B on field-grown barley 63
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
Figure 1. Effects of solar ultraviolet-B (UV-B) on the elongationof the primary leaf in two lines of barley.
CAT-deficient mutant (P ≤ 0·02), but it was not signifi-cantly affected by solar UV-B (data not shown).
Solar UV-B increased the content of UV-absorbing com-pounds per unit of leaf dry mass in both barley lines(Fig. 3, upper panel). The increase in UV-absorbing com-pounds did not appear to result from a general upregulationof flavonoid synthesis, as the concentration of antho-cyanins per unit dry mass fell (in Maris Mink) in responseto solar UV-B (Fig. 3, lower panel).
Solar UV increases the CPD burden in field-grown plants
Leaf tissue harvested around noon on sunny days in latewinter/early spring had measurable levels of CPDs (Fig. 4).A significant fraction of the total DNA damage was causedby the UV-B component of sunlight, as indicated by the sig-nificant difference in damage density between + UV-B and– UV-B plots. Additional experiments suggest that part of
64 C. A. Mazza et al.
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
Figure 2. Effects of solar ultraviolet-B (UV-B) on tiller production in two lines of barley. Factorial analysis: T, main effect of UV-Btreatment; G, main effect of genotype (i.e. Maris Mink versus RPr 79/4); Τ×G, UV-B by genotype interaction; the figures above the columnsindicate the significance of the UV-B effect in each genotype, which is reported only when the relevant interaction term was significant. Thefinal harvest data were analysed together for the two sowings.
Figure 3. Effects of solar ultraviolet-B (UV-B) on theaccumulation of UV-absorbing compounds (A305) andanthocyanins (A546) in two lines of barley. The results are forsamples collected on 20 October 1997; qualitatively similar resultswere obtained earlier in the spring. Factorial analysis: see details inlegend to Fig. 2.
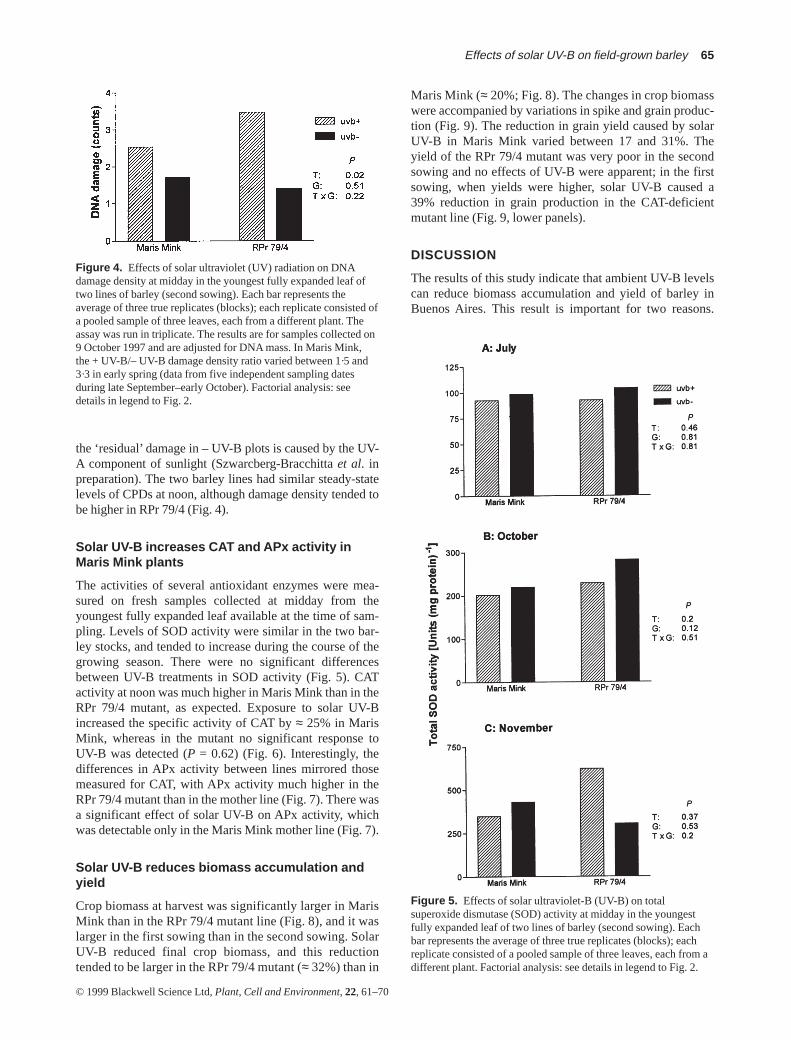
the ‘residual’ damage in – UV-B plots is caused by the UV-A component of sunlight (Szwarcberg-Bracchitta et al. inpreparation). The two barley lines had similar steady-statelevels of CPDs at noon, although damage density tended tobe higher in RPr 79/4 (Fig. 4).
Solar UV-B increases CAT and APx activity inMaris Mink plants
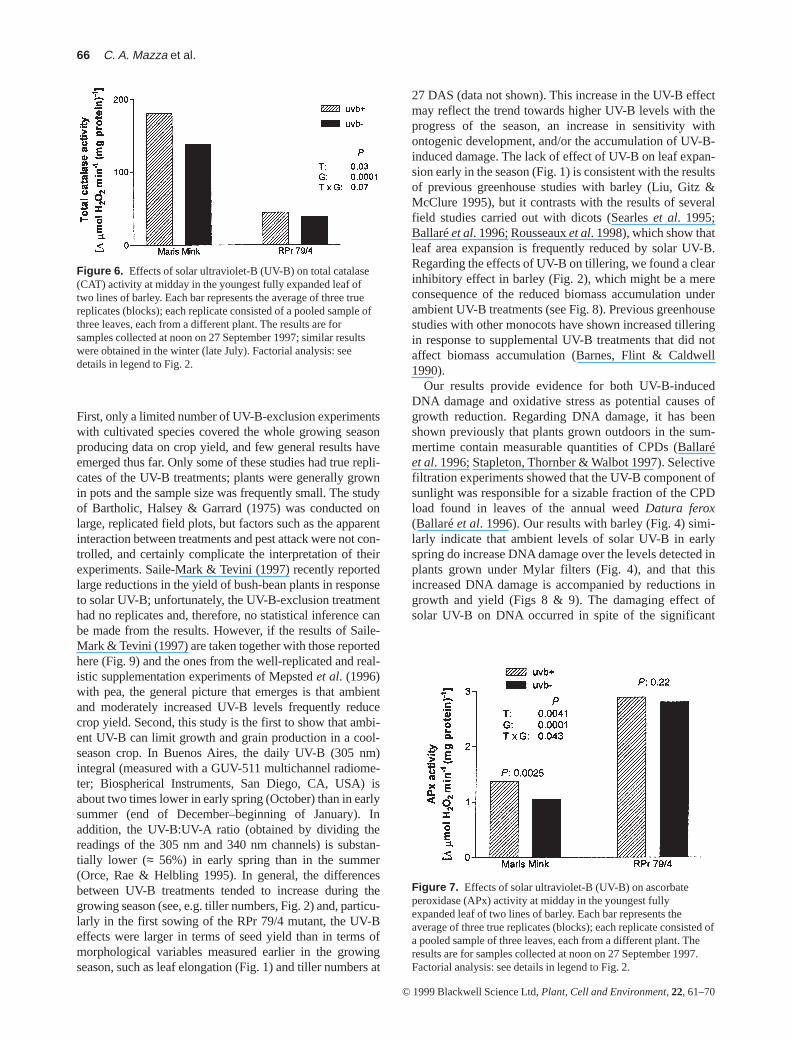
The activities of several antioxidant enzymes were mea-sured on fresh samples collected at midday from theyoungest fully expanded leaf available at the time of sam-pling. Levels of SOD activity were similar in the two bar-ley stocks, and tended to increase during the course of thegrowing season. There were no significant differencesbetween UV-B treatments in SOD activity (Fig. 5). CATactivity at noon was much higher in Maris Mink than in theRPr 79/4 mutant, as expected. Exposure to solar UV-Bincreased the specific activity of CAT by ≈ 25% in MarisMink, whereas in the mutant no significant response toUV-B was detected (P = 0.62) (Fig. 6). Interestingly, thedifferences in APx activity between lines mirrored thosemeasured for CAT, with APx activity much higher in theRPr 79/4 mutant than in the mother line (Fig. 7). There wasa significant effect of solar UV-B on APx activity, whichwas detectable only in the Maris Mink mother line (Fig. 7).
Solar UV-B reduces biomass accumulation andyield
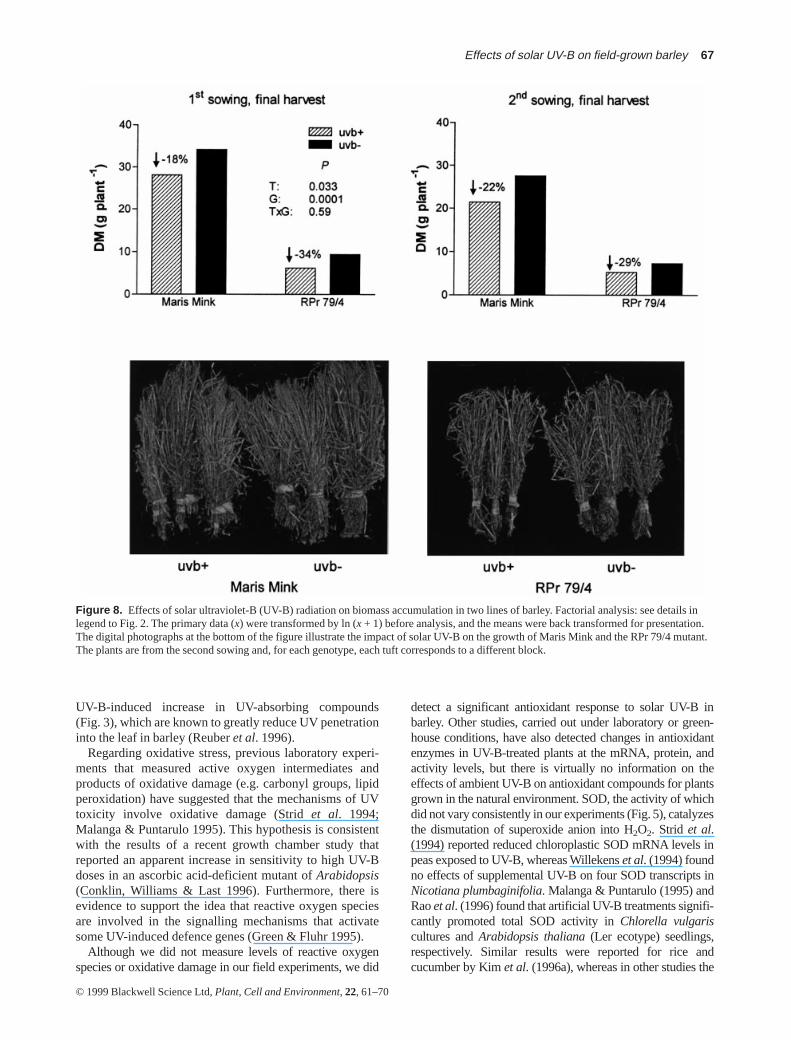
Crop biomass at harvest was significantly larger in MarisMink than in the RPr 79/4 mutant line (Fig. 8), and it waslarger in the first sowing than in the second sowing. SolarUV-B reduced final crop biomass, and this reductiontended to be larger in the RPr 79/4 mutant (≈ 32%) than in
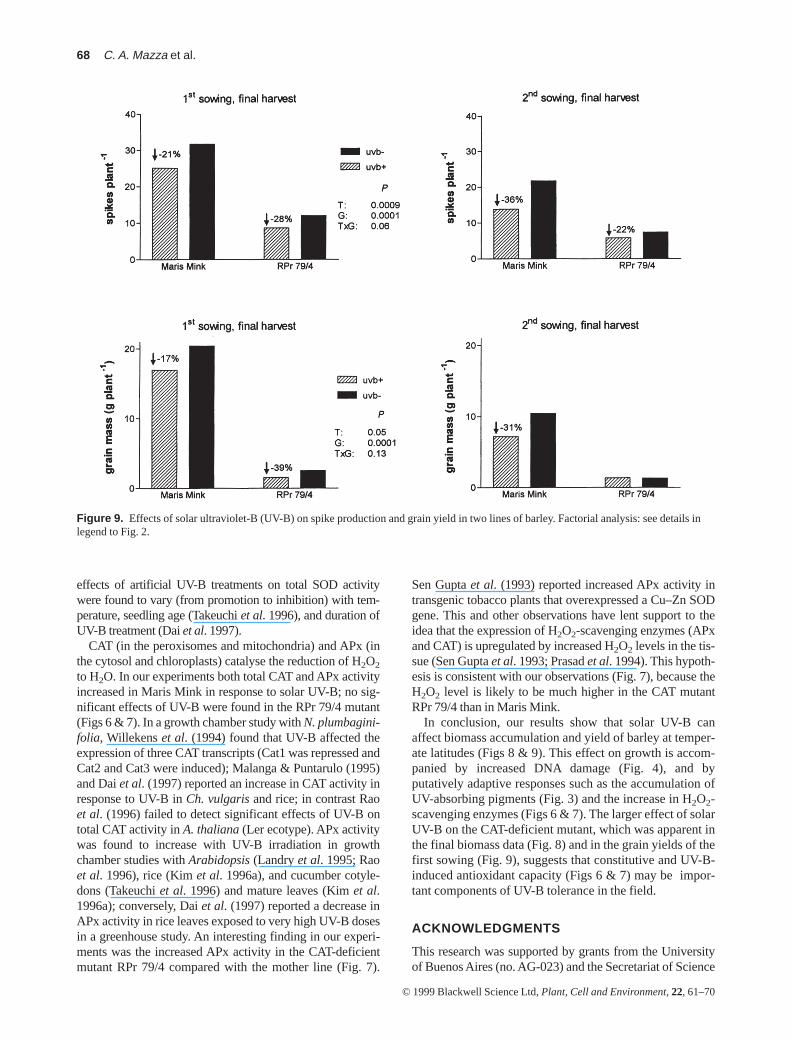
Maris Mink (≈ 20%; Fig. 8). The changes in crop biomasswere accompanied by variations in spike and grain produc-tion (Fig. 9). The reduction in grain yield caused by solarUV-B in Maris Mink varied between 17 and 31%. Theyield of the RPr 79/4 mutant was very poor in the secondsowing and no effects of UV-B were apparent; in the firstsowing, when yields were higher, solar UV-B caused a39% reduction in grain production in the CAT-deficientmutant line (Fig. 9, lower panels).
DISCUSSION
The results of this study indicate that ambient UV-B levelscan reduce biomass accumulation and yield of barley inBuenos Aires. This result is important for two reasons.
Effects of solar UV-B on field-grown barley 65
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
Figure 4. Effects of solar ultraviolet (UV) radiation on DNAdamage density at midday in the youngest fully expanded leaf oftwo lines of barley (second sowing). Each bar represents theaverage of three true replicates (blocks); each replicate consisted ofa pooled sample of three leaves, each from a different plant. Theassay was run in triplicate. The results are for samples collected on9 October 1997 and are adjusted for DNA mass. In Maris Mink,the + UV-B/– UV-B damage density ratio varied between 1·5 and3·3 in early spring (data from five independent sampling datesduring late September–early October). Factorial analysis: seedetails in legend to Fig. 2.
Figure 5. Effects of solar ultraviolet-B (UV-B) on totalsuperoxide dismutase (SOD) activity at midday in the youngestfully expanded leaf of two lines of barley (second sowing). Eachbar represents the average of three true replicates (blocks); eachreplicate consisted of a pooled sample of three leaves, each from adifferent plant. Factorial analysis: see details in legend to Fig. 2.
First, only a limited number of UV-B-exclusion experimentswith cultivated species covered the whole growing seasonproducing data on crop yield, and few general results haveemerged thus far. Only some of these studies had true repli-cates of the UV-B treatments; plants were generally grownin pots and the sample size was frequently small. The studyof Bartholic, Halsey & Garrard (1975) was conducted onlarge, replicated field plots, but factors such as the apparentinteraction between treatments and pest attack were not con-trolled, and certainly complicate the interpretation of theirexperiments. Saile-Mark & Tevini (1997) recently reportedlarge reductions in the yield of bush-bean plants in responseto solar UV-B; unfortunately, the UV-B-exclusion treatmenthad no replicates and, therefore, no statistical inference canbe made from the results. However, if the results of Saile-Mark & Tevini (1997) are taken together with those reportedhere (Fig. 9) and the ones from the well-replicated and real-istic supplementation experiments of Mepsted et al. (1996)with pea, the general picture that emerges is that ambientand moderately increased UV-B levels frequently reducecrop yield. Second, this study is the first to show that ambi-ent UV-B can limit growth and grain production in a cool-season crop. In Buenos Aires, the daily UV-B (305 nm)integral (measured with a GUV-511 multichannel radiome-ter; Biospherical Instruments, San Diego, CA, USA) isabout two times lower in early spring (October) than in earlysummer (end of December–beginning of January). Inaddition, the UV-B:UV-A ratio (obtained by dividing thereadings of the 305 nm and 340 nm channels) is substan-tially lower (≈ 56%) in early spring than in the summer(Orce, Rae & Helbling 1995). In general, the differencesbetween UV-B treatments tended to increase during thegrowing season (see, e.g. tiller numbers, Fig. 2) and, particu-larly in the first sowing of the RPr 79/4 mutant, the UV-Beffects were larger in terms of seed yield than in terms ofmorphological variables measured earlier in the growingseason, such as leaf elongation (Fig. 1) and tiller numbers at
27 DAS (data not shown). This increase in the UV-B effectmay reflect the trend towards higher UV-B levels with theprogress of the season, an increase in sensitivity withontogenic development, and/or the accumulation of UV-B-induced damage. The lack of effect of UV-B on leaf expan-sion early in the season (Fig. 1) is consistent with the resultsof previous greenhouse studies with barley (Liu, Gitz &McClure 1995), but it contrasts with the results of severalfield studies carried out with dicots (Searles et al. 1995;Ballaré et al. 1996; Rousseaux et al. 1998), which show thatleaf area expansion is frequently reduced by solar UV-B.Regarding the effects of UV-B on tillering, we found a clearinhibitory effect in barley (Fig. 2), which might be a mereconsequence of the reduced biomass accumulation underambient UV-B treatments (see Fig. 8). Previous greenhousestudies with other monocots have shown increased tilleringin response to supplemental UV-B treatments that did notaffect biomass accumulation (Barnes, Flint & Caldwell1990).
Our results provide evidence for both UV-B-inducedDNA damage and oxidative stress as potential causes ofgrowth reduction. Regarding DNA damage, it has beenshown previously that plants grown outdoors in the sum-mertime contain measurable quantities of CPDs (Ballaréet al. 1996; Stapleton, Thornber & Walbot 1997). Selectivefiltration experiments showed that the UV-B component ofsunlight was responsible for a sizable fraction of the CPDload found in leaves of the annual weed Datura ferox(Ballaré et al. 1996). Our results with barley (Fig. 4) simi-larly indicate that ambient levels of solar UV-B in earlyspring do increase DNA damage over the levels detected inplants grown under Mylar filters (Fig. 4), and that thisincreased DNA damage is accompanied by reductions ingrowth and yield (Figs 8 & 9). The damaging effect ofsolar UV-B on DNA occurred in spite of the significant
66 C. A. Mazza et al.
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
Figure 6. Effects of solar ultraviolet-B (UV-B) on total catalase(CAT) activity at midday in the youngest fully expanded leaf oftwo lines of barley. Each bar represents the average of three truereplicates (blocks); each replicate consisted of a pooled sample ofthree leaves, each from a different plant. The results are forsamples collected at noon on 27 September 1997; similar resultswere obtained in the winter (late July). Factorial analysis: seedetails in legend to Fig. 2.
Figure 7. Effects of solar ultraviolet-B (UV-B) on ascorbateperoxidase (APx) activity at midday in the youngest fullyexpanded leaf of two lines of barley. Each bar represents theaverage of three true replicates (blocks); each replicate consisted ofa pooled sample of three leaves, each from a different plant. Theresults are for samples collected at noon on 27 September 1997.Factorial analysis: see details in legend to Fig. 2.
UV-B-induced increase in UV-absorbing compounds(Fig. 3), which are known to greatly reduce UV penetrationinto the leaf in barley (Reuber et al. 1996).
Regarding oxidative stress, previous laboratory experi-ments that measured active oxygen intermediates andproducts of oxidative damage (e.g. carbonyl groups, lipidperoxidation) have suggested that the mechanisms of UVtoxicity involve oxidative damage (Strid et al. 1994;Malanga & Puntarulo 1995). This hypothesis is consistentwith the results of a recent growth chamber study thatreported an apparent increase in sensitivity to high UV-Bdoses in an ascorbic acid-deficient mutant of Arabidopsis(Conklin, Williams & Last 1996). Furthermore, there isevidence to support the idea that reactive oxygen speciesare involved in the signalling mechanisms that activatesome UV-induced defence genes (Green & Fluhr 1995).
Although we did not measure levels of reactive oxygenspecies or oxidative damage in our field experiments, we did
detect a significant antioxidant response to solar UV-B inbarley. Other studies, carried out under laboratory or green-house conditions, have also detected changes in antioxidantenzymes in UV-B-treated plants at the mRNA, protein, andactivity levels, but there is virtually no information on theeffects of ambient UV-B on antioxidant compounds for plantsgrown in the natural environment. SOD, the activity of whichdid not vary consistently in our experiments (Fig. 5), catalyzesthe dismutation of superoxide anion into H2O2. Strid et al.(1994) reported reduced chloroplastic SOD mRNA levels inpeas exposed to UV-B, whereas Willekens et al. (1994) foundno effects of supplemental UV-B on four SOD transcripts inNicotiana plumbaginifolia. Malanga & Puntarulo (1995) andRao et al. (1996) found that artificial UV-B treatments signifi-cantly promoted total SOD activity in Chlorella vulgariscultures and Arabidopsis thaliana(Ler ecotype) seedlings,respectively. Similar results were reported for rice andcucumber by Kim et al. (1996a), whereas in other studies the
Effects of solar UV-B on field-grown barley 67
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
Figure 8. Effects of solar ultraviolet-B (UV-B) radiation on biomass accumulation in two lines of barley. Factorial analysis: see details inlegend to Fig. 2. The primary data (x) were transformed by ln (x + 1) before analysis, and the means were back transformed for presentation.The digital photographs at the bottom of the figure illustrate the impact of solar UV-B on the growth of Maris Mink and the RPr 79/4 mutant.The plants are from the second sowing and, for each genotype, each tuft corresponds to a different block.
effects of artificial UV-B treatments on total SOD activitywere found to vary (from promotion to inhibition) with tem-perature, seedling age (Takeuchi et al. 1996), and duration ofUV-B treatment (Dai et al. 1997).
CAT (in the peroxisomes and mitochondria) and APx (inthe cytosol and chloroplasts) catalyse the reduction of H2O2
to H2O. In our experiments both total CAT and APx activityincreased in Maris Mink in response to solar UV-B; no sig-nificant effects of UV-B were found in the RPr 79/4 mutant(Figs 6 & 7). In a growth chamber study with N. plumbagini-folia, Willekens et al. (1994) found that UV-B affected theexpression of three CAT transcripts (Cat1 was repressed andCat2 and Cat3 were induced); Malanga & Puntarulo (1995)and Dai et al. (1997) reported an increase in CAT activity inresponse to UV-B in Ch. vulgarisand rice; in contrast Raoet al. (1996) failed to detect significant effects of UV-B ontotal CAT activity in A. thaliana(Ler ecotype). APx activitywas found to increase with UV-B irradiation in growthchamber studies withArabidopsis(Landry et al. 1995; Raoet al. 1996), rice (Kim et al. 1996a), and cucumber cotyle-dons (Takeuchi et al. 1996) and mature leaves (Kim et al.1996a); conversely, Dai et al. (1997) reported a decrease inAPx activity in rice leaves exposed to very high UV-B dosesin a greenhouse study. An interesting finding in our experi-ments was the increased APx activity in the CAT-deficientmutant RPr 79/4 compared with the mother line (Fig. 7).
Sen Gupta et al. (1993) reported increased APx activity intransgenic tobacco plants that overexpressed a Cu–Zn SODgene. This and other observations have lent support to theidea that the expression of H2O2-scavenging enzymes (APxand CAT) is upregulated by increased H2O2 levels in the tis-sue (Sen Gupta et al. 1993; Prasad et al. 1994). This hypoth-esis is consistent with our observations (Fig. 7), because theH2O2 level is likely to be much higher in the CAT mutantRPr 79/4 than in Maris Mink.
In conclusion, our results show that solar UV-B canaffect biomass accumulation and yield of barley at temper-ate latitudes (Figs 8 & 9). This effect on growth is accom-panied by increased DNA damage (Fig. 4), and byputatively adaptive responses such as the accumulation ofUV-absorbing pigments (Fig. 3) and the increase in H2O2-scavenging enzymes (Figs 6 & 7). The larger effect of solarUV-B on the CAT-deficient mutant, which was apparent inthe final biomass data (Fig. 8) and in the grain yields of thefirst sowing (Fig. 9), suggests that constitutive and UV-B-induced antioxidant capacity (Figs 6 & 7) may be impor-tant components of UV-B tolerance in the field.
ACKNOWLEDGMENTS
This research was supported by grants from the Universityof Buenos Aires (no. AG-023) and the Secretariat of Science
68 C. A. Mazza et al.
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
Figure 9. Effects of solar ultraviolet-B (UV-B) on spike production and grain yield in two lines of barley. Factorial analysis: see details inlegend to Fig. 2.

and Technology (BID 802 OC-AR, PID no. 394) to C. L. B.and A. L. S., and also in part by a grant from the E. U.(INCO-DC: IC18-CT96–0124) to A. A. We thank Dr ToshioMori for the antibodies used for the detection of CPDs, DrAnn Stapleton for her advice on DNA damage analysis, andGabriela Malanga and Dr Susana Puntarulo for their adviceon the determination of antioxidant enzymes. AndrésArakelian, M. Laura Federico, María Irianni, and SebastiánMunilla provided excellent technical assistance.
REFERENCES
Aebi H. (1984) Catalase in vitro.Methods in Enzymology105,121–126.
Ballaré C.L., Barnes P.W. & Flint S.D. (1995a) Inhibition ofhypocotyl elongation by ultraviolet-B radiation in de-etiolatingtomato seedlings. I. The photoreceptor.Physiologia Plantarum93,584–592.
Ballaré C.L., Barnes P.W., Flint S.D. & Price S. (1995b) Inhibitionof hypocotyl elongation by ultraviolet-B radiation in de-etiolat-ing tomato seedlings. II. Time-course, comparisons withflavonoid responses, and adaptive significance.PhysiologiaPlantarum93,593–601.
Ballaré C.L., Scopel A.L., Stapleton A.L. & Yanovsky M.J. (1996)Solar ultraviolet-B radiation affects seedling emergence, DNAintegrity, plant morphology, growth rate, and attractiveness toherbivore insects in Datura ferox. Plant Physiology 112,161–170.
Barnes P.W., Flint S.D. & Caldwell M.M. (1990) Morphologicalresponses of crop and weed species of different growth forms toultraviolet-B radiation.American Journal of Botany77,1354–1360.
Bartholic J.F., Halsey L.H. & Garrard I.A. (1975) Field trials withfilters to test for effects of UV radiation on agricultural produc-tivity. In Climatic Impact Assessment Program Monograph No. 5(eds D.S. Nachtwey, M.M. Caldwell & R.H. Biggs), pp.4/61–4/71. US Department of Transportation no. DOT-TST-76–55, National Technical Information Service, Springfield, VA.
Beggs C.J. & Wellmann E. (1994) Photocontrol of flavonoidbiosynthesis. In Photomorphogenesis in Plants, 2nd edn (edsR.E. Kendrick & G.H.M. Kronenberg), pp. 733–751. Kluwer,Dordrecht.
Beuchamp C.H. & Fridovich I. (1971) Superoxide dismutase:improved assays and an assay applicable to acrylamide gels.Analytical Biochemistry44,276–287.
Beyschlag W., Barnes P.W., Flint S.D. & Caldwell M.M. (1988)Enhanced UV-B radiation has no effect on photosynthetic char-acteristics of wheat (Triticum aestivumL.) and wild oat (AvenafatuaL.) under glasshouse and field conditions.Photosynthetica22,516–525.
Bornman J.F. & Teramura A.H. (1993) Effects of ultraviolet-B radi-ation on terrestrial plants. In Environmental UV Photobiology(eds A.R. Young, L.O. Björn, J. Moan & W. Nultsch), pp.427–471. Plenum Press, New York.
Bradford M. (1976) A rapid and sensitive method for the quantifica-tion of microgram quantities of protein utilizing the principle ofprotein–dye binding.Analytical Biochemistry72,248–254.
Britt A.B. (1995) Repair of DNA damage induced by ultravioletradiation.Plant Physiology108,891–896.
Caldwell M.M., Teramura A.H., Tevini M., Bornman J.F., BjörnL.O. & Kulandaivelu G. (1995) Effects of increased solar ultravi-olet radiation on terrestrial plants.Ambio24,166–173.
Conklin P.L., Williams E.H. & Last R.L. (1996) Environmentalstress sensitivity of an ascorbic acid-deficient Arabidopsismutant.Proceedings of the National Academy of Sciences of theUSA93,9970–9974.
Dai Q., Yan B., Huang S., Liu X., Peng S., Miranda M.L.L., ChavezA.Q., Vergara B.S. & Olszyk D.M. (1997) Response of oxidativestress defense systems in rice (Oryza sativa) leaves with supple-mental UV-B radiation.Physiologia Plantarum101,301–308.
Doyle J.J. & Doyle J.L. (1987) A rapid DNA isolation procedure forsmall quantities of fresh leaf tissue. Phytochemistry Bulletin19,11–15.
Ensminger P.A. (1993) Control of development in plants and fungiby far-UV radiation.Physiologia Plantarum88,501–508.
Gallagher S.R. (1994) Quantitation of DNA and RNA with absorp-tion and fluorescence spectroscopy. In Current Protocols inMolecular Biology, Vol. 3 (eds D.M. Ausubel, R. Brent, R.E.Kingston, D.D. Moore, J.G. Seidman, J.A. Smith & K. Struhl),(Suppl. 28). J. Wiley & Sons, New York.
Green R. & Fluhr R. (1995) UV-B-induced PR-1 accumulation ismediated by active oxygen species.Plant Cell7, 203–212.
Kendall A.C., Keys A.J., Turner J.C., Lea P.J. & Miflin B.J. (1983)The isolation and characterization of a catalase-deficient mutantof barley (Hordeum vulgareL.). Planta159,501–511.
Kim H.Y., Kobayashi K., Nouchi I. & Yoneyama T. (1996a)Differential influences of UV-B radiation on antioxidants andrelated enzymes between rice (Oryza sativaL.) and cucumber(Cucumis sativusL.) leaves.Environmental Science9, 55–63.
Kim H.Y., Kobayashi K., Nouchi I. & Yoneyama T. (1996b) Changesin antioxidant levels and activities of related enzymes in rice(Oryza sativaL.) leaves irradiated with enhanced UV-B radiationunder field conditions.Environmental Science9,55–63.
Kramer G.F., Norman H.A., Krizek D.T. & Mireki R.M. (1991)Influence of UV-B radiation on polyamines, lipid peroxidationand membrane lipids in cucumber.Phytochemistry 30,2101–2108.
Landry L.G., Chapple C.C.S. & Last R.L. (1995) Arabidopsismutants lacking phenolic sunscreens exhibit enhanced ultra-violet-B injury and oxidative damage.Plant Physiology109,1159–1166.
Liu L., Gitz D.C. III & McClure J.W. (1995) Effects of UV-B onflavonoids, ferulic acid, growth and photosynthesis in barley pri-mary leaves.Physiologia Plantarum93,725–733.
Malanga G. & Puntarulo S. (1995) Oxidative responses and antiox-idant content in Chlorella vulgarisafter exposure to ultraviolet-Bradiation.Physiologia Plantarum94,672–679.
Mark U., Saile-Mark M. & Tevini M. (1996) Effects of solar UVBradiation on growth, flowering and yield of Central and SouthernEuropean maize cultivars (Zea maysL.). Photochemistry andPhotobiology64,457–463.
Mepsted R., Paul N.D., Stephen J., Corlett J.E., Nogués S., BakerN.R., Jones H.G. & Ayres P.G. (1996) Effects of enhanced UV-Bradiation on pea (Pisum sativumL.) grown under field conditionsin the UK.Global Change Biology2, 325–334.
Nakano Y. & Asada K. (1981) Hydrogen peroxide is scavenged byascorbate-specific peroxidase in spinach chloroplasts.Plant CellPhysiology22,539–549.
Orce L.V., Rae G. & Helbling E.W. (1995) Latitudinal UVR-PARmonitoring network in Argentina: data from October 1994 toSeptember 1995. Contribución Científica no. 17 of the CentroAustral de Investigaciones Científicas (CADIC).Ushuaia,Argentina.
Prasad T.K., Anderson M.D., Martin B.A. & Stewart C.R. (1994)Evidence for chilling-induced oxidative stress in maize seedlingsand a regulatory role for hydrogen peroxide.Plant Cell6,65–74.
Rao M.V., Paliyath G. & Ormrod D.P. (1996) Ultraviolet-B andozone-induced biochemical changes in antioxidant enzymes ofArabidopsis thaliana. Plant Physiology110,125–136.
Reuber S., Bornman J.F. & Weissenböck G. (1996) A flavonoidmutant of barley (Hordeum vulgareL.) exhibits increased sensi-tivity to UV-B radiation in the primary leaf.Plant Cell andEnvironment19,593–601.
Effects of solar UV-B on field-grown barley 69
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70

Rousseaux M.C., Ballaré C.L., Scopel A.L., Searles P.S. &Caldwell M.M. (1998) Solar ultraviolet-B radiation affectsplant–insect interactions in a natural ecosystem of Tierra delFuego (southern Argentina).Oecologia116, 528–535.
Rozema J., van de Staaij J., Björn L.O. & Caldwell M.M. (1997)UV-B as an environmental factor in plant life: stress and regula-tion. Trends in Ecology and Evolution12,22–28.
Saile-Mark M. & Tevini M. (1997) Effects of solar UV-B radiationon growth, flowering and yield of central and southern Europeanbush bean cultivars (Phaseolus vulgarisL.). Plant Ecology128,114–125.
Searles P.S., Caldwell M.M. & Winter K. (1995) The response offive tropical species to solar ultraviolet-B radiation.AmericanJournal of Botany82,445–453.
Sen Gupta A., Webb R.P., Holaday A.S. & Allen R.D. (1993)Overexpression of superoxide dismutase protects plants fromoxidative stress. Induction of ascorbate peroxidase in superoxidedismutase-overexpressing plants.Plant Physiology 103,1067–1073.
Stapleton A.E., Mori T. & Walbot V. (1993) A simple and sensitiveantibody-based method to measure UV-induced DNA damage inZea mays. Plant Molecular Biology Reporter11,230–236.
Stapleton A.E., Thornber C.S. & Walbot V. (1997) UV-B compo-nent of sunlight causes measurable damage in field-grown maize
(Zea maysL.): developmental and cellular heterogeneity of dam-age and repair.Plant, Cell and Environment20,279–290.
Strid Å., Chow W.S. & Anderson J.M. (1994) UV-B damage andprotection at the molecular level in plants.PhotosynthesisResearch39,475–489.
Takeuchi Y., Fukumoto R., Kasahara H., Sakaki T. & Kitao M.(1995) Peroxidation of lipids and growth inhibition induced byUV-B irradiation.Plant Cell Reports14,566–670.
Takeuchi Y., Kubo H., Kasahara H. & Sakaki T. (1996) Adaptivealterations in the activities of scavengers of active oxygen incucumber cotyledons irradiated with UV-B.Journal of PlantPhysiology147,589–592.
Taylor R.M., Nikaido O., Jordan B.R., Rosamond J., Bray C.M. &Tobin K. (1996) Ultraviolet-B induced DNA lesions and theirremoval in wheat (Triticum aestivumL.) leaves.Plant, Cell andEnvironment19,171–181.
Willekens H., Van Camp W., Van Montagu M., Inzé D.,Langebartels C. & Sandermann H. Jr (1994) Ozone, sulfur diox-ide and ultraviolet B have similar effects on mRNA accumula-tion of antioxidant genes in Nicotiana plumbaginifoliaL. PlantPhysiology106,1007–1014.
Received 25 May 1998; received in revised form 8 August 1998;accepted for publication 8 August 1998
70 C. A. Mazza et al.
© 1999 Blackwell Science Ltd, Plant, Cell and Environment,22, 61–70
Top Related
Copyright © 2022 FDOKUMEN

