







Bahasa
Halaman
Hukum


Abstract Poor seedling establishment is a major
deterrent in adopting direct seeding of rice. Seed
priming to obtain better crop stand could be an
attractive approach. The objective of this study
was to determine the effectiveness of seed priming
strategies on the improved agronomic characters
of direct-sown rice. Seed priming strategies were:
hydropriming for 48 h, osmohardening with KCl
or CaCl2 for 24 h, ascorbate priming for 48 h and
seed hardening for 24 h, pre-germination (tradi-
tional soaking for nursery raising) and untreated
control. Seed priming improved germination and
emergence, allometry, kernel yield, and its quality,
whilst pre-germination displayed poor and erratic
emergence of seedling followed by poor plant
performance. Faster and uniform emergence was
due to improved a-amylase activity, which in-
creased the level of soluble sugars in the primed
kernels. Osmohardening with KCl gave greater
kernel and straw yield and harvest index, followed
by that of CaCl2, hardening and ascorbate prim-
ing. Improved yield was attributed principally to
number of fertile tillers and 1000 kernel weight. A
positive correlation between mean emergence
time and days to heading, while a negative one
between kernel yield and harvest index suggested
long-term effects of seed priming on plant growth
and development. The results suggest that physi-
ological changes produced by osmohardening
enhanced the starch hydrolysis and made more
sugars available for embryo growth, vigorous
seedling production and, later on, improved
allometric, kernel yield and quality attributes.
Keywords a-Amylase Æ Allometry Æ Direct
seeding Æ Osmohardening Æ Seedling vigor Æ Paddy
quality Æ Rice
Introduction
Traditionally, rice is transplanted after puddling,
while wheat cultivation followed by rice, requires
pulverized soil. This reflects an edaphic conflict in
traditional soil management for rice and its del-
eterious effects on the soil environment for the
succeeding wheat and other upland crops. Pud-
dling requires an excess of water at a time when
the reservoirs are low. Late onset of monsoon and
drudgery of operations often delay rice trans-
plantation, which leads to late vacation of fields,
forcing the sowing of wheat when the appropriate
time has passed. Furthermore, in view of the
M. Farooq Æ S. M. A. BarsaDepartment of Crop Physiology, University ofAgriculture, Faisalabad 38040, Pakistan
A. Wahid (&)Department of Botany, University of Agriculture,Faisalabad 38040, Pakistane-mail: [email protected]
Plant Growth Regul (2006) 49:285–294
DOI 10.1007/s10725-006-9138-y
123
ORIGINAL PAPER
Priming of field-sown rice seed enhances germination,seedling establishment, allometry and yield
Muhammad Farooq Æ Shahzad M. A. Barsa ÆAbdul Wahid
Received: 23 February 2006 / Accepted: 12 April 2006 / Published online: 3 November 2006� Springer Science+Business Media B.V. 2006

depleting water resources, it is desirable that rice
culture should also be like wheat so that it can
continually benefit the cropping system in
improving productivity. Direct seeding of rice,
may have certain benefits. Firstly, it eliminates
puddling and labor of nursery growing and
transplantation, and provides an option to resolve
the edaphic conflict. Secondly, it ensures the rice–
wheat cropping system and facilitates timely
establishment of succeeding winter crops. Lastly,
unlike puddled, direct-seeded fields show no soil
crack problems, saving irrigation water. In grow-
ing a successful direct-seeded crop, issues like
weed management and erratic emergence require
serious attention (Balasubramanian and Hill
2002). This necessitates finding strategies to
ensure faster and uniform crop stand.
Improved seed priming techniques are used to
reduce emergence time, accomplish uniform
emergence, better allometric (changes in growth of
plant parts over time) attributes and requisite stand
in many horticultural and field crops (Ashraf and
Foolad 2005; Farooq et al. 2005). These include
hydropriming, osmoconditioning, osmohardening,
hardening, and hormonal priming or soaking prior
to sowing (Basra et al. 2005; Ashraf and Foolad
2005). Effects of priming or pre-treatment of seed
persist under suboptimum field conditions, such as
salinity (Muhyaddin and Weibe 1989; Wahid et al.
2006), low or high temperature (Bradford et al.
1990; Pill and Finch-Savage 1988; Wahid and
Shabbir 2005) and low soil moisture availability
(Lee et al. 1998; Du and Tuong 2002). Different
seed priming tools have been successfully inte-
grated (Taylor et al. 1998; Basra et al. 2004; Farooq
et al. 2006b). Seed hardening is done in water (Lee
et al. 1998; Basra et al. 2005) and priming is per-
formed in a single cycle of wetting and drying (Lee
and Kim 1999). Until recently, Farooq et al.
(2006b) introduced a new technique of osmohar-
dening for rice seed invigoration, in which both
hardening and osmoconditioning were integrated.
Rice seeds were hardened in various salt solutions
instead of tap or distilled water. Osmohardening in
CaCl2 (ws –1.25 MPa) solution was more effective
for vigor enhancement than simple hardening.
Seed priming is beneficial in many respects.
For instance, it increases the activities of the
enzymes amylase and dehydrogenase in soybean
(Saha et al. 1990), and counteracts the adverse
effects on peroxidation of membrane lipids
(Bailly et al. 2000; Hsu et al. 2003). Seed priming
induces de novo biosynthesis of a-amylase (Lee
and Kim 2000), a key metabolic event in pro-
ducing vigorous seedlings. In a greenhouse study,
osmopriming (with CaCl2 and CaCl2 + NaCl)
improved seedling vigor and stand establishment
in flooded soil (Ruan et al. 2002). Likewise,
priming with 4% KCl solution or a saturated
CaHPO4 solution, increased plant density, fertile
tillers, and grain yield compared with unprimed
treatment when sown in soil with low moisture
content. This suggests that in drought-prone
areas, seed priming can economize seed rate, but
priming could be detrimental if seeding is done
when soil is at or near saturation (Du and Tuong
2002).
Although reports are available on the physio-
logical enhancements of direct-seeded rice (Du
and Tuong 2002; Ruan et al. 2002), no compre-
hensive study has evaluated the response of wide-
ranging seed priming treatments for enhancing
seedling establishment, plant allometry or the
quality of harvested paddy. Information is also
scarce on the physiological implications of prim-
ing-triggered enhancement in germination,
growth or yield, and their inter-relationships
using primed direct-seeded rice. It is surmised
that the priming of seed is beneficial in improving
the agronomic characters of rice under aerobic
conditions without compromising the quality of
harvested paddy. Here we tested this prediction
by evaluating the effects of a range of seed
priming strategies on germination, growth,
allometry and quality of harvested paddy as well
as some physiological determinants of growth
promotion in direct field-sown rice.
Material and methods
Experimental details and seed priming
treatments
Coarse rice (Oryza sativa L. cv. KS-282) seed for
this study was obtained from the Rice Research
286 Plant Growth Regul (2006) 49:285–294
123

Institute, Kala Shah Kakoo, Pakistan. Moisture
content of the seed was ca 8%. The study was
conducted in plots (6.5 m · 4.5 m) at a farm in
the rice growing belt during in the years 2004 and
2005. The experiment was laid out as a random-
ized complete block design (RCBD) with three
replications.
Seed priming treatments, chosen from the pre-
vious experience (Basra et al. 2005; Farooq et al.
2005, 2006a, b), were: (a) hydropriming, soaking
seed in aerated distilled water for 48 h, (b) hard-
ening, soaking seeds in tap water at 27�C ± 3 for
24 h and redrying to initial moisture content and
this cycle repeated once (Lee et al. 1998; Basra
et al. 2005; Farooq et al. 2005); (c, d) osmohar-
dening, similar to hardening but in the presence of
CaCl2 or KCl solutions of ws = –1.25 MPa
(Farooq et al. 2006b) and (e) ascorbate priming,
soaking seeds in an aerated solution of 10 mg l–1
ascorbic acid for 48 h. Pre-germination, soaking
seeds in water for 24 h followed by placing them
between two layers of saturated gunny bags up to
radicle appearance (chitting stage), was used to
compare the traditional rice sowing strategy for
raising nursery, while controls were seeds receiv-
ing no prior treatment. Primed seeds were given
three washings with water and re-dried closer to
original moisture (ca. 8%) under forced air at
27�C ± 3 (except for pre-emergence). These seeds
were put in polythene bags and stored in a
refrigerator at 5 ± 1�C until used.
Seed sowing and agronomic practices
Field soil was sandy clay loam with pH 8.1, elec-
trical conductivity 0.30 dS m–1 and organic matter
0.75%. Land was ploughed five times with tractor
drawn implements to achieve the required seed-
bed. Fertilizer was applied as urea (46%), single
superphosphate (18% P2O5), sulphate of potash
(50% K2O) and ZnSO4 (35% Zn) at recom-
mended doses. Whole quantities of phosphorus,
potash and zinc, and a half-dose of nitrogen were
applied prior to sowing. Remaining nitrogen was
applied in two equal splits, one at tillering and the
other at panicle initiation. Seed was hand drilled
at 65 kg ha–1 in 22 cm spaced rows on June 1,
2004. Plots were irrigated when the soil moisture
was slightly below field capacity. For weed
control, a mixture of ethoxy sulphuran and
phenoxyprop-p-ethyl at 200 g and 370 ml ha–1,
respectively was applied 20 days after sowing in
saturated soil. Ten irrigations were applied during
the crop growth period. Irrigation was halted
10 days prior to harvesting.
Emergence, growth and yield data
Days to start of emergence were recorded and
mean emergence time (MET), days to 50%
emergence (E50) and final emergence percentage
(FEP) were computed (Ellis and Roberts 1981).
Days taken from emergence to heading and from
heading to maturity were noted. Early at physi-
ological maturity (25 Aug), data on agronomic
characters were recorded four times at 15-day
intervals while spike and kernel characteristics
and yield components were recorded at full
maturity (25 Oct). Leaf area was measured using
a leaf area meter (Licor, Model 3100). Leaf area
index (LAI), crop growth rate (CGR), and net
assimilation rate (NAR) were calculated using
the formulae of Hunt (1978). The crop was har-
vested when fully ripe to determine paddy and
straw yield, and harvest index.
a-Amylase activity and soluble sugar content
For a-amylase activity, 1 g ground seed sample
was mixed with 10 ml phosphate buffer (pH 7.0)
and left for 24 h at 4�C. The enzyme activity was
determined from the supernatant by the DNS
method (Bernfeld 1955). For soluble sugars, 1 g
ground seed sample was mixed with 10 ml dis-
tilled water and left for 24 h at 25�C (Lee and
Kim 2000). The mixture was filtered (with
Whatman No. 42) and the final volume made to
10 ml with distilled water. Total soluble sugars
were determined by the phenol sulfuric method
(Dubois et al. 1956).
Spikelet and kernel quality characteristics
A common electric lamp with an adjustable stand
was used as a light source to determine the pan-
icle and kernel characteristics. A panicle was
positioned in front of the lamp so as to pass light
through it. This enabled the separation of sterile
Plant Growth Regul (2006) 49:285–294 287
123
spikelets or abortive (kernels that do not develop
after fertilization and look dull under light) and
opaque kernels within a spikelet. The chalky
kernels were separated by visually observing
chalky areas on them with a magnifying glass.
Length and width were taken of 100 kernels in
replicate with a digital caliper to determine
length:width ratio. Crude proteins from fresh
kernels were determined from total nitrogen
estimated by the microKjeldahl method (multi-
plied by the factor 5.95). Amylose content of the
fresh milled kernel and kernel water absorption
ratio was determined as described by Juliano
et al. (1965) and Juliano (1971).
Statistical analysis
Data were statistically analyzed using the software
MSTAT-C. Analysis of variance was used to test
the significance of variance sources, while LSD test
(p = 0.05) was used to compare the differences
among treatment means. Trend lines were set and
linear correlation coefficients were determined to
find the relationship of different attributes.
Results
Characteristics of primed seeds
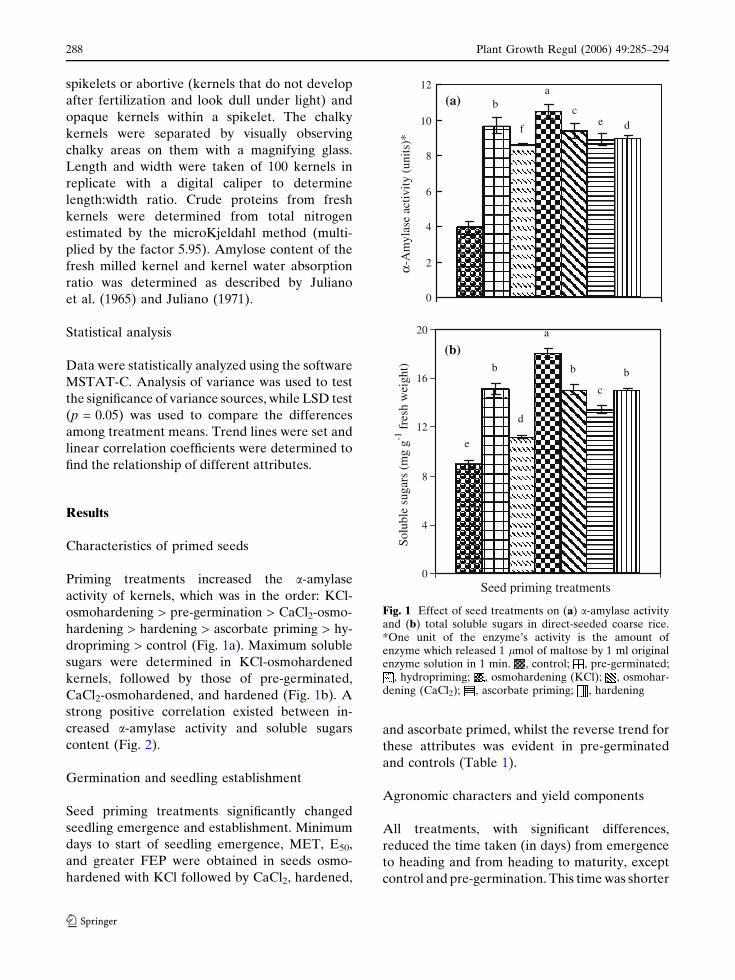
Priming treatments increased the a-amylase
activity of kernels, which was in the order: KCl-
osmohardening > pre-germination > CaCl2-osmo-
hardening > hardening > ascorbate priming > hy-
dropriming > control (Fig. 1a). Maximum soluble
sugars were determined in KCl-osmohardened
kernels, followed by those of pre-germinated,
CaCl2-osmohardened, and hardened (Fig. 1b). A
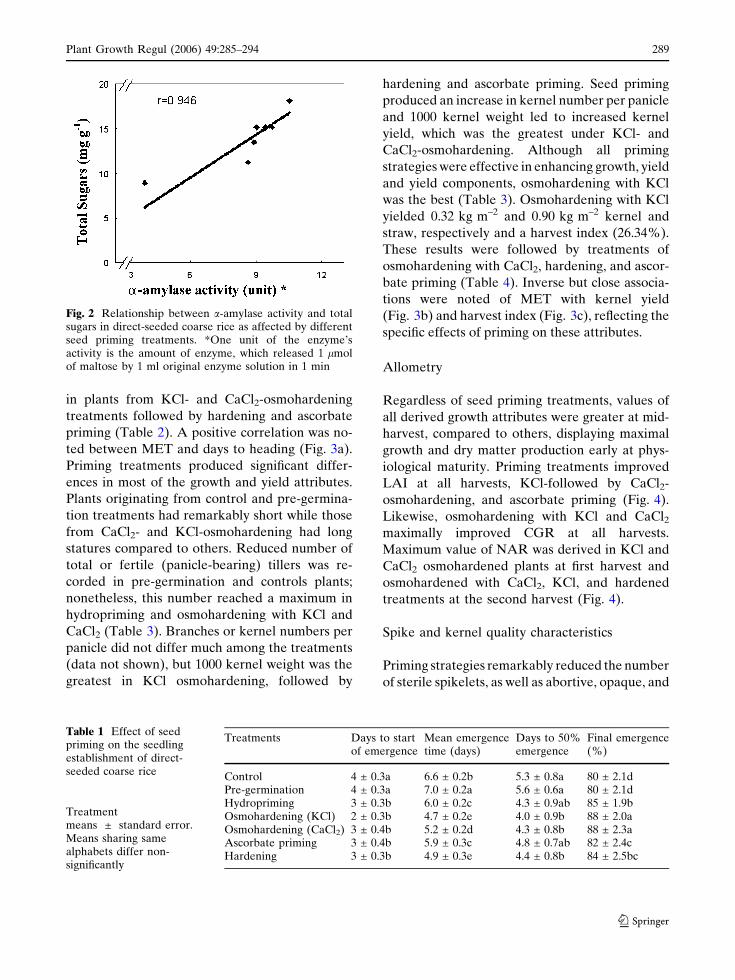
strong positive correlation existed between in-
creased a-amylase activity and soluble sugars
content (Fig. 2).
Germination and seedling establishment
Seed priming treatments significantly changed
seedling emergence and establishment. Minimum
days to start of seedling emergence, MET, E50,
and greater FEP were obtained in seeds osmo-
hardened with KCl followed by CaCl2, hardened,
and ascorbate primed, whilst the reverse trend for
these attributes was evident in pre-germinated
and controls (Table 1).
Agronomic characters and yield components
All treatments, with significant differences,
reduced the time taken (in days) from emergence
to heading and from heading to maturity, except
control and pre-germination. This time was shorter
α-A
myl
ase
activ
ity (
units
)*
(a) b
f
a
ce d
0
2
4
6
8
10
12
Solu
ble
suga
rs (
mg
g-1 f
resh
wei
ght)
(b)
e
b
d
a
b
c
b
0
4
8
12
16
20
Seed priming treatments
Fig. 1 Effect of seed treatments on (a) a-amylase activityand (b) total soluble sugars in direct-seeded coarse rice.*One unit of the enzyme’s activity is the amount ofenzyme which released 1 lmol of maltose by 1 ml originalenzyme solution in 1 min. , control; , pre-germinated;
, hydropriming; , osmohardening (KCl); , osmohar-dening (CaCl2); , ascorbate priming; , hardening
288 Plant Growth Regul (2006) 49:285–294
123
in plants from KCl- and CaCl2-osmohardening
treatments followed by hardening and ascorbate
priming (Table 2). A positive correlation was no-
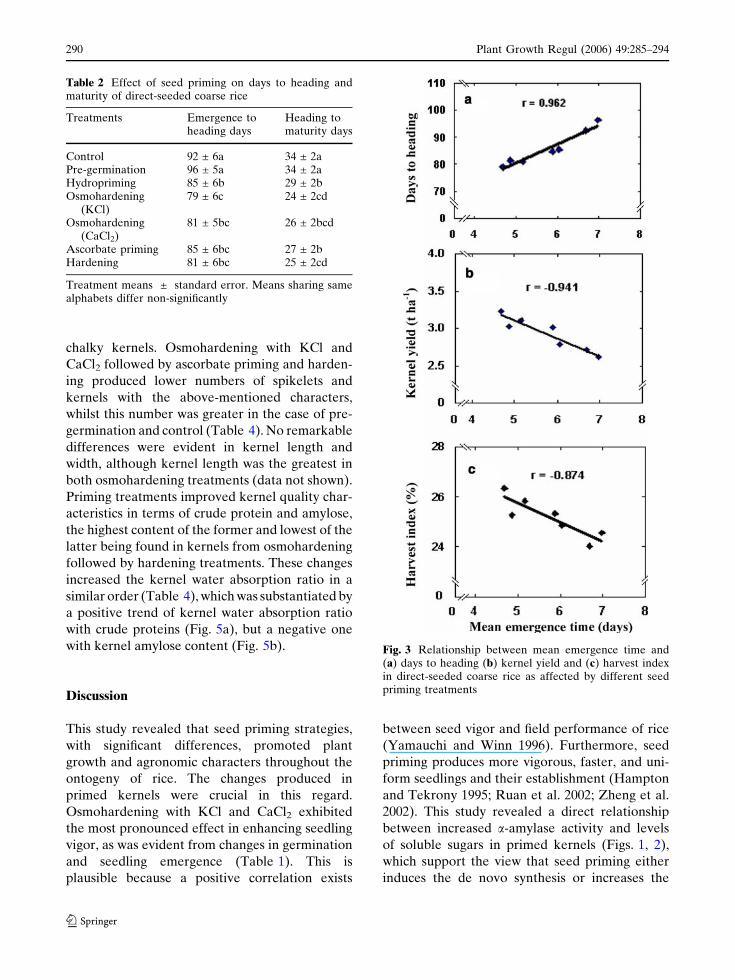
ted between MET and days to heading (Fig. 3a).
Priming treatments produced significant differ-
ences in most of the growth and yield attributes.
Plants originating from control and pre-germina-
tion treatments had remarkably short while those
from CaCl2- and KCl-osmohardening had long
statures compared to others. Reduced number of
total or fertile (panicle-bearing) tillers was re-
corded in pre-germination and controls plants;
nonetheless, this number reached a maximum in
hydropriming and osmohardening with KCl and
CaCl2 (Table 3). Branches or kernel numbers per
panicle did not differ much among the treatments
(data not shown), but 1000 kernel weight was the
greatest in KCl osmohardening, followed by
hardening and ascorbate priming. Seed priming
produced an increase in kernel number per panicle
and 1000 kernel weight led to increased kernel
yield, which was the greatest under KCl- and
CaCl2-osmohardening. Although all priming
strategies were effective in enhancing growth, yield
and yield components, osmohardening with KCl
was the best (Table 3). Osmohardening with KCl
yielded 0.32 kg m–2 and 0.90 kg m–2 kernel and
straw, respectively and a harvest index (26.34%).
These results were followed by treatments of
osmohardening with CaCl2, hardening, and ascor-
bate priming (Table 4). Inverse but close associa-
tions were noted of MET with kernel yield
(Fig. 3b) and harvest index (Fig. 3c), reflecting the
specific effects of priming on these attributes.
Allometry
Regardless of seed priming treatments, values of
all derived growth attributes were greater at mid-
harvest, compared to others, displaying maximal
growth and dry matter production early at phys-
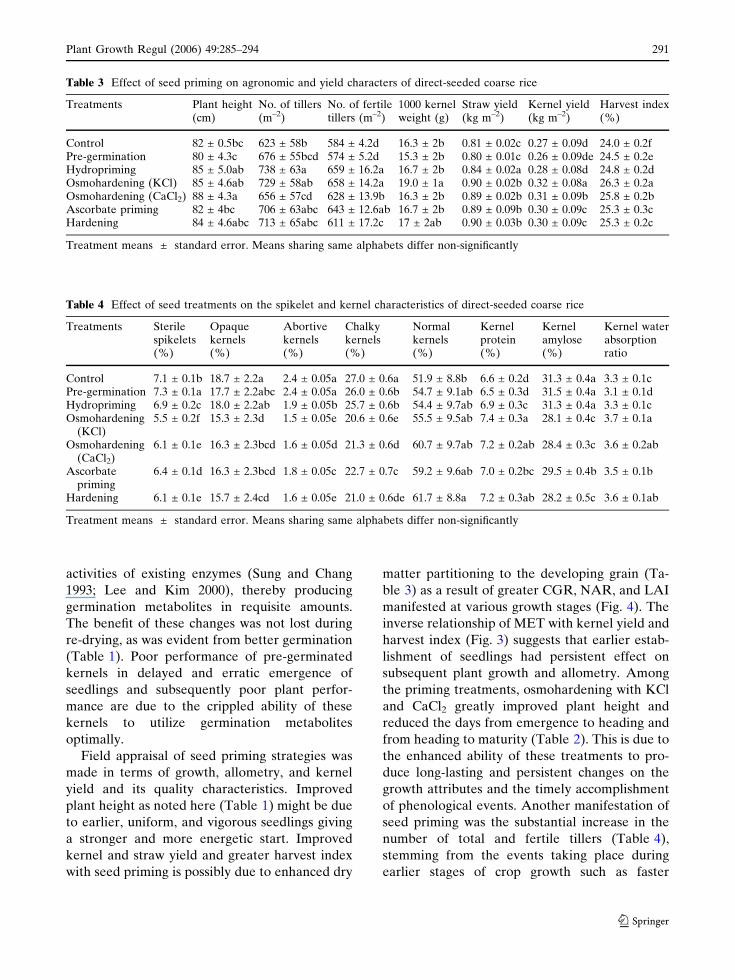
iological maturity. Priming treatments improved
LAI at all harvests, KCl-followed by CaCl2-
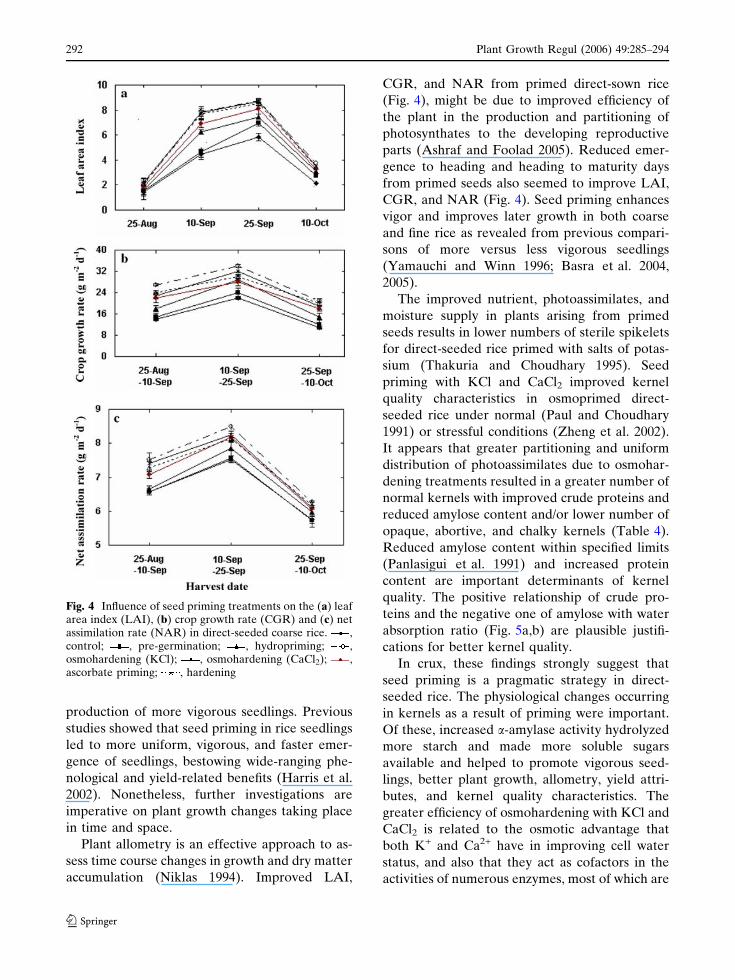
osmohardening, and ascorbate priming (Fig. 4).
Likewise, osmohardening with KCl and CaCl2maximally improved CGR at all harvests.
Maximum value of NAR was derived in KCl and
CaCl2 osmohardened plants at first harvest and
osmohardened with CaCl2, KCl, and hardened
treatments at the second harvest (Fig. 4).
Spike and kernel quality characteristics
Priming strategies remarkably reduced the number
of sterile spikelets, as well as abortive, opaque, and
Fig. 2 Relationship between a-amylase activity and totalsugars in direct-seeded coarse rice as affected by differentseed priming treatments. *One unit of the enzyme’sactivity is the amount of enzyme, which released 1 lmolof maltose by 1 ml original enzyme solution in 1 min
Table 1 Effect of seedpriming on the seedlingestablishment of direct-seeded coarse rice
Treatmentmeans ± standard error.Means sharing samealphabets differ non-significantly
Treatments Days to startof emergence
Mean emergencetime (days)
Days to 50%emergence
Final emergence(%)
Control 4 ± 0.3a 6.6 ± 0.2b 5.3 ± 0.8a 80 ± 2.1dPre-germination 4 ± 0.3a 7.0 ± 0.2a 5.6 ± 0.6a 80 ± 2.1dHydropriming 3 ± 0.3b 6.0 ± 0.2c 4.3 ± 0.9ab 85 ± 1.9bOsmohardening (KCl) 2 ± 0.3b 4.7 ± 0.2e 4.0 ± 0.9b 88 ± 2.0aOsmohardening (CaCl2) 3 ± 0.4b 5.2 ± 0.2d 4.3 ± 0.8b 88 ± 2.3aAscorbate priming 3 ± 0.4b 5.9 ± 0.3c 4.8 ± 0.7ab 82 ± 2.4cHardening 3 ± 0.3b 4.9 ± 0.3e 4.4 ± 0.8b 84 ± 2.5bc
Plant Growth Regul (2006) 49:285–294 289
123
chalky kernels. Osmohardening with KCl and
CaCl2 followed by ascorbate priming and harden-
ing produced lower numbers of spikelets and
kernels with the above-mentioned characters,
whilst this number was greater in the case of pre-
germination and control (Table 4). No remarkable
differences were evident in kernel length and
width, although kernel length was the greatest in
both osmohardening treatments (data not shown).
Priming treatments improved kernel quality char-
acteristics in terms of crude protein and amylose,
the highest content of the former and lowest of the
latter being found in kernels from osmohardening
followed by hardening treatments. These changes
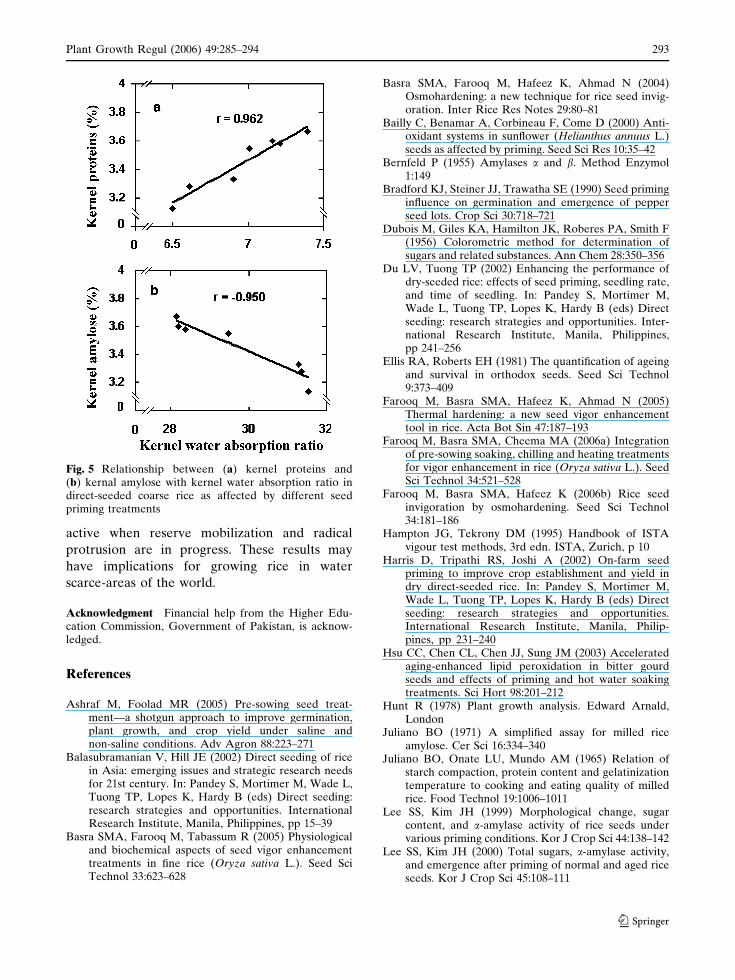
increased the kernel water absorption ratio in a
similar order (Table 4), which was substantiated by
a positive trend of kernel water absorption ratio
with crude proteins (Fig. 5a), but a negative one
with kernel amylose content (Fig. 5b).
Discussion
This study revealed that seed priming strategies,
with significant differences, promoted plant
growth and agronomic characters throughout the
ontogeny of rice. The changes produced in
primed kernels were crucial in this regard.
Osmohardening with KCl and CaCl2 exhibited
the most pronounced effect in enhancing seedling
vigor, as was evident from changes in germination
and seedling emergence (Table 1). This is
plausible because a positive correlation exists
between seed vigor and field performance of rice
(Yamauchi and Winn 1996). Furthermore, seed
priming produces more vigorous, faster, and uni-
form seedlings and their establishment (Hampton
and Tekrony 1995; Ruan et al. 2002; Zheng et al.
2002). This study revealed a direct relationship
between increased a-amylase activity and levels
of soluble sugars in primed kernels (Figs. 1, 2),
which support the view that seed priming either
induces the de novo synthesis or increases the
Table 2 Effect of seed priming on days to heading andmaturity of direct-seeded coarse rice
Treatments Emergence toheading days
Heading tomaturity days
Control 92 ± 6a 34 ± 2aPre-germination 96 ± 5a 34 ± 2aHydropriming 85 ± 6b 29 ± 2bOsmohardening
(KCl)79 ± 6c 24 ± 2cd
Osmohardening(CaCl2)
81 ± 5bc 26 ± 2bcd
Ascorbate priming 85 ± 6bc 27 ± 2bHardening 81 ± 6bc 25 ± 2cd
Treatment means ± standard error. Means sharing samealphabets differ non-significantly
Fig. 3 Relationship between mean emergence time and(a) days to heading (b) kernel yield and (c) harvest indexin direct-seeded coarse rice as affected by different seedpriming treatments
290 Plant Growth Regul (2006) 49:285–294
123
activities of existing enzymes (Sung and Chang
1993; Lee and Kim 2000), thereby producing
germination metabolites in requisite amounts.
The benefit of these changes was not lost during
re-drying, as was evident from better germination
(Table 1). Poor performance of pre-germinated
kernels in delayed and erratic emergence of
seedlings and subsequently poor plant perfor-
mance are due to the crippled ability of these
kernels to utilize germination metabolites
optimally.
Field appraisal of seed priming strategies was
made in terms of growth, allometry, and kernel
yield and its quality characteristics. Improved
plant height as noted here (Table 1) might be due
to earlier, uniform, and vigorous seedlings giving
a stronger and more energetic start. Improved
kernel and straw yield and greater harvest index
with seed priming is possibly due to enhanced dry
matter partitioning to the developing grain (Ta-
ble 3) as a result of greater CGR, NAR, and LAI
manifested at various growth stages (Fig. 4). The
inverse relationship of MET with kernel yield and
harvest index (Fig. 3) suggests that earlier estab-
lishment of seedlings had persistent effect on
subsequent plant growth and allometry. Among
the priming treatments, osmohardening with KCl
and CaCl2 greatly improved plant height and
reduced the days from emergence to heading and
from heading to maturity (Table 2). This is due to
the enhanced ability of these treatments to pro-
duce long-lasting and persistent changes on the
growth attributes and the timely accomplishment
of phenological events. Another manifestation of
seed priming was the substantial increase in the
number of total and fertile tillers (Table 4),
stemming from the events taking place during
earlier stages of crop growth such as faster
Table 3 Effect of seed priming on agronomic and yield characters of direct-seeded coarse rice
Treatments Plant height(cm)
No. of tillers(m–2)
No. of fertiletillers (m–2)
1000 kernelweight (g)
Straw yield(kg m–2)
Kernel yield(kg m–2)
Harvest index(%)
Control 82 ± 0.5bc 623 ± 58b 584 ± 4.2d 16.3 ± 2b 0.81 ± 0.02c 0.27 ± 0.09d 24.0 ± 0.2fPre-germination 80 ± 4.3c 676 ± 55bcd 574 ± 5.2d 15.3 ± 2b 0.80 ± 0.01c 0.26 ± 0.09de 24.5 ± 0.2eHydropriming 85 ± 5.0ab 738 ± 63a 659 ± 16.2a 16.7 ± 2b 0.84 ± 0.02a 0.28 ± 0.08d 24.8 ± 0.2dOsmohardening (KCl) 85 ± 4.6ab 729 ± 58ab 658 ± 14.2a 19.0 ± 1a 0.90 ± 0.02b 0.32 ± 0.08a 26.3 ± 0.2aOsmohardening (CaCl2) 88 ± 4.3a 656 ± 57cd 628 ± 13.9b 16.3 ± 2b 0.89 ± 0.02b 0.31 ± 0.09b 25.8 ± 0.2bAscorbate priming 82 ± 4bc 706 ± 63abc 643 ± 12.6ab 16.7 ± 2b 0.89 ± 0.09b 0.30 ± 0.09c 25.3 ± 0.3cHardening 84 ± 4.6abc 713 ± 65abc 611 ± 17.2c 17 ± 2ab 0.90 ± 0.03b 0.30 ± 0.09c 25.3 ± 0.2c
Treatment means ± standard error. Means sharing same alphabets differ non-significantly
Table 4 Effect of seed treatments on the spikelet and kernel characteristics of direct-seeded coarse rice
Treatments Sterilespikelets(%)
Opaquekernels(%)
Abortivekernels(%)
Chalkykernels(%)
Normalkernels(%)
Kernelprotein(%)
Kernelamylose(%)
Kernel waterabsorptionratio
Control 7.1 ± 0.1b 18.7 ± 2.2a 2.4 ± 0.05a 27.0 ± 0.6a 51.9 ± 8.8b 6.6 ± 0.2d 31.3 ± 0.4a 3.3 ± 0.1cPre-germination 7.3 ± 0.1a 17.7 ± 2.2abc 2.4 ± 0.05a 26.0 ± 0.6b 54.7 ± 9.1ab 6.5 ± 0.3d 31.5 ± 0.4a 3.1 ± 0.1dHydropriming 6.9 ± 0.2c 18.0 ± 2.2ab 1.9 ± 0.05b 25.7 ± 0.6b 54.4 ± 9.7ab 6.9 ± 0.3c 31.3 ± 0.4a 3.3 ± 0.1cOsmohardening
(KCl)5.5 ± 0.2f 15.3 ± 2.3d 1.5 ± 0.05e 20.6 ± 0.6e 55.5 ± 9.5ab 7.4 ± 0.3a 28.1 ± 0.4c 3.7 ± 0.1a
Osmohardening(CaCl2)
6.1 ± 0.1e 16.3 ± 2.3bcd 1.6 ± 0.05d 21.3 ± 0.6d 60.7 ± 9.7ab 7.2 ± 0.2ab 28.4 ± 0.3c 3.6 ± 0.2ab
Ascorbatepriming
6.4 ± 0.1d 16.3 ± 2.3bcd 1.8 ± 0.05c 22.7 ± 0.7c 59.2 ± 9.6ab 7.0 ± 0.2bc 29.5 ± 0.4b 3.5 ± 0.1b
Hardening 6.1 ± 0.1e 15.7 ± 2.4cd 1.6 ± 0.05e 21.0 ± 0.6de 61.7 ± 8.8a 7.2 ± 0.3ab 28.2 ± 0.5c 3.6 ± 0.1ab
Treatment means ± standard error. Means sharing same alphabets differ non-significantly
Plant Growth Regul (2006) 49:285–294 291
123
production of more vigorous seedlings. Previous
studies showed that seed priming in rice seedlings
led to more uniform, vigorous, and faster emer-
gence of seedlings, bestowing wide-ranging phe-
nological and yield-related benefits (Harris et al.
2002). Nonetheless, further investigations are
imperative on plant growth changes taking place
in time and space.
Plant allometry is an effective approach to as-
sess time course changes in growth and dry matter
accumulation (Niklas 1994). Improved LAI,
CGR, and NAR from primed direct-sown rice
(Fig. 4), might be due to improved efficiency of
the plant in the production and partitioning of
photosynthates to the developing reproductive
parts (Ashraf and Foolad 2005). Reduced emer-
gence to heading and heading to maturity days
from primed seeds also seemed to improve LAI,
CGR, and NAR (Fig. 4). Seed priming enhances
vigor and improves later growth in both coarse
and fine rice as revealed from previous compari-
sons of more versus less vigorous seedlings
(Yamauchi and Winn 1996; Basra et al. 2004,
2005).
The improved nutrient, photoassimilates, and
moisture supply in plants arising from primed
seeds results in lower numbers of sterile spikelets
for direct-seeded rice primed with salts of potas-
sium (Thakuria and Choudhary 1995). Seed
priming with KCl and CaCl2 improved kernel
quality characteristics in osmoprimed direct-
seeded rice under normal (Paul and Choudhary
1991) or stressful conditions (Zheng et al. 2002).
It appears that greater partitioning and uniform
distribution of photoassimilates due to osmohar-
dening treatments resulted in a greater number of
normal kernels with improved crude proteins and
reduced amylose content and/or lower number of
opaque, abortive, and chalky kernels (Table 4).
Reduced amylose content within specified limits
(Panlasigui et al. 1991) and increased protein
content are important determinants of kernel
quality. The positive relationship of crude pro-
teins and the negative one of amylose with water
absorption ratio (Fig. 5a,b) are plausible justifi-
cations for better kernel quality.
In crux, these findings strongly suggest that
seed priming is a pragmatic strategy in direct-
seeded rice. The physiological changes occurring
in kernels as a result of priming were important.
Of these, increased a-amylase activity hydrolyzed
more starch and made more soluble sugars
available and helped to promote vigorous seed-
lings, better plant growth, allometry, yield attri-
butes, and kernel quality characteristics. The
greater efficiency of osmohardening with KCl and
CaCl2 is related to the osmotic advantage that
both K+ and Ca2+ have in improving cell water
status, and also that they act as cofactors in the
activities of numerous enzymes, most of which are
Fig. 4 Influence of seed priming treatments on the (a) leafarea index (LAI), (b) crop growth rate (CGR) and (c) netassimilation rate (NAR) in direct-seeded coarse rice. ,control; , pre-germination; , hydropriming; ,osmohardening (KCl); , osmohardening (CaCl2); ,ascorbate priming; , hardening
292 Plant Growth Regul (2006) 49:285–294
123
active when reserve mobilization and radical
protrusion are in progress. These results may
have implications for growing rice in water
scarce-areas of the world.
Acknowledgment Financial help from the Higher Edu-cation Commission, Government of Pakistan, is acknow-ledged.
References
Ashraf M, Foolad MR (2005) Pre-sowing seed treat-ment—a shotgun approach to improve germination,plant growth, and crop yield under saline andnon-saline conditions. Adv Agron 88:223–271
Balasubramanian V, Hill JE (2002) Direct seeding of ricein Asia: emerging issues and strategic research needsfor 21st century. In: Pandey S, Mortimer M, Wade L,Tuong TP, Lopes K, Hardy B (eds) Direct seeding:research strategies and opportunities. InternationalResearch Institute, Manila, Philippines, pp 15–39
Basra SMA, Farooq M, Tabassum R (2005) Physiologicaland biochemical aspects of seed vigor enhancementtreatments in fine rice (Oryza sativa L.). Seed SciTechnol 33:623–628
Basra SMA, Farooq M, Hafeez K, Ahmad N (2004)Osmohardening: a new technique for rice seed invig-oration. Inter Rice Res Notes 29:80–81
Bailly C, Benamar A, Corbineau F, Come D (2000) Anti-oxidant systems in sunflower (Helianthus annuus L.)seeds as affected by priming. Seed Sci Res 10:35–42
Bernfeld P (1955) Amylases a and b. Method Enzymol1:149
Bradford KJ, Steiner JJ, Trawatha SE (1990) Seed priminginfluence on germination and emergence of pepperseed lots. Crop Sci 30:718–721
Dubois M, Giles KA, Hamilton JK, Roberes PA, Smith F(1956) Colorometric method for determination ofsugars and related substances. Ann Chem 28:350–356
Du LV, Tuong TP (2002) Enhancing the performance ofdry-seeded rice: effects of seed priming, seedling rate,and time of seedling. In: Pandey S, Mortimer M,Wade L, Tuong TP, Lopes K, Hardy B (eds) Directseeding: research strategies and opportunities. Inter-national Research Institute, Manila, Philippines,pp 241–256
Ellis RA, Roberts EH (1981) The quantification of ageingand survival in orthodox seeds. Seed Sci Technol9:373–409
Farooq M, Basra SMA, Hafeez K, Ahmad N (2005)Thermal hardening: a new seed vigor enhancementtool in rice. Acta Bot Sin 47:187–193
Farooq M, Basra SMA, Cheema MA (2006a) Integrationof pre-sowing soaking, chilling and heating treatmentsfor vigor enhancement in rice (Oryza sativa L.). SeedSci Technol 34:521–528
Farooq M, Basra SMA, Hafeez K (2006b) Rice seedinvigoration by osmohardening. Seed Sci Technol34:181–186
Hampton JG, Tekrony DM (1995) Handbook of ISTAvigour test methods, 3rd edn. ISTA, Zurich, p 10
Harris D, Tripathi RS, Joshi A (2002) On-farm seedpriming to improve crop establishment and yield indry direct-seeded rice. In: Pandey S, Mortimer M,Wade L, Tuong TP, Lopes K, Hardy B (eds) Directseeding: research strategies and opportunities.International Research Institute, Manila, Philip-pines, pp 231–240
Hsu CC, Chen CL, Chen JJ, Sung JM (2003) Acceleratedaging-enhanced lipid peroxidation in bitter gourdseeds and effects of priming and hot water soakingtreatments. Sci Hort 98:201–212
Hunt R (1978) Plant growth analysis. Edward Arnald,London
Juliano BO (1971) A simplified assay for milled riceamylose. Cer Sci 16:334–340
Juliano BO, Onate LU, Mundo AM (1965) Relation ofstarch compaction, protein content and gelatinizationtemperature to cooking and eating quality of milledrice. Food Technol 19:1006–1011
Lee SS, Kim JH (1999) Morphological change, sugarcontent, and a-amylase activity of rice seeds undervarious priming conditions. Kor J Crop Sci 44:138–142
Lee SS, Kim JH (2000) Total sugars, a-amylase activity,and emergence after priming of normal and aged riceseeds. Kor J Crop Sci 45:108–111
Fig. 5 Relationship between (a) kernel proteins and(b) kernal amylose with kernel water absorption ratio indirect-seeded coarse rice as affected by different seedpriming treatments
Plant Growth Regul (2006) 49:285–294 293
123

Lee SS, Kim JH, Hong SB, Yun SH (1998) Effect ofhumidification and hardening treatment on seedemergence of rice. Kor J Crop Sci 43:157–160
Muhyaddin T, Weibe HJ (1989) Effect of seed treatmentswith polyethyleneglycol (PEG) on emergence ofvegetable seeds. Seed Sci Technol 17:49–56
Niklas KJ (1994) Plant allometry. The University ofChicago Press, Chicago, Illinois
Panlasigui LN, Thompson LU, Juliano BO, Perez CM,Yiu SH, Greenberg GR (1991) Rice varieties withsimilar amylose content differ in starch digestibilityand glycemic response in humans. Amer J Clinic Nutr54:871–877
Paul SR, Choudhary AK (1991) Effect of seed primingwith salts on growth and yield of wheat under rainfedconditions. Ann Agric Res 12:415–418
Pill WG, Finch-Savage WE (1988) Effects of combiningpriming and plant growth regulator treatments on thesynchronization of carrot seed emergence. Ann ApplBiol 113:383–389
Ruan S, Xue Q, Tylkowska K (2002) Effects of seedpriming on emergence and health of rice (Oryzasativa L.) seeds. Seed Sci Technol 30:451–458
Saha R, Mandal AK, Basu RN (1990) Physiology of seedinvigoration treatments in soybean (Glycine max L.).Seed Sci Technol 18:269–276
Sung FJ, Chang YH (1993) Biochemical activities associ-ated with priming of sweet corn seeds to improvevigor. Seed Sci Technol 21:97–105
Taylor AG, Allen PS, Bennett MA, Bradford JK, BurrisJS, Misra MK (1998) Seed enhancements. Seed SciRes 8:245–256
Thakuria RK, Choudhary JK (1995) Effect of seed prim-ing, potassium and anti-transpirant on dry-seededrainfed ahu rice (Oryza sativa). Ind J Agron 40:412–414
Wahid A, Shabbir A (2005) Induction of heat stresstolerance in barley seedlings by pre-sowing seedtreatment with glycinebetaine. Plant Growth Regul46:133–141
Wahid A, Parveen M, Gelani S, Basra SMA (2006) Pre-treatment of seeds with H2O2 improves salt toleranceof wheat seedling by alleviation of oxidative damageand expression of stress proteins. J Plant Physiol(published online)
Yamauchi M, Winn T (1996) Rice seed vigor and seedlingestablishment in anaerobic soil. Crop Sci 36:680–686
Zheng HC, Jin HU, Zhi Z, Ruan SL, Song WJ (2002)Effect of seed priming with mixed-salt solution onemergence and physiological characteristics of seed-ling in rice (Oryza sativa L.) under stress conditions.J Zhejiang Uni 28:175–178
294 Plant Growth Regul (2006) 49:285–294
123
Top Related
Copyright © 2022 FDOKUMEN

