
Ecology of a coastal lagoon to dune forest sequence, south Westland, New Zealand
14
New Zealand Journal of Botany, 1991, Vol. 29: 17-30 0028-825X/91/2901-0017$2.50/0 © Crown copyright 1991 17 Ecology of a coastal lagoon to dune forest sequence, south Westland, New Zealand ALASTAIR W. ROBERTSON* ALAN F.MARK J.BASTOW WILSON Botany Department University of Otago P.O. Box 56 Dunedin, New Zealand *Present address: Hatherly Laboratories, Department of Biological Sciences, University of Exeter, Exeter, EX4 4PS, U.K. Abstract The vegetation and floristic patterns in and around a coastal lagoon are described from quadrat sampling and related to several environmental factors (elevation, water depth, soil organic content, and pH). A cluster analysis was used to identify 14 communities while a detrended correspondence analysis ordinated the quadrats. The ordination's Axis 1 was correlated with water depth/ elevation, while Axis 2 was correlated with soil organic content and, in the opposite direction, with soil pH. Two vegetation development sequences are proposed for the dune hollow-ridge system: one in the hollows involving stages of aquatic, bog, and forest vegetation; the second on the ridges where forest composition changes in response to increasing soil development. Ratesof succession arepresumably very slow. Formal protection of the lagoon within the World Heritage area is justified because of its ecological, conservation, and scenic values; in light of diminishing natural wetlands; and the common occurrence of a vulnerable macrophyte, Myriophyllum robustum, in the lagoon. B90030 Received 16 July 1990; accepted 18 October 1990 Keywords vegetation sequence; vegetation analysis; wetland; dune forest; ecological factors; Haast Ecological District INTRODUCTION The lowland outwash gravel and coastal terraces of south Westland are replete with permanent wetlands and adjoining sequences of shrubland, woodland, and forest vegetation associated with increased elevation and lower water tables (Mark & Smith 1975; Dawson 1988, fig. 68; Norton 1989). Distinctive examples of such wetland sequences, unlike any of those previously described, are associated with the series of coastal and near-coastal sand dune-hollow alternations on the South Wesdand coastal plain of the Paringa and Haast Ecological Districts. One example is alongside State Highway 6 in Mataketake Forest c. 3 km north of the Waita River. Here, a coastal lagoon merges with mature mixed podocarp-broadleaved dune forest via zones of sedgeland and shrubland (Fig. 1). The aims of the study were to describe the vegetation zonation pattern and relate this to environmental parameters. The communities described by Wardle (1977) from Westland National Park, although some distance to the north, can be compared in some cases, though Wardle did not define aquatic communities, and any coastal dune forest in his area had been either disturbed or removed. THE STUDY AREA The lagoon studied (Fig. 1) is between old dunes within c. 10 m elevation and 100 m distance of the coast (Fig. 2), lying in a depression bounded on the seaward side by an active dune front and on the inland side by a stabilised dune crest. Located between the Waita River and Ship Creek (Fig. 2) the lagoon forms a typical unit of the long, narrow coastal plain between the Arawata River in the south and the coastal hills north of the Waita River. As such.adetailed study of the vegetation andassociated
Transcript of Ecology of a coastal lagoon to dune forest sequence, south Westland, New Zealand

New Zealand Journal of Botany, 1991, Vol. 29: 17-300028-825X/91/2901-0017$2.50/0 © Crown copyright 1991
17
Ecology of a coastal lagoon to dune forest sequence,south Westland, New Zealand
ALASTAIR W. ROBERTSON*ALAN F.MARKJ.BASTOW WILSON
Botany DepartmentUniversity of OtagoP.O. Box 56Dunedin, New Zealand
*Present address: Hatherly Laboratories,Department of Biological Sciences, Universityof Exeter, Exeter, EX4 4PS, U.K.
Abstract The vegetation and floristic patterns inand around a coastal lagoon are described fromquadrat sampling and related to severalenvironmental factors (elevation, water depth, soilorganic content, and pH). A cluster analysis wasused to identify 14 communities while a detrendedcorrespondence analysis ordinated the quadrats. Theordination's Axis 1 was correlated with water depth/elevation, while Axis 2 was correlated with soilorganic content and, in the opposite direction, withsoil pH.
Two vegetation development sequences areproposed for the dune hollow-ridge system: one inthe hollows involving stages of aquatic, bog, andforest vegetation; the second on the ridges whereforest composition changes in response to increasingsoil development. Ratesof succession arepresumablyvery slow.
Formal protection of the lagoon within the WorldHeritage area is justified because of its ecological,conservation, and scenic values; in light ofdiminishing natural wetlands; and the commonoccurrence of a vulnerable macrophyte,Myriophyllum robustum, in the lagoon.
B90030Received 16 July 1990; accepted 18 October 1990
Keywords vegetation sequence; vegetationanalysis; wetland; dune forest; ecological factors;Haast Ecological District
INTRODUCTION
The lowland outwash gravel and coastal terraces ofsouth Westland are replete with permanent wetlandsand adjoining sequences of shrubland, woodland,and forest vegetation associated with increasedelevation and lower water tables (Mark & Smith1975; Dawson 1988, fig. 68; Norton 1989).Distinctive examples of such wetland sequences,unlike any of those previously described, areassociated with the series of coastal and near-coastalsand dune-hollow alternations on the South Wesdandcoastal plain of the Paringa and Haast EcologicalDistricts. One example is alongside State Highway6 in Mataketake Forest c. 3 km north of the WaitaRiver. Here, a coastal lagoon merges with maturemixed podocarp-broadleaved dune forest via zonesof sedgeland and shrubland (Fig. 1).
The aims of the study were to describe thevegetation zonation pattern and relate this toenvironmental parameters. The communitiesdescribed by Wardle (1977) from Westland NationalPark, although some distance to the north, can becompared in some cases, though Wardle did notdefine aquatic communities, and any coastal duneforest in his area had been either disturbed or removed.
THE STUDY AREA
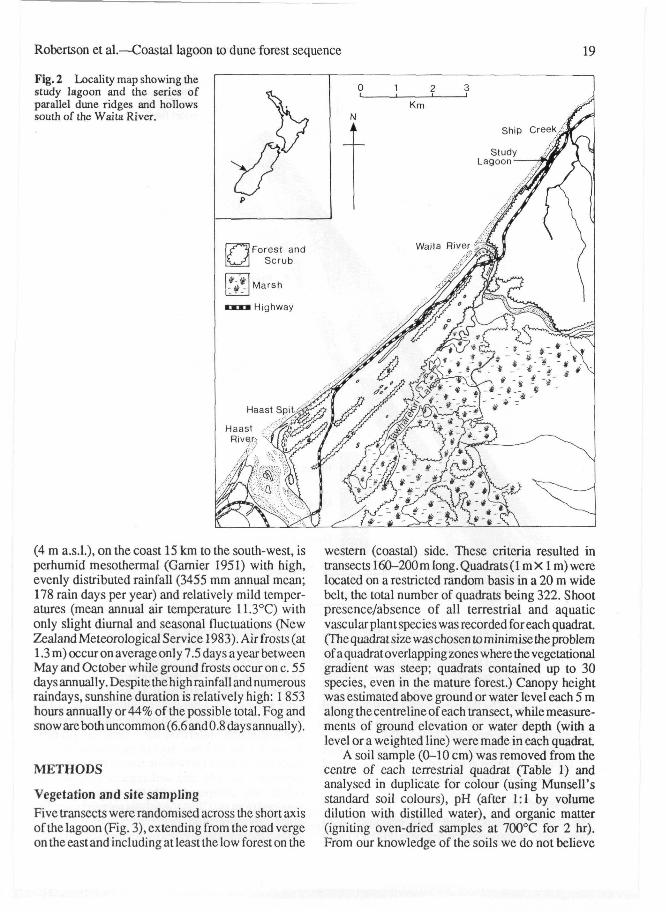
The lagoon studied (Fig. 1) is between old duneswithin c. 10 m elevation and 100 m distance of thecoast (Fig. 2), lying in a depression bounded on theseaward side by an active dune front and on theinland side by a stabilised dune crest. Locatedbetween the Waita River and Ship Creek (Fig. 2) thelagoon forms a typical unit of the long, narrowcoastal plain between the Arawata River in the southand the coastal hills north of the Waita River. Assuch.adetailed study of the vegetation andassociated

18 New Zealand Journal of Botany, 1991, Vol. 29
Fig. 1 View south-west from the north-eastern end of the lagoon showing the vegetation zonation pattern. Reed bedsof Eleocharis sphacelata stand in moderately deep open water and adjoin a zone of stunted Leptocarpus similis (jointedrush) which in rum merges with stunted Leptospermum scoparium (manuka) that increases in height with progressiontowards a low forest dominated by Dacrydium cupressinwn (rimu) and Weinmannia racemosa (kamahi). The positionfrom which the photo was taken is indicated in Fig. 3.
environmental factors seemed relevant to a fullerunderstanding of patterns andprocesses in and aroundthe lagoon. Mutch & McKellar (1975) characterisethe coastal plain as having "steep-sided, dome-shaped, ice-smoothed hills and coastal swampsfronted by active and fixed sand dunes and broadgravel and sand beaches". The beach deposits arederived from marine and fluvial material of veryrecent origin, certainly < 5000 yr. At the northernend of the lagoon is an outcrop corresponding to thesouthern-most extension of a much oldertransgressive series of marine and alluvial depositsdating back to the Paleocene (Mutch & McKellar1975).
The lagoon, c. 1.5 km X 0.1 km, is elongatedparallel to the shoreline with only a narrow vegetateddune separating it from the beach proper. The beachhas a steep small-bouldered foreshore topped with abroad dune ridge and with a sandy backslope
colonised largely by Desmoschoenus spiralis*(pingao) that is bounded by aPhormium tenax (NewZealand flax) tall-graminoid community which inturn grades into podocarp-broadleaved dune forest.The presence of tracks indicated occasional entry bycattle, though none were seen and there was no signof grazing impact. The canopy plants are obviouslywind-shorn with a general pattern similar to thatdescribed at Cole Creek c. 4 km northwards bySykes & Wilson (1991). The parallel series of duneridges and wet inter-dune hollows is particularlywell developed immediately south of the WaitaRiver.
The climate of the coastal plain area, ascharacterised by long-term records from Haast
"For pteridophytes nomenclature follows Brownsey &Smith-Dodsworth (1989), for other vascular plantsConnor & Edgar (1987), and references therein.
Robertson et al.—Coastal lagoon to dune forest sequence 19
Fig. 2 Locality map showing thestudy lagoon and the series ofparallel dune ridges and hollowssouth of the Waita River.
(4 m a.s.l.), on the coast 15 km to the south-west, isperhumid mesothermal (Gamier 1951) with high,evenly distributed rainfall (3455 mm annual mean;178 rain days per year) and relatively mild temper-atures (mean annual air temperature 11.3°C) withonly slight diurnal and seasonal fluctuations (NewZealand Meteorological Service 1983). Air frosts (at1.3 m) occur on average only 7.5 days a year betweenMay and October while ground frosts occur on c. 55days annually. Despite the high rainfall and numerousraindays, sunshine duration is relatively high: 1 853hours annually or 44% of the possible total. Fog andsnow are both uncommon (6.6 and 0.8 day s annually).
METHODS
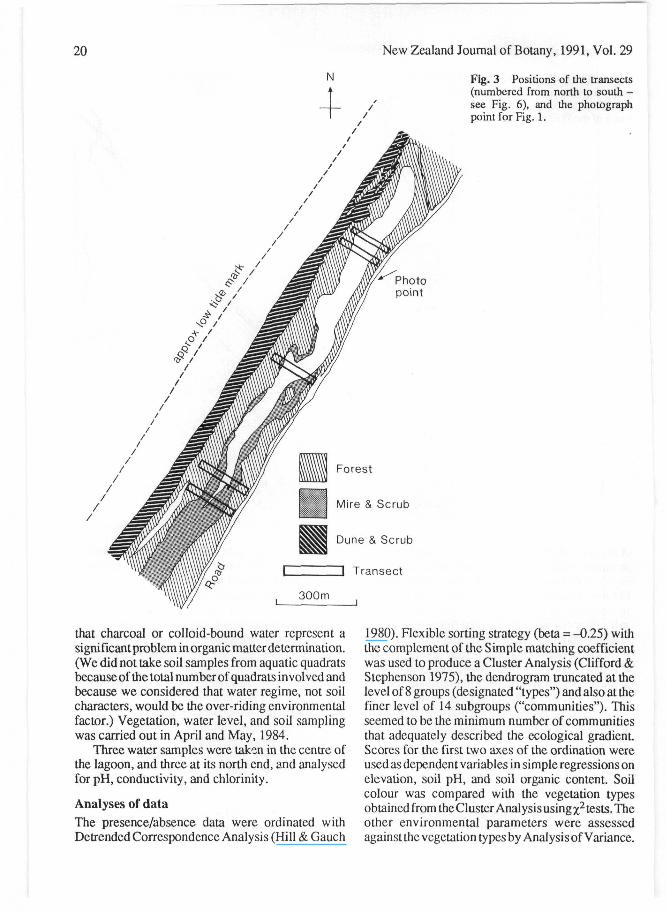
Vegetation and site samplingFive transects were randomised across the short axisof the lagoon (Fig. 3), extending from the road vergeon the east and including at least the low forest on the
western (coastal) side. These criteria resulted intransects 160-200 m long. Quadrats (1 m X1 m) werelocated on a restricted random basis in a 20 m widebelt, the total number of quadrats being 322. Shootpresence/absence of all terrestrial and aquaticvascular plant species was recorded for each quadrat.(The quadrat size was chosen to minimise the problemof aquadratoverlappingzones where the vegetationalgradient was steep; quadrats contained up to 30species, even in the mature forest.) Canopy heightwas estimated above ground or water level each 5 malong the centreline of each transect, while measure-ments of ground elevation or water depth (with alevel or a weighted line) were made in each quadrat.
A soil sample (0-10 cm) was removed from thecentre of each terrestrial quadrat (Table 1) andanalysed in duplicate for colour (using Munsell'sstandard soil colours), pH (after 1:1 by volumedilution with distilled water), and organic matter(igniting oven-dried samples at 700°C for 2 hr).From our knowledge of the soils we do not believe
20 New Zealand Journal of Botany, 1991, Vol. 29
Fig. 3 Positions of the transects(numbered from north to south -see Fig. 6), and the photographpoint for Fig. 1.
Forest
Mire & Scrub
Dune & Scrub
] Transect
that charcoal or colloid-bound water represent asignificantproblem in organic matter determination.(We did not take soil samples from aquatic quadratsbecause of the total number of quadrats involved andbecause we considered that water regime, not soilcharacters, would be the over-riding environmentalfactor.) Vegetation, water level, and soil samplingwas carried out in April and May, 1984.
Three water samples were taken in the centre ofthe lagoon, and three at its north end, and analysedfor pH, conductivity, and chlorinity.
Analyses of dataThe presence/absence data were ordinated withDetrended Correspondence Analysis (Hill & Gauch
1980). Flexible sorting strategy (beta = -0.25) withthe complement of the Simple matching coefficientwas used to produce a Cluster Analysis (Clifford &Stcphenson 1975), the dendrogram truncated at thelevel of 8 groups (designated "types") and also at thefiner level of 14 subgroups ("communities"). Thisseemed to be the minimum number of communitiesthat adequately described the ecological gradient.Scores for the first two axes of the ordination wereused as dependent variables in simple regressions onelevation, soil pH, and soil organic content. Soilcolour was compared with the vegetation typesobtained from the Cluster Analysisusingx2tests. Theother environmental parameters were assessedagainst the vegetation types by Analysis of Variance.
Robertson et al.—Coastal lagoon to dune forest sequence 21
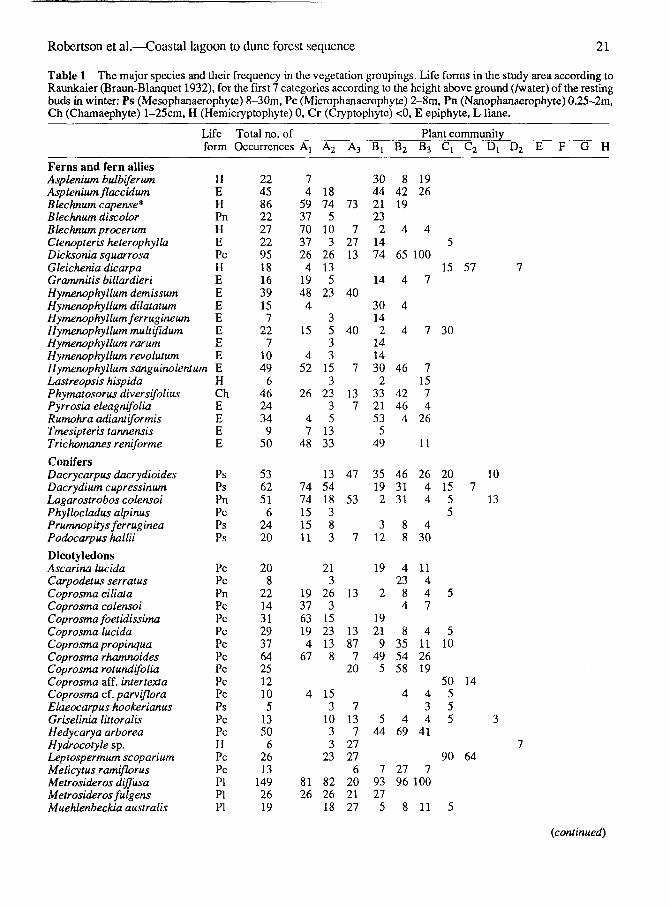
Table 1 The major species and their frequency in the vegetation groupings. Life forms in the study area according toRaunkaier (Braun-Blanquet 1932), for the first 7 categories according to the height above ground (/water) of the restingbuds in winter: Ps (Mesophanaerophyte) 8-30m, Pc (Microphanaerophyte) 2-8m, Pn (Nanophanaerophyte) 0.25-2m,Ch (Chamaephyte) l-25cm, H (Hemicryptophyte) 0, Cr (Cryptophyte) <0, E epiphyte, L liane.
Life Total no. of Plant communityform Occurrences Aj A2 A3 Bj B2 B3 Cl C2 Dl D2 E F G H
Ferns and fern alliesAsplenium bulbiferumAsp lenium flaccidumBlechnum capense*Blechnum discolorBlechnum procerumCtenopteris heterophyllaDicksonia squarrosaGleichenia dicarpaGrammitis billardieriHymenophyllum demissumHymenophyllum dilatatumHymenophyllum ferrugineumHymenophyllum multifidumHymenophyllum rarumHymenophyllum revolutumHymenophyllum sanguinolentumLastreopsis hispidaPhymatosorus diversifoliusPyrrosia eleagnifoliaRumohra adiantiformisTmesipteris tannensisTrichomanes reniforme
ConifersDacrycarpus dacrydioidesDacrydium cupressinumLagarostrobos colensoiPhyllocladus alpinusPrumnopitys ferrug ineaPodocarpus hallii
DicotyledonsAscarina lucidaCarpodetus serratusCoprosma ciliataCoprosma colensoiCoprosma foetidissimaCoprosma lucidaCoprosma propinquaCoprosma rhamnoidesCoprosma rotundifoliaCoprosma aff. intertextaCoprosma cf. parvifloraElaeocarpus hookerianusGriselinia littoralisHedycarya arboreaHydrocotyle sp.Leptospermum scopariumMelicytus ramiflorusMetrosideros diffusaMetrosideros fulgensMuehlenbeckia australis
HEHPnHEPcHEEEEEEEEHChEEEE
PsPsPnPcPsPs
PcPcPnPcPcPcPcPcPcPcPcPsPcPcHPcPcPIPIPI
22458622272295181639157
227
1049
64624349
50
536251
62420
208
2214312937642512105
1350
62613
1492619
74
59377037264
19484
15
452
26
47
48
7474151511
193763194
67
4
8126
18745
103
26135
23
3533
153
2335
1333
135418383
213
263
1523138
153
1033
23
822618
73
72713
40
40
7
137
47
53
7
13
13877
20
7137
2727
6202127
304421232
1474
14
30142
141430
2332153
549
35192
312
19
2
1921
949
5
544
79327
5
84219
4
65
4
4
4
46
4246
4
463131
88
423
84
8355458
4
469
2796
8
1926
4
100
7
7
71574
26
11
2644
430
11447
4112619
434
41
7100
11
5
15 57
30
2015 755
5
510
50 14555
90 64
5
10
13
{continued)

22 New Zealand Journal of Botany, 1991, Vol. 29
Table 1 (continued)
Myriophyllwn pedunculatumMyriophyllum robustumMyrsine australisMyrsine divaricataNeomyrtus pedunculataNertera depressaNertera scapanioidesNertera cf. dichondrifoliaParsonsia heterophyllaPseudopanax colensoiPseudopanax crassifoliusPseudowintera colorataRubus australisWeinmannia racemosaUnidentified aquatic weed
MonocotyledonsAstelia grandisBaumea tenaxCarex virgataCentrolepis ciliataEarina autumnalisEleocharis sphacelataFreycinetia baueriana'Gahnia xanthocarpaLepidosperma australeLeptocarpus similisLuzuriaga parvifloraMicrolaena avenaceaPhormium tenaxPotamogeton cheesemaniiRipogonum scandensUncinia langustifoliaUncinia luncinata
Lifeform
CrCrPcPcPcChChChPIPcPcPcPIPsCr
HHHHECrPIHHHEHHCrPIHH
Total no. ofOccurrences
546194051165
47231127
69
14413
9121
877
715349152558
128
462018
Ai
15375215
634
1111
93
74
4
474
19
441
A2
51833
8
15
815
87
15
13
8715
3
563
A3
7
401313
7
7
32713
5
27
1353
27131357
Bi
122123
2
265
121255
95
15
2
3323
7
535
19
B2
19154615
5038
354
46
4
4
48
154
508
19
Plant communityB3
114
4
33
711
4
4
19
11
C'i
1530
55
510
5
15
5100
55
155
45
30
21
100
14
10029
Di D2
13
100 100
29
7
3 86
E F G
100
100
100100 91
43 9
H
44
100
86
3
*sensu Allan (1961)fssp. banksii
The Teddybear (Wilson 1975) multivariate packagewas used for both the regressions and analysis ofvariance.
RESULTS AND INTERPRETATION
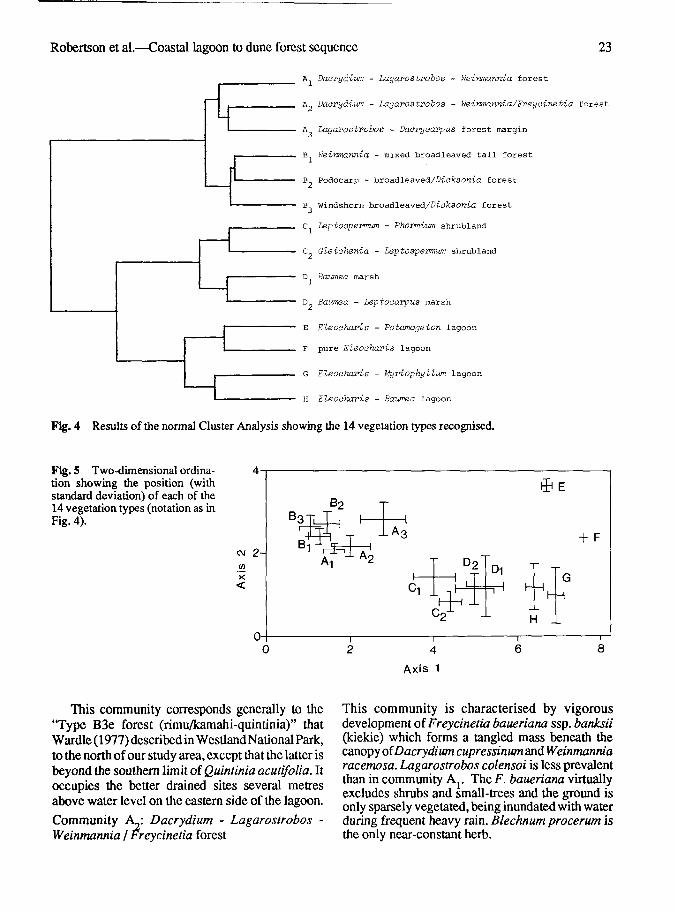
The vegetation types and communities derived fromCluster Analysis (Fig. 4, Table 1) are confirmed bythe ordination diagram (Fig. 5). The types aredescribed in sequence. Comparisons with Wardle's(1977) communities are made only when there werecomparable communities in his area.
Type A: podocarp - Weinmannia forestCommunity A}: Dacrydium - Lagarostrobos -Weinmannia forest
Dacrydium cupressinum (rimu) and Lagarostroboscolensoi (silver pine) along with Weinmanniaracemosa (kamahi), form a low forest c. 9 m tall, ofrelatively uniform diameter trees. A shrub layer ofCoprosma foetidissima, C. rhamnoides, andNeomyrtus pedunculata is associated with a herblayer in which Gahnia xanthocarpa, Blechnumcapense (sensu Allan 1961), B. procerum, and B.discolor (crown fern) are all conspicuous. Beneaththese is a ground cover of a fine-leaved Uncinialangustifolia,Nertera cf. dichondrifolia,Luzuriagaparviflora (lantern berry), Trichomanes reniforme(kidney fern) and Hymenophyllum demissum, with//. sanguinolentum a common epiphyte on podocarptrunks. Lianes of Metrosideros diffusa are presenton most trees with the larger leaved M.fulgens lessfrequent.
Robertson et al.—Coastal lagoon to dune forest sequence 23
A, Daovyd-ium - Lagavostvobos - Weinmannia forest
A? Dacrydium - Lagarostrobos - Weirmannia/Freycinetia forest
A Lagarostrobos - Daorycarpus forest margin
B. Weinmannia - mixed broadleaved tall forest
B Podocarp - broadleaved/D-icfcsonia forest
B Windshorn broadleaved/Diyfesonia forest
C Leptospermum - Phormium shrubland
C Gleiohenia - Leptospermum shrubland
D Banmea marsh
D Bawnea - Leptocarpus marsh
E Eleochari-s - Potamogeton lagoon
F pure Eleooharis lagoon
G Eleoohari-s - Myiriophyllum lagoon
H Eleocharis - Baumea lagoon
Fig. 4 Results of the normal Cluster Analysis showing the 14 vegetation types recognised.
Fig. 5 Two-dimensional ordina-tion showing the position (withstandard deviation) of each of the14 vegetation types (notation as inFig. 4).
Axis 1
This community corresponds generally to the"Type B3e forest (rimu/kamahi-quintinia)" thatWardle (1977) described in Westland National Park,to the north of our study area, except that the latter isbeyond the southern limit of Quintinia acutifolia. Itoccupies the better drained sites several metresabove water level on the eastern side of the lagoon.
Community A •. Dacrydium - Lagarostrobos -Weinmannia / Freycinetia forest
This community is characterised by vigorousdevelopment of Freycinetia baueriana ssp. banksii(kiekie) which forms a tangled mass beneath thecanopy of Dacrydium cupressinum and Weinmanniaracemosa. Lagarostrobos colensoi is less prevalentthan in community Ay The F. baueriana virtuallyexcludes shrubs and small-trees and the ground isonly sparsely vegetated, being inundated with waterduring frequent heavy rain. Blechnum procerum isthe only near-constant herb.

24 New Zealand Journal of Botany, 1991, Vol. 29
Stands occupy the small terrace adjacent to thelagoon and close to its water level. Largely restrictedto the lagoon's eastern side, they are exposed toonshore winds and are windshom. The canopy isusually somewhat lower than in community A r
Community Ag: Lagarostrobos -Dacrycarpus forestmarginRestricted to the forest edge on both sides of thelagoon, this community is characterised by immatureLagarostrobos co lensoi and saplings oWacrycarpusdacrydioides (kahikatea). Weinmannia racemosa isless common than in other types and Dacrydiumcupressinum is absent. It corresponds to Wardle's(1977) type Bl forest (open kahikatea foresttransitional to swamps). Shrubs are common,especially Coprosma propinqua and Myrsinedivaricata, while the large herbs Gahniaxanthocarpaand Blechnum procerum are also conspicuous.Leptospermum scoparium (manuka) occurs near themargin of the lagoon together with Carex virgataand occasional Phormiumtenax. Ground covervarieswith the amount of free water; Hymenophyllumdemissum is usually present but Nertera cf.dichondrifolia is rarely so.Themorelight-demandingH.multifidumTeplacesH.sanguinolentumoftiieA^community as the most frequent epiphyte.
Type B: Hedycarya - Weinmannia I DicksoniaforestCommunity By Weinmannia - mixed broadleavedtall forestWeinmannia racemosa is the main canopy dominantat c. 8 m in this community, with only sparsepodocarps - Dacrydium cupressinum andDacrycarpus dacrydioides. Subcanopy speciesinclude Dicksonia squarrosa and the broadleavedAscarina lucida, Hedycarya arborea (pigeonwood),and Coprosma lucida. Coprosma rhamnoides, C.foetidissima, Myrsine divaricata, and Neomyrtuspedunculata predominate among the shrubs withoccasional Freycinetia baueriana and Gahniaxanthocarpa in the herb layer. Rumohraadiantiformis and Trichomanes reniforme arecommon ground ferns while Asplenium flaccidumand several filmy ferns (Hymenophyllum spp.) areepiphytic. Lianes of Metrosideros diffusa andRipogonum scandens (supplejack) are abundant.
This community occupies the sloping banks onthe lagoon's eastern side that are better drained thanthe flatter terraces above and below it, or as tallerdune forest on the coastal side of the lagoon.Community B2: podocarp - broadleaved I DicksoniaforestThis community differs from the Bj community in
its dominant species rather than its physiognomy.Hedycarya arborea is common in the canopy,Weinmannia racemosa is less common, andDacrycarpusdacrydioides, Dacrydiumcupressinum,and Lagarostrobos colensoi are all of minor butconsistent occurrence. Dicksonia squarrosa is againcommon in the subcanopy stratum but Carpodetusserratus and Melicytus ramiflorus may occur ascanopy or subcanopy species. Shrubs of Coprosmarotundifolia are characteristic with less of C.rhamnoides, C. propinqua, or Neomyrtuspedunculata. Hymenophyllum sanguinolentum iscommon as an epiphyte with Phymatosorusdiver sifolius and Pyrrosia eleagnifolia as climbers.Other ferns found in the Bj community are largelyabsent. Ground cover is composed of Nertera cf.dichondrifolia with occasionalMicrolaenaavenacea(bush rice grass) and Uncinia spp. Lianes (Ripogonumscandens, Metrosideros diffusa, and less Parsonsiaheterophylla) are again abundant.
This community is restricted to the lagoon'scoastal side in the forest zone least exposed to salt-laden winds and sufficiently elevated to avoidwaterlogged soils.
Community B3: windshorn broadleaved I DicksoniaforestThis community, occurring near the foreshore marginof the coastal forest, is depressed and sculptured bysalt-spray. It includes the podocarps Podocarpushallii (thin-barked totara) and Dacrycarpusdacrydioides as well as Hedycarya arborea.Weinmannia racemosa is absent. Dicksoniasquarrosa alone forms the sub-canopy layer with theshrub species of Coprosma listed for the B,community much icsscommon.Lastreopsis hispidais a common ground fern while other ferns are moreminor than in communities B, and B,. Lianes ofMuehlenbeckia australis, Lycopodium volubile,Ripogonum scandens, and Parsonsia heterophyllaare frequent. This community is relatively poorfloristically, perhaps due to occasional grazing bycattle.
Type C: Leptospermum shrublandCommunity Cji Leptospermum - PhormiumshrublandA 2 m tall Leptospermum scoparium shrubland withoccasional co-dominant Phormium tenax and twocommon graminoid subdominants, Baumea tenaxand Leptocarpus similis (jointed rush), plusCoprosma cf. intertexta, is located alongside andgrades into the margin of forest community AJ. This

Robertson et al.—Coastal lagoon to dune forest sequence 25
shrubland contains occasional seedlings (< 30 cm)oiWeinmannia racemosa.Dacrydium cupressinum,and Dacrycarpus dacrydioides.
Community C2: Gleichenia - Leptospermumshrubland.Beyond the Leptospermum - Phormium shrubland,L. scoparium becomes more sporadic and smaller (c.1 m) and is commonly associated with Gleicheniadicarpa (tangle fern). Three graminoids are alsopresent, with Lepidosperma australe and Baumeatenax more common than Leptocarpus similis.
Type D: Baumea - Leptocarpus marshCommunity Dji Baumea marshIncreasingly extensive areas of almost pure Baumeatenax occur towards the southern end of the lagoon.Community D2: Baumea - Leptocarpus marshThis community is generally similar to communityDj but with the addition of Leptocarpus similis. Thesmall graminoid cushion Centrolepis ciliata isfrequently present. This community occurs in isolatedpatches among the Baumea marsh community andseems to be associated with slightly shallower water.
Type (Community) E: Eleocharis - PotamogetonlagoonThis community was found towards the northernend of the lagoon and consists of Eleocharissphacelata, frequent plants of Potamogetoncheesemanii, and an unidentified aquatic macrophyte.
Type (Community) F: pure Eleocharis lagoonOccasional sites throughout the lagoon contain purestands of E. sphacelata.
Type (Community) G: Eleocharis -Myriophyllum lagoonIn the central parts of the lagoon, in deep water, E.sphacelata and Myriophyllum spp. are accompaniedby occasional plants of Potamogeton cheesemanii.
Type (Community) H: Eleocharis - BaumealagoonIn water more shallow than that associated with theEleocharis -Potamogeton community, mostly at thesouthern end of the lagoon, E. sphacelata and B. tenaxco-dominate. Myriophyllum robustum is lessfrequent, occurring in about half the quadrats.
ZonationResults of three of the transects are summarised asprofile diagrams (Fig. 6) which also show thedistribution of the 14 communities.
In the ordination (Fig. 5), Axis 1 accounts formost of the variation in species composition and
reveals a gradient of increasing wetness withincreasing ordination score. Axis 2 is less obviouslyrelated to the communities.
Correlations with environmental parametersMean values for four environmental variables amongthe plant communities previously described areshown in Table 2, together with results of Duncan'sNew Multiple Range Tests.
ElevationBoth analysis of variance and Duncan's test (Table2) indicate that elevation is a significant factor indifferentiating many of the 14 communitiesrecognised. Moreover, a regression of mean elevationof a quadrat against its ordination scores showed astrong relationship with Axis 1 scores for quadratsless than 2 m above the lagoon surface (negative; R2
= 0.75; n = 281; P < 0.001). The importance of water(depth or drainage) is again implied in determiningthe vegetation pattern, though other effects ofelevation, e.g., on salt spray, may also beimportant.
There was no significant relation betweenelevation of quadrats more than 2 m above thelagoon and ordination values on either axis. A weakerrelation occurred between quadrat elevation andAxis2(positive;fl2=0.13;«=281;P<0.001),againexcluding those quadrats more than 2 m above thelagoon.
Soil organic contentThere were significant differences in the organiccontent (% of dry weight) of soil associated with thefive forest types that occur above the water level(Table 2).
The lower values in the three B-type communitiesare related to their location on sandier and steepersites than the more or less permanently waterloggedsites that characterise A-type forest. Significantregressions were found between soil organic contentand ordination scores on both Axis 1 (positive; R2 =0.28; n = 141; P < 0.001) and Axis 2 (negative;^?2 =0.25; n= 141; P< 0.001).
Soil aciditySoil pH varied significantly with vegetation, the A-type forest associations being more acidic than theB-type (Table 2). Only Axis 2 of the ordination,however, relates to pH (positive; R2 = 0.28; n = 141;P < 0.001).Soil colourX2 tests showed significant differences in soil colourbetween communities (Table 2), confirmed for bothaxes of the ordination with ANOVA.

26 New Zealand Journal of Botany, 1991, Vol. 29
Water analysisThere was no indication of a difference betweenwater samples taken from the two different parts ofthe lagoon. Overall means are therefore presented(Table 3).
Species richnessDefined as the number of species per quadrat, this isa simple but meaningful measure of species diversity.
It showed overall a significant decrease (negative;R2 = 0.77; n = 322; P < 0.001) towards the positive(dry) end of ordination Axis 1.
CONCLUSIONS
Ordination Axis 1 has been shown to represent thepattern of the dune-hollow sequence which is
Table 2 Mean values for four environmental parameters associated with the 14 plant associationsrecognised. Elevation is shown in relation to the lagoon surface. Letters shared within each factor indicateno significant difference with Duncan's NewMultiple Range Tests atP < 0.05. For soil colour, "brownish-black" was the most frequent in all communities and the "characteristic soil colour" indicates the nextmost frequent.
CommunityNo. of Elevation Soil organic Soil Characteristic
quadrats (cm) content (%) pH soil colour
A Dacrydium -Lagarostrobos -Weinmannia forest
A Dacrydium -Lagarostrobos -Weinmannia /
Freycinetia forest
A Lagarostrobos -Dacrycarpusforest margin
B Weinmannia -mixed broadleavedtall forest
B_ Podocarp - broadleaved /Dicksonia forest
B Windshom broadleaved /Dicksonia forest
C^Leplospermum -rhormium shrubland
28
35
15
43
27
25
20
C Gleichenia - Leptospermumshrubland 14
+473 a
+106 b
+llcde
+129 b
+42 cd
+41 c
-26
-16
30.2 b
56.0 a
23.8 b
9.7 c
12.1c
def
cdef
Dj Baumea marsh
D Baumea - Leptocarpus
28
-29 def
lagoonF pure Eleocharis lagoon
G Eleocharis -Myriophyllum lagoon
H Eleocharis - Baumealagoon
711
21
34
-68 efg-148 g
-143 g
-78 fg
4.09 a olive-black
4.30 b black
4.47 be black/dark brown
4.86 d olive-black
4.67 cd dark olive brown
Fig. 6 Profile diagrams for three of the belt transects sampled across the lagoon. Vegetation types (notation as in Fig. 4)and important species (dominant or frequent) are shown. All profiles are drawn with the inland side to the left.

Robertson et al.—Coastal lagoon to dune forest sequence 27
Profile 2
50 100D I S T A N C E - m
1 5 0
Weinmanniaracemoaa
Shrubs of *-3jC
/Voprosma f) sft^V spp. \ y Leptospermum /
\ Neomyrtus 1 BCOpariUm / Dicksonia
fl LagarostroboIcolenaoi Y1- Gahnia —
I xatithocarpa xy]
' Baumeatenax
' Dacrydium cupres
/ i'>|,'';'('Leptocarpus
F r . y c in . t i . '•"" = i " i l i S
DacrycarpusJ,acrydioides

28 New Zealand Journal of Botany, 1991, Vol. 29
correlated with site elevation, water depth and soilorganic content. For purely terrestial sites, this willrepresent soil water availability; for aquatic sites itwill represent depth of water, possibly acting vialight limitation on the bed; for intermediate sites itrepresents the frequency and duration ofwaterlogging. Sites at higher elevation will also bemore subject to salt spray.
The zonation we recorded suggests thatsuccession within the lagoon starts with establishmentof abedof tall emergcntEleocharis sphacelata. Thiscontributes organic matter while probably trappingwind-blown sand and other material. The substrateis gradually built up, we suggest, until at a waterdepth of c. 1 m (Table 2) the site comes within thetolerance range of Baumea tenax. Later, withcontinued accumulation of substrate, Leptocarpussimilis and Lepidosperma australe invade, then, inapproximate order, Leptospermum scoparium,Gleicheniadicarpa.Phormiumtenax.andBlechnumprocerum. Pedestals of turf build up around thebases of L. scoparium and P. tenax plants andprovide sites for seedling establishment of foresttree species such as Lagarostrobos colensoi,DacrycarpusdacrydioidesandPhyllocladusalpinus.These species also dominate the first forest stand onthe dunes.
Examination of species richness indicates thatthe deepest water is colonised by a few dominantspecies. These are replaced by an increasing numberof species with narrower alpha niches as water depthdecreases. As species diversity increases so alsodoes plant stature and presumably also biomass.
The suggested developmental sequence isexpressed as concentric zones around the lagoon.Development appears to have occurred fastest at thesouthern end of the lagoon, where water enters. HereLeptospermum shrubland (Type C) and Baumea-Leptocarpus marsh (D) are well established, whilesmall clumps of trees, particularly of Lagarostroboscolensoi and Dacrycarpus dacrydioides, up to2-3 m tall are common. The northern end shows theleast development with small patches oiEleocharissphacelata only 2-3 m wide bordering large expansesof open water.
Table 3 Analysis of lagoon water.
mean s.e. of mean
pHConductivity (m SrrT1)Chlorinity (mg H Cl)
5.63.8813.5
0.150.174
0.13
Development of the wetland sequence proceedsthrough a pakihi-likephase (community types C andD) with occasional small forest trees. ("Pakihi" is aregional name for an oligotrophic mire amid forest:Mew 1983.) The fate of this community is indicatedby forest stands, mostly of communities A^ and A,,on the flat areas adjacent to the marsh vegetation. Anarrow fringe oiGahniaxanthocarpa andBlechnumprocerum, characteristic of theA^ommunity.formsthe initial understorey but as the podocarps increasein size Weinmannia racemosa and Dacrydiumcupressinum become established. Freycinetiabaueriana largely replaces most other subcanopyspecies and forest community A^ apparently persistsfor an extended period, though a broadleaved forest(Community type B) may substitute for the mixedpodocarp-Weinmannia type (A).
Results of waterlogging experiments (Robertson,unpublished) indicated that most local species arelimited less by decreased waterlogging than bydecreased light levels under the developing forestcanopy. Absenc&ofDacrycarpus dacrydioides fromthe sequence possibly is due to low soil fertility(Wardle 1974).
Vegetation sequences similar to those describedhere have received some attention from Cockayne(1928), and most recently from Wardle (1980) andNorton (1989). Wardle examined a range of primarysuccessions in Westland, including a sequence verysimilar to that described here. The mire-forestsequence described by Norton from SaltwaterEcological Area, 130 km to the north, bears someresemblance to that on the lagoon margin, but thesubstrates and the pH differ and the Saltwatersequence is considered to be essentially static (Nortonop. cit).
The dune ridges obviously support a differentdevelopment sequence. Soils here derive from sandand are well-drained with the crest freely exposed tosalt-laden winds. The vegetation on such sites,towards the top end of ordination Axis 2, is abroadleaved forest community, predominantly ofWeinmannia racemosa and Hedycarya arborea witha Dicksonia squarrosa understorey. Such duneforest was not present in or near Westland NationalPark, some 80 km to the north, presumably becauseof human disturbance (Wardle 1977). Observationsindicate that podocarps become more prominent onthe older dunes further from the coast, beyond thestudy area. We suggest this follows podzolisationand associated impeded drainage. Eventually a forestdevelops, dominated by Dacrydium cupressinum.Decreasing scores on Axis 2 are correlated with

Robertson et al.—Coastal lagoon to dune forest sequence 29
decreasing pH and increasing organic content, bothtrends associated with soil development (Smith &Lee 1984). Soil colour also changes, with olive-brown soils of the early stages darkening to black asdevelopment continues. Several generations of mor-forming forest species presumably would beinvolved.
There is evidence that the vegetation sequencedescribed represents two separate processes ofvegetational development occurring in the vicinityof the lagoon. Establishment of successive dunecrests and their associated hollows has initiatedforest development on the well-drained crests, andhydrarch succession in the hollows. While the twotemporal sequences are somewhat convergent thereis no indication that full convergence is ever achieved.
The ecological, conservation, and scenic valuesof the lagoon plus its proximity to a main highway alljustify the area being given formal protection. Thecaseforpreservation is greatly enhancedby the presenceof the aquatic herb Myriophyllum robustum, listed asvulnerableby Given (1981). Williams & Given (1981)contend that its future survival outside Fiordlandseems doubtful, and report that attempts to relocatepreviously reported wild populations elsewhere havebeen largely unsuccessful. Orchard (1979) considersthat the combination of draining and burning of thespecies' habitat will result in its extinction unlessconservation measures are taken soon. The species iscommon in this lagoon. Large areas of open waterremain available for it to inhabit and it would seem thatsurvival here could be ensured with the site protected.Formal protection is now assured since the area isincluded in the South-west New Zealand WorldHeritage area recently approved by the InternationalUnion for the Conservation of Nature and NaturalResources (IUCN). The actual designation of thewetland area has not yet been decided.
ACKNOWLEDGMENTS
The authors thank Sharon Quinn and Murray Grant forfield assistance and the Zoology Department, Universityof Otago for loaning levelling equipment and a dinghy. D.Kelly and two anonymous referees provided usefulcomments on an earlier version.
REFERENCES
Allan, H. H. 1961. Flora of New Zealand. Vol. I.Wellington, Government Printer,
Braun-Blanquet, J. 1932: Plant sociology: the study ofplant communities. Fuller and Conard.
Brownsey, P. J.; Smith-Dodsworth, J. C. 1989: NewZealand ferns and allied plants. Auckland,Bateman.
Clifford, H. T.; Stephenson, W. 1975: An introduction tonumerical classification. New York, AcademicPress.
Cockayne, L. 1928: The vegetation of New Zealand. Ed.2. Leipzig, Engelmann.
Connor, H. E.; Edgar, E. 1987: Name changes in theindigenous New Zealand flora, 1960-1986 andnominanovalV, 19S3-19&6.NewZealandjournalof botany 25: 115-170.
Dawson, J. 1988: Forest vines to snow tussocks: the storyof New Zealand plants. Wellington, VictoriaUniversity Press.
Gamier, B. J. 1951: Thomthwaite 's new system of climateclassification in its application to New Zealand.Transactions oft he Royal Society of New Zealand79: 87-103.
Given, D. R. 1981: Rare and endangered plants of NewZealand. Wellington, Reed.
Hill,M.O.;Gauch,H.G. 1980: Detrended correspondenceanalysis: an improved ordination technique.Vegetatio42: 47-58.
Mark, A. F.; Smith, P. M. F. 1975: A lowland vegetationsequence in south Westland: pakihi bog to mixedbeech-podocarp forest. Part 1: the principal strata.Proceedings of the New Zealand EcologicalSociety 22: 76-92.
Mew, G. 1983: Application of the term "pakihi" in NewZealand - a review. Journal of the Royal Societyof New Zealand 13: 175-198.
Mutch, A. R.; McKellar, I. C. 1975: Sheet 19 Haast (Ed.1) Geological Map of New Zealand 1:250 000.,Wellington, New Zealand Department of Scientificand Industrial Research.
New Zealand Meteorological Service. 1983: Summariesof climatological observations to 1980. NewZealand Meteorological Service miscellaneouspublication 177. Wellington, Government Printer.
Norton, D. A. 1989: Floristics and structure of mire-foresteco tones, West Coast, Southlsland, New Zealand.Journal of the Royal Society of New Zealand 19:31-42.
Orchard, A. E. 1979: Myriophyllum (Haloragaceae) inAustralasia 1. New Zealand: a revision of thegenus and synopsis of the family. Brunonia 2:247-287.
Smith, S. M.; Lee, W. G. 1984: Vegetation and soildevelopment on a Holocene river terrace sequence,Arawata Valley, South Westland, New Zealand.New Zealand journal of science 27: 187-196.
Sykes, M. T.; Wilson, J. B. 1991: Vegetation patterns ondunes of southern New Zealand. Submitted tolournal of Vegetation Science.

30 New Zealand Journal of Botany, 1991, Vol. 29
Wardle, P. 1974: The kahikatea (Dacrycarpus 1980: Primary succession in Westland National Parkdacrydioides) forest of South Westland. and its vicinity, New Zealand. NewZealandjournalProceedingsoftheNewZealandEcologicalSociety of botany 18: 221-332.21: 62-71. Williams, G. R.; Given, D. R. 1981: The red data book of
1977: Plant communities of WestlandNational Park New Zealand. Wellington, Nature Conservation(New Zealand) and neighbouring lowland and Council.coastal areas. New Zealand journal of botany 15: Wilson, J. B. 1975: Teddybear - a statistical system. New323-398. Zealand statistician 10 (3): 36-42.




















