







Bahasa
Halaman
Hukum


Neuron
Perspective
Striatal Local Circuitry:A New Framework for Lateral Inhibition
Dennis A. Burke,1,2 Horacio G. Rotstein,3,4 and Veronica A. Alvarez1,5,*1Laboratory on Neurobiology of Compulsive Behaviors, Intramural Research Program, National Institute on Alcohol Abuse and Alcoholism,NIH, Bethesda, MD 20892, USA2Department of Neuroscience, Brown University, Providence, Providence, RI 02912, USA3Federated Department of Biological Sciences, New Jersey Institute of Technology and Rutgers University, Newark, NJ 07102, USA4Institute for Brain and Neuroscience Research, New Jersey Institute of Technology, Newark, NJ 07102, USA5Intramural Research Program, National Institute on Drug Abuse, NIH, Baltimore, MD 21224, USA*Correspondence: [email protected]://doi.org/10.1016/j.neuron.2017.09.019
This Perspective will examine the organization of intrastriatal circuitry, review recent findings in this area, anddiscuss how the pattern of connectivity between striatal neurons might give rise to the behaviorally observedsynergism between the direct/indirect pathway neurons. The emphasis of this Perspective is on theunderappreciated role of lateral inhibition between striatal projection cells in controlling neuronal firingand shaping the output of this circuit. We review some classic studies in combination with more recentanatomical and functional findings to lay out a framework for an updated model of the intrastriatal lateral in-hibition, where we explore its contribution to the formation of functional units of processing and theintegration and filtering of inputs to generate motor patterns and learned behaviors.
IntroductionThe striatum, as the input nucleus to the basal ganglia, is func-
tionally implicated in motor planning, action selection, reward
guided learning, and other motivated behaviors and cognitive
processes. Not surprisingly given these broad functions, striatal
dysfunction is involved in a variety of diverse neurological and
psychiatric diseases including, but not limited to, Parkinson’s
disease, Huntington’s disease, schizophrenia, depression, and
addiction. By receiving glutamatergic inputs from cortical,
thalamic, and limbic regions combined with dopaminergic input
from the midbrain, the striatum acts as an integrative hub that
assists in the selection of appropriate behaviors through its
outputs to downstream basal ganglia structures (Redgrave
et al., 1999). This integrative function of the striatum that is crit-
ical for the processing of input information is thought to happen
at the level of the local intrastriatal circuitry. However, it is still
unclear exactly how local striatal circuitry transforms information
flow and ultimately affects behavioral output. This Perspective
summarizes the most recent finding on the properties and
organization of the local circuitry in the striatum with special
emphasis on the connectivity between the striatal projection
neurons. This component of the circuitry has been largely
ignored in the past because the connectivity is considered rela-
tively sparse. However, recent studies suggest that this synaptic
connection between projection neurons, which is mediated by
their local axon collaterals within the striatum, can exert potent
lateral inhibition to the striatal circuitry, which can profoundly
regulate the excitability of other projection neurons. Focusing
on this underappreciated component of the striatal circuitry,
this Perspective further speculates that these connections are
functionally organized to aid in the information processing that
shapes striatal output. In doing so, we provide a conceptual
model in which to test how particular patterns of connectivity be-
tween projection neurons can affect the firing. This model pro-
vides a framework for understanding the role of striatal lateral
inhibition with the hope of inspiring future connectomics
research into the organization of the intrastriatal connectivity
and how it ultimately assists in coordinating behavior and
learning both in health and in disease.
The layout of the Perspective is as follows: General Overview
provides a general overview of the different cell types within the
striatum as well as its functional role and inputs. Local Striatal
Circuitry focuses on the local connectivity within the striatum
and describes the wide variety of synaptic connections among
striatal neurons. SPN Collateral Transmission Drives Lateral
Inhibition concentrates on the synapses formed between projec-
tion neurons and offers a historical view from their initial dis-
covery to current findings. It discusses possible functional roles
for this lateral inhibition and presents evidence for the existence
of striatal ensembles. Finally, A Conceptual Model of Synaptic
Organization of the Lateral Inhibition builds on this empirical
data and lays out a theoretical framework for how the synapses
between striatal projection neurons might be organized to aid in
formation of ensembles or functional units. Under this hypothet-
ical model, the pattern of connectivity of the lateral inhibition
determines the degree of co-activation of striatal projection neu-
rons and, as such, shapes the striatal output. It also demon-
strates that different patterns of intrastriatal connectivity will
affect the degree of neuronal co-activation, determining which
units of cells fire together, ultimately contributing to each
ensemble. Further, it opens the door to propositions involving
modulators of lateral inhibition in dynamically re-shaping the
ensembles, which might underlie forms of striatal learning.
This last section aims to stimulate discussion and generate
testable hypotheses that drive research on the topic of striatal
organization.
Neuron 96, October 11, 2017 Published by Elsevier Inc. 267

Neuron
Perspective
General OverviewHeterogeneity of Striatal Subregions and Cell Types
Striatal Subregions. The rodent striatum is traditionally subdi-
vided into two regions, dorsal and ventral, which contain subdi-
visions based on their inputs and/or immunohistochemical
markers. The dorsal striatum, or neostriatum, can be divided
into the dorsal lateral striatum (DLS) and the dorsal medial
striatum (DMS) based on glutamatergic inputs. The ventral
striatum includes the olfactory tubercule and the nucleus
accumbens (NAc), which can be further subdivided into ‘‘core’’
and ‘‘shell’’ subregions based on inputs and immunohistochem-
ical markers.
Another dimension along which the striatum can be divided is
the ‘‘patch’’ (striosome) and ‘‘matrix’’ compartments. First iden-
tified by the expression patterns of proteins such as mu-opioid
receptors and acetylcholinesterase, there are now over 60 genes
known to be differentially expressed in patch versus matrix
compartments (Crittenden and Graybiel, 2011). Though patches
only represent a small proportion of the striatum by volume,
many of the differentially expressed genes are neuromodulators
and their receptors suggesting that the patch and matrix have
distinct pharmacological properties (Banghart et al., 2015; Brim-
blecombe and Cragg, 2017). Afferents to patch and matrix are
also thought to arise from different sources, with patches
receiving limbic inputs and matrix neurons receiving sensori-
motor inputs (Gerfen, 1984). However, recent viral tracing
methods have cast doubt on this strict segregation (Smith
et al., 2016).
The striatal subregions are also functionally segregated. In
general terms, the dorsal striatum is involved in motor planning,
action selection, and stimulus-response habit learning, while the
NAc plays a role in processing-motivated behavior and reward-
related learning. However, there is much overlap in the known
roles of the dorsal and ventral striatum in controlling motor and
motivated behaviors and learning (Isomura et al., 2013). Tech-
nical limitations make it difficult to distinguish motivation from
motor ability and to discriminate learning and memory from
performance. Also the overlapping functions between these
striatal subregions is hypothesized to be embedded within the
circuit connectivity in the form of ‘‘ascending spiral loops’’ that
allow for information flow from the ventral to the dorsal striatum
via the circuitry within the basal ganglia (Haber et al., 2000).
Spiny Projection Neurons. In addition to potential functional
overlaps, the dorsal and ventral striatum share multiple features
including their main cell-type composition. The principal projec-
tion neurons in the striatum are the GABAergic spiny projection
neurons (SPNs; also referred to as ‘‘medium spiny neurons’’),
which comprise over 95% of the neurons in this region (Gravel-
and and DiFiglia, 1985) and are the only projection neurons of
the striatum. SPNs are classically divided in two subclasses
based on their projection targets and expression of dopamine
receptors (Gerfen et al., 1990). Direct pathway SPNs (dSPNs)
represent approximately half of the SPN population, mainly ex-
press Gs/olf-coupled dopamine D1 receptors, and send dense
projections to the ventral tegmental area (VTA) or the substantia
nigra pars reticulata and internal globus pallidus/entopeduncular
nucleus through the ‘‘direct’’ pathway of the basal ganglia. Indi-
rect pathway SPNs (iSPNs) mainly express Gi/o-coupled dopa-
268 Neuron 96, October 11, 2017
mine D2 receptors and project through the ‘‘indirect’’ pathway
to the external globus pallidus (GP) and the ventral pallidum
(VP). It is clear now that this dichotomy is an over-simplification
of the existing subtypes of SPNs and that the segregation is
imperfect. For example, a subset of SPNs co-express both
D1 and D2 receptors in the adult striatum (�2%–5% dorsal
striatum,�6%–7% in the NAc core,�12%–15% in the NAc shell)
that are morphologically distinct from other SPNs and have a
poorly understood function (Bertran-Gonzalez et al., 2008; Gag-
non et al., 2017). Furthermore, a recent study shows large cell-
to-cell variability in the pattern of overall gene expression within
each subclass of SPN (Gokce et al., 2016). Another consider-
ation is that the projections are not fully segregated in the sense
that D1R expressing SPNs in the dorsal striatum also send collat-
eral axon projections to the GP (Cazorla et al., 2014; Kawaguchi
et al., 1990) and those in the NAc send numerous projections the
VP (Kupchik et al., 2015). While the classic segregation of SPNs
into direct- and indirect-pathway neurons is incomplete and
oversimplified, it has proven extremely useful for advancing
our understanding of this brain region and the function of the
basal ganglia as a whole. With these caveats in mind, keeping
with field nomenclature, throughout this manuscript we will use
dSPN to refer to SPNs expressing D1 receptors and iSPNs to
refer to those expressing D2 receptors.
Interneurons. The remaining 5% of neurons in the striatum are
either cholinergic or GABAergic interneurons. Cholinergic inter-
neurons (CIN) are large, tonically active neurons that make up
about 1% of the cells in the striatum and, as their name implies,
release acetylcholine (Zhou et al., 2002). These cells have been
classically referred to as TANs for ‘‘tonically active neurons’’
(but see below that they are not the only type of spontaneously
active interneuron). Despite their relatively low abundance,
CINs extend wide axonal arborizations throughout the region
profoundly influencing striatal function through a variety of
muscarinic and nicotinic acetylcholine receptors (reviewed in
Gonzales and Smith, 2015). Recently, CINs were shown to also
co-release glutamate (Guzman et al., 2011; Higley et al., 2011;
Nelson et al., 2014a). Further experiments using paired record-
ings will be useful in confirming this finding.
TheGABAergic interneurons of the striatum can be subdivided
in three classical groups and at least three new additional re-
ported subpopulations based on a combination of molecular,
physiological, and pharmacological properties (Figure 1).
The three classically described GABAergic interneurons are
the fast-spiking (FSI), the low-threshold spiking (LTSI), and the
calretinin expressing (CR) interneuron. FSIs, the best-studied
subtype of the GABAergic interneurons, express parvalbumin
(PV), can fire action potentials at high frequencies, and mediates
strong feedforward inhibition onto SPNs. LTSIs are tonically
active, and although striatal ‘‘TANs’’ is used in the literature to
refer to presumed cholinergic interneurons, this is an important
consideration for in vivo electrophysiology experiments. LTSIs
exhibit low-threshold Ca2+ spikes and can be identified based
on the expression of somatostatin (SOM), nitric oxide synthase
(NOS), and neuropeptide Y (NPY). Not much is known with re-
gards to the physiology or synaptic function of CR interneurons
likely due to their low abundance in mouse compared to primate
striatum (Petryszyn et al., 2014).

GABA
ACh
PV
SOM/NOS/NPY
NPY
CR
FS ChAT
TAN
LTS*
FA5HT3R
TH
?
IIIIIIIV
NGF-like
striatal interneurons
PV/5HT3R
Figure 1. Diversity of Cell Types of Striatal InterneuronsThe plot represents the estimated proportions of each subtype of striatalinterneuron identified based on their expression of neurotransmitter (innerdonut), other molecular markers (middle donut), and electrophysiologicalproperties (outer donut). Abbreviations: ACh, acetylcholine; ChAT, choline-acetyl transferase; TAN, tonically active neurons; TH, tyrosine hydroxlase;CR, calretinin; SOM, somatostatin; NOS, nitric oxide synthase; NPY, neuro-peptide Y; LTS, low-threshold spiking; NGF, neurogliaform; 5HT3R, serotonintype-3 receptor; FA, fast adapting; PV, parvalbumin; FS, fast spiking. *, SOM/NOS LTS neurons are also tonically active, although not classically referredas TANs.
Neuron
Perspective
Recently, BAC transgenic mice have facilitated further classi-
fication of novel GABAergic striatal interneuron subtypes based
on molecular, physiological, and pharmacological properties
(Figure 1). For example, a recently characterized population of
striatal interneurons expresses tyrosine hydroxlase (TH). TH
interneurons all release GABA, and this group does not appear
to overlap with other known subtypes of striatal interneurons.
Characterization of these neurons has revealed four distinct
subtypes based on electrophysiological properties, termed
Type I-IV (Ibanez-Sandoval et al., 2010). Interestingly, though
tyrosine hydroxylase is the rate limiting enzyme in dopamine
synthesis and is used as a marker for dopamine neurons, TH
interneurons do not appear to release dopamine (Xenias
et al., 2015).
Two recently identified novel interneuron subtypes are the
NPY-neurogliaform (NPY-NGF) interneuron and the fast-adapt-
ing interneuron (FAI). NPY-NGF interneurons express NPY, but
not SOM and NOS, as LTSIs do, and share similarities with
cortical NPY-expressing neurogliaform cells (Ibanez-Sandoval
et al., 2011). NPY-NGF neurons are excited by acetylcholine
through nicotinic receptors and differ substantially from SOM/
NOS/NPY-expressing LTS neurons in both electrophysiological
and anatomical characteristics, as well as in the synaptic
response that they elicit in SPNs.
FAIs have been identified by expression of the serotonin
5-HT3 receptor (5HT3R) and given their name due to a distinct
spike-frequency adaptation during repetitive firing (Faust et al.,
2015). FAIs express nicotinic acetylcholine receptors. The
5HT3R is not a perfect marker for the FAI, however, as in many
interneurons in the striatum the 5HT3R is coexpressedwith other
interneuron markers, in particular PV. Neurons that express both
PV and 5HT3R appear to have FSI electrophysiological charac-
teristics similar to those neurons that express PV alone (Munoz-
Manchado et al., 2016). For reviews of striatal interneurons, see
Tepper et al. (2010) and Tepper and Koos (2017).
Functional Overview
Classical models of basal ganglia function posit that activation
of dSPNs disinhibits principal thalamic neurons to facilitate
behavior, while activation of iSPNs ultimately inhibits thalamic
neurons to suppress motor and reward-based behaviors. Dysre-
gulation in the balance of dSPN and iSPN activity is thought to be
the underlying cause of several basal ganglia movement disor-
ders (Albin et al., 1989; DeLong, 1990). This model is supported
by experiments using cell-specific genetic perturbation, cell
ablation, or optogenetic activation (Bateup et al., 2010; Bock
et al., 2013; Durieux et al., 2009; Kravitz et al., 2010, 2012;
Lobo et al., 2010; Tai et al., 2012).
Glutamate inputs drive SPN firing and, without these excit-
atory inputs, SPNs are quiescent and their membrane potential
is very hyperpolarized. During a desirable action, dopamine is
thought to modulate the neuronal excitability to acutely raise
the signal-to-noise ratio through activation of Gs/olf-coupled D1
receptors (D1Rs) on dSPNs and the Gi/o-coupled D2 receptors
(D2Rs) on iSPNs. Also, by triggering synaptic plasticity and
inducing changes in gene expression, dopamine biases future
behavior toward repeating desirable actions, mediating pro-
cesses that underlie learning (Alexander and Crutcher, 1990;
Cohen and Frank, 2009; Gerfen and Surmeier, 2011; Surmeier
et al., 2014; Tritsch and Sabatini, 2012).
There is accumulating evidence for a more nuanced view of
the interaction between the direct and indirect pathways in
driving behavior. Recent work in the dorsal striatum using cal-
cium imaging with fiber photometry (Cui et al., 2013; Tecuapetla
et al., 2014), deep brain calcium imaging (Barbera et al., 2016),
and in vivo electrophysiology, in which SPN subtypes were iden-
tified with optogenetic tagging (Jin et al., 2014) or post hoc in situ
hybridization (Isomura et al., 2013), have all demonstrated that
both dSPNs and iSPNs are more active when an animal is
performing a task or freely moving and less active when the
animal is not moving or engaged in a task (though dSPNs and
iSPNs have differing activity patterns). These data imply that
iSPNs and dSPNs are coactive during action initiation and that
the direct and indirect pathways are both necessary to facilitate
actions. Further support for this idea comes from studies
showing that inhibition of either iSPNs or dSPNs can disrupt
action initiation or ongoing behavior (Tecuapetla et al.,
2014, 2016).
Extending this idea to the ventral striatum, optogenetic inhibi-
tion of either iSPN or dSPNs in the NAc after trial start cue in an
operant food task reduced motivated responding on the task
(Natsubori et al., 2017), and inhibiting synaptic transmission
from either iSPNs or dSPNs prevents the acute locomotor
response to cocaine and methamphetamine, a behavior known
to involve the NAc (Hikida et al., 2010). These studies provide
evidence that both dSPNs and iSPNs are more active during
actions than at rest, and activity in both SPN subtypes is needed
for driving normal motivated behavior.
Neuron 96, October 11, 2017 269

Neuron
Perspective
In addition, fiber photometry recordings that examine the
activity of dSPNs and iSPNs in the NAc during an operant food
responding task show that both SPN subtypes are more active
after a trial start cue and reward delivery than at rest (Natsubori
et al., 2017). Furthermore photometry recordings in the NAc dur-
ing a risky decision task have demonstrated that iSPN activity is
highest following a risky choice that led to no reward and that this
activity predicts future ‘‘safe’’ choices (Zalocusky et al., 2016).
When recording calcium transients in iSPNs during a cocaine-
conditioned place preference paradigm, iSPNs show marked
decreases in their activity after the animal moves into the drug
paired side of the chamber, similar to their response to cocaine
(Calipari et al., 2016). Three main considerations with fiber
photometry experiments are: (1) this technique measures cal-
cium signals, an indirect assessment of spikes, and while it is
capable of detecting single spikes under the right conditions,
this is almost rarely verified, (2) the time course of the photometry
signal is largely determined by properties of the calcium indica-
tor, not the spiking, and (3) the spatial information is lost when
collecting photons through a fiber and as such photometry
measures bulk calcium signals that are amplified when there is
simultaneous activation. Thus, photometry signals in vivo are
most likely a reflection of 1–3 action potentials and the signal
detection is biased toward synchronized events that can sum-
mate in time. Photometry signals might reflect the activity of
ensembles rather than single neurons (Alvarez, 2016). While it
is important to consider these factors, neither of these consider-
ations undermines the main findings that co-activation of iSPNs
and dSPNs is seen during a wide range of behaviors. Altogether
these studies suggest that both the direct and indirect pathways
are necessary to drive behavioral output in both the dorsal and
the ventral striatum.
An important question then is how to reconcile the findings
from optogenetic stimulation experiments suggesting that
iSPN activation inhibits movement (Kravitz et al., 2010) with
those from calcium dynamics showing iSPN activation during
action initiation and behavior (Barbera et al., 2016; Cui et al.,
2013; Natsubori et al., 2017). The most parsimonious explana-
tion is that during behavior, dSPNs facilitate the desired
behavior, while iSPNs act simultaneously to inhibit all other
competing undesirable behaviors (see more in Local Striatal
Circuitry). Indeed, this concept of the basal ganglia serving as
a selection mechanism between competing behavioral pro-
grams was proposed over 20 years ago by Jonathon Mink
(Mink, 1996, 2003; Mink and Thach, 1993) and Okihide Hikosaka
(Hikosaka, 1991, 1998), and it has been incorporated into
computational models of basal ganglia function (Collins and
Frank, 2014; Frank, 2005). Consistent with this, global optoge-
netic stimulation of iSPNs would therefore suppress all behav-
ioral programs and inhibit movement.
If this model were correct, precise regulation of the activity of
dSPNs and iSPNs that control competing behavioral programs
would be required (Tecuapetla et al., 2016), making it important
to determine how striatal circuitry can give rise to this coordi-
nated activity. In order for the model to work properly, coordi-
nated activation of dSPNs that facilitate a desired behavior and
activation of iSPNs that suppress competing behaviors should
be achieved. This coordination could be dictated by the inputs
270 Neuron 96, October 11, 2017
to the striatum, the organization of the local intrastriatal connec-
tivity, or a combination of both. In the next sections, we will first
review inputs into the striatum and then discuss the local intra-
striatal circuity.
Main Inputs to the Striatum
The striatum receives two main types of inputs: glutamatergic
and dopaminergic. Glutamatergic projections arise from most
cortical regions, as well as thalamic and limbic regions. Cortical
inputs descend in a topographic fashion with neighboring
regions of cortex projecting to adjacent regions of the striatum
(Bolam et al., 2000; Dudman and Gerfen, 2015). The DLS
receives inputs from sensorimotor cortex, while the DMS is
innervated by associative regions of cortex. Both DMS and
DLS receive inputs from the thalamus. The NAc is targeted by
associative and limbic regions of cortex, as well as amygdala,
hippocampal, and thalamic nuclei (Britt et al., 2012; Bubser
and Deutch, 1998; MacAskill et al., 2012). Cortical inputs target
both subtypes of SPNs, with single axons making synapses
onto both dSPNs and iSPNs (Doig et al., 2010). While anatomical
studies have suggested that dSPNs and iSPNs receive inputs
from different types of cortical neurons, with intertelencephalic
(IT) neurons more likely to synapse onto dSPNs and pyramidal
tract neurons (PT) more likely to synapse onto iSPNs (Lei et al.,
2004), physiological studies have shown that both types of
cortical neurons synapse onto both iSPNs and dSPNs (Kress
et al., 2013). Retrograde viral tracing methods have suggested
that there may be differences in dSPN and iSPN innervation by
cortical region (Wall et al., 2013). However, a subsequent study,
in which the number of presynaptic neurons labeled outside the
striatum was an order of magnitude greater, found no difference
in dSPN or iSPN innervation by cortical region (Guo et al., 2015).
This suggests that the initial findings of segregated inputs
could have suffered from sampling bias and cautions against
drawing quantitative conclusions from viral methods with low
infection rates.
In the dorsal striatum, both dSPN and iSPNs receive glutama-
tergic inputs from not only cortex, but also thalamus, and single
axons of both types contact both dSPNs and iSPNs (Doig et al.,
2010). However, cortical and thalamic synapses differ in their
properties: cortical synapses onto SPNs have amuch higher fail-
ure rate and are facilitating, while thalamic synapses have a
lower rate of failure and are depressing (Ding et al., 2008). Syn-
apses from different thalamic nuclei display a great deal of het-
erogeneity in their strength, short-term dynamics, and SPN sub-
type targets (Ellender et al., 2013; Smith et al., 2009).
Furthermore, through their interactions with cholinergic interneu-
rons, thalamic neurons appear to ‘‘gate’’ cortical information
flow, suggesting an interesting interaction between these two in-
formation streams for striatal processing (Ding et al., 2010).
The striatum is densely innervated by dopamine neurons.
Dopamine neurons of the substantia nigra pars compacta
target the dorsal striatum, while those of the VTA target the
ventral striatum. Dopamine’s influence on striatal function is
vast and complex due to the variety of D1-like (D1 and D5)
and D2-like (D2, D3, and D4) receptors located on both axon
terminals and postsynaptic sites in the striatum. In addition to
differential effects in directly modulating D1 containing dSPN
and D2 containing iSPNs, dopamine powerfully influences

Neuron
Perspective
striatal processing through its actions on cholinergic and
GABAergic interneurons as well as presynaptic terminals of
inputs to the striatum. These actions of dopamine on the inputs
can be acute and reversible or expressed in the form of long-
term changes in synaptic strength (for reviews, see Gerfen
and Surmeier, 2011; Kreitzer and Malenka, 2008). For reviews
covering afferents to GABAergic interneurons, see Tepper
et al. (2010) and Tepper and Koos (2017).
It is now known that the midbrain provides another source of
glutamate to these regions. The VTA contains a population of
glutamate neurons that projects to the striatum (Hnasko et al.,
2012; Yamaguchi et al., 2007, 2011, 2015) aswell as a population
of neurons that co-release dopamine and glutamate (Hnasko
et al., 2010; Stuber et al., 2010; Tecuapetla et al., 2010). VTA
glutamate neurons projecting to the NAc shell synapse onto
FSIs and mediate behavioral aversion (Qi et al., 2016). In addi-
tion, it has recently been reported that nondopaminergic
glutamate neurons in the VTA can also co-release GABA and
that stimulation of these neurons can be transiently reinforcing
(Yoo et al., 2016).
Further, dopamine neurons have been shown to co-release
GABA along with dopamine in the dorsal striatum (Tritsch
et al., 2012) and the NAc, though dopamine neuron IPSCs re-
corded in the NAc are smaller than in the dorsal striatum (Kim
et al., 2015). Interestingly, dopamine neurons do not synthesize
GABA through canonical GAD65 and GAD67 enzymes but rather
uptake it through the plasmamembrane (Tritsch et al., 2014) and
synthesize it through a non-canonical aldehyde dehydrogenase
1a1 pathway (Kim et al., 2015).
Functional GABA projections from cortex have recently been
described in both NAc and dorsal striatum. The mPFC sends
GABA projections to the NAc shell, and activation of these
afferents produces avoidance behavior in a real-time place-pref-
erence paradigm (Lee et al., 2014). In the dorsal striatum, cortical
GABA projections from layer 5 and 6 SOM+ cells to both dSPNs
and iSPNs have recently been described in both auditory and
motor cortex, implying that this may be a general feature of
cortex (Rock et al., 2016). These findings are in agreement with
anatomical studies that identified cortical GABA projections to
the striatum (Tomioka et al., 2015) and challenge the common
assumption that cortical projections to the striatum are exclu-
sively glutamatergic.
Another large source of GABA to the striatum comes from
back projections from the GP. Recently identified arkypallidal,
or GP-TA, neurons in the GP exclusively target the striatum
with �10,000 boutons per cell and synapse onto SPNs as well
as both cholinergic and GABAergic (PV+ and NOS+) interneu-
rons, suggesting that these cells may provide the largest
extrinsic source of GABA to the striatum (Mallet et al., 2012).
Functionally, in vivo recordings of these neurons during a
behavior task, in which animals must abruptly abort an action,
imply that arkypallidal cells may be responsible for sending a
‘‘stop’’ signal to the striatum to cancel evolving action plans
(Mallet et al., 2016). In addition to arkypallidal cells, GP neurons
that project to downstream structures send GABA projections to
the striatum that target PV interneurons (Bevan et al., 1998). VTA
contains nondopaminergic GABA neurons that project to the
NAc (Van Bockstaele and Pickel, 1995). In addition to locally
modulating the excitability of VTA dopamine neurons (Tan
et al., 2012; van Zessen et al., 2012), these neurons project to
cholinergic interneurons in the NAc and can pause their firing,
which plays a role in associative learning (Brown et al., 2012).
Thus, in addition to GABA co-released from midbrain dopamine
neurons, GABA projection neurons in midbrain also influence
striatal circuits via their projections to CINs.
Another novel inhibitory projection from the bed nucleus of the
stria terminalis to the striatum has also been described, terminat-
ing on patch and exo-patch SPNs in the dorsal striatum (Smith
et al., 2016). Using tract tracing, another study identified an
extrinsic source of acetylcholine arising from brainstem nuclei,
the pedunculopontine nucleus and the lateral dorsal tegmental
nucleus (Dautan et al., 2014). The function of these newly
identified projections still needs to be determined. Other minor
afferents to the striatum release neuromodulators, such as sero-
tonin from the dorsal raphe (Miguelez et al., 2014) and histamine
and oxytocin from hypothalamus (Bolam and Ellender, 2016;
Dolen et al., 2013).
3. Local Striatal CircuitryThere is increasing appreciation for the role that local striatal
circuitry plays in allowing the striatum to compute sensory,
motor, and limbic information into behavioral or cognitive output.
It can filter, modulate, amplify, and ultimately transform informa-
tion flow through this region and produce an output that could
not be predicted by the summation/integration of the inputs to
SPNs alone.
The local connectivity is mediated by the striatal interneurons
and by the synapses formed by collateral axons of SPNs that
linger within the striatum and contact other SPNs. While some
of these local connections are reciprocal, most of them are
unidirectional in nature. Local connectivity can shape the output
by directly affecting the activity of projection neurons via classic
axo-somatic and axo-dendritic synapses onto SPNs that alter
neuronal excitability, cell signaling, dendritic integration, and
gene expression. Alternatively, local connectivity can act indi-
rectly to affect striatal output via axo-axonal synapses, which
can modulate the synaptic inputs to SPNs or the synaptic trans-
mission from SPNs (Figure 2). Below, we will briefly outline the
different synaptic connections between local striatal interneu-
rons and projection neurons (Figure 3).
Axo-axonal Connections
In axo-axonal synapses, neurotransmitter release from one
synaptic bouton activates receptors located presynaptically on
another bouton or axon fiber and act directly to suppress, facil-
itate, or even trigger neurotransmitter release from the ‘‘postsyn-
aptic’’ axon terminal, sometimes without affecting the firing of
action potentials from the postsynaptic neuron (Figure 2). While
anatomical evidence for axo-axonal synapses in the striatum is
still lagging behind, functional data indicate that presynaptic
heteroreceptors can be activated quickly and reliably upon
neurotransmitter release from other terminals (Cachope et al.,
2012; Kosillo et al., 2016; Threlfell et al., 2012; J.H. Shin, M.F.
Adrover, and V.A.A., unpublished data). A recent example of
an axo-axonal connection that was functionally identified in the
striatum is between CINs and dopamine (DA) fibers. Synchro-
nized release of ACh from cholinergic interneurons can activate
Neuron 96, October 11, 2017 271

DA
GABA
ACh
D2R
GABA
Glut
5 M
D2
2D4/2
M
D2M2/4
nAR
M4
2D
DA
GABA
ACh
D2R
GABA
Glut
5 M
D2
2D4/2
M
D2M2/4
nAR
M4
2D?
Figure 2. Axo-axonal Modulation in theStriatumSimple diagram of known axo-axonal presynapticmodulation by dopamine and acetylcholine (ACh)in the striatum. D2, dopamine D2 receptor; M5,M5muscarinic ACh receptor; M2/4, M2 and M4muscarinic ACh receptors; nAR, nicotinic AChreceptor.
Neuron
Perspective
presynaptic nicotinic acetylcholine receptors (nAChRs) located
on DA axons and evoke DA release in both ventral and dorsal
striatum (Cachope et al., 2012; Threlfell et al., 2012). Via presyn-
aptic muscarinic M5 receptors on DA axon fibers, ACh can
further enhance DA transmission in the striatum (Shin et al.,
2015). Through activation of presynaptic muscarinic M2/M4 re-
ceptors on glutamatergic terminals from cortex and thalamus,
ACh can also suppress glutamate transmission onto SPNs
(Ding et al., 2010). GABA transmission from FSIs onto SPNs is
also inhibited by ACh through presynaptic muscarinic receptors
(Koos and Tepper, 2002). In addition, the existence of nAChRs
on glutamatergic terminals has been suggested (Kaiser and
Wonnacott, 2000). Further confirmation that CIN stimulation
and/or ACh release from these interneurons can produce these
modulatory effects on GABA and glutamate transmission will
be important for the understanding the spatial and temporal lim-
itation of ACh transmission
Other neurotransmitters act presynaptically in the striatum as
well: through activation of presynaptic D2 receptors, DA inhibits
its own release from DA terminals through autoreceptors,
GABA transmission from iSPNs (Dobbs et al., 2016; Tecuapetla
et al., 2009), and acetylcholine release from CINs. It is proposed
to inhibit glutamate transmission fromcortical afferents (Bamford
et al., 2004), though because cortical D2 receptor expression is
sparse, postsynaptic effects of dopamineonglutamate transmis-
sion likely playa large role in observedmodulation (HigleyandSa-
batini, 2010). Further, single NPY-NGF interneuron firing can
inhibit cortical glutamate release through presynaptic GABAB re-
ceptors, suggesting an axo-axonal synapse (Logie et al., 2013).
Axo-dendritic and Axo-somatic Connections
CIN-GABA Interneuron. Cholinergic interneurons form synapses
onto FSIs and release both glutamate and acetylcholine, though
these synapses are weak and sometimes undetected (English
et al., 2011; Nelson et al., 2014a). ACh has been shown to both
depolarize FSIs through nicotinic receptors and tomodulate their
synaptic transmission by inhibiting GABA transmission onto
SPNs through presynaptic muscarinic receptors (Koos and Tep-
per, 2002; Luo et al., 2013). Interestingly, despite this evidence of
nicotinic stimulation and depolarizing responses to cholinergic
stimulation, FSIs do not appear to play a major role in the disy-
naptic inhibition of SPNs through cholinergic interneurons
(Nelson et al., 2014b). CIN to FSI synaptic connections are not
reciprocal as it has been shown that FSI cells do not form
272 Neuron 96, October 11, 2017
synapses onto cholinergic interneurons
(Straub et al., 2016; Szydlowski
et al., 2013).
Cholinergic interneurons excite both
5HT3R-expressing and NPY-NGF neu-
rons through nicotinic receptors (English et al., 2011; Faust
et al., 2015). It is unclear whether these connections are recip-
rocal. However, stimulation of a single CIN is capable of eliciting
polysynaptic GABAA-mediated synaptic responses both in itself
and in nearby cholinergic interneurons (Sullivan et al., 2008). This
recurrent feedback inhibition requires activation of nAChRs and
while the cell-type identity of this source of GABA is unknown, it
is possible that GABA interneurons are responsible for the
recurrent inhibition. Though generally not detected in paired
recordings between cells in close proximity (Gittis et al., 2010;
Ibanez-Sandoval et al., 2011), optogenetic experiments have
shown that LTSIs form GABAergic synapses onto distant cholin-
ergic interneurons inhibiting them through GABAA receptors
(Straub et al., 2016). In addition, there is evidence that LTSIs
can induce slow depolarizations in cholinergic interneurons
mediated by nitric oxide (Elghaba et al., 2016).
TH interneurons are depolarized by nicotinic agonists, indi-
cating the presence of nicotinic receptors and suggesting that
TH interneurons receive synaptic contacts from cholinergic
interneurons or afferents (Ibanez-Sandoval et al., 2015; Luo
et al., 2013). However, direct synaptic responses from cholin-
ergic neurons have not yet been recorded.
CIN-SPN. Cholinergic neurons acutely modulate SPN excit-
ability and synaptic function directly though muscarinic recep-
tors (for review, see Oldenburg and Ding, 2011) and have a
role in controlling striatal plasticity (Wang et al., 2006). Indirectly
cholinergic interneurons can modulate SPNs through disynaptic
release of DA from midbrain terminals (Cachope et al., 2012;
Threlfell et al., 2012) and release of GABA from FAIs or NPY-
NGF interneurons (English et al., 2011; Faust et al., 2015, 2016)
as well as from DA terminals (Nelson et al., 2014b).
SPNs are known to synapse onto cholinergic interneurons and
release GABA (Chuhma et al., 2011). There is some anatomical
evidence that dSPNs in the dorsal striatum make more synaptic
contacts onto cholinergic interneurons than iSPNs do (Martone
et al., 1992); however, rigorous quantification is lacking, and
whether this is generalizable to the ventral striatum is unknown.
GABA Interneurons-GABA Interneurons. FSIs weakly and
sparsely inhibit LTSIs (Gittis et al., 2010; Lee et al., 2017; Szy-
dlowski et al., 2013). Paired recordings have failed to detect
LTSI synapses onto FSIs; however, as LTSI synapses onto
distant CINs and SPNs have also been missed by paired record-
ings, it remains possible that LTSIs synapse onto distant FSIs.

dSPN
iSPN dSPN
iSPN
ChAT
ChATNPY-NGF
PV
5HTR3
PV
SOM/NOS/NPY
TH
dSPN
iSPN
local circuitry
Figure 3. Main Identified SynapticConnections between Local StriatalNeuronsSchematic showing intrastriatal connectivity be-tween interneurons and projection neurons. In-terneurons displayedwithmolecular markers usedto identify cells during experiments. Abbrevia-tions: dSPN, direct pathway projecting mediumspiny neuron; iSPN, indirect pathway projectingmedium spiny neuron; ChAT, choline-acetyltransferase; TH, tyrosine hydroxlase; SOM, so-matostatin; NOS, nitric oxide synthase; NPY,neuropeptide Y; NGF, neurogliaform; 5HT3R,serotonin 3 receptor; PV, parvalbumin;
Neuron
Perspective
FSIsstronglysynapseonotherFSIs, communicatingchemically
through GABA synapses as well as electrically through gap junc-
tions (Gittis et al., 2010; Koos and Tepper, 1999). NPY-NGF inter-
neurons receive GABAergic inhibition from FSIs (Lee et al., 2017).
GABA Interneurons-SPNs. FSIs strongly synapse on SPNs
(Gittis et al., 2010; Koos and Tepper, 1999; Planert et al.,
2010). In vivo, FSIs respondmore quickly and to lower thresholds
of cortical stimulation than SPNs, providing a powerful mecha-
nism for feedforward inhibition from cortex (Mallet et al., 2005).
FSIs appear to have a slight bias in targeting dSPNs over iSPNs
(Gittis et al., 2010; Planert et al., 2010). There is no evidence for
SPNs synapsing onto FSIs (Chuhma et al., 2011). LTSIs have
been found in paired recordings with SPNs to evoke sparse
GABA responses on SPNs that were much weaker than those
evoked from FSIs to SPNs (Gittis et al., 2010; Luo et al., 2013).
Recent work using viral tracing and optogenetics has shown
that this difference in connectivity detected with paired record-
ings from cells in close proximity is likely due to differences in
spatial organization of these circuits: LTSIs target distant
SPNs, while FSIs target SPNs in close proximity (Straub et al.,
2016). Further, LTSIs synapse on distal dendrites of SPNs, while
FSIs target the soma and proximal dendrites of SPNs.
FAIs provide fast GABAA-mediated inhibition to SPNs, while
NPY-NGF neurons mediate slow GABAA synaptic responses
(English et al., 2011; Faust et al., 2015). NPY-NGF activation
inhibits SPN firing in vivo (Lee et al., 2017). Because NPY-NGF
interneurons and FAIs are excited by cholinergic stimulation,
this provides a both quick and prolonged mechanism for disy-
naptic cholinergic inhibition of SPNS. TH interneurons synapse
onto SPNs and release GABA. Interestingly, TH interneurons
are the only identified GABAergic interneurons to receive inputs
form SPNs (Ibanez-Sandoval et al., 2010), suggesting a unique
role for these interneurons in striatal processing.
Collateral Transmission between SPNs
In addition to their long-range projection targets, SPNs extend
dense axon collaterals that stay within the striatum or nucleus
accumbens and target other SPNs (Chang and Kitai, 1985;
Pennartz et al., 1991; Somogyi et al., 1981; Wilson and Groves,
1980). These GABAergic synapses are mostly formed on distal
dendrites of SPNs (Wilson and Groves,
1980) and are relatively weak compared
to FSI inputs that are made at the soma
(Czubayko and Plenz, 2002; Koos et al.,
2004; Tunstall et al., 2002). The connec-
tivity is sparse in the sense that only �30% of paired recordings
between SPNs produce a synaptic response, and these connec-
tions are all unidirectional.
The network of collateral axons can provide the circuit with a
sufficient degree of lateral inhibition to shape the output of the
striatum. Indeed, despite the early work showing weak synaptic
connections between discrete pairs of SPNs, synchronized
stimulation of axon collateral from multiple SPN can provide a
strong source of inhibition to other striatal SPNs (Dobbs et al.,
2016). The properties and regulation of this network of lateral
inhibition are discussed in more detail below. In addition, we
present some models for the organization of the lateral inhibition
and its implications for circuit level function.
4. SPN Collateral Transmission Drives Lateral InhibitionAnatomical and Functional Evidence
Early anatomical studies identifying SPNs in the dorsal striatum
as the projection neurons of this region made note of their dense
axons collaterals within the striatum (Grofova, 1975; Kemp and
Powell, 1971). Indeed, some of the earliest studies identifying
SPNs, including those by Ramon y Cajal (1911), identified them
as interneurons due to their intrastriatal axon branches and the
limitations of the Golgi technique (Tepper et al., 2010). By
combining antidromic stimulation recordings with cell filling
and anatomical reconstruction, it became clearer that SPNs
have both projections out of the striatum and an extensive collat-
eral plexus within the striatum (Preston et al., 1980). The use of
light and electron microscopy in the early 1980s allowed both
Wilson and Groves and Somogyi and colleagues to show that
SPN axons, in addition to projecting out of the striatum, target
other SPNs within the striatum (Somogyi et al., 1981; Wilson
and Groves, 1980). As the idea of separate direct/indirect
pathway SPN subpopulations began to emerge, Kawaguchi
and colleagues demonstrated that SPNs form intrastriatal
axon collaterals regardless of the downstream projection target
(i.e., whether that cell was an iSPN or dSPN) (Kawaguchi et al.,
1990). Electron microscopy was combined with immunocyto-
chemistry to show that iSPNs and dSPNs were synaptically
interconnected and synapses were formed between SPNs of
Neuron 96, October 11, 2017 273

Neuron
Perspective
different subclasses (Yung et al., 1996). Anatomical studies in the
NAc confirmed that similar to the dorsal striatum, NAc SPNs
have dense axon collaterals that remain within the accumbens
(Chang and Kitai, 1985; Pennartz et al., 1991) in addition to the
long-range axon projections to the midbrain and pallidum.
Despite the wealth of anatomical evidence, the resulting
functional effect of the high density of local synapses between
SPNs has been harder to grasp. Electrophysiological studies us-
ing antidromic stimulation hinted at early evidence of functional
GABAergic synapses from the axon collaterals in both dorsal
striatum (Park et al., 1980) and nucleus accumbens (Chang
and Kitai, 1985). Despite this encouraging functional evidence,
one of the first studies directly addressing the existence of func-
tional synapses using sharp intracellular electrode recordings in
pairs of SPNs in the dorsal striatum of rats found no evidence for
functional synaptic connections (Jaeger et al., 1994). Another
study using paired whole-cell recordings between fast-spiking
PV interneurons and SPNs found that PV cells form strong
GABAergic connections to SPNs (Koos and Tepper, 1999).
Together, these findings suggested that PV interneurons provide
a larger source of GABAergic inhibition to SPNs than SPN collat-
erals do, contrary to previous hypotheses based on the anatom-
ical data (Tepper et al., 2008).
In 2002, the first evidence of monosynaptic GABAergic con-
nections between SPNs was demonstrated by Tunstall and
colleagues, followed shortly by Czubayko and Plenz (Czubayko
and Plenz, 2002; Tunstall et al., 2002). Tunstall and colleagues
used sharp intracellular electrode recordings in adult rat striatal
slices and found that 9/45 pairs of SPNs formed unidirectional
synapses that were blocked by GABAA antagonists. In addition
to the sparse connectivity (20% of pairs), the synaptic responses
were of small size (IPSP amplitudes of a few hundred microvolts)
and showed high rate of failure. As a consequence, averaging
hundreds of events was required in order to resolve IPSPs,
possibly explaining the discrepancy with the previous study of
SPN pairs. Similarly, using young rats and organotypic cultures,
Czubayko and Plenz found further evidence for the existence of
functional synapses between SPNs. They found synaptic con-
nections between 5/38 (13%) of pairs of SPNs with high failure
rates (�0.5) in rat slices, in agreement with the connectivity rates
reported by Tunstall et al. IPSPs reported in Czubayko and Plenz
were an order of magnitude larger than those reported in Tunstall
et al. (�2 mV versus �250 mV), likely due to differences in the
recording configurations (sharp electrode versus whole cell).
Two years later, Taverna and colleagues showed that SPNs in
the NAc also form functional collateral synapses with each other
(Taverna et al., 2004). Like in the dorsal striatum, these connec-
tions are relatively sparse (13/38 pairs, 34%), mediated by
GABAA receptors, and mainly unidirectional in nature.
Properties and Organization of the Inhibitory Collateral
Transmission
The creation of BAC transgenic mice expressing fluorescent
proteins under the control of the Drd1 or Drd2 promotor (Gong
et al., 2003) allowed for the examination of collateral synapses
in identified dSPNs and iSPNs. In both the dorsal striatum
and the NAc, Taverna and colleagues found asymmetries in
both the connection probability and strength of the synapses be-
tween the SPN subclasses (Taverna et al., 2008). They found that
274 Neuron 96, October 11, 2017
while iSPNs form synapses on postsynaptic dSPN (13/47, 28%)
and iSPNs (14/39, 36%) with a similar probability, dSPNs were
much more likely to form synapse onto other dSPNs (5/19,
26%) than onto iSPNs (3/47, 6%). In addition, presynaptic iSPNs
generate larger inhibitory postsynaptic currents (IPSCs) than
dSPNs do. In similar experiments, Planert and colleagues found
slightly different connection probabilities between the sub-
classes but largely supported the finding that iSPNs are more
likely to form collateral synapses than dSPNs are (Planert
et al., 2010). Future studies will determine whether the asymme-
try in the connection strength and probability is observed in vivo.
These findings are important because they imply that the pattern
of collateral synapses between SPNs is organized in a specific
nonrandom fashion, and they suggest that there is an asymmetry
in the strength and probability of contacts among the sub-
classes.
Though new hypotheses emerged on how SPN collaterals
could shape information flow through the striatum (Plenz,
2003), the concept that PV interneurons are the main source of
GABAergic inhibition in the striatum persisted in the field (Tepper
et al., 2004, 2008). This was due in large part to the fact that PV/
SPN synapses are more likely formed on the SPN soma and
generate larger IPSPs than SPN /SPN synapses, which form
mainly on distal dendrites (Koos et al., 2004). As a result, many
broad theories and overviews of basal ganglia function do not
incorporate a possible role for SPN collateral synapses in influ-
encing basal ganglia output (Calabresi et al., 2014; Cazorla
et al., 2015; Sesack and Grace, 2010; Silberberg and Bolam,
2015). This is in large part because their function remains unclear
but also because collaterals are thought to be weak and infre-
quent. However, estimates suggest that there are >2,800 SPNs
within the dendritic field of a given SPN (Oorschot, 1996) and
thus, 28%–36% of them represent more than 790–1,000 likely
synaptic partners for each SPN. A clever analysis of synapses
in the striatum based on electron microscopy data and synaptic
density estimates that each SPN receives around 1,200 to 1,800
synaptic contacts from other SPNs, implying that lateral inhibi-
tion could strongly affect SPNs and thus shape output (Wil-
son, 2007).
Indeed, recent studies from our laboratory have demonstrated
an important functional role for SPN collaterals in controlling the
excitability of SPNs and locomotion. Simultaneous activation of
iSPN axon collaterals using channelrhodopsin caused notice-
able inhibition of dSPN firing (Dobbs et al., 2016), proving further
confirmation of the anatomical predictions. Important to note
with this study is that synchronous activation with channelrho-
dopsin in a slice does not necessarily recapitulate the in vivo
behavior of the network. However, these experiments do shed
light on what is possible given the existing synapses within the
striatal network. Altogether, the anatomical and functional find-
ings show that while connection probability between isolated
pairs of SPNs is relatively low, and it involves only a handful of
synapses, concerted activation of axon collaterals from multiple
connected SPNs can have a significant inhibitory effect on the
firing of their synaptic partners. Also, activation of D2Rs ex-
pressed in iSPNs suppresses the collateral GABA transmission,
and as such, dopamine and D2-like receptor agonists reduce the
inhibition from iSPNs to dSPNs (Dobbs et al., 2016; Lemos et al.,

Neuron
Perspective
2016). By controlling the degree of lateral inhibition from iSPNs to
dSPNs, D2Rs can gate dSPN action potential firing and output.
In animals with targeted deletion of D2Rs in iSPNs, neither eleva-
tion of endogenous dopamine by cocaine nor a D2-like agonist is
able to suppress GABAergic transmission from iSPNs /
dSPNs, and mice show impaired cocaine-induced locomotion
(Dobbs et al., 2016). In addition, animals lacking D2Rs in iSPNs
display hypolocomotion, bradykinesia, and reduced in vivo firing
of SPNs throughout the striatum. In slices prepared from these
animals, SPNs were shown to receive more potent inhibition
from GABA transmission, likely caused in part by an enhanced
collateral transmission that followed the chronic loss of D2R-
mediated suppression of iSPN collaterals (Lemos et al., 2016).
Taken together, these studies suggest an important functional
role for the network of collateral axons and synapses in regu-
lating the excitability of SPNs within the striatum and in gating
behavioral output of the basal ganglia. The behavioral implica-
tions of the lateral inhibition are multiple and seem to involve
locomotion and stimulant-induced activity.
Other groups have visualized SPN network activity using cal-
cium indicators both in vitro and in vivo to provide evidence
that SPNs are organized into neuronal clusters based on syn-
chronized activation that might represent functional ensembles
(Barbera et al., 2016; Carrillo-Reid et al., 2008; Carrillo-Reid
et al., 2011). In light of this, many computational models have at-
tempted to explain how feedforward inhibition, mediated by PV
cells, and lateral inhibition, mediated by SPN collaterals, could
generate these ensembles and thereby transform information
flow through the striatum (Humphries et al., 2009, 2010; Moyer
et al., 2014; Ponzi and Wickens, 2010, 2012, 2013; Yim et al.,
2011). These models suggest that lateral inhibition mediated
by SPN collaterals might play a role in driving ensemble coher-
ence on slower timescales or in synchronizing active ensembles
and suppressing unsynchronized cells. However, these models
all assume a random connection probability between SPNs
and deserve to be reconsidered in light of the recent evidence
of asymmetries in both strength and connection probability of
the collateral connections between SPN subclasses and the
possible existence of neuronal clusters or ensembles.
Evidence of Striatal Functional Units
An important question to ask is whether there exists a funda-
mental unit of information processing in the striatum. Though
not easily visualized, are there functional units throughout the
striatum analogous to the canonical cortical microcircuit
columns? A convergence of evidence suggests that ensemble
activity in the striatum is important for information transfer
through the region and the resulting behavior.
On one hand, little evidence of synchronized firing is found be-
tween SPNs during in vivo electrophysiological recordings in
anesthetized animals (Stern et al., 1998). This could speak
against the existence of ensembles but given the low firing rate
of SPNs in vivo, little spontaneous synchrony is expected. On
the other hand, in vivo recordings from freely moving animals
show greater evidence of synchronized firing among SPNs
(Miller et al., 2008). In behaving animals, SPN firing is seen
time locked to specific aspects of the behavioral task, suggest-
ing synchronized spiking of functionally related populations
(Adler et al., 2012; Barnes et al., 2005; Cacciapaglia et al.,
2011; Carelli, 2002; Jin et al., 2014; West and Carelli, 2016)
that are not necessarily in close proximity (Bakhurin et al.,
2016). Ensembles are likely formed by sparsely distributed rather
than tightly clustered neurons, and as such, the chances of
simultaneously recording their firing from the same electrode
array might be low. In support of this concept, a recent study
that used a large-scale recording approach found higher
correlation in the resting-state firing of neurons that were respon-
sive to reward predictive cue compared to non-responsive
neurons when investigating the overall firing of a large number
of striatal neurons during a Pavlovian reward conditioning task
(Bakhurin et al., 2016).
Fiber photometry recordings of SPNs in the NAc and dorsal
striatum also showed large transients of calcium concentrations
time locked to cues and specific behaviors during behavioral
tasks (Calipari et al., 2016; Cui et al., 2013; Natsubori et al.,
2017; Zalocusky et al., 2016), suggestive of population syn-
chrony during actions, though again not necessarily due to cells
in close proximity. Moreover, deep brain calcium imaging, which
provides awide field of view, shows clusters of SPNs that display
synchronized calcium transients during free locomotion. The
clusters are observed in both the dorsomedial and dorsolateral,
and they involve both dSPNs and iSPNs, which supports the
concept of striatal functional units where iSPNs and dSPNs
work together to produce a behavioral output (Barbera
et al., 2016).
Further, in the ventral striatum, which has long been hypothe-
sized to be organized into functionally distinct ensembles of cells
(Pennartz et al., 1994), further support for the idea that sparse en-
sembles of cells work together to control striatal behaviors
comes from the analysis of immediate-early gene expression.
For example, examination of immediate-early gene c-fos expres-
sion after NAc-dependent behaviors, such as context-based
cocaine-induced locomotor sensitization has suggested that
sparse ensembles of SPNs are necessary for behavioral expres-
sion (Hope et al., 2006). Lending support to this, causal manipu-
lations of c-fos expression or cell excitability in the sparsely
labeled cells that express c-fos during locomotor sensitization
or conditioned place preference has been shown to disrupt
that behavior (Koya et al., 2009; Tolliver et al., 2000).
How are these ensembles formed? Are the cells in the striatum
simply following cortical commands or does intrastriatal circuitry
play a role in their activity? The topographical organization of the
cortex is conveyed through its projections to the striatum and is
proposed to persist through the basal ganglia in segregated
information streams (Alexander et al., 1986; Dudman and Ger-
fen, 2015). Thus, cortical inputs could contribute to the creation
of discrete functional units in part defined by their inputs.
However, while cortical inputs are critical for generating spikes
in SPNs, it is unlikely that they are defining the units, based on
the current experimental evidence. Systematic tracing and map-
ping of cortical projections into the dorsal striatum has identified
29 distinct subregions, suggesting the existence of numerous
discrete zones of striatal processing (Hintiryan et al., 2016).
In congruence with the anatomy, the in vivo firing of neurons
in the dorsolateral striatum, which receives sensorimotor input,
correlates with specific body sensations, movements, or even
direction of movement of a specific limb (Carelli and West,
Neuron 96, October 11, 2017 275

Neuron
Perspective
1991; West et al., 1990). However, since cortical axons in the
striatum display a great deal of divergence and nearby SPNs in
general receive few shared cortical inputs (Kincaid et al., 1998),
it is unlikely that these inputs completely define striatal en-
sembles.
Furthermore, in vitro calcium imaging experiments have iden-
tified synchronized cell assemblies in the striatum active in the
presence of tonic excitatory input (Carrillo-Reid et al., 2008; Car-
rillo-Reid et al., 2011), demonstrating that glutamate is neces-
sary to generate activity in these cell assemblies. However,
blocking GABA abolishes much of the sequential activation
of the assemblies in the network, implying that local circuitry
within the striatum is important for the observed network
behavior of the cell assemblies. Note that similar sequentially
switching cell assemblies have been identified in the primate
striatum in vivo during behavior, suggesting that there is behav-
ioral relevance for the sequential activation pattern of the cell as-
semblies (Adler et al., 2012). Taken together, all these studies
support the idea that SPNs in the striatum are organized into
discrete functional units responsible for driving specific behav-
iors and that local striatal circuitry plays a role in their dynamics.
5. A Conceptual Model of Synaptic Organization of theLateral InhibitionIf indeed the striatum operates as a collection of functional units
that are responsible for driving specific behaviors, as the most
recent evidence seems to indicate, then how are these functional
units created and organized? What is the synaptic organization
of the functional units both within and between units that allow
for the selection of appropriate behaviors and other striatal func-
tions?We hypothesize that the lateral inhibition between SPNs is
critical in defining the functional units and that the organization of
the SPN collateral transmission within the striatum shapes this
lateral inhibition to define behaviorally relevant ensembles of
SPNs. This Perspective will mainly focus on the role of the
most abundant striatal neurons, SPNs; however, each functional
unit is likely to also contain interneurons that regulate activity
both within and between functional units.
In this section, we will use the current anatomical and
functional data on SPN collateral transmission to draw some
conceptual models and speculate how the connectivity pattern
of collateral transmission can bring functional relevance to the
lateral inhibition within the striatum in order to effectively aid in
the integration and filtering of inputs. A variety of theories on
striatal function emerged early on that incorporated the idea
that lateral inhibition shaped information flow through the
striatum (Parent et al., 2000; Smith and Bolam, 1990). Groves,
drawing on ‘‘winner-take-all’’ principles of lateral inhibition first
established in the retina, hypothesized that SPN collaterals proj-
ect to all their neighbors allowing the network to temporally and
spatially sharpen the broad input from the cortex and that dysre-
gulation in this system leads to movement disorders (Groves,
1983). Similarly, Penney and Young posited that lateral inhibition
between projection neurons (SPNs) served to suppress or main-
tain desired cortical input and hypothesized that the extent and
patterning of these axon collaterals may develop during acquisi-
tion of new motor and cognitive capacities (Penney and Young,
1983). Extending the idea that SPN collaterals sharpen and filter
276 Neuron 96, October 11, 2017
information into the ventral striatum, Swerdlow and Koob pro-
posed that in the NAc, a disruption of this circuitry, and the re-
sulting inability to filter emotional or cognitive inputs, could be
an underlying cause of psychiatric diseases like depression
and schizophrenia (Swerdlow and Koob, 1987).
Despite the fact that there is no easily visualized pattern of
SPN connectivity, several pieces of evidence point to an overall
structure: (1) the connection probability is asymmetrical between
SPN subclasses where iSPNs are more likely than dSPNs to be
connected to both dSPNs and other iSPNs, while dSPNs are
more likely to synapse on other dSPNs (Planert et al., 2010;
Taverna et al., 2008); (2) the synaptic contacts made by iSPNs
produce larger responses than those from dSPNs (Planert
et al., 2010; Taverna et al., 2004, 2008; Tunstall et al., 2002);
(3) connections are unidirectional in nature and sparse so they
do not just contact every nearby SPN (Planert et al., 2010;
Taverna et al., 2004, 2008; Tunstall et al., 2002); and (4) SPNs
synapse mainly onto distal dendrites of other SPNs, although
there is some diversity on location suggesting different types
of collateral synapses (Oorschot et al., 2013). These properties
suggest a more complex, nonrandom functional organization
of lateral inhibition in the striatum. They also argue strongly
against the idea of an all-to-all/winner-take-all network of lateral
inhibition in which the first, or most strongly activated, neuron is
able to inhibit all of its neighboring SPNs, like early models of
striatal function hypothesized (Groves, 1983).
The wiring diagram in Figure 4A illustrates a possible pattern of
connectivity between SPNs in an attempt to merge the current
findings at the behavioral level with the circuit level/anatomical
data. We know that (1) dSPN activation facilitates behavior, (2)
iSPN activation suppresses behavior, (3) both dSPNs and iSPNs
are active during behavior, and (4) there is collateral transmission
between SPNs. This supports a conceptual model in which each
behavior is controlled by a functional unit (FU, rectangles) con-
sisting of both dSPN and iSPN clusters (green and red circles),
where the dSPN cluster that drives the desired behavior (say,
behavior A) and the iSPN cluster that inhibits the competing
behavior (say, behavior B) are simultaneously active during a
task (dark colored circles), while the complementary clusters
are inactive (light colored circles) (Mink, 1996).
The model then incorporates recent evidence showing that
iSPN collateral transmission within the striatum is important for
setting the magnitude of the lateral inhibition onto dSPNs. It
proposes that the active cluster of iSPNs within FU2, which sup-
presses behavior B through long-range indirect-pathway projec-
tions (into the globus pallidus and ventral pallidum), also extends
intrastriatal collateral axons to inhibit the dSPN cluster (in the
same functional unit) and diminish its output through long-range
direct pathway projections to the midbrain, which would other-
wise promote the competing behavior B.
Paired recording data suggest that iSPNs are connected to a
third of nearby iSPNs, and so we hypothesize that the active
iSPN cluster in each functional unit also extends axon collaterals
that synapse onto the iSPN cluster in the competing functional
unit and therefore exerts lateral inhibition that limits any possible
inhibition of, in this case, the desired behavior A. Thus, under this
proposed organization of the collateral transmission, the active
iSPN cluster not only suppresses the activation of competing

A
B
Figure 4. Proposed Model of Organization of Striatal FunctionalUnits with Lateral Inhibition within and between Units(A) Colored circles represent multiple SPNs from each subclass that areactivated together during behavior forming a cluster or ensemble. Eachfunctional unit contains one dSPN cluster and one iSPN cluster. Clusters thatare active during execution of behavior A are highlighted and have strongoutput that enhances behavior A (green arrow) and suppresses competingbehavior B (red brake). Lateral inhibition from the active clusters further limitsactivity of the other silent dSPN and iSPN clusters.(B) Basic connectivity of the lateral inhibition within and between the twofunctional units described. Note that all clusters receive excitatory inputsas well. For simplicity, all interneurons are not included in this early concep-tual model.
Neuron
Perspective
behaviors but also suppresses the inhibition of the desired
behavior A.
With regards to the active dSPN cluster in FU1, the model also
incorporates the reported dSPN/dSPN collateral transmission
and the existence of weaker dSPN/iSPN transmission. The
model proposes that the active dSPN cluster in FU1 exerts
lateral inhibition onto the dSPN cluster in FU2 to further limit
the competing behavior B, and via the weaker connections to
the iSPN cluster in FU1, the lateral inhibition dampens the sup-
pression of the desired behavior A. In summary, the active
dSPN cluster in this FU1 promotes the desired behavior A via
the canonical long-range direct pathway projections, and, at
the same time via the axon collaterals that remainwithin the stria-
tum, it provides lateral inhibition within and across functional
units to limit competing behaviors and dampen the inhibition of
the desired behavior. This conceptual model will provide a
framework for future studies.
Recent work has shown the strength of local GABA transmis-
sion provided by iSPN axon collaterals can be modulated by
dopamine via activation of D2 receptors (Dobbs et al., 2016,
2017; Lemos et al., 2016). Acute activation of D2 receptors in
iSPNs suppresses the lateral inhibition onto dSPNs and chronic
loss of the D2 receptors in these neurons causes an increase in
the strength of the GABA-mediated tonic current on dSPNs,
which reduces the in vivo firing of SPNs and impairs locomotion
(Lemos et al., 2016). If we hypothesize that iSPN collaterals then
‘‘gate’’ dSPNs, and when collateral transmission is inhibited by
dopamine, dSPNs are better able to fire action potentials and
drive behavior (Dobbs et al., 2016).
How Can This Hypothesized Connectivity Pattern Aid in
the Processing of Inputs?
The impact that this proposed pattern of axon collateral trans-
mission has on the activity of the SPN clusters will have to be
determined and tested in future studies. To illustrate the possible
ways in which the connectivity patterns in this conceptual model
can aid in the processing and filtering of the excitatory inputs to
the striatum, we applied a standard conductance-based
formalism to the individual neurons and synapses that reflects
the main network properties rather than reproducing the under-
lying biology. In this simple model, all cells are identical and
receive identical inputs. We consider two patterns of synaptic
connectivity and determine how the different network scenarios
affect the firing of action potentials of the SPN clusters in the two
representative functional units.
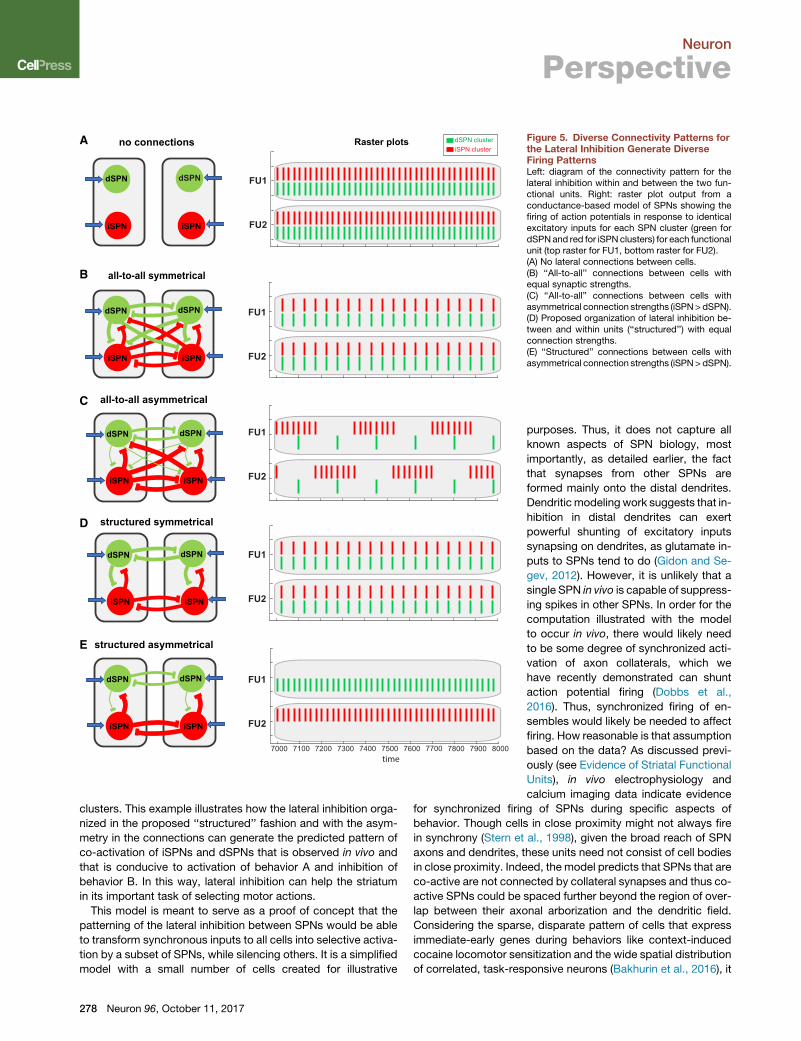
In a model without lateral inhibition, each SPN cluster fires ac-
tion potentials at relatively high frequency in response to excit-
atory inputs, and the clusters fire in synchrony because the
inputs are identical and the cells are homogeneous (Figure 5A).
Addition of lateral inhibition with an all-to-all connectivity pattern
and equal strength among connections (symmetric connectivity
patterns) results in synchronous firing at a lower frequency
(Figure 5B). Weakening transmission from dSPNs to introduce
the experimentally reported asymmetry in the connectivity
strength among SPN subclasses results in non-trivial firing pat-
terns where iSPN clusters become more dominant (Figure 5C).
iSPN clusters fire in anti-phase bursts and dSPN clusters fire a
single spike at every cycle. While it is interesting to see that func-
tional units are taking turns, the low activity of SPN clusters will
not be conducive to the generation of motor behaviors. Going
back to the pattern of Figure 5B, removal of the crossed connec-
tions between the functional leaves the originally proposed orga-
nization of the lateral inhibition (structured, Figure 5D), which
produces synchrony in all clusters, similar to the firing pattern
observed in Figure 5B (with slightly lower frequency). This illus-
trates the different balances that can be operating in the network
to produce the same type of patterns. However, when these
structured connections are no longer symmetrical between
SPN subclasses, the firing patterns can change drastically. As
shown in the example of Figure 5E, the asymmetric structured
pattern can result in synchronized activation of dSPN and iSPN
clusters belonging to different competing functional units where
iSPN and dSPN clusters fire in phase, while the other clusters are
silent. In other words, some clusters are shut down while others
are activated in phase. Note that the excitatory inputs to the si-
lent SPN clusters are still present and have not changed, and
as such this shows that, at least in this conceptual model, the
lateral inhibition can indeed filter excitatory inputs to SPNs to
shut down the firing of a selected subset of iSPN and dSPN
Neuron 96, October 11, 2017 277
no connections
all-to-all symmetrical
7000 7100 7200 7300 7400 7500 7600 7700 7800 7900 8000time
Raster plots dSPN clusteriSPN cluster
FU1
all-to-all asymmetrical
structured asymmetrical
structured symmetrical
A
B
E
D
C
FU2
FU1
FU2
FU1
FU2
FU1
FU2
FU1
FU2
dSPN dSPN
dSPN dSPN
dSPN dSPN
dSPN dSPN
dSPN dSPN
iSPN iSPN
iSPN iSPN
iSPN iSPN
iSPN iSPN
iSPN iSPN
Figure 5. Diverse Connectivity Patterns forthe Lateral Inhibition Generate DiverseFiring PatternsLeft: diagram of the connectivity pattern for thelateral inhibition within and between the two fun-ctional units. Right: raster plot output from aconductance-based model of SPNs showing thefiring of action potentials in response to identicalexcitatory inputs for each SPN cluster (green fordSPNand red for iSPN clusters) for each functionalunit (top raster for FU1, bottom raster for FU2).(A) No lateral connections between cells.(B) ‘‘All-to-all’’ connections between cells withequal synaptic strengths.(C) ‘‘All-to-all’’ connections between cells withasymmetrical connection strengths (iSPN> dSPN).(D) Proposed organization of lateral inhibition be-tween and within units (‘‘structured’’) with equalconnection strengths.(E) ‘‘Structured’’ connections between cells withasymmetrical connection strengths (iSPN> dSPN).
Neuron
Perspective
clusters. This example illustrates how the lateral inhibition orga-
nized in the proposed ‘‘structured’’ fashion and with the asym-
metry in the connections can generate the predicted pattern of
co-activation of iSPNs and dSPNs that is observed in vivo and
that is conducive to activation of behavior A and inhibition of
behavior B. In this way, lateral inhibition can help the striatum
in its important task of selecting motor actions.
This model is meant to serve as a proof of concept that the
patterning of the lateral inhibition between SPNs would be able
to transform synchronous inputs to all cells into selective activa-
tion by a subset of SPNs, while silencing others. It is a simplified
model with a small number of cells created for illustrative
278 Neuron 96, October 11, 2017
purposes. Thus, it does not capture all
known aspects of SPN biology, most
importantly, as detailed earlier, the fact
that synapses from other SPNs are
formed mainly onto the distal dendrites.
Dendriticmodeling work suggests that in-
hibition in distal dendrites can exert
powerful shunting of excitatory inputs
synapsing on dendrites, as glutamate in-
puts to SPNs tend to do (Gidon and Se-
gev, 2012). However, it is unlikely that a
single SPN in vivo is capable of suppress-
ing spikes in other SPNs. In order for the
computation illustrated with the model
to occur in vivo, there would likely need
to be some degree of synchronized acti-
vation of axon collaterals, which we
have recently demonstrated can shunt
action potential firing (Dobbs et al.,
2016). Thus, synchronized firing of en-
sembles would likely be needed to affect
firing. How reasonable is that assumption
based on the data? As discussed previ-
ously (see Evidence of Striatal Functional
Units), in vivo electrophysiology and
calcium imaging data indicate evidence
for synchronized firing of SPNs during specific aspects of
behavior. Though cells in close proximity might not always fire
in synchrony (Stern et al., 1998), given the broad reach of SPN
axons and dendrites, these units need not consist of cell bodies
in close proximity. Indeed, the model predicts that SPNs that are
co-active are not connected by collateral synapses and thus co-
active SPNs could be spaced further beyond the region of over-
lap between their axonal arborization and the dendritic field.
Considering the sparse, disparate pattern of cells that express
immediate-early genes during behaviors like context-induced
cocaine locomotor sensitization and the wide spatial distribution
of correlated, task-responsive neurons (Bakhurin et al., 2016), it

Neuron
Perspective
seems likely that the cells in these units would, in fact, not be in
close proximity (but note Barbera et al., 2016).
Another possibility that has been suggested (Koos et al., 2004;
Plenz, 2003) is that local collaterals of SPNs are important for
controlling transition to depolarized, plateau ‘‘upstates,’’ which
prime SPNs for firing, rather than affecting the spiking itself. As
SPN collaterals synapse mainly on distal dendrites (Oorschot
et al., 2013), and plateau upstate potentials are generated by
regenerative events in the distal dendrites (Plotkin et al., 2011),
GABA released from SPN could provide sufficient shunting to
prevent this transition. Indeed, in vitro studies show that GABA
antagonists prolong SPN upstate duration, suggesting that
GABA inputs play a role in upstate timing (Vergara et al., 2003).
By controlling the upstate transitions that precede spiking and
bursting, as well as directly shunting excitatory inputs, inhibition
in the dendrites would still provide a powerful means of effecting
cell output (Gidon and Segev, 2012; Lovett-Barron et al., 2012).
Thus, the local SPN synapses between and within units could be
controlling the excitability of neighboring cells in the way pro-
posed by the model, without necessarily directly suppressing
spiking.
Open Questions
This proposed model is intended as a preliminary proposition to
stimulate discussion and research regarding how local circuitry
within the striatum might be organized given known functional
and behavioral data. As a result, there are many open questions
regarding this framework. Howmany cells do these units consist
of? How much overlap is there between units? How might these
units arise? Are they fixed developmentally, completely plastic
based on learning, or somewhere in between? Given the known
role of the striatum in certain forms of learning and the numerous
examples of plasticity in the region, it is likely that any organiza-
tion in the region is malleable. Dopamine release during behavior
could modulate the strength of the lateral inhibition to alter the
connectivity pattern in ways that it favors the activation of
selected FUs and the inhibition of others to ultimately raise the
signal-to-noise ratio and reinforce desirable behaviors. To what
degree is this the case, and are certain units more flexible and
plastic than others? Furthermore, how would the various inter-
neurons and their numerous known synapses detailed earlier
fit within this functional unit framework?
Concluding RemarksGiven the growing diversity of striatal inputs, cell types, and
connections described in recent years, it is important to develop
a framework for how the circuitry within the striatum is wired to
drive processing of information and affect output. Here we pre-
sent a hypothesis on how one aspect of the striatal circuitry, the
lateral inhibition between SPNs, could be organized given avail-
able data. It is our hope that this will drive the future work needed
tobothdetermine the validity of thismodel and to understandhow
other elements of striatal circuitry could fit within this framework.
ACKNOWLEDGMENTS
This work was supported by the Intramural Programs of National Institute onAlcohol Abuse (ZIA-AA000421) and Alcoholism and National Institute ofNeurological Disorders and Stroke Grant (ZIA-AA000421) to V.A.A. and bythe National Science Foundation (DMS-1313861) to H.G.R. We are grateful
to Dr. Miriam E. Bocarsly and the other members of the Alvarez laboratoryfor their valuable comments on this Perspective.
REFERENCES
Adler, A., Katabi, S., Finkes, I., Israel, Z., Prut, Y., and Bergman, H. (2012).Temporal convergence of dynamic cell assemblies in the striato-pallidalnetwork. J. Neurosci. 32, 2473–2484.
Albin, R.L., Young, A.B., and Penney, J.B. (1989). The functional anatomy ofbasal ganglia disorders. Trends Neurosci. 12, 366–375.
Alexander, G.E., and Crutcher, M.D. (1990). Functional architecture of basalganglia circuits: neural substrates of parallel processing. Trends Neurosci.13, 266–271.
Alexander, G.E., DeLong, M.R., and Strick, P.L. (1986). Parallel organization offunctionally segregated circuits linking basal ganglia and cortex. Annu. Rev.Neurosci. 9, 357–381.
Alvarez, V.A. (2016). Clues on the coding of reward cues by the nucleusaccumbens. Proc. Natl. Acad. Sci. USA 113, 2560–2562.
Bakhurin, K.I., Mac, V., Golshani, P., and Masmanidis, S.C. (2016). Temporalcorrelations among functionally specialized striatal neural ensembles inreward-conditioned mice. J. Neurophysiol. 115, 1521–1532.
Bamford, N.S., Zhang, H., Schmitz, Y., Wu, N.P., Cepeda, C., Levine, M.S.,Schmauss, C., Zakharenko, S.S., Zablow, L., and Sulzer, D. (2004). Heterosy-naptic dopamine neurotransmission selects sets of corticostriatal terminals.Neuron 42, 653–663.
Banghart, M.R., Neufeld, S.Q., Wong, N.C., and Sabatini, B.L. (2015). Enkeph-alin disinhibits mu opioid receptor-rich striatal patches via delta opioidreceptors. Neuron 88, 1227–1239.
Barbera, G., Liang, B., Zhang, L., Gerfen, C.R., Culurciello, E., Chen, R., Li, Y.,and Lin, D.-T. (2016). Spatially compact neural clusters in the dorsal striatumencode locomotion relevant information. Neuron 92, 202–213.
Barnes, T.D., Kubota, Y., Hu, D., Jin, D.Z., andGraybiel, A.M. (2005). Activity ofstriatal neurons reflects dynamic encoding and recoding of proceduralmemories. Nature 437, 1158–1161.
Bateup, H.S., Santini, E., Shen, W., Birnbaum, S., Valjent, E., Surmeier, D.J.,Fisone, G., Nestler, E.J., and Greengard, P. (2010). Distinct subclasses ofmedium spiny neurons differentially regulate striatal motor behaviors. Proc.Natl. Acad. Sci. USA 107, 14845–14850.
Bertran-Gonzalez, J., Bosch, C., Maroteaux, M., Matamales, M., Herve, D.,Valjent, E., and Girault, J.-A. (2008). Opposing patterns of signaling activationin dopamine D1 and D2 receptor-expressing striatal neurons in response tococaine and haloperidol. J. Neurosci. 28, 5671–5685.
Bevan, M.D., Booth, P.A., Eaton, S.A., and Bolam, J.P. (1998). Selective inner-vation of neostriatal interneurons by a subclass of neuron in the globus pallidusof the rat. J. Neurosci. 18, 9438–9452.
Bock, R., Shin, J.H., Kaplan, A.R., Dobi, A., Markey, E., Kramer, P.F., Gremel,C.M., Christensen, C.H., Adrover, M.F., and Alvarez, V.A. (2013). Strength-ening the accumbal indirect pathway promotes resilience to compulsivecocaine use. Nat. Neurosci. 16, 632–638.
Bolam, J.P., and Ellender, T.J. (2016). Histamine and the striatum. Neurophar-macology 106, 74–84.
Bolam, J.P., Hanley, J.J., Booth, P.A.C., and Bevan, M.D. (2000). Synapticorganisation of the basal ganglia. J. Anat. 196, 527–542.
Brimblecombe, K.R., and Cragg, S.J. (2017). The striosome and matrix com-partments of the striatum: a path through the labyrinth from neurochemistrytowards function. ACS Chem. Neurosci. 8, 235–242.
Britt, J.P., Benaliouad, F., McDevitt, R.A., Stuber, G.D., Wise, R.A., and Bonci,A. (2012). Synaptic and behavioral profile of multiple glutamatergic inputs tothe nucleus accumbens. Neuron 76, 790–803.
Brown, M.T.C., Tan, K.R., O’Connor, E.C., Nikonenko, I., Muller, D., andL€uscher, C. (2012). Ventral tegmental area GABA projections pause accumbal
Neuron 96, October 11, 2017 279

Neuron
Perspective
cholinergic interneurons to enhance associative learning. Nature 492,452–456.
Bubser, M., and Deutch, A.Y. (1998). Thalamic paraventricular nucleus neu-rons collateralize to innervate the prefrontal cortex and nucleus accumbens.Brain Res. 787, 304–310.
Cacciapaglia, F., Wightman, R.M., and Carelli, R.M. (2011). Rapid dopaminesignaling differentially modulates distinct microcircuits within the nucleusaccumbens during sucrose-directed behavior. J. Neurosci. 31, 13860–13869.
Cachope, R., Mateo, Y., Mathur, B.N., Irving, J., Wang, H.L., Morales, M.,Lovinger, D.M., and Cheer, J.F. (2012). Selective activation of cholinergic inter-neurons enhances accumbal phasic dopamine release: setting the tone forreward processing. Cell Rep. 2, 33–41.
Calabresi, P., Picconi, B., Tozzi, A., Ghiglieri, V., and Di Filippo, M. (2014).Direct and indirect pathways of basal ganglia: a critical reappraisal. Nat.Neurosci. 17, 1022–1030.
Calipari, E.S., Bagot, R.C., Purushothaman, I., Davidson, T.J., Yorgason, J.T.,Pena, C.J., Walker, D.M., Pirpinias, S.T., Guise, K.G., Ramakrishnan, C., et al.(2016). In vivo imaging identifies temporal signature of D1 and D2 mediumspiny neurons in cocaine reward. Proc. Natl. Acad. Sci. USA 113, 2726–2731.
Carelli, R.M. (2002). Nucleus accumbens cell firing during goal-directed be-haviors for cocaine vs. ‘natural’ reinforcement. Physiol. Behav. 76, 379–387.
Carelli, R.M., and West, M.O. (1991). Representation of the body by singleneurons in the dorsolateral striatum of the awake, unrestrained rat. J. Comp.Neurol. 309, 231–249.
Carrillo-Reid, L., Tecuapetla, F., Tapia, D., Hernandez-Cruz, A., Galarraga, E.,Drucker-Colin, R., and Bargas, J. (2008). Encoding network states by striatalcell assemblies. J. Neurophysiol. 99, 1435–1450.
Carrillo-Reid, L., Hernandez-Lopez, S., Tapia, D., Galarraga, E., and Bargas, J.(2011). Dopaminergic modulation of the striatal microcircuit: receptor-specificconfiguration of cell assemblies. J. Neurosci. 31, 14972–14983.
Cazorla, M., de Carvalho, F.D., Chohan, M.O., Shegda, M., Chuhma, N.,Rayport, S., Ahmari, S.E., Moore, H., and Kellendonk, C. (2014). DopamineD2 receptors regulate the anatomical and functional balance of basal gangliacircuitry. Neuron 81, 153–164.
Cazorla, M., Kang, U.J., and Kellendonk, C. (2015). Balancing the basal gangliacircuitry: a possible new role for dopamine D2 receptors in health and disease.Mov. Disord. 30, 895–903.
Chang, H.T., and Kitai, S.T. (1985). Projection neurons of the nucleus accum-bens: an intracellular labeling study. Brain Res. 347, 112–116.
Chuhma, N., Tanaka, K.F., Hen, R., and Rayport, S. (2011). Functional connec-tome of the striatal medium spiny neuron. J. Neurosci. 31, 1183–1192.
Cohen, M.X., and Frank, M.J. (2009). Neurocomputational models of basalganglia function in learning, memory and choice. Behav. Brain Res. 199,141–156.
Collins, A.G.E., and Frank, M.J. (2014). Opponent actor learning (OpAL):modeling interactive effects of striatal dopamine on reinforcement learningand choice incentive. Psychol. Rev. 121, 337–366.
Crittenden, J.R., and Graybiel, A.M. (2011). Basal Ganglia disorders associ-atedwith imbalances in the striatal striosome andmatrix compartments. Front.Neuroanat. 5, 59.
Cui, G., Jun, S.B., Jin, X., Pham, M.D., Vogel, S.S., Lovinger, D.M., and Costa,R.M. (2013). Concurrent activation of striatal direct and indirect pathways dur-ing action initiation. Nature 494, 238–242.
Czubayko, U., and Plenz, D. (2002). Fast synaptic transmission between stria-tal spiny projection neurons. Proc. Natl. Acad. Sci. USA 99, 15764–15769.
Dautan, D., Huerta-Ocampo, I., Witten, I.B., Deisseroth, K., Bolam, J.P., Gerd-jikov, T., and Mena-Segovia, J. (2014). A major external source of cholinergicinnervation of the striatum and nucleus accumbens originates in the brainstem.J. Neurosci. 34, 4509–4518.
DeLong, M.R. (1990). Primate models of movement disorders of basal gangliaorigin. Trends Neurosci. 13, 281–285.
280 Neuron 96, October 11, 2017
Ding, J., Peterson, J.D., and Surmeier, D.J. (2008). Corticostriatal and thala-mostriatal synapses have distinctive properties. J. Neurosci. 28, 6483–6492.
Ding, J.B., Guzman, J.N., Peterson, J.D., Goldberg, J.A., and Surmeier, D.J.(2010). Thalamic gating of corticostriatal signaling by cholinergic interneurons.Neuron 67, 294–307.
Dobbs, L.K., Kaplan, A.R., Lemos, J.C., Matsui, A., Rubinstein, M., and Al-varez, V.A. (2016). Dopamine regulation of lateral inhibition between striatalneurons gates the stimulant actions of cocaine. Neuron 90, 1100–1113.
Dobbs, L.K., Lemos, J.C., and Alvarez, V.A. (2017). Restructuring of basalganglia circuitry and associated behaviors triggered by low striatal D2 receptorexpression: implications for substance use disorders. Genes Brain Behav.16, 56–70.
Doig, N.M., Moss, J., and Bolam, J.P. (2010). Cortical and thalamic innervationof direct and indirect pathway medium-sized spiny neurons in mouse striatum.J. Neurosci. 30, 14610–14618.
Dolen, G., Darvishzadeh, A., Huang, K.W., and Malenka, R.C. (2013). Socialreward requires coordinated activity of nucleus accumbens oxytocin and se-rotonin. Nature 501, 179–184.
Dudman, J.T., and Gerfen, C.R. (2015). The basal ganglia. In The Rat NervousSystem, G. Paxinos, ed. (Elsevier), pp. 391–440.
Durieux, P.F., Bearzatto, B., Guiducci, S., Buch, T., Waisman, A., Zoli, M.,Schiffmann, S.N., and de Kerchove d’Exaerde, A. (2009). D2R striatopallidalneurons inhibit both locomotor and drug reward processes. Nat. Neurosci.12, 393–395.
Elghaba, R., Vautrelle, N., and Bracci, E. (2016). Mutual control of cholinergicand low-threshold spike interneurons in the striatum. Front. Cell. Neurosci.10, 111.
Ellender, T.J., Harwood, J., Kosillo, P., Capogna, M., and Bolam, J.P. (2013).Heterogeneous properties of central lateral and parafascicular thalamic syn-apses in the striatum. J. Physiol. 591, 257–272.
English, D.F., Ibanez-Sandoval, O., Stark, E., Tecuapetla, F., Buzsaki, G.,Deisseroth, K., Tepper, J.M., and Koos, T. (2011). GABAergic circuits mediatethe reinforcement-related signals of striatal cholinergic interneurons. Nat. Neu-rosci. 15, 123–130.
Faust, T.W., Assous,M., Shah, F., Tepper, J.M., and Koos, T. (2015). Novel fastadapting interneurons mediate cholinergic-induced fast GABAA inhibitorypostsynaptic currents in striatal spiny neurons. Eur. J. Neurosci. 42,1764–1774.
Faust, T.W., Assous, M., Tepper, J.M., and Koos, T. (2016). NeostriatalGABAergic interneuronsmediate cholinergic inhibition of spiny projection neu-rons. J. Neurosci. 36, 9505–9511.
Frank, M.J. (2005). Dynamic dopamine modulation in the basal ganglia: a neu-rocomputational account of cognitive deficits in medicated and nonmedicatedParkinsonism. J. Cogn. Neurosci. 17, 51–72.
Gagnon, D., Petryszyn, S., Sanchez, M.G., Bories, C., Beaulieu, J.M.,De Koninck, Y., Parent, A., and Parent, M. (2017). Striatal neurons expressingD1 and D2 Receptors are morphologically distinct and differently affected bydopamine denervation in mice. Sci. Rep. 7, 41432.
Gerfen, C.R. (1984). The neostriatal mosaic: compartmentalization of cortico-striatal input and striatonigral output systems. Nature 311, 461–464.
Gerfen, C.R., and Surmeier, D.J. (2011). Modulation of striatal projectionsystems by dopamine. Annu. Rev. Neurosci. 34, 441–466.
Gerfen, C.R.C., Engber, T.M., Mahan, L.C., Susel, Z., Chase, T.N., Monsma,F.J., Jr., and Sibley, D.R. (1990). D1 and D2 dopamine receptor-regulatedgene expression of striatonigral and striatopallidal neurons. Science 250,1429–1432.
Gidon, A., and Segev, I. (2012). Principles governing the operation of synapticinhibition in dendrites. Neuron 75, 330–341.
Gittis, A.H., Nelson, A.B., Thwin, M.T., Palop, J.J., and Kreitzer, A.C. (2010).Distinct roles of GABAergic interneurons in the regulation of striatal outputpathways. J. Neurosci. 30, 2223–2234.

Neuron
Perspective
Gokce, O., Stanley, G., Treutlein, B., Neff, N.F., Camp, G.J., Malenka, R.C.,Rothwell, P.E., Fuccillo, M.V., S€udhof, T.C., and Quake, S.R. (2016). Cellulartaxonomy of the mouse striatum as revealed by single-cell RNA-seq. CellRep. 16, 1126–1137.
Gong, S., Zheng, C., Doughty,M.L., Losos, K., Didkovsky, N., Schambra, U.B.,Nowak, N.J., Joyner, A., Leblanc, G., Hatten, M.E., and Heintz, N. (2003). Agene expression atlas of the central nervous system based on bacterial artifi-cial chromosomes. Nature 425, 917–925.
Gonzales, K.K., and Smith, Y. (2015). Cholinergic interneurons in the dorsaland ventral striatum: anatomical and functional considerations in normal anddiseased conditions. Ann. N Y Acad. Sci. 1349, 1–45.
Graveland, G.A., and DiFiglia, M. (1985). The frequency and distribution ofmedium-sized neurons with indented nuclei in the primate and rodent neo-striatum. Brain Res. 327, 307–311.
Grofova, I. (1975). The identification of striatal and pallidal neurons projectingto substantia nigra. An experimental study by means of retrograde axonaltransport of horseradish peroxidase. Brain Res. 91, 286–291.
Groves, P.M. (1983). A theory of the functional organization of the neostriatumand the neostriatal control of voluntary movement. Brain Res. 286, 109–132.
Guo, Q., Wang, D., He, X., Feng, Q., Lin, R., Xu, F., Fu, L., and Luo, M. (2015).Whole-brain mapping of inputs to projection neurons and cholinergic interneu-rons in the dorsal striatum. PLoS ONE 10, e0123381.
Guzman, M.S., De Jaeger, X., Raulic, S., Souza, I.A., Li, A.X., Schmid, S.,Menon, R.S., Gainetdinov, R.R., Caron, M.G., Bartha, R., et al. (2011). Elimina-tion of the vesicular acetylcholine transporter in the striatum reveals regulationof behaviour by cholinergic-glutamatergic co-transmission. PLoS Biol. 9,e1001194.
Haber, S.N., Fudge, J.L., and McFarland, N.R. (2000). Striatonigrostriatalpathways in primates form an ascending spiral from the shell to the dorsolat-eral striatum. J. Neurosci. 20, 2369–2382.
Higley, M.J., and Sabatini, B.L. (2010). Competitive regulation of synapticCa2+ influx by D2 dopamine and A2A adenosine receptors. Nat. Neurosci.13, 958–966.
Higley, M.J., Gittis, A.H., Oldenburg, I.A., Balthasar, N., Seal, R.P., Edwards,R.H., Lowell, B.B., Kreitzer, A.C., and Sabatini, B.L. (2011). Cholinergic inter-neurons mediate fast VGluT3-dependent glutamatergic transmission in thestriatum. PLoS ONE 6, e19155.
Hikida, T., Kimura, K.,Wada, N., Funabiki, K., and Nakanishi, S. (2010). Distinctroles of synaptic transmission in direct and indirect striatal pathways to rewardand aversive behavior. Neuron 66, 896–907.
Hikosaka, O. (1991). Basal ganglia–possible role in motor coordination andlearning. Curr. Opin. Neurobiol. 1, 638–643.
Hikosaka, O. (1998). Neural systems for control of voluntary action–a hypoth-esis. Adv. Biophys. 35, 81–102.
Hintiryan, H., Foster, N.N., Bowman, I., Bay, M., Song, M.Y., Gou, L., Yama-shita, S., Bienkowski, M.S., Zingg, B., Zhu, M., et al. (2016). The mousecortico-striatal projectome. Nat. Neurosci. 19, 1100–1114.
Hnasko, T.S., Chuhma, N., Zhang, H., Goh, G.Y., Sulzer, D., Palmiter, R.D.,Rayport, S., and Edwards, R.H. (2010). Vesicular glutamate transport pro-motes dopamine storage and glutamate corelease in vivo. Neuron 65,643–656.
Hnasko, T.S., Hjelmstad, G.O., Fields, H.L., and Edwards, R.H. (2012). Ventraltegmental area glutamate neurons: electrophysiological properties and pro-jections. J. Neurosci. 32, 15076–15085.
Hope, B.T., Simmons, D.E., Mitchell, T.B., Kreuter, J.D., and Mattson, B.J.(2006). Cocaine-induced locomotor activity and Fos expression in nucleus ac-cumbens are sensitized for 6 months after repeated cocaine administrationoutside the home cage. Eur. J. Neurosci. 24, 867–875.
Humphries, M.D., Wood, R., and Gurney, K. (2009). Dopamine-modulated dy-namic cell assemblies generated by the GABAergic striatal microcircuit. Neu-ral Netw. 22, 1174–1188.
Humphries, M.D., Wood, R., and Gurney, K. (2010). Reconstructing the three-dimensional GABAergic microcircuit of the striatum. PLoS Comput. Biol. 6,e1001011.
Ibanez-Sandoval, O., Tecuapetla, F., Unal, B., Shah, F., Koos, T., and Tepper,J.M. (2010). Electrophysiological and morphological characteristics and syn-aptic connectivity of tyrosine hydroxylase-expressing neurons in adult mousestriatum. J. Neurosci. 30, 6999–7016.
Ibanez-Sandoval, O., Tecuapetla, F., Unal, B., Shah, F., Koos, T., and Tepper,J.M. (2011). A novel functionally distinct subtype of striatal neuropeptideY interneuron. J. Neurosci. 31, 16757–16769.
Ibanez-Sandoval, O., Xenias, H.S., Tepper, J.M., and Koos, T. (2015). Dopami-nergic and cholinergic modulation of striatal tyrosine hydroxylase interneu-rons. Neuropharmacology 95, 468–476.
Isomura, Y., Takekawa, T., Harukuni, R., Handa, T., Aizawa, H., Takada, M.,and Fukai, T. (2013). Reward-modulated motor information in identified stria-tum neurons. J. Neurosci. 33, 10209–10220.
Jaeger, D., Kita, H., andWilson, C.J. (1994). Surround inhibition among projec-tion neurons is weak or nonexistent in the rat neostriatum. J. Neurophysiol. 72,2555–2558.
Jin, X., Tecuapetla, F., and Costa, R.M. (2014). Basal ganglia subcircuitsdistinctively encode the parsing and concatenation of action sequences.Nat. Neurosci. 17, 423–430.
Kaiser, S., and Wonnacott, S. (2000). a-bungarotoxin-sensitive nicotinic re-ceptors indirectly modulate [(3)H]dopamine release in rat striatal slices viaglutamate release. Mol. Pharmacol. 58, 312–318.
Kawaguchi, Y., Wilson, C.J., and Emson, P.C. (1990). Projection subtypes ofrat neostriatal matrix cells revealed by intracellular injection of biocytin.J. Neurosci. 10, 3421–3438.
Kemp, J.M., and Powell, T.P. (1971). The structure of the caudate nucleus ofthe cat: light and electron microscopy. Philos. Trans. R. Soc. Lond. B Biol.Sci. 262, 383–401.
Kim, J.-I., Ganesan, S., Luo, S.X., Wu, Y.-W., Park, E., Huang, E.J., Chen, L.,and Ding, J.B. (2015). Aldehyde dehydrogenase 1a1mediates aGABA synthe-sis pathway in midbrain dopaminergic neurons. Science 350, 102–106.
Kincaid, A.E., Zheng, T., and Wilson, C.J. (1998). Connectivity and conver-gence of single corticostriatal axons. J. Neurosci. 18, 4722–4731.
Koos, T., and Tepper, J.M. (1999). Inhibitory control of neostriatal projectionneurons by GABAergic interneurons. Nat. Neurosci. 2, 467–472.
Koos, T., and Tepper, J.M. (2002). Dual cholinergic control of fast-spiking in-terneurons in the neostriatum. J. Neurosci. 22, 529–535.
Koos, T., Tepper, J.M., and Wilson, C.J. (2004). Comparison of IPSCs evokedby spiny and fast-spiking neurons in the neostriatum. J. Neurosci. 24,7916–7922.
Kosillo, P., Zhang, Y.-F., Threlfell, S., and Cragg, S.J. (2016). Cortical Control ofStriatal Dopamine Transmission via Striatal Cholinergic Interneurons. Cereb.Cortex. Published online October 17, 2016. https://doi.org/10.1093/cercor/bhw252.
Koya, E., Golden, S.A., Harvey, B.K., Guez-Barber, D.H., Berkow, A., Sim-mons, D.E., Bossert, J.M., Nair, S.G., Uejima, J.L., Marin, M.T., et al. (2009).Targeted disruption of cocaine-activated nucleus accumbens neurons pre-vents context-specific sensitization. Nat. Neurosci. 12, 1069–1073.
Kravitz, A.V., Freeze, B.S., Parker, P.R.L., Kay, K., Thwin, M.T., Deisseroth, K.,and Kreitzer, A.C. (2010). Regulation of parkinsonian motor behaviours byoptogenetic control of basal ganglia circuitry. Nature 466, 622–626.
Kravitz, A.V., Tye, L.D., and Kreitzer, A.C. (2012). Distinct roles for direct andindirect pathway striatal neurons in reinforcement. Nat. Neurosci. 15, 816–818.
Kreitzer, A.C., and Malenka, R.C. (2008). Striatal plasticity and basal gangliacircuit function. Neuron 60, 543–554.
Kress, G.J., Yamawaki, N., Wokosin, D.L., Wickersham, I.R., Shepherd,G.M.G., and Surmeier, D.J. (2013). Convergent cortical innervation of striatalprojection neurons. Nat. Neurosci. 16, 665–667.
Neuron 96, October 11, 2017 281

Neuron
Perspective
Kupchik, Y.M., Brown, R.M., Heinsbroek, J.A., Lobo, M.K., Schwartz, D.J.,and Kalivas, P.W. (2015). Coding the direct/indirect pathways by D1 and D2receptors is not valid for accumbens projections. Nat. Neurosci. 18,1230–1232.
Lee, A.T., Vogt, D., Rubenstein, J.L., and Sohal, V.S. (2014). A class ofGABAergic neurons in the prefrontal cortex sends long-range projections tothe nucleus accumbens and elicits acute avoidance behavior. J. Neurosci.34, 11519–11525.
Lee, K., Holley, S.M., Shobe, J.L., Chong, N.C., Cepeda, C., Levine, M.S., andMasmanidis, S.C. (2017). Parvalbumin interneurons modulate striatal outputand enhance performance during associative learning. Neuron 93, 1451–1463.e4.
Lei, W., Jiao, Y., Del Mar, N., and Reiner, A. (2004). Evidence for differentialcortical input to direct pathway versus indirect pathway striatal projection neu-rons in rats. J. Neurosci. 24, 8289–8299.
Lemos, J.C., Friend, D.M., Kaplan, A.R., Shin, J.H., Rubinstein, M., Kravitz,A.V., and Alvarez, V.A. (2016). EnhancedGABA transmission drives bradykine-sia following loss of dopamine D2 receptor signaling. Neuron 90, 824–838.
Lobo, M.K., Covington, H.E., 3rd, Chaudhury, D., Friedman, A.K., Sun, H.,Damez-Werno, D., Dietz, D.M., Zaman, S., Koo, J.W., Kennedy, P.J., et al.(2010). Cell type-specific loss of BDNF signaling mimics optogenetic controlof cocaine reward. Science 330, 385–390.
Logie, C., Bagetta, V., and Bracci, E. (2013). Presynaptic control of cortico-striatal synapses by endogenous GABA. J. Neurosci. 33, 15425–15431.
Lovett-Barron, M., Turi, G.F., Kaifosh, P., Lee, P.H., Bolze, F., Sun, X.-H., Nic-oud, J.-F., Zemelman, B.V., Sternson, S.M., and Losonczy, A. (2012). Regula-tion of neuronal input transformations by tunable dendritic inhibition. Nat.Neurosci. 15, 423–430, S1–S3.
Luo, R., Janssen, M.J., Partridge, J.G., and Vicini, S. (2013). Direct andGABA-mediated indirect effects of nicotinic ACh receptor agonists on striatalneurones. J. Physiol. 591, 203–217.
MacAskill, A.F., Little, J.P., Cassel, J.M., and Carter, A.G. (2012). Subcellularconnectivity underlies pathway-specific signaling in the nucleus accumbens.Nat. Neurosci. 15, 1624–1626.
Mallet, N., Le Moine, C., Charpier, S., and Gonon, F. (2005). Feedforward inhi-bition of projection neurons by fast-spiking GABA interneurons in the rat stria-tum in vivo. J. Neurosci. 25, 3857–3869.
Mallet, N., Micklem, B.R., Henny, P., Brown, M.T., Williams, C., Bolam, J.P.,Nakamura, K.C., and Magill, P.J. (2012). Dichotomous organization of theexternal globus pallidus. Neuron 74, 1075–1086.
Mallet, N., Schmidt, R., Leventhal, D., Chen, F., Amer, N., Boraud, T., andBerke, J.D. (2016). Arkypallidal Cells Send a Stop Signal to Striatum. Neuron89, 308–316.
Martone, M.E., Armstrong, D.M., Young, S.J., and Groves, P.M. (1992). Ultra-structural examination of enkephalin and substance P input to cholinergic neu-rons within the rat neostriatum. Brain Res. 594, 253–262.
Miguelez, C., Morera-Herreras, T., Torrecilla, M., Ruiz-Ortega, J.A., andUgedo, L. (2014). Interaction between the 5-HT system and the basal ganglia:functional implication and therapeutic perspective in Parkinson’s disease.Front. Neural Circuits 8, 21.
Miller, B.R., Walker, A.G., Shah, A.S., Barton, S.J., Rebec, G.V., Br, M., Ag, W.,As, S., Sj, B., and Gv, R. (2008). Dysregulated information processing bymedium spiny neurons in striatum of freely behaving mouse models ofHuntington’s disease. J. Neurophysiol. 100, 2205–2216.
Mink, J.W. (1996). The basal ganglia: focused selection and inhibition ofcompeting motor programs. Prog. Neurobiol. 50, 381–425.
Mink, J.W. (2003). The Basal Ganglia and involuntary movements: impaired in-hibition of competing motor patterns. Arch. Neurol. 60, 1365–1368.
Mink, J.W., and Thach, W.T. (1993). Basal ganglia intrinsic circuits and theirrole in behavior. Curr. Opin. Neurobiol. 3, 950–957.
Moyer, J.T., Halterman, B.L., Finkel, L.H., and Wolf, J.A. (2014). Lateraland feedforward inhibition suppress asynchronous activity in a large,
282 Neuron 96, October 11, 2017
biophysically-detailed computational model of the striatal network. Front.Comput. Neurosci. 8, 152.
Munoz-Manchado, A.B., Foldi, C., Szydlowski, S., Sjulson, L., Farries, M., Wil-son, C., Silberberg, G., and Hjerling-Leffler, J. (2016). Novel striatal GABAergicinterneuron populations labeled in the 5HT3a(EGFP) mouse. Cereb. Cortex26, 96–105.
Natsubori, A., Tsutsui-Kimura, I., Nishida, H., Bouchekioua, Y., Sekiya, H.,Uchigashima, M., Watanabe, M., de Kerchove d’Exaerde, A., Mimura, M.,Takata, N., and Tanaka, K.F. (2017). Ventrolateral striatal medium spiny neu-rons positively regulate food-incentive, goal-directed behavior independentlyof D1 and D2 selectivity. J. Neurosci. 37, 2723–2733.
Nelson, A.B., Bussert, T.G., Kreitzer, A.C., and Seal, R.P. (2014a). Striatalcholinergic neurotransmission requires VGLUT3. J. Neurosci. 34, 8772–8777.
Nelson, A.B., Hammack, N., Yang, C.F., Shah, N.M., Seal, R.P., and Kreitzer,A.C. (2014b). Striatal cholinergic interneurons Drive GABA release from dopa-mine terminals. Neuron 82, 63–70.
Oldenburg, I.A., and Ding, J.B. (2011). Cholinergic modulation of synaptic inte-gration and dendritic excitability in the striatum. Curr. Opin. Neurobiol. 21,425–432.
Oorschot, D.E. (1996). Total number of neurons in the neostriatal, pallidal, sub-thalamic, and substantia nigral nuclei of the rat basal ganglia: a stereologicalstudy using the cavalieri and optical disector methods. J. Comp. Neurol.366, 580–599.
Oorschot, D.E., Lin, N., Cooper, B.H., Reynolds, J.N.J., Sun, H., and Wickens,J.R. (2013). Synaptic connectivity between rat striatal spiny projection neuronsin vivo: Unexpected multiple somatic innervation in the context of overallsparse proximal connectivity. Basal Ganglia 3, 93–108.
Parent, A., Sato, F., Wu, Y., Gauthier, J., Levesque, M., and Parent, M. (2000).Organization of the basal ganglia: the importance of axonal collateralization.Trends Neurosci. 23 (10, Suppl), S20–S27.
Park, M.R., Lighthall, J.W., and Kitai, S.T. (1980). Recurrent inhibition in the ratneostriatum. Brain Res. 194, 359–369.
Pennartz, C.M.A., Boeijinga, P.H., Kitai, S.T., and Lopes da Silva, F.H. (1991).Contribution of NMDA receptors to postsynaptic potentials and paired-pulsefacilitation in identified neurons of the rat nucleus accumbens in vitro. Exp.Brain Res. 86, 190–198.
Pennartz, C.M.A., Groenewegen, H.J., and Lopes da Silva, F.H. (1994). The nu-cleus accumbens as a complex of functionally distinct neuronal ensembles: anintegration of behavioural, electrophysiological and anatomical data. Prog.Neurobiol. 42, 719–761.
Penney, J.B., Jr., and Young, A.B. (1983). Speculations on the functional anat-omy of basal ganglia disorders. Annu. Rev. Neurosci. 6, 73–94.
Petryszyn, S., Beaulieu, J.M., Parent, A., and Parent, M. (2014). Distributionandmorphological characteristics of striatal interneurons expressing calretininin mice: a comparison with human and nonhuman primates. J. Chem. Neuro-anat. 59-60, 51–61.
Planert, H., Szydlowski, S.N., Hjorth, J.J.J., Grillner, S., and Silberberg, G.(2010). Dynamics of synaptic transmission between fast-spiking interneuronsand striatal projection neurons of the direct and indirect pathways. J. Neurosci.30, 3499–3507.
Plenz, D. (2003).When inhibition goes incognito: feedback interaction betweenspiny projection neurons in striatal function. Trends Neurosci. 26, 436–443.
Plotkin, J.L., Day, M., and Surmeier, D.J. (2011). Synaptically driven state tran-sitions in distal dendrites of striatal spiny neurons. Nat. Neurosci. 14, 881–888.
Ponzi, A., and Wickens, J. (2010). Sequentially switching cell assemblies inrandom inhibitory networks of spiking neurons in the striatum. J. Neurosci.30, 5894–5911.
Ponzi, A., andWickens, J. (2012). Input dependent cell assembly dynamics in amodel of the striatal medium spiny neuron network. Front. Syst. Neurosci. 6, 6.
Ponzi, A., and Wickens, J.R. (2013). Optimal balance of the striatal mediumspiny neuron network. PLoS Comput. Biol. 9, e1002954.

Neuron
Perspective
Preston, R.J., Bishop, G.A., and Kitai, S.T. (1980). Medium spiny neuron pro-jection from the rat striatum: an intracellular horseradish peroxidase study.Brain Res. 183, 253–263.
Qi, J., Zhang, S., Wang, H.-L., Barker, D.J., Miranda-Barrientos, J., and Mo-rales, M. (2016). VTA glutamatergic inputs to nucleus accumbens drive aver-sion by acting on GABAergic interneurons. Nat. Neurosci. 19, 725–733.
Redgrave, P., Prescott, T.J., and Gurney, K. (1999). The basal ganglia: a verte-brate solution to the selection problem? Neuroscience 89, 1009–1023.
Rock, C., Zurita, H., Wilson, C., and Apicella, A.J. (2016). An inhibitory cortico-striatal pathway. eLife 5, 1–17.
Sesack, S.R., and Grace, A.A. (2010). Cortico-Basal Ganglia reward network:microcircuitry. Neuropsychopharmacology 35, 27–47.
Shin, J.H., Adrover, M.F., Wess, J., and Alvarez, V.a. (2015). Muscarinic regu-lation of dopamine and glutamate transmission in the nucleus accumbens.Proc. Natl. Acad. Sci. USA 112, 8124–8249.
Silberberg, G., and Bolam, J.P. (2015). Local and afferent synaptic pathways inthe striatal microcircuitry. Curr. Opin. Neurobiol. 33, 182–187.
Smith, A.D., and Bolam, J.P. (1990). The neural network of the basal ganglia asrevealed by the study of synaptic connections of identified neurones. TrendsNeurosci. 13, 259–265.
Smith, Y., Raju, D., Nanda, B., Pare, J.F., Galvan, A., andWichmann, T. (2009).The thalamostriatal systems: anatomical and functional organization in normaland parkinsonian states. Brain Res. Bull. 78, 60–68.
Smith, J.B., Klug, J.R., Ross, D.L., Howard, C.D., Hollon, N.G., Ko, V.I.,Hoffman, H., Callaway, E.M., Gerfen, C.R., and Jin, X. (2016). Genetic-baseddissection unveils the inputs and outputs of striatal patch and matrix compart-ments. Neuron 91, 1069–1084.
Somogyi, P., Bolam, J.P., and Smith, A.D. (1981). Monosynaptic cortical inputand local axon collaterals of identified striatonigral neurons. A light andelectron microscopic study using the Golgi-peroxidase transport-degenera-tion procedure. J. Comp. Neurol. 195, 567–584.
Stern, E.A., Jaeger, D., and Wilson, C.J. (1998). Membrane potentialsynchrony of simultaneously recorded striatal spiny neurons in vivo. Nature394, 475–478.
Straub, C., Saulnier, J.L., Begue, A., Feng, D.D., Huang, K.W., and Sabatini,B.L. (2016). Principles of Synaptic Organization of GABAergic Interneuronsin the Striatum. Neuron 92, 84–92.
Stuber, G.D., Hnasko, T.S., Britt, J.P., Edwards, R.H., and Bonci, A. (2010).Dopaminergic terminals in the nucleus accumbens but not the dorsal striatumcorelease glutamate. J. Neurosci. 30, 8229–8233.
Sullivan,M.A., Chen, H., andMorikawa, H. (2008). Recurrent inhibitory networkamong striatal cholinergic interneurons. J. Neurosci. 28, 8682–8690.
Surmeier, D.J., Graves, S.M., and Shen, W. (2014). Dopaminergic modulationof striatal networks in health and Parkinson’s disease. Curr. Opin. Neurobiol.29, 109–117.
Swerdlow, N.R., and Koob, G.F. (1987). Dopamine, schizophrenia, mania, anddepression: Toward a unified hypothesis of cortico-striato- pallido-thalamicfunction. Behav. Brain Sci. 10, 197–245.
Szydlowski, S.N., Pollak Dorocic, I., Planert, H., Carlen, M., Meletis, K., andSilberberg, G. (2013). Target selectivity of feedforward inhibition by striatalfast-spiking interneurons. J. Neurosci. 33, 1678–1683.
Tai, L.-H., Lee, A.M., Benavidez, N., Bonci, A., and Wilbrecht, L. (2012). Tran-sient stimulation of distinct subpopulations of striatal neurons mimics changesin action value. Nat. Neurosci. 15, 1281–1289.
Tan, K.R., Yvon, C., Turiault, M., Mirzabekov, J.J., Doehner, J., Labouebe, G.,Deisseroth, K., Tye, K.M., and L€uscher, C. (2012). GABA neurons of the VTAdrive conditioned place aversion. Neuron 73, 1173–1183.
Taverna, S., van Dongen, Y.C., Groenewegen, H.J., and Pennartz, C.M. (2004).Direct physiological evidence for synaptic connectivity between medium-sized spiny neurons in rat nucleus accumbens in situ. J. Neurophysiol. 91,1111–1121.
Taverna, S., Ilijic, E., and Surmeier, D.J. (2008). Recurrent collateral connec-tions of striatal medium spiny neurons are disrupted in models of Parkinson’sdisease. J. Neurosci. 28, 5504–5512.
Tecuapetla, F., Koos, T., Tepper, J.M., Kabbani, N., and Yeckel, M.F. (2009).Differential dopaminergic modulation of neostriatal synaptic connections ofstriatopallidal axon collaterals. J. Neurosci. 29, 8977–8990.
Tecuapetla, F., Patel, J.C., Xenias, H., English, D., Tadros, I., Shah, F., Berlin,J., Deisseroth, K., Rice, M.E., Tepper, J.M., and Koos, T. (2010). Glutamatergicsignaling by mesolimbic dopamine neurons in the nucleus accumbens.J. Neurosci. 30, 7105–7110.
Tecuapetla, F., Matias, S., Dugue, G.P., Mainen, Z.F., and Costa, R.M. (2014).Balanced activity in basal ganglia projection pathways is critical for contraver-sive movements. Nat. Commun. 5, 4315.
Tecuapetla, F., Jin, X., Lima, S.Q., and Costa, R.M. (2016). Complementarycontributions of striatal projection pathways to action initiation and execution.Cell 166, 703–715.
Tepper, J.M., and Koos, T. (2017). GABAergic Interneurons of the Striatum.Handb. Behav. Neurosci. 24, 157–178.
Tepper, J.M., Koos, T., andWilson, C.J. (2004). GABAergic microcircuits in theneostriatum. Trends Neurosci. 27, 662–669.
Tepper, J.M., Wilson, C.J., and Koos, T. (2008). Feedforward and feedback in-hibition in neostriatal GABAergic spiny neurons. Brain Res. Brain Res. Rev. 58,272–281.
Tepper, J.M., Tecuapetla, F., Koos, T., and Ibanez-Sandoval, O. (2010). Het-erogeneity and diversity of striatal GABAergic interneurons. Front. Neuroanat.4, 150.
Threlfell, S., Lalic, T., Platt, N.J., Jennings, K.A., Deisseroth, K., and Cragg, S.J.(2012). Striatal dopamine release is triggered by synchronized activity incholinergic interneurons. Neuron 75, 58–64.
Tolliver, B.K., Sganga, M.W., and Sharp, F.R. (2000). Suppression of c-fos in-duction in the nucleus accumbens prevents acquisition but not expression ofmorphine-conditioned place preference. Eur. J. Neurosci. 12, 3399–3406.
Tomioka, R., Sakimura, K., and Yanagawa, Y. (2015). Corticofugal GABAergicprojection neurons in the mouse frontal cortex. Front. Neuroanat. 9, 133.
Tritsch, N.X., and Sabatini, B.L. (2012). Dopaminergic modulation of synaptictransmission in cortex and striatum. Neuron 76, 33–50.
Tritsch, N.X., Ding, J.B., and Sabatini, B.L. (2012). Dopaminergic neuronsinhibit striatal output through non-canonical release of GABA. Nature 490,262–266.
Tritsch, N.X., Oh, W.J., Gu, C., and Sabatini, B.L. (2014). Midbrain dopamineneurons sustain inhibitory transmission using plasma membrane uptake ofGABA, not synthesis. eLife 3, e01936.
Tunstall, M.J., Oorschot, D.E., Kean, A., and Wickens, J.R. (2002). Inhibitoryinteractions between spiny projection neurons in the rat striatum.J. Neurophysiol. 88, 1263–1269.
Van Bockstaele, E.J., and Pickel, V.M. (1995). GABA-containing neurons in theventral tegmental area project to the nucleus accumbens in rat brain. BrainRes. 682, 215–221.
van Zessen, R., Phillips, J.L., Budygin, E.A., and Stuber, G.D. (2012). Activationof VTA GABA neurons disrupts reward consumption. Neuron 73, 1184–1194.
Vergara, R., Rick, C., Hernandez-Lopez, S., Laville, J.A., Guzman, J.N.,Galarraga, E., Surmeier, D.J., and Bargas, J. (2003). Spontaneous voltage os-cillations in striatal projection neurons in a rat corticostriatal slice. J. Physiol.553, 169–182.
Wall, N.R., De La Parra, M., Callaway, E.M., and Kreitzer, A.C. (2013). Differen-tial innervation of direct- and indirect-pathway striatal projection neurons.Neuron 79, 347–360.
Wang, Z., Kai, L., Day, M., Ronesi, J., Yin, H.H., Ding, J., Tkatch, T., Lovinger,D.M., and Surmeier, D.J. (2006). Dopaminergic control of corticostriatal long-term synaptic depression in medium spiny neurons is mediated by cholinergicinterneurons. Neuron 50, 443–452.
Neuron 96, October 11, 2017 283

Neuron
Perspective
West, E.A., and Carelli, R.M. (2016). Nucleus accumbens core and shelldifferentially encode reward-associated cues after reinforcer devaluation.J. Neurosci. 36, 1128–1139.
West, M.O., Carelli, R.M., Pomerantz, M., Cohen, S.M., Gardner, J.P., Chapin,J.K., andWoodward, D.J. (1990). A region in the dorsolateral striatum of the ratexhibiting single-unit correlations with specific locomotor limb movements.J. Neurophysiol. 64, 1233–1246.
Wilson, C.J. (2007). GABAergic inhibition in the neostriatum. Prog. Brain Res.160, 91–110.
Wilson, C.J., andGroves, P.M. (1980). Fine structure and synaptic connectionsof the common spiny neuron of the rat neostriatum: a study employing intracel-lular inject of horseradish peroxidase. J. Comp. Neurol. 194, 599–615.
Xenias, H.S., Ibanez-Sandoval, O., Koos, T., and Tepper, J.M. (2015). Arestriatal tyrosine hydroxylase interneurons dopaminergic? J. Neurosci. 35,6584–6599.
Yamaguchi, T., Sheen, W., and Morales, M. (2007). Glutamatergic neurons arepresent in the rat ventral tegmental area. Eur. J. Neurosci. 25, 106–118.
Yamaguchi, T., Wang, H.-L., Li, X., Ng, T.H., andMorales, M. (2011). Mesocor-ticolimbic glutamatergic pathway. J. Neurosci. 31, 8476–8490.
284 Neuron 96, October 11, 2017
Yamaguchi, T., Qi, J.,Wang, H.L., Zhang, S., andMorales,M. (2015). Glutama-tergic and dopaminergic neurons in the mouse ventral tegmental area. Eur. J.Neurosci. 41, 760–772.
Yim, M.Y., Aertsen, A., and Kumar, A. (2011). Significance of input correlationsin striatal function. PLoS Comput. Biol. 7, e1002254.
Yoo, J.H., Zell, V., Gutierrez-Reed, N., Wu, J., Ressler, R., Shenasa, M.A.,Johnson, A.B., Fife, K.H., Faget, L., and Hnasko, T.S. (2016). Ventral tegmentalarea glutamate neurons co-release GABA and promote positive reinforce-ment. Nat. Commun. 7, 13697.
Yung, K.K., Smith, A.D., Levey, A.I., and Bolam, J.P. (1996). Synapticconnections between spiny neurons of the direct and indirect pathways inthe neostriatum of the rat: evidence from dopamine receptor and neuropeptideimmunostaining. Eur. J. Neurosci. 8, 861–869.
Zalocusky, K.A., Ramakrishnan, C., Lerner, T.N., Davidson, T.J., Knutson, B.,and Deisseroth, K. (2016). Nucleus accumbens D2R cells signal prioroutcomes and control risky decision-making. Nature 531, 642–646.
Zhou, F.M., Wilson, C.J., and Dani, J.A. (2002). Cholinergic interneuroncharacteristics and nicotinic properties in the striatum. J. Neurobiol. 53,590–605.
Top Related
Copyright © 2022 FDOKUMEN

