







Bahasa
Halaman
Hukum


DOI: 10.1126/science.1179386 , 572 (2009); 326Science
et al.Robert L. Yauch,Hedgehog Pathway Inhibitor in Medulloblastoma
Mutation Confers Resistance to aSmoothened
www.sciencemag.org (this information is current as of December 7, 2009 ):The following resources related to this article are available online at
http://www.sciencemag.org/cgi/content/full/326/5952/572version of this article at:
including high-resolution figures, can be found in the onlineUpdated information and services,
http://www.sciencemag.org/cgi/content/full/1179386/DC1 can be found at: Supporting Online Material
found at: can berelated to this articleA list of selected additional articles on the Science Web sites
http://www.sciencemag.org/cgi/content/full/326/5952/572#related-content
http://www.sciencemag.org/cgi/content/full/326/5952/572#otherarticles, 4 of which can be accessed for free: cites 16 articlesThis article
http://www.sciencemag.org/cgi/collection/medicineMedicine, Diseases
: subject collectionsThis article appears in the following
http://www.sciencemag.org/about/permissions.dtl in whole or in part can be found at: this article
permission to reproduce of this article or about obtaining reprintsInformation about obtaining
registered trademark of AAAS. is aScience2009 by the American Association for the Advancement of Science; all rights reserved. The title
CopyrightAmerican Association for the Advancement of Science, 1200 New York Avenue NW, Washington, DC 20005. (print ISSN 0036-8075; online ISSN 1095-9203) is published weekly, except the last week in December, by theScience
on
Dece
mbe
r 7, 2
009
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from

5. W. C. Feldman et al., J. Geophys. Res. 106, 23231(2001).
6. D. H. Crider, R. R. Vondrak, Adv. Space Res. 30, 1869 (2002).7. L. Starukhina, Y. Shkuratov, Icarus 147, 585 (2000).8. Materials and methods are available as supporting
material on Science Online.9. R. G. Burns, Mineralogical Applications of Crystal Field
Theory (Cambridge Univ. Press, ed. 2, Cambridge, 1993).10. L. P. Keller, D. S. McKay, Geochim. Cosmochim. Acta 61,
2331 (1997).11. S. K. Noble, C. M. Pieters, L. P. Keller, Icarus 192, 629
(2007).12. L. A. Taylor, C. M. Pieters, L. P. Keller, R. V. Morris,
D. S. McKay, J. Geophys. Res. 106, 27985 (2001).13. C. M. Pieters et al., Meteorit. Planet. Sci. 35, 1101 (2000).14. R. N. Clark, Icarus 40, 94 (1979).15. We have used the approach described in (14). Briefly,
our thermal removal algorithm estimates the thermalemission as an excess signal compared with a valuederived from the reflectance trend of unaffected shorterwavelengths. The thermal emission component includesan estimate of emissivity, and our procedure cannotartificially create the appearance of an absorptionfeature. See also (8).
16. Bright feldspathic (highland) soils appear relatively redin this image. The green and blue displayed channels(green–light blue–dark blue colors in Fig. 1D) capturethe relative band strength of diagnostic absorptions ofseveral ferrous minerals. Regions containing abundantand diverse mafic minerals are highlighted by these twoparameters. Both are calculated relative to a continuumand are integrated across M3 channels within the broadmineral absorption bands (see fig. S1). The strength ofthese particular parameters is largely sensitive to therelative abundance of pyroxene, but the integrated1000-nm band-strength parameter (green) is alsosensitive to other mafic minerals present (e.g., olivine).The band strengths of all absorptions are modulated bythe abundance of opaque phases (ilmenite, chromite)and npFe0.
17. R. O. Green et al., 40th Lunar and Planetary ScienceConference, abstract 2307, Houston, TX, 23 to 27 March2009.
18. The reflectance standards used in the calibration ofM3 are 30-by-30–cm panels of Spectralon SN:50119-1-1 and Infragold SN: 50205-1-1 fromLabsphere (North Sutton, NH).
19. R. N. Clark, Science 326, 562 (2009); published online24 September 2009 (10.1126/science.1178105).
20. J. M. Sunshine et al., Science 326, 565 (2009);published online 24 September 2009 (10.1126/science.1179788).
21. A measured depth of the 3-mm absorption band of~7% would be equivalent to an effective thickness of~0.09 mm for such surficial water, which could beaccounted for by a single molecular layer (or a fewlayers) of water/ion on individual grains, given aneffective particle size for the interacting upper lunarregolith of ~1 mm and an overall optical depth of~300 mm. A monolayer of water/ion would not tend tomigrate into the grains, which would therefore remaindry (i.e., the overall abundance of water in the surfacewould remain low but potentially stable). If this surficialwater/ion were photolytically dissociated, the resultingH ions may simply migrate along the surface untilinteracting with another O atom in the grain to againform OH or H2O, resulting in an effective pseudostabilityor temporary steady state. If the regolith grains areanorthositic plagioclase, a monolayer of water on a 1-mmspherical grain would equate to a molar abundance of0.6% and a ~770-ppm mass fraction, given a 1/1 ratioof water molecule/ion to the anorthositic surfacemolecules. However, if the effective particle size ofregolith grains carrying the water is larger than 1 mm,then the surface water proportion will be less.
22. J. R. Johnson et al., J. Geophys. Res. 107, 5008 (2002).23. S. Maurice, D. J. Lawrence, W. C. Feldman, R. C. Elphic,
O. Gasnault, J. Geophys. Res. 109, E07S04 (2004).24. D. J. Lawrence et al., J. Geophys. Res. 111, E08001
(2006).
25. J. W. Boardman et al., paper presented at the 6th EARSeLSIG IS Workshop on Imaging Spectroscopy, Tel Aviv,Israel, 16 to 19 March 2009.
26. Apparent reflectance is the radiance at the sensormultiplied by pi and divided by a solar distance–normalizedsolar spectrum. Currently, M3 often also uses a cos(i)scalar for simple photometric approximations, where i isthe solar incidence angle relative to a smooth sphere.
27. D. M. Wieliczka, S. Weng, M. T. Querry, Appl. Opt. 28,1714 (1989).
28. S. G. Warren, Appl. Opt. 23, 1206 (1984).29. S. J. Seaman, M. D. Dyar, N. Marinkovic, N. Dunbar,
Am. Mineral. 91, 12 (2006).30. M3 is funded as a Mission of Opportunity under NASA’s
Discovery program contract NNM05AB26C to BrownUniversity. Lunar reflectance spectra were acquired usingthe NASA/Keck Reflectance Experiment Laboratory, amultiuser facility supported by NASA grant NNG06GJ31G.A portion of this research was carried out at the JPL,California Institute of Technology, under a contract withNASA. We thank the team of engineers at JPL whodesigned and built M3 (led by T. Glavich and M. White)and the Chandrayaan-1 mission operations team (led byN. S. Hegde, with M3 implementation largely byS. Gomathi), whose mission support has made M3 datapossible. The M3 team is honored to be a guestinstrument on India’s first mission to the Moon.
Supporting Online Materialwww.sciencemag.org/cgi/content/full/1178658/DC1Materials and MethodsSOM TextFigs. S1 to S5References
5 July 2009; accepted 15 September 2009Published online 24 September 2009;10.1126/science.1178658Include this information when citing this paper.
Smoothened Mutation ConfersResistance to a Hedgehog PathwayInhibitor in MedulloblastomaRobert L. Yauch,1 Gerrit J. P. Dijkgraaf,1 Bruno Alicke,1 Thomas Januario,1 Christina P. Ahn,1Thomas Holcomb,1 Kanan Pujara,1 Jeremy Stinson,1 Christopher A. Callahan,1 Tracy Tang,1J. Fernando Bazan,1 Zhengyan Kan,1 Somasekar Seshagiri,1 Christine L. Hann,2Stephen E. Gould,1 Jennifer A. Low,1 Charles M. Rudin,2 Frederic J. de Sauvage1*
The Hedgehog (Hh) signaling pathway is inappropriately activated in certain human cancers,including medulloblastoma, an aggressive brain tumor. GDC-0449, a drug that inhibits Hhsignaling by targeting the serpentine receptor Smoothened (SMO), has produced promisinganti-tumor responses in early clinical studies of cancers driven by mutations in this pathway. Toevaluate the mechanism of resistance in a medulloblastoma patient who had relapsed after aninitial response to GDC-0449, we determined the mutational status of Hh signaling genes in thetumor after disease progression. We identified an amino acid substitution at a conserved asparticacid residue of SMO that had no effect on Hh signaling but disrupted the ability of GDC-0449 tobind SMO and suppress this pathway. A mutation altering the same amino acid also arose in aGDC-0449–resistant mouse model of medulloblastoma. These findings show that acquiredmutations in a serpentine receptor with features of a G protein–coupled receptor can serve as amechanism of drug resistance in human cancer.
The Hh signaling pathway has been im-plicated in the pathogenesis of humanbasal cell carcinoma (BCC) and medullo-
blastoma (1, 2). Constitutive Hh signaling, which
is most often due to underlying loss-of-functionmutations in the gene encoding the inhibitory re-ceptor Patched 1 (PTCH1), occurs in a majorityof BCCs and approximately 30% of sporadic
medulloblastoma cases (2, 3). Mice that areheterozygous forPtch1 (Ptch1+/−) spontaneouslydevelopmedulloblastoma, and treatment with Hhpathway inhibitors suppresses tumor growth andprolongs survival (4, 5). The Hh pathway in-hibitor GDC-0449, a 2-pyridyl amide initiallyidentified in a high-throughput screen, targets theG protein–coupled-like receptor Smoothened(SMO), which becomes activated after loss ofPTCH1 function (3, 6–8). In early-stage clinicalstudies, administration of GDC-0449 to patientswith advanced BCC has produced promisingresults, highlighted by a 55% overall responserate (6, 7). In addition, treatment of a medullo-blastoma patient harboringwidespreadmetastaticdisease with GDC-0449 resulted in a rapid anddramatic tumor regression (8).
Molecular profiling of the medulloblastomapatient’s primary and metastatic tumor takenbefore treatment with GDC-0449 revealed anunderlying somatic mutation in PTCH1 (PTCH1-W844C) as well as up-regulated expression ofHh pathway target genes, supporting the hypoth-esis that the tumor was driven by dysregulated
1Genentech, South San Francisco, CA 94080, USA. 2SidneyKimmel Comprehensive Cancer Center, Johns Hopkins Uni-versity, Baltimore, MD 21231, USA.
*To whom correspondence should be addressed. E-mail:[email protected]
23 OCTOBER 2009 VOL 326 SCIENCE www.sciencemag.org572
REPORTS
on
Dece
mbe
r 7, 2
009
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from
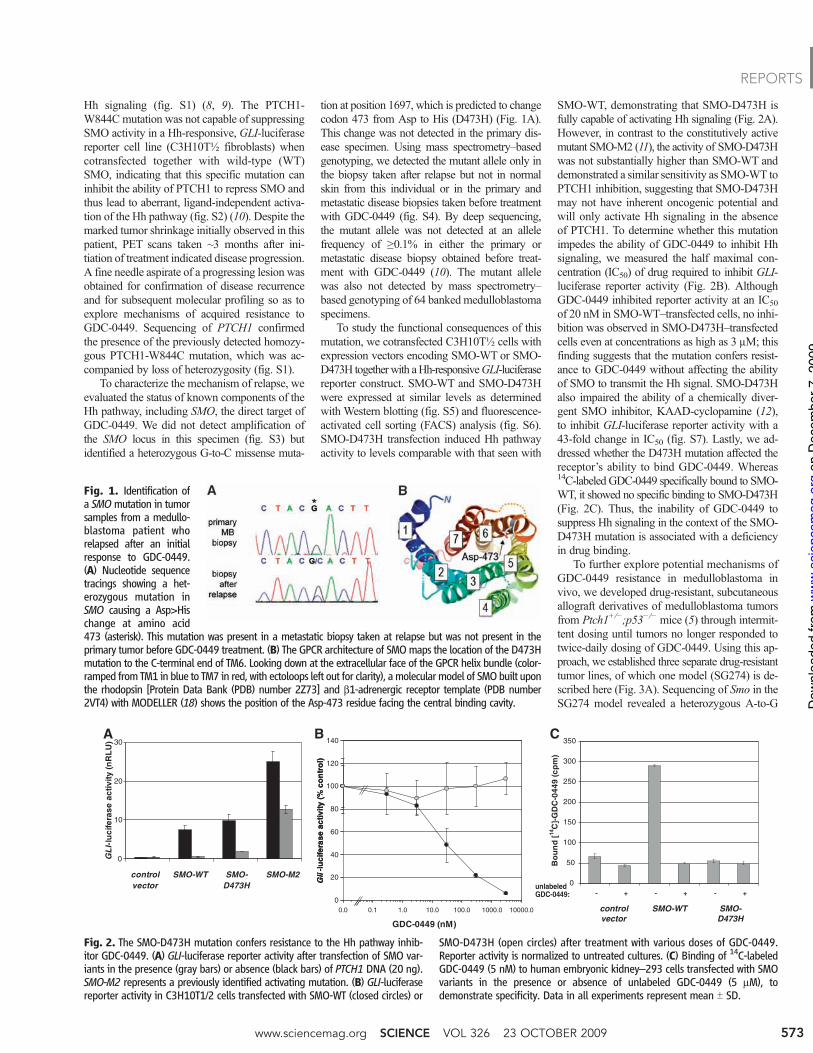
Hh signaling (fig. S1) (8, 9). The PTCH1-W844Cmutation was not capable of suppressingSMO activity in a Hh-responsive, GLI-luciferasereporter cell line (C3H10T½ fibroblasts) whencotransfected together with wild-type (WT)SMO, indicating that this specific mutation caninhibit the ability of PTCH1 to repress SMO andthus lead to aberrant, ligand-independent activa-tion of the Hh pathway (fig. S2) (10). Despite themarked tumor shrinkage initially observed in thispatient, PET scans taken ~3 months after ini-tiation of treatment indicated disease progression.A fine needle aspirate of a progressing lesion wasobtained for confirmation of disease recurrenceand for subsequent molecular profiling so as toexplore mechanisms of acquired resistance toGDC-0449. Sequencing of PTCH1 confirmedthe presence of the previously detected homozy-gous PTCH1-W844C mutation, which was ac-companied by loss of heterozygosity (fig. S1).
To characterize the mechanism of relapse, weevaluated the status of known components of theHh pathway, including SMO, the direct target ofGDC-0449. We did not detect amplification ofthe SMO locus in this specimen (fig. S3) butidentified a heterozygous G-to-C missense muta-
tion at position 1697, which is predicted to changecodon 473 from Asp to His (D473H) (Fig. 1A).This change was not detected in the primary dis-ease specimen. Using mass spectrometry–basedgenotyping, we detected the mutant allele only inthe biopsy taken after relapse but not in normalskin from this individual or in the primary andmetastatic disease biopsies taken before treatmentwith GDC-0449 (fig. S4). By deep sequencing,the mutant allele was not detected at an allelefrequency of !0.1% in either the primary ormetastatic disease biopsy obtained before treat-ment with GDC-0449 (10). The mutant allelewas also not detected by mass spectrometry–based genotyping of 64 banked medulloblastomaspecimens.
To study the functional consequences of thismutation, we cotransfected C3H10T½ cells withexpression vectors encoding SMO-WT or SMO-D473H togetherwith aHh-responsiveGLI-luciferasereporter construct. SMO-WT and SMO-D473Hwere expressed at similar levels as determinedwith Western blotting (fig. S5) and fluorescence-activated cell sorting (FACS) analysis (fig. S6).SMO-D473H transfection induced Hh pathwayactivity to levels comparable with that seen with
SMO-WT, demonstrating that SMO-D473H isfully capable of activating Hh signaling (Fig. 2A).However, in contrast to the constitutively activemutant SMO-M2 (11), the activity of SMO-D473Hwas not substantially higher than SMO-WT anddemonstrated a similar sensitivity as SMO-WT toPTCH1 inhibition, suggesting that SMO-D473Hmay not have inherent oncogenic potential andwill only activate Hh signaling in the absenceof PTCH1. To determine whether this mutationimpedes the ability of GDC-0449 to inhibit Hhsignaling, we measured the half maximal con-centration (IC50) of drug required to inhibit GLI-luciferase reporter activity (Fig. 2B). AlthoughGDC-0449 inhibited reporter activity at an IC50
of 20 nM in SMO-WT–transfected cells, no inhi-bition was observed in SMO-D473H–transfectedcells even at concentrations as high as 3 mM; thisfinding suggests that the mutation confers resist-ance to GDC-0449 without affecting the abilityof SMO to transmit the Hh signal. SMO-D473Halso impaired the ability of a chemically diver-gent SMO inhibitor, KAAD-cyclopamine (12),to inhibit GLI-luciferase reporter activity with a43-fold change in IC50 (fig. S7). Lastly, we ad-dressed whether the D473H mutation affected thereceptor’s ability to bind GDC-0449. Whereas14C-labeled GDC-0449 specifically bound to SMO-WT, it showed no specific binding to SMO-D473H(Fig. 2C). Thus, the inability of GDC-0449 tosuppress Hh signaling in the context of the SMO-D473H mutation is associated with a deficiencyin drug binding.
To further explore potential mechanisms ofGDC-0449 resistance in medulloblastoma invivo, we developed drug-resistant, subcutaneousallograft derivatives of medulloblastoma tumorsfrom Ptch1+/−;p53−/− mice (5) through intermit-tent dosing until tumors no longer responded totwice-daily dosing of GDC-0449. Using this ap-proach, we established three separate drug-resistanttumor lines, of which one model (SG274) is de-scribed here (Fig. 3A). Sequencing of Smo in theSG274 model revealed a heterozygous A-to-G
Fig. 1. Identification ofa SMOmutation in tumorsamples from a medullo-blastoma patient whorelapsed after an initialresponse to GDC-0449.(A) Nucleotide sequencetracings showing a het-erozygous mutation inSMO causing a Asp>Hischange at amino acid473 (asterisk). This mutation was present in a metastatic biopsy taken at relapse but was not present in theprimary tumor before GDC-0449 treatment. (B) The GPCR architecture of SMOmaps the location of the D473Hmutation to the C-terminal end of TM6. Looking down at the extracellular face of the GPCR helix bundle (color-ramped from TM1 in blue to TM7 in red, with ectoloops left out for clarity), amolecularmodel of SMObuilt uponthe rhodopsin [Protein Data Bank (PDB) number 2Z73] and b1-adrenergic receptor template (PDB number2VT4) with MODELLER (18) shows the position of the Asp-473 residue facing the central binding cavity.
0
10
20
30
controlvector
SMO-WT SMO-D473H
SMO-M2
GL
I-lu
cife
rase
act
ivity
(nR
LU
)
0
20
40
60
80
100
120
140
0.0 0.1 1.0 10.0 100.0 1000.0 10000.0
GDC-0449 (nM)
Gli
-luci
fera
se a
ctiv
ity (%
co
ntr
ol)
Gli
-luci
fera
se a
ctiv
ity (%
co
ntr
ol)
0
50
100
150
200
250
300
350
- + - + - +
Bo
un
d [14
C]-
GD
C-0
449
(cpm
)
unlabeled GDC-0449:
controlvector
SMO-WT SMO-D473H
A B C
Fig. 2. The SMO-D473H mutation confers resistance to the Hh pathway inhib-itor GDC-0449. (A) GLI-luciferase reporter activity after transfection of SMO var-iants in the presence (gray bars) or absence (black bars) of PTCH1 DNA (20 ng).SMO-M2 represents a previously identified activating mutation. (B) GLI-luciferasereporter activity in C3H10T1/2 cells transfected with SMO-WT (closed circles) or
SMO-D473H (open circles) after treatment with various doses of GDC-0449.Reporter activity is normalized to untreated cultures. (C) Binding of 14C-labeledGDC-0449 (5 nM) to human embryonic kidney–293 cells transfected with SMOvariants in the presence or absence of unlabeled GDC-0449 (5 mM), todemonstrate specificity. Data in all experiments represent mean T SD.
www.sciencemag.org SCIENCE VOL 326 23 OCTOBER 2009 573
REPORTS
on
Dece
mbe
r 7, 2
009
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from
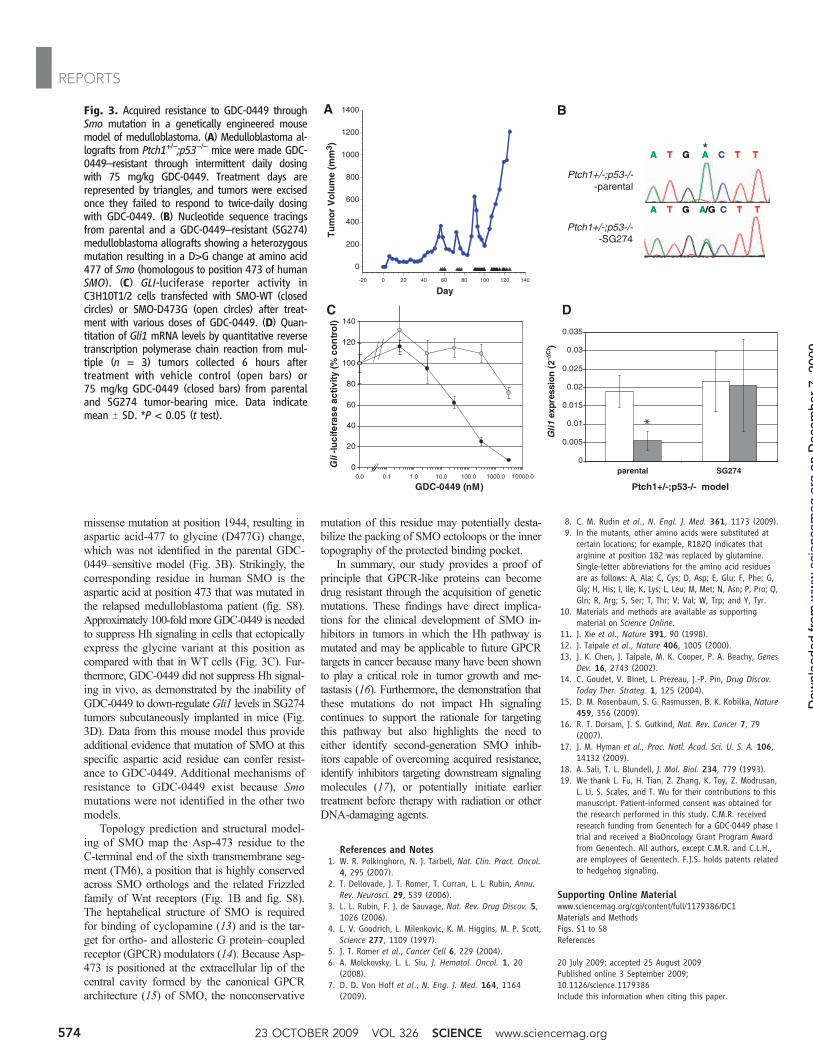
missense mutation at position 1944, resulting inaspartic acid-477 to glycine (D477G) change,which was not identified in the parental GDC-0449–sensitive model (Fig. 3B). Strikingly, thecorresponding residue in human SMO is theaspartic acid at position 473 that was mutated inthe relapsed medulloblastoma patient (fig. S8).Approximately 100-fold more GDC-0449 is neededto suppress Hh signaling in cells that ectopicallyexpress the glycine variant at this position ascompared with that in WT cells (Fig. 3C). Fur-thermore, GDC-0449 did not suppress Hh signal-ing in vivo, as demonstrated by the inability ofGDC-0449 to down-regulate Gli1 levels in SG274tumors subcutaneously implanted in mice (Fig.3D). Data from this mouse model thus provideadditional evidence that mutation of SMO at thisspecific aspartic acid residue can confer resist-ance to GDC-0449. Additional mechanisms ofresistance to GDC-0449 exist because Smomutations were not identified in the other twomodels.
Topology prediction and structural model-ing of SMO map the Asp-473 residue to theC-terminal end of the sixth transmembrane seg-ment (TM6), a position that is highly conservedacross SMO orthologs and the related Frizzledfamily of Wnt receptors (Fig. 1B and fig. S8).The heptahelical structure of SMO is requiredfor binding of cyclopamine (13) and is the tar-get for ortho- and allosteric G protein–coupledreceptor (GPCR) modulators (14). Because Asp-473 is positioned at the extracellular lip of thecentral cavity formed by the canonical GPCRarchitecture (15) of SMO, the nonconservative
mutation of this residue may potentially desta-bilize the packing of SMO ectoloops or the innertopography of the protected binding pocket.
In summary, our study provides a proof ofprinciple that GPCR-like proteins can becomedrug resistant through the acquisition of geneticmutations. These findings have direct implica-tions for the clinical development of SMO in-hibitors in tumors in which the Hh pathway ismutated and may be applicable to future GPCRtargets in cancer because many have been shownto play a critical role in tumor growth and me-tastasis (16). Furthermore, the demonstration thatthese mutations do not impact Hh signalingcontinues to support the rationale for targetingthis pathway but also highlights the need toeither identify second-generation SMO inhib-itors capable of overcoming acquired resistance,identify inhibitors targeting downstream signalingmolecules (17), or potentially initiate earliertreatment before therapy with radiation or otherDNA-damaging agents.
References and Notes1. W. R. Polkinghorn, N. J. Tarbell, Nat. Clin. Pract. Oncol.
4, 295 (2007).2. T. Dellovade, J. T. Romer, T. Curran, L. L. Rubin, Annu.
Rev. Neurosci. 29, 539 (2006).3. L. L. Rubin, F. J. de Sauvage, Nat. Rev. Drug Discov. 5,
1026 (2006).4. L. V. Goodrich, L. Milenkovic, K. M. Higgins, M. P. Scott,
Science 277, 1109 (1997).5. J. T. Romer et al., Cancer Cell 6, 229 (2004).6. A. Molckovsky, L. L. Siu, J. Hematol. Oncol. 1, 20
(2008).7. D. D. Von Hoff et al., N. Eng. J. Med. 164, 1164
(2009).
8. C. M. Rudin et al., N. Engl. J. Med. 361, 1173 (2009).9. In the mutants, other amino acids were substituted at
certain locations; for example, R182Q indicates thatarginine at position 182 was replaced by glutamine.Single-letter abbreviations for the amino acid residuesare as follows: A, Ala; C, Cys; D, Asp; E, Glu; F, Phe; G,Gly; H, His; I, Ile; K, Lys; L, Leu; M, Met; N, Asn; P, Pro; Q,Gln; R, Arg; S, Ser; T, Thr; V, Val; W, Trp; and Y, Tyr.
10. Materials and methods are available as supportingmaterial on Science Online.
11. J. Xie et al., Nature 391, 90 (1998).12. J. Taipale et al., Nature 406, 1005 (2000).13. J. K. Chen, J. Taipale, M. K. Cooper, P. A. Beachy, Genes
Dev. 16, 2743 (2002).14. C. Goudet, V. Binet, L. Prezeau, J.-P. Pin, Drug Discov.
Today Ther. Strateg. 1, 125 (2004).15. D. M. Rosenbaum, S. G. Rasmussen, B. K. Kobilka, Nature
459, 356 (2009).16. R. T. Dorsam, J. S. Gutkind, Nat. Rev. Cancer 7, 79
(2007).17. J. M. Hyman et al., Proc. Natl. Acad. Sci. U. S. A. 106,
14132 (2009).18. A. Sali, T. L. Blundell, J. Mol. Biol. 234, 779 (1993).19. We thank L. Fu, H. Tian, Z. Zhang, K. Toy, Z. Modrusan,
L. Li, S. Scales, and T. Wu for their contributions to thismanuscript. Patient-informed consent was obtained forthe research performed in this study. C.M.R. receivedresearch funding from Genentech for a GDC-0449 phase Itrial and received a BioOncology Grant Program Awardfrom Genentech. All authors, except C.M.R. and C.L.H.,are employees of Genentech. F.J.S. holds patents relatedto hedgehog signaling.
Supporting Online Materialwww.sciencemag.org/cgi/content/full/1179386/DC1Materials and MethodsFigs. S1 to S8References
20 July 2009; accepted 25 August 2009Published online 3 September 2009;10.1126/science.1179386Include this information when citing this paper.
Fig. 3. Acquired resistance to GDC-0449 throughSmo mutation in a genetically engineered mousemodel of medulloblastoma. (A) Medulloblastoma al-lografts from Ptch1+/−;p53−/− mice were made GDC-0449–resistant through intermittent daily dosingwith 75 mg/kg GDC-0449. Treatment days arerepresented by triangles, and tumors were excisedonce they failed to respond to twice-daily dosingwith GDC-0449. (B) Nucleotide sequence tracingsfrom parental and a GDC-0449–resistant (SG274)medulloblastoma allografts showing a heterozygousmutation resulting in a D>G change at amino acid477 of Smo (homologous to position 473 of humanSMO). (C) GLI-luciferase reporter activity inC3H10T1/2 cells transfected with SMO-WT (closedcircles) or SMO-D473G (open circles) after treat-ment with various doses of GDC-0449. (D) Quan-titation of Gli1 mRNA levels by quantitative reversetranscription polymerase chain reaction from mul-tiple (n = 3) tumors collected 6 hours aftertreatment with vehicle control (open bars) or75 mg/kg GDC-0449 (closed bars) from parentaland SG274 tumor-bearing mice. Data indicatemean T SD. *P < 0.05 (t test).
0
20
40
60
80
100
120
140
0.0 0.1 1.0 10.0 100.0 1000.0 10000.0
GDC-0449 (nM)
Gli
-luci
fera
se a
ctiv
ity (%
con
tro
l)
0
200
400
600
800
1000
1200
1400
-20 0 20 40 60 80 100 120 140
Tum
or V
olum
e (m
m3 )
Day
A T G A C T T
A T G A/G C T T
A T G A C T T
A T G A/G C T T
Ptch1+/-;p53-/--parental
Ptch1+/-;p53-/--SG274
*
A B
DC
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
Gli1
exp
ress
ion
(2-∆
Ct )
Ptch1+/-;p53-/- model
*
parental SG274
23 OCTOBER 2009 VOL 326 SCIENCE www.sciencemag.org574
REPORTS
on
Dece
mbe
r 7, 2
009
www.
scie
ncem
ag.o
rgDo
wnlo
aded
from

www.sciencemag.org/cgi/content/full/1179386/DC1
Supporting Online Material for
Smoothened Mutation Confers Resistance to a Hedgehog Pathway Inhibitor in Medulloblastoma
Robert L. Yauch, Gerrit J. P. Dijkgraaf, Bruno Alicke, Thomas Januario, Christina P. Ahn, Thomas Holcomb, Kanan Pujara, Jeremy Stinson,
Christopher A. Callahan, Tracy Tang, J. Fernando Bazan, Zhengyan Kan, Somasekar Seshagiri, Christine L. Hann, Stephen E. Gould, Jennifer A. Low,
Charles M. Rudin, Frederic J. de Sauvage*
*To whom correspondence should be addressed. E-mail: [email protected]
Published 3 September 2009 on Science Express DOI: 10.1126/science.1179386
This PDF file includes:
Materials and Methods
Figs. S1 to S8
References

Supporting Online Material MATERIALS & METHODS !Reagents and constructs.! "##$%&'()*+,-./0! 1,2! +34(5,206! 74*-! 8*4*/9*!:020,4(5! &50-.(,)2! ;/(<! =(,9<! >! "?@?AAAB<! C$&%ADDE! 1,2! -,60! ,9!C0/0/90(5=8B<! #))! F5! ./5.G.9*42!1040! 29*406! ,2! HA!-I! 29*(J2! ./! ?AAK!$ILM!=L.N-,B!,9!%OA!!&<!!F3-,/!SMOP!53-,/!PTCH1!=94,/2(4.+9!Q,4.,/9!?GP!C0/R,/J!SITAAAOUD<HB! ,/6! eGFP! 1040! ()*/06! ./9*! +:"V! =R$! R.*2(.0/(02B! ,/6!0W+402206! 74*-! 950! &IX! +4*-*904<! Y*./9! -39,9.*/2! 1040! ./94*63(06! 1.95! 950!Z3.J&5,/N0! ;;! L.90%$.40(906!I39,N0/02.2! ".9! 74*-!L94,9,N0/0! =(,9<! >! OAAVODBP!,/6!,![\#C!9,N!1,2! ./94*63(06!,9! 950!(,4G*W'! 904-./.!*7!1.)6%9'+0!,/6!-39,/9!53-,/!SMO! G'! Y&:! 32./N! Y),9./3-!Taq! $S#!Y*)'-04,20!F.N5! [.60).9'! 74*-!;/Q.94*N0/!=(,9<!>!??HAD%A??B<!I34*/0!et al<!=?EEEB!+40Q.*32)'!602(4.G06!950!F5!)3(.704,20!40+*4904!C).%RL!,/6! 950!Renilla! 94,/270(9.*/!(*/94*)!+),2-.6!=+:\%8"B!,40!74*-!Y4*-0N,!=(,9<!>!]OOD?B<!#))!(*/2943(92!1040!(*/7.4-06!G'!20^30/(./N<!!!!Hh pathway gene status.! ! ]W*/2! (*Q04./N! 950! *+0/! 40,6./N! 74,-0! *7!-*320!,/6! 53-,/! SMO/Smo, ./! ,66.9.*/! 9*! 0W*/?V! *7! PTCH1,! 1040! Y&:%,-+).7.06!74*-!N0/*-.(!$S#!32./N!,!+,.4!*7!/02906!+4.-042<! !850! ./904/,)!+,.4!*7!+4.-042!3206! ./! 950! ,-+).7.(,9.*/! (*/9,./06! -?H! 7*41,46! *4! -?H! 40Q0420! +4.-04!20^30/(02<!!#7904!Y&:P!7400!/3()0*9.602!,/6!0W(022!+4.-04!1040!40-*Q06!32./N!]W*L#Y%;8! J.9! =_LRB`! Y&:! +4*63(92! 1040! 20^30/(06! ./! G*95! 6.40(9.*/2! 32./N!-?H! 20^30/(./N! +4.-042<! Y&:! +4*63(92! 1040! ('()0%20^30/(06! 32./N!R.N804-./,9*4!".9!=#++).06!R.*2'290-2B<!!#))!20^30/(./N!+4*63(92!1040!402*)Q06!*/! ,! H@HAW)! 20^30/(./N!-,(5./0! =#++).06! R.*2'290-2B<! ! L0^30/(0! 94,(0! 7.)02!1040! ,/,)'a06! 32./N! L0^30/(504! =C0/0&*602B! ,/6b*4! I39,9.*/! L34Q0'*4!=L*79C0/09.(2! \\&B<! ! 850!LIM!$D@HF!-39,9.*/!1,2! ,66.9.*/,))'! (*/7.4-06! G'!+4.-04! 0W90/2.*/! ,/6! I#\$;%8M[! -,22! 2+0(94*2(*+'! *7! 950! ,-+).7.06! $S#!=I,22#::#cP!L0^30/*-P!L,/!$.0N*P!&,).7*4/.,B<!!!!!850!7*))*1./N!+4.-042!1040!39.).a06`!!]W90/6!Y4.-04!=_]Y<$D@HFBd!!8&#C&8C&&#&88&8#&!!=VAe?<H!$,BP!!#/,)'90!Cd!!8&#C&8C&&#&88&8#&C!=VHUe<V!$,BP!!#/,)'90!&d!!8&#C&8C&&#&88&8#&&!=VHOe<V!$,B<!!!850!LIM!$D@HF!-39,9.*/!1,2!+40Q.*32)'!40+*4906!,2!,!4,40!LSY!=407!LSY!;$d!42?@@?AeE?BP!5*10Q04!10!1040!3/,G)0!9*!(*/7.4-!95.2!N0/*9'+0!./!$S#!*G9,./06!74*-!950!,770(906!./6.Q.63,)!*4!950.4!+06.N400<!!! Deep sequencing. !850!40N.*/!2344*3/6./N!950!SMO!0W*/!e!-39,9.*/,)!2.90!1,2!Y&:!,-+).7.06!74*-!950!+4.-,4'!6.20,20!G.*+2'!$S#P!-09,29,9.(!6.20,20!G.*+2'!$S#!,/6!,!(*/94*)!/*4-,)!$S#!=Y4*-0N,P!f;BP!+**)06!,/6!,/,)'a06!*/!,/!;))3-./,!C0/*-0!#/,)'a04<!!#!20^30/(0!G,4(*60!g##&C&#h!7*4!950!+4.-,4'!6.20,20!$S#P!g##&8C&h!7*4!-09,29,9.(!6.20,20!$S#!,/6!g##C&&8h!7*4!/*4-,)!$S#!1,2!,6606!,2!+,49!*7!950!Y&:!+4*(022!,/6!95.2!20^30/(0!1,2!3206!9*!2*49!

950!20^30/(02!./9*!950!95400!(,90N*4.02!+*29!20^30/(./N<!!#!9*9,)!*7!V@!-.)).*/!HU%G+!40,62!(*Q04./N!950!9,4N09!40N.*/!1,2!,/,)'a06!7*4!950!+4020/(0!*7!950!-39,906!,))0)0!=Ci&B<!!f0!23((02273))'!,).N/06!EH<VK!*7!950!25*49!40,62!9*!950!9,4N09P!40^3.4./N!,!+0470(9!-,9(5!9*!*/0!*7!950!95400!20^30/(0!G,4(*602P!,/6!,))*1./N!7*4!91*!-.2-,9(502!,7904!-,2J./N!950!+*2.9.*/!*7!950!-39,/9!,))0)0<!!#).N/-0/92!1040!+047*4-06!32./N!I#Z!1.95!950!607,3)9!(4.904.,!7*4!7.)904./N!+**4!^3,).9'!,).N/-0/92!=2?B<!!#/,)'2.2!1,2!40294.(906!9*!G,202!5,Q./N!,!^3,).9'!2(*40!!!HAP!,/6!,9!0,(5!+*2.9.*/P!950!+4*+*49.*/!*7!40,62!(*/9,././N!Q,4.,/9!,))0)02!1,2!(*-+3906<!!850!-39,/9!Ci&!,))0)0!*7!./904029!*((34406!./!A<AOEK!*7!950!40,62!./!950!+4.-,4'!2,-+)02!=*39!*7!??<O!-.)).*/!40,62!1.95!^3,).9'!!!!HABP!A<AOK!./!950!-09,29,9.(!2,-+)0!=*39!*7!?H<@!-.)).*/!40,62BP!,/6!A<AOK!./!950!/*4-,)!2,-+)0!=*39!*7!?A<O!-.)).*/!40,62B<!!_2./N!950!G,4(*60!40N.*/!,/6!950!20^30/(0!*392.60!*7!950!+*2.9.*/!*7!./904029!10!029.-,906!950!G,(JN4*3/6!20^30/(./N!044*4!4,90!9*!G0!A<AOKP!15.(5!40+4020/92!950!954025*)6!*7!6090(9.*/!32./N!95.2!90(5/*)*N'<!!#!G./*-.,)!9029!=+jAB!0W()3602!950!+4020/(0!*7!950!-39,/9!,))0)0!,9!,!A<?K!*4!5.N504!)0Q0)!./!,/'!*7!950!2,-+)02<!!Luciferase reporter assays.! &HF?A8?bO! (0))2! =#8&&P! (,9<! >! &&\%OOUB! 1040!200606!./9*!2.W%10))!+),902!,9!?<V!W!?AV!(0))2b10))! ./!$I]I!F.N5!C)3(*20!1.95!D!-I! N)39,-./0P! ?A! -I! F0+02! +F! @<O! ,/6! ?AK! [RL! 950! ,7904/**/! G07*40!94,/270(9.*/<!&0))2!1040!94,/270(906!950!/0W9!-*4/./N!1.95!DAA!/N!*7!0W+4022.*/!(*/2943(9P! DAA! /N! *7! C).%RL! ,/6! OAA! /N! *7! +:\%8"! +04! 10))! 32./N! C0/0k3.(0!84,/270(9.*/! :0,N0/9! =S*Q,N0/P! (,9<! >! @AEU@B<! [*4! 950! Y8&F?! ./5.G.9.*/!0W+04.-0/92P!(0))2!1040!94,/270(906!1.95!,/!,66.9.*/,)!OAA!/N!*7!$S#!(*/9,././N!Q,4'./N! 4,9.*2! *7!PTCH1! 9*! eGFP! 0W+4022.*/! (*/2943(92<! L.W! 5*342! ),904P! (0))2!1040! (*))0(906! G'! 94'+2./! 940,9-0/9! ,/6! 0,(5! 10))! 1,2! 406.294.G3906! *Q04! 7*34!10))2!*7!,!?O%10))!+),90<!850![RL!(*/90/9!*7!950!(3)9340!-06.3-!1,2!4063(06!9*!A<VK! 950! 7*))*1./N! -*4/./N! 9*! ./63(0! 7*4-,9.*/! *7! +4.-,4'! (.).,P! ,/6! 2-,))!-*)0(3)0! F5! ./5.G.9*42! 1040! ,6606! ,9! ./6.(,906! (*/(0/94,9.*/2<! \3(.704,20!,(9.Q.9'! 1,2! 60904-./06! De! 5*342! ),904! 1.95! 950! $3,)%C)*! \3(.704,20! #22,'!L'290-! =Y4*-0N,P! (,9<! >! ]OEDAB<! X,)302! 1040! 6.Q.606! G'! Renilla! )3(.704,20!,(9.Q.9.02! 9*!/*4-,).a0! 7*4! 94,/270(9.*/!077.(.0/('!,/6!,40!25*1/!,2! 950!-0,/!*7!95400!20+,4,90!0W+04.-0/92!"!?!29,/6,46!60Q.,9.*/<!!!Western Blot and FACS for expression of Smo and Flag in 10T1/2 cells ?A8?bO!(0))2!1040! 94,/2.0/9)'! 94,/270(906!32./N!C0/0l3.(0!84,/270(9.*/!:0,N0/9!=S*Q,N0/!@AEU@B!1.95!0.9504!f8!L-*P!-39!L-*P!*4!0-+9'!+:"V!Q0(9*4! 7*4!De!5*342<!!f02904/!G)*9!1,2!(,44.06!*39!32./N!29,/6,46!+3G).2506!-095*62!39.).a./N!,!D%?OK!84.2%C)'(./0!N0)! !,/6!,! !/.94*(0))3)*20!-0-G4,/0<! !Y4*90./!1,2!6090(906!32./N! 950! ,/9.%[),N! ,/9.G*6'!IO! =L.N-,! [H?UVB!1.95! ,/! #)0W,! Dee! ,/9.%-*320!20(*/6,4'! =I*)0(3),4! Y4*G02! #??AA?B<! ! R)*92! 1040! .-,N06! G'! 7)3*402(0/(0!32./N!,/!#-0425,-!R.*2(.0/(02!8'+5**/!84.*<!![*4![#&L!,/,)'2.2P!94,/270(906!(0))2!1040!6.2)*6N06! ./!?-I!]$8#bYRL!,/6!23G20^30/9)'! ./(3G,906!1.95!L-*!,/9.G*6'P! O$??! =?3Nb-)BP! 7*))*106! G'! OA! -./! ./(3G,9.*/2! 1.95! ! G.*9./%LY%(*/l3N,906! #77./.Y340! N*,9! ,/9.%-*320! ;NC! =?d?AAP! k,(J2*/! ;--3/*4020,4(5!\,G2!??V%AUV%A@?B!,/6!:%Y5'(*04'954./%(*/l3N,906!L940+9,Q.6./!=?dVAP!k,(J2*/!

;--3/*4020,4(5!\,G2!A?U%??A%AeDB<!!#7904!40232+0/2.*/!./!Y;!=VAA/Nb-)BP!(0))2!1040!,/,)'a06!32./N!,!R$!R.*2(.0/(02!F8L![,(2&,).G34<!!!14C-GDC-0449 Binding and Competition OEH! (0))2! 1040! 94,/2.0/9)'! 94,/270(906! 32./N! C0/0l3.(0! 1.95! 0.9504!f8! L-*! *4!-39!L-*! 7*4!De!5*342<! !#7904!5,4Q029./N! ./!?-I!]$8#! ./!YRLP!?A!-.)).*/!(0))2!1040! 7.W06! 1.95! DKY[#! 7*4! ?A!-./3902! ,9! 4**-! 90-+04,9340P! 1,2506! Hm! 1.95!?-I!]$8#!./!YRL!950/!./(3G,906!7*4!?!5*34!,9!4**-!90-+04,9340!1.95!?D&%C$&%ADDE!=V/IBP!1.95!*4!1.95*39!VA#I!(*)6!C$&%ADDE<!!&0))2!1040!1,2506!Hm!1.95!YRL! 950/! 94,/2704406! 9*!,! 2(./9.)),9.*/! Q.,)<! ! ?V-)!Y.(*[)3*4! DA!1,2!,6606!,/6!(*3/92!+04!-./390!1040!,2202206!32./N!,!Y04J./])-04!).^3.6!2(./9.)),9.*/!,/,)'a04!84.%&,4G!OEAA8:<!!#))!2,-+)02!1040!6*/0!./!94.+).(,90<!!!Generation of mouse medulloblastoma models of GDC-0449 resistance L9,49./N!,9!D!100J2!*7!,N0P!Y9(5?%bn`+VH%b%!-.(0!*/!,!&V@R\bU!G,(JN4*3/6!1040!-*/.9*406!100J)'! 7*4! 950!+4020/(0!*7!-063))*G),29*-,2!G'!I:;<!#))!-.(0!1040!-*/.9*406! 6,.)'! 7*4! ,/'! 2.N/2! *7! ,G/*4-,)! G05,Q.*4! ./6.(,9.Q0! *7! &SL!./Q*)Q0-0/9<!I.(0!1.95!10))%607./06!93-*42!6090(906!G'!I:;!1040!2,(4.7.(06!,/6!93-*42!6.220(906!74*-!/*4-,)!(040G0))3-P!-0(5,/.(,))'!6.22*(.,906P!,/6!V!W?AU!(0))2! ./l0(906! ./9*! 950! ),904,)! 95*4,(.(! 40N.*/!*7!&$%?!/360!-.(0! =&:\B<!83-*42!1040!,))*106! 9*!+4*N4022! 9*!,++4*W.-,90)'!DAA!--H! ./!2.a0!,9!15.(5! 9.-0!-.(0!1040! 940,906!1.95! @V!-NbJN!C$&%ADDE! =7400! G,20! 0^3.Q,)0/92B! */(0! 6,.)'! 3/9.)!93-*42! 60(40,206! ./! 2.a0! 9*! ,++4*W.-,90)'! ?AA!--H<! I.(0! 1040! 950/! N.Q0/! ,!6*2./N!5*).6,'!3/9.)! 40%N4*195!*((34406<!L0^30/9.,)! 4*3/62!*7! 940,9-0/9!,/6! 40%N4*195!1040!,++).06!3/9.)!950!93-*42!1040!/*!)*/N04!402+*/2.Q0!9*!6,.)'!6*2./N!,9!15.(5! 9.-0! 6*2./N! 740^30/('! 1,2! ./(40,206! 9*! 91.(0! 6,.)'! ,9! @V! -NbJN<!:02.29,/(0!60Q0)*+06!*Q04!,!+04.*6!*7!@%?A!100J2! ./! 95020!-*60)2!23NN029./N!95,9!23G*+9.-,)!6*2./N!!*7!C$&%ADDE!(,/!4,+.6)'!)0,6!9*!402.29,/(0<!!83-*42!95,9!7,.)06! 9*!402+*/6!9*! 95.2!6*20!,/6!740^30/('!1040! 950/!5,4Q02906!7*4!-*)0(3),4!,/,)'2.2! ,/6! +,22,N06! =7.429! +,22,N0B! 23G(39,/0*32)'! ./9*! ,66.9.*/,)! -.(0! 7*4!0W+,/2.*/<! ;/! *4604! 9*! ,/,)'a0! +,951,'!-*63),9.*/! ./! 402+*/20! 9*!C$&%ADDEP!93-*4%G0,4./N!-.(0! ,9! 20(*/6%+,22,N0!1040! 6*206! */(0!1.95! @V!-NbJN!C$&%ADDE!,/6!5,4Q02906!U!5*342!+*29!6*20! 7*4!,/,)'2.2<!#))!-.(0!1040!5*3206!,/6!-,./9,./06!,((*46./N!9*!950!,/.-,)!320!N3.60)./02!*7!C0/0/90(5P!;/(P!(*/7*4-./N!9*!&,).7*4/.,!L9,90!)0N,)!,/6!095.(,)!+4,(9.(02<!!:07040/(02!!2?<! F<!\.P!k<!:3,/P!:<!$34G./P!Genome Res!18P!?eV?!=OAAeB<
GT
GC
C T
G
GT
GG
C T
G
GT
GC
C T
G
GT
GC
C T
G
GT
GG
C T
G
GT
GC
C T
G
primary
MB
biopsy
biopsyafter
relapse
control
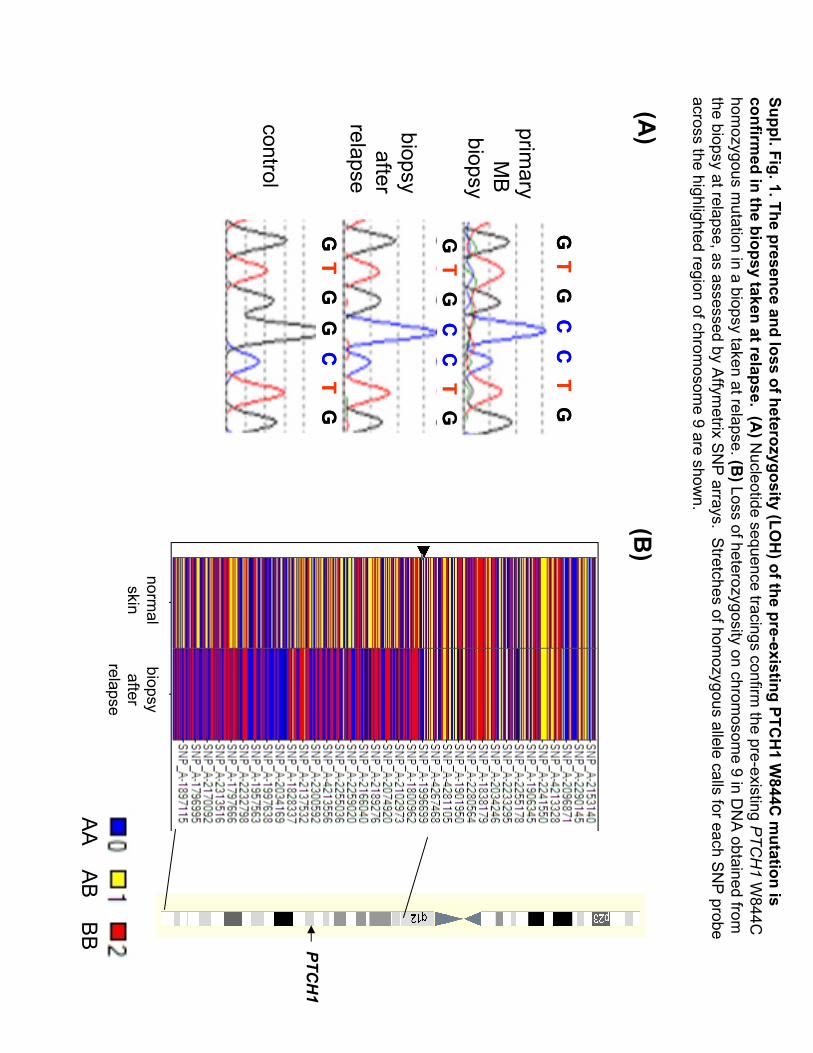
Suppl. Fig. 1.The presence and loss of heterozygosity(LO
H) of the pre-existing PTC
H1 W
844C m
utation is confirm
ed in the biopsy taken at relapse.(A
)Nucleotide sequence tracings confirm
the pre-existing PTCH1
W844C
hom
ozygous mutation in a biopsy taken at relapse. (B
)Loss of heterozygosityon chrom
osome 9 in D
NA
obtained from
the biopsy at relapse, as assessed by Affym
etrixS
NP
arrays.S
tretches of homozygous
allele calls for each SNP
probe across the highlighted region of chrom
osome 9 are show
n.
AA
AB B
B PTCH1
(A)
(B)
normal
skinbiopsyafter
relapse

Suppl. Fig. 2.PTCH
1-W844C
is unable to suppress Hh
pathway activity. G
LI-luciferasereporter activity follow
ing cotransfection
of various input ratios of SMO
and either WT (closed circles) or W
844C (open circles) P
TCH1
DN
A in
C3H
10T1/2 cells.
0 2 4 6 8 10 12
no PTC
H1
40:1 20:1
10:1 5:1
2:1
SMO:PTC
H1
ratio
GLI- luciferase activity (nRLU)
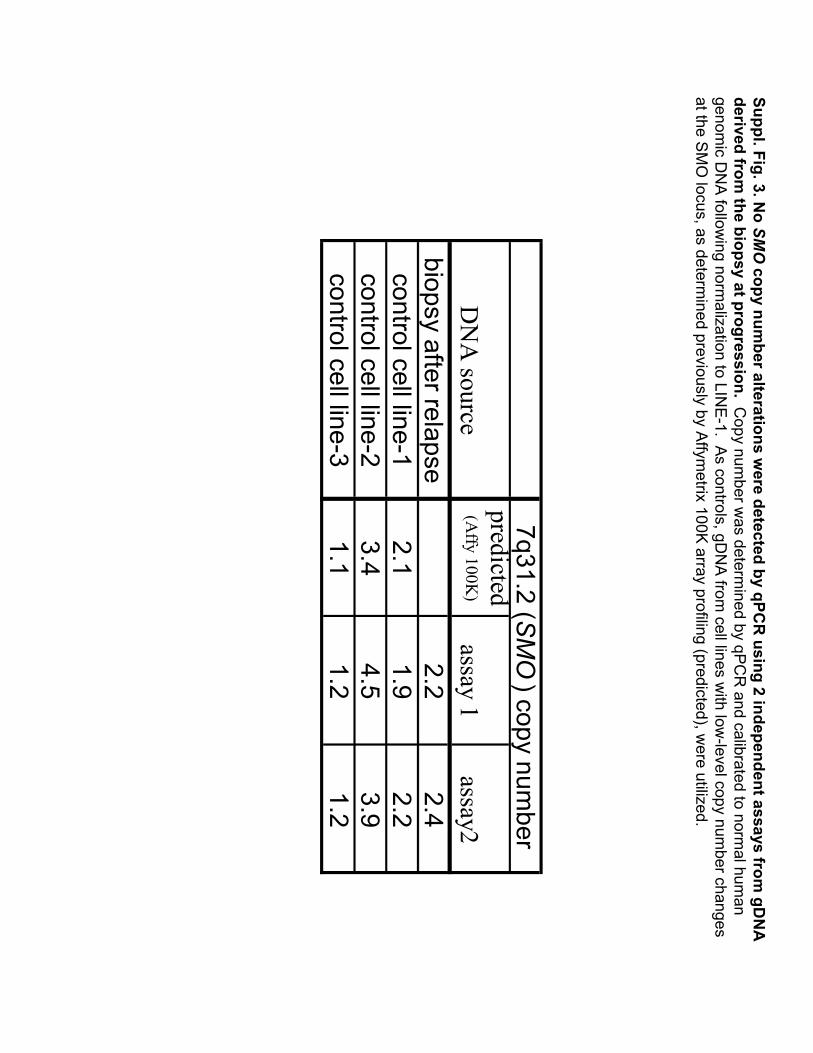
Suppl. Fig. 3.No SM
Ocopy num
ber alterations were detected by qPC
Rusing 2 independent assays from
gDN
Aderived from
the biopsy at progression. Copy num
ber was determ
ined by qPCR
and calibrated to normal hum
an genom
ic DN
A following norm
alization to LINE-1. A
s controls, gDN
Afrom
cell lines with low
-level copy number changes
at the SM
O locus, as determ
ined previously by Affym
etrix100K
array profiling (predicted), were utilized.
DN
A source
predicted (A
ffy 100K)
assay 1assay2
biopsy after relapse2.2
2.4control cell line-1
2.11.9
2.2control cell line-2
3.44.5
3.9control cell line-3
1.11.2
1.2
7q31.2 (SMO
) copy number

Suppl. Fig. 4.Mass spectra of extended D
NA
products for SMO
WT and the D
473H variant (asterisk) from
m
ultiple biopsies.The prim
ary and metastatic m
edulloblastoma
(MB
) biopsies were both taken prior to G
DC
-0449 treatm
ent. Mass spectra intensities represent arbitrary units.
48004900
50005100
52005300
54005500
56004800
49005000
51005200
53005400
55005600
Mass
*
Intensity
primary M
B biopsy
metastatic M
B biopsy
normal skin
biopsy after relapse

Suppl. Fig. 5.Im
munoblotanalysis of FLA
G-tagged, SM
O-W
T and SMO
-D473H
transfectedC
3H10T1/2 cells
probed with anti-FLA
G or anti-actin
as a loading control
-"# -
-%&'-
-"# -
-%&'-
control
SMO-WT
SMO-D473H
IB:
anti-FLAGIB
:anti-actin

0200
400600
8001000
FSC
-H: Forw
ard Scatter
100
101
102
103
104
FL2-H: PE FL2-Height
0.57
0200
400600
8001000
FSC
-H: Forw
ard Scatter
100
101
102
103
104
FL2-H: PE FL2-Height
16.8
0200
400600
8001000
FSC
-H: Forw
ard Scatter
100
101
102
103
104
FL2-H: PE FL2-Height
18.2
Em
pty S
MO
-WT
SM
O-D
473H
Suppl. Fig. 6. Flow cytom
etryanalysis of FLA
G-tagged, SM
O-W
T and SMO
-D473H
transfectedC
3H10T1/2 cells

0 20 40 60 80
100
120
140
0.00.1
1.010.0
100.01000.0
10000.0
KAAD
-cyclopamine (nM
)
Gli -luciferase activity (% control)
0 20 40 60 80
100
120
140
0.00.1
1.010.0
100.01000.0
10000.0
KAAD
-cyclopamine (nM
)
Gli -luciferase activity (% control)
Suppl. Fig. 7. SMO
-D473H
impairs the ability of K
AA
D-cyclopam
ineto suppress H
hsignaling. G
LI-luciferasereporter activity in C
3H10T1/2 cells transfected
with S
MO
-WT (closed circles) or S
MO
-D473H
(open circles) after treatm
ent with various doses of K
AA
D-cyclopam
ine
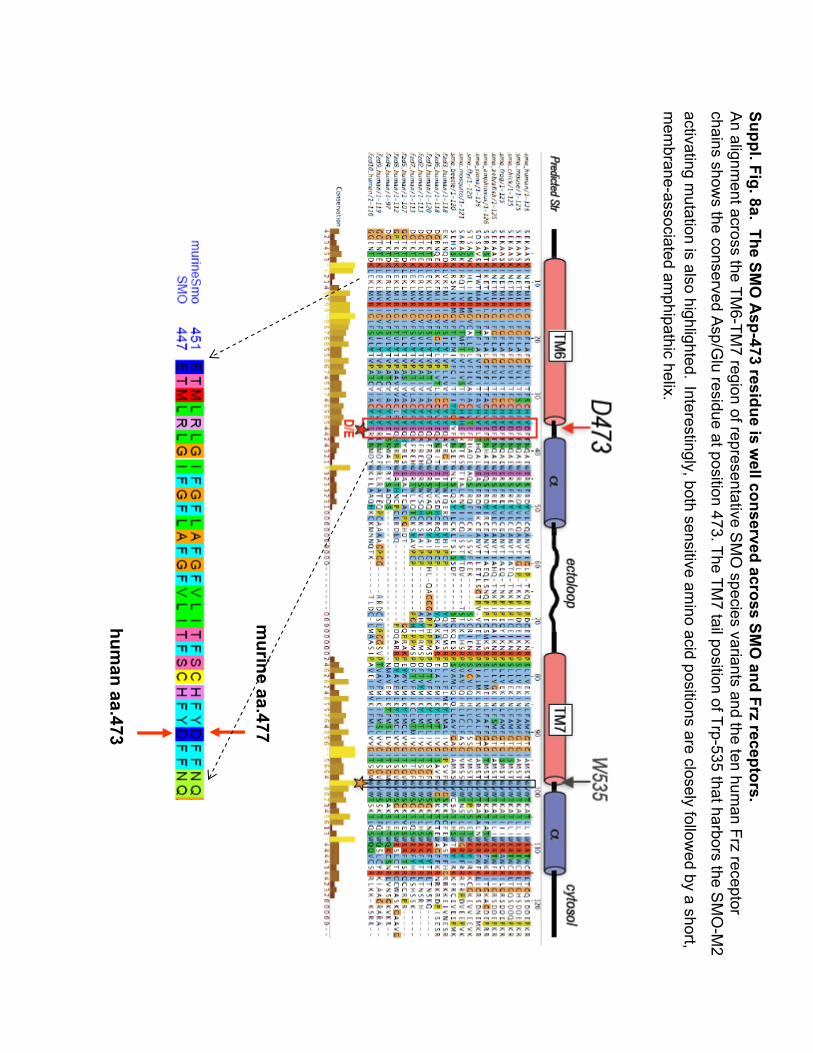
Suppl. Fig. 8a.The SM
O A
sp-473 residue is well conserved across SM
O and Frz
receptors.A
n alignment across the TM
6-TM7 region of representative S
MO
species variants and the ten human Frz
receptor chains show
s the conserved Asp/G
luresidue at position 473. The TM
7 tail position of Trp-535 that harbors the SM
O-M
2
activating mutation is also highlighted.
Interestingly, both sensitive amino acid positions are closely follow
ed by a short, m
embrane-associated am
phipathichelix.
human aa.473
murine aa.477
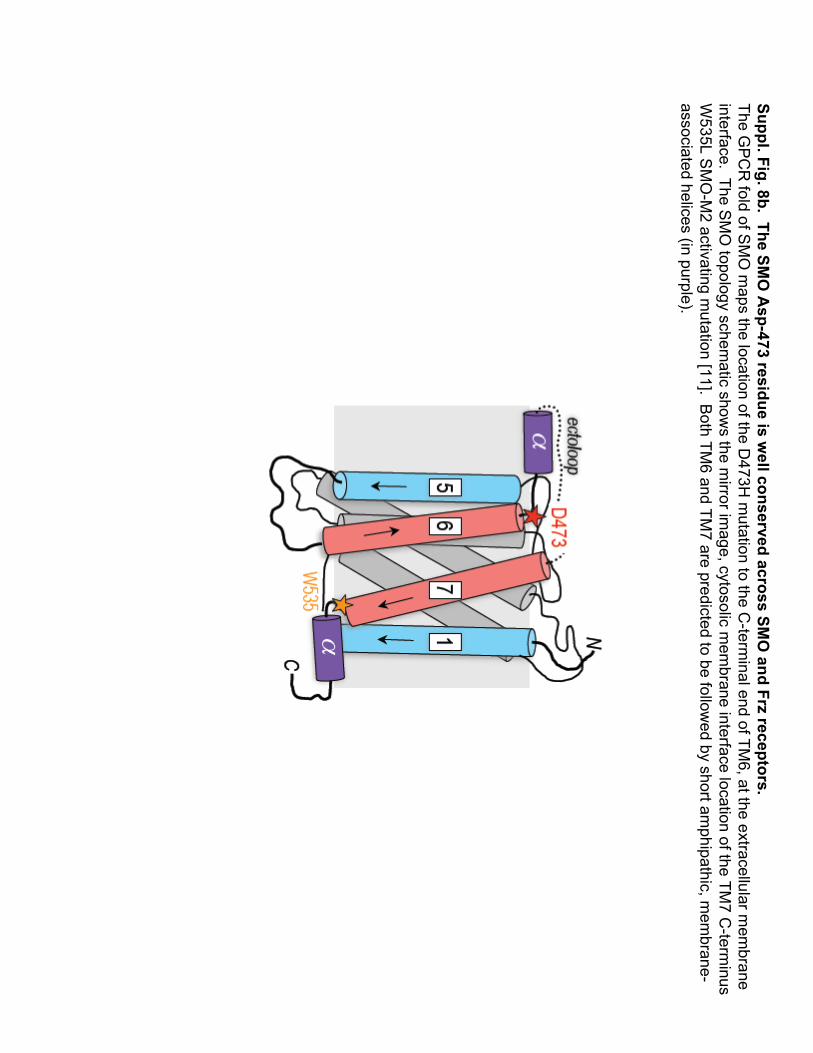
Suppl. Fig. 8b.The SM
O A
sp-473 residue is well conserved across SM
O and Frz
receptors.The G
PC
R fold of S
MO
maps the location of the D
473H m
utation to the C-term
inal end of TM6, at the extracellular m
embrane
interface. The SM
O topology schem
atic shows the m
irror image, cytosolic
mem
brane interface location of the TM7 C
-terminus
W535L S
MO
-M2 activating m
utation [11]. Both TM
6 and TM7 are predicted to be follow
ed by short amphipathic, m
embrane-
associated helices (in purple).
Top Related
Copyright © 2022 FDOKUMEN

