







Bahasa
Halaman
Hukum


lable at ScienceDirect
Quaternary Science Reviews 112 (2015) 181e196
Contents lists avai
Quaternary Science Reviews
journal homepage: www.elsevier .com/locate/quascirev
Prehistoric fires and the shaping of colonial transported landscapes insouthern California: A paleoenvironmental study at Dune Pond, SantaBarbara County
Ana Ejarque a, b, *, R. Scott Anderson a, b, Alexander R. Simms c, Beau J. Gentry c
a School of Earth Sciences & Environmental Sustainability, Northern Arizona University, Flagstaff, AZ 86011, USAb Laboratory of Paleoecology, Bilby Research Center, Northern Arizona University, Flagstaff, AZ, USAc Department of Earth Science, University of California, Santa Barbara, Santa Barbara, CA 93106, USA
a r t i c l e i n f o
Article history:Received 21 July 2014Received in revised form19 November 2014Accepted 19 January 2015Available online
Keywords:Colonial transported landscapesFire historyPollenNon-pollen palynomorphsSpheroidal Carbonaceous ParticlesCalifornia
* Corresponding author. Present address: SeminaResearch (SERP), Department of Prehistory, AncienUniversity of Barcelona, Montalegre 6, 08001 Barcelo
E-mail address: [email protected] (A. Ejarque).
http://dx.doi.org/10.1016/j.quascirev.2015.01.0170277-3791/© 2015 Elsevier Ltd. All rights reserved.
a b s t r a c t
Using a novel combination of paleoecologic proxies including pollen, non-pollen palynomorphs (NPPs),macroscopic charcoal, and Spheroidal Carbonaceous Particles (SCPs), 5000 years of landscape change, firehistory and land-use have been reconstructed from Dune Pond, Santa Barbara County, California. Thepond was sensitive to Holocene regional climatic variability, showing different phases of lower (4600e3700 cal yr BP, 2100e700 cal yr BP, historical period) and higher (3700e2100 cal yr BP, 700e150 cal yr BP) local moisture availability. During this period the landscape was dominated by a coastalmosaic vegetation including dune mats, coastal scrub and salt marshes on the dunes and backdunes,with chaparral and oak woodland growing in the valley plains and foothills. Fire was intimately linkedwith such dominating mosaic vegetation, and the combination of wet conditions and the presence ofnearby human settlement were a trigger favoring coastal fires for at least two periods: from 3100 to1500 cal yr BP and from 650 cal yr BP until the 18th century. In both cases fire was an important tool tokeep an open coastal landscape attractive to hunting wildlife. Finally, matching this varied range of high-resolution paleoecological proxies with historical records we could characterize the development ofcolonial transported landscapes following the Euro-American settlement of Santa Barbara. The intro-duction of livestock grazing by Spanish colonists favored erosive processes and the introduction of fecal-borne parasites in freshwater bodies, negatively impacted salt and brackish coastal marshes, and pro-moted the invasion of alien grasses and ruderals. This agro-pastoral landscape was consolidated duringthe American period, with a greater role for cultivation, the development of industrial activities andincreased population. Despite negative environmental consequences such as the loss of native habitats,exotic land-uses and plants introduced during the historical period significantly contributed to theconfiguration of a cultural landscape which forms part of the cultural heritage of California.
© 2015 Elsevier Ltd. All rights reserved.
1. Introduction
Our knowledge about the long-term vegetation and fire historyof coastal regions of southern California has been limited by therestricted availability of preserved wetlands suitable for paleoeco-logical studies in this dry and highly developed area of the westernUS. Such widely dispersed studies such as Soledad Pond and
r of Prehistoric Study andt History and Archaeology,na, Spain.
Abalone Rocks Marsh on Santa Rosa Island (Cole and Liu, 1994;Anderson et al., 2010), San Joaquin Marsh (Davis, 1992), LosPe~nasquitos lagoon (Cole and Wahl, 2000), Mod Pond (Andersonet al., 2015) and the Santa Barbara Basin marine record (Heusser,1978), and most recently Zaca Lake (Dingemans et al., 2014) haveprovided an introduction to late Holocene paleoenvironments ofthe region. Fire history studies have been evenmore limited. In thiscase, only two high-resolution fire reconstructions are available forthe mainland southern California coast at the Santa Barbara Basin(Mensing et al., 1999) and at Los Padres National Forest (Lombardoet al., 2009), spanning the last 600 and 400 years, respectively.

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196182
Here we present a multi-proxy paleoecological study of aninterdunal swale pond located just a few kmwest of Santa Barbara,California, spanning the last 5000 years. The unusual length of theDune Pond record offers new insights into the presently poorlyknown vegetation and fire history of southwestern coastal areas ofCalifornia. Dune Pond is located within the Coal Oil Point Reserve(COPR). Established in 1970, this reserve attempts to protect one ofthe best remaining examples of a coastal-strand environment insouthern California. Together with the analysis of pollen, our studyincludes non-pollen palynomorphs (NPPs). These are micro-organisms of diverse origin, such as fungal and algal spores orzoological remains, which provide additional and independent on-site paleoenvironmental and land-use information to pollen anal-ysis (van Geel, 2001). Despite being increasingly important in-dicators in paleoenvironmental analyses, their systematic use isstill scarce, especially in North America.
The southern California coast has long been occupied by pre-historic peoples. It includes the earliest human remains thus farfound on the Pacific coast, dating back to ~13,000 cal yr BP(Erlandson et al., 2011), and over the last 20 years numerousadditional archaeological sites have been recorded in the SantaBarbara basin area (Glassow et al., 2007). However, little is knownregarding the impact of prehistoric forager communities on theirimmediate environment. Our knowledge increases considerablyduring the late 18th century historic period, as European explorersproduced written ethnographic accounts about the way of life ofNative American communities and their relationship to theirenvironment. Ethnohistoric accounts attest to the frequent andintentional use of fire by Chumash societies in the Santa BarbaraBasin area in one of the best examples of intentional burning byprehistoric hunteregatherers (Timbrook et al., 1982). Additionalexamples exist in the paleoecological record relating prehistoricfires to anthropogenic triggers in the north-central California coast(Anderson et al., 2013) as well as the northern Channel Islands(Anderson et al., 2010). However, little is known about the possibleuse of fire by prehistoric communities in the mainland coast ofsouthern California because of the lack of high-resolution and long-term charcoal analysis. Our study includes a high-resolution char-coal analysis, which contributes significantly to understanding therole of natural and/or anthropogenic fires in the Holocene shapingof Mediterranean coastal ecosystems.
One of the most significant observations of changing culturaland environmental history of coastal Californian landscapes beganwith Spanish colonization in the late 18th century. Societies place ahigh value on their local ecosystems for their economic, symbolicand cultural properties. When people move to a new territory, theyoften take basic elements of their landscapes with them, creating inthis way the ‘so called’ transported landscapes. These imply theintroduction of new land-uses, plants and land organization sys-tems, which colonists use to transform local landscapes to theirown socio-economic and cultural patterns (Anderson, 1952; Kirch,1982; Palet and Orengo, 2011). The colonization of the Alta Cali-fornia involved the development of agro-pastoral land-use systemswhich were new to this part of the New World, leading to theconfiguration of a new cultural landscape in coastal California(Anderson et al., 2013). It is estimated that ~20% of the currentCalifornia flora is composed of nonnative species introduced duringthe last 150 years (Bossard and Randall, 2007). Nonnative plantshave forever changed the landscape of California, reflected in cur-rent management policies of natural parks and reserves, whichseek to control the expansion of many introduced taxa and restoreindigenous vegetation (Rick et al., 2014). However, currentlyimportant introduced plants and land-uses are also part of thecultural landscape heritage of modern California, which includes,among other things, exotic crops such as grape vineyards and olive
orchards. Despite its significance, understanding the role of colonialtransported landscapes in shaping modern cultural landscapes hasbeen poorly addressed so far (Sluyter, 2002). The paleoecologicalstudy of Dune Pond contributes to a characterization of this processfor southern California. This is essential in order to develop land-scape management strategies that take into account both the valueof restoring native landscapes and the cultural dimension of thecolonial landscapes which developed during the historical period inthis region.
The study of colonial landscapes from a paleoecologicalperspective is hampered by the limitations of constructing reliableageedepth models for the historical period. This includes thelimited accuracy of radiocarbon dating for recent periods, andproblems related with radiometric dating such as downwash anddisplacement effects through the sediment (Oldfield et al., 1995).Here we present an innovative methodology which includes thecoupling of radiometric dating (239þ240Pu), stratigraphic changes inpollen and NPPs, historical accounts and a novel dating techniquefor the first time employed in western North America: SpheroidalCarbonaceous Particles (SCP). SCPs result from the industrialcombustion of fossil fuels and are unambiguous indicators of in-dustrial processes. Due to a characteristic morphology they can beidentified and compared with historic records of fossil-fuel com-bustion to generate reliable chronologies for the industrial period(Rose, 2001). Dune Pond is perfectly positioned for this analysis as itis not only located in one of the historically most productive areasof oil exploitation in California (Schmitt et al., 2003), but is itselfsituated adjacent to the former Elwood oil field (Fig. 1). The his-torical significance of this region as an oil production center isunderlined by the fact that the Elwood oil field became a target ofthe 1942 Japanese shelling of the US during WWII (Baker, 2003).
The main objectives of this paper are (1) to provide paleoeco-logical data regarding vegetation history and environmentalchange in Southern California for the last 5000 years; (2) to assessthe role of fires in the Holocene development of California coastalvegetation habitats and evaluate those natural and anthropogenictriggers behind prehistoric fires; and (3) to characterize thedevelopment of colonial transported landscapes in Southern Cali-fornia coastal areas, including distinct landscape transformationsrelated to the Spanish, Mexican and American settlement of SantaBarbara.
2. Archaeo-historical context
The Santa Barbara coastal area holds one of the most ancientand intense prehistoric occupations in California. During the Mid-dle Holocene, between 4500 and 3500 cal yr BC, an increase inarchaeological sites is reported, with the occupation of sites likeSBA-78, 83, 84 and 53 (Fig. 1). These were frequented by mobileforager societies who relied on the combined exploitation of ma-rine and land resources (Glassow et al., 2007). Very few archaeo-logical sites are identified between ~3000 and ~2000 cal yr BC inthe Santa Barbara Channel area, but increased settlement is docu-mented after this, and increasingly so since ~800 cal yr BC in areasneighboring Dune Pond with the occupation of SBA-81 and 71[Fig. 1, Table 1]. This increase in the number of archaeological sitesincluded significant socioeconomic and cultural changes, such asincreased sedentism, larger sites and burials, village layout, cere-monial practices or a resource diversification with a major role forfishing, and sea and land mammal hunting than before (Erlandsonand Rick, 2002). Radiocarbon dates support the occupation of manysites during the late Holocene until the contact period or shortlybefore during the 15the16th centuries (Table 1, Fig. 1). At the timeof Spanish contact the mainland Santa Barbara coast was occupiedby Chumash populations. These were hunteregatherers with a

Fig. 1. Location of the COPR with main vegetation types and photograph of Dune Pond. Archaeological sites mentioned in the text have been included in the map, together with thelocation of some historic features. The shapefile of Rancho Dos Pueblos was obtained from the Santa Barbara County Public Works, Office of the County Surveyor, CA.
A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196 183
complex socio-political and economic system which included,among other traits, sedentism, a monetized trade system based onshell beads, occupational specialization, and the processing andstorage of large quantities of food resources (Gamble, 2008). Overeighteen Chumash villages were located in the coast betweenCarpinteria and Point Conception (McLendon and Johnson, 1999:31). Archaeological digging has been performed at some of thevillages located nearby Dune Pond such as SBA-78 (Mikiw) or SBA-46 (Helo') (Table 1, Fig. 1). The largest population center in the re-gion was located around Goleta Slough (Fig. 1), where according tothe late 18th century accounts of Father Crespi, about 2000 peoplelived in large villages such as Helo' (Gamble, 2008: 86e87).
Spanish exploration of Alta California began with Juan Rodrí-guez Cabrillo's expedition in 1542 followed by the several othersduring the 17th and 18th centuries. Permanent settlement did notbegan until 1769, when Spanish authorities ordered the occupationof the northwestern-most region of New Spain to secure the king'sdomains from Russian advances along the Pacific coast (Geiger,1965). The occupation of the Alta California was performedexpanding north the network of Missions established in the LowerCalifornia one century ago, together with the founding of Presidios(forts) and Pueblos (towns). The founding of the Presidio in 1782and of the Santa Barbara Mission in 1786 documents the earliestSpanish settling of the study area (Fig. 1). Cultivation and grazingwere carried out in the Mission as well as in outlying ranches andIndian rancherias extending along the coast from the Gaviota areaon the west to the Rinc�on on the east (Geiger, 1965). A combinationof Eurasian and Mesoamerican crops such as grapes, olives, wheat,barley, chickpeas, oranges, beans or maize were introduced andgrown in the Mission lands (Hardwick, 2005). After gaining inde-pendence from Spain in 1821, M�exico ruled over Alta California and,since 1833, Mission lands were secularized and transformed intoprivate ranchos. Our study area was included within Rancho Dos
Pueblos (Fig. 1), whose title was granted in 1842 to Nicholas A. Denand latter honored by the U.S. government (Tompkins, 1960). TheAmerican expansion of Santa Barbara in the mid-19th century ledto additional major socio-economic changes in this region, such asincreases in population and urban development, with both thedevelopment of the oil industry and an intensive agricultural socio-economic system.
3. Study site
Dune Pond (5m a.s.l.) occupies a small basin (<0.5 km2) within afield of secondary dune ridges and low-lying swales in the westernpart of the Coal Oil Point Reserve (COPR), at 34�240 4900N, 119� 520
5500 W. The COPR lies along the southern coastal strip of the Goletavalley, ~11 km west of the city of Santa Barbara in Santa BarbaraCounty (Fig. 1). The Goleta valley is a flat alluvial plain bordered onthe south by the Pacific Ocean and on the north by the eastewesttrending Santa Ynez Mountains. Dune Pond is a fresh waterwetland but can be exposed to occasional salt spray. The pondformed after the damming of a small ephemeral drainage by a dunefield on the edge of a larger estuary, Devereux Slough (Fig. 1).Devereux Slough and Dune Pond occupy a flooded stream valleythat presumably formed due to sea-level fall during the last glacialmaximum (Minor et al., 2009). The flooded valley is incised into an~20e25 m high marine isotope stage 3 (MIS3) marine terrace datedto ~45 ka by a combination of UeTh dates from isolated corals,optically stimulated luminescence dates from buried sands, andnear-infinite 14C ages (Gurrola et al., 2014). TheMIS3marine terracedeposits are composed of 1e3 m of marine sands overlying foldedpetroleum-bearing Miocene siltstones and shales of the underlyingMiocene Monterey and Sisquioc Formations (Minor et al., 2009).The marine sands are discontinuously overlain by late Quaternary/

Table 1Occupation periods of contemporaneous and nearby sites to Dune Pond. Periods ofoccupation have been established on the base of published calibrated radiocarbondates (Harrison, 1965; Erlandson and Rick, 2002) and archaeological material(Gamble, 2008) obtained at these sites.
Sitecode
Occupation period (age cal BC/AD) Reference
SBA-81 BC 790e520/BC 730e410 Erlandson and Rick 2002SBA-71 BC 90e130 AD/AD 270e680 Erlandson and Rick 2002SBA-72 AD 1e160/AD 390e720/AD 830e1040/AD
1170e1450Erlandson and Rick 2002
SBA-1731
AD 400e570/AD 1050e1650 Erlandson and Rick 2002
SBA-2149
AD 1480e1640 Erlandson and Rick 2002
SBA-78 AD 1710e1780 Harrison, 1965 inGamble, 2008
SBA-46 AD 1770e1803 Gamble 2008
A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196184
Holocene eolian dune sands and interdune marshy sediments(Minor et al., 2009).
COPR experiences a maritime Mediterranean climate charac-teristic of coastal California, with moist winters, dry summers andoverall mild temperatures. Long-term climatic series (72 years)from the nearby Santa Barbara airport climate station show littletemperature fluctuations from month to month, with maximummonthly average temperatures of 17.7 �C in January to 23.8 �C inAugust. Average monthly precipitations vary from a high of98.8 mm in January to near absence during July (Western RegionalClimate Center, http://www.wrcc.dri.edu, accessed November2013).
Scirpus sp. (bulrush), Juncus phaeocephalus (brown-headedrush) and Juncus mexicanus (Mexican rush) dominate the fresh-water marsh, but other species include Distichlis spicata (saltgrass), Salix lasiolepis (arroyo willow), Sambucus mexicana (elder-berry) and members of the Fabaceae (Lotus purshianus, Lupinus sp.)and Polygonaceae (Rumex sp.) families (Plants of Coal Oil PointReserve, Fig. 1). Salicornia pacifica (pickleweed) is dominant onDevereux Slough and scattered around Dune Pond and duneswales, while Lythrum californicum (California loose-strife),Xanthium strumarium (cocklebur) and Spergularia marina (sand-spurrey) grow in fresh-water wetlands and marshes. Groves ofEucalyptus sp. (gum tree) and Cupressus macrocarpa (Montereycypress) also grow within 50 and 150 m to the SE of the pond.Common perennial plants growing on dunes are Ambrosia cha-missonis (bear burr), Chenopodium californicum (soap plant), Abro-nia umbellata (pink sand verbena) and Cirsium rhothophilum (surfthistle), while species more tolerant of soil salinity, such as Atriplexleucophylla (beach saltbush), Cakile maritima (sea rocket) or Abroniamaritima (red sand verbena) are found in foredunes. Located southand west of Dune Pond are small areas of coastal bluff scrub, whichinclude species tolerant to constant salt sprays such as Suaedataxifolia (wooly sea-blite) or Isocoma menziesii (coast goldenbush).Back dunes are covered by grasslands, mainly exotic annuals (e.g.,Bromus sp., Phalaris aquatica), and dune scrub. The latter is domi-nated by Baccharis pilularis (coyote brush) and mixed with otherplants such as Eriogonum parvifolium (wild buckwheat), Scrophu-laria californica floribunda (California figwort), Verbena lasiostachys(verbena) or Phacelia ramosissima austrolitoralis (phacelia). Plantterminology follows the CALflora database and Barbour et al.(2007).
4. Material and methods
4.1. Coring
Three ~4 m long 7.6 cm diameter vibracores where taken fromDune Pond. The longest and best preserved core, DP12-1, waschosen for detailed analysis. The other two cores contained thesame stratigraphy as DP12-1. Cores were split and photographed inthe sedimentology laboratory at the University of California, SantaBarbara. The core was described according to sedimentary texture,grain-size, and the presence/absence of macrofossils and sedi-mentary structures.
4.2. Pollen and NPPs
82 samples were prepared for pollen and NPP analysis followinga modified standard procedure (Faegri and Iversen, 1989). We used2 cm3 of sediment except for the uppermost meter where micro-fossil concentration was lower and 6 cm3 were used. Samples weresuspended in Na4P207 and sieved, then treated with HCL, HF, ace-tolysis, density separation in ZnBr2 (specific gravity 1.9) andmounted in glycerine jelly. Pollen and NPPs were identified on a
light microscope at 400� and 1000� magnification. Pollen identi-fication followed published illustrations and morphological keys(Punt et al., 1976e2009; Reille, 1992) and the LOP-NAU pollenreference collection. Asteraceace family was divided into 6 pollensubgroups based on the morphological description of Andersonet al. (2010), and Cerealia-type grains were identified followingmorphological criteria of Faegri and Iversen (1989). At least 700terrestrial pollen grains were counted per sample. Pollen values areexpressed as a percentages of the total terrestrial pollen (TTP) sum,which excludes Cyperaceae, fern spores, aquatic plants and NPPs.Lycopodium clavatum spore tablets (Stockmarr, 1971) were added inorder to calculate pollen concentration. NPPs were counted aspollen was tallied and identification followed published de-scriptions and illustrations van (e.g. van Geel, 2001; van Geel andAptroot, 2006). NPP types were assigned to an existing codewhen already described in the catalogue of the Hugo de VriesLaboratory (HdV), University of Amsterdam (Netherlands). Un-known NPPs types were numbered using the code LOP (Laboratoryof Paleoecology, Northern Arizona University). 37 NPP morpho-types were identified and selected pictures are shown in Fig. 2. NPPvalues are calculated as percentages relative to the TTP sum, whichacts as an independent parameter to normalize NPP taxa. Pollenand NPPs stratigraphic diagrams were graphed using C2 software v.1.7.2 (Juggins, 2011). Pollen zones were established using theCONstrained Incremental Sum-of-Square cluster analysis (CONISS;Grimm, 1987).
4.3. Charcoal analysis
High-resolution charcoal analysis was performed on contiguous1-cc sediment samples following the method of Whitlock andAnderson (2003). Macroscopic charcoal particles �125 mm andmacroremains encountered were tallied using a binocular micro-scope at 10e40� magnification. We followed the decompositionapproach to identify local fire events from regional fire activity(Gavin et al., 2006). Statistical treatment of charcoal data was per-formed using CharAnalysis v.1.1 software (https://code.google.com/p/charanalysis; Higuera et al., 2009). Charcoal accumulation rates(CHAR; # cm�2 yr�1) were calculated from charcoal concentrationand sediment accumulation rates. CHARs were resampled toequally-spaced intervals approximating the mean sampling reso-lution of the record (10 years) and log-transformed to minimizevariance. CHARs were then decomposed into (1) background CHAR(Cback), a low-frequency component resulting from secondarycharcoal deposition, distant/regional fires and/or changes in char-coal production; and (2) the peaks component (Cpeak), a high-frequency and high-magnitude component representing
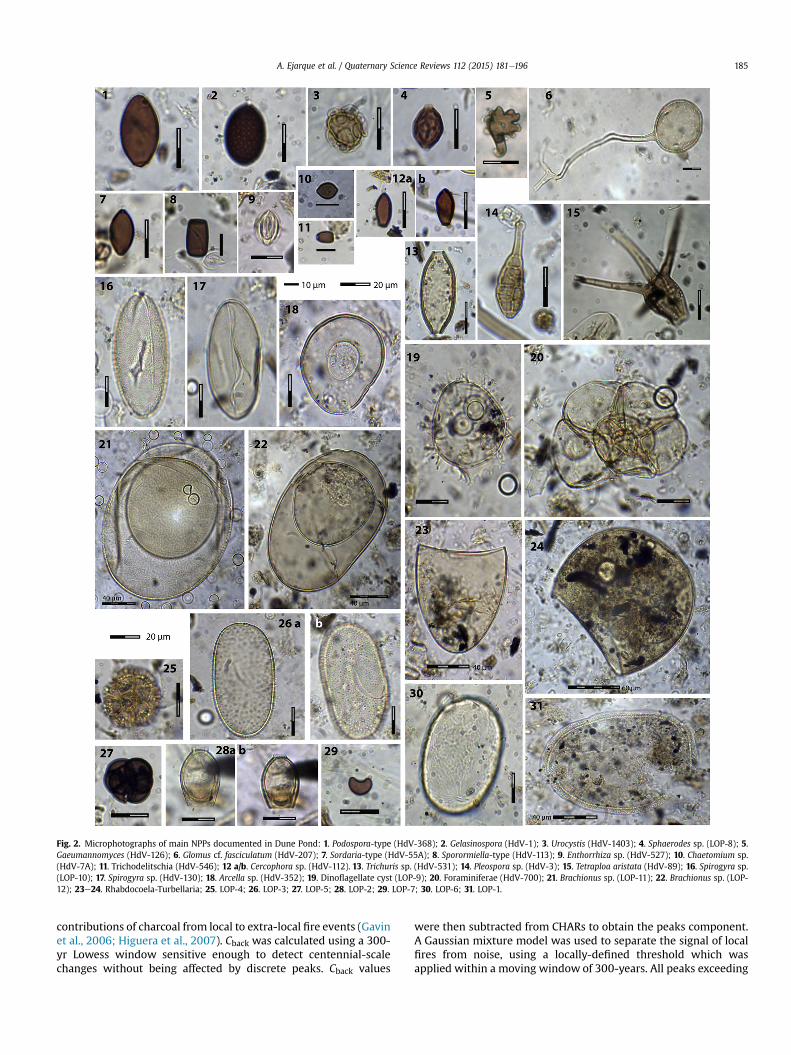
Fig. 2. Microphotographs of main NPPs documented in Dune Pond: 1. Podospora-type (HdV-368); 2. Gelasinospora (HdV-1); 3. Urocystis (HdV-1403); 4. Sphaerodes sp. (LOP-8); 5.Gaeumannomyces (HdV-126); 6. Glomus cf. fasciculatum (HdV-207); 7. Sordaria-type (HdV-55A); 8. Sporormiella-type (HdV-113); 9. Enthorrhiza sp. (HdV-527); 10. Chaetomium sp.(HdV-7A); 11. Trichodelitschia (HdV-546); 12 a/b. Cercophora sp. (HdV-112). 13. Trichuris sp. (HdV-531); 14. Pleospora sp. (HdV-3); 15. Tetraploa aristata (HdV-89); 16. Spirogyra sp.(LOP-10); 17. Spirogyra sp. (HdV-130); 18. Arcella sp. (HdV-352); 19. Dinoflagellate cyst (LOP-9); 20. Foraminiferae (HdV-700); 21. Brachionus sp. (LOP-11); 22. Brachionus sp. (LOP-12); 23e24. Rhabdocoela-Turbellaria; 25. LOP-4; 26. LOP-3; 27. LOP-5; 28. LOP-2; 29. LOP-7; 30. LOP-6; 31. LOP-1.
A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196 185
contributions of charcoal from local to extra-local fire events (Gavinet al., 2006; Higuera et al., 2007). Cback was calculated using a 300-yr Lowess window sensitive enough to detect centennial-scalechanges without being affected by discrete peaks. Cback values
were then subtracted from CHARs to obtain the peaks component.A Gaussian mixture model was used to separate the signal of localfires from noise, using a locally-defined threshold which wasapplied within a moving window of 300-years. All peaks exceeding
A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196186
the locally-defined threshold were then subjected to a minimum-count test at the 95% confidence level to remove peaks resultingfrom statistically insignificant changes in charcoal abundance(Higuera et al., 2010). Local fire event frequency (FEF) was calcu-lated on the base of number of local fires (peaks) per 300 years.
4.4. SCP analysis
23 samples in the upper 90 cm of the record were processed forSCP concentration using 2 cm3 of sediment. A modified Rose (1990)procedure was used in which samples were boiled in HNO3 toeliminate organic matter, then treated with HCL, HF and sievedusing a 5-mm filter. SCP were identified and tallied using a lightmicroscope at 400� magnification. SCPs �25-mm were countedseparately in order to account for a more local signal (Inoue et al.,2013). Lycopodium spore tablets were added in order to calculateSCP concentration. SCP results have been calibrated with thefollowing long-term instrumental series: (1) US Co2 emissions offossil-fuel burning (110 years, source: Boden et al., 2013); (2) U.S.Field Production of Crude Oil (153 years, source: U.S. Energy In-formation Administration, http://www.eia.gov, accessedSeptember 2013); and (3) the Field Production of Crude Oil of theCoastal Region of California (135 years), including the coastalcounties of Santa Barbara, Ventura and San Luis Obispo (multiplesources: Nehring (1975); Schmitt et al. (2003); U.S. Energy Infor-mation Administration).
4.5. Ageedepth model
The age model for Dune Pond was built using different meth-odologies. For the older part of the record 3 14C AMS dates wereobtained from Accium Biosciences on macrocharcoal samples.Dates were calibrated with CLAM v. 2.2 (Blaauw, 2010) using theIntCal13 14C calibration curve (Reimer et al., 2013). For the youngestportion of the record we used radiometric dating (239þ240Pu),
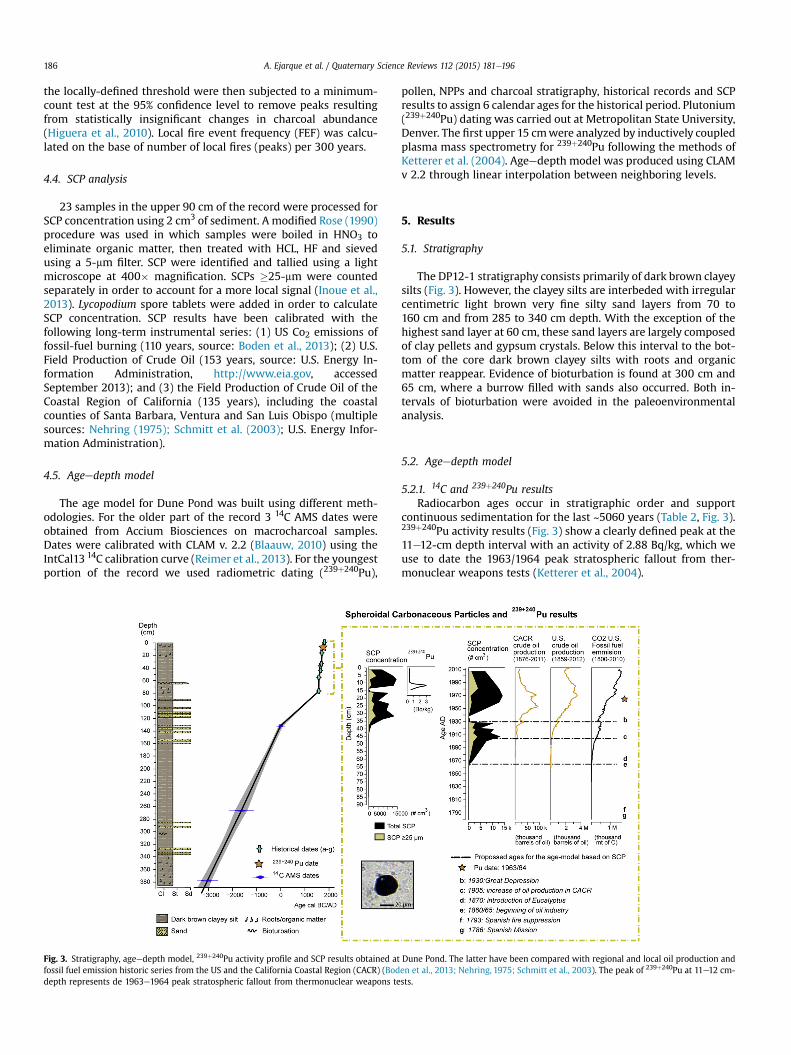
Fig. 3. Stratigraphy, ageedepth model, 239þ240Pu activity profile and SCP results obtained atfossil fuel emission historic series from the US and the California Coastal Region (CACR) (Boddepth represents de 1963e1964 peak stratospheric fallout from thermonuclear weapons te
pollen, NPPs and charcoal stratigraphy, historical records and SCPresults to assign 6 calendar ages for the historical period. Plutonium(239þ240Pu) dating was carried out at Metropolitan State University,Denver. The first upper 15 cmwere analyzed by inductively coupledplasma mass spectrometry for 239þ240Pu following the methods ofKetterer et al. (2004). Ageedepth model was produced using CLAMv 2.2 through linear interpolation between neighboring levels.
5. Results
5.1. Stratigraphy
The DP12-1 stratigraphy consists primarily of dark brown clayeysilts (Fig. 3). However, the clayey silts are interbeded with irregularcentimetric light brown very fine silty sand layers from 70 to160 cm and from 285 to 340 cm depth. With the exception of thehighest sand layer at 60 cm, these sand layers are largely composedof clay pellets and gypsum crystals. Below this interval to the bot-tom of the core dark brown clayey silts with roots and organicmatter reappear. Evidence of bioturbation is found at 300 cm and65 cm, where a burrow filled with sands also occurred. Both in-tervals of bioturbation were avoided in the paleoenvironmentalanalysis.
5.2. Ageedepth model
5.2.1. 14C and 239þ240Pu resultsRadiocarbon ages occur in stratigraphic order and support
continuous sedimentation for the last ~5060 years (Table 2, Fig. 3).239þ240Pu activity results (Fig. 3) show a clearly defined peak at the11e12-cm depth interval with an activity of 2.88 Bq/kg, which weuse to date the 1963/1964 peak stratospheric fallout from ther-monuclear weapons tests (Ketterer et al., 2004).
Dune Pond. The latter have been compared with regional and local oil production anden et al., 2013; Nehring, 1975; Schmitt et al., 2003). The peak of 239þ240Pu at 11e12 cm-sts.

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196 187
5.2.2. Historical pollen, NPPs and charcoal datesThe lowermost historical date “g” at 77 cm (Fig. 3) was assigned
to the establishment of the Spanish Mission of Santa Barbara in1786. This was determined on the basis of the synchronousoccurrence of dung-related fungal spores indicative of grazing (i.e.Sporormiella-type, Sordaria-type) and of pollen indicators of land-disturbance likely to represent cultural and introduced taxa (i.e.Rumex, Erodium, Plantago, Cerealia-type). Several Mission ranchesand rancherias holding farming activities, such as San Miguel orTecolote (O'Keefe, 1886: 17), were located nearby Dune Pond atGoleta Slough and Tecolote canyon (Fig. 1). Date “f” at 55 cm cor-responds to the drop of CHAR in Dune Pond, which we relate to the1793 fire suppression edict of Governor Jos�e Joaquin de Arrillagabanning ‘all kind of burning’ by the Indians in the Alta Californiaunder Spanish jurisdiction (Car, 1959). Finally we have assignedanother historical date “d” to the first occurrences of Eucalyptuspollen in the record at 36 cm. We relate that to 1870, when the firstgum trees in Santa Barbara were introduced by Ellwood Cooper,who planted about 50,000 Eucalyptus on his property in Goleta(Esau, 2006: 254e255), a few kilometers north of Dune Pond.
5.2.3. SCP results and datesThe comparison of SCP results in Dune Pond with historical
data-sets of both the local oil industry in California and the US CO2fossil fuel emissions have furnished a robust age-model for the late19th and early 20th centuries (Fig. 3). SCP results obtained at DunePond show abundant concentration values of fly-ash particles inthe uppermost 44 cm of the record varying from ~120 SCP cm3 to~14,500 SCP cm3. The SCP trend at Dune Pond is characterized by(1) the beginning of the recording of SCP at 44 cm; (2) a first andconstant increase in SCP concentration from 43 cmwhich includesthe SCP large fraction�25-mm; (3) the decline and disappearance offly-ash particles from 16 to 14 cm; and (4) a final recovery from12 cm arriving to maximum concentrations of SCP in the record.
We attribute the first occurrence of SCP in the profile at 44 cm(date “e”) to the beginning of the oil-industry in Ventura County,California at about 1860e65 (Nehring, 1975; Schmitt et al., 2003;Fig. 3). The first occurrences of Eucalyptus pollen in the record a fewcentimeters above, dated at 1870 (“d”), supports the accuracy ofthis age. The increase of SCP concentration recorded from 43 cmwas assigned to the beginning of oil-exploitation in Santa BarbaraCounty at the beginning of the 20th century. The crude oil pro-duction accelerated in the California Coastal Region (CACR) since1905 with the discovery of very productive oil wells in the Santa
Table 2Radiocarbon dates results and historic dating established in Dune Pond through pollen,
Designated date event (aeg)/14Claboratory code
Depth(cm)
Age historic, age14C yr BP
Historic event
a 0 2012 e
b 16 1930 Great Depressioc 34 1905 Beginning oil pr
Santa Barbarad 36 1870 Eucalyptus introe 44 1865 Beginning oil pr
Californiaf 55 1793 Spanish fire sup
g 77 1786 Spanish Mission
UCIAMS11562 132e134
1895 ± 30 e
DAMS001750 268e269
3294 ± 99 e
DAMS001751 380e381
4388 ± 75 e
Maria Valley (Schmitt et al., 2003), ~65 km NW of Dune Pond, andwe have assigned this date to this initial rise of SCP (point “c”). Thisincrease includes large SCP �25-mm. Studies on current SCPdeposition in industrial areas show that a relationship exists be-tween the size of SCPs and their spatial distribution from theemission source (Larsen, 2003). Inoue et al. (2013) stressed thatconcentrations of SCP �20-mm representing more than 40% of thetotal SCP are locally generated within a few kilometers. Eventhough SCP �25-mm at Dune Pond do not reach those percentages,their increase up to 26% of the total SCP counts may well indicate acloser source related to the beginning and rise of the oil industry inthe CACR at that time (Fig. 3). Our last dating point “b” correspondsto the sharp drop of SCP at 16 cm. We attribute this event to thedecrease in oil production documented in the US during the eco-nomic stagnation of the Great Depression in 1930. Decreasing in-dustrial activity is evident in both the US fossil fuel emission andthe overall crude oil production of the US at this time including asharp decline in oil production in the CACR. The subsequent 1963/1964 Pu date at 11e12-cm depth supports this interpretation(Fig. 3).
5.2.4. Sedimentation ratesSedimentation rates for the lower ~3.0 m of the core are defined
by 3 radiocarbon dates. The ageedepth model shows a low sedi-ment accumulation rate (SAR) for the lower ~3.0 m of the core, withSARs below 80 cm-depth ranging from ~0.07 to ~0.03 cm/yr(~13e30 yr/cm). Constant sedimentation rates of ~0.07 cm/yr be-tween the three radiocarbon dates performed on the lower part ofthe record support the reliability of the dates obtained on macro-charcoal particles. A lower SAR is suggested for the ca 134e80 cmdepth core section. Lowered sediment accumulation during a drierperiod is supported not only by changes in pollen and spores, butalso the occurrence of gypsum sand layers (see section 7.1 below).Above 80 cm-depth SAR increases significantly to mean values of~0.9 cm/yr (~6 yr/cm). The change towards rapid sedimentation at80 cm most probably results from increasing erosion followingEuro-American settlement and the introduction of grazing andfarming activities in the area. Similar abrupt SAR increases at thebeginning of the historic period are frequently reported at othercoastal ponds and salt marshes studied in California (Davis, 1992;Cole and Wahl, 2000; Anderson et al., 2010, 2013). Increasing SARallowed for high-resolution pollen and NPP analyses for the his-torical period, with sampling intervals ranging from 1 to 35 years.
NPP stratigraphy and SCP results.
Calibrated age BC/AD (2s)
Mean agevalues
Date type Materialdated
e e Surface e
n e e SCP/historic e
oduction in e e SCP/historic e
duction e e Pollen/historic e
oduction in e e SCP/historic e
pression e e Charcoal/historic
e
e e Pollen, NPPs/historic
e
53e213 AD AD 133 AMS Charcoal
1876e1325 BC BC 1600 AMS Charcoal
3335e2892 BC BC 3113 AMS Charcoal
A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196188
6. Results
6.1. Pollen, NPPs and macrocharcoal results
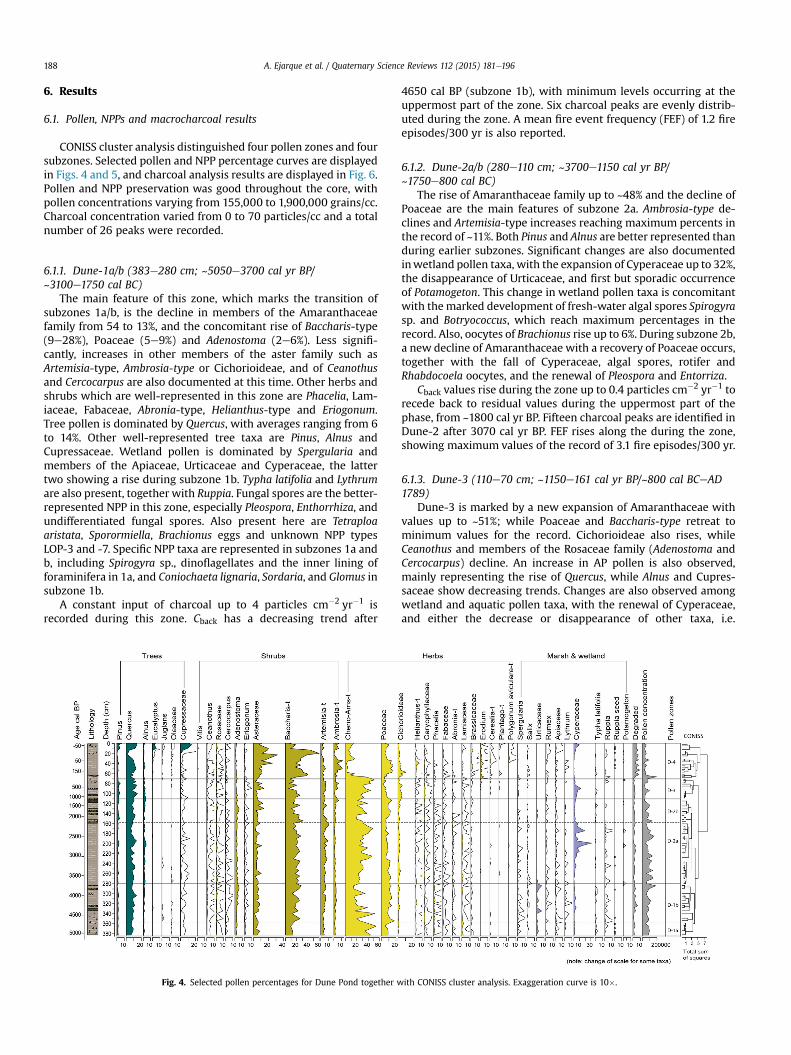
CONISS cluster analysis distinguished four pollen zones and foursubzones. Selected pollen and NPP percentage curves are displayedin Figs. 4 and 5, and charcoal analysis results are displayed in Fig. 6.Pollen and NPP preservation was good throughout the core, withpollen concentrations varying from 155,000 to 1,900,000 grains/cc.Charcoal concentration varied from 0 to 70 particles/cc and a totalnumber of 26 peaks were recorded.
6.1.1. Dune-1a/b (383e280 cm; ~5050e3700 cal yr BP/~3100e1750 cal BC)
The main feature of this zone, which marks the transition ofsubzones 1a/b, is the decline in members of the Amaranthaceaefamily from 54 to 13%, and the concomitant rise of Baccharis-type(9e28%), Poaceae (5e9%) and Adenostoma (2e6%). Less signifi-cantly, increases in other members of the aster family such asArtemisia-type, Ambrosia-type or Cichorioideae, and of Ceanothusand Cercocarpus are also documented at this time. Other herbs andshrubs which are well-represented in this zone are Phacelia, Lam-iaceae, Fabaceae, Abronia-type, Helianthus-type and Eriogonum.Tree pollen is dominated by Quercus, with averages ranging from 6to 14%. Other well-represented tree taxa are Pinus, Alnus andCupressaceae. Wetland pollen is dominated by Spergularia andmembers of the Apiaceae, Urticaceae and Cyperaceae, the lattertwo showing a rise during subzone 1b. Typha latifolia and Lythrumare also present, together with Ruppia. Fungal spores are the better-represented NPP in this zone, especially Pleospora, Enthorrhiza, andundifferentiated fungal spores. Also present here are Tetraploaaristata, Sporormiella, Brachionus eggs and unknown NPP typesLOP-3 and -7. Specific NPP taxa are represented in subzones 1a andb, including Spirogyra sp., dinoflagellates and the inner lining offoraminifera in 1a, and Coniochaeta lignaria, Sordaria, and Glomus insubzone 1b.
A constant input of charcoal up to 4 particles cm�2 yr�1 isrecorded during this zone. Cback has a decreasing trend after
Fig. 4. Selected pollen percentages for Dune Pond together w
4650 cal BP (subzone 1b), with minimum levels occurring at theuppermost part of the zone. Six charcoal peaks are evenly distrib-uted during the zone. A mean fire event frequency (FEF) of 1.2 fireepisodes/300 yr is also reported.
6.1.2. Dune-2a/b (280e110 cm; ~3700e1150 cal yr BP/~1750e800 cal BC)
The rise of Amaranthaceae family up to ~48% and the decline ofPoaceae are the main features of subzone 2a. Ambrosia-type de-clines and Artemisia-type increases reaching maximum percents inthe record of ~11%. Both Pinus and Alnus are better represented thanduring earlier subzones. Significant changes are also documentedinwetland pollen taxa, with the expansion of Cyperaceae up to 32%,the disappearance of Urticaceae, and first but sporadic occurrenceof Potamogeton. This change in wetland pollen taxa is concomitantwith themarked development of fresh-water algal spores Spirogyrasp. and Botryococcus, which reach maximum percentages in therecord. Also, oocytes of Brachionus rise up to 6%. During subzone 2b,a new decline of Amaranthaceae with a recovery of Poaceae occurs,together with the fall of Cyperaceae, algal spores, rotifer andRhabdocoela oocytes, and the renewal of Pleospora and Entorriza.
Cback values rise during the zone up to 0.4 particles cm�2 yr�1 torecede back to residual values during the uppermost part of thephase, from ~1800 cal yr BP. Fifteen charcoal peaks are identified inDune-2 after 3070 cal yr BP. FEF rises along the during the zone,showing maximum values of the record of 3.1 fire episodes/300 yr.
6.1.3. Dune-3 (110e70 cm; ~1150e161 cal yr BP/~800 cal BCeAD1789)
Dune-3 is marked by a new expansion of Amaranthaceae withvalues up to ~51%; while Poaceae and Baccharis-type retreat tominimum values for the record. Cichorioideae also rises, whileCeanothus and members of the Rosaceae family (Adenostoma andCercocarpus) decline. An increase in AP pollen is also observed,mainly representing the rise of Quercus, while Alnus and Cupres-saceae show decreasing trends. Changes are also observed amongwetland and aquatic pollen taxa, with the renewal of Cyperaceae,and either the decrease or disappearance of other taxa, i.e.
ith CONISS cluster analysis. Exaggeration curve is 10�.
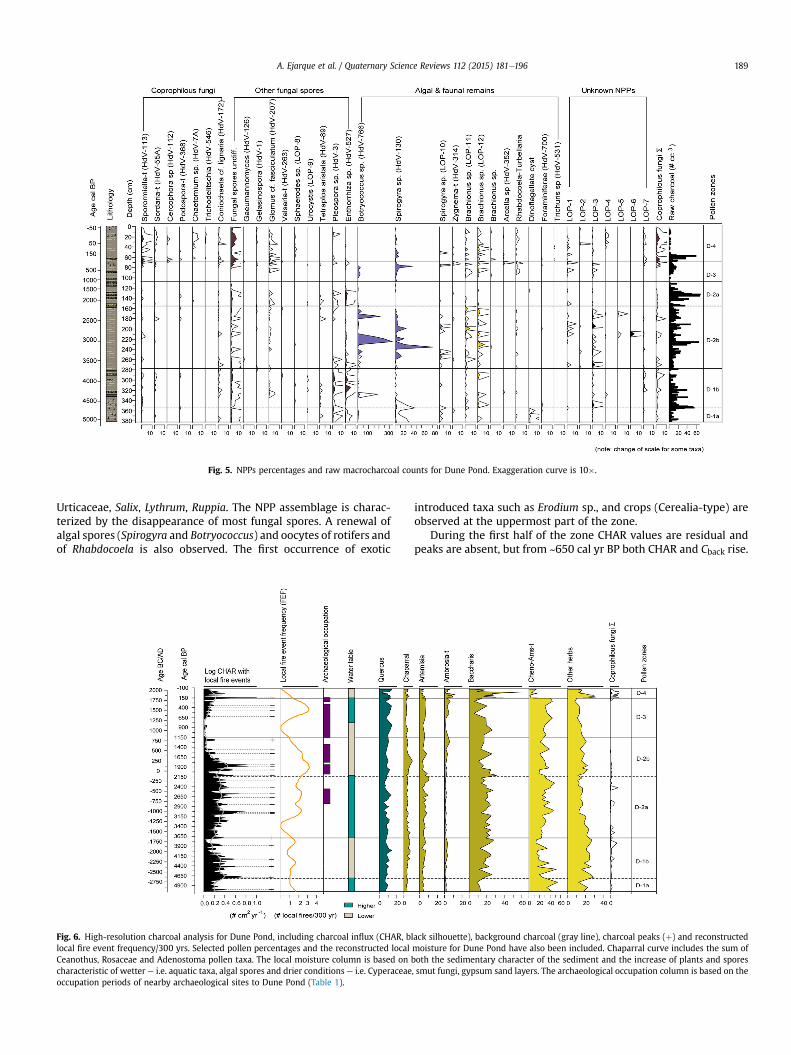
Fig. 5. NPPs percentages and raw macrocharcoal counts for Dune Pond. Exaggeration curve is 10�.
A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196 189
Urticaceae, Salix, Lythrum, Ruppia. The NPP assemblage is charac-terized by the disappearance of most fungal spores. A renewal ofalgal spores (Spirogyra and Botryococcus) and oocytes of rotifers andof Rhabdocoela is also observed. The first occurrence of exotic
Fig. 6. High-resolution charcoal analysis for Dune Pond, including charcoal influx (CHAR, bllocal fire event frequency/300 yrs. Selected pollen percentages and the reconstructed localCeanothus, Rosaceae and Adenostoma pollen taxa. The local moisture column is based on bcharacteristic of wetter e i.e. aquatic taxa, algal spores and drier conditions e i.e. Cyperaceaeoccupation periods of nearby archaeological sites to Dune Pond (Table 1).
introduced taxa such as Erodium sp., and crops (Cerealia-type) areobserved at the uppermost part of the zone.
During the first half of the zone CHAR values are residual andpeaks are absent, but from ~650 cal yr BP both CHAR and Cback rise.
ack silhouette), background charcoal (gray line), charcoal peaks (þ) and reconstructedmoisture for Dune Pond have also been included. Chaparral curve includes the sum ofoth the sedimentary character of the sediment and the increase of plants and spores, smut fungi, gypsum sand layers. The archaeological occupation column is based on the

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196190
Four peaks are reported after ~650 cal yr BP and FEF reaches similarlevels as the end of zone Dune 2b.
6.1.4. Dune-4 (70e0 cm; 1789 to present)Zone Dune-4 attests to the marked decline of members of the
Amaranthaceae family (51%e2%), which only recover in the upperpart of the zone, and the expansion of Baccharis-type, which risesup to 60%. Poaceae also expands during the first half of the phase,together with either the appearance or rise of species consideredintroduced or exotic e i.e. Erodium, Brassicaceae, Helianthus-type,Rumex, Plantago e and fruit trees or crops such as Juglans, Cerealia-type, Oleaceae or Vitis. The latter mainly rise during the second halfof the zone, when new exotic species are documented for the firsttime, such as Eucalyptus and Polygonum aviculare-type. AP showsan oscillating trend during the zone, with a strong decline ofQuercus down to minimum rates in the record of 2% and the ulti-mate recovery of Quercus, Cupressaceae and Pinus during the latterpart of the zone. This zone documents the rise and reappearance ofa varied range of dung-related fungi such as Sordaria, Coniochaetalignaria, Cercophora and most remarkably, Sporormiella, whichreaches maximum rates in the record of 3%. Maximum values ofundifferentiated fungal spores, continuous recording of Glomus,rotifer eggs and the first notations of eggworms Trichuris sp. arealso observed in the zone. The disappearance of the algal spores,Spirogyra and Zygnema-type also occurs.
A single large charcoal peak (14 particles cm�2 peak�1) occurs at~1890 AD. In addition, Dune-4 is marked by low levels of Cback of0.1e0.2 particles cm�2 yr�1 and the general absence of charcoalpeaks.
6.2. Reconstruction of fires
The results of both simulation models (Higuera et al., 2007) andempirical evidence (Gavin et al., 2003; Higuera et al., 2005) ofcharcoal production, transport and deposition in natural basins arecritical for interpretation of paleofire records. These results suggestthat for small basins like Dune Pond, identified charcoal peaksrepresent local fires that have occurred within an area <1 kmradius, while Cback values are indicative of more distant, perhapsregional fires. Comparison of the historical record of nearby firesand our recent sedimentary charcoal record is consistent with thismodel. Evidence of historic fires occurred on the coast and theSanta Inez Mountains at a distance of ~2e10 km from Dune Pond,but not directly at the pond (111-yr record, GIS source: CALFIRE,Department of Forestry and Fire Protection of California). Thelargest and closest fires are 1) the AD 1979 Eagle Canyon fire,~1500 ha burned ~2 km NWof Dune Pond; 2) the AD 1955 Refugioand AD 2008 fires, ~33.000 and ~4000 ha respectively burned~5 kmNof the pond; and 3) the AD 1990 Painted Cave fire, ~1800 haburned ~9 km E to the pond. Our reconstructed fire-history for thelast 120 years, shows only the presence of low background valuesand no peaks, and is consistent with the recent history of regionalfires (>2e10 km) documented in the area. Although CHAR valuesshow three increases higher than Cback since AD 1912, one of whichat AD 1952 coincides with the Refugio fire, these increases were notsignificant enough to be detected as peaks in our statistical analysis.This supports the accuracy of our peak signal for reconstructinglocal fires within 1e2 km from the pond, and of Cback representingmore regional fires. Finally, the position of the pond adjacent to thePacific Ocean, with predominant winds from the west, makes thesite more suitable for reflecting local coastal fires. Indeed, althoughexposed to occasional northern down-slope Sundowner events, theSanta Barbara region is sheltered from extreme Santa Ana events,long-distance winds blowing westward to the southern California
coast that could favor the transport of charcoal particles from extra-regional fires to the pond (Moritz, 2003).
7. Discussion
7.1. Coastal vegetation and environmental change during theMiddle and late Holocene
During the Middle Holocene, between 5000 and 4600 cal yr BP[Dune-1a], Dune Pond developed as a shallow freshwater tobrackish pond exposed to saltwater influence. This is indicated bythe co-occurrence of Zygnemataceae algae spores (Spirogyra-typeand Zygnema-type), inhabiting freshwater shallow wetlands (vanGeel, 2001), and marine-brackish organisms such as foraminifera(their testate linings) and dinoflagellates. The highest ratios ofAmaranthaceae at this time indicate the predominance of salt andbrackish marsh vegetation growing around the pond and in thenearby salt flats, coastal bluffs and foredunes. These were probablydominated by Salicornia pacifica and other halophytic and salt-tolerant plants included within the Amaranthaceae, Spergularia,Ruppia and Abronia pollen taxa (Munz, 1974; Macdonald, 1988).Freshwater patches of Cyperaceae (sedges), Typha latifolia (cattails)and Urticaceae (nettles) probably grew around the basin. EnhancedEl Ni~no/Southern Oscillation (ENSO) activity documented betweenca 5000 and 4650 cal yr BP (Moy et al., 2002) could have led tohigher water tables here, as they apparently did in the SierraNevada to the east (Anderson and Smith, 1994) and elsewhere. Itwould also have favored storm-generated washover and thus salt-exposure in Dune Pond.
Between 4600 and 3700 cal yr BP [Dune-1b], the disappearanceof algae spores, the increase of grasslands (Poaceae), marshes,wetland vegetation e i.e. Cyperaceae, Salix, Urticaceae, Lythrum,Rumex e and either the increase or more regular recording ofspores of saprophytic fungi growing on decaying herbaceous re-mains and sedges such as Pleospora sp., Tetraploa aristata orEnthorrhiza sp. (Ellis, 1971; V�anky, 1994; Kodsueb et al., 2006)suggest lower water tables and increased organic matter decom-position on the pond's moister soils. Sand layers of gypsumdeposited during this time in the record probably reflect drierconditions more conducive to evaporate formation. Lower watertables or decreased moisture conditions documented at Dune Pondare concomitant with a period of general arid conditions inSouthern California and warm sea temperatures recorded in theSanta Barbara basin during the Middle Holocene (Kennett et al.,2007; Barron and Anderson, 2011; Kirby et al., 2007). Also, thedisappearance of marine organisms and the decline of Amar-anthaceae pollen indicate less saline influence. For the rest of theHolocene, freshwater conditions prevailed at Dune Pond as sug-gested by the sparse occurrence of foraminifera and dinoflagellatesindicating only occasional exposure to salt sprays. Less saline con-ditions allowed the expansion of grasses (Poaceae) and coastalscrub dominated by B. pilularis in dunes and backdunes adjacent toDune Pond from ~4600 cal yr BP through the late Holocene.
Unlike the modern vegetation composition at the COPR, saltmarshes and coastal bluff scrub communities dominated bymembers of the Amaranthaceae would continue to be an increas-ingly significant part of this coastal open environment during thelate Holocene, as shown by the rise of this family from~3700 cal yr BP. Likewise, Artemisia-type was an importantcomponent of the coastal scrub and of dune mats, together withother members of the Asteraceae family like Ambrosia-type orHelianthus-type. More regionally, oak woodlands and chaparral(e.g. Adenostoma, Ceanothus) increased their presence dominatingnearby coastal plains and foothills. This mosaic setting including

Fig. 7. Simplified diagram of Dune Pond for the historical period plotted against mean calibrated ages. It shows main pollen curves, introduced and cultural taxa, selected NPPs, SCPresults and local population census. The latter has been obtained from the California State Data Center, Demographic Research Unit, and includes the population of the Goleta andSanta Barbara cities. Exaggeration curve is 10�.
A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196 191
elements of dune mats, coastal scrub, chaparral and oak woodlanddominated this Southern California coastal landscape during theMiddle and late Holocene, a pattern which is also reported in otherSouthern California paleoenvironmental records such as SoledadPond and Abalone Rocks Marsh in Santa Rosa Island (Cole and Liu,1994; Anderson et al., 2010), San Joaquin Marsh in Orange County(Davis, 1992), Los Pe~nasquitos lagoon in San Diego County (Cole andWahl, 2000) and the Santa Barbara Basin marine record (Heusser,1978).
During the late Holocene, changes in the pollen and NPP recordindicate successive phases of higher and lower water levels in DunePond (Figs. 4, 5 and 7). Between 3700 and 2100 cal yr BP the in-crease of algal spores (Spirogyra sp., Botryococcus), sedges andaquatic pollen (Potamogeton), together with the disappearance ofsmut fungi and gypsum, suggest generally higher groundwatertables and perhaps the existence of a more permanent water bodythan before (Figs. 4 and 5). Variability within this period does exist,with Spirogyra and Botryococcus suggesting wetter phases fromca 3400e2800 and 2500e2100 cal yr BP. This coincides withenhanced ENSO activity between 3400 and 2400 cal yr BP (Moyet al., 2002), which could have resulted in increased winter pre-cipitation and thus higher moisture availability in coastal wetlandssuch as Dune Pond. A similar context of increased freshwater runoffor higher groundwater tables in coastal marshes of southern Cali-fornia is found in many other records, such as at Abalone RocksMarsh (Cole and Liu, 1994; Anderson et al., 2010) and San JoaquinMarsh (Davis, 1992).
These coastal environmental conditions contrast with paleocli-matic reconstructions from more inland records from southernCalifornia. Both Dingemans et al. (2014) and Kirby et al. (2014)suggest predominantly dry environmental conditions at ZacaLake between 2700 and 2100 cal BP (the late Holocene Dry Periodof Mensing et al., 2013), although interpretations of Kirby et al. aremore complex, with a wetter phase ca 2800e2500 cal yr BP. Kirbyet al. (2010, 2012) suggested dry conditions at Lake Elsinore3200e2000 cal BP and at Lower Bear Lake ca 3000e2000 cal BP,respectively, in relation to a phase of aridity reported in the centralGreat basin between the 3rd and 2nd millennia cal BP (Mensinget al., 2013). We suggest that local climate conditions may play arole in the difference in climate interpretations, or that the records
have identified a gradient of effective precipitation for this periodfrom coastal to inland sites. For instance, Zaca Lake occurs ~50 kmfrom the ocean at 730 m elevation, outside the fog belt. However,Dune Pond sits only 250 m from the ocean and within the coastalfog belt. Support for this comes from other coastal locations furtherto the north, where evidence for dry conditions was absent orequivocal (Anderson et al., 2013; Cowart and Byrne, 2013).
Following this period, increased drying at Dune Pond occurredfrom 2100 to 700 cal yr BP. Lowered groundwater tables are re-flected in the paleoenvironmental record through changes in pollenand NPPs e i.e. retreat and near disappearance of algal spores andaquatic taxa, and an increase in smut fungi e but also in the sedi-mentary character of the deposits through the reappearance ofgypsum sand layers. Recurrent or periodic drying of the pond mayhave ultimately affected pollen and NPP preservation, suggestedhere by decreasing pollen concentration and the disappearance ofmost NPP taxa from 1500 cal yr BP. Using these indicators, wetterconditions are again found in Dune Pond from 700 cal yr BP untilthe historical period, where drying conditions may have prevailed.Such changes inmoisture availability are in general agreement withincreased regional climatic variability during the late Holocene(Barron et al., 2010; Barron and Anderson, 2011).
7.2. Prehistoric fire history
Charcoal particles are found in almost every sample analyzedfrom the Dune Pond record, signifying that fire was a regularfeature intimately linked with the dominating coastal mosaicvegetation over the last 5000 years. These results agree with pre-vious fire reconstructions from the Santa Rosa Island records, whichshow a general increase in charcoal influx through the Holoceneafter ~4500 cal yr BP associated with greater biomass developmentand human influence (Anderson et al., 2010).
Local low frequency fires are interpreted from 5000 to3650 cal yr BP in a context of reduced regional fire activity(decreasing trends of Cback) since 4600 cal BP (Fig. 6). Warm and dryclimatic conditions documented in Dune Pond and elsewhere insouthern California during this period may have favored lowerwinter precipitation and thus low fuel production for burning. Thiscoupled with low human settlement documented in the Santa

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196192
Barbara Channel area between ~4950 and 3950 cal yr BP (Glassowet al., 2007) probably explains the low frequency fire regime at thistime. Indeed, the beginning of the Dune Pond record coincides withthe abandonment of various nearby coastal sites (SBA-83, 84, 78,53) at the transition from the 4th to the 3rd millennia BC (Glassow,1997). Natural lighting-induced fires are indeed scarce in coastalCalifornia (Keeley, 2006; van Wagtendonk and Cayan, 2008) and,unlike other regions in southern California, this area has a lowexposure to extreme fireweather events related to Santa Anawinds(Moritz, 2003).
This pattern changes at ~3100 cal yr BP (~BC 1150), when localfire events become significantly more frequent, as shown by theincrease in charcoal peaks and the increase in FEF, which reachesmaximum values for the record at ~2000 cal yr BP (~BC 50). Theincreasing trend of background CHAR since 3700 cal BP also sig-nifies a rise in regional fires. The occurrence of cooler and local wetconditions revealed at Dune Pond (Fig. 6) and other paleoclimaticseries could have favored an increased fuel load during wintermonths and thus more frequent occurrences of local fires duringdry summer months. However, the fact that local fires persistedafter ~2100 cal BP, when a change towards lowered table tables isobserved (Fig. 6), suggests that forces other than climate and/orincreased biomass were probably involved. We suggest that thisrise in FEF occurs due to increased mainland coastal settlement,sedentism and cultural and socio-economic complexity during thelate Holocene. Evidence for this change in human activities isdocumented since ~4000e3500 cal yr BP (~BC 2000e1550) in themainland Santa Barbara coast and since ~2750 cal yr BP (~BC 800)in areas adjacent to Dune Pond, with the first settlement at sitesSBA-81 and 71 (Fig. 1, Table 1). Large clusters of semisubterraneanhouses documented at SBA-81 between 800 and 400 cal BC provideevidence of sedentism, architecture and village organization whichare typical of later Chumash sites (Erlandson and Rick, 2002). Thecoincidence of FEF and archaeological evidence at Dune Pond since~3100 cal yr BP supports our contention of the influence of NativeAmerican communities in increased coastal fires in Dune Pondduring the late Holocene. Even though more archaeological andhigh-resolution fire analyses are needed to confirm this hypothesis,recent coastal fire-reconstructions from Santa Rosa Island andnorth-central coastal California also stress the important relation-ship between increased fire activity and expanded settlement andsedentism of native populations during the late Holocene, espe-cially since 3500e3000 cal yr BP (Anderson et al., 2010, 2013;Cowart and Byrne, 2013).
Based on ethnographic data (Timbrook et al., 1982) the use offire must have been important for coastal prehistoric communitiesin promoting open areas and maintaining vegetation mosaics,especially in a wetter period, which may have favored theexpansion of mesic taxa such as oak woodland, chaparral andcoyote bush over sage scrub and grasslands. The fact that no suchchanges are evidenced in the pollen record between ~3700 and2100 cal yr BP argues in favor of the anthropogenic use of local andregional fires to maintain a coastal open and mosaic-like land-scape (Fig. 6). The increase of Poaceae in favor of Baccharis-type atthe peak of maximum FEF at ~2000 cal yr BP further support therelationship between fires and the opening of the landscape, asrecurrent fires preclude scrub invasion and promote grasslands incoastal California (Ford and Hayes, 2007). However drier condi-tions documented since 2100 cal yr BP could also have favored therecession of Baccharis and the expansion of grasslands. In any case,prehistoric societies benefited from an open landscape for multi-ple reasons, including enhancing the presence of wildlife forhunting. The regular presence of wild herbivores is confirmed inDune Pond by the presence of coprophilous fungi between~3000and 1200 cal yr BP. The Dune Pond vicinity would have been
attractive to prehistoric communities due to the availability of adiverse resource base, known to be valuable for later Chumashsocieties (Timbrook et al., 1982; Gamble, 2008). Proxies from ourrecord document resources such as freshwater, abundant grass-lands (Poaceae), coastal salt and freshwater marsh communities(Amaranthaceae, Cyperaceae) and wild herbivores (coprophilousfungi).
Despite local fire events persisting until ~1200 cal yr BP (~AD750), the sharp drop in reconstructed Cback documented after1900 cal yr BP could be related to decreased regional fire activity.The increases first in chaparral and then oak concomitant with thedecline in Cback may reflect the recovery of these communities innearby inland plains and foothills after the decrease in regional fireactivity. The development of these mesic vegetation groups duringa period of lower moisture availability would support thishypothesis.
Renewed local fire event frequency similar to that observedbetween ~3100 and 1500 cal yr BP also occurred from 650 cal yr BP(AD 1300) until the 18th century, when the largest CHAR accu-mulation is documented immediately prior to the 1793 Spanish firesuppression (Fig. 6). This local fire activity occurs within a period oflocal wetter conditions as reconstructed through pollen and NPPindicators in Dune Pond. As previously suggested, fires may haveprimarily impacted Baccharis-type communities growing in dunesand backdunes, favoring the expansion of herbaceous and scrubcommunities as indicated by the lowered ratios of Baccharis-typeand the rise of Amaranthaceae. This fire activity should be related tocontemporaneous Native American settlement documented nearbyDune Pond during this time (Fig. 1, Table 1), with sites SBA-78 andSBA-46 being amongst the largest villages in the Chumash regionduring the historic period (McLendon and Johnson, 1999; Gamble,2008). Charcoal and pollen analyses performed in domestic con-texts of SBA-46 support that the historic Chumash relied on theexploitation of fresh and salt marshes, mainly plants includedwithin the Amaranthaceae and Poaceae families and the Cichor-ioideae subfamily (Gamble, 2008). The use of fire by Chumash inthe Santa Barbara mainland region is furthermore well-documented by Spanish ethnohistoric accounts. Father Juan Cres-pi's journals during the Portol�a expeditions (1769e1770) stressesthat fires by native populations were mainly focused in coastal“grasses” and “salt-grasses” within a remarkably open landscape(Timbrook et al., 1982: 166e167). Crespi's historical account fitswell with the documentation from Dune Pond of a pre-historicallyburned, open landscape dominated by herbaceous communitiesgrowing in dunes and back dunes, especially belonging to theAmaranthaceae family. These results also coincide with nearbyhigh-resolution fire reconstructions from Santa Barbara Basinsediments (Mensing et al., 1999) and in chaparral of the Los PadresNational Forest (Lombardo et al., 2009), which document theoccurrence of regular fires in this region since AD 1425 and 1634,respectively.
Governor Jos�e Joaquin de Arrillaga's fire suppression procla-mation in 1793 banning the “harmful practice of setting fire topasture land” by the Indians in Alta California (Car, 1959) led to acomplete change in fire regimes, well documented in the DunePond record as a decline in CHAR and essential disappearance oflocal fires since the mid-18th century (Fig. 6). Our sedimentarycharcoal study thus agrees with the fire-scar and tree-ring study ofLombardo et al. (2009) from Los Padres National Forest, whichdocuments more regular fires during pre-historical times than afterthe years following Spanish settlement in the region. However,Mensing et al. (1999) questioned the effect of Spanish fire sup-pression on a larger scale in Southern California, suggesting nosignificant change in the fire regime between pre-historical andlater periods. Additional paleoenvironmental studies are of course

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196 193
needed to further enlighten this topic, but results obtained heresupport that the Mission period impacted the burning pattern ofcoastal areas in this region. The differences between our study andthose drawn byMensing et al. (1999) may be ascribed to scale, witha local, small-scale record in the case of Dune Pond, and primarilyregional scale in the case of the Santa Barbara Basin marine varvedsediment record. The close location of the pond to the Santa Bar-bara Mission and ranches (Fig. 1) perhaps encouraged the colonists'control over Native populations in supporting the fire suppressionproclamation.
7.3. Historical period: the shaping of transported coloniallandscapes
Our pollen and NPP record documents that the most significantenvironmental change within the Santa Barbara region occurredduring the historical period, beginning in the late 18th century.Beginning at a depth of ~77 cm in our core, the recording and in-crease in coprophilous fungal spores points towards an increasedpresence of herbivores at Dune Pond (Figs. 5 and 7). Many docu-mented fungal species are commonly found, and even specialize,on herbivore dunge e.g., Sordaria-type, Cercophora sp., Coniochaetacf. lignaria- (Ejarque et al., 2011) e and some, such as Sporormiella-type (Ahmed and Cain, 1972) e are dung obligates. This increasesignifies the introduction and expansion of grazing activities in theregion after establishment of the Santa Barbara Mission in 1786.Livestock population on the Mission increased rapidly, reachingover 16,600 animals by 1814 (Geiger, 1965). By the beginning of the19th century, nearly all the ranches of the Mission were fullystocked with grazing animals. This includes the San Miguel ranch,located in Goleta Slough near to Dune Pond, which had over 2700sheep in 1804 (O'Keefe, 1886: 17e18). Dune Pond should have beenan attractive water source for herds at this time.
The presence of livestock also impacted the nutrient status ofthe pond waters, as suggested by the increase of resting eggs ofBrachionus sp., a rotifer species indicator of eutrophic lakes in NorthAmerica (Gannon and Stemberger, 1978). The occurrence of whip-worm eggs (Trichuris sp.) at this time is also significant. Members ofTrichuris sp. are fecal-borne parasitic worms infecting animals andhumans, causing among other diseases, diarrhea and dysentery inhosts (Griffiths, 1978; Reinhard et al., 2013). Its presence here couldbe attributed to livestock and their dung in the Dune Pond water-shed. Despite the fact that Dune Pond was frequented by wildherbivores during prehistoric times, parasitic eggs only appear withthe introduction of livestock. Grazing activities thus contributed tothe contamination of freshwater bodies and, probably, the spread ofdiseases among the Native population staffing the ranchos. Epi-demics and diseases were common amongst missionized Indiansand dysentery was reported in Spanish archives as one of the mostdeadly (Archibald, 1978).
The introduction of grazing had amajor impact on native coastalvegetation, with the contemporary near disappearance of Amar-anthaceae growing on coastal salt and brackish marshes, coastalbluffs and foredunes (Fig. 7). However, a coastal scrub vegetation,dominated by Baccharis and grasses (Poaceae) with Ambrosia wasfavored under these conditions. This occurs within the context ofdrier climate conditions and continued erosion, as shown byincreased SARs and elevated percentages of Glomus (Fig. 5), a ve-sicular arbuscular mycorrhizal fungal spore found in soils, butfrequently associated with erosive processes in lakes and ponds(Marinova and Atanassova, 2006). Disturbance associated with theincrease in exposed soils due to grazing, and the absence of localfires, are factors that favor the expansion of Baccharis scrub incoastal California (Ford and Hayes, 2007). Research on modernvegetation communities at Elkhorn Slough farther north along the
California coast has shown that human trampling and cattle grazingcontributed to an increase in bare ground, the loss of marsh habitatand a change in community composition, with the decrease ofpickleweed (Salicornia), an increase of grasses and the invasion ofnon-native species (Woolfolk,1999). All these changes are observedin Dune Pond at the same time as the introduction of grazing,including the introduction of non-native herbs such as Erodium.The rise of Erodium during the Mission period should be attributedto Erodium cicutarium, a Eurasian weed native to Mediterraneancoastal sands and grasslands (Mensing and Byrne, 1998). Likewise,increasing trends of other herbaceous pollen taxa, such as Poaceae,Brassicaceae or Cichorioideae observed at this time reflect thespread of exotic-dominated grasslands with ruderal taxa (Fig. 7).Some Eurasian species in these groups e i.e. Bromus sp, Phalarisaquatica, Raphanus sp., Sonchus sp. e are currently common in theCOPR. Changes in the pollen record showing either the introductionor increase of many exotic taxa have been identified and related toearly European settlement in other paleoenvironmental records incoastal California (Mudie and Byrne, 1980; Davis, 1992; Mensingand Byrne, 1998; Anderson et al., 2010, 2013).
The development of grazing activities in the late 18th centuryseems to dominantly have affected coastal areas, while oakwoodland and chaparral communities located farther away fromthe coast in the foothills did not experience significant or long-lasting changes (Fig. 7). Likewise, cultivation activities seemed tohave played a minor role in this coastal area as minimal amounts ofOleaceae (olive) trees, Cerealia-type (cereals) and Vitis (grapes)(Fig. 7) are reported during the Mission period. Probably, morefertile soils in themission lands were selected for cultivation. This isconsistent with the late 19th century account of Father J.J. O'Keefe,who stressed Dos Pueblos, San Miguel, San Estevan and San Jos�eranches, the last two located west of Goleta between the present-day Hope Ranch and Arroyo Burro, as the preferred areas forgrowing wheat, corn, and other crops (1886: 18e19).
The American period during the mid-19th century documentsthe consolidation of the agro-pastoral landscape instituted by theSpanish missionaries. The continuous occurrence of coprophilousfungi underlines continued grazing in Dune Pond during the lasttwo centuries, which is attributed to ranches such as Rancho DosPueblos under Mexican and American rule (Tompkins, 1960).Cropping activities are better represented during the Americanthan under the Mission period, with the regular occurrence ofOleaceae and Juglans (walnut) trees, Vitis and increases in Cerealia-type. Low percentages of crops recorded at Dune Pond probablyreflect agricultural activities in the surrounding ranches. Olive andwalnut trees became popular in Santa Barbara during the late 19thcentury and groves were planted in the Goleta valley (Ruhge, 1991;Soleri et al., 2010). Together with the consolidation of an agro-pastoral system, the American period was also a period of signifi-cant population growth, urban development and the onset of in-dustrial activities, most importantly the development of thepetroleum industry, as shown in the paleoenvironmental recordthrough the rise of SCP. Development and urbanization negativelyimpacted oak woodlands and chaparral communities, which seri-ously declined during the first half of the 20th century (Fig. 7).
American immigrants also introduced a new suite of herbs andtrees. A case in point is the alien Eucalyptus (gum) trees, whichwerefirst introduced from Australia to Santa Barbara in 1870 (Esau,2006), and of native taxa such as Monterrey cypress (Cupressa-ceae), naturally growing in Monterey County farther north butwidely planted as an ornamental in coastal California. Increases ofEucalyptus and Cupressaceae in the pollen record show the modernintroduction of these trees in the vicinity of Dune Pond, wherestands of both gum trees and Monterey Cypress currently grow(Fig. 1). Likewise, pollen taxa including exotic weeds such as

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196194
Plantago sp. or Rumex sp., increases, and others, like P. aviculare-type are first documented and specifically related to the Americansettlement.
8. Conclusion
Using a combination of multi-proxy paleoecologic techniques,some well-established but others being novel approaches, 5000years of landscape change and fire history have been reconstructedfrom a small coastal pond in southern California. Dune Pond wasformed during the Middle Holocene as a shallow freshwater bodysensitive to regional climatic variability. Changes in the composi-tion of aquatic and marsh pollen and NPPs between 4600 and3700 cal yr BP indicate that relatively dry conditions prevailed atthe pond during the Middle Holocene, when generally arid condi-tions and warm sea temperatures are recorded in Southern Cali-fornia (Kennett et al., 2007; Barron and Anderson, 2011). During thelate Holocene, successive phases of higher (3700 e 2100 cal yr BP,700e150 cal yr BP) and lower (2100e700 cal yr BP, historicalperiod) local moisture availability reflect increased climatic vari-ability (Barron et al., 2010). Despite this climatic variability duringthe late Holocene, no major changes are observed in the mainterrestrial vegetation groups until the historic period, with thelandscape continuously dominated by a coastal mosaic vegetation.This mosaic included dune mats, coastal scrub, salt marshes andcoastal bluff scrub on and near the coastal dunes and backdunes,with chaparral and oak woodland growing in the valley plains andfoothills.
The high-resolution charcoal analysis performed at Dune Pondshows a complex interaction with vegetation mosaic, climate andhuman settlement over the last 5000 years. The low frequency oflocal fires and decreasing regional fire activity during the MiddleHolocene probably resulted from sparse wildfires which happenedin a context of low biomass production favored by prevailing warmand dry conditions. However the increase in local fires documentedduring the late Holocene since 3100 cal yr BP seems to be related toa combination of factors. Two factors are important in southernCalifornia e a change towards wetter conditions during wintermonths favoring increased fuel loads, and increased mainlandcoastal settlement, sedentism and cultural and socio-economiccomplexity since ~4000e3500 cal yr BP. Greater impact on thelandscape is consistent with the development of a more sedentarysettlement and the diversified subsistence pattern of late Holoceneprehistoric coastal communities. Comparison of the pollen and NPPwith the fire record shows that fire was an important tool forkeeping amosaic-like open landscape attractive to huntingwildlife.This may have been particularly important for the period of wetterconditions between 3700 and 2100 cal yr BP, where mesic com-munities such as oak woodland, chaparral and coyote bush scrubwould have expanded. The anthropogenic use of fire by NativeAmerican societies to maintain an open mosaic of coastal vegeta-tion is also suggested from 650 cal yr BP (AD 1300) until the 18thcentury, when renewed fire activity is again coupled with wetterconditions and documented human settlement near the pond. Inthis case, the use of fire by Chumash societies in this region isfurther corroborated by ethnographic accounts from the Spanishcolonists. In both cases the combination of wet conditions and thepresence of prehistoric settlement in the region seem to have beena trigger favoring fires in this coastal area. The Euro-Americanpeopling of the Santa Barbara region contributed to a significantchange in this pattern, as fire suppression e first by Spanish andlater by Americans e and subsequent urbanization of the areaprecluded further fire activity around Dune Pond during the his-torical period.
The primary environmental e vegetation and land use e changefor the Dune Pond record occurs during the historic period. Thehigh-resolution paleoecological analyses, coupled with a multi-proxy chronological model has allowed us to characterize distinctlandscape changes linked to the Spanish e Mexican e Americansettlement in the region. During the late 18th century Spanishcolonists brought to Santa Barbara a cultural landscape whichmirrored the agro-pastoral model of contemporaneous Europeancountries. Introduced livestock grazing activities, determined bythe presence of coprophilous fungal spores in the record, played amajor role in this process. They favored erosive processes andaltered the composition of coastal vegetation, degrading salt andbrackish marsh vegetation (Amaranthaceae) and promoting theexpansion of coyote bush and the invasion of alien ruderal andgrassland taxa. They also favored the introduction of fecal-borneparasites in freshwater bodies like Dune Pond and potentially thespread of diseases among both animals and human populations.The Mission period also led to the banning of fires by native pop-ulations and the first introduction of cropping activities. Introducedagro-pastoral practices were primarily focused on coastal ratherthan inland areas, as suggested by the greater modificationobserved on the coastal vegetation in relation to inland oakwoodlands and chaparral, which remained unaltered during theSpanish and Mexican periods. This agro-pastoral landscape wasconsolidated during the succeeding American period of SantaBarbara, with a greater role for cultivation, the development ofindustrial activities, increased human population and urbandevelopment. This later period also witnessed the introduction ofdistinct alien herbs and trees e i.e. gum tree, Monterrey cypress, P.aviculare-type- and amore extended impact on the landscape, withthe clearing of inland woodlands and chaparral.
This study stresses some negative environmental consequencesthat followed the Euro-American colonization of southern Califor-nia, such as a loss of native coastal vegetation habitats. However,with the introduction of new land-uses and plants, Euro-Americansettlers gave place to a newly born cultural landscape, which notonly suited their own socio-economic and cultural patterns but wasconsistent with those of the current California population. Thisstudy reflects how deeply bound many human societies are to theirsurrounding landscapes, so that when societies move, they oftentake basic elements of their landscapes with them leading to sig-nificant landscape changes in colonized territories. It also showsthat high-resolution paleoecological analyses using multiple in-dicators such as pollen, NPPs, macrocharcoal or SCP can effectivelytrace transported landscapes in the past.
Acknowledgments
We would like to thank Lisa Stratton (Cheadle Center forBiodiversity & Ecological Restoration) for her help in providinginformation and cartographic material about the vegetationcomposition at the COPR. Coring was made possible through fundsfrom the Santa Barbara Coastal Long Term Ecological ResearchProject. This research was supported by a Beatriu de Pin�os post-doctoral fellowship (2010 BP_A 00006) from the Agency for Man-agement of University and Research Grants of the Government ofCatalonia to A.E. Laboratory of Paleoecology Contribution Number161.
References
Ahmed, S.E., Cain, R.F., 1972. Revision of the genera Sporormia and Sporormiella.Can. J. Bot. 50, 419e477.
Anderson, E., 1952. Plants, Man, and Life. University of California Press, Berkeley.

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196 195
Anderson, R.S., Ejarque, A., Brown, P.M., Hallett, D., 2013. Holocene and historicvegetation change and fire history in the mid-coastal region of California. Ho-locene 23 (12), 1795e1808.
Anderson, R.S., Smith, S.J., 1994. Paleoclimatic interpretations of meadow sedimentand pollen stratigraphies from California. Geology 22, 723e726.
Anderson, R.S., Ejarque, A., Rice, J., Smith, S.J., Lebow, C.G., 2015. Historic and Ho-locene environmental change in the San Antonio Creek Basin, Mid-coastalCalifornia. Quat. Res. (in press).
Anderson, R.S., Starratt, S., Jass, R.B., Pinter, N., 2010. Fire and vegetation history onSanta Rosa Island, Channel Islands, and long-term environmental change inSouthern California. J. Quat. Sci. 25, 782e797.
Archibald, R., 1978. The Economics Aspects of the California Missions. Academy ofAmerican Franciscan History, Washington D.C.
Baker, G., 2003. Santa Barbara. Another Harbour Town History. Hignell BookPrinting, Canada.
Barbour, M.G., Keeler-Wolf, T., Schoenherr, A.A., 2007. Terrestrial Vegetation ofCalifornia, third ed. University of California Press, Berkeley.
Barron, J.A., Anderson, L., 2011. Enhanced Late Holocene ENSO/PDO expressionalong the margins of the eastern North Pacific. Quat. Int. 235, 3e12.
Barron, J.A., Bukry, D., Field, D., 2010. Santa Barbara Basin diatom and silicoflagellateresponse to global climate anomalies during the past 2200 years. Quat. Int. 215,34e44.
Blaauw, M., 2010. Methods and code for ‘classical’ age-modelling of radiocarbonsequences. Quat. Geochronol. 5, 512e518.
Boden, T.A., Marland, G., Andres, R.J., 2013. Global, Regional, and National Fossil-Fuel CO2 Emissions. Carbon Dioxide Information Analysis Center, Oak RidgeNational Laboratory, Tennessee, USA. http://dx.doi.org/10.3334/CDIAC/00001_V2010.
Bossard, C.C., Randall, J.M., 2007. Nonnative plants of California. In: Barbour, M.G.,Keeler-Wolf, T., Schoenherr, A.A. (Eds.), Terrestrial Vegetation of California, thirded. University of California Press, Berkeley, pp. 107e123.
Car, C.R., 1959. California Government and Forestry from Spanish Days Until theCreation of the Department of Natural Resources in 1927. Division of Forestry,Department of Natural Resources, State of California Sacramento, CA.
Cole, K.L., Liu, G., 1994. Holocene paleoecology of an estuary on Santa Rosa Island,California. Quat. Res. 41, 326e335.
Cole, K.L., Wahl, E., 2000. A Late Holocene paleoecological record from Torrey PinesState Reserve, California. Quat. Res. 53, 341e351.
Cowart, A., Byrne, R., 2013. A paleolimnological record of late Holocenevegetation change from the central California coast. Calif. Archaeol. 5 (2),337e352.
Davis, O.K., 1992. Rapid climatic change in coastal southern California inferred frompollen analysis of San Joaquin Marsh. Quat. Res. 37, 89e100.
Dingemans, T., Mensing, S.A., Feakins, S.J., Kirby, M.E., Zimmerman, S.R.H., 2014.3000 years of environmental change at Zaca Lake, California, USA. Front. Ecol.Evol. 2 (34), 1e16.
Ejarque, A., Miras, Y., Riera, S., 2011. Pollen and non-pollen palynomorphs indicatorsof vegetation and highland grazing activities obtained frommodern surface anddung datasets in the eastern Pyrenees. Rev. Palaeobot. Palynol. 167, 123e139.
Ellis, M.B., 1971. Dematiaceous Hyphomycetes. Commonwealth Mycological Insti-tute, Kew.
Erlandson, J.M., Rick, T.C., 2002. Late Holocene cultural developments along theSanta Barbara Coast. In: Erlandson, J.M., Jones, T.L. (Eds.), Catalysts toComplexity: Late Holocene Societies of the California Coast. Cotsen Institute ofArchaeology, University of California, Los Angeles, pp. 166e182.
Erlandson, J.M., Rick, T.C., Braje, T.J., Casperson, M., Culleton, B., Fulfrost, B., Garcia, T.,Guthrie, D.A., Jew, N., Kenneth, D.J., Moss, M.L., Reeder, L., Skinner, C., Watts, J.,Willis, L., 2011. Paleoindian seafaring, maritime technologies, and coastalforaging on California's Channel Islands. Science 331, 1181e1184.
Esau, E., 2006. Images of the Pacific Rim: Australia and California, 1850e1935. Po-wer Publications, Australia, Sydney.
Faegri, K., Iversen, J., 1989. Textbook of Pollen Analysis (revised by K. Faegri, P. E.Kaland, and K. Krzwinski), fourth ed. Wiley, Chichester.
Ford, L.D., Hayes, G.F., 2007. Northern coastal scrub. In: Barbour, M.G., Keeler-Wolf, T., Schoenherr, A.A. (Eds.), Terrestrial Vegetation of California, third ed.University of California Press, Berkeley, pp. 180e207.
Gamble, L.H., 2008. The Chumash World at European Contact. Power, Trade, andFeasting Among Complex Hunter-Gatherers. University of California Press,Berkeley.
Gannon, J.E., Stemberger, R.S., 1978. Zooplankton (especially crustaceans and roti-fers) as indicators of water quality. Trans. Am. Microsc. Soc. 97, 16e35.
Gavin, D.G., Brubaker, L.B., Lertzman, K.P., 2003. An 1800-year record of the spatialand temporal distribution of fire from the west coast of Vancouver Island,Canada. Can. J. For. Res. 33, 573e586.
Gavin, D.G., Hu, F.S., Lertzman, K., Corbett, P., 2006. Weak climatic control ofstandscale fire history during the Late Holocene in southeastern BritishColumbia. Ecology 87, 1722e1732.
Geiger, M., 1965. Mission Santa Barbara, 1782-1965. Franciscan Fathers of California,Santa Barbara.
Glassow, M.A., 1997. Middle Holocene cultural development in the central SantaBarbara Channel region. In: Erlandson, J.M., Glassow, M.A. (Eds.), The Archae-ology of the California Coast During the Middle Holocene. Cotsen Institute ofArchaeology, University of California, Los Angeles, pp. 73e90.
Glassow, M.A., Gamble, L.H., Perry, J.E., Russell, G.S., 2007. Prehistory of theNorthern California Bight and the adjacent transverse ranges. In: Jones, T.L.,
Klar, K.A. (Eds.), California Prehistory. Colonization, Culture, and Complexity.Altamira Press, Lanham, MA, pp. 191e213.
Griffiths, H.J., 1978. A Handbook of Veterinary Parasitology. Domestic Animals ofNorth America. University of Minnesota Press, Minneapolis.
Grimm, E.C., 1987. Coniss - a Fortran-77 Program for stratigraphically constrainedcluster-analysis by the method of incremental sum of squares. Comput. Geosci.13, 13e35.
Gurrola, L.D., Keller, E.A., Chen, J.H., Owen, L.A., Spencer, J.Q., 2014. Tectonic geo-morphology of marine terraces: Santa Barbara fold belt, California. Geol. Soc.Am. Bull. 126, 219e233.
Harrison, W.M., 1965. Mikiw: a Coastal Chumash Village. Annual Reports of theUniversity of California Archaeological Survey 7, pp. 91e178.
Hardwick, M.R., 2005. Changes in Landscape. The Beginnings of Horticulture in theCalifornia Missions, second ed. The Paragon Agency, California.
Heusser, L., 1978. Pollen in Santa Barbara Basin, California: a 12,000-yr record. Geol.Soc. Am. Bull. 89, 673e678.
Higuera, P.E., Sprugel, D.G., Brubaker, L.B., 2005. Reconstructing fire regimes withcharcoal from small-hollow sediments: a calibration with tree-ring records offire. Holocene 15, 238e251.
Higuera, P.E., Peters, M.E., Brubaker, L.B., Gavin, D.G., 2007. Understanding the originand analysis of sediment-charcoal records with a simulation model. Quat. Sci.Rev. 26, 1790e1809.
Higuera, P.E., Brubaker, L.B., Anderson, P.M., Hu, F.S., Brown, T.A., 2009. Vegetationmediated the impacts of postglacial climatic change on fire regimes in thesouthcentral Brooks Range, Alaska. Ecol. Monogr. 79, 201e219.
Higuera, P.E., Gavin, D.G., Bartlein, P.J., Hallett, D.J., 2010. Peak detection insediment-charcoal records: impacts of alternative data analysis methods onfire-history interpretations. Int. J. Wildland Fire 19, 996e1014.
Inoue, J., Tomozawa, A., Okudaira, T., 2013. The use of size distributions of sphe-roidal carbonaceous particles in swimming pool deposits for evaluating at-mospheric particle behaviour. Water Air Soil Pollut. 224, 1580.
Juggins, S., 2011. C2 Data Analysis Version 1.7.2. University of Newcastle, Newcastleupon Tyne.
Keeley, J.F., 2006. South coast bioregion. In: Sugihara, N.G., van Wagtendonk, J.,Shaffer, K.E., Fites-Kaufman, J., Thode, A.E. (Eds.), Fire in California's Ecosystems.University of California Press, Berkeley, CA, pp. 350e390.
Kennett, D.J., Kennett, J.P., Erlandson, J.M., Cannariato, K.G., 2007. Human responsesto Middle Holocene climate change on California's Channel Islands. Quat. Sci.Rev. 26, 351e367.
Ketterer, M.E., Hafer, K.M., Jones, V.J., Appleby, P.G., 2004. Rapid dating of recentsediments in Loch Ness: inductively coupled plasma mass spectrometric mea-surements of global fallout plutonium. Sci. Total Environ. 322, 221e229.
Kirby, M.E., Lund, S.P., Anderson, M.A., Bird, B.W., 2007. Insolation forcing of Ho-locene climate change in southern California: a sediment study from LakeElsinore. J. Paleolimnol. 28, 395e417.
Kirby, M., Lund, S., Patterson, W., Anderson, M., Bird, B., Ivanovici, L., Monarrez, P.,Nielsen, S., 2010. A Holocene record of Pacific Decadal Oscillation (PDO)-relatedhydrologic variability in southern California (Lake Elsinore, CA). J. Paleolimnol.44, 819e839.
Kirby, M.E., Zimmerman, S.R.H., Patterson, W.P., Rivera, J.J., 2012. A 9170-year recordof decadal-to-multi-centennial scale pluvial episodes from the coastal South-west United States: a role for atmospheric rivers? Quat. Sci. Rev. 46, 57e65.
Kirby, M.E., Feakins, S.J., Hiner, C.A., Fantozzi, J., Zimmerman, S.R.H., Dingemans, T.,Mensing, S.A., 2014. Tropical Pacific forcing of Late-Holocene hydrologic vari-ability in the coastal southwest United States. Quat. Sci. Rev. 102, 27e38.
Kirch, P.V., 1982. The impact of the prehistoric Polynesians on the Hawaiianecosystem. Pac. Sci. 36, 1e14.
Kodsueb, R., Dhanasekaran, V., Aptroot, A., Lumyong, S., McKenzie, E.H.C.,Hyde, K.D., Jeewon, R., 2006. The family Pleosporaceae: intergeneric relation-ships and phylogenetic perspectives based on sequence analyses of partial 28SrDNA. Mycologia 98 (4), 571e583.
Larsen, J., 2003. Size distributions and concentrations of spheroidal carbonaceousfly-ash particles (SCPs) in lake sediments as an aid to detecting locally depos-ited atmospheric pollution. Water Air Soil Pollut. 149, 163e175.
Lombardo, K.J., Swetnam, T.W., Baisan, C.H., Borchert, M.I., 2009. Using bigconeDouglas-fir fire scars and tree rings to reconstruct interior chaparral fire history.Fire Ecol. 5 (3), 32e52.
Macdonald, K.B., 1988. Coastal salt marsh. In: Barbour, M.G., Major, J. (Eds.),Terrestrial Vegetation of California. California Native Plant Society, Sacramento,pp. 263e294.
McLendon, S., Johnson, J.R., 1999. Cultural Affiliation and Lineal Descent of ChumashPeoples in the Channel Islands and Santa Monica Mountains, vol. I. Reportsubmitted to the Archaeology and Ethnography Program, National Park Service.Santa Barbara Museum of Natural History, Santa Barbara.
Marinova, E., Atanassova, J., 2006. Anthropogenic impact on vegetation and envi-ronment during the Bronze Age in the area of lake Durankulak, NE Bulgaria:pollen, microscopic charcoal, non-pollen palynomorphs and plant macrofossils.Rev. Palaeobot. Palynol. 141, 165e178.
Mensing, S.A., Byrne, R., 1998. Pre-mission invasion of Erodium cicutarium in theCalifornia grassland. J. Biogeogr. 25, 757e762.
Mensing, S.A., Michaelsen, J., Byrne, R., 1999. A 560-year record of Santa Ana firesreconstructed from charcoal deposited in the Santa Barbara Basin, California.Quat. Res. 51, 295e305.
Mensing, S.A., Sharpe, S.E., Tunno, I., Sada, D.W., Thomas, J.M., Starratt, S., Smith, J.,2013. The Late Holocene Dry period: multiproxy evidence for an extended

A. Ejarque et al. / Quaternary Science Reviews 112 (2015) 181e196196
drought between 2800 and 1850 cal yr BP across the central Great Basin, USA.Quat. Sci. Rev. 78, 266e282.
Minor, S.A., Kellogg, K.S., Stanley, R.G., Gurrola, L.D., Keller, E.A., Brandt, T.R., 2009.Geologic Map of the Santa Barbara Coastal Plain Area, Santa Barbara County,California. U.S. Geological Survey Special Investigations Map 3001. Scale 1:25,000; sheet one of one.
Moritz, M.A., 2003. Spatiotemporal analysis of controls on shrubland fire regimes:age-dependency and fire hazard. Ecology 84 (2), 351e361.
Moy, C.M., Seltzer, G.O., Rodbell, D.T., Anderson, M.D., 2002. Variability of El Ni~no/Southern Oscillation activity at millennial timescales during the Holoceneepoch. Nature 240 (6912), 162e165.
Mudie, P.J., Byrne, R., 1980. Pollen evidence for historic sedimentation rates inCalifornia salt marshes. Estuar. Coast. Mar. Sci. 10, 305e316.
Munz, P.A., 1974. A Flora of Southern California. University of California Press,Berkeley.
Nehring, R., 1975. Oil and Gas Supplies for California: Past and Future. Reportprepared for the California State Assemby from the Rockefeller foundation.RAND Corporation, California.
O'Keefe, J.J., 1886. The Buildings and Churches of the Mission of Santa Barbara. AHandbook of Authentic Information on the Mission of Santa Barbara from ItsFoundation to the Present Day. Independent Job Printing House, Santa Barbara.
Oldfield, F., Richardson, N., Appleby, P.G., 1995. Radiometric dating (210Pb, 137Cs,241Am) of recent ombrotrophic peat accumulation and evidence for changes inmass balance. Holocene 5, 141e148.
Palet, J.M., Orengo, H.A., 2011. The Roman centuriated landscape: conception,genesis, and development as inferred from the Ager Tarraconensis Case. Am. J.Archaeol. 115 (3), 383e402.
Plants of Coal Oil Point Reserve. http://coaloilpoint.ucnrs.org/Species/COPR%20Plant%20List.htm, (accessed 10.07.13.).
Punt, W., Blackmore, S., Hoen, P., Stafford, P., 1976e2009. The Northwest EuropeanPollen Flora, vols. IeIX. Elsevier, Amsterdam.
Reimer, P.J., Bard, E., Bayliss, A., Beck, J.W., Blackwell, P.G., Bronk Ramsey, C.,Grootes, P.M., Guilderson, T.P., Haflidason, H., Hajdas, I., Hatte, C., Heaton, T.J.,Hoffmann, D.L., Hogg, A.G., Hughen, K.A., Kaiser, K.F., Kromer, B., Manning, S.W.,Niu, M., Reimer, R.W., Richards, D.A., Scott, E.M., Southon, J.R., Staff, R.A.,Turney, C.S.M., van der Plicht, J., 2013. IntCal13 and Marine13 radiocarbon agecalibration curves 0-50,000 ears cal BP. Radiocarbon 55 (4), 1869e1887.
Reinhard, K.J., Ferreira, L.F., Bouchet, F., Sianto, L., Dutra, J.M.F., Iniguez, A., Leles, D., LeBailly, M., Fugassa, M., Pucu, E., Araújo, A., 2013. Food, parasites, and epidemio-logical transitions: a broad perspective. Int. J. Paleopathol. 3 (3), 150e157.
Reille, M., 1992. Pollen et spores d'Europe et d'Afrique du nord. Marseille. Labo-ratoire de Botanique historique et Palynologie, Universit�e d'Aix-Marseille III.
Rick, T.C., Sillett, S.T., Ghalambor, C.K., Hofman, C.A., Ralls, K., Anderson, R.S.,Boser, C., Braje, T.J., Cayan, D., Chesser, T., Collins, P.W., Erlandson, J.M.,Faulkner, K.R., Funk, W.C., Fleischer, R., Galipeau, R., Huston, A., Junak, S., King, J.,Maldonado, J., McEachern, K., Muhs, D.R., Newsome, S.D., Reeder-Myers, L.,
Still, C., Morrison, S.A., 2014. From the Pleistocene to the Anthropocene: 20,000years of ecological change and the future of biodiversity on California's ChannelIslands. Bioscience (in press).
Rose, N., 1990. A method for the extraction of carbo-naceous particles from lakesediment. J. Paleolimnol. 3, 45e53.
Rose, N.L., 2001. Fly-ash particles. In: Last, W.M., Smol, J.P. (Eds.), Tracking Envi-ronmental Change Using Lake Sediments, Physical and Geochemical Methods,vol. 2. Kluwer Academic Publishers, Dordrecht, pp. 319e349.
Ruhge, J.M., 1991. Looking Back. A History of Goleta's Historic Structures and Sitesand the Pioneer Families who made Them. Getting in Touch with Our Heritage.Quantum Imaging Associates, Lompoc, CA.
Schmitt, R.J., Dugan, J.E., Adamson, M.R., 2003. Industrial Activity and Its Socio-economic Impacts: Oil and Three Coastal California Counties. MMS OCS Study2002-049. Coastal Research Center, Marine Science Institute, University ofCalifornia, Santa Barbara, CA. MMS Cooperative Agreement Number 14-35-01-00-CA-31603. 24.
Soleri, D., Koehmstedt, A., Aradhya, M.K., Polito, V., Pinney, K., 2010. Comparing thehistoric olive trees (Olea europaea L.) of Santa Cruz Island with contempora-neous trees in the Santa Barbara, CA area: a case study of diversity and structurein an introduced agricultural species conserved in situ. Genet. Resour. CropEvol. 57, 973e984.
Sluyter, A., 2002. Colonialism and Landscape. Postcolonial Theory and Applications.Rowman and Littlefield Publishers, Inc, Maryland.
Stockmarr, J., 1971. Tablets with spores used in absolute pollen analysis. PollenSpores 13, 615e621.
Timbrook, J., Johnson, J.R., Earle, D.D., 1982. Vegetation burning by the Chumash.J. Calif. Gt. Basin Anthropol. 4 (2), 163e186.
Tompkins, W.A., 1960. Santa Barbara's Royal Rancho. The Fabulous History of LosDos Pueblos. Howell-North Books, Berkeley.
van Geel, B., 2001. Non-pollen palynomorphs. In: Smol, J.P., Birks, H.J.B., Last, W.M.(Eds.), Tracking Environmental Change Using Lake Sediments, Terrestrial, Algaland Silicaceaous Indicators, vol. 3. Kluwer Academic Publishers, Dordrecht,pp. 99e119.
van Geel, B., Aptroot, A., 2006. Fossil ascomycetes in Quaternary deposits. NovaHedwig. 82, 313e329.
Van Wagtendonk, J.W., Cayan, D.R., 2008. Temporal and spatial distribution oflightning strikes in California in relation to large-scale weather patterns. FireEcol. 4, 34e56.
V�anky, K., 1994. European Smut Fungi. Gustav Fischer Verlag, Stuttgart.Whitlock, C., Anderson, R.S., 2003. Fire history reconstructions based on sediment
records from lakes and wetlands. In: Veblen, T.T., Baker, W.L., Montenegro, G.,Swetnam, T.W. (Eds.), Fire and Climatic Change in Temperate Ecosystems of theAmericas, Ecological Studies, vol. 160. Springer-Verlag, New York, pp. 3e31.
Woolfolk, A.M., 1999. Effects of Human Trampling and Cattle Grazing on Salt MarshAssemblages in Elkhorn Slough, California (M.S. thesis). Moss Landing MarineLaboratories and California State University Sacramento.
Top Related
Copyright © 2022 FDOKUMEN

