







Bahasa
Halaman
Hukum


The Distinct Agonistic Properties of thePhenylpyrazolosteroid Cortivazol RevealInterdomain Communication within theGlucocorticoid Receptor
Noritada Yoshikawa, Keiko Yamamoto, Noriaki Shimizu, Sachiko Yamada, Chikao Morimoto, andHirotoshi Tanaka
Division of the Clinical Immunology, the Advanced Clinical Research Center, the Institute of MedicalScience, the University of Tokyo, Tokyo 108-8639; and the Institute of Biomaterials andBioengineering, Tokyo Medical and Dental University, Tokyo 101-0062, Japan
Recent structural analyses of the nuclear recep-tors establish a paradigm of receptor activation, inwhich agonist binding induces the ligand bindingdomain (LBD)/activation function-2 helix to form acharge clamp for coactivator recruitment. How-ever, these analyses have not sufficiently ad-dressed the mechanisms for differential actions ofvarious synthetic steroids in terms of fine tuning ofmultiple functions of whole receptor molecules. Inthe present study, we used the glucocorticoid re-ceptor (GR)-specific agonist cortivazol (CVZ) toprobe the plasticity and functional modularity ofthe GR. Structural docking analysis revealed thatalthough CVZ is more bulky than other agonists, itcan be accommodated in the ligand binding pocketof the GR by reorientation of several amino acidside chains but without major alterations in theactive conformation of the LBD. In this induced fit
model, the phenylpyrazole A-ring of CVZ estab-lishes additional contacts with helices 3 and 5 ofthe LBD that may contribute to a more stable LBDconfiguration. Structural and functional analysisrevealed that CVZ is able to compensate for thedeleterious effects of a C-terminal deletion of theLBD in a manner that mimics the stabilizing influ-ence of the F602S point mutation. CVZ-mediatedproductive recruitment of transcriptional interme-diary factor 2 to the C-terminally deleted LBD re-quires the receptor’s own DNA binding domain andis positively influenced by the N-terminal regions ofGR or progesterone receptor. These results sup-port a model where ligand-dependent conforma-tional changes in the LBD play a role in GR-medi-ated gene regulation via modular interaction withthe DBD and activation function-1. (Molecular En-docrinology 19: 1110–1124, 2005)
GLUCOCORTICOIDS ARE PRODUCED in the ad-renal cortex under the strict control of the hypo-
thalamus-pituitary-adrenal axis and exert a variety ofbiological actions including the regulation of glucoseand lipid metabolism, electrolyte balance, and modu-lation of the immune, cardiovascular, and central ner-vous system (1, 2). Multiple compounds with glu-cocorticoid activity including dexamethasone (DEX),prednisolone, and cortivazol (CVZ) are widely used asan antiinflammatory and/or immunosuppressiveagents (3). At pharmacological doses, however, pa-
tients often suffer from side effects of glucocorticoids,the molecular basis for which have not been fully clar-ified. Indeed, dissociation of their therapeutic effectsand adverse reactions is still one of the most challeng-ing clinical issues to be solved (4, 5).
Glucocorticoids act by the binding to their cognatereceptor, the glucocorticoid receptor (GR), the proto-typic member of the nuclear receptor superfamily,which also includes the receptors for the mineralocor-ticoids (MR), estrogens, progestins (PR), and andro-gens (AR), as well as those for peroxisome prolif-erators, vitamin D, and thyroid hormones (6, 7).Phylogenetic and sequence analysis indicate that theGR, MR, PR, and AR form a subfamily of oxosteroidreceptors (6, 7). Like most nuclear receptors, the GR isa modular protein that is organized into three majordomains: an N-terminal regulatory domain harboring astrong transcriptional activation function (AF)-1, a cen-tral DNA binding domain (DBD), and a C-terminal li-gand binding domain (LBD) (6, 8). The LBD harbors asecond AF-2 directly regulated by ligands. Agonistbinding to the LBD induces the reorientation of a crit-ical �-helix (AF-2 helix) and the formation of a bindingpocket for a family of coactivator proteins that play
First Published Online January 27, 2005Abbreviations: AF, Activation function; AR, androgen re-
ceptor; CVZ, cortivazol; DBD, DNA binding domain; DEX,dexamethasone; GFP, green fluorescent protein; GR, glu-cocorticoid receptor; GRE, glucocorticoid response element;hsp90, heat shock protein 90; LBD, ligand binding domain;MR, mineralocorticoid receptor; NF-�B, nuclear factor-� B;NLS, nuclear localization signal; PR, progesterone receptor;SDS, sodium dodecyl sulfate; TIF, transcriptional intermedi-ary factor.
Molecular Endocrinology is published monthly by TheEndocrine Society (http://www.endo-society.org), theforemost professional society serving the endocrinecommunity.
0888-8809/05/$15.00/0 Molecular Endocrinology 19(5):1110–1124Printed in U.S.A. Copyright © 2005 by The Endocrine Society
doi: 10.1210/me.2004-0264
1110

essential roles in transactivation (9, 10). Among nu-clear receptors, the AF-2 pocket is highly conserved,whereas the molecular size and amino acid composi-tion of AF-1 is much more diverse (6, 9). In the absenceof ligand, the GR is retained in the cytoplasm in asso-ciation with chaperone proteins such as heat shockprotein 90 (hsp90) (11, 12). Hormone binding initiatesthe release of the chaperone proteins and transloca-tion of the receptor into the nucleus where GR binds toDNA promoter elements termed glucocorticoid re-sponse element (GRE) from which it can either activateor repress transcription depending on the context ofthe target promoter (6, 8). In addition, the GR alsointeracts with other transcription factors such as nu-clear factor-�B (NF-�B) to repress their transcriptionalactivities. This GR-mediated repression has been pos-tulated to be one of the major mechanisms for thetherapeutic antiinflammatory and immunosuppressiveactivities of glucocorticoids (13, 14).
Recent crystallographic analyses of the nuclear re-ceptors have clarified the relationship between recep-tor structure and function and established a paradigmof receptor activation. The GR LBD, similar to othernuclear receptor LBDs, is composed of �-helices and�-strands folded into a three-layer helical sandwich.The ligand binding pocket is composed of residuesfrom helices 3, 4, 5, 6, 7, 10, and the AF-2 helix as wellas residues from �-strands between helices 5 and 6.Following AF-2 helix is an extended strand that formsa conserved �-sheet with a �-strand between helices8 and 9. This C-terminal �-strand also appears to playan important role in receptor activation by stabilizingAF-2 helix in an active conformation (15, 16). ManyAF-2 coactivators for the GR have been identified todate, including steroid receptor coactivator-1, tran-scriptional intermediary factor (TIF) 2/GR-interactingprotein-1 and cAMP response element binding pro-tein-binding protein/p300 (17–21). These coactivatorsdirectly associate with the GR LBD via their LXXLLmotif. For example, the LLRYLL sequence in the TIF2forms a two-turn �-helix that orients the hydrophobicleucine side chains into a groove formed in part by theAF-2 helix and residues from helices 3, 3�, 4, and 5.The N- and C-terminal ends of the coactivator helix areclamped by Glu-755 from the AF-2 helix and Lys-579in helix 3, respectively (15). Mutations that disrupteither the first (Glu-755) or the second (Arg-585 andAsp-590) charge clamp dramatically reduce activationmediated by the GR LBD, demonstrating that they arecritical for transactivation function of the GR (15). Onthe other hand, AF-1 coactivators have only recentlybeen described. For example, basal transcription fac-tors including TBP and TFIID have been shown toassociate with the AF-1 of GR (22, 23). TSG101 andDRIP150 have also been reported to interact with GRAF-1 and regulate GR function in a reciprocal manner;GR transcriptional activities are repressed by TSG101but enhanced by DRIP150 (24). These cofactors areshown to interact with distinct regions of AF-1 (22–24).Although we now have at hand a large number of
regulatory proteins that interact directly or indirectlywith the various modular domains of nuclear recep-tors, how ligands differentially regulate the functionalinterplay between them remains poorly understood.
The phenylpyrazologlucocorticoid CVZ is a uniquesynthetic glucocorticoid agonist with complex bindingproperties and is more potent than DEX (25). We pre-viously demonstrated that CVZ selectively binds to theGR but not to the MR and, based on two criteria, weproposed that the functional interaction of CVZ withthe GR LBD is different from that of DEX. Firstly, de-letion of the last 12 amino acids of GR severely com-promises DEX but not CVZ binding and secondly, thepoint mutant L753F, in which Leu-753 in AF-2 is sub-stituted to Phe, can efficiently recruit TIF2 to the LBDwhen bound to CVZ but not when bound to DEX (26).These results prompted us to propose that occupancyof the GR LBD by CVZ might lead to a more stableactive conformation that can tolerate the disruptingeffects of LBD mutations and may have unique effectson the structure and function of the whole GR mole-cule. In the present study, we explore the distinctproperties of CVZ-bound GR by modeling its struc-ture, analyzing the influence of both destabilizing andstabilizing LBD mutations, and probing the role playedby other receptor domains. We provide evidence thatthe CVZ-specific LBD conformation allows efficientTIF2 recruitment to the receptor at least in part throughintrareceptor communication between the LBD andDBD, as well as through collaboration with N-terminalsequences.
RESULTS
Docking Model of CVZ-Bound GR LBD
Because we and others showed that CVZ-bound GRhas distinct functional characteristics when comparedwith DEX-bound GR (see introductory section), we firstexamined whether CVZ could be accommodated intothe classical ligand binding pocket of the GR LBD orwhether a distinct binding mode must be invoked. Forthis purpose, we modeled the three-dimensionalstructure of CVZ-bound GR using the coordinates ofthe crystal structure between the F602S mutant LBDand DEX (15). In brief, we docked CVZ in silico into theligand binding pocket of the GR LBD by superimpos-ing its steroid backbone with that of DEX. The calcu-lated volume of DEX and CVZ is 386 Å3 and 541 Å3,respectively, reflecting that CVZ has a bulky phe-nylpyrazole ring attached to the A-ring of steroid back-bone as well as a C21-acetoxyl group at the D-ring(Fig. 1A). The estimated volume of the ligand bindingpocket of the GR LBD is 600 Å3, which appears to belarge enough for binding either ligand. Energy minimi-zation of the CVZ/GR LBD complex suggested thatfavorable configurations between GR and CVZ couldbe reached by the induced fit mechanism. The result-ant model for CVZ-GR LBD is shown in Fig. 1B. CVZ,
Yoshikawa et al. • LBD-DBD Functional Interactions in the GR Mol Endocrinol, May 2005, 19(5):1110–1124 1111

as well as DEX, is completely enclosed within thebottom half of the GR LBD and spatial position of the�-helices and �-strands of the CVZ-bound GR LBD isalmost identical with that of DEX-bound one, includingthe orientation of helix 12. One or more hydrophobicresidues within the GR protein contact nearly everyatom of the steroid core of DEX and CVZ (Fig. 1C). Inaddition, the model allows for all of the hydrophilicgroups of CVZ to form hydrogen bonds with the pro-tein, which is what is observed in the DEX structure(Fig. 1C). A similar and extensive hydrogen bond net-work between GR and either DEX or CVZ is likely tocontribute to their high affinity binding. Moreover, bothligands make direct contacts with AF-2 helix at Leu-753 and the loop preceding AF-2 helix at Ile-747 andPhe-749 (Fig. 1C). These interactions are likely to sta-bilize the AF-2 helix in the active conformation in CVZ-bound LBD and are consistent with the strong ago-nistic activities of CVZ.
Given the extra volume of CVZ, several differencesbetween the DEX-bound GR LBD complex and ourmodel are also evident. To accommodate the bulkyphenylpyrazole ring and 21-acetoxyl group of CVZ, theposition of the side chain of Arg-611 is shifted outwardof the ligand binding pocket and side chain conforma-tions of Asn-564, Gln-570, Met-604, Leu-608, Met-646, and Phe-749 are altered, resulting in distinct hy-drogen bond network (Fig. 1, C and D). Notably, all ofthese amino acids except for Phe-749 originate fromhelices 3 and 5. Taken together, the modeling sug-gests that CVZ might stably bind with the GR and elicitits distinct effects on GR function via minor local al-teration in the conformation of these helices.
Discrimination of Glucocorticoid Ligands by GRVariants with Destabilizing and StabilizingMutations
We have previously shown that deletion of the last 12amino acids severely compromises DEX binding andDEX-dependent activity but CVZ binding activity ispreserved (26). In contrast to this deleterious deletion,the Phe-602 to Ser substitution in the GR LBD hasbeen shown to stabilize the binding of various ligandsand was instrumental in the successful crystallizationof the GR LBD (15, 16, 27). We therefore hypothesizedthat this substitution might counteract the deleteriouseffects of C-terminal deletion and restore DEX func-tion. We thus introduced this mutation in the context ofthe C-terminally deleted GR-(1–765) (resultant mutantis GR-(1–765)/F602S, Fig. 2A). These mutants shouldbe useful tools to contrast the characteristics of theinteraction between CVZ and DEX with the GR.
Initially, we characterized these mutants biochemi-cally. Because hsp90 is essential for ligand binding,we addressed whether these mutant GR could bindhsp90 in situ (Fig. 2B). To this end, COS7 cells weretransfected with expression plasmids for the wild-typeand mutant GR forms, and cellular lysates were pre-pared in the absence of ligand. Protein expression of
Fig. 1. Putative Three-Dimensional Structure of the GR LBDDocked with DEX and CVZ
A, Structure of DEX and CVZ. B, Overall arrangement ofDEX- and CVZ-bound GR LBD. Ribbons represent �-helicesand �-strands, and tubes represent loops of the protein. TheGR-LBD docked with DEX or CVZ is depicted with space fillmodel. C, Ligand binding pocket of GR-DEX complex super-imposed with that of GR-CVZ one (StereoView 20/20, 3DExperience, Hertfordshire, UK). Structures are drawn withstick model. Amino acid residues of GR-DEX and GR-CVZcomplex are drawn by green and orange sticks, respectively.DEX and CVZ are depicted with blue and red, respectively. D,Amino acid sequence and secondary structure of the GRLBD. Amino acid sequence of the GR LBD and the location of�-helices (H1 to H12, bold bars) and �-strands (�1 to �4,arrows) are schematically illustrated. Number depicts the po-sition of amino acids. Open and filled circles indicate the GRresidues closer than 4.5 Å to DEX and CVZ, respectively.
1112 Mol Endocrinol, May 2005, 19(5):1110–1124 Yoshikawa et al. • LBD-DBD Functional Interactions in the GR
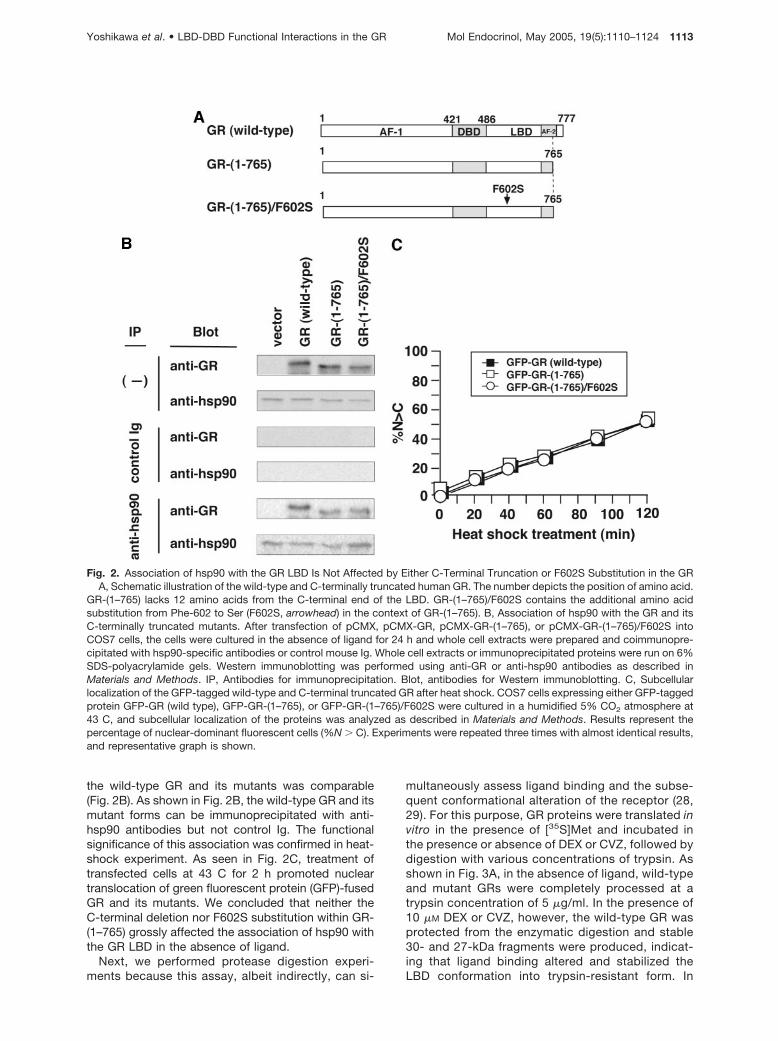
the wild-type GR and its mutants was comparable(Fig. 2B). As shown in Fig. 2B, the wild-type GR and itsmutant forms can be immunoprecipitated with anti-hsp90 antibodies but not control Ig. The functionalsignificance of this association was confirmed in heat-shock experiment. As seen in Fig. 2C, treatment oftransfected cells at 43 C for 2 h promoted nucleartranslocation of green fluorescent protein (GFP)-fusedGR and its mutants. We concluded that neither theC-terminal deletion nor F602S substitution within GR-(1–765) grossly affected the association of hsp90 withthe GR LBD in the absence of ligand.
Next, we performed protease digestion experi-ments because this assay, albeit indirectly, can si-
multaneously assess ligand binding and the subse-quent conformational alteration of the receptor (28,29). For this purpose, GR proteins were translated invitro in the presence of [35S]Met and incubated inthe presence or absence of DEX or CVZ, followed bydigestion with various concentrations of trypsin. Asshown in Fig. 3A, in the absence of ligand, wild-typeand mutant GRs were completely processed at atrypsin concentration of 5 �g/ml. In the presence of10 �M DEX or CVZ, however, the wild-type GR wasprotected from the enzymatic digestion and stable30- and 27-kDa fragments were produced, indicat-ing that ligand binding altered and stabilized theLBD conformation into trypsin-resistant form. In
Fig. 2. Association of hsp90 with the GR LBD Is Not Affected by Either C-Terminal Truncation or F602S Substitution in the GRA, Schematic illustration of the wild-type and C-terminally truncated human GR. The number depicts the position of amino acid.
GR-(1–765) lacks 12 amino acids from the C-terminal end of the LBD. GR-(1–765)/F602S contains the additional amino acidsubstitution from Phe-602 to Ser (F602S, arrowhead) in the context of GR-(1–765). B, Association of hsp90 with the GR and itsC-terminally truncated mutants. After transfection of pCMX, pCMX-GR, pCMX-GR-(1–765), or pCMX-GR-(1–765)/F602S intoCOS7 cells, the cells were cultured in the absence of ligand for 24 h and whole cell extracts were prepared and coimmunopre-cipitated with hsp90-specific antibodies or control mouse Ig. Whole cell extracts or immunoprecipitated proteins were run on 6%SDS-polyacrylamide gels. Western immunoblotting was performed using anti-GR or anti-hsp90 antibodies as described inMaterials and Methods. IP, Antibodies for immunoprecipitation. Blot, antibodies for Western immunoblotting. C, Subcellularlocalization of the GFP-tagged wild-type and C-terminal truncated GR after heat shock. COS7 cells expressing either GFP-taggedprotein GFP-GR (wild type), GFP-GR-(1–765), or GFP-GR-(1–765)/F602S were cultured in a humidified 5% CO2 atmosphere at43 C, and subcellular localization of the proteins was analyzed as described in Materials and Methods. Results represent thepercentage of nuclear-dominant fluorescent cells (%N � C). Experiments were repeated three times with almost identical results,and representative graph is shown.
Yoshikawa et al. • LBD-DBD Functional Interactions in the GR Mol Endocrinol, May 2005, 19(5):1110–1124 1113

contrast to the wild-type GR, the C-terminally de-leted receptor discriminated between the two li-gands because CVZ was able to protect the LBDfrom digestion with 50 and 100 �g/ml of trypsin(generating 25- and 28-kDa fragments), whereasDEX was substantially less effective. Consistent withour hypothesis, addition of the F602S mutationwithin the context of the C-terminal deletion partiallyrestored the ability of DEX to protect against 50 and
100 �g/ml of trypsin. This suggests that this aminoacid substitution enabled DEX to more efficientlybind and stabilize the deleted LBD just as CVZ cando in the absence of the point mutation (Fig. 3A).
These results were further supported in separateexperiments using constructs expressing a fusion pro-tein between the simian virus 40 nuclear localizationsignal (NLS) and the GR LBD alone (Fig. 3B). In theabsence of ligands, in vitro-translated wild-type and
Fig. 3. Effects of DEX and CVZ on Proteolytic Digestion of in Vitro-Translated Wild-Type and C-Terminal Truncated GRSchematic illustration of the wild-type GR, GR-(1–765), and GR-(1–765)/F602S (A), and NLS-fused GR LBD, GRLBD-(499–765),
and GRLBD-(499–765)/F602S (B) are shown in the left. The predicted trypsin-resistant fragments are depicted as bold lines withtheir calculated molecular weights (45, 61). Representative results from trypsin digestion assays are presented in the right. In brief,in vitro-synthesized [35S]GR and its mutants (A), or [35S]GRLBD and its mutants (B) was incubated with either vehicle or 10 �M
of DEX or CVZ for 30 min at 20 C; then the indicated concentrations of trypsin were added and digestion was preceded for 10min at 20 C as described in Materials and Methods. Samples were denatured and separated on 12% SDS-polyacrylamide gels.Gels were processed as described in Materials and Methods for visualization and representative results are shown.
1114 Mol Endocrinol, May 2005, 19(5):1110–1124 Yoshikawa et al. • LBD-DBD Functional Interactions in the GR

mutant LBDs were completely digested (Fig. 3B). Inthe presence of DEX or CVZ, however, trypsin treat-ment of the wild-type LBD generated stable 30- and27-kDa fragments (Fig. 3B). As in the case of thefull-length GR, the C-terminally deleted LBD [LBD-(499–765)] was resistant to trypsin in the presence ofCVZ but not DEX. Moreover, the F602S substitutionenabled DEX to protect this LBD. Taken together,these results indicate that CVZ-bound GR mimics themore stable conformation achieved by DEX in thepresence of the F602S point mutation.
Distinct Effects of DEX and CVZ on the Functionof C-Terminally Truncated GR
We next studied the effect of treatment with DEX andCVZ on nuclear translocation of GR and its mutantsusing GFP-tagged forms. The mutations did not alterthe behavior of the receptor in the absence of ligandsbecause GFP-tagged GR and its mutants are all local-ized in the cytoplasm. In the case of wild-type GFP-GR, both DEX and CVZ promoted efficient nucleartranslocation (Fig. 4A). In contrast, whereas CVZ wasable to induce nuclear translocation of GFP-GR-(1–765), DEX failed to do so (Fig. 4A). Notably, introduc-tion of the F602S mutation restored the ability of DEXto induce the translocation of the C-terminally deletedreceptor and eliminated the difference between theligands (Fig. 4A). These results again suggested thatthe F602S substitution appears to stabilize DEX-bind-ing to GR-(1–765).
To assess the effect of DEX and CVZ on the trans-activation function of wild-type and mutant GRs, weused a GRE-driven luciferase reporter plasmid inCOS7 cells. As previously reported, DEX and CVZinduced GRE-dependent transactivation by the wild-type GR, whereas reporter gene activation by GR-(1–765) was only observed for CVZ (Fig. 4B). Surprisingly,although DEX and CVZ were able to induce nucleartranslocation and DNA binding activities of GR-(1–765)/F602S (Fig. 4A and data not shown), DEXscarcely induced transactivation even at high concen-trations, whereas CVZ elicited gene activation in aconcentration-dependent manner (Fig. 4B). Because ithas been shown that ligand binding affects stability ofthe GR (30), we examined GR protein levels after treat-ment with DEX or CVZ. As previously reported, treat-ment with DEX or CVZ decreased the protein levels ofthe GFP-fused full-length GR (Fig. 4C). In clear con-trast, DEX treatment did not lead to reduced proteinlevels of either GFP-GR-(1–765) or GFP-GR-(1–765)/F602S (Fig. 4C). Moreover, CVZ showed only marginaldecrease in the protein levels of these mutant GRs(Fig. 4C). These results reveal a role for the C-terminal12 amino acids in down-regulation of the receptor butindicate that the differential effects of DEX and CVZ onGR-mediated reporter gene activation is not simplydue to ligand-induced decrease of GR protein stability.The striking difference in transactivation betweenDEX- and CVZ-bound GR mutants coupled with our
structural prediction that CVZ-bound LBD might leadto only minor conformational alteration in the LBDwhen compared with DEX suggests that such subtleconformational changes might affect ligand-depen-dent cofactor recruitment to the receptor, especiallybecause it has already been reported that in additionto AF-2, N-terminal segments of the LBD are importantfor stable interaction with coactivators (15, 31). Wetherefore examined the effect of exogenous expres-sion of TIF2 on the ability of the various GR mutants toinduce transcription in response to DEX and CVZ.Overexpression of TIF2-enhanced CVZ-inducedtransactivation by the wild-type GR as well as GR-(1–765) and GR-(1–765)/F602S (Fig. 5B). On the otherhand, TIF2 only marginally rescued DEX-dependenttransactivation by either mutant (Fig. 5B).
To test the possibility that TIF2 recruitment is influ-enced by sequences within the N-terminal domain ofthe receptor, we exchanged this region of the GR andits mutants with those of the MR and PR (Fig. 5A).Deletion of AF-1 (GR�AF-1) resulted in a 60% de-crease in activity, which was still enhanced by TIF2.As previously reported (32), replacement of the N-terminal region of the GR with that of the MR didnot significantly enhance activity compared withGR�AF-1. Enhancement of CVZ-mediated transacti-vation by TIF2 was observed in M/GG/765 and M/GG/765/F602S as seen in GR�AF-1 and M/GG. The N-terminal region of PR functionally substituted for thatof GR and TIF2 enhancement is observed for bothP/GG/765 and P/GG/765/F602S (Fig. 5B). These re-sults argue that although AF-1 cooperates with AF-2,it does not play a critical role in CVZ-mediated recruit-ment of TIF2 to the receptor.
To confirm the effects of these ligands on the inter-action between the GR and TIF2 in situ, we performedimmunofluorescence colocalization assays in whichGFP-GR and TIF2 were coexpressed. As previouslyreported (26), wild-type GR displace a speckled local-ization in the nucleus in the presence of TIF2 (Fig. 5Cand data not shown). Consistent with the transactiva-tion data, CVZ induced this speckled pattern for bothGR-(1–765) and GR-(1–765)/F602S, whereas DEXtreatment failed to translocate GR-(1–765) and re-sulted in a diffuse nuclear localization of GR-(1–765)/F602S (Fig. 5C). Thus, although DEX-bound GR-(1–765)/F602S accumulates in the nucleus, it does notappear to form transcriptionally productive complexwith TIF2.
Ligand-Dependent Scenario for Communicationbetween DBD and the LBD
To further delineate the receptor domains involved inthe differential properties of DEX- vs. CVZ-bound GR,we next examined the ability of CVZ to counteract thedeleterious effects of the C-terminal deletion and sup-port the recruitment of TIF2 in the context of the GRLBD alone. To test this, we constructed plasmids forthe expression of GAL4 DBD-GR LBD fusion proteins
Yoshikawa et al. • LBD-DBD Functional Interactions in the GR Mol Endocrinol, May 2005, 19(5):1110–1124 1115

Fig. 4. C-Terminal Truncation of the LBD with F602S Substitution Unveils Differential Effects of DEX and CVZ on TransactivationFunction of the GR
A, Effects of ligands on subcellular localization of the GR and its C-terminally truncated mutants. GFP-tagged wild-type GR,GR-(1–765), and GR-(1–765)/F602S were transiently expressed in COS7 cells and the cells were cultured in the presence orabsence of 1 �M of DEX or CVZ for 2 h, then digital images were taken as described in Materials and Methods and representativeresults are shown. B, Differential effects of DEX and CVZ on GRE-driven reporter gene expression. COS7 cells were cotransfectedwith 2 �g of GRE-driven reporter plasmid pGRE-LUC and 100 ng of expression plasmids for either wild-type GR, GR-(1–765), orGR-(1–765)/F602S, and cultured in the absence or presence of the indicated concentrations of DEX or CVZ for 24 h. Cellularluciferase activities were measured as described in Materials and Methods. Experiments were performed in triplicate and resultsare expressed as relative light units (RLU) per microgram of protein in the extract, and the means � SD are shown. C, Effects ofDEX and CVZ on receptor protein levels. COS7 cells were transiently transfected with expression plasmids for GFP-tagged
1116 Mol Endocrinol, May 2005, 19(5):1110–1124 Yoshikawa et al. • LBD-DBD Functional Interactions in the GR

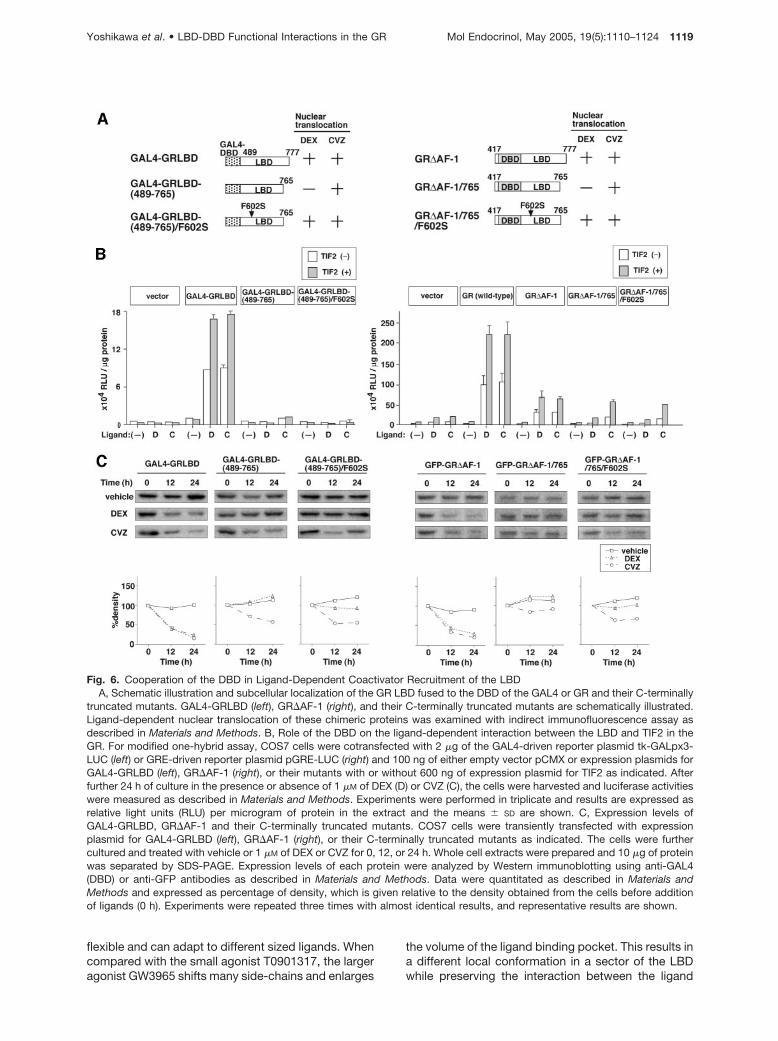
[GAL4-GRLBD, GAL4-GRLBD-(489–765), and GAL4-GRLBD-(489–765)/F602S; see Fig. 6A], and testedthem in a mammalian one-hybrid assay using GAL4-responsive reporter plasmid in COS7 cells. Nucleartranslocation assays paralleled the observations usingthe intact GR and confirmed that CVZ could achievenuclear entry of all the chimeric proteins, whereas DEXinduced the translocation of only GAL4-GRLBD andGAL4-GRLBD-(489–765)/F602S (Fig. 6A). The GAL4-GRLBD chimera activated the reporter gene in re-sponse to both DEX and CVZ and heterologous ex-pression of TIF2 enhanced this response, indicatingthat binding of either ligand facilitated the recruitmentof TIF2 to the LBD alone. Deletion of the last 12 aminoacids of the LBD severely compromised CVZ as wellas DEX induction even in the presence of TIF2 or aftercombining it with the F602S substitution (Fig. 6B).Both ligands induced substantial down-regulation ofthe wild-type LBD fusion chimera, whereas the levelsof GAL4-GRLBD-(489–765) and GAL4-GRLBD-(489–765)/F602S were mildly affected or unchanged (Fig.6C). These results indicate that the deleterious effectsof the C-terminal truncation cannot be counteractedby CVZ binding or the F602S substitution in the con-text of the LBD alone and suggest that additionaldomains are required.
To examine the involvement of the GR DBD, wecarried out parallel one-hybrid assays in the context ofN-terminally deleted GR�AF-1 construct where theLBD is recruited to the promoter through the GR’s ownDBD. As in the case of the GAL4 fusions, CVZ couldachieve nuclear entry of all three mutant GR forms,whereas DEX could do so only for GR�AF-1 andGR�AF-1/765/F602S (Fig. 6A). In clear contrast to theGAL4 fusion chimeras, CVZ was able to support trans-activation after deletion of the last 12 amino acids andTIF2-mediated enhancement was preserved (Fig. 6B).Because the behavior of these N-terminally deletedforms resembled that of the full-length receptor (in-cluding the ligand-induced changes in protein level;see Fig. 6C), the results indicate that the DBD of GR issufficient to support the ability of CVZ to counteractthe deleterious effects of the C-terminal deletion andthat productive recruitment of TIF2 in this contextlikely involves a ligand-based functional communica-tion between the LBD and the DBD.
The Functional Differences between CVZ andDEX Are Also Evident during GR Transrepressionof NF-�B
To examine whether the induced structural change ofthe LBD upon CVZ binding affects other receptor func-
tions, we tested the ability of both DEX and CVZ tosupport GR mediated transrepression of NF-�B-stimulated transcription. Although the mechanism ofNF-�B repression by the GR remains to be fully de-fined, it is known that the p65 subunit of NF-�B phys-ically interact with the GR DBD (33–37). Wild-type GRsuppressed NF-�B-dependent transcription in re-sponse to both DEX and CVZ (Fig. 7). Deletion of thelast 12 amino acids again revealed a marked differ-ence between ligands because CVZ-mediated repres-sion was preserved, whereas the DEX response wasessentially abrogated. As in the case of activation,inclusion of the F602S substitution did not restoreDEX-mediated inhibition (Fig. 7). These results sug-gest that CVZ differentially modulates receptor func-tion not only in transactivation but also in transrepres-sion, likely through the establishment of a distinct LBDconformation and functional interaction with the DBD.
DISCUSSION
In the present study, we have explored the mode ofbinding of the unique glucocorticoid agonist CVZ andhave taken advantage of C-terminally deleted GR vari-ants to unveil otherwise hidden differences betweenCVZ and the prototypic agonist DEX. Ligand binding isbelieved to give the receptor a cue for activation by theadoption of a conformation that is conducive to inter-action with coactivator proteins (38). Data from multi-ple members of this family indicate that the chemicalstructure of individual agonists can produce distinctconformations of the LBD with unique regulatory prop-erties (9, 39). Given bulky phenylpyrazole substituentat the A-ring of CVZ, it is thus reasonable to hypoth-esize that CVZ-bound GR is likely to have a distinctstructure when compared with the DEX-bound one.According to our modeling data, CVZ could fit in theligand binding pocket through an induced fit mecha-nism (Fig. 1). The additional volume contributed by thephenylpyrazole ring of CVZ provides additional con-tacts with the protein and can be accommodatedthrough minor alterations in the receptor structure in-volving the reorientation of amino acid side chainsemerging mainly from helices 3 and 5. These alter-ations may result in the generation of a more stablebound conformation and distinct functional surfaces.Although it is obvious that the direct determination ofthe structure of CVZ-bound LBD is required to deter-mine the actual mode of binding, data from existingstructures are compatible with the model. In the caseof the liver X receptor �, the structure of the LBD is
wild-type GR, GR-(1–765), or GR-(1–765)/F602S. The cells were further cultured and treated with vehicle or 1 �M of DEX or CVZfor 0, 12, or 24 h. Whole cell extracts were prepared and 10 �g of protein was separated by SDS-PAGE. Expression levels of eachprotein were analyzed by Western immunoblotting using anti-GFP antibodies as described in Materials and Methods. Data werequantitated and expressed as percentage of density, which was given relative to the density obtained from the cells treated witheach ligand for 0 h as described in Materials and Methods. Experiments were repeated three times with almost identical results,and representative results are shown.
Yoshikawa et al. • LBD-DBD Functional Interactions in the GR Mol Endocrinol, May 2005, 19(5):1110–1124 1117
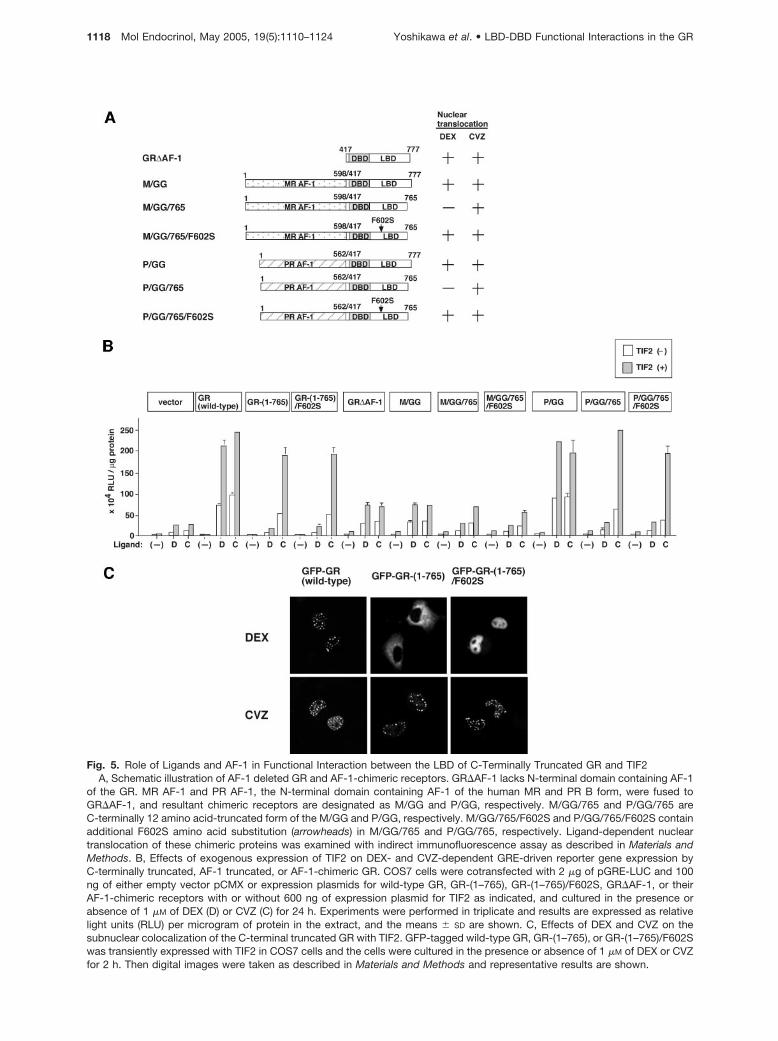
Fig. 5. Role of Ligands and AF-1 in Functional Interaction between the LBD of C-Terminally Truncated GR and TIF2A, Schematic illustration of AF-1 deleted GR and AF-1-chimeric receptors. GR�AF-1 lacks N-terminal domain containing AF-1
of the GR. MR AF-1 and PR AF-1, the N-terminal domain containing AF-1 of the human MR and PR B form, were fused toGR�AF-1, and resultant chimeric receptors are designated as M/GG and P/GG, respectively. M/GG/765 and P/GG/765 areC-terminally 12 amino acid-truncated form of the M/GG and P/GG, respectively. M/GG/765/F602S and P/GG/765/F602S containadditional F602S amino acid substitution (arrowheads) in M/GG/765 and P/GG/765, respectively. Ligand-dependent nucleartranslocation of these chimeric proteins was examined with indirect immunofluorescence assay as described in Materials andMethods. B, Effects of exogenous expression of TIF2 on DEX- and CVZ-dependent GRE-driven reporter gene expression byC-terminally truncated, AF-1 truncated, or AF-1-chimeric GR. COS7 cells were cotransfected with 2 �g of pGRE-LUC and 100ng of either empty vector pCMX or expression plasmids for wild-type GR, GR-(1–765), GR-(1–765)/F602S, GR�AF-1, or theirAF-1-chimeric receptors with or without 600 ng of expression plasmid for TIF2 as indicated, and cultured in the presence orabsence of 1 �M of DEX (D) or CVZ (C) for 24 h. Experiments were performed in triplicate and results are expressed as relativelight units (RLU) per microgram of protein in the extract, and the means � SD are shown. C, Effects of DEX and CVZ on thesubnuclear colocalization of the C-terminal truncated GR with TIF2. GFP-tagged wild-type GR, GR-(1–765), or GR-(1–765)/F602Swas transiently expressed with TIF2 in COS7 cells and the cells were cultured in the presence or absence of 1 �M of DEX or CVZfor 2 h. Then digital images were taken as described in Materials and Methods and representative results are shown.
1118 Mol Endocrinol, May 2005, 19(5):1110–1124 Yoshikawa et al. • LBD-DBD Functional Interactions in the GR
flexible and can adapt to different sized ligands. Whencompared with the small agonist T0901317, the largeragonist GW3965 shifts many side-chains and enlarges
the volume of the ligand binding pocket. This results ina different local conformation in a sector of the LBDwhile preserving the interaction between the ligand
Fig. 6. Cooperation of the DBD in Ligand-Dependent Coactivator Recruitment of the LBDA, Schematic illustration and subcellular localization of the GR LBD fused to the DBD of the GAL4 or GR and their C-terminally
truncated mutants. GAL4-GRLBD (left), GR�AF-1 (right), and their C-terminally truncated mutants are schematically illustrated.Ligand-dependent nuclear translocation of these chimeric proteins was examined with indirect immunofluorescence assay asdescribed in Materials and Methods. B, Role of the DBD on the ligand-dependent interaction between the LBD and TIF2 in theGR. For modified one-hybrid assay, COS7 cells were cotransfected with 2 �g of the GAL4-driven reporter plasmid tk-GALpx3-LUC (left) or GRE-driven reporter plasmid pGRE-LUC (right) and 100 ng of either empty vector pCMX or expression plasmids forGAL4-GRLBD (left), GR�AF-1 (right), or their mutants with or without 600 ng of expression plasmid for TIF2 as indicated. Afterfurther 24 h of culture in the presence or absence of 1 �M of DEX (D) or CVZ (C), the cells were harvested and luciferase activitieswere measured as described in Materials and Methods. Experiments were performed in triplicate and results are expressed asrelative light units (RLU) per microgram of protein in the extract and the means � SD are shown. C, Expression levels ofGAL4-GRLBD, GR�AF-1 and their C-terminally truncated mutants. COS7 cells were transiently transfected with expressionplasmid for GAL4-GRLBD (left), GR�AF-1 (right), or their C-terminally truncated mutants as indicated. The cells were furthercultured and treated with vehicle or 1 �M of DEX or CVZ for 0, 12, or 24 h. Whole cell extracts were prepared and 10 �g of proteinwas separated by SDS-PAGE. Expression levels of each protein were analyzed by Western immunoblotting using anti-GAL4(DBD) or anti-GFP antibodies as described in Materials and Methods. Data were quantitated as described in Materials andMethods and expressed as percentage of density, which is given relative to the density obtained from the cells before additionof ligands (0 h). Experiments were repeated three times with almost identical results, and representative results are shown.
Yoshikawa et al. • LBD-DBD Functional Interactions in the GR Mol Endocrinol, May 2005, 19(5):1110–1124 1119

and helix 12 intact (40). In our model, the bulky A-ringof CVZ intimately contacts with and modulates theconformation of helices 3 and 5 (Fig. 1). Along this line,Ali et al. (41) have reported on compounds that have aphenylpyrazole group at the A-ring. Replacement ofthe C- and D-ring of the steroid backbone by alkyl,alkenyl, or benzyl groups yields compounds that se-lectively bind to the GR and induce transactivation(41). As in the case of CVZ and DEX, when the phe-nylpyrazole group in such compounds is replaced by aC3-ketone, their GR binding ability is completely abol-ished (41). This suggests that the mode of bindingbetween these compounds is similar and is consistentwith the idea that contacts between the phenylpyra-zole A-ring and helices 3 and 5 are compatible with astable active conformation. Interestingly, a similarmodeling and functional approach in the MR arguesthat the Ser-810 to Leu substitution in helix 5 of theMR, which renders progesterone and the antagonistspironolactone into an agonist, is due to the creationof new interactions between these ligands and helices3 and 5 (42). Concerning the GR, the conformation ofhelix 3 is likely to be important for agonism becauseLys-579 within this helix is involved in clamping theC-terminal end of the coactivator helix and mutation ofthis residue reduces GR transactivation without affect-ing ligand binding (15, 31).
The C-terminal end of the GR LBD forms an ex-tended �-sheet that is important for binding of certainagonists and stable interaction with coactivators (15,16, 38, 43). Deletion of the C-terminal 12 amino acidsfrom GR severely compromises the ability of DEX tobind, to induce a trypsin-resistant conformation of theLBD, and to support transactivation. Because this de-letion does not extend into the AF-2 helix and pre-serves the essential constituents of the AF-2 pocket,the observations suggest that the role of these resi-dues is to stabilize an active conformation of the re-
ceptor. Our current results indicate that the deleteriouseffects of this deletion can be surmounted when CVZoccupies the binding pocket. This indicates that thedeleted receptor possesses all the necessary featuresfor an active conformation as long as an appropriateligand is bound. In this view, it is likely that the addi-tional contacts provided by the bulky phenylpyrazolesubstituent yield a stable active conformation withoutthe contributions provided by the C-terminal residues.The fact that the stabilizing F602S substitution canrestore the ability of DEX to bind and induce the nu-clear translocation of the C-terminally deleted GR isalso consistent with CVZ stabilizing an active confor-mation. The fact that DEX-dependent transcriptionalactivity of the deletion mutants is not restored by theF602S mutation suggests that, in this context, thestabilizing influence of the F602S is not sufficient andthat conformations other than an active one are fa-vored. A similar behavior is also observed in the caseof the antagonist RU486 because it is capable of bind-ing and inducing the nuclear translocation of GR evenafter deletion of the last 28 C-terminal amino acids (44,45). Our results, thus, clearly show that the C-terminal12 amino acids are not absolutely essential for ago-nistic activity and highlight the critical role played bythe ligand in sculpting the functional surfaces of thereceptor.
The unique ability of CVZ to support transactivationby the C-terminally deleted receptor allowed us toexamine the role played by domains other than theLBD. Our results indicate that, at least in terms ofligand-dependent transactivation, the N-terminal re-gion of the receptor is not essential but contributes tooverall activity, most likely through cooperation be-tween AF-1 and AF-2 (46). More importantly, directtargeting of the C-terminally deleted LBD to the DNAvia the GAL4 DBD does not support CVZ-driven ac-tivity and TIF-2 recruitment. In contrast, when the de-
Fig. 7. CVZ Elicits Transrepression Function of the GR Despite the Lack of the C-Terminal EndHeLa cells were cotransfected with 2 �g of pNF�BHL reporter plasmid and 1 �g of either empty vector pCMX or expression
plasmids for the wild-type GR, GR-(1–765), or GR-(1–765)/F602S. The cells were cultured and treated with or without 10 nM
phorbol 12-myristate acetate (PMA) in the presence or absence of 1 �M of DEX or CVZ for 24 h. Assays were performed intriplicate and results are expressed as percentage of induction, which is given relative to the luciferase activity obtained from thecells treated with PMA alone, and the means � SD are shown.
1120 Mol Endocrinol, May 2005, 19(5):1110–1124 Yoshikawa et al. • LBD-DBD Functional Interactions in the GR

leted LBD is brought to the promoter by the GR’s ownDBD, CVZ is able to support transactivation and TIF-2recruitment. Together with the protease digestionstudies, these results indicate that, although CVZ canpromote a trypsin-resistant conformation of the C-terminally deleted LBD alone, the transcriptional ef-fects of CVZ require the DBD of GR. The unique role ofthe GR DBD might involve an appropriate orientationof the two LBDs upon DNA binding-induced dimeriza-tion or more likely, reflects a direct contribution of theDBD to the active conformation of the CVZ-boundLBD. Several laboratories have shown that the DBDstabilizes the LBD and enhances ligand-dependentnuclear translocation (47–50). Moreover, ligand-drivenconformations of the LBD can influence both DNAbinding (51) and anti-NF-�B activities (52). Kumar et al.(53) have suggested that the DBD plays an importantrole in the structural stabilization of the GR. Impor-tantly, interdomain communication is not exclusive toGR and has been observed in other receptors as well,especially in AR (54). Steroid pharmacology is increas-ingly focused in the development of ligands with se-lective modulatory activities. Because the mode ofinterdomain communication may be distinct for eachreceptor and may be modulated in a ligand-, tissue-,and promoter-context-dependent manner, ligandssuch as CVZ and other phenylpyrazole analogs thatmanipulate this regulatory avenue will not only providea better understanding of the mechanisms of interdo-main communication but also provide novel leads inthe development of selective GR modulators.
MATERIALS AND METHODS
Reagents and Antibodies
DEX was purchased from Sigma (St. Louis, MO). CVZ was akind gift from Aventis Pharma (Strasbourg, France). Otherchemicals were obtained from Wako Pure Chemical (Osaka,Japan) unless otherwise specified. Monoclonal anti-hsp90antibodies were obtained from Affinity Bioreagents, Inc.(Golden, CO). Goat antimouse IgM and control mouse IgMTEPC183 were obtained from Sigma. Monoclonal anti-GFPantibodies were obtained from CLONTECH (Palo Alto, CA).Polyclonal anti-GAL4 (DBD) and antipolyhistidine antibodieswere obtained from Santa Cruz Biotechnologies (SantaCruz, CA).
Plasmids
The expression plasmids for the wild-type and C-terminal 12amino acid-truncated human GR, pCMX-GR, and pCMX-GR-(1–765) have been previously described (26). Constructionof pCMX-GR-(1–765)/F602S was carried out using theQuikChange site-directed mutagenesis kit (Stratagene, LaJolla, CA) using pCMX-GR-(1–765) as a template. The ex-pression plasmids for the fusion between the simian virus 40NLS and the human GR LBD (amino acids 499–777) and forthe fusion of GFP and wild-type human GR, pCMX-NLS-GRLBD and pCMX-GFP-GR have been previously described(55, 56). The expression plasmid for the chimera between theGAL4 DBD and the human GR LBD (amino acids 489–777),pCMX-GAL4-GRLBD, was a kind gift of Dr. K. Umesono
(Kyoto University, Kyoto, Japan). To construct the expressionplasmid for GFP- or polyhistidine-tagged AF-1-deleted hu-man GR (amino acids 417–777), pCMX-GFP-GR�AF-1 orpCMX-GR�AF-1, the DNA fragments encoding human GRDBD and LBD were inserted into the parent pCMX-GFP orpCMX-6xHis vectors, respectively. To construct the expres-sion plasmids for chimeric proteins of the AF-1 of either thehuman MR (amino acids 1–598) or PR B form (amino acids1–562) and GR�AF-1 (resultant plasmids are pCMX-MRAF-1/GR�AF-1 and pCMX-PRBAF-1/GR�AF-1, respectively),the N-terminal domain of each receptor was amplified byPCR using pRShMR or pEGFP-PRB as templates with ap-propriate flanking sequences [these template plasmids werea kind gift of Dr. R. M. Evans (Salk Institute, La Jolla, CA) andDr. G. L. Hager (National Cancer Institute, Bethesda, MD),respectively], and inserted into the parent pCMX-GR�AF-1.To exchange the mutations within the LBD, PstI-BamHI frag-ments encoding a part of the human GR LBD (amino acids596–765) from pCMX-GR-(1–765) or pCMX-GR-(1–765)/F602S were inserted into the same sites of the recipientexpression plasmids. The expression plasmid for TIF2 pSG5-TIF2 was kindly provided by Dr. P. Chambon (Institut deGenetique et de Biologie Moleculaire et Cellulaire, Stras-bourg, France). The GRE-driven reporter plasmid pGRE-LUC, GAL4-responsive reporter plasmid tk-GALpx3-LUC,and NF-�B-responsive reporter plasmid pNF�BHL have beendescribed previously (57).
Cell Culture and Heat Shock Treatment
COS7 and HeLa cells were obtained from the RIKEN CellBank (Tsukuba, Japan) and maintained in DMEM (Sigma)supplemented with 10% fetal calf serum and antibiotics. In allexperiments, serum steroids were stripped with dextran-coated charcoal, and cells were cultured in a humidifiedatmosphere at 37 C with 5% CO2. Heat shock treatment forCOS7 cells was achieved by shifting flasks to another 5%CO2 incubator set at 43 C.
Graphical Manipulations and Ligand Docking
Graphical manipulations were performed using SYBYL 6.9(Tripos, St. Louis, MO). The atomic coordinates of the crystalstructure of human GR LBD (amino acids 523–777) wereretrieved from Protein Data Bank (entry 1M2Z) (15). Wedocked CVZ into the ligand binding pocket manually by su-perimposing its steroid backbone with that of DEX (58–60).Energy minimization of CVZ/GR-LBD complex was per-formed until the energy gradient was lower than 0.1 kcal/(mol)(Å) on Tripos force field by using subset minimizationcommand.
Immunoprecipitation and Western Immunoblot Assay
For analysis of the interaction between the GR and hsp90, wetransiently transfected expression plasmids for wild-type GRor its mutants in COS7 cells and the assays were performedas described previously (56). In brief, whole cell extracts wereprepared by lysing cells and immunoprecipitating with eitherthe anti-hsp90 IgM antibody 3G3 or control mouse IgMantibody TEPC 183 as follows. We first prepared goat anti-mouse IgM coupled to CNBr-activated Sepharose 4B(Amersham Biosciences, Piscataway, NJ) as described pre-viously (56). Seventy micrograms of cellular protein wasadded to the goat antimouse IgM-coupled Sepharose. Thereaction mixtures were incubated on ice for 90 min, afterwhich Sepharose beads were pelleted by centrifugation andwashed three times with MENG buffer [25 mM Mops (pH 7.5),1 mM EDTA, 0.02% NaN3, 10% glycerol] containing 20 mM
sodium molybdate and 2 mM dithiothreitol. Immunoprecipi-tated proteins were eluted by boiling in sample buffer and
Yoshikawa et al. • LBD-DBD Functional Interactions in the GR Mol Endocrinol, May 2005, 19(5):1110–1124 1121

analyzed by SDS-PAGE and electrophoretically transferredto an Immobilon-NC Pure nitrocellulose membrane (Millipore,Bedford, MA). Subsequently, Western immunoblot analysiswas performed with polyclonal anti-GR antibodies diluted at1:1000, followed by horseradish peroxidase-conjugated an-tirabbit Ig (Amersham Biosciences) diluted at 1:2000. Afterstripping off the immune complexes, the same membranewas probed for detection of hsp90, using monoclonal mouseanti-hsp90 IgG antibodies 3B6 (1:500), followed by horserad-ish peroxidase-conjugated antimouse Ig diluted at 1:1000. Inparallel, 20 �g of whole cell extracts were independentlyused for immunodetection of wild-type and mutant GR orhsp90. Antibody-protein complexes were visualized usingthe enhanced chemiluminescence method according to themanufacturer’s protocol (Amersham Biosciences).
Visualization of GFP Fusion Proteins
For analysis of subcellular localization of the human GR andits mutants, we transiently expressed GFP-tagged receptorsin COS7 cells and assays were performed as described pre-viously (26). Briefly, after 6 h of transient transfection of theexpression plasmids for GFP-fusion proteins, the mediumwas replaced with phenol red-free DMEM supplemented with2% dextran-coated charcoal-treated fetal calf serum, and thecells were cultured at 37 C for at least 24 h. After varioustreatments, cells were examined using an IX70 microscope(Olympus, Tokyo, Japan) enclosed by an incubator andequipped with a heating-stage and a fluorescein isothiocya-nate filter set. Digital images were randomly taken in eightviews and analyzed on FLUOVIEW FV 500 systems(Olympus).
Limited Proteolysis Assay
The expression plasmids for the GR and its mutants, whichcontain the coding sequences under control of the T7 pro-moter, were transcribed and translated with the TNTT7-cou-pled reticulocyte lysate system (Promega Corp., Madison,WI) in the presence of [35S]Met (1000 Ci/mmol, AmershamBiosciences) according to the manufacturer’s instruction.Three microliters of [35S]Met-labeled translation mixtures in-cluding in vitro-translated GR were incubated for 30 min at 20C with 1 �l of vehicle (0.4% ethanol) or 10 �M of DEX or CVZ.Limited proteolysis was performed by the addition of 1 �l oftrypsin solution to the translation mixtures (final trypsin con-centrations were 5–100 �g/ml). Digestion was conducted for10 min at 20 C and stopped by cooling in ice, followed by theaddition of 5 �l of sodium dodecyl sulfate (SDS) samplebuffer and boiling for 5 min. The proteolysis products wereseparated on a 1.5-mm thick 12% SDS-polyacrylamide gels.After electrophoresis, the gels were vacuum-dried for 60 minat 80 C and autoradiographed.
Transfection and Reporter Gene Assay
Cells were plated on 6-cm diameter culture dishes (IwakiGlass, Chiba, Japan) to 30–50% confluence and cell culturemedium was replaced with Opti-MEM lacking phenol red(Invitrogen, Carlsbad, CA) before transfection. Plasmid cock-tail was mixed with TransIT-LT1 transfection reagent (Pan-vera Corp., Madison, WI) and added to the culture. Totalamount of the plasmids was kept constant by adding anirrelevant plasmid (pGEM3Z was used unless otherwisespecified). After 6 h of incubation, the medium was replacedwith fresh DMEM supplemented with 2% dextran-coatedcharcoal-treated fetal calf serum, and the cells were culturedin the presence or absence of various ligands for 24 h at 37C. Luciferase enzyme activity was determined using a lumi-nometer (Promega) essentially as described (26).
Indirect Immunofluorescence Assay
For assessment of subcellular localization of chimeric GRproteins, indirect immunofluorescence assay was performedas described previously (26). After transfection of expressionplasmids for various GR mutants into COS7 cells, the cellswere grown on eight-chambered sterile glass slides (NipponBecton Dickinson, Tokyo, Japan) for 24 h and were treatedwithout or with l �M of DEX or CVZ for 2 h. The cells werefixed in ice-cold acetone for 2 min and air-dried. After fixation,the cells were washed with PBS and incubated with anti-GRpolyclonal rabbit antibody at a dilution of 1:100 in PBS con-taining 0.1% Triton X-100 for 1 h at 37 C. Then, the cells werewashed three times with PBS and incubated with fluoresceinisothiocyanate-conjugated antirabbit IgG (Santa Cruz Bio-technology) at a dilution of 1:200 in PBS containing 0.1%Triton X-100 for 1 h at 37 C. The cells were finally washedthree times with PBS and mounted with GEL/MOUNT(Biomeda Corp., Foster City, CA) for examination on a con-focal laser-scanning microscope IX70. Digital images wererandomly taken in eight views and analyzed on FLUOVIEW FV500 systems.
Quantitative Analysis of Chimeric Proteins
For determination of expression levels of GFP-, GAL4-, andpolyhistidine-tagged proteins, we transiently expressed eachchimeric protein in COS7 cells and the cells were cultured inthe presence or absence of ligands for 0, 12, or 24 h. Aftervarious treatments, whole cell extracts were prepared and 10�g of proteins ware separated by SDS-PAGE and transferredto nitrocellulose membranes. Subsequently, Western immu-noblot analysis was performed with anti-GFP, -GAL4 (DBD),and -polyhistidine antibodies followed by appropriate sec-ondary horseradish peroxidase-conjugated antibodies. Anti-body-protein complexes were visualized using the enhancedchemiluminescence method. Expression levels of each chi-meric protein were quantified by scanning the blot and usingimage analysis software from the National Institutes of Health(NIH Image 1.62).
Acknowledgments
We thank Drs. H. Ogawa, P. Chambon, R. M. Evans, G. L.Hager, and K. Umesono for plasmids and the members ofMorimoto laboratory for fruitful discussions and help.
Received June 30, 2004. Accepted January 20, 2005.Address all correspondence and requests for reprints to:
Hirotoshi Tanaka, M.D., Ph.D., the Division of the ClinicalImmunology, the Advanced Clinical Research Center, theInstitute of Medical Science, the University of Tokyo, 4-6-1,Shirokanedai, Minato-ku, Tokyo 108-8639, Japan. E-mail:[email protected].
This work was supported in part by the grants from theMinistry of Education, Science, Technology, Sports, and Cul-ture, the Ministry of Health, Labour, and Welfare, the TakedaScience Foundation, the Uehara Memorial Foundation, theVehicle Racing Commemorative Foundation, and Japan So-ciety for the Promotion of Science.
REFERENCES
1. Sapolsky RM, Romero LM, Munck AU 2000 How doglucocorticoids influence stress responses? Integratingpermissive, suppressive, stimulatory, and preparative ac-tions. Endocr Rev 21:55–89
1122 Mol Endocrinol, May 2005, 19(5):1110–1124 Yoshikawa et al. • LBD-DBD Functional Interactions in the GR

2. Reichardt HM, Tronche F, Berger S, Kellendonk C,Schutz G 2000 New insights into glucocorticoid and min-eralocorticoid signaling: lessons from gene targeting.Adv Pharmacol 47:1–21
3. Adcock IM 2003 Glucocorticoids: new mechanisms andfuture agents. Curr Allergy Asthma Rep 3:249–257
4. Resche-Rigon M, Gronemeyer H 1998 Therapeutic po-tential of selective modulators of nuclear receptor action.Curr Opin Chem Biol 2:501–507
5. Miner JN 2002 Designer glucocorticoids. Biochem Phar-macol 64:355–361
6. Mangelsdorf DJ, Thummel C, Beato M, Herrlich P,Schutz G, Umesono K, Blumberg B, Kastner P, Mark M,Chambon P, Evans RM 1995 The nuclear receptorsuperfamily: the second decade. Cell 83:835–839
7. Nuclear Receptors Nomenclature Committee 1999 A uni-fied nomenclature system for the nuclear receptor super-family. Cell 97:161–163
8. Beato M, Herrlich P, Schutz G 1995 Steroid hormonereceptors: many actors in search of a plot. Cell 83:851–857
9. Bourguet W, Germain P, Gronemeyer H 2000 Nuclearreceptor ligand-binding domains: three-dimensionalstructures, molecular interactions and pharmacologicalimplications. Trends Pharmacol Sci 21:381–388
10. Rosenfeld MG, Glass CK 2001 Coregulator codes oftranscriptional regulation by nuclear receptors. J BiolChem 276:36865–36868
11. Pratt WB, Toft DO 1997 Steroid receptor interactionswith heat shock protein and immunophilin chaperones.Endocr Rev 18:306–360
12. Freeman BC, Yamamoto KR 2001 Continuous recycling:a mechanism for modulatory signal transduction. TrendsBiochem Sci 26:285–290
13. McKay LI, Cidlowski JA 1999 Molecular control of im-mune/inflammatory responses: interactions between nu-clear factor-� B and steroid receptor-signaling pathways.Endocr Rev 20:435–459
14. De Bosscher K, Vanden Berghe W, Haegeman G 2003The interplay between the glucocorticoid receptor andnuclear factor-�B or activator protein-1: molecularmechanisms for gene repression. Endocr Rev 24:488–522
15. Bledsoe RK, Montana VG, Stanley TB, Delves CJ, ApolitoCJ, McKee DD, Consler TG, Parks DJ, Stewart EL, Will-son TM, Lambert MH, Moore JT, Pearce KH, Xu HE 2002Crystal structure of the glucocorticoid receptor ligandbinding domain reveals a novel mode of receptor dimer-ization and coactivator recognition. Cell 110:93–105
16. Kauppi B, Jakob C, Farnegardh M, Yang J, Ahola H,Alarcon M, Calles K, Engstrom O, Harlan J, Muchmore S,Ramqvist AK, Thorell S, Ohman L, Greer J, GustafssonJA, Carlstedt-Duke J, Carlquist M 2003 The three-dimen-sional structures of antagonistic and agonistic forms ofthe glucocorticoid receptor ligand-binding domain: RU-486 induces a transconformation that leads to activeantagonism. J Biol Chem 278:22748–22754
17. Onate SA, Tsai SY, Tsai MJ, O’Malley BW 1995 Se-quence and characterization of a coactivator for the ste-roid hormone receptor superfamily. Science 270:1354–1357
18. Voegel JJ, Heine MJ, Zechel C, Chambon P, Grone-meyer H 1996 TIF2, a 160 kDa transcriptional mediatorfor the ligand-dependent activation function AF-2 of nu-clear receptors. EMBO J 15:3667–3675
19. Hong H, Kohli K, Trivedi A, Johnson DL, Stallcup MR1996 GRIP1, a novel mouse protein that serves as atranscriptional coactivator in yeast for the hormone bind-ing domains of steroid receptors. Proc Natl Acad SciUSA 93:4948–4952
20. Arany Z, Sellers WR, Livingston DM, Eckner R 1994E1A-associated p300 and CREB-associated CBP belongto a conserved family of coactivators. Cell 77:799–800
21. Robyr D, Wolffe AP, Wahli W 2000 Nuclear hormonereceptor coregulators in action: diversity for sharedtasks. Mol Endocrinol 14:329–347
22. Ford J, McEwan IJ, Wright AP, Gustafsson JA 1997Involvement of the transcription factor IID protein com-plex in gene activation by the N-terminal transactivationdomain of the glucocorticoid receptor in vitro. Mol En-docrinol 11:1467–1475
23. Kumar R, Lee JC, Bolen DW, Thompson EB 2001 Theconformation of the glucocorticoid receptor af1/�1 do-main induced by osmolyte binds co-regulatory proteins.J Biol Chem 276:18146–18152
24. Hittelman AB, Burakov D, Iniguez-Lluhi JA, FreedmanLP, Garabedian MJ 1999 Differential regulation of glu-cocorticoid receptor transcriptional activation via AF-1-associated proteins. EMBO J 18:5380–5388
25. Schlechte JA, Simons Jr SS, Lewis DA, Thompson EB1985 [3H]cortivazol: a unique high affinity ligand for theglucocorticoid receptor. Endocrinology 117:1355–1362
26. Yoshikawa N, Makino Y, Okamoto K, Morimoto C,Makino I, Tanaka H 2002 Distinct interaction of cortivazolwith the ligand binding domain confers glucocorticoidreceptor specificity: cortivazol is a specific ligand for theglucocorticoid receptor. J Biol Chem 277:5529–5540
27. Garabedian MJ, Yamamoto KR 1992 Genetic dissectionof the signaling domain of a mammalian steroid receptorin yeast. Mol Biol Cell 3:1245–1257
28. Allan GF, Leng X, Tsai SY, Weigel NL, Edwards DP, TsaiMJ, O’Malley BW 1992 Hormone and antihormone in-duce distinct conformational changes which are centralto steroid receptor activation. J Biol Chem 267:19513–19520
29. Benkoussa M, Nomine B, Mouchon A, Lefebvre B, Ber-nardon JM, Formstecher P, Lefebvre P 1997 Limitedproteolysis for assaying ligand binding affinities of nu-clear receptors. Recept Signal Transduct 7:257–267
30. Wallace AD, Cidlowski JA 2001 Proteasome-mediatedglucocorticoid receptor degradation restricts transcrip-tional signaling by glucocorticoids. J Biol Chem 276:42714–42721
31. Keeton EK, Fletcher TM, Baumann CT, Hager GL, SmithCL 2002 Glucocorticoid receptor domain requirementsfor chromatin remodeling and transcriptional activationof the mouse mammary tumor virus promoter in differentnucleoprotein contexts. J Biol Chem 277:28247–28255
32. Rupprecht R, Arriza JL, Spengler D, Reul JM, Evans RM,Holsboer F, Damm K 1993 Transactivation and synergis-tic properties of the mineralocorticoid receptor: relation-ship to the glucocorticoid receptor. Mol Endocrinol7:597–603
33. Caldenhoven E, Liden J, Wissink S, Van de Stolpe A,Raaijmakers J, Koenderman L, Okret S, Gustafsson JA,Van der Saag PT 1995 Negative cross-talk between RelAand the glucocorticoid receptor: a possible mechanismfor the antiinflammatory action of glucocorticoids. MolEndocrinol 9:401–412
34. Ray A, Prefontaine KE 1994 Physical association andfunctional antagonism between the p65 subunit of tran-scription factor NF-� B and the glucocorticoid receptor.Proc Natl Acad Sci USA 91:752–756
35. Scheinman RI, Gualberto A, Jewell CM, Cidlowski JA,Baldwin Jr AS 1995 Characterization of mechanisms in-volved in transrepression of NF-� B by activated glu-cocorticoid receptors. Mol Cell Biol 15:943–953
36. Nissen RM, Yamamoto KR 2000 The glucocorticoid re-ceptor inhibits NF�B by interfering with serine-2 phos-phorylation of the RNA polymerase II carboxy-terminaldomain. Genes Dev 14:2314–2329
37. McKay LI, Cidlowski JA 1998 Cross-talk between nuclearfactor-� B and the steroid hormone receptors: mecha-nisms of mutual antagonism. Mol Endocrinol 12:45–56
Yoshikawa et al. • LBD-DBD Functional Interactions in the GR Mol Endocrinol, May 2005, 19(5):1110–1124 1123

38. Bledsoe RK, Stewart EL, Pearce KH 2004 Structure andfunction of the glucocorticoid receptor ligand bindingdomain. Vitam Horm 68:49–91
39. Li Y, Lambert MH, Xu HE 2003 Activation of nuclearreceptors: a perspective from structural genomics.Structure (Camb) 11:741–746
40. Farnegardh M, Bonn T, Sun S, Ljunggren J, Ahola H,Wilhelmsson A, Gustafsson JA, Carlquist M 2003 Thethree dimensional structure of the liver X receptor �reveals a flexible ligand binding pocket that can accom-modate fundamentally different ligands. J Biol Chem278:38821–38828
41. Ali A, Thompson CF, Balkovec JM, Graham DW, Ham-mond ML, Quraishi N, Tata JR, Einstein M, Ge L, HarrisG, Kelly TM, Mazur P, Pandit S, Santoro J, Sitlani A,Wang C, Williamson J, Miller DK, Thompson CM, ZallerDM, Forrest MJ, Carballo-Jane E, Luell S 2004 NovelN-arylpyrazolo[3,2-c]-based ligands for the glucocorti-coid receptor: receptor binding and in vivo activity.J Med Chem 47:2441–2452
42. Geller DS, Farhi A, Pinkerton N, Fradley M, Moritz M,Spitzer A, Meinke G, Tsai FT, Sigler PB, Lifton RP 2000Activating mineralocorticoid receptor mutation in hyper-tension exacerbated by pregnancy. Science 289:119–123
43. Dey R, Roychowdhury P, Mukherjee C 2001 Homologymodelling of the ligand-binding domain of glucocorticoidreceptor: binding site interactions with cortisol and cor-ticosterone. Protein Eng 14:565–571
44. Lanz RB, Rusconi S 1994 A conserved carboxy-terminalsubdomain is important for ligand interpretation andtransactivation by nuclear receptors. Endocrinology 135:2183–2195
45. Modarress KJ, Opoku J, Xu M, Sarlis NJ, Simons Jr SS1997 Steroid-induced conformational changes at ends ofthe hormone-binding domain in the rat glucocorticoidreceptor are independent of agonist versus antagonistactivity. J Biol Chem 272:23986–23994
46. Warnmark A, Treuter E, Wright AP, Gustafsson JA 2003Activation functions 1 and 2 of nuclear receptors: mo-lecular strategies for transcriptional activation. Mol En-docrinol 17:1901–1909
47. Simons Jr SS, Sistare FD, Chakraborti PK 1989 Steroidbinding activity is retained in a 16-kDa fragment of thesteroid binding domain of rat glucocorticoid receptors.J Biol Chem 264:14493–14497
48. Segard-Maurel I, Jibard N, Schweizer-Groyer G, Cade-pond F, Baulieu EE 1992 Mutations in the “zinc fingers”or in the N-terminal region of the DNA binding domain ofthe human glucocorticosteroid receptor facilitate its salt-induced transformation, but do not modify hormonebinding. J Steroid Biochem Mol Biol 41:727–732
49. Xu M, Chakraborti PK, Garabedian MJ, Yamamoto KR,Simons SS 1996 Modular structure of glucocorticoid re-ceptor domains is not equivalent to functional indepen-dence. Stability and activity of the steroid binding do-
main are controlled by sequences in separate domains.J Biol Chem 271:21430–21438
50. Picard D, Yamamoto KR 1987 Two signals mediatehormone-dependent nuclear localization of the glucocor-ticoid receptor. EMBO J 6:3333–3340
51. Pandit S, Geissler W, Harris G, Sitlani A 2002 Allostericeffects of dexamethasone and RU486 on glucocorticoidreceptor-DNA interactions. J Biol Chem 277:1538–1543
52. Garside H, Stevens A, Farrow S, Normand C, Houle B,Berry A, Maschera B, Ray D 2004 Glucocorticoid LigandsSpecify Different Interactions with NF-{�}B by allostericeffects on the glucocorticoid receptor DNA binding do-main. J Biol Chem 279:50050–50059
53. Kumar R, Baskakov IV, Srinivasan G, Bolen DW, Lee JC,Thompson EB 1999 Interdomain signaling in a two-do-main fragment of the human glucocorticoid receptor.J Biol Chem 274:24737–24741
54. Ghali SA, Gottlieb B, Lumbroso R, Beitel LK, Elhaji Y, WuJ, Pinsky L, Trifiro MA 2003 The use of androgen receptoramino/carboxyl-terminal interaction assays to investi-gate androgen receptor gene mutations in subjects withvarying degrees of androgen insensitivity. J Clin Endo-crinol Metab 88:2185–2193
55. Kodama T, Shimizu N, Yoshikawa N, Makino Y, OuchidaR, Okamoto K, Hisada T, Nakamura H, Morimoto C,Tanaka H 2003 Role of the glucocorticoid receptor forregulation of hypoxia-dependent gene expression. J BiolChem 278:33384–33391
56. Okamoto K, Tanaka H, Ogawa H, Makino Y, Eguchi H,Hayashi S, Yoshikawa N, Poellinger L, Umesono K,Makino I 1999 Redox-dependent regulation of nuclearimport of the glucocorticoid receptor. J Biol Chem 274:10363–10371
57. Miura T, Ouchida R, Yoshikawa N, Okamoto K, Makino Y,Nakamura T, Morimoto C, Makino I, Tanaka H 2001Functional modulation of the glucocorticoid receptor andsuppression of NF-�B-dependent transcription by ur-sodeoxycholic acid. J Biol Chem 276:47371–47378
58. Yamamoto K, Masuno H, Choi M, Nakashima K, Taga T,Ooizumi H, Umesono K, Sicinska W, VanHooke J, De-Luca HF, Yamada S 2000 Three-dimensional modeling ofand ligand docking to vitamin D receptor ligand bindingdomain. Proc Natl Acad Sci USA 97:1467–1472
59. Choi M, Yamamoto K, Masuno H, Nakashima K, Taga T,Yamada S 2001 Ligand recognition by the vitamin Dreceptor. Bioorg Med Chem 9:1721–1730
60. Choi M, Yamamoto K, Itoh T, Makishima M, MangelsdorfDJ, Moras D, DeLuca HF, Yamada S 2003 Interactionbetween vitamin D receptor and vitamin D ligands. Two-dimensional alanine scanning mutational analysis. ChemBiol 10:261–270
61. Roux S, Terouanne B, Couette B, Rafestin-Oblin ME,Nicolas JC 1999 Conformational change in the humanglucocorticoid receptor induced by ligand binding is al-tered by mutation of isoleucine 747 by a threonine. J BiolChem 274:10059–10065
Molecular Endocrinology is published monthly by The Endocrine Society (http://www.endo-society.org), the foremostprofessional society serving the endocrine community.
1124 Mol Endocrinol, May 2005, 19(5):1110–1124 Yoshikawa et al. • LBD-DBD Functional Interactions in the GR
Top Related
Copyright © 2022 FDOKUMEN

