






Bahasa
Halaman
Hukum


Potential and Opportunities for Use of Recombinant
Lactic Acid Bacteria in Human Health
SEAN HANNIFFY,* URSULA WIEDERMANN,{ ANDREAS REPA, {
ANNICK MERCENIER,{ CATHERINE DANIEL,{ JEAN FIORAMONTI,}
HELENA TLASKOLOVA,k HANA KOZAKOVA, k HANS ISRAELSEN, { SØREN MADSEN,{
ASTRID VRANG,{ PASCAL HOLS,# JEAN DELCOUR,# PETER BRON,**MICHIEL KLEEREBEZEM,** AND JERRY WELLS*, {{
*Institute of Food Research,Norwich Research Park, Colney,
Norwich, NR4 7UA, United Kingdom{University of Vienna, Department of Pathophysiology
A-1090 Vienna, Austria{Institut Pasteur de Lille,
Department of Microbiology of EcosystemsF59019 Lille, France
}Neurogastroenterology and Nutrition Unit, INRA,F31931 Toulouse 9, FrancekInstitute of Microbiology,
Department of Immunology and GnotobiologyAcademy of Sciences of the Czech Republic
142 20 Prague 4, Czech Republic{Bioneer A/S, DK-2970 Horsholm, Denmark
#Universite Catholique de Louvain, Unite de GenetiqueB1348 Louvain-la-Neuve, Belgium
**Wageningen Centre for Food Sciences—NIZO Food Research6710 BA Ede, The Netherlands
{{Author for correspondence. E-mail: [email protected]
I. Introduction 2
II. Use of Recombinant LAB to Prevent Infectious Diseases 3
A. Recombinant LAB as Vaccine Delivery Vehicles 4
B. Infections of the Respiratory Tract 7
C. Infections of the Gastrointestinal Tract 9
D. Infections of the Urogenital Tract 11
III. Potential for Immune Modulation of Type I Allergy
Using Recombinant LAB 14
A. Role of the Indigenous Microflora 15
B. Animal Model of Type I Allergy 16
C. Use of LAB for Prophylaxis and Therapy of Type I Allergy 17
IV. Opportunities for the Treatment of Inflammatory Bowel
Diseases Using Recombinant LAB 18
A. Role of LAB in Intestinal Barrier Function 19
1
ADVANCES IN APPLIED MICROBIOLOGY, VOLUME 56Copyright 2004, Elsevier Inc.
All rights reserved.0065-2164/04 $35.00

2 HANNIFFY et al.
B. LAB and Intestinal Inflammation 20
C. Recombinant LAB as a Therapeutic Option 22
V. LAB as Cell Factories for the Manufacturing of
Pharmaceutical Proteins 23
A. The Choice of Microbial Host 23
B. The Expression Vector 24
C. Propagation, Fermentation, and Initial Downstream Processing 26
VI. Engineering LAB for Their Safe Use in Humans 28
A. Food-Grade Systems in LAB for Plasmid Maintenance and
Chromosomal Insertion 28
B. Biological Containment Systems 32
VII. Opportunities and Potential Applications of Future Research 33
A. Insights from Genome Sequencing and Comparative Genomics 33
B. The Behavior of LAB in the Host 35
C. The Host Response to LAB 39
VIII. Concluding Remarks 43
References 45
I. Introduction
Dietary lactic acid bacteria (LAB) are mostly known for their wide-spread use in the production and preservation of fermented foods andas such have obtained the ‘‘generally regarded as safe’’ (GRAS) statuswithin the food industry (Adams and Marteau, 1995). Some membersof this diverse group of bacteria are components of the indigenous gutmicroflora of both animals and humans and have long been recognizedfor their health-promoting properties. Indeed, specific strains of LAB,and in particular lactobacilli, have been used as probiotics (Fuller,1989; Holzapfel et al., 1998; Isolauri et al., 2001) because they arethought to play a crucial role in maintaining a healthy microflora aswell as contributing to an expanding list of health-promoting activities(Mercenier et al., 2003). Probiotic LAB have been shown to be benefi-cial in the treatment of gastrointestinal disorders such as lactoseintolerance, travelers’ diarrhea, antibiotic-associated diarrhea, and in-fections caused by various bacterial and viral pathogens (Heyman,2000). Clinical trials and animal studies indicate that LAB may alsobe used to treat the symptoms of atopy and may prevent/reducethe development of allergy (Cross et al., 2001; Kalliomaki and Isolauri,2003; Kalliomaki et al., 2003; Lodinova-Zadnikova et al., 2003;Tlaskalova-Hogenova et al., 2002). Furthermore, evidence that LABplay a role in controlling intestinal microflora, restoring intestinal

USE OF RECOMBINANT LAB IN HUMAN HEALTH 3
barrier function, and alleviating inflammatory responses has led totheir proposed use for therapy and management of immunopathologi-cal disorders such as Crohn’s disease, ulcerative colitis, and pouchitis(Campieri and Gionchetti, 1999; Marteau et al., 2003; Shanahan, 2001).
Over the last few years, a coordinated effort involving several Euro-pean laboratories and combining a number of interdisciplinaryresearch strategies has set out to provide experimental evidence forthe efficacy of different prototype health products based on the muco-sal administration of recombinant LAB (http://www.labdel.eu.com).This consortium also aims to further advance technology for LABdelivery and the safe containment of genetically modified organismsin order to increase the range of potential applications for recombinantLAB and to accelerate their commercial development. In this review,the consortium discusses the potential and future opportunities for theuse of recombinant lactic acid bacteria in human health.
II. Use of Recombinant LAB to Prevent Infectious Diseases
Although vaccines against several major pathogens are in commonuse, morbidity and mortality from infectious disease remains a consid-erable burden worldwide. According to World Health Organization(WHO) estimates, infectious diseases caused 14 million deaths in2001, accounting for 26% of global mortality. This situation is steadilybecoming worse because of increasing microbial resistance to antimi-crobial drugs worldwide, especially Streptococcus pneumoniae, enter-ococci, and Gram-negative enteric pathogens (Hakenbeck et al., 1999;Kariuki and Hart, 2001; Threlfall, 2002; Walsh, 2000). Recent evidenceis also linking a growing number of infectious agents to an increasedrisk of cancer, blurring the traditional distinctions between chronicand communicable diseases. In addition, behavioral changes over thelast two decades have seen the emergence of new pathogens as well asthe re-emergence of ‘‘old’’ infectious diseases thought to be extinct (vanGinkel et al., 2000). Some of the existing vaccines are also not withoutproblems, and more-effective versions need to be developed. In some ofthe poorest countries, existing vaccines are unaffordable to those mostin need, a situation that might be helped by strategies to reduce the costof vaccination (e.g., through avoidance of the use of syringes andneedles). Orally administered vaccines are also an attractive goal indeveloped countries where combined childhood vaccination sche-dules generally administered via nonmucosal or percutaneous injec-tions are becoming increasingly more complex to formulate andcombine. The main advantage of mucosal immunization (e.g., oral,

4 HANNIFFY et al.
nasal, rectal) is the potential to elicit both mucosal and systemic im-mune responses that would enhance the efficacy of many vaccines. It isnot surprising therefore that the development of effective strategies forthe delivery of vaccine antigens to the mucosal tissues has receivedconsiderable attention over the past decade (Michalek et al., 1994;O’Hagan, 1994; Wells and Pozzi, 1997). The use of recombinant bacte-ria as carrier systems has received particular attention, most vectorsunder development being derived from attenuated pathogenic bacteria(Bumann et al., 2000; Gicquel, 1995; Levine et al., 1996; Michalek et al.,1994; Roberts et al., 1994; Sirard et al., 1999; Stahl et al., 1997). Inaddition to the potential to revert to virulence with associated risk ofinfection and a public opinion sensitive to the use of recombinantorganisms, variation in the immunogenicity of the different attenuatedstrains has constituted a major problem, and it has been difficult toreach the right balance between the level of attenuation (i.e., lack ofdisease symptoms) and immunogenicity (i.e., efficacy). An additionalconcern with the use of attenuated pathogens is that they may still besufficiently virulent to cause disease in infants, the elderly, or immu-nocompromised individuals. Therefore the use of noninvasive andnonpathogenic lactic acid bacteria as vehicles for mucosal deliveryof vaccine antigens and other therapeutic molecules is an attractiveconcept.
In addition to their application as vaccine vehicles (discussedbelow), lactic acid bacteria can be used to deliver anti-infectives orantimicrobial products in situ. An example is the use of recombinantLactobacillus strains expressing anti-idiotypic single-chain Fv anti-body to a Streptococcus mutans adhesin (SAI/II) in the prevention ofdental caries in a rat model (Kruger et al., 2002). Similarily, recombi-nant Streptococcus gordonii expressing the microbiocidal moleculeH6, which is an anti-idiotypic single chain antibody mimicking a yeastkiller toxin, was shown to demonstrate candidacidal activity in a ratmodel (Oggioni et al., 2001).
A. RECOMBINANT LAB AS VACCINE DELIVERY VEHICLES
Many lactic acid bacteria are used by the food industry and have ahistory of safe use or are classified as nonpathogenic. They can there-fore be given orally in relatively large doses without risk of potentialside effects such as those associated with live attenuated bacterialvectors (Medina and Guzman, 2001). In addition, many LAB strainsare acid resistant (Kociubinski et al., 1999) and adhere to mucosalepithelium (Morita et al., 2002), properties that may be beneficial for

USE OF RECOMBINANT LAB IN HUMAN HEALTH 5
their use as mucosal delivery vehicles. Expressing vaccine antigensand other therapeutic molecules in recombinant LAB would obviatethe need to purify the active component, reducing the overall produc-tion and delivery costs. Moreover, these bacteria would ideally begiven orally and thus would be amenable to large-scale vaccinationprograms in populations at risk. In addition to providing protection,LAB-based vaccines could also potentially elicit protective mucosalimmune responses against pathogens that are also present as compo-nents of the normal microflora in asymptomatic carriers, thus havingan impact on carriage rates.
While evidence suggests that LAB exhibit low intrinsic immunoge-nicity when administered by the mucosal route, there is no doubt thatspecific strains of LAB do possess immunoadjuvant properties and canenhance antigen-specific immune responses when administered incombination with antigen (Link-Amster et al., 1994). It is also clearthat the immunoadjuvant properties of LAB vary for different species,which has obvious implications for the selection of specific LAB asvaccine vehicles (Maassen et al., 2000; Repa et al., 2003). In addition, itis important to consider that not all LAB are commensals in humansand that specific strains may be associated with different mucosal sitesand environmental niches within the host (Wells and Mercenier,2003). While selection of an appropriate model species is thereforecritical, their inherent diversity can also be seen as an advantage. Theirability to survive at different mucosal surfaces may increase opportu-nities for use of recombinant LAB as vaccine vehicles against a widerrange of diseases. Similarly, differences in the immunomodulatorycapacities of different LAB create additional possibilities for tailoringthe choice of vehicle to meet the requirements for immunity orto modulate immune outcomes in the treatment of various immuno-pathological diseases. A theoretical model of the immunomodulatoryeffects of LAB is shown in Fig. 1.
Importantly, the last decade has seen the development of the genetictools necessary for expression of heterologous proteins in an increasingnumber of LAB (for reviews see de Vos, 1999a; Langella and Le Loir,1999; Nouaille et al., 2003; Pouwels et al., 2001; Reuter et al., 2003;Wells and Mercenier, 2003; Wells and Schofield, 1996). Expressionsystems were first developed in the model LAB strain Lactococcuslactis, a noninvasive, noncolonizing bacterium widely used to producecheese curds by the fermentation of milk. These included constitutiveand inducible promoters that allowed efficient high-level production ofantigen under various conditions (de Vos, 1999a; Israelsen et al.,1995a; Kuipers et al., 1997; Wells and Schofield, 1996; Wells et al.,

FIG. 1. Theoretical model for Th1-enhancing effects of antigen expressing lactic acid
bacteria on immune responses based on a previous illustration by Cross et al. (2001). The
production of pro-interferon cytokines IL-12 and IL-18 and interferon alpha (INF�) by the
interaction of certain strains of lactic acid bacteria with antigen presenting cells (APC)
(e.g., dendritic cells and macrophages) is supported by in vitro co-culture studies and
in vivo animal models. Cell wall components of the Gram-positive lactic acid bacteria
may induce pro-Th1 cytokine production through Toll-like receptors (TLRs) and
possibly also through other surface receptors. The interaction of LAB with other cell
types, including epithelial cells, may also influence the outcome of exposure to LAB and
prevent tissue inflammation.
6 HANNIFFY et al.
1993a). In more recent times, constitutive and regulated expres-sion systems, some of which have been adapted from Lc. lactis(Kleerebezem et al., 1997; Pavan et al., 2000), have also been usedto express heterologous proteins in other LAB, including species ofLactobacillus that have been shown to colonize different niches in bothanimals and humans (Chang et al., 2003; Grangette et al., 2001, 2002;Hols et al., 1997b; Maassen et al., 1999; Oliveira et al., 2003; Pouwelset al., 1996; Rush et al., 1997; Scheppler et al., 2002). These systemshave been further adapted such that the expressed protein can besecreted by LAB or anchored to the bacterial surface (Bernasconiet al., 2002; Dieye et al., 2001; Grangette et al., 2002; Kruger et al.,2002; Oliveira et al., 2003; Reveneau et al., 2002; Ribeiro et al., 2002;Wells et al., 1993b). These improvements have ensured that LAB canbe used to effectively deliver antigen to mucosal surfaces either inparticulate form or as soluble antigen secreted into the surroundinglumen (Norton et al., 1996). The development of efficient expressionsystems for LAB has enabled several vaccine antigens derived from avariety of mucosal pathogens to be successfully expressed in differentLAB (Mercenier et al., 2000; Seegers, 2002; Thole et al., 2000; Wellsand Mercenier, 2003; Wells et al., 1996).

USE OF RECOMBINANT LAB IN HUMAN HEALTH 7
Pioneering research carried out by several European laboratoriesdemonstrated that immunization with Lc. lactis and selected speciesof lactobacilli expressing the tetanus toxin fragment C (TTFC) antigenwere capable of eliciting antigen-specific secretory IgA and serumantibodies that were protective against lethal challenge with tetanustoxin (Grangette et al., 2001, 2002; Norton et al., 1997; Robinson et al.,1997; Wells et al., 1993a). In a continuation of this work, it was alsoshown that Lc. lactis could express and secrete the murine cytokinesIL-2 and IL-6, which enhanced antigen-specific immune responseswhen co-expressed with TTFC (Steidler et al., 1998). These experi-ments demonstrated that biologically active molecules such as cyto-kines, enzymes and other molecules could be effectively delivered tomucosal surfaces by using recombinant LAB. This application wasfurther exploited by Steidler et al. (2000), who showed that Lc. lactisstrains expressing and secreting murine IL-10 could be used to treatinflammation in two different mouse colitis models. This work hasnow led to the development of a biologically contained Lc. lactis strainsecreting human IL-10 (Steidler et al., 2003), which has been approvedby Dutch authorities for use in a small clinical trial as an experimentaltherapy for use in humans with inflammatory bowel diseases (IBD) (seealso Section IV). The aforementioned discoveries demonstrate thatthere is considerable potential to develop health-based products basedon oral delivery of vaccine and other therapeutics when using LAB.Several recent publications have now documented the use of prototypeLAB-based vaccines for production and delivery of various moleculestargeting a range of mucosal pathogens (Bermudez-Humaran et al.,2002, 2003; Enouf et al., 2001; Gil et al., 2001; Gilbert et al., 2000; Leeet al., 2001; Ribeiro et al., 2002; Xin et al., 2003; Zegers et al., 1999),some of which are discussed in the following sections.
B. INFECTIONS OF THE RESPIRATORY TRACT
As well as being a reservoir for potentially pathogenic bacteria suchas Haemophilus influenzae, Morexella catarrhalis, Pseudomonas aer-uginosa, Staphylococcus aureus, S. pneumoniae, and beta-hemolyticstreptococci, the respiratory tract is also susceptible to serious infec-tions caused by Bordetella pertussis, Mycobacterium tuberculosis,Mycoplasma pneumoniae, influenza virus, and respiratory syncitialvirus (RSV). All these organisms initiate disease at the mucosal surfaceof the respiratory tract, and thus the efficacy of the host’s responseto these infections is dependent on optimal local immune responsesat this site. However, vaccines available for diseases caused by many

8 HANNIFFY et al.
of these pathogens have limitations in accessibility or efficacy, high-lighting the need for improvements in approaches and products. Theneed for more-effective control strategies is heightened by reports ofincreasing antibiotic resistance among isolates from studies on carriagein healthy individuals and from clinical specimens (Metlay and Singer,2002).
Until recently there has been little evidence that probiotic LAB mighthelp prevent respiratory infections despite numerous studies showingtheir beneficial effects against gastrointestinal infections. However, anumber of studies have now shown that the oral application of variousLAB and/or their bioactive components can enhance the antimicrobialactivity of pulmonary natural killer cells (NK) and macrophages, in-crease IgAþ cells at the bronchial level, and induce cytokine produc-tion by nasal lymphocytes (Hori et al., 2002; Matar et al., 2001;Moineau and Goulet, 1997; Perdigon et al., 1999). Moreover, a numberof studies carried out in animals and in humans have shown that LABmay attenuate infections caused by respiratory pathogens. When usinganimal models, the oral application of different LAB strains increasedphagocytic activity in macrophages and enhanced clearance ofS. pneumoniae (Cangemi de Gutierrez et al., 2001) and P. aeruginosa(Alvarez et al., 2001). Similarly, LAB were also shown to enhancecellular immunity and reduce influenza virus titers in aged mice (Horiet al., 2001, 2002). These results suggest that selected strains of LAB arecapable of preventing respiratory tract infections, perhaps by microbialexclusion and/or by mediating nonspecific immune responses. Inter-estingly, Cangemi de Gutierrez et al. (2001) also showed that protectedmice had increased numbers of lymphocytes in their lamina propria, aswell as higher levels of antibodies binding to S. pneumoniae, indicatingthat a specific immune response may have been elicited. Some studiescarried out in humans have further demonstrated the potential of usingprobiotics to confer protection against respiratory diseases, particularlyin young children. In a randomized, double-blind, placebo-controlledstudy, the consumption of milk containing the probiotic strain Lacto-bacillus rhamnosus GG modestly reduced the incidence of respiratoryinfections as well as their severity in young children (1–6 yr) (Hatakkaet al., 2001). In a similar study, a live dietary supplement containingLactobacillus acidophilus and Lactobacillus casei suppressed pneumo-nia and decreased bronchitis in 6- to 24-month-old children (Rio et al.,2002). These studies have been supported by Gluck et al. (2003), whodemonstrated that nasal colonization by pathogenic bacteria (S. aureus,S. pneumoniae, and beta-hemolytic streptococci) was reduced inindividuals who ingested milk supplemented with a cocktail of

USE OF RECOMBINANT LAB IN HUMAN HEALTH 9
probiotic LAB as compared with individuals fed standard yogurt(Gluck and Gebbers, 2003). While only preliminary, these studies dosuggest that oral administration of probiotic LAB may help to reducethe occurrence and severity of respiratory infections in both childrenand adults. These findings have important implications for the devel-opment of recombinant LAB vaccines against respiratory pathogens,because they suggest that LAB themselves may have an effect onimmune function that could potentially act as an adjuvant duringvaccination.
Previous work has already demonstrated that intranasal administra-tion of recombinant Lc. lactis and Lb. plantarum strains expressingTTFC could elicit significant antigen-specific IgA in broncheoalveolarfluids as well as antigen-specific serum antibody and T cell responsesin mice (Grangette et al., 2001; Norton et al., 1997). Moreover, theseimmune responses, which were dose dependent, were sufficient toprotect against lethal challenge with injected tetanus toxin. Since then,reports of the construction of LAB strains expressing antigens from anumber of respiratory pathogens including S. pneumoniae (Gilbertet al., 2000), Bacillus anthracis (Zegers et al., 1999) and Bordetellapertussis (Lee et al., 1999, 2002) were reported but await proper testingin relevant animal models of disease. More recently, prototype vaccinestrains of Lc. lactis and Lb. plantarum expressing pneumococcal PspAantigen were constructed and evaluated in a respiratory challengemodel for S. pneumoniae (Hanniffy et al., 2004). PspA, a surfaceprotein and virulence factor found on all isolates of S. pneumoniae(Briles et al., 1998; Crain et al., 1990), is highly immunogenic andis considered a vaccine candidate because it has been shown to conferprotection against virulent isolates in different animal models of pneu-mococcal infection (Bosarge et al., 2001; Briles et al., 1996, 2000).Intranasal administration of LAB expressing PspA have now beenshown to elicit mucosal and systemic immune responses that protectagainst lethal challenge with virulent pneumococcci (Hanniffy et al.,2004).
C. INFECTIONS OF THE GASTROINTESTINAL TRACT
As constituents of the normal microflora, LAB have long been recog-nized as playing a role in maintaining oral tolerance and homeostasisin the gut. Additionally, LAB have been used therapeutically to protectagainst gastrointestinal infections, the most notable example being theuse of Lactobacillus GG to treat acute infectious diarrhea in children(Rosenfeldt et al., 2002a,b; Szajewska and Mrukowicz, 2001). The exact

10 HANNIFFY et al.
mechanisms that confer significant clinical benefit following adminis-tration of probiotic LAB are uncertain, but various intrinsic propertiesof the bacteria have been proposed as an explanation for the beneficialeffects (Heyman, 2000). In addition to restoring normal intestinalmicroflora, LAB may help to eliminate enteric pathogens by reinforcingintestinal barrier function, increasing mucosal secretory IgA andhumoral immune responses, and boosting specific and nonspecificimmunity. In vitro experiments with intestinal epithelial cell lineshave demonstrated that LAB probiotic strains can prevent adhesionand invasion by pathogenic bacteria and enhance barrier function innaıve epithelial cells (Lee et al., 2003; Resta-Lenert and Barrett, 2003).Various studies have also shown that LAB can intervene by binding toreceptors on gastric and intestinal epithelium typically used by patho-gens to gain entry into host cells (Mukai et al., 2002; Neeser et al.,2000). In addition, certain LAB produce relatively large amounts oforganic acids (Aiba et al., 1998; Koga et al., 1998; Midolo et al., 1995)and/or bacteriocin-like substances (Kim et al., 2003; Lee et al., 2003;Strus et al., 2001) that have been implicated in the inhibition ofHelicobacter pylori, Campylobacter spp., and Clostridium difficile.While the extent to which these components can cause an effect inthe gastrointestinal (GI) environment remains questionable, it is be-coming increasingly clear that by modulating immune responses,LAB play a vital role in protecting against pathogens. Experimentscarried out in animal models have demonstrated the protective cap-abilities of different LAB against pathogenic Escherichia coli (Ogawaet al., 2001; Shu and Gill, 2001), Salmonella typhimurium (Shu et al.,2000), Helicobacter pylori, and Clostridial species. Shu et al. (2000)demonstrated that compared with control mice, Lb. rhamnosus HN001-fed mice exhibited lower morbidity and bacterial translocation rateswhen challenged with E. coli O157:H7 (Shu et al., 2000), which wasassociated with significantly higher levels of pathogen-specific IgA andblood leukocyte phagocytic activity in the intestines of these mice. In aseparate study, mice administered dietary Bifidobacterium lactis de-monstrated increased specific and nonspecific immune responses andreduced intestinal infection by S. typhimurium (Shu and Gill, 2001).These results would suggest that LAB may be capable of enhancinglocal immune responses that might play an important role in protectingagainst infectious disease.
Using genetic engineering to further enhance probiotic LAB strainsholds much promise in the prevention and treatment of infecti-ous diseases of the GI tract. Prototype recombinant Lc. lactis vaccinestrains expressing antigen have already been developed against

USE OF RECOMBINANT LAB IN HUMAN HEALTH 11
H. pylori (Lee et al., 2001) and rotavirus (Enouf et al., 2001; Gil et al.,2001). Oral administration of recombinant Lc. lactis expressing the H.pylori urease subunit B antigen did stimulate low levels of antigen-specific immune responses in serum, but no protection was observedduring challenge with H. pylori (Lee et al., 2001). The influence ofdifferent LAB strains and choice of antigen is likely to be critical toany vaccine delivery strategy, particularly when dealing with the GItract, where persistence and survival of different strains can differwidely (Vesa et al., 2000). Other variables such as interaction of thestrain with mucus, intestinal epithelium, and lymphoid cells, as wellas the location and amount of antigen expressed in the bacteria, are alsolikely to influence immune outcomes. Except for recent studies onthe effect of antigen quantity on level of antibody response, theimpact of many of the aforementioned variables has yet to be assessedexperimentally (Wells and Mercenier, 2003).
D. INFECTIONS OF THE UROGENITAL TRACT
LAB-based vaccine delivery systems may also be appropriate forpreventing and treating bacterial vaginosis (BV) as well as specificpathogen-related infections of the urogenital tract (Reid and Bruce,2001; Reid and Burton, 2002). These include human immunodeficien-cy virus (HIV), Chlamydia, herpes simplex virus (HSV), papillomavi-rus, Treponema palladium, and Trichomonas vaginalis, as well asdiseases caused by bacterial pathogens such as Neisseria gonorrhoeae,group B Streptococcus, and enteropathogenic E. coli. Lactobacilli aredominant among microflora associated with the urogenital tract ofhealthy women but are almost completely absent in patients whodevelop most forms of urinary tract infections. Depletion or distur-bance of vaginal Lactobacillus sp. has been associated with the devel-opment of bacterial vaginosis as well as increased risk of acquiring HIVand other sexually transmitted diseases (Hillier et al., 1993; Taha et al.,1998; Wiesenfeld et al., 2003). There is evidence that lactobacilli canprevent urinary tract infections by microbial exclusion; by producingbacteriocins, biosurfactants, hydrogen peroxide, and coaggregation mo-lecules; by maintaining a low pH; or a combination of these factors(Reid, 2002). In vitro studies have shown that adhesive lactobacilli caninhibit growth and attachment of uropathogenic bacteria to uroepithe-lial cells in a strain-specific manner (Boris et al., 1998; McGroarty andReid, 1988; Osset et al., 2001). Similarly in mice, an endogenouspopulation of Lb. casei in the urinary tract prevented colonization byuropathogenic bacteria in the absence of pathogen-specific immune

12 HANNIFFY et al.
responses (Reid et al., 1985). More recently, a randomized, placebo-controlled trial of 64 healthy women showed that daily consumptionof a capsule containing the probiotic strains Lb. rhamnosus GR-1 andLactobacillus fermentum RC-14 reduced colonization of the vagina bypotential pathogenic bacteria and yeast (Reid et al., 2003). While thisobserved probiotic effect appeared to be mediated predominantlythrough microbial exclusion and/or production of antagonistic bypro-ducts, an immunological component cannot be discounted. Althoughfurther studies are required to determine if LAB can induce protectiveimmune responses in the vagina and bladder, these findings do suggestthat daily intake of probiotic LAB could provide a natural, safe, andeffective means of stabilizing the continually fluctuating vaginal floraand lower the risk of infection in healthy individuals as well as thoseprone to urogenital disease.
For women, direct application of a LAB-based vaccine by the intra-vaginal route leading to the induction of local antigen-specific immuneresponses could provide increased protection against urogenital tractinfections, reduce pathogen carriage, and help block sexual transmis-sion of pathogens. Previous findings, however, suggest that the urogen-ital tract may have a limited ability to mount immune responses toepithelial infections. This is evident from clinical observations thatpathogens such as N. gonorrhoeae can be contracted repeatedly in theabsence of effective immunity from previous infections (Russell, 2002).In addition, local immunization in order to induce local immuneresponses would also be effected by hormonal variations that are morepronounced in the urogenital tract than at other mucosal sites and areknown to have profound effects on susceptibility to sexually transmit-ted diseases (Gallichan and Rosenthal, 1996; Gillgrass et al., 2003;Kaushic et al., 2000). While mucosal immunization usually results inhigher IgA at the site where immunization occurs, there is evidencethat this is not the case in the vagina and that other routes of adminis-tration may be more effective. Experiments carried out in mice by Wuet al. (2000) showed that intranasal immunization could induce sub-stantially higher levels of IgG and IgA in vaginal fluids and serum ascompared with intravaginal immunization. Such compartmentaliza-tion within the common mucosal immune system occurs also in hu-mans, where the route of administration can significantly influenceimmune responses at remote mucosal surfaces (Kantele et al., 1998).This has been demonstrated by Kutteh et al. (2001), who showed thatinduction of antibodies (S-IgA) in the female genital tract was bestachieved by oral immunization followed by a rectal administration(Kutteh et al., 2001).

USE OF RECOMBINANT LAB IN HUMAN HEALTH 13
LAB would offer greater flexibility as vaccine vectors in that they canbe administered by whatever route is deemed most suitable for a givenclinical situation. The nasal route of administration, for example, maybe the best inductive site for a common mucosal immune response andcould provide a useful strategy for inducing more-potent, longer-last-ing immune responses not just in saliva but in remote secretions (e.g.,vaginal fluids) as well as in serum (Wu et al., 2000). Recombinant Lc.lactis and S. gordonii vaccine strains have already been developedagainst human papillomavirus type 16 (HPV-16) (Bermudez-Humaranet al., 2002, 2003; Cortes-Perez et al., 2003) and HIV (Oggioni et al.,2001; Xin et al., 2003). Similarly, Lactobacillus vaccine strains againstChlamydia psittaci and HIV have also been engineered (Zegers et al.,1999). Xin et al. (2003) also showed that Lc. lactis expressing theenvelope protein of HIV protected mice against intraperitoneal chal-lenge by an HIV Env-expressing vaccinia virus. While oral and intrana-sal administration of a number of these LAB vaccine strains has beeneffective in inducing antigen-antibody responses in serum, their abilityto induce local and remote mucosal responses has not yet been inves-tigated (Cortes-Perez et al., 2003; Xin et al., 2003). Interestingly, vaginalcolonization of mice with recombinant strains of S. gordonii, expres-sing HPV and HIV antigens, has been shown to induce antigen-specificvaginal IgA as well as serum IgG (Medaglini et al., 1998). These strainswere also able to induce local and systemic immune responses whenrepeatedly administered (three inoculations) to the vagina of Cynomo-logus monkeys (Di Fabio et al., 1998). These findings would indicatethat LAB possess adjuvant properties that may be capable of elicitingantigen-specific local as well as systemic immune responses evenwhen applied directly to the vagina. If persistence and colonizationof the vagina are essential components of a LAB-based vaccine ortherapy, intravaginal immunization would therefore be optimal. Sucha strategy would be particularly preferable where the aim is to providepassive immunity in situ. This has recently been addressed by Changet al. (2003), who engineered Lactobacillus jensenii, a natural humanvaginal isolate, to express and secrete the HIV binding protein CD4,which bound the HIV gp120 protein and inhibited HIV-1 entry intotarget cells in vitro. Lb. jensenii efficiently colonizes the vaginal muco-sa of women where it exists as part of a natural ‘‘biofilm’’ composed ofbacteria and extracellular matrix materials. It is hoped that residentLAB engineered to express HIV binding proteins that are either surfaceassociated or secreted into the surrounding ‘‘biofilm’’ and mucus layercould effectively neutralize HIV particles by impeding viral accessto epithelial cells and prolonging exposure to viral inactivating

14 HANNIFFY et al.
substances that are produced naturally. The potential of this ap-proach has been demonstrated by reports detailing the developmentof prototype S. gordonii vaccine strains expressing molecules thatare microbiocidal against candidiasis (Beninati et al., 2000) andHIV (Giomarelli et al., 2002). Such LAB vaccines, perhaps in tabletform (Bonetti et al., 2003; Maggi et al., 2000; Mastromarino et al., 2002),could be self-administered intermittently to provide ongoing pro-tection against these pathogens. Vaccines such as this could potentiallybe further improved by combining different LAB strains with dif-ferent antimicrobial, adhesive, and biochemical characteristics to in-crease their effectiveness in preventing and treating urogenitaldiseases.
Recombinant LAB vaccines would also be suitable for maternal vac-cination strategies aimed at preventing disease in neonates, such asthose caused by group B Streptococcus (GBS) and selected strains of E.coli. While antibiotic therapy has proved effective in reducing theincidence of neonatal disease in developed countries, other complica-tions such as premature rupture of the membranes, premature birth,low-birth-weight babies, or stillbirth have been attributed to GBS andother infections of the urogenital tract. A LAB-based vaccine againstGBS and other pathogens could reduce the level of colonization in themother and provide an important first line of defense against thepathogen at mucosal surfaces. In addition, colostrum antibody andtransplacentally transferred serum IgG antibodies against GBS couldconfer immune protection to the newborn. Recombinant strains of Lc.lactis and Lb. plantarum have already been developed that expressantigen that has been shown to be protective in an invasive animalmodel of GBS infection (Seepersaud et al., 2004). These strainsare now being tested in animal models for their ability to elicit protec-tive mucosal and systemic responses (Hanniffy and Wells, personalcommunication).
III. Potential for Immune Modulation of Type I Allergy Using
Recombinant LAB
The prevalence of type I allergy has constantly increased withinrecent years, leading to the fact that currently up to 25% of the popula-tion in industrialized countries suffer from allergic symptoms such asallergic rhinoconjunctivities, allergic asthma, or atopic dermatitis. Be-sides atopic disposition (Marsh et al., 1994) and allergenic molecules(Scheiner and Kraft, 1995), there are various environmental factorssuch as air pollutants (von Mutius et al., 1994), changes in nutrition

USE OF RECOMBINANT LAB IN HUMAN HEALTH 15
(Bottcher et al., 2000), or ‘‘sterile’’ lifestyles in industrialized coun-tries without bacterial epidemics (Ring, 1997) that may have a causalrelationship with the increasing prevalence of the disease.
The pathway of IgE production is well described for mice and manand is basically influenced by the reciprocal relationship betweenTh2 cytokines, mainly IL-4, the switching factor for IgE class, andTh1 cytokines (e.g., interferon gamma [IFN-�], which counteracts theactivity of IL-4) (Mosmann and Coffman, 1989). In atopic individualsan imbalance favors the production of IL-4, leading to a bias of Th2responses with an overproduction of IgE against allergenic proteins(Romagnani, 1994). These molecules lead to cross-linking of the aller-gen-specific IgE bound to the surface of mast cells via specific FC-�-receptors, thereby triggering the release of anaphylactic mediators,leading to the characteristic allergic symptoms mentioned previously.
Until now, specific immunotherapy (SIT) has been the treatment ofchoice against type I allergy, performed by injecting increasing doses ofallergens (Bousquet et al., 1998). Although SIT is effective in themajority of treated patients, there are certain drawbacks, such as fre-quent injections and long duration of the treatment, leading to poorcompliance in patients. Moreover, aluminium salts, which are used asadjuvants in SIT, are potent inducers of Th2 responses, which mightreduce the efficacy of the treatment. Thus, there is increasing interestin improving immunotherapy by using Th1-promoting adjuvants(Wheeler and Woroniecki, 2001) and/or administration via a lessinvasive route, such as mucosal delivery (Morris, 1999).
A. ROLE OF THE INDIGENOUS MICROFLORA
The indigenous microflora plays an important role in anti-infectiousresistance by competitive interaction with pathogenic bacteria but isalso important for directly influencing immune responses. This hasbeen demonstrated in animals reared under sterile conditions (germ-free animals), in which it has been shown that systemic and localimmune responses are more difficult to establish and that, in particu-lar, the induction of oral tolerance is unstable and short-lived (Cebraet al., 1999). Based on these findings, an imbalance of the compositionof the indigenous microflora is believed to play a role in the devel-opment of inflammatory diseases, such as intestinal bowel disease(Duchmann et al., 1995) and allergies (Rautava and Isolauri, 2002;Wiedermann, 2003). Indeed, differences in the intestinal colonizationpattern between children of ‘‘Western lifestyle countries’’ with a highprevalence of allergies and of economically low developed countries

16 HANNIFFY et al.
where allergies are less common has also been reported (Bottcher et al.,2000). The ‘‘hygiene theory’’ proposes that an increasingly hygieniclifestyle has contributed to the increasing prevalence of allergic diseasecaused by intestinal colonization, with a limited range of microbes(Strachan, 1989).
In the last few years, the possible role of specific LAB strains in theprevention of allergic diseases has become more evident. In particularthe relationship between the composition of the intestinal flora and theprevalence of allergic diseases has been epidemiologically documen-ted. One study showed that infants from countries with a high preva-lence of allergy, such as Sweden and England, have a lower level ofintestinal colonization with certain LAB strains than children fromcountries where allergic diseases are less prevalent, such as Estoniaand Nigeria (Bjorksten et al., 1999; Sepp et al., 1997; Simhon et al.,1982). Moreover, it was recently demonstrated that oral administrationof a particular LAB strain (Lb. rhamnosus GG) led to reduced atopicdermatitis in children with a positive family history of type I allergy(Kalliomaki et al., 2001), indicating that LAB can directly or indirectlyexert an anti-allergic effect.
B. ANIMAL MODEL OF TYPE I ALLERGY
Among the numerous inhalant allergens, tree pollen of the whitebirch Betula verrucosa is one of the most important sources responsiblefor eliciting allergic symptoms. The major allergen of birch pollen isBet v 1, a 17-kD molecule, to which 95% of birch pollen allergicpatients (and 60% exclusively) display IgE binding reactivity. Bet v 1was the first pollen allergen to be cloned, sequenced, and produced as arecombinant protein in E. coli (Breiteneder et al., 1989) and to have itscrystalline structure determined (Gajhede et al., 1996). RecombinantBet v 1 has also been shown to possess biological properties equivalentto those exhibited by the natural Bet v 1 molecule (Ferreira et al., 1993).Moreover, immune responses to Bet v 1 have been characterized at theB and T cell level in atopic and nonatopic individuals (Ebner et al.,1995).
An animal model of allergic sensitization to birch pollen and itsmajor allergen Bet v 1 has been developed in BALB/c mice that havebeen identified as high IgE responders to this allergen (Bauer et al.,1997; Wiedermann et al., 1998). The established standard sensitizationscheme is based on an intraperitoneal injection of recombinant (r) Bet v1 adsorbed to aluminium hydroxide (Al(OH)3), followed by an aerosoltreatment with natural birch pollen extract (BP). This sensitization

USE OF RECOMBINANT LAB IN HUMAN HEALTH 17
procedure leads to high allergen-specific IgE/IgG1 antibody levels,positive immediate type skin reactions in vivo and Th2-like immuneresponses in vitro, eosinophilic infiltration within the lungs, as wellas broncheoalveolar and airway hyper-responsiveness. Thus the im-mune responses represent an immunological state comparable to thatof human type I allergy/asthma (Wiedermann et al., 1998). Using thismodel, several mucosal adjuvants have been tested for their capacity tomodulate an allergic immune response. In this respect it has beendemonstrated that immunomodulation can be achieved by using mu-cosal adjuvants such as cholera toxin (Wiedermann et al., 1998), chol-era toxin subunit B (Wiedermann et al., 1999), or certain bacterialcomponents (CpG-motifs) (Jahn-Schmid et al., 1999).
C. USE OF LAB FOR PROPHYLAXIS AND THERAPY OF TYPE I ALLERGY
Experiments carried out in mice have shown that IgG1 and IgEantibody levels could be reduced when certain LAB strains were in-jected or orally applied with a particular antigen/allergen (Matsuzakiet al., 1998; Shida et al., 2002). In light of these studies, experimentswere carried out to investigate the capacity of two LAB strains (Lacto-coccus lactis MG1363 and Lactobacillus plantarum NCIMB8826) toprevent or modulate allergic immune responses. Both LAB strainsinduced high levels of Th1 cytokines IL-12 and IFN-� in naıve murinespleen cell cultures possibly via the mechanisms depicted in Fig. 1. Inthe murine birch pollen allergy model, intranasal or oral co-applicationof Lc. lactis or Lb. plantarum with recombinant Bet v 1, prior to or afterallergic sensitization, led to increased levels of allergen-specific IgG2aantibodies and in vitro IFN-� production, indicating a shift towardTh1 responses. Successful immunomodulation by the mucosal pre-treatment was further demonstrated by suppression of allergen-inducedbasophil degranulation (Repa et al., 2003). These results, based oninduction of counter-regulatory Th1 responses, indicated that com-bined mucosal application of LAB with a specific allergen could providean effective prophylactic and therapeutic strategy against allergy.
The construction of recombinant LAB for local delivery of an aller-gen could further enhance this protective/immunostimulatory effectand could represent a useful tool for mucosal vaccination againsttype I allergy. Recently it has been shown that treatment with a recom-binant LAB strain expressing the house dust mite allergen Der p1reduced levels of allergen-specific IL-5 (a Th2 cytokine) in sensi-tized mice (Kruisselbrink et al., 2001). Similarly, recombinant Lc. lactisand Lb. plantarum strains expressing Bet v 1 have now also been

18 HANNIFFY et al.
constructed and preliminary data indicate that mucosal admin-istration of these strains prior to or after sensitization results in asignificant suppression of allergic immune responses (Mercenier andWiedermann, personal communication).
IV. Opportunities for the Treatment of Inflammatory Bowel
Diseases Using Recombinant LAB
IBDs, as typified by ulcerative colitis and Crohn’s disease, are char-acterized by chronic dysregulation of inflammatory immune responsesin the gastrointestinal tract. While the pathogenesis of IBD remainsunclear, it is thought to involve complex interactions combining hostgenetic susceptibility, intestinal bacteria, and gut mucosal immuneresponses (Farrell and Peppercorn, 2002; Rath, 2003). It is now gener-ally accepted that intestinal microflora provide the antigenic stimuli toderegulate mucosal immune responses in genetically susceptible hostssuch that they become overly aggressive with reduced tolerance towardthe indigenous microflora.
The proposed use of probiotics including LAB for therapy and man-agement of IBD has arisen from increasing evidence implicating indig-enous bacteria in the pathogenesis of these diseases. Much of thisevidence comes from transgenic animal models in which immunopath-ological disease is induced by the absence of immunologically impor-tant molecules but is dependent on the presence of a normal bacterialmicroflora (Blum and Schiffrin, 2003; Hudcovic et al., 2001; Stroberet al., 2002). For example, mice with disrupted IL-2 and IL-10 genes or�/� T cell receptor mutants that normally develop chronic intestinalinflammation resembling ulcerative colitis in humans remain healthyin germ-free conditions (Strober et al., 2002). These studies confirmedthat IBD is likely to result from abnormal immune responses to normalintestinal microflora and demonstrated the importance of regulatorycytokines such as IL-10 in maintaining immune homeostasis at muco-sal sites (Blum and Schiffrin, 2003). As a result, there has beenincreasing interest in using probiotics, particularly those with immu-nomodulatory capacities including the ability to induce immunoregu-latory cytokines such as IL-10 and TGF-�. This has led to suggestionsthat LAB and their associated anti-inflammatory effects may be capableof reinstating mucosal immune homeostasis and may provide an alter-native strategy for intervention in IBD patients (Blum and Schiffrin,2003).

USE OF RECOMBINANT LAB IN HUMAN HEALTH 19
A. ROLE OF LAB IN INTESTINAL BARRIER FUNCTION
One task of the gut is to act as a barrier between the external andinternal environments to prevent the entrance of potentially harmfulcomponents. This barrier, which can be considered both as physical(paracellular permeability and protective action derived from mucus)and functional (mucosal immune system), is strongly disrupted ininflammatory states. A decrease in mucosal barrier function consistent-ly occurs in experimental colitis as well as in human IBD (Rath, 2003).One of the main actions of probiotics concerns the reinforcementof the intestinal mucosal barrier, an activity which can confer intesti-nal anti-inflammatory properties to some probiotics (see review,Fioramonti et al., 2003). For example, treatment with Lb. reuteri orLb. plantarum reduced the level of intestinal permeability in a ratmodel of methotrexate-induced enterocolitis (Mao et al., 1996).
In addition, as well as decreasing intestinal myeloperoxidase levels,the administration of LAB re-established the intestinal microecologyand reduced bacterial translocation to extra-intestinal sites. The abilityof LAB to enhance barrier function and reduce bacterial translocationmay play an important role in preventing subsequent activation ofinflammatory responses. Bifidobacterial supplementation has beenshown to reduce the incidence of necrotizing enterocolitis in miceby preventing bacterial translocation and subsequent activation ofinflammatory mediators such as plasma endotoxin and intestinalphospholipase A2 (Caplan et al., 1999). In another study, Shiba et al.(2003) provided evidence that treatment of Bifidobacterium vulgatus–implanted mice with Bifidobacterium infantis abrogated increases inplasma B cells in the Peyer’s patch, probably by protecting the epithe-lium layer (including Peyer’s Patch) from invasion by B. vulgatus. LABmay also have a trophic action on colonic mucosa. This has beenelegantly shown in rats, where an elemental liquid diet induced anatrophy of colonic mucosa (assessed by the rate of crypt cell produc-tion) that was significantly improved by treatment with Lb. casei orClostridium butyricum (Ichikawa et al., 1999). Strong interactions existbetween mucus and colonic bacteria, and some actions of probioticsmay involve this protective glycoprotein layer. The inability to degrademucus by bacteria such as Lactobacillus acidophilus or Bifidobacter-ium bifidum could itself be seen as protective (Ruseler-van Embdenet al., 1995). Indeed, in vitro experiments have shown that adherenceby specific strains of Lactobacillus induces mucin gene expression andextracellular secretion of MUC3 by intestinal epithelial cells (Macket al., 2003). Moreover, there was a direct correlation between

20 HANNIFFY et al.
increased mucin secretion and reduced adherence by enteropathogenicEscherichia coli. Similar interactions between LAB and host epithe-lial and other host cells may play an important role in protect-ing against translocation of antigenic stimuli that contribute to thepathogenesis of IBD.
B. LAB AND INTESTINAL INFLAMMATION
A number of studies have shown that a select number of probioticstrains can reduce experimental colonic inflammation in animals.A study carried out by Fabia et al. (1993) was one of the first to showthat treatment with LAB could prevent the development of acetic acid–induced colitis in rats. Intracolonic administration of Lb. reuteri pro-duced normal myeloperoxidase (MPO) activity levels and mucosalpermeability and prevented the development of morphologic lesions(Fabia et al., 1993). These pioneering experiments were subsequentlyreproduced with other bacteria and in other models of experimentalcolitis. Lb. reuteri, but not Lb. rhamnosus, was shown to be effective inattenuating acetic acid–induced colitis in rats (Holma et al., 2001).Treatment with Bifidobacterium longum in mice (Fujiwara et al.,2003) and C. butyricum in rats (Araki et al., 2000) both reduced theseverity of a colitis induced by dextran sulfate sodium (DSS). In addi-tion, Lb. plantarum and Lb. reuteri ameliorated methotrexate-inducedentercolitis in rats (Mao et al., 1996). The spontaneous development ofcolitis in IL-10–deficient mice has also been ameliorated by treatmentwith Lb. plantarum (Schultz et al., 2002) or Lb. reuteri (Madsen et al.,1999a). In a recent study it has been shown that treatment with asolution of lysed E. coli ameliorated a colitis induced by DSS in mice(Konrad et al., 2003). To date, there is no evidence from animal modelsthat LAB can adversely effect the development of colitis or augmentseverity of associated symptoms. While the mechanisms remain un-clear, evidence from some of these studies indicates that administrationof LAB may induce tolerance to bacterial antigens by down-regulatingTh1 inflammatory cytokines (Konrad et al., 2003; Schultz et al., 2002).However, it is also possible that LAB may beneficially interact withother commensal bacteria that may play a pathogenic role in IBD.A recent study carried out by Shiba et al. (2003) showed that B. infantisinhibited the growth of B. vulgatus, a putative pathogenic microbe inIBD. In addition, B. infantis also suppressed systemic immune re-sponses to B. vulgatus in a gnotobiotic murine model (Shiba et al.,2003). This study would indicate that LAB-like B. infantis may protect

USE OF RECOMBINANT LAB IN HUMAN HEALTH 21
epithelial layers from invasion by other commensal microbes that arebelieved to play a pathogenic role in IBD.
In recent years we have seen probiotics including LAB being used asan experimental therapy against IBD in humans. In a double-blind,placebo-controlled trial, a probiotic preparation (VSL#3), a cocktail ofBifidobacterium, Lactobacillus, and Streptococcus species, was shownto be effective in preventing flare-ups of chronic ileal pouchitis(Gionchetti et al., 2000). The VSL#3 preparation has also been usedfor maintenance treatment of ulcerative colitis (Venturi et al., 1999) andin preventing postoperative recurrence of Crohn’s disease. More recent-ly, Gionchetti et al. (2003) showed in double-blind, placebo-controlledtrial that treatment with VSL#3 was effective in preventing the onset ofacute pouchitis in patients with ileal pouch-anal anastomosis.Promising data have been also obtained with E. coli strain Nissle, whichwas found to have an equivalent to traditional treatment with mesala-zine in maintaining remission of ulcerative colitis (Kruis et al., 1997;Rembacken et al., 1999). On a cautionary note, other studies carried outin humans have shown LAB/probiotic therapy to have little or no effectin the treatment of IBD. While feeding Lactobacillus GG to patients witha history of pouchitis and endoscopic inflammation did change thepouch bacterial flora, the treatment proved ineffective as a primarytherapy for a clinical or endoscopic response (Kuisma et al., 2003).Similarly, Lactobacillus GG treatment did not prevent endoscopic re-currence 1 year after curative resection for Crohn’s disease nor did itreduce the severity of recurrent lesions (Prantera et al., 2002).
While some studies indicate that probiotics do have potentialas therapies against IBD, they have provided little insight into themicrobiological and immunological mechanisms that underlie thesediseases. A recent study showed that ex vivo production of the pro-inflammatory cytokine tumor necrosis factor alpha (TNF-�) by ileal–mucosal explants surgically removed from Crohn’s patients weredown-regulated in the presence of Lb. casei and Lactobacillus bulgar-icus but not by E. coli and Lactobacillus crispatus (Borruel et al., 2002).This would indicate that LAB interact differently with immunocompe-tent cells and have different capacities in modulating the production ofpro-inflammatory cytokines such as TNF-�, which play a key role inthe pathogenesis of IBD. More clinical trials are therefore needed toevaluate different LAB strains as therapeutic preparations against thevarious manifestations of IBD. These studies must also establishthe correct placement and dosage of probiotic required for any treat-ment regimen. Clearly more research is also needed to characterizeand establish the mechanisms underlying the interactions of probiotic

22 HANNIFFY et al.
bacteria with the immune system and the role of intestinal microflorain homeostasis. The generated understanding will be of crucial impor-tance for the future screening of lactic acid bacteria affecting immunefunctions and the development of recombinant strains with enhancedproperties.
C. RECOMBINANT LAB AS A THERAPEUTIC OPTION
While results from experiments carried out in animals are encourag-ing, the potential of probiotic therapy against IBD in humans remainsuncertain. Various studies have now shown that in addition to provenefficacy against experimental colitis in animals, probiotics includingLAB also have other beneficial actions on intestinal mucosa. Unfortu-nately, these studies have yet to establish a link between the two sets ofdata, and the mechanisms that underlie the anti-inflammatory actionof probiotics remain ill-defined. Until more is known about mucosa-associated microflora and the mechanisms that underlie inflammatorydiseases, the use of probiotics in therapy of IBD will remain largelyempirical. An alternative approach is to genetically engineer LAB toproduce and deliver to the intestinal mucosa, molecules that have atherapeutic activity against IBD. Steidler and colleagues (2000) re-ported the construction of a recombinant Lb. lactis strain geneticallyengineered to secrete interleukin-10. Intravenous administration of IL-10 has previously shown clinical efficacy in the treatment of Crohn’sdisease (van Deventer et al., 1997) but can cause side effects thatprevent long-term use. In addition, IL-10 is sensitive to acid; thereforeintestinal delivery is not an option. In addressing this problem Steidleret al. (2000) showed that Lb. lactis genetically engineered to secreteIL-10 could reduce DSS-induced colitis and prevent spontaneous coli-tis in IL-10–deficient mice. Unfortunately, only a limited number ofmolecules or compounds display efficacy against intestinal inflamma-tion when infused in the colonic lumen. Nitric oxide (NO) is one suchcompound and was shown to reduce inflammation when infused intothe colonic lumen of rats (Perner and Rask-Madsen, 1999). Lactobacil-lus farciminis, which produces NO in vitro, has also been shown toproduce NO in the colon and to reduce experimental colitis whengiven orally to rats (Lamine et al., 2004). Another possibility wouldbe to engineer LAB-secreting antioxidant enzymes such as catalase orsuperoxide dismutase (SOD), which may prove effective in removingfree radicals such as superoxides and hydrogen peroxide producedby leukocytes, which are thought to play a role in amplifying inflam-matory responses and subsequent mucosal damage in IBD patients

USE OF RECOMBINANT LAB IN HUMAN HEALTH 23
(Babbs, 1992). Indeed, some LAB possess significant antioxidative ac-tivity and are intrinsically resistant to oxidative stress, properties thatmay help some isolates of LAB to serve as defensive components in theintestinal microbial ecosystem (Kullisaar et al., 2002).
While use of recombinant LAB in functional foods would not beacceptable, their use as therapeutics may be foreseen where the poten-tial benefits greatly outweigh any potential risks. This is particularlytrue of IBD for which current treatments are unsatisfactory and thedevelopment of new and innovative therapies is urgently needed.The recent approval by Dutch authorities to use genetically engineeredLc. lactis secreting IL-10 as an experimental therapy for humans withIBD gives encouragement for the further development of therapiesbased on recombinant LAB (Steidler et al., 2003).
V. LAB as Cell Factories for the Manufacturing
of Pharmaceutical Proteins
Several aspects are considered when choosing a host and a geneexpression system for the production of proteins for pharmaceuticaluse (‘‘pharmaceutical proteins’’). These include cost of production,yield, purity, formation of biologically active molecules, and possiblecontamination by toxic substances either produced by the host cell orpresent in the growth medium. In addition, commercial production of apharmaceutical protein and its production process (including appliedtools and unit operations) requires approval from the regulatory autho-rities. These processes must, therefore, follow good manufacturingpractices (GMP), ensuring that the production organism and associatedmanufacturing procedures and materials are well-characterized anddocumented. When considering LAB for commercial production ofpharmaceutical proteins, most issues that need to be addressed arecommon to all biological production systems and can be divided intofour categories relating to (1) the microbial host; (2) the expressionvector; (3) propagation, fermentation and initial downstream proces-sing; and (4) quality control of the product. The latter includes testingfor the proper activity as well as stability and purity and will not bediscussed here, because it is specific to each product.
A. THE CHOICE OF MICROBIAL HOST
E. coli was the first microorganism to be utilized for the production ofpharmaceutical proteins with recombinant DNA technology (Goeddelet al., 1979a,b; Villa-Komaroff et al., 1978), primarily because this bacte-rium had been extensively studied in laboratories throughout the world.

24 HANNIFFY et al.
E. coli demonstrates several disadvantages from a technological point ofview, such as inefficient protein secretion, formation of inclusion bodies,and production of endotoxin. However, because of precedent from itsprevious use in producing a long list of approved pharmaceutical pro-teins, E. coli continues to be the preferred bacterial production host forindustrial applications. This tendency to revert to a proven and acceptedproduction system seems to have hindered the adoption of LAB as aproduction host. Although LAB demonstrate several features that makethem ideal for producing certain products, pharmaceutical proteins pro-duced by these bacteria have yet to be approved and brought to market.However, it is expected that this bottleneck will be eliminated in thepresent decade, since several proteins produced in lactic acid bacteriaare currently being tested in clinical trials.
For regulatory approval of the recombinant product, a historical de-scription of the host strain is required (i.e., description of how the strainwas isolated, characterized, and subsequently treated and manipulatedin the laboratory prior to use). This should include a risk evaluationdescribing known and potential application risks and documenting anyrecord of infections and diseases caused by the same species as theselected host strain. Finally, the evaluation should also deal with hoststrain stability and potential for genetic exchange with other bacteria(e.g., natural competence for DNA uptake). As many LAB are alreadyused in food production and preservation and more recently as probio-tics, many of the aforementioned requirements for regulatory approvalare already in place for selected strains. Genome sequences are nowavailable for an increasing number of LAB (Klaenhammer et al., 2002),and various postgenomic studies involving DNA arrays (van de Guchteet al., 2002) and 2-D gel electrophoresis (Champomier-Verges et al.,2002; Guillot et al., 2003) have already been carried out with differentLAB under different conditions. These approaches can be used to com-pare production strains with their corresponding source strain, thuscontributing to more-robust risk assessment procedures capable of pre-dicting an undesirable effect. Similarly, comparative genomics andother bioinformatic approaches can also be used to identify potentialconjugation or mobilization genes, which can be subsequently deletedin order to minimize gene transfer and its associated risks.
B. THE EXPRESSION VECTOR
The vector for expression of a desired gene can be chromosomallyintegrated or plasmid borne. In addition to unnecessary and redun-dant DNA, it is essential that all genes dedicated to gene transfer are

USE OF RECOMBINANT LAB IN HUMAN HEALTH 25
eliminated. Similarly, if a plasmid is used, the origin of replicationshould ideally replicate in one or few species only. The entire DNAsequence of the plasmid or any integrated DNA must be determined totrace recombinant DNA during downstream processing as well as inthe final product. In addition, the segregational and structural stabilityof the chromosomal insert or expression plasmid in a production hostmust be analyzed to determine if it is stable. Usually a selection markeris required for maintenance of the plasmid carrying the desired gene.Previously the regulatory authorities have approved the use of antibi-otic selection markers. However, future approvals are expected to in-clude nonantibiotic selection markers only (see Section VI) because ofthe growing concerns regarding resistant bacteria whose emergenceappears to correlate with the extensive use of antibiotics.
The ideal promoter for gene expression should fulfil several require-ments (Makrides, 1996). While in most cases the activity of the promot-er should be as high as possible to produce the greatest amount ofprotein, high levels of constitutive expression can be lethal to the hostcell or result in inhibition of growth, leading to loss of the expressionvector or structural instability of the recombinant DNA. An induciblepromoter is therefore preferable to coordinate protein production withcell growth in a way that maximizes the ratio of protein yield relative tobiomass. Background transcription should be minimal during thegrowth phase to produce sufficient biomass, at which point inductionof the promoter should initiate a burst of gene expression.
The last decade has seen the development of elegant and efficienttools for genetic manipulation and gene expression in Lc. lactis, themost extensively studied member of the entire LAB group (reviewed inSection II). For example, the P170 expression system (Fig. 2) utilizes theregulated promoter P170, which is activated in Lc. lactis on transitionfrom exponential to stationary phase (Israelsen et al., 1995b; Madsenet al., 1999b). Consequently, the growth phase is separated from theprotein production phase that occurs only when biomass has reacheda maximum. The P170 system also utilizes the lactococcal replicon(repB) and the lactococcal hom and thrB genes, thus permiting auxotro-phic selection when using a hom-thrB-deficient production strain(Glenting et al., 2002; Madsen et al., 1996). The system has been furtheradapted to include DNA encoding the SP310mut2* lactococcal secre-tion signal (Ravn et al., 2000, 2003). The secretion signal directs the pre-protein to the translocation apparatus, where it is cleaved off thusreleasing the mature protein into the growth medium. Pharmaceuticalproteins secreted into the growth medium can be easily purified, therebyreducing downstream processing and potential cost of production.

FIG. 2. Expression vector pAMJ1223. The P170 promoter, signal peptide SP310mut2*,
the auxotrophic selection marker hom-thrB, and the Lc. lactis replicon repB are
indicated. The multiple cloning site is located between the signal peptide and the
transcription terminator.
26 HANNIFFY et al.
DNA microarray and proteomic characterization of global regu-latory responses to different environmental stimuli in Lc. lactis(Champomier-Verges et al., 2002; Guillot et al., 2003; van de Guchteet al., 2002) should help the development of new expression systemsthat are activated under industrial conditions. In addition, Lc. lactisstrains are now available that lack extracellular proteases, resultingin reduced degradation of secreted proteins (Madsen, personal com-munication; Miyoshi et al., 2002). Combining appropriate expressionsystems with such mutant strains could significantly improve pro-duction and secretion of pharmaceutical proteins, making Lc. lactisincreasingly attractive as an industrial production host.
C. PROPAGATION, FERMENTATION, AND INITIAL DOWNSTREAM PROCESSING
The production strain must be propagated into growth medium(propagation medium) that contains no toxic or harmful substances andensures the stability of the strain. Once the production strain has beencharacterized with respect to critical parameters (e.g., composition ofheterologous DNA, genetic stability, productivity, and byproducts),minimally passaged cultures are used to generate master and workingcell banks that are checked at regular intervals.
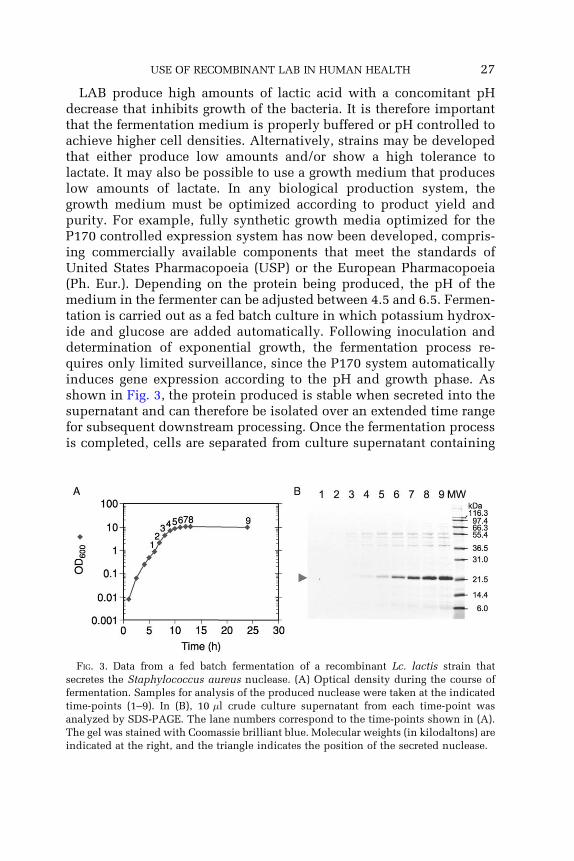
USE OF RECOMBINANT LAB IN HUMAN HEALTH 27
LAB produce high amounts of lactic acid with a concomitant pHdecrease that inhibits growth of the bacteria. It is therefore importantthat the fermentation medium is properly buffered or pH controlled toachieve higher cell densities. Alternatively, strains may be developedthat either produce low amounts and/or show a high tolerance tolactate. It may also be possible to use a growth medium that produceslow amounts of lactate. In any biological production system, thegrowth medium must be optimized according to product yield andpurity. For example, fully synthetic growth media optimized for theP170 controlled expression system has now been developed, compris-ing commercially available components that meet the standards ofUnited States Pharmacopoeia (USP) or the European Pharmacopoeia(Ph. Eur.). Depending on the protein being produced, the pH of themedium in the fermenter can be adjusted between 4.5 and 6.5. Fermen-tation is carried out as a fed batch culture in which potassium hydrox-ide and glucose are added automatically. Following inoculation anddetermination of exponential growth, the fermentation process re-quires only limited surveillance, since the P170 system automaticallyinduces gene expression according to the pH and growth phase. Asshown in Fig. 3, the protein produced is stable when secreted into thesupernatant and can therefore be isolated over an extended time rangefor subsequent downstream processing. Once the fermentation processis completed, cells are separated from culture supernatant containing
FIG. 3. Data from a fed batch fermentation of a recombinant Lc. lactis strain that
secretes the Staphylococcus aureus nuclease. (A) Optical density during the course of
fermentation. Samples for analysis of the produced nuclease were taken at the indicated
time-points (1–9). In (B), 10 �l crude culture supernatant from each time-point was
analyzed by SDS-PAGE. The lane numbers correspond to the time-points shown in (A).
The gel was stained with Coomassie brilliant blue. Molecular weights (in kilodaltons) are
indicated at the right, and the triangle indicates the position of the secreted nuclease.

28 HANNIFFY et al.
the pharmaceutical protein. The supernatant can then be concentratedby dia-filtration in readiness for downstream purification and formula-tion. A risk evaluation of the P170 Expression System and host strainshas already been carried out by the independent Danish ToxicologyCentre, which is now being used as a guide by a number of commercialclients to meet with demands from regulatory authorities (Israelsen,personal communication).
VI. Engineering LAB for Their Safe Use in Humans
The use of genetically modified microbes (GMMs) in humans raisesconcerns about their survival and proliferation in the environment.Microorganisms have evolved highly efficient systems for horizontalgene transfer such as transformation, conjugation, retromobilization,and transduction to improve their adaptation to changes in their eco-logical niche. Therefore the transfer of recombinant DNA such asantibiotic resistance markers or other genetic modifications from awell-characterized transgenic microorganism to uncharacterized indig-enous species is perceived as a significant risk that must be minimized(Gruzza et al., 1993; Netherwood et al., 1999; Ramos et al., 1995; vonWright and Bruce, 2003). In the context of recombinant live vaccines,for example, a possible scenario could be the transfer of a clonedvirulence determinant from a live vaccine strain to a pathogen poten-tially reinforcing its ability to cause infection. Apart from the transgeneof interest, the ideal GMM for use in humans should therefore containthe minimal amount of foreign DNA and must not include an antibioticresistance marker. Furthermore, the possibilities of transgene horizon-tal transfer must be minimized, and GMM lethality should be achievedin an unconfined environment.
A. FOOD-GRADE SYSTEMS IN LAB FOR PLASMID MAINTENANCE AND
CHROMOSOMAL INSERTION
A large number of safe and sustainable food-grade systems for genet-ic modifications of LAB have been developed (initially for Lc. lactis),including endogenous cloning vectors, selection markers for plasmidmaintenance, inducible high-level expression systems, and chromo-somal insertion strategies (for a review, see de Vos, 1999b; Renault,2002). The transgene of interest can either be inserted into the chromo-some or cloned into a multicopy plasmid, the latter being sometimesnecessary to achieve very high levels of expression such as thoserequired for intracellular expression of antigen in live recombinant

USE OF RECOMBINANT LAB IN HUMAN HEALTH 29
LAB-based vaccines (Grangette et al., 2001). Complete food-gradeplasmid-based expression systems have been successfully developedin LAB that minimize the introduction of foreign DNA (self-clonedsystems) (de Vos, 1999b). In these systems, all components ideallyoriginate from the cloning host including replicons, the inducible orconstitutive expression/secretion cassette, as well as a reliable selec-tion system for plasmid introduction and maintenance.
The selection system is of crucial importance to ensure the stabilityof the transgenic strain during the fermentation processes that precedeadministration. Nonantibiotic food-grade markers developed for LABcan be grouped in two categories based on the selection method. Thefirst category includes so-called ‘‘dominant markers’’ that do not relyon specific host genes (versatile) and can be readily compared withantibiotic resistance markers. For example, the catabolism of specificsugars (melibiose, sucrose, xylose, starch, and inulin) offers variouspossibilities for the development of dominant selection markers in Lc.lactis and various lactobacilli (Boucher et al., 2002; Fitzsimons et al.,1994; Hols et al., 1994; Leenhouts et al., 1998; Posno et al., 1991).A second group in this category are genes conferring resistance orimmunity (e.g., immunity to bacteriocins [nisI, lafI]) and resistance tocycloserine (alr) and heavy metals (Cdr) (Allison and Klaenhammer,1996; Bron et al., 2002; Froseth and McKay, 1991; Liu et al., 1996;Takala and Saris, 2002; von Wright et al., 1990). Although dominantmarkers are convenient, most cannot be used at the industrial scale forsafety or economic reasons. Furthermore, their implementation some-times requires the transfer of numerous genes that can strongly affectplasmid stability or could result in slower growing recombinantstrains.
The second category includes the complementation markers result-ing from specific mutations in the host chromosome, thus permitting aspecific plasmid-host combination. Two types of complementationmarkers can be distinguished in LAB based either on sugar utilization(lactose) or on a specific auxotrophy (pyrimidine, thymine, D-alanine)(Bron et al., 2002; Fu and Xu, 2000; Hashiba et al., 1992; MacCormicket al., 1995; Platteeuw et al., 1996; Ross et al., 1990; Sorensen et al.,2000; Takala et al., 2003). A complementation system for lactoseutilization based on the lacF gene has been successfully applied toLc. lactis on an industrial scale (Kleerebezem, personal communica-tion; Platteeuw et al., 1996). This system, however, uses a growthmedium that utilizes lactose as a carbon source, which is not alwaysdesirable for implementation in other LAB. Complementation markersbased on pyrimidine or thymine auxotrophy have also been described

30 HANNIFFY et al.
that exploit the amber suppressor tRNA gene (supD) and thymidilatesynthase gene (thyA), respectively (Fu and Xu, 2000; Sorensen et al.,2000), but again, the requirement for dedicated growth media withoutthese specific compounds represents a major drawback. More re-cently the use of the alanine racemase gene (alr) as a complementationmarker was reported as functional in both Lc. lactis and Lb. plantarum(Bron et al., 2002). Alanine racemase converts L-alanine (L-Ala) intoD-alanine (D-Ala), which is essential in cell wall biosynthesis as itis involved in the cross-linking of peptidoglycan. Alanine racemase-deficient strains of both LAB are strictly auxotrophic for D-Ala andundergo cell lysis when starved of D-Ala (Fig. 4) (Bron et al., 2002; Holset al., 1997a, 1999). Because D-Ala is mainly present in the cell wallsof bacteria and concentrations of this compound are extremely lowin most growth media, the alr gene offers greater potential as a com-plementation marker. Significantly, the alr gene can also be usedas a dominant marker, since high alanine racemase expression con-fers cycloserine resistance (Bron et al., 2002). In addition to D-Ala, thepeptidoglycan contains other unique compounds (D-glutamate,D-aspartate, meso-diaminopimelate) that are essential and can alsobe used to supplement growth medium. In the future these compoundsand their genes could potentially represent a range of complementationmarkers that could be used for the development of new food-gradeplasmid maintenance or chromosomal integration systems.
Although complete food-grade systems based on plasmids have beensuccessfully developed, they often demonstrate intrinsic structural in-stability and display a high level of potential horizontal gene transferthrough mechanisms such as plasmid co-integration and conjugation.To minimize these problems, several food-grade chromosome deliverysystems have now been developed for LAB based on homologous recom-bination (recA-dependent), site-specific recombination (phage-derivedsystems), retrotransposition (group II intron-derived systems), andtransposition (insertion sequence-based systems) (de Vos, 1999b;Frazier et al., 2003; Hols et al., 1994; Maguin et al., 1996; Martin et al.,2000; Romero and Klaenhammer, 1991). With the exception of homolo-gous recombination, all of these delivery systems are based on geneticelements that contribute to genome plasticity and/or horizontal trans-fer and must be avoided for safe use. Chromosomal integration byhomologous recombination can be achieved by a single crossoverevent (Campbell integration with one homologous region) with a non-replicating plasmid (suicide vector). Food-grade systems based on thisapproach have been developed for Lc. lactis by using endogenousplasmids devoid of their repA replication protein (pORIþ plasmids)
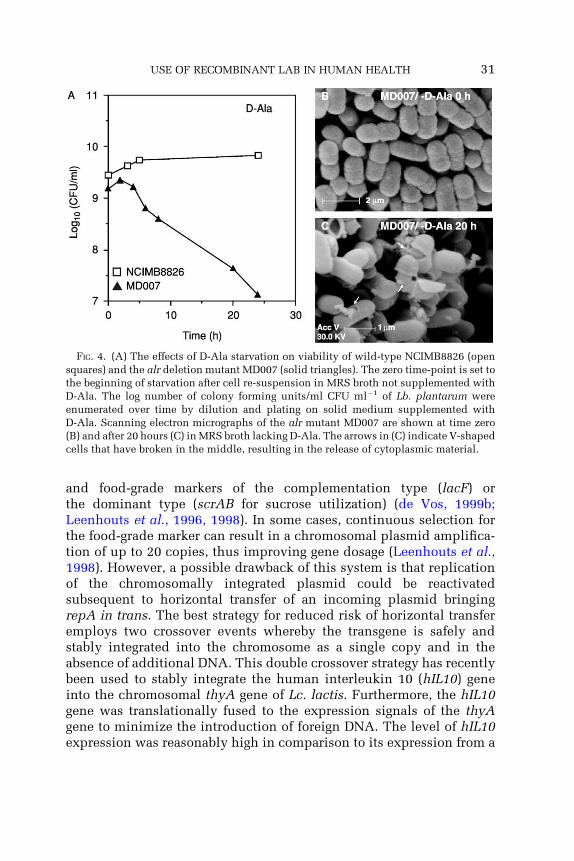
FIG. 4. (A) The effects of D-Ala starvation on viability of wild-type NCIMB8826 (open
squares) and the alr deletion mutant MD007 (solid triangles). The zero time-point is set to
the beginning of starvation after cell re-suspension in MRS broth not supplemented with
D-Ala. The log number of colony forming units/ml CFU ml�1 of Lb. plantarum were
enumerated over time by dilution and plating on solid medium supplemented with
D-Ala. Scanning electron micrographs of the alr mutant MD007 are shown at time zero
(B) and after 20 hours (C) in MRS broth lacking D-Ala. The arrows in (C) indicate V-shaped
cells that have broken in the middle, resulting in the release of cytoplasmic material.
USE OF RECOMBINANT LAB IN HUMAN HEALTH 31
and food-grade markers of the complementation type (lacF) orthe dominant type (scrAB for sucrose utilization) (de Vos, 1999b;Leenhouts et al., 1996, 1998). In some cases, continuous selection forthe food-grade marker can result in a chromosomal plasmid amplifica-tion of up to 20 copies, thus improving gene dosage (Leenhouts et al.,1998). However, a possible drawback of this system is that replicationof the chromosomally integrated plasmid could be reactivatedsubsequent to horizontal transfer of an incoming plasmid bringingrepA in trans. The best strategy for reduced risk of horizontal transferemploys two crossover events whereby the transgene is safely andstably integrated into the chromosome as a single copy and in theabsence of additional DNA. This double crossover strategy has recentlybeen used to stably integrate the human interleukin 10 (hIL10) geneinto the chromosomal thyA gene of Lc. lactis. Furthermore, the hIL10gene was translationally fused to the expression signals of the thyAgene to minimize the introduction of foreign DNA. The level of hIL10expression was reasonably high in comparison to its expression from a

32 HANNIFFY et al.
multicopy plasmid (Steidler et al., 2000, 2003). It is now likely thatsuch a strategy will be used increasingly for the genetic engineering ofLAB for other medical applications.
B. BIOLOGICAL CONTAINMENT SYSTEMS
Biological containment systems can be subdivided into two groups:active systems and passive systems. Active containment is based onthe conditional genetic control of either a killing gene (activation) or anessential gene (repression). Various killing genes have been identifiedin LAB such as those encoded by phage lytic cassettes (lysin or lysin/holin combination) and DNA restriction systems (de Ruyter et al.,1997; Djordjevic and Klaenhammer, 1996, Djordjevic et al., 1997;Sanders et al., 1997). While a large number of essential genes couldpotentially be exploited, peptidoglycan biosynthetic genes are particu-larly attractive, because their inactivation usually results in cell lysis(Curtiss III et al., 1977). Proof of the concept for lysis inductionwith peptidoglycan hydrolases (lysins) and biosynthetic genes (alr;D-ala-D-ala ligase, ddl) tightly regulated by the nisin-inducible systemhas already been achieved in the laboratory by using Lc. lactis and Lb.plantarum (Bron et al., 2002; LABDEL project; Prozzi and Fontaine,unpublished data). To be implemented in humans, these active contain-ment systems would require expression signals that are tightly con-trolled by environmental parameters. These expression signals couldeither be activated/repressed in the host or subsequent to release of therecombinant strain into the environment. A range of expression cassetteshave been isolated from Lb. plantarum that are specifically activated inthe GI tract of mice (Bron et al., 2004a; LABDEL project) (see Section VII).These expression systems are now being exploited in Lb. plantarum tospecifically induce lysis and consequent release of a biomolecule ofinterest (therapeutic molecules) into the GI tract via the control of lyticcassettes. A drawback to using active containment systems is that muta-tions can occur that can inactivate a killing gene/compound or result inconstitutive expression of the essential gene (Hols, P., unpublished data;Molina et al., 1998; Szafranski et al., 1997). However, it may be possibleto minimize these problems by combining more than one system in adefined recombinant strain (Ronchel and Ramos, 2001).
Passive containment systems, by contrast, are robust and very sim-ple and based mainly on complementation of an auxotrophy by sup-plementing an essential metabolite. This metabolite is ideally notavailable or occurs in extremely low amounts in the environment.Furthermore, if the compound must be added to growth medium for

USE OF RECOMBINANT LAB IN HUMAN HEALTH 33
inoculum preparation, it is preferable that it occurs at a low cost tofacilitate future industrial applications. Several mutant strains used forplasmid maintenance via complementation markers also offer potentialas passive containment systems. These include two auxotrophies thathave a high potential for human application since they are bactericidal.Alanine racemase mutants that demonstrate a requirement for D-Alacan be obtained in a large number of bacteria including LAB (Bronet al., 2002; Hols et al., 1997a). D-Ala starvation results in cell death viaa lysis process, and experiments are currently underway for contain-ment of S. mutans GMMs designed for anti-caries therapy (Hillman,2002). The second bactericidal auxotrophy is based on a thyA mutantof Lc. lactis that has recently been evaluated for its efficacy in a pigmodel (Steidler et al., 2003). Thymine starvation results in activation ofthe SOS repair system and DNA fragmentation, thus constituting anintrinsic suicide system (Curtiss et al., 1977). Exploiting the thyA geneas a passive containment system by replacing it with a transgene ofinterest offers additional advantages in terms of biosafety (Steidleret al., 2003). While any reversion of the thyA mutation through homol-ogous recombination with a Lc. lactis thyA gene that might be acquiredby horizontal transfer would restore a wild-type phenotype, it shouldalso result in loss of the transgene. Furthermore, thyA mutants of Lc.lactis are severely impaired in phage replication, further reducing therisk of horizontal transfer by phage transduction (Pedersen et al., 2002).This elegant system has recently been applied to the delivery of hIL10by live Lc. lactis bacteria and has now been approved for use in humans(Steidler et al., 2003).
VII. Opportunities and Potential Applications of Future Research
A. INSIGHTS FROM GENOME SEQUENCING AND COMPARATIVE GENOMICS
The relatively small genome size of bacteria, combined with the avail-ability of high-throughput sequencing facilities, has stimulated the de-termination of many bacterial genomes. Over the past decade, thesequences of more than 90 bacterial genomes have become available inthe public domain. These efforts have focused primarily on pathogenicbacteria and have included the completion of several genome se-quences of food-borne pathogens (Schoolnik, 2002; Wells and Bennik,2003). However, in more recent times, the genomes of food-gradebacteria have received considerable attention and genome sequences(including partial sequences) are now available for more than 20 LABas well as a number of related species (Klaenhammer et al., 2002).

FIG. 5. Phylogenetic tree constructed by using 16 S rDNA sequences of LAB and related
bacteria that have been completely or partially analyzed at the genomic level. It should be
noted that for a number of species the genome sequence has (partially) been determined
for more than one strain of this species. The estimated genome sizes are indicated
between brackets. Complete genomic sequences (and thus their exact sizes) are available
for the paradigm Gram-positive bacterium Bacillus subtilis (Kunst et al., 1997) and the
LAB, Lactococcus lactis subspecies lactis IL1403 (Bolotin et al., 2001) and Lactobacillusplantarum WCFS1 (Kleerebezem et al., 2003).
34 HANNIFFY et al.
The first complete LAB genome sequence to be published was that ofLc. lactis subspecies lactis strain IL1403 (Bolotin et al., 2001). Morerecently, a high-fidelity genome sequence has also been published forLb. plantarum strain WCFS1 (Kleerebezem et al., 2003). In addition,more LAB genomes are nearing completion (Fig. 5; for a review seeKlaenhammer et al., 2002) including draft genome information for anumber of LAB that was made available in the public domain in2002 by the Joint Genome Institute (ftp://ftp.jgi-psf.org/pub/JGI_data/Microbial/) in collaboration with the lactic acid bacteria genomicsconsortium.
Comparative genomics will be used to determine the evolutionaryrelationships between different species of LAB and allow analysis ofgenomic diversity among different strains and closely related species.Studies on pathogenic bacteria have already shown that comparativegenome analysis and microarray-based studies of genome composi-tion can indeed reveal genetic factors linked to specific functions or

USE OF RECOMBINANT LAB IN HUMAN HEALTH 35
plasticity regions associated with lifestyle properties of specific strains(Israel et al., 2001; Pearson et al., 2003; Porwollik et al., 2002; Salamaet al., 2000). In the case of LAB, it is already well established that somespecies are genetically diverse and that strain adaptation is rapid.Moreover, horizontal gene transfer appears to be the rule rather thanthe exception and is especially important for genes or functions relatedto mobile genetic elements such as plasmids, transposons, and bacter-iophages. Recently, genomic variation within a single LAB species wasexemplified by a detailed genome diversity study of Lb. plantarumisolates using DNA microarrays (Molenaar et al., 2003). In addition tohigh levels of genome diversity, Molenaar et al. (2003) also identifiedspecific genomic regions of relatively high plasticity that may representlifestyle adaptation islands. These findings show that genomic diversi-ty is a key determinant of bacterial functionality and indicate thatspecific approaches are required to fully exploit the collective genomicpotential of LAB.
B. THE BEHAVIOR OF LAB IN THE HOST
The human GI tract is colonized by a vast, complex, and dynamicpopulation of commensal microorganisms including food-associatedbacteria that contribute to nutrient processing, affect the host’s immunefunction, and stimulate a variety of other host activities. Molecularapproaches based on 16 S rDNA sequencing and profiling of dominantGI tract microbiota have revealed that the majority of commensalsbelong to unknown, ‘‘novel’’ bacterial species that so far have not beenstudied under laboratory conditions (Zoetendal et al., 1998). In addi-tion, these studies revealed that while the microbial composition wasrelatively stable in individual adults, it varies significantly betweendifferent individuals (Zoetendal et al., 1998, 2002). Intriguingly, thecomposition of dominant GI tract microbiota was also shown to beaffected by specific host-microbe interactions that appear to be relatedto the host genotype (Zoetendal et al., 2001). Similar molecular ap-proaches as well as classical cultivation experiments have also shownthat several food-grade bacteria are encountered as natural inhabitantsof the human GI tract. In particular, certain Lactobacillus species arefrequently found among the natural intestinal microbiota and wouldappear to be relatively numerous in the upper regions of the smallintestine (Ahrne et al., 1998; Heilig et al., 2002; Walter et al., 2000,2001). The increasing availability of genome sequences for severalintestinal species of LAB opens new avenues to study their functional-ity in the gut, including interactions with host cells. For example,

36 HANNIFFY et al.
when ingested, Lb. plantarum WCFS1 was shown to effectively survivepassage of the human stomach and to reach the ileum in high numbersas compared with other LAB strains (Vesa et al., 2000). In addition, thebacterium could also be detected in the colon. These characteristicsand the availability of its genome sequence (Kleerebezem et al., 2003)render this strain a suitable candidate for the study of bacterial behav-ior in the GI tract.
To date, DNA microarray and proteomic studies carried out inLAB have been confined to in vitro experiments investigating globalregulatory responses to different environmental stimuli (Champomier-Verges et al., 2002; Guillot et al., 2003; van de Gucht et al., 2002).Similarly, studies investigating host-LAB interaction have focused pri-marily on physiological aspects such as intrinsic levels of acid andbile tolerance (Chou and Weimer, 1999; Hyronimus et al., 2000) andthe development of complex media to selectively enrich for LAB spe-cies that are tolerant to digestive stress (Shah, 2000). For example, agenome-wide genetic screen carried out in Lb. plantarum identified31 open reading frames (ORFs) whose expression would appear to beinduced by bile acids, including genes encoding efflux pumps as wellas several membrane and cell wall–associated functions (Bron et al.,2004b,c). While such in vitro experiments might unravel responsesby specific microorganisms to certain GI-tract conditions, they willnot suffice in portraying bacterial behavior in the GI tract. The fullresponse repertoire will be triggered only in vivo, where all physico-chemical conditions are combined with specific host-microbe andmicrobe-microbe interactions. This notion has led to development ofmore sophisticated in vivo approaches aimed at identifying bacterialgenes that are important during residence in the GI tract.
Three main strategies have been developed for the identification ofgenes that are highly expressed in vivo as compared to laboratoryconditions, that is, (recombination-based) in vivo expression technolo-gy ((R-)IVET), signature tagged mutagenesis (STM), and selective cap-ture of transcribed sequences (SCOTS). The basic characteristics andrelative (dis)advantages of each of these approaches have recently beenreviewed (Mahan et al., 2000), but because of their recent applicationin LAB, (R-)IVET approaches will be explored in somewhat more detailhere. To date, four variations of IVET utilizing different reporter geneshave evolved that allow the selection of genomic sequences that harborpromoters that are specifically induced in situ, for example, duringGI-tract passage (for a review see Angelichio and Camilli, 2002; Mahanet al., 2000). The (R-)IVET approach relies on the generation of tran-scriptional fusions of random genomic fragments to a promoterless

USE OF RECOMBINANT LAB IN HUMAN HEALTH 37
reporter gene encoding an enzymatic activity. In the decade followingthe first report describing IVET technology, this technique has solelybeen used for the identification of genes that were up-regulated duringthe infection processes of a number of pathogenic bacteria (Angelichioand Camilli, 2002; Mahan et al., 2000). Recently, however, two reportshave described for the first time, the application of (R-)IVET technologyto food-grade LAB. An IVET strategy based on in vivo selection of anantibiotic-resistant phenotype was used to identify three in vivo in-duced (ivi) genes in Lb. reuteri that were expressed at induced levelsduring colonization of the GI tract of reconstituted Lactobacillus-freemice (Walter et al., 2003). Interestingly, one of these genes encoding apeptide methionine sulfoxide reductase has previously been identifiedby using IVET in the non-food-associated bacterium S. gordonii duringexperimental endocarditis (Kili et al., 1999). Although not noticed bythe authors, this finding gives a first glimpse into potential overlap thatmay exist between genetic responses triggered by both pathogenic andnon-pathogenic bacteria subsequent to host interaction. A disadvan-tage of this approach, however, is that it required antibiotic selectionpressure during host transit, which was achieved by administeringantibiotic to the mice during the course of the experiment. This treat-ment would have been detrimental to native intestinal microbiotaalready present in these mice, and it is highly likely that the in vivoconditions encountered by Lb. reuteri would have been significantlychanged.
More recently, Bron et al. (2004a) have reported the successful ap-plication of a second (R-)IVET approach for the identification of ivigenes in Lb. plantarum. In contrast to other IVET systems based onselection of an antibiotic marker, this approach employs the irrevers-ible enzymatic activity of resolvases as a reporter and screening tool.In this system, an antibiotic resistance marker flanked by two resol-vase-recognition sites (Fig. 6: loxP-ery-loxP cassette) is integrated intothe chromosome of the bacterium of interest. In addition, a promoter-less copy of the corresponding resolvase-encoding gene (Fig. 6: cre) isintroduced on a plasmid and used as a reporter to trap transcriptionalactivation (promoter activity) by monitoring changes in the antibioticresistance phenotype. Since promoter activation leads to irreversibleexcision of the antibiotic resistance marker, this strategy does not relyon selective pressure during animal experiments. This form of (R-)IVETcan therefore be used to monitor induction of bacterial gene expressionin a live host possessing an intact unmodified GI tract. Moreover,this strategy is the only IVET approach that functions as a geneticscreen as it discards all promoters displaying activity under laboratory

FIG. 6. Schematic representation describing the (R)-IVET screen performed with
Lactobacillus plantarum (Bron et al., 2004a). A loxP-ery-loxP cassette is introduced in
the Lactobacillus plantarum chromosome. A library of random Lactobacillus plantarum
chromosomal fragments is constructed in pIVET-cre, upstream of a promoterless copy of
the cre resolvase gene. Expression of cre leads to excision of the loxP-ery-loxP cassette
and erythromycin sensitivity. An erythromycin-resistant sub-library is selected under
laboratory conditions and administered to conventional mice. The complete sub-library
is recovered from fecal samples by selection for the pIVET-cre encoded chloramphenicol
resistance and replicated to select erythromycin-sensitive clones (CmR, EmR). These
clones contain pIVET-cre derivatives that harbor a GI tract activated promoter in the
Lactobacillus plantarum chromosomal fragment cloned upstream of cre, since loxP-ery-
loxP excision has occurred during transit through the host intestine. Sequencing of the
pIVET-cre inserts is used to identify the corresponding ivi genes of Lactobacillusplantarum (Bron et al., 2004a).
38 HANNIFFY et al.
conditions, since the (R-)IVET library is prepared under antibioticselection pressure prior to administration to the host (Fig. 6). Applica-tion of this genetic screen in Lb. plantarum resulted in the identifica-tion of 72 genes that were induced during passage of the GI tract inconventional mice (Bron et al., 2004a). A diverse range of functions waspredicted for these 72 ivi-genes, including nutrient acquisition, inter-mediate and/or co-factor biosynthesis, stress response, cell surfaceproteins, and a number of (conserved) hypothetical proteins. Remark-ably, one of the hypothetical proteins identified in this study displayssignificant homology (32% identity) to the conserved hypothetical pro-tein that was identified by using IVET in Lb. reuteri (Bron et al., 2004a).Moreover, a large number of the functions and pathways identified

USE OF RECOMBINANT LAB IN HUMAN HEALTH 39
by (R-)IVET in Lb. plantarum have also been described in pathogenicbacteria, where they have also been shown to play an important roleduring infection. The striking parallels that exist between ivi responsesexhibited by pathogens and nonpathogens in vivo may well be ex-plained by the importance of these genes for survival rather thanpathogenesis during host residence (Bron et al., 2004a).
These studies represent an important step toward elucidating thebehavior of food-grade LAB in the complex environment they encoun-ter after ingestion by the consumer. Approaches such as IVET willprovide the genetic tools necessary, including the required promoters,that will allow the development of dedicated LAB-based delivery ve-hicles that will express only desired pharmaceutical proteins in situ.Moreover, these approaches should provide a more geographical andprecise insight into the exact locations of ivi-gene activation in the GItract that may allow the construction of a new generation of site-specif-ic delivery vehicles. Eventually, ivi-promoters could be combined withcertain genes (e.g., bacteriophage-derived lytic cassettes) to developLAB delivery systems that exhibit controlled release of their cellularcontent (including desirable molecules) at a specific location in the GItract. IVET and other genomic-based approaches will also provide abetter understanding of LAB behavior in the GI tract as well as in otherenvironmental niches. They should facilitate the genetic engineering ofmutant strains that can be compared with their wild-type counterpartsin appropriate in vitro and in vivo models. Ultimately, it is hoped thatthese approaches will help define potential probiotic functions andprovide a molecular basis to support and explain health benefits thatare associated with LAB and related species.
C. THE HOST RESPONSE TO LAB
The ability of LAB to alter the function of the systemic and immuneresponses has been the subject of intense research over the last fewyears. Some LAB have been shown to translocate across the mucosalepithelium of the GI tract, allowing them to come into direct contactwith the underlying immune cells and influence immune responses.There is also evidence that some LAB can directly stimulate the im-mune system at the gut mucosal surface via localized GI tract lymphoidcell loci, increasing lymphocyte populations and cell surface expres-sion in the gut-associated lymphoid tissue (GALT) environment, andfacilitate increased immunoglobulin output into the intestinal lumen.It is also becoming increasingly clear that interactions of different LABwith host cells in the mucosa can have different effects on immune

40 HANNIFFY et al.
function. Administration of both viable and/or heat-killed preparationsof the probiotic Lb. acidophilus strain UFV-H2b20 has been shownto enhance phagocytic activity in germ-free mice and to improve clear-ance capacity against circulating E. coli as compared with controls(Neumann et al., 1998). Experiments carried out in mice have shownthat oral administration of different LAB increased IgA-producing cellsassociated with the lamina propria (Vitini et al., 2000). However, thisincrease did not always correlate with an increase in CD4 þ T lympho-cytes, suggesting that these bacteria only induced clonal expansion ofcells in the lamina propria that were already triggered to produce IgA.In addition, while some LAB did increase macrophages, neutrophilsand eosinophils indicative of an inflammatory response, CD8 þ T cellpopulations were diminished or not affected. Other studies carried outin animals and humans have also shown that the administration ofvarious Lactobacilli and Bifidobacteria can significantly increase totalIgA as well as antigen or pathogen-specific IgA, thereby contributing tomucosal resistance to gastrointestinal pathogens (Cukrowska et al.,2002; Fang et al., 2000; Fukushima et al., 1998; Herias et al., 1999;Tejada-Simon et al., 1999). In one of these studies, co-administration ofLb. plantarum and an E. coli strain to germ-free rats resulted in signifi-cantly increased densities of CD25 þ cells in the lamina propria anddecreased proliferative spleen cell responses to E. coli (Herias et al.,1999). CD 25 þ cells have been shown to be involved in tolerance anddown-regulation of immune responses to self and nonself antigens.While the production of IgA may be important in preventing infectionsby pathogens, it may also be considered anti-inflammatory because itinvolves mechanisms controlled by Th2 effector cytokines that aremore associated with tolerance. These studies have been supportedby work carried out by Kirjavainen et al. (1999a) that showedthat administration of selected LAB strains could reduce T cell reactiv-ity in mice in a dose-dependent fashion (Kirjavainen et al., 1999a,b).Similarily, in vitro experiments have shown that LAB or their compo-nents can inhibit mitogen- and antigen-induced T lymphocyte prolif-eration and down-regulate both Th1 and Th2 effector cytokines (Chenet al., 2002; Pessi et al., 1999; von der Weid et al., 2001). In addition,LAB have been reported to induce IL-10 production in gut epitheliumcells (Haller et al., 2000) and in peripheral blood mononuclearcells isolated from both animals and humans (Miettinen et al., 1996;Vinderola et al., 2004). Interestingly, von der Weid et al. (2001)also identified a Tr1-like population of CD4þ T cells with lowproliferative capacity that produced substantial levels of the regu-latory cytokines IL-10 and TGF-�. These studies would indicate that

USE OF RECOMBINANT LAB IN HUMAN HEALTH 41
mechanisms controlled by LAB may play an important role in main-taining immune homeostasis and, in particular, maintaining toleranceto the indigenous microbial population. This is supported by experi-ments in animals that show that select strains of LAB induce IL-10 andsuppress Th1-related immunopathologies such as IBD.
Conversely, LAB have also been shown to increase lymphocyte prolif-eration as well as the production of inflammatory cytokines. Experi-ments carried out by Aattour et al. (2002) in pathogen-free rats showedthat oral ingestion of yogurt containing S. thermophilus and Lb. bulgar-icus resulted in increased in vitro proliferation of lymphocytes isolatedfrom peripheral blood, spleen, and Peyer’s patches that, for the two latterlymphoid compartments, were accompanied by increased interferon- �production. Similarly, Gill et al. (2000) showed that feeding various LABstrains to mice significantly enhanced mitogen-induced proliferation ofsplenocytes and production of IFN- �, increased phagocytic activityof peripheral blood leukocytes and peritoneal macrophages, and en-hanced serum antibody responses to antigen administered by the oraland systemic routes. Moreover, numerous studies have also shown thatcertain LAB or their components are potent inducers of Th1 cytokines(Cleveland et al., 1996; Kato et al., 1999; Maassen et al., 2000) and candecrease Th2 responses (Marshall et al., 1995; Murosaki et al., 1998;Repa et al., 2003; Shida et al., 2002). Similarly, studies carried out inanimals and humans have demonstrated that ingestion of dietary LABcan prevent atopy and reduce the development of allergy (Bjorkstenet al., 1999; Kalliomaki et al., 2001; Repa et al., 2003; Sepp et al., 1997;Simhon et al., 1982). These studies would indicate that LAB canenhance natural and acquired immunity and may be beneficial in opti-mizing or enhancing immune responses in healthy and immunocom-promised individuals (Gill et al., 2000). In addition, counter-regulatoryproperties based on their Th1-inducing capacities could provide a use-ful strategy against Type I allergy (Repa et al., 2003) (see also Section III).
It has been demonstrated that different strains of LAB have substan-tially different capacities to induce IL-12 and TNF-� production indendritic cells (DCs) (Christensen et al., 2002). In addition to exertingless-pronounced effects on IL-6 and IL-10 production, all strains up-regulated expression of surface MHC class II and B7-2 indicative of DCactivation and maturation. In a separate study, significant functionaland phenotypic dichotomy was also observed in DCs exposed eitherto Lb. rhamnosus or Klebsiella pneumoniae (Braat et al., 2003). Inter-estingly, compared to K. pneumoniae, Lb. rhamnosus induced lowerTNF-�, IL-6, and IL-8 production in immature DCs as well as lower IL-12 and IL-18 production in mature DCs that were also associated

42 HANNIFFY et al.
with the development of T cells without a typical Th phenotype.Significantly, Christensen et al. (2002) also showed that at least onespecies of Lactobacillus was capable of inhibiting the activities of otherLactobacillus species with respect to DC activation and production ofcytokines. These studies demonstrated that gut microflora, and inparticular LAB, can differentially modulate immune responses by sti-mulating DC activation and function, which in turn may determinewhether the DC population may favor a Th1 or Th2 response as well astolerance.
It is now clear that different LAB possess intrinsically differentimmunoadjuvant capacities that can play a significant role in modulat-ing immune responses. Germ-free animal models will help elucidatethe interactions that occur between lactic acid bacteria and the host.This includes their ability to induce innate immunity that has not yetbeen appropriately compared in germ-free and conventional animals.Like other commensals, lactic acid bacteria will express various com-mensal associated molecular patterns (CAMPs) that are able to recog-nize and activate mucosal cells (including epithelial cells) throughpattern recognition receptors (Funda et al., 2001; Janeway andMedzhitov, 2002). Recombinant mutants could be compared withwild-type strains to identify specific components, lipoteichoic acidand peptidoglycan for example, which may be responsible for distinctimmune responses. The availability of genomic data will acceleratethese efforts by facilitating the identification of other key bioactivecomponents such as those displayed on the bacterial surface. Combin-ing post-genomic approaches such as microarrays and proteomics withGF animal models and selective colonization strategies could improveunderstanding of host-commensal interactions and bring new insightsinto the mechanisms of mucosal immunity. These efforts can also becombined with ex vivo approaches such as laser microdissection tech-niques that will allow us to examine individual cellular components aswell as general cell populations from different anatomical regionsof the immune system. Similarly, these tools can be applied to animalmodels of human diseases developed under defined gnotobioticconditions and may help elucidate the etiology of frequent disorderssuch as those associated with several infectious, inflammatory, auto-immune and neoplastic diseases. These approaches may help to ex-plain why lactic acid bacteria and other commensals do not triggerpathological inflammatory responses in mucosal tissues in normalhosts and may answer intriguing questions regarding their associatedbenefits.

USE OF RECOMBINANT LAB IN HUMAN HEALTH 43
VIII. Concluding Remarks
The last decade has seen increasing interest in the use of recombinantLAB as mucosal delivery vehicles for vaccines and other therapeuticmolecules. It is now clear that sufficient advances in the genetics of LABhave made it possible to construct safe LAB-based recombinant vaccinesthat are capable of eliciting protection against lethal challenge withtoxin or a human pathogen in a relevant disease model. Although multi-ple doses of LAB vaccines are currently needed to afford immune pro-tection by the intranasal and oral routes of administration, this shouldnot preclude their suitability for use in humans. Multiple doses of theinjected DTP and oral polio vaccines (OPV) are already given to infantsin the first several months of life and have the potential to be combinedwith LAB vaccines. Moreover, this technology is relatively inexpensiveand could be applied successfully to mass-vaccination programs that areparticularly relevant in developing countries. There are also opportu-nities to enhance the efficacy of LAB vaccines through increased antigenexpression or through the combined delivery of multiple immunogensand specific adjuvants. Further insights may be gained through directcomparisons of LAB strains with different persistence and survivalcharacteristics or immunostimulatory properties with different immu-nization routes or schedules against a selected target disease. Geneticengineering clearly has the potential to further optimize the survivalcharacteristics of selected LAB, define optimal placement and dosageregimes in different clinical settings, and enhance their ability to delivera pharmaceutical protein. Ultimately, we hope that LAB-based deliverytechnology will add a new dimension to vaccine development withrespect to safety, potential for use in continual vaccination stratagemsthat encompass both control and eradication of a disease, and increasedflexibility and potential use against a wide range of diseases.
Opportunities for developing more effective LAB vaccines may alsoarise from ongoing research on the fate of LAB in the human host andthe interaction of LAB with the cells and tissues of the immune system.Genetic engineering may help elucidate the mechanisms by whichprobiotic strains of LAB exert positive effects on human health andfacilitate the identification of the key bioactive components responsi-ble for conferring probiotic traits to these bacteria. These efforts willbe further supported by ongoing developments in genomic andpostgenomic-based approaches and use of appropriate animal models.As well as providing intriguing answers to questions regarding theirtherapeutic role in conditions such as allergy and IBD, these studiesmay help clarify the effect of nutrition and genetic background on the

44 HANNIFFY et al.
development of these diseases to establish the true role of these bacte-ria in human health. It is foreseen that these approaches will enable theselection of appropriate species that might prove most beneficial undercertain conditions and facilitate the identification of new probiotics.In addition, it should be possible to determine optimal probioticcompositions, ensuring that they demonstrate synergistic rather thancompeting activities. Most importantly, these approaches will helpestablish proven criteria to substantiate the sometimes ‘‘vague’’ healthclaims of currently available probiotic preparations. In doing so, thesecriteria will lead to the possibility of creating custom-designed probio-tics that are scientifically proven, increase their range of potentialapplications, and accelerate their development into health products.
Currently,many probiotic strainsofLABaremarketedandregulatedasconventional foods. The regulatory environment surrounding these pro-ducts is diverse, but national differences are being harmonized by ongo-ing European legislation. For example, the 1997 novel foods regulation ofthe EU parliament governs new food organisms and makes recommenda-tions for assessment of foods containing genetically modified organisms(GMOs). In principle, this framework governs new recombinant strainsof probiotic microorganisms and requires information on toxicology,genetic stability, potential for genetic transfer in the host, and the effectof the GMO in humans. Specific health claims for the product would alsoneed to be substantiated by studies in humans resembling what is re-quired by regulatory frameworks governing human pharmaceutical pro-ducts. The future of recombinant LAB as novel therapies for humans canbe foreseen where the potential benefits are significant, particularlywhere the absence of satisfactory treatments for a particular diseasenecessitates the developmentofnew andinnovative therapies. However,there is an urgent need for clarification to differentiate food supplementsfrom medicines for product applications based on probiotic organisms.Aswellasensuring theexpansion ofboth these applications, appropriatelegislation would allay fears within the food supplement industry andaccelerate the development of health products based on recombinantLAB. It is now clear that biotechnological applications of LAB over thepast decade has meant that many food-grade systems for genetic modifi-cation are in fact already available, including methods for chromosomalintegration and controlled high-level gene expression. Moreover, therecent approval of a recombinant strain of Lc. lactis expressing IL-10 foruse in trials with human IBD patients provides further encouragement.We are therefore optimistic that as well as the continued development ofprototype health products based on recombinant LAB, the next decadewill see more of these reach the marketplace as finished products.

USE OF RECOMBINANT LAB IN HUMAN HEALTH 45
ACKNOWLEDGMENTS
The Partners of the LABDEL consortium are grateful for financial support from
The European Framework 5 Programme (EC contract QLK3-CT-QLK3-CT-2000-00340).
The authors also wish to thank Andy Carter, Wendy Glennison, Sally M. Hoffer, Maria
Marco, Esther van Mullekom, Douwe Molenaar, Willem M. de Vos, Anne Mette Wolff,
Astrid Vrang, Renata Stepankova, Deborah Prozzi, Marie Deghorain, Laetitia Fontaine,
Vassilia Therodorou, Heimo Breiteneder, and Michael Hisbergues for their contribution
to the work described. We are also grateful to Therese Hall for help with coordination of
the LABDEL project. Pascal Hols is scientific collaborator at FNRS.
REFERENCES
Aattour, N., Bouras, M., Tome, D., Marcos, A., and Lemonnier, D. (2002). Oral ingestion of
lactic-acid bacteria by rats increases lymphocyte proliferation and interferon-gamma
production. Br. J. Nutr. 87, 367–373.
Adams, M. R., and Marteau, P. (1995). On the safety of lactic acid bacteria from food. Int.
J. Food Microbiol. 27, 263–264.
Ahrne, S., Nobaek, S., Jeppsson, B., Adlerberth, I., Wold, A. E., and Molin, G. (1998). The
normal Lactobacillus flora of healthy human rectal and oral mucosa. J. Appl.Microbiol. 85, 88–94.
Aiba, Y., Suzuki, N., Kabir, A. M., Takagi, A., and Koga, Y. (1998). Lactic acid-mediated
suppression of Helicobacter pylori by the oral administration of Lactobacillus sali-varius as a probiotic in a gnotobiotic murine model. Am. J. Gastroenterol. 93,2097–2101.
Allison, G. E., and Klaenhammer, T. R. (1996). Functional analysis of the gene encoding
immunity to lactacin F, lafI, and its use as a Lactobacillus-specific, food-grade
genetic marker. Appl. Environ. Microbiol. 62, 4450–4460.
Alvarez, S., Herrero, C., Bru, E., and Perdigon, G. (2001). Effect of Lactobacillus casei and
yogurt administration on prevention of Pseudomonas aeruginosa infection in young
mice. J. Food Prot. 64, 1768–1774.
Angelichio, M. J., and Camilli, A. (2002). In vivo expression technology. Infect. Immun.
70, 6518–6523.
Araki, Y., Fujiyama, Y., Andoh, A., Koyama, S., Kanauchi, O., and Bamba, T. (2000). The
dietary combination of germinated barley foodstuff plus Clostridium butyricumsuppresses the dextran sulfate sodium-induced experimental colitis in rats. Scand.
J. Gastroenterol. 35, 1060–1067.
Babbs, C. F. (1992). Oxygen radicals in ulcerative colitis. Free. Radic. Biol. Med. 13,169–181.
Bauer, L., Bohle, B., Jahn-Schmid, B., Wiedermann, U., Daser, A., Renz, H., Kraft, D., and
Ebner, C. (1997). Modulation of the allergic immune response in BALB/c mice by
subcutaneous injection of high doses of the dominant T cell epitope from the major
birch pollen allergen Bet v 1. Clin. Exp. Immunol. 107, 536–541.
Beninati, C., Oggioni, M. R., Boccanera, M., Spinosa, M. R., Maggi, T., Conti, S., Magliani,
W., De Bernardis, F., Teti, G., Cassone, A., Pozzi, G., and Polonelli, L. (2000).
Therapy of mucosal candidiasis by expression of an anti-idiotype in human com-
mensal bacteria. Nat. Biotechnol. 18, 1060–1064.
Bermudez-Humaran, L. G., Cortes-Perez, N. G., Le Loir, Y., Gruss, A., Rodriguez-Padilla,
C., Saucedo-Cardenas, O., Langella, P., and Montes de Oca-Luna, R. (2003). Fusion to

46 HANNIFFY et al.
a carrier protein and a synthetic propeptide enhances E7 HPV-16 production and
secretion in Lactococcus lactis. Biotechnol. Prog. 19, 1101–1104.
Bermudez-Humaran, L. G., Langella, P., Miyoshi, A., Gruss, A., Guerra, R. T., Montes de
Oca-Luna, R., and Le Loir, Y. (2002). Production of human papillomavirus type 16 E7
protein in Lactococcus lactis. Appl. Environ. Microbiol. 68, 917–922.
Bernasconi, E., Germond, J. E., Delley, M., Fritsche, R., and Corthesy, B. (2002). Lactoba-
cillus bulgaricus proteinase expressed in Lactococcus lactis is a powerful carrier for
cell wall-associated and secreted bovine beta-lactoglobulin fusion proteins. Appl.
Environ. Microbiol. 68, 2917–2923.
Bjorksten, B., Naaber, P., Sepp, E., and Mikelsaar, M. (1999). The intestinal microflora in
allergic Estonian and Swedish 2-year-old children. Clin. Exp. Allergy 29, 342–346.
Blum, S., and Schiffrin, E. J. (2003). Intestinal microflora and homeostasis of the mucosal
immune response: Implications for probiotic bacteria? Curr. Issues Intest. Microbiol.
4, 53–60.
Bolotin, A., Wincker, P., Mauger, S., Jaillon, O., Malarme, K., Weissenbach, J., Ehrlich,
S. D., and Sorokin, A. (2001). The complete genome sequence of the lactic acid
bacterium Lactococcus lactis ssp. lactis IL1403. Genome Res. 11, 731–753.
Bonetti, A., Morelli, L., Campominosi, E., Ganora, E., and Sforza, F. (2003). Adherence of
Lactobacillus plantarum P 17630 in soft-gel capsule formulation versus Doderlein’s
bacillus in tablet formulation to vaginal epithelial cells. Minerva. Ginecol. 55,279–284.
Boris, S., Suarez, J. E., Vazquez, F., and Barbes, C. (1998). Adherence of human vaginal
lactobacilli to vaginal epithelial cells and interaction with uropathogens. Infect.
Immun. 66, 1985–1989.
Borruel, N., Carol, M., Casellas, F., Antolin, M., de Lara, F., Espin, E., Naval, J., Guarner,
F., and Malagelada, J. R. (2002). Increased mucosal tumour necrosis factor alpha
production in Crohn’s disease can be downregulated ex vivo by probiotic bacteria.
Gut 51, 659–664.
Bosarge, J. R., Watt, J. M., McDaniel, D. O., Swiatlo, E., and McDaniel, L. S. (2001).
Genetic immunization with the region encoding the alpha-helical domain of PspA
elicits protective immunity against Streptococcus pneumoniae. Infect. Immun. 69,5456–5463.
Bottcher, M. F., Nordin, E. K., Sandin, A., Midtvedt, T., and Bjorksten, B. (2000). Micro-
flora-associated characteristics in faeces from allergic and nonallergic infants. Clin.
Exp. Allergy 30, 1590–1596.
Boucher, I., Parrot, M., Gaudreau, H., Champagne, C. P., Vadeboncoeur, C., and Moineau,
S. (2002). Novel food-grade plasmid vector based on melibiose fermentation for the
genetic engineering of Lactococcus lactis. Appl. Environ. Microbiol. 68, 6152–6161.
Bousquet, J., Lockey, R., Malling, H. J., Alvarez-Cuesta, E., Canonica, G. W., Chapman, M. D.,
Creticos, P. J., Dayer, J. M., Durham, S. R., Demoly, P., Goldstein, R. J., Ishikawa, T., Ito,
K., Kraft, D., Lambert, P. H., Lowenstein, H., Muller, U., Norman, P. S., Reisman, R. E.,
Valenta, R., Valovirta, E., and Yssel, H. (1998). Allergen immunotherapy: Therapeutic
vaccines for allergic diseases. World Health Organization. American Academy of Aller-
gy, Asthma and Immunology. Ann. Allergy Asthma Immunol. 81, 401–405.
Braat, H., De Jong, E. C., Van Den Brande, J. M., Kapsenberg, M. L., Peppelenbosch, M. P.,
Van Tol, E. A., and Van Deventer, S. J. (2003). Dichotomy between Lactobacillus
rhamnosus and Klebsiella pneumoniae on dendritic cell phenotype and function.
J. Mol. Med. 82(3), 197–205.
Breiteneder, H., Pettenburger, K., Bito, A., Valenta, R., Kraft, D., Rumpold, H., Scheiner,
O., and Breitenbach, M. (1989). The gene coding for the major birch pollen allergen

USE OF RECOMBINANT LAB IN HUMAN HEALTH 47
Betv1, is highly homologous to a pea disease resistance response gene. EMBO J. 8,1935–1938.
Briles, D. E., Hollingshead, S. K., King, J., Swift, A., Braun, P. A., Park, M. K., Ferguson,
L. M., Nahm, M. H., and Nabors, G. S. (2000). Immunization of humans with recom-
binant pneumococcal surface protein A (rPspA) elicits antibodies that passively
protect mice from fatal infection with Streptococcus pneumoniae bearing heterolo-
gous PspA. J. Infect. Dis. 182, 1694–1701.
Briles, D. E., King, J. D., Gray, M. A., McDaniel, L. S., Swiatlo, E., and Benton, K. A.
(1996). PspA, a protection-eliciting pneumococcal protein: Immunogenicity of
isolated native PspA in mice. Vaccine 14, 858–867.
Briles, D. E., Tart, R. C., Swiatlo, E., Dillard, J. P., Smith, P., Benton, K. A., Ralph, B. A.,
Brooks-Walter, A., Crain, M. J., Hollingshead, S. K., and McDaniel, L. S. (1998).
Pneumococcal diversity: Considerations for new vaccine strategies with emphasis
on pneumococcal surface protein A (PspA). Clin. Microbiol. Rev. 11, 645–657.
Bron, P. A., Benchimol, M. G., Lambert, J., Palumbo, E., Deghorain, M., Delcour, J., De
Vos, W. M., Kleerebezem, M., and Hols, P. (2002). Use of the alr gene as a food-grade
selection marker in lactic acid bacteria. Appl. Environ. Microbiol. 68, 5663–5670.
Bron, P. A., Grangette, C., Mercenier, A., de Vos, W. M., and Kleerebezem, M. (2004a).
Identification of Lactobacillus plantarum genes that are induced in the gastrointesti-
nal tract of mice. J. Bacteriol. 186, 5721–5729.
Bron, P. A., Hoffer, S. M., van Swam, I. I., de Vos, W. M., and Kleerebezem, M. (2004b).
Selection and characterisation of conditionally active promoters in Lactobacillusplantarum, using alanine racemase as a promoter probe. Appl. Env. Microbiol. 70,310–317.
Bron, P. A., Marco, M., Hoffer, S. M., van Mullekom, E., de Vos, W. M., and Kleerebezem,
M. (2004c). In vitro analysis of the bile salt response in Lactobacillus plantarumWCFS1 and its translation to in situ conditions in the gastrointestinal tract.
Submitted for publication.
Bumann, D., Hueck, C., Aebischer, T., and Meyer, T. F. (2000). Recombinant live Salmo-
nella spp. for human vaccination against heterologous pathogens. FEMS Immunol.
Med. Microbiol. 27, 357–364.
Campieri, M., and Gionchetti, P. (1999). Probiotics in inflammatory bowel disease: New
insight to pathogenesis or a possible therapeutic alternative? Gastroenterology 116,1246–1249.
Cangemi de Gutierrez, R., Santos, V., and Nader-Macias, M. E. (2001). Protective effect of
intranasally inoculated Lactobacillus fermentum against Streptococcus pneumoniae
challengeon themouse respiratory tract. FEMSImmunol.Med. Microbiol. 31, 187–195.
Caplan, M. S., Miller-Catchpole, R., Kaup, S., Russell, T., Lickerman, M., Amer, M., Xiao,
Y., and Thomson, R., Jr. (1999). Bifidobacterial supplementation reduces the inci-
dence of necrotizing enterocolitis in a neonatal rat model. Gastroenterology 117,577–583.
Cebra, J., Han-Quiang, J., Sterzl, J., and Tlaskalova, H. (1999). The role of mucosal micro-
biota in the development and maintenance of the mucosal immune system. In ‘‘Hand-
book of Mucosal Immunology’’ (P. L. Ogra, J. Mestecky, M. E. Lamm, W. Strober, J. R.
McGhee, and J. Bienenstock, eds.), pp. 267–280. Academic Press, New York.
Champomier-Verges, M. C., Maguin, E., Mistou, M. Y., Anglade, P., and Chich, J. F.
(2002). Lactic acid bacteria and proteomics: Current knowledge and perspectives.
J. Chromatogr. B. Analyt. Technol. Biomed. Life Sci. 771, 329–342.
Chang, T. L., Chang, C. H., Simpson, D. A., Xu, Q., Martin, P. K., Lagenaur, L. A.,
Schoolnik, G. K., Ho, D. D., Hillier, S. L., Holodniy, M., Lewicki, J. A., and Lee, P. P.

48 HANNIFFY et al.
(2003). Inhibition of HIV infectivity by a natural human isolate of Lactobacillus
jensenii engineered to express functional two-domain CD4. Proc. Natl. Acad. Sci.
USA 100, 11672–11677.
Chen, T., Toivanen, P., and Vainio, O. (2002). Suppression of antigen-specific lympho-
cyte proliferation by Gram-positive bacterial cell walls. Apmis 110, 490–498.
Chou, L. S., and Weimer, B. (1999). Isolation and characterization of acid- and bile-
tolerant isolates from strains of Lactobacillus acidophilus. J. Dairy Sci. 82, 23–31.
Christensen, H. R., Frokiaer, H., and Pestka, J. J. (2002). Lactobacilli differentially modu-
late expression of cytokines and maturation surface markers in murine dendritic
cells. J. Immunol. 168, 171–178.
Cleveland, M. G., Gorham, J. D., Murphy, T. L., Tuomanen, E., and Murphy, K. M. (1996).
Lipoteichoic acid preparations of Gram-positive bacteria induce interleukin-12
through a CD14-dependent pathway. Infect. Immun. 64, 1906–1912.
Cortes-Perez, N. G., Bermudez-Humaran, L. G., Le Loir, Y., Rodriguez-Padilla, C., Gruss,
A., Saucedo-Cardenas, O., Langella, P., and Montes-de-Oca-Luna, R. (2003). Mice
immunization with live lactococci displaying a surface anchored HPV-16 E7
oncoprotein. FEMS Microbiol. Lett. 229, 37–42.
Crain, M. J., Waltman, W. D., 2nd, Turner, J. S., Yother, J., Talkington, D. F., McDaniel,
L. S., Gray, B. M., and Briles, D. E. (1990). Pneumococcal surface protein A (PspA) is
serologically highly variable and is expressed by all clinically important capsular
serotypes of Streptococcus pneumoniae. Infect. Immun. 58, 3293–3299.
Cross, M. L., Stevenson, L. M., and Gill, H. S. (2001). Anti-allergy properties of fermen-
ted foods: An important immunoregulatory mechanism of lactic acid bacteria?
Int. Immunopharmacol. 1, 891–901.
Cukrowska, B., LodInova-ZadnIkova, R., Enders, C., Sonnenborn, U., Schulze, J., and
Tlaskalova-Hogenova, H. (2002). Specific proliferative and antibody responses of
premature infants to intestinal colonization with nonpathogenic probiotic E. coli
strain Nissle 1917. Scand J. Immunol. 55, 204–209.
Curtiss III, R., Pereira, D. A., Hsu, J. C., Hull, S. C., Clark, J. E., Maturin, Sr., L. J.,
Goldschmidt, R., Moody, R., Inoue, M., and Alexander, L. (1977). Biological contain-
ment: The subordination of Escherichia coli K12. In ‘‘Recombinant Molecules: Impact
on Science and Society’’ (R. F. Bers, Jr. and E. G. Bassett, eds.), pp. 45–56. Raven
Press, New York.
de Ruyter, P. G., Kuipers, O. P., Meijer, W. C., and de Vos, W. M. (1997). Food-grade
controlled lysis of Lactococcus lactis for accelerated cheese ripening. Nat. Biotechnol.
15, 976–979.
de Vos, W. M. (1999a). Gene expression systems for lactic acid bacteria. Curr. Opin.Microbiol. 2, 289–295.
de Vos, W. M. (1999b). Safe and sustainable systems for food-grade fermentations by
genetically modified lactic acid bacteria. International Dairy Journal 9, 3–10.
Di Fabio, S., Medaglini, D., Rush, C. M., Corrias, F., Panzini, G. L., Pace, M., Verani, P.,
Pozzi, G., and Titti, F. (1998). Vaginal immunization of Cynomolgus monkeys with
Streptococcus gordonii expressing HIV-1 and HPV 16 antigens. Vaccine 16, 485–492.
Dieye, Y., Usai, S., Clier, F., Gruss, A., and Piard, J. C. (2001). Design of a protein-targeting
system for lactic acid bacteria. J. Bacteriol. 183, 4157–4166.
Djordjevic, G. M., and Klaenhammer, T. R. (1996). Positive selection, cloning vectors for
Gram-positive bacteria based on a restriction endonuclease cassette. Plasmid 35, 37–45.
Djordjevic, G. M., O’Sullivan, D. J., Walker, S. A., Conkling, M. A., and Klaenhammer,
T. R. (1997). A triggered-suicide system designed as a defense against bacteriophages.
J. Bacteriol. 179, 6741–6748.

USE OF RECOMBINANT LAB IN HUMAN HEALTH 49
Duchmann, R., Kaiser, I., Hermann, E., Mayet, W., Ewe, K., and Meyer zum Buschenfelde,
K. H. (1995). Tolerance exists towards resident intestinal flora but is broken in active
inflammatory bowel disease (IBD). Clin. Exp. Immunol. 102, 448–455.
Ebner, C., Schenk, S., Najafian, N., Siemann, U., Steiner, R., Fischer, G. W., Hoffmann, K.,
Szepfalusi, Z., Scheiner, O., and Kraft, D. (1995). Nonallergic individuals recognize
the same T cell epitopes of Bet v 1, the major birch pollen allergen, as atopic patients.
J. Immunol. 154, 1932–1940.
Enouf, V., Langella, P., Commissaire, J., Cohen, J., and Corthier, G. (2001). Bovine rotavi-
rus nonstructural protein 4 produced by Lactococcus lactis is antigenic and immu-
nogenic. Appl. Environ. Microbiol. 67, 1423–1428.
Fabia, R., Ar’Rajab, A., Johansson, M. L., Willen, R., Andersson, R., Molin, G., and
Bengmark, S. (1993). The effect of exogenous administration of Lactobacillus reuteri
R2LC and oat fiber on acetic acid-induced colitis in the rat. Scand. J. Gastroenterol.
28, 155–162.
Fang, H., Elina, T., Heikki, A., and Seppo, S. (2000). Modulation of humoral immune
response through probiotic intake. FEMS Immunol. Med. Microbiol. 29, 47–52.
Farrell, R. J., and Peppercorn, M. A. (2002). Ulcerative colitis. Lancet 359, 331–340.
Ferreira, F. D., Hoffmann-Sommergruber, K., Breiteneder, H., Pettenburger, K., Ebner, C.,
Sommergruber, W., Steiner, R., Bohle, B., Sperr, W. R., Valent, P., et al. (1993).
Purification and characterization of recombinant Bet v I, the major birch pollen
allergen. Immunological equivalence to natural Bet v I. J. Biol. Chem. 268,19574–19580.
Fioramonti, J., Theodorou, V., and Bueno, L. (2003). Probiotics: What are they?
What are their effects on gut physiology? Best Pract. Res. Clin. Gastroenterol. 17,711–724.
Fitzsimons, A., Hols, P., Jore, J., Leer, R. J., O’Connell, M., and Delcour, J. (1994).
Development of an amylolytic Lactobacillus plantarum silage strain expressing the
Lactobacillus amylovorus alpha-amylase gene. Appl. Environ. Microbiol. 60,3529–3535.
Frazier, C. L., San Filippo, J., Lambowitz, A. M., and Mills, D. A. (2003). Genetic
manipulation of Lactococcus lactis by using targeted group II introns: Generation of
stable insertions without selection. Appl. Environ. Microbiol. 69, 1121–1128.
Froseth, B. R., and McKay, L. L. (1991). Molecular characterization of the nisin resistance
region of Lactococcus lactis subsp. lactis biovar diacetylactis DRC3. Appl. Environ.
Microbiol. 57, 804–811.
Fu, X., and Xu, J. G. (2000). Development of a chromosome-plasmid balanced lethal
system for Lactobacillus acidophilus with thyA gene as selective marker. Microbiol.Immunol. 44, 551–556.
Fujiwara, M., Kaneko, T., Iwana, H., Taketomo, N., Tsunoo, H., Kanno, J., Ohkusa, T., and
Okayasu, I. (2003). Inhibitory effects of Bifidobacterium longum on experimental
ulcerative colitis induced in mice by synthetic dextran sulfate sodium. Digestion
67, 90–95.
Fukushima, Y., Kawata, Y., Hara, H., Terada, A., and Mitsuoka, T. (1998). Effect of a
probiotic formula on intestinal immunoglobulin A production in healthy children.
Int. J. Food Microbiol. 42, 39–44.
Fuller, R. (1989). Probiotics in man and animals. J. Appl. Bacteriol. 66, 365–378.
Funda, D. P., Tuckova, L., Farre, M. A., Iwase, T., Moro, I., and Tlaskalova-Hogenova, H.
(2001). CD14 is expressed and released as soluble CD14 by human intestinal epithe-
lial cells in vitro: Lipopolysaccharide activation of epithelial cells revisited. Infect.
Immun. 69, 3772–3781.

50 HANNIFFY et al.
Gajhede, M., Osmark, P., Poulsen, F. M., Ipsen, H., Larsen, J. N., Joost van Neerven, R. J.,
Schou, C., Lowenstein, H., and Spangfort, M. D. (1996). X-ray and NMR structure of
Bet v 1, the origin of birch pollen allergy. Nat. Struct. Biol. 3, 1040–1045.
Gallichan, W. S., and Rosenthal, K. L. (1996). Effects of the estrous cycle on local humoral
immune responses and protection of intranasally immunized female mice against
herpes simplex virus type 2 infection in the genital tract. Virology 224, 487–497.
Gicquel, B. (1995). BCG as a vector for the construction of multivalent recombinant
vaccines. Biologicals 23, 113–118.
Gil, M. T., Perez-Arellano, I., Buesa, J., and Perez-Martinez, G. (2001). Secretion of the
rotavirus VP8* protein in Lactococcus lactis. FEMS Microbiol. Lett. 203, 269–274.
Gilbert, C., Robinson, K., Le Page, R. W., and Wells, J. M. (2000). Heterologous expression
of an immunogenic pneumococcal type 3 capsular polysaccharide in Lactococcus
lactis. Infect. Immun. 68, 3251–3260.
Gill, H. S., Rutherfurd, K. J., Prasad, J., and Gopal, P. K. (2000). Enhancement of natural
and acquired immunity by Lactobacillus rhamnosus (HN001), Lactobacillus aci-
dophilus (HN017) and Bifidobacterium lactis (HN019). Br. J. Nutr. 83, 167–176.
Gillgrass, A. E., Ashkar, A. A., Rosenthal, K. L., and Kaushic, C. (2003). Prolonged
exposure to progesterone prevents induction of protective mucosal responses follow-
ing intravaginal immunization with attenuated herpes simplex virus type 2. J. Virol.
77, 9845–9851.
Giomarelli, B., Provvedi, R., Meacci, F., Maggi, T., Medaglini, D., Pozzi, G., Mori, T.,
McMahon, J. B., Gardella, R., and Boyd, M. R. (2002). The microbicide cyanovirin-N
expressed on the surface of commensal bacterium Streptococcus gordonii captures
HIV-1. AIDS 16, 1351–1356.
Gionchetti, P., Rizzello, F., Venturi, A., Brigidi, P., Matteuzzi, D., Bazzocchi, G., Poggioli,
G., Miglioli, M., and Campieri, M. (2000). Oral bacteriotherapy as maintenance
treatment in patients with chronic pouchitis: A double-blind, placebo-controlled
trial. Gastroenterology 119, 305–309.
Gionchetti, P., Rizzello, F., Helwig, U., Venturi, A., Lammers, K. M., Brigidi, P., Vitali, P.,
Poggioli, G., and Campieri, M. (2003). Prophylaxis of pouchitis onset with
probiotic therapy: A double-blind, placebo-controlled trial. Gastroenterology 124,1202–1209.
Glenting, J., Madsen, S. M., Vrang, A., Fomsgaard, A., and Israelsen, H. (2002). A plasmid
selection system in Lactococcus lactis and its use for gene expression in L. lactis and
human kidney fibroblasts. Appl. Environ. Microbiol. 68, 5051–5056.
Gluck, U., and Gebbers, J. O. (2003). Ingested probiotics reduce nasal colonization with
pathogenic bacteria (Staphylococcus aureus, Streptococcus pneumoniae, and beta-
hemolytic streptococci). Am. J. Clin. Nutr. 77, 517–520.
Goeddel, D. V., Heyneker, H. L., Hozumi, T., Arentzen, R., Itakura, K., Yansura, D. G., Ross,
M. J., Miozzari, G., Crea, R., and Seeburg, P. H. (1979a). Direct expression in Escher-ichia coli of a DNA sequence coding for human growth hormone. Nature 281, 544–548.
Goeddel, D. V., Kleid, D. G., Bolivar, F., Heyneker, H. L., Yansura, D. G., Crea, R., Hirose,
T., Kraszewski, A., Itakura, K., and Riggs, A. D. (1979b). Expression in Escherichia
coli of chemically synthesized genes for human insulin. Proc. Natl. Acad. Sci. USA76, 106–110.
Grangette, C., Muller-Alouf, H., Geoffroy, M., Goudercourt, D., Turneer, M., and Mercenier,
A. (2002). Protection against tetanus toxin after intragastric administration of two
recombinant lactic acid bacteria: Impact of strain viability and in vivo persistence.
Vaccine 20, 3304–3309.

USE OF RECOMBINANT LAB IN HUMAN HEALTH 51
Grangette, C., Muller-Alouf, H., Goudercourt, D., Geoffroy, M. C., Turneer, M., and
Mercenier, A. (2001). Mucosal immune responses and protection against tetanus
toxin after intranasal immunization with recombinant Lactobacillus plantarum.
Infect. Immun. 69, 1547–1553.
Gruzza, M., Langella, P., Duval-Iflah, Y., and Ducluzeau, R. (1993). Gene transfer from
engineered Lactococcus lactis strains to Enterococcus faecalis in the digestive tract of
gnotobiotic mice. Microb. Releases 2, 121–125.
Guillot, A., Gitton, C., Anglade, P., and Mistou, M. Y. (2003). Proteomic analysis of
Lactococcus lactis, a lactic acid bacterium. Proteomics 3, 337–354.
Hakenbeck, R., Grebe, T., Zahner, D., and Stock, J. B. (1999). Beta-lactam resistance in
Streptococcus pneumoniae: Penicillin-binding proteins and non-penicillin-binding
proteins. Mol. Microbiol. 33, 673–678.
Haller, D., Bode, C., Hammes, W. P., Pfeifer, A. M., Schiffrin, E. J., and Blum, S. (2000).
Non-pathogenic bacteria elicit a differential cytokine response by intestinal epithelial
cell/leucocyte co-cultures. Gut 47, 79–87.
Hanniffy, S., Carter, A. T., and Wells, J. M. (2004). Protection from respiratory infection
with Streptococcus pneumoniae by intranasal vaccination with Lc. lactis expressing
PspA. Submitted for publication.
Hashiba, H., Takiguchi, R., Jyoho, K., and Aoyama, K. (1992). Establishment of a host-
vector system in Lactobacillus helveticus with beta-galactosidase activity as a
selection marker. Biosci. Biotechnol. Biochem. 56, 190–194.
Hatakka, K., Savilahti, E., Ponka, A., Meurman, J. H., Poussa, T., Nase, L., Saxelin, M.,
and Korpela, R. (2001). Effect of long term consumption of probiotic milk on infec-
tions in children attending day care centres: Double blind, randomised trial. BMJ
322, 1327.
Heilig, H. G., Zoetendal, E. G., Vaughan, E. E., Marteau, P., Akkermans, A. D., and de Vos,
W. M. (2002). Molecular diversity of Lactobacillus spp. and other lactic acid bacteria
in the human intestine as determined by specific amplification of 16S ribosomal
DNA. Appl. Environ. Microbiol. 68, 114–123.
Herias, M. V., Hessle, C., Telemo, E., Midtvedt, T., Hanson, L. A., and Wold, A. E. (1999).
Immunomodulatory effects of Lactobacillus plantarum colonizing the intestine of
gnotobiotic rats. Clin. Exp. Immunol. 116, 283–290.
Heyman, M. (2000). Effect of lactic acid bacteria on diarrheal diseases. J. Am. Coll. Nutr.19, 137S–146S.
Hillier, S. L., Krohn, M. A., Rabe, L. K., Klebanoff, S. J., and Eschenbach, D. A. (1993).
The normal vaginal flora, H2O2-producing lactobacilli, and bacterial vaginosis in
pregnant women. Clin. Infect. Dis. 16, S273–S281.
Hillman, J. D. (2002). Genetically modified Streptococcus mutans for the prevention of
dental caries. Antonie Van Leeuwenhoek 82, 361–366.
Holma, R., Salmenpera, P., Lohi, J., Vapaatalo, H., and Korpela, R. (2001). Effects of
Lactobacillus rhamnosus GG and Lactobacillus reuteri R2LC on acetic acid-induced
colitis in rats. Scand. J. Gastroenterol. 36, 630–635.
Hols, P., Defrenne, C., Ferain, T., Derzelle, S., Delplace, B., and Delcour, J. (1997a). The
alanine racemase gene is essential for growth of Lactobacillus plantarum. J. Bacteriol.179, 3804–3807.
Hols, P., Ferain, T., Garmyn, D., Bernard, N., and Delcour, J. (1994). Use of homologous
expression-secretion signals and vector-free stable chromosomal integration in engi-
neering of Lactobacillus plantarum for alpha-amylase and levanase expression.
Appl. Environ. Microbiol. 60, 1401–1413.

52 HANNIFFY et al.
Hols, P., Kleerebezem, M., Schanck, A. N., Ferain, T., Hugenholtz, J., Delcour, J., and de
Vos, W. M. (1999). Conversion of Lactococcus lactis from homolactic to homoalanine
fermentation through metabolic engineering. Nat. Biotechnol. 17, 588–592.
Hols, P., Slos, P., Dutot, P., Reymund, J., Chabot, P., Delplace, B., Delcour, J., and
Mercenier, A. (1997b). Efficient secretion of the model antigen M6-gp41E in Lactoba-
cillus plantarum NCIMB 8826. Microbiology 143, 2733–2741.
Holzapfel, W. H., Haberer, P., Snel, J., Schillinger, U., and Huis in’t Veld, J. H. (1998).
Overview of gut flora and probiotics. Int. J. Food Microbiol. 41, 85–101.
Hori, T., Kiyoshima, J., Shida, K., and Yasui, H. (2001). Effect of intranasal administration
of Lactobacillus casei Shirota on influenza virus infection of upper respiratory tract
in mice. Clin. Diagn. Lab. Immunol. 8, 593–597.
Hori, T., Kiyoshima, J., Shida, K., and Yasui, H. (2002). Augmentation of cellular immu-
nity and reduction of influenza virus titer in aged mice fed Lactobacillus casei strain
Shirota. Clin. Diagn. Lab. Immunol. 9, 105–108.
Hudcovic, T., Stepankova, R., Cebra, J., and Tlaskalova-Hogenova, H. (2001). The role of
microflora in the development of intestinal inflammation: Acute and chronic colitis
induced by dextran sulfate in germ-free and conventionally reared immunocompe-
tent and immunodeficient mice. Folia Microbiol (Praha) 46, 565–572.
Hyronimus, B., Le Marrec, C., Sassi, A. H., and Deschamps, A. (2000). Acid and bile
tolerance of spore-forming lactic acid bacteria. Int. J. Food Microbiol. 61, 193–197.
Ichikawa, H., Kuroiwa, T., Inagaki, A., Shineha, R., Nishihira, T., Satomi, S., and Sakata,
T. (1999). Probiotic bacteria stimulate gut epithelial cell proliferation in rat. Dig. Dis.Sci. 44, 2119–2123.
Isolauri, E., Sutas, Y., Kankaanpaa, P., Arvilommi, H., and Salminen, S. (2001). Probio-
tics: Effects on immunity. Am. J. Clin. Nutr. 73, 444S–450S.
Israel, D. A., Salama, N., Krishna, U., Rieger, U. M., Atherton, J. C., Falkow, S., and Peek,
R. M., Jr. (2001). Helicobacter pylori genetic diversity within the gastric niche of a
single human host. Proc. Natl. Acad. Sci. USA 98, 14625–14630.
Israelsen, H., Madsen, S. M., Johansen, E., Vrang, A., and Hansen, E. B. (1995a). Environ-
mentally regulated promoters in lactococci. Dev. Biol. Stand. 85, 443–448.
Israelsen, H., Madsen, S. M., Vrang, A., Hansen, E. B., and Johansen, E. (1995b). Cloning
and partial characterization of regulated promoters from Lactococcus lactis Tn917-
lacZ integrants with the new promoter probe vector, pAK80. Appl. Environ. Micro-biol. 61, 2540–2547.
Jahn-Schmid, B., Wiedermann, U., Bohle, B., Repa, A., Kraft, D., and Ebner, C. (1999).
Oligodeoxynucleotides containing CpG motifs modulate the allergic TH2 response of
BALB/c mice to Bet v 1, the major birch pollen allergen. J. Allergy Clin. Immunol.104, 1015–1023.
Janeway, C. A., Jr., and Medzhitov, R. (2002). Innate immune recognition. Annu. Rev.
Immunol. 20, 197–216. Epub 2001 Oct 2004.
Kalliomaki, M., and Isolauri, E. (2003). Role of intestinal flora in the development of
allergy. Curr. Opin. Allergy Clin. Immunol. 3, 15–20.
Kalliomaki, M., Salminen, S., Arvilommi, H., Kero, P., Koskinen, P., and Isolauri, E.
(2001). Probiotics in primary prevention of atopic disease: A randomised placebo-
controlled trial. Lancet 357, 1076–1079.
Kalliomaki, M., Salminen, S., Poussa, T., Arvilommi, H., and Isolauri, E. (2003). Probiotics
and prevention of atopic disease: 4-year follow-up of a randomised placebo-controlled
trial. Lancet 361, 1869–1871.
Kantele, A., Hakkinen, M., Moldoveanu, Z., Lu, A., Savilahti, E., Alvarez, R. D., Michalek,
S., and Mestecky, J. (1998). Differences in immune responses induced by oral and

USE OF RECOMBINANT LAB IN HUMAN HEALTH 53
rectal immunizations with Salmonella typhi Ty21a: Evidence for compartmentaliza-
tion within the common mucosal immune system in humans. Infect. Immun. 66,5630–5635.
Kariuki, S., and Hart, C. A. (2001). Global aspects of antimicrobial-resistant enteric
bacteria. Curr. Opin. Infect. Dis. 14, 579–586.
Kato, I., Tanaka, K., and Yokokura, T. (1999). Lactic acid bacterium potently induces the
production of interleukin-12 and interferon-gamma by mouse splenocytes. Int. J.Immunopharmacol. 21, 121–131.
Kaushic, C., Zhou, F., Murdin, A. D., and Wira, C. R. (2000). Effects of estradiol and
progesterone on susceptibility and early immune responses to Chlamydia trachoma-
tis infection in the female reproductive tract. Infect. Immun. 68, 4207–4216.
Kili, A. O., Herzberg, M. C., Meyer, M. W., Zhao, X., and Tao, L. (1999). Streptococcal
reporter gene-fusion vector for identification of in vivo expressed genes. Plasmid 42,67–72.
Kim, T. S., Hur, J. W., Yu, M. A., Cheigh, C. I., Kim, K. N., Hwang, J. K., and Pyun, Y. R.
(2003). Antagonism of Helicobacter pylori by bacteriocins of lactic acid bacteria.
J. Food Prot. 66, 3–12.
Kirjavainen, P. V., El-Nezami, H. S., Salminen, S. J., Ahokas, J. T., and Wright, P. F.
(1999a). Effects of orally administered viable Lactobacillus rhamnosus GG
and Propionibacterium freudenreichii subsp. shermanii JS on mouse lymphocyte
proliferation. Clin. Diagn. Lab. Immunol. 6, 799–802.
Kirjavainen, P. V., El-Nezami, H. S., Salminen, S. J., Ahokas, J. T., and Wright, P. F.
(1999b). The effect of orally administered viable probiotic and dairy lacto-
bacilli on mouse lymphocyte proliferation. FEMS Immunol. Med. Microbiol. 26,131–135.
Klaenhammer, T., Altermann, E., Arigoni, F., Bolotin, A., Breidt, F., Broadbent, J., Cano,
R., Chaillou, S., Deutscher, J., Gasson, M., van de Guchte, M., Guzzo, J., Hartke, A.,
Hawkins, T., Hols, P., Hutkins, R., Kleerebezem, M., Kok, J., Kuipers, O., Lubbers, M.,
Maguin, E., McKay, L., Mills, D., Nauta, A., Overbeek, R., Pel, H., Pridmore, D., Saier,
M., van Sinderen, D., Sorokin, A., Steele, J., O’Sullivan, D., de Vos, W., Weimer, B.,
Zagorec, M., and Siezen, R. (2002). Discovering lactic acid bacteria by genomics.
Antonie Van Leeuwenhoek 82, 29–58.
Kleerebezem, M., Beerthuyzen, M. M., Vaughan, E. E., de Vos, W. M., and Kuipers, O. P.
(1997). Controlled gene expression systems for lactic acid bacteria: Transferable
nisin-inducible expression cassettes for Lactococcus, Leuconostoc, and Lactobacillus
spp. Appl. Environ. Microbiol. 63, 4581–4584.
Kleerebezem, M., Boekhorst, J., van Kranenburg, R., Molenaar, D., Kuipers, O. P., Leer, R.,
Tarchini, R., Peters, S. A., Sandbrink, H. M., Fiers, M. W., Stiekema, W., Lankhorst,
R. M., Bron, P. A., Hoffer, S. M., Groot, M. N., Kerkhoven, R., de Vries, M., Ursing, B.,
de Vos, W. M., and Siezen, R. J. (2003). Complete genome sequence of Lactobacillusplantarum WCFS1. Proc. Natl. Acad. Sci. USA 100, 1990–1995.
Kociubinski, G., Perez, P., and De Antoni, G. (1999). Screening of bile resistance and bile
precipitation in lactic acid bacteria and bifidobacteria. J. Food Prot. 62, 905–912.
Koga, T., Mizobe, T., and Takumi, K. (1998). Antibacterial activity of Lactobacillusspecies against Vibrio species. Microbiol. Res. 153, 271–275.
Konrad, A., Mahler, M., Flogerzi, B., Kalousek, M. B., Lange, J., Varga, L., and Seibold, F.
(2003). Amelioration of murine colitis by feeding a solution of lysed Escherichia coli.
Scand. J. Gastroenterol. 38, 172–179.
Kruger, C., Hu, Y., Pan, Q., Marcotte, H., Hultberg, A., Delwar, D., van Dalen, P. J., Pouwels,
P. H., Leer, R. J., Kelly, C. G., van Dollenweerd, C., Ma, J. K., and Hammarstrom, L.

54 HANNIFFY et al.
(2002). In situ delivery of passive immunity by lactobacilli producing single-chain
antibodies. Nat. Biotechnol. 20, 702–706.
Kruis, W., Schutz, E., Fric, P., Fixa, B., Judmaier, G., and Stolte, M. (1997). Double-
blind comparison of an oral Escherichia coli preparation and mesalazine in
maintaining remission of ulcerative colitis. Aliment. Pharmacol Ther. 11,853–858.
Kruisselbrink, A., Heijne Den Bak-Glashouwer, M. J., Havenith, C. E., Thole, J. E., and
Janssen, R. (2001). Recombinant Lactobacillus plantarum inhibits house dust mite-
specific T-cell responses. Clin. Exp. Immunol. 126, 2–8.
Kuipers, O. P., de Ruyter, P. G., Kleerebezem, M., and de Vos, W. M. (1997). Controlled
over-production of proteins by lactic acid bacteria. Trends Biotechnol. 15, 135–140.
Kuisma, J., Mentula, S., Jarvinen, H., Kahri, A., Saxelin, M., and Farkkila, M. (2003).
Effect of Lactobacillus rhamnosus GG on ileal pouch inflammation and microbial
flora. Aliment. Pharmacol. Ther. 17, 509–515.
Kullisaar, T., Zilmer, M., Mikelsaar, M., Vihalemm, T., Annuk, H., Kairane, C., and Kilk,
A. (2002). Two antioxidative lactobacilli strains as promising probiotics. Int. J. Food
Microbiol. 72, 215–224.
Kunst, F., Ogasawara, N., Moszer, I., Albertini, A. M., Alloni, G., Azevedo, V., Bertero,
M. G., Bessieres, P., Bolotin, A., Borchert, S., Borriss, R., Boursier, L., Brans, A.,
Braun, M., Brignell, S. C., Bron, S., Brouillet, S., Bruschi, C. V., Caldwell, B.,
Capuano, V., Carter, N. M., Choi, S. K., Codani, J. J., Connerton, I. F., Danchin, A.,
et al. (1997). The complete genome sequence of the Gram-positive bacterium Bacillussubtilis. Nature 390, 249–256.
Kutteh, W. H., Kantele, A., Moldoveanu, Z., Crowley-Nowick, P. A., and Mestecky, J.
(2001). Induction of specific immune responses in the genital tract of women
after oral or rectal immunization and rectal boosting with Salmonella typhi Ty
21a vaccine. J. Reprod. Immunol. 52, 61–75.
Lamine, F., Fioramonti, J., Bueno, L., Nepveu, F., Cauquil, E., Lobysheva, I., Eutamene,
H., and Theodorou, V. (2004). Nitric oxide released by Lactobacillus farciminisimproves TNBS-induced colitis in rats. Scand. J. Gastroenterol. 39, 37–45.
Langella, P., and Le Loir, Y. (1999). Heterologous protein secretion in Lactococcus lactis:
A novel antigen delivery system. Braz. J. Med. Biol. Res. 32, 191–198.
Lee, M. H., Roussel, Y., Wilks, M., and Tabaqchali, S. (2001). Expression of Helicobacterpylori urease subunit B gene in Lactococcus lactis MG1363 and its use as a vaccine
delivery system against H. pylori infection in mice. Vaccine 19, 3927–3935.
Lee, S. F., Halperin, S. A., Knight, J. B., and Tait, A. (2002). Purification and immunoge-
nicity of a recombinant Bordetella pertussis S1S3FHA fusion protein expressed by
Streptococcus gordonii. Appl. Environ. Microbiol. 68, 4253–4258.
Lee, S. F., March, R. J., Halperin, S. A., Faulkner, G., and Gao, L. (1999). Surface
expression of a protective recombinant pertussis toxin S1 subunit fragment in
Streptococcus gordonii. Infect. Immun. 67, 1511–1516.
Lee, Y. K., Puong, K. Y., Ouwehand, A. C., and Salminen, S. (2003). Displacement of
bacterial pathogens from mucus and Caco-2 cell surface by lactobacilli. J. Med.
Microbiol. 52, 925–930.
Leenhouts, K., Bolhuis, A., Venema, G., and Kok, J. (1998). Construction of a food-grade
multiple-copy integration system for Lactococcus lactis. Appl. Microbiol. Biotechnol.
49, 417–423.
Leenhouts, K., Buist, G., Bolhuis, A., ten Berge, A., Kiel, J., Mierau, I., Dabrowska, M.,
Venema, G., and Kok, J. (1996). A general system for generating unlabelled gene
replacements in bacterial chromosomes. Mol. Gen. Genet. 253, 217–224.

USE OF RECOMBINANT LAB IN HUMAN HEALTH 55
Levine, M. M., Galen, J., Barry, E., Noriega, F., Chatfield, S., Sztein, M., Dougan, G., and
Tacket, C. (1996). Attenuated Salmonella as live oral vaccines against typhoid fever
and as live vectors. J. Biotechnol. 44, 193–196.
Link-Amster, H., Rochat, F., Saudan, K. Y., Mignot, O., and Aeschlimann, J. M. (1994).
Modulation of a specific humoral immune response and changes in intestinal
flora mediated through fermented milk intake. FEMS Immunol. Med. Microbiol. 10,55–63.
Liu, C. Q., Leelawatcharamas, V., Harvey, M. L., and Dunn, N. W. (1996). Cloning vectors
for lactococci based on a plasmid encoding resistance to cadmium. Curr. Microbiol.
33, 35–39.
Lodinova-Zadnikova, R., Cukrowska, B., and Tlaskalova-Hogenova, H. (2003). Oral ad-
ministration of probiotic Escherichia coli after birth reduces frequency of allergies
and repeated infections later in life (after 10 and 20 years). Int. Arch. Allergy
Immunol. 131, 209–211.
Maassen, C. B., Laman, J. D., den Bak-Glashouwer, M. J., Tielen, F. J., van Holten-Neelen,
J. C., Hoogteijling, L., Antonissen, C., Leer, R. J., Pouwels, P. H., Boersma, W. J., and
Shaw, D. M. (1999). Instruments for oral disease-intervention strategies: Recombi-
nant Lactobacillus casei expressing tetanus toxin fragment C for vaccination or
myelin proteins for oral tolerance induction in multiple sclerosis. Vaccine 17,2117–2128.
Maassen, C. B., van Holten-Neelen, C., Balk, F., den Bak-Glashouwer, M. J., Leer, R. J.,
Laman, J. D., Boersma, W. J., and Claassen, E. (2000). Strain-dependent induction of
cytokine profiles in the gut by orally administered Lactobacillus strains. Vaccine 18,2613–2623.
MacCormick, C. A., Griffin, H. G., and Gasson, M. J. (1995). Construction of a food-grade
host/vector system for Lactococcus lactis based on the lactose operon. FEMSMicrobiol. Lett. 127, 105–109.
Mack, D. R., Ahrne, S., Hyde, L., Wei, S., and Hollingsworth, M. A. (2003). Extracellular
MUC3 mucin secretion follows adherence of Lactobacillus strains to intestinal
epithelial cells in vitro. Gut 52, 827–833.
Madsen, K. L., Doyle, J. S., Jewell, L. D., Tavernini, M. M., and Fedorak, R. N. (1999a).
Lactobacillus species prevents colitis in interleukin 10 gene-deficient mice.
Gastroenterology 116, 1107–1114.
Madsen, S. M., Albrechtsen, B., Hansen, E. B., and Israelsen, H. (1996). Cloning and
transcriptional analysis of two threonine biosynthetic genes from Lactococcus lactis
MG1614. J. Bacteriol. 178, 3689–3694.
Madsen, S. M., Arnau, J., Vrang, A., Givskov, M., and Israelsen, H. (1999b). Molecular
characterization of the pH-inducible and growth phase-dependent promoter P170 of
Lactococcus lactis. Mol. Microbiol. 32, 75–87.
Maggi, L., Mastromarino, P., Macchia, S., Brigidi, P., Pirovano, F., Matteuzzi, D., and
Conte, U. (2000). Technological and biological evaluation of tablets containing
different strains of lactobacilli for vaginal administration. Eur. J. Pharm. Biopharm.
50, 389–395.
Maguin, E., Prevost, H., Ehrlich, S. D., and Gruss, A. (1996). Efficient insertional muta-
genesis in lactococci and other Gram-positive bacteria. J. Bacteriol. 178, 931–935.
Mahan, M. J., Heithoff, D. M., Sinsheimer, R. L., and Low, D. A. (2000). Assessment of
bacterial pathogenesis by analysis of gene expression in the host. Annu. Rev. Genet.
34, 139–164.
Makrides, S. C. (1996). Strategies for achieving high-level expression of genes in Escher-
ichia coli. Microbiol. Rev. 60, 512–538.

56 HANNIFFY et al.
Mao, Y., Nobaek, S., Kasravi, B., Adawi, D., Stenram, U., Molin, G., and Jeppsson, B.
(1996). The effects of Lactobacillus strains and oat fiber on methotrexate-induced
enterocolitis in rats. Gastroenterology 111, 334–344.
Marsh, D. G., Neely, J. D., Breazeale, D. R., Ghosh, B., Freidhoff, L. R., Ehrlich-Kautzky, E.,
Schou, C., Krishnaswamy, G., and Beaty, T. H. (1994). Linkage analysis of IL-4 and
other chromosome 5q31.1 markers and total serum immunoglobulin E concentra-
tions. Science 264, 1152–1156.
Marshall, J. D., Secrist, H., DeKruyff, R. H., Wolf, S. F., and Umetsu, D. T. (1995). IL-12
inhibits the production of IL-4 and IL-10 in allergen-specific human CD4þ T lym-
phocytes. J. Immunol. 155, 111–117.
Marteau, P., Seksik, P., and Shanahan, F. (2003). Manipulation of the bacterial flora in
inflammatory bowel disease. Best. Pract. Res. Clin. Gastroenterol. 17, 47–61.
Martin, M. C., Alonso, J. C., Suarez, J. E., and Alvarez, M. A. (2000). Generation of food-
grade recombinant lactic acid bacterium strains by site-specific recombination.
Appl. Environ. Microbiol. 66, 2599–2604.
Mastromarino, P., Brigidi, P., Macchia, S., Maggi, L., Pirovano, F., Trinchieri, V., Conte,
U., and Matteuzzi, D. (2002). Characterization and selection of vaginal Lactobacillus
strains for the preparation of vaginal tablets. J. Appl. Microbiol. 93, 884–893.
Matar, C., Valdez, J. C., Medina, M., Rachid, M., and Perdigon, G. (2001). Immunomodu-
lating effects of milks fermented by Lactobacillus helveticus and its non-proteolytic
variant. J. Dairy Res. 68, 601–609.
Matsuzaki, T., Yamazaki, R., Hashimoto, S., and Yokokura, T. (1998). The effect of oral
feeding of Lactobacillus casei strain Shirota on immunoglobulin E production in
mice. J. Dairy Sci. 81, 48–53.
McGroarty, J. A., and Reid, G. (1988). Detection of a Lactobacillus substance that inhibits
Escherichia coli. Can. J. Microbiol. 34, 974–978.
Medaglini, D., Oggioni, M. R., and Pozzi, G. (1998). Vaginal immunization with recombi-
nant gram-positive bacteria. Am. J. Reprod. Immunol. 39, 199–208.
Medina, E., and Guzman, C. A. (2001). Use of live bacterial vaccine vectors for antigen
delivery: Potential and limitations. Vaccine 19, 1573–1580.
Mercenier, A., Muller-Alouf, H., and Grangette, C. (2000). Lactic acid bacteria as live
vaccines. Curr. Issues. Mol. Biol. 2, 17–25.
Mercenier, A., Pavan, S., and Pot, B. (2003). Probiotics as biotherapeutic agents: Present
knowledge and future prospects. Curr. Pharm. Des. 9, 175–191.
Metlay, J. P., and Singer, D. E. (2002). Outcomes in lower respiratory tract infections
and the impact of antimicrobial drug resistance. Clin. Microbiol. Infect. 8(Suppl. 2),
1–11.
Michalek, S. M., Eldridge, J. H., Curtiss III, R., and Rosenthal, K. L. (1994). Antigen
delivery systems: New approaches to mucosal immunization. In ‘‘Handbook
of Mucosal Immunology’’ (P. L. Ogra, J. Mestecky, M. E. Lamm, W. Strober, J. R.
McGhee, and J. Bienenstock, eds.), pp. 373–380. Academic Press, Inc., Boston.
Midolo, P. D., Lambert, J. R., Hull, R., Luo, F., and Grayson, M. L. (1995). In vitro
inhibition of Helicobacter pylori NCTC 11637 by organic acids and lactic acid
bacteria. J. Appl. Bacteriol. 79, 475–479.
Miettinen, M., Vuopio-Varkila, J., and Varkila, K. (1996). Production of human tumor
necrosis factor alpha, interleukin-6, and interleukin-10 is induced by lactic acid
bacteria. Infect. Immun. 64, 5403–5405.
Miyoshi, A., Poquet, I., Azevedo, V., Commissaire, J., Bermudez-Humaran, L., Domakova,
E., Le Loir, Y., Oliveira, S. C., Gruss, A., and Langella, P. (2002). Controlled production

USE OF RECOMBINANT LAB IN HUMAN HEALTH 57
of stable heterologous proteins in Lactococcus lactis. Appl. Environ. Microbiol. 68,3141–3146.
Moineau, S., and Goulet, J. (1997). Effect of feeding feremented milks on the pulmonary
macrophage activity of mice. Milchwissenshaft 46, 551–554.
Molenaar, D., Bron, P. A., Siezen, R. J., Bringel, F., de Vos, W. M., and Kleerebezem, M.
(2003). Comparative analysis of 20 strains of Lactobacillus plantarum using micro-
arrays. Manuscript in preparation.
Molina, L., Ramos, C., Ronchel, M. C., Molin, S., and Ramos, J. L. (1998). Construction of
an efficient biologically contained Pseudomonas putida strain and its survival in
outdoor assays. Appl. Environ. Microbiol. 64, 2072–2078.
Morita, H., He, F., Fuse, T., Ouwehand, A. C., Hashimoto, H., Hosoda, M., Mizumachi, K.,
and Kurisaki, J. (2002). Adhesion of lactic acid bacteria to caco-2 cells and their effect
on cytokine secretion. Microbiol. Immunol. 46, 293–297.
Morris, D. L. (1999). WHO position paper on oral (sublingual) immunotherapy.
Ann. Allergy Asthma Immunol. 83, 423–424.
Mosmann, T. R., and Coffman, R. L. (1989). TH1 and TH2 cells: Different patterns of
lymphokine secretion lead to different functional properties. Annu. Rev. Immunol. 7,145–173.
Mukai, T., Asasaka, T., Sato, E., Mori, K., Matsumoto, M., and Ohori, H. (2002). Inhibition
of binding of Helicobacter pylori to the glycolipid receptors by probiotic Lactobacil-
lus reuteri. FEMS Immunol. Med. Microbiol. 32, 105–110.
Murosaki, S., Yamamoto, Y., Ito, K., Inokuchi, T., Kusaka, H., Ikeda, H., and Yoshikai, Y.
(1998). Heat-killed Lactobacillus plantarum L-137 suppresses naturally fed antigen-
specific IgE production by stimulation of IL-12 production in mice. J. Allergy Clin.
Immunol. 102, 57–64.
Neeser, J. R., Granato, D., Rouvet, M., Servin, A., Teneberg, S., and Karlsson, K. A. (2000).
Lactobacillus johnsonii La1 shares carbohydrate-binding specificities with several
enteropathogenic bacteria. Glycobiology 10, 1193–1199.
Netherwood, T., Bowden, R., Harrison, P., O’Donnell, A. G., Parker, D. S., and Gilbert, H. J.
(1999). Gene transfer in the gastrointestinal tract. Appl. Environ. Microbiol. 65,5139–5141.
Neumann, E., Oliveira, M. A., Cabral, C. M., Moura, L. N., Nicoli, J. R., Vieira, E. C., Cara,
D. C., Podoprigora, G. I., and Vieira, L. Q. (1998). Monoassociation with Lactobacillusacidophilus UFV-H2b20 stimulates the immune defense mechanisms of germfree
mice. Braz. J. Med. Biol. Res. 31, 1565–1573.
Norton, P. M., Brown, H. W., Wells, J. M., Macpherson, A. M., Wilson, P. W., and
Le Page, R. W. (1996). Factors affecting the immunogenicity of tetanus toxin
fragment C expressed in Lactococcus lactis. FEMS Immunol. Med. Microbiol. 14,167–177.
Norton, P. M., Wells, J. M., Brown, H. W., Macpherson, A. M., and Le Page, R. W. (1997).
Protection against tetanus toxin in mice nasally immunized with recombinant
Lactococcus lactis expressing tetanus toxin fragment C. Vaccine 15, 616–619.
Nouaille, S., Ribeiro, L. A., Miyoshi, A., Pontes, D., Le Loir, Y., Oliveira, S. C., Langella,
P., and Azevedo, V. (2003). Heterologous protein production and delivery systems
for Lactococcus lactis. Genet. Mol. Res. 2, 102–111.
Ogawa, M., Shimizu, K., Nomoto, K., Takahashi, M., Watanuki, M., Tanaka, R., Tanaka,
T., Hamabata, T., Yamasaki, S., and Takeda, Y. (2001). Protective effect of Lactoba-
cillus casei strain Shirota on Shiga toxin-producing Escherichia coli O157:H7
infection in infant rabbits. Infect. Immun. 69, 1101–1108.

58 HANNIFFY et al.
Oggioni, M. R., Beninati, C., Boccanera, M., Medaglini, D., Spinosa, M. R., Maggi, T.,
Conti, S., Magliani, W., De Bernardis, F., Teti, G., Cassone, A., Pozzi, G., and
Polonelli, L. (2001). Recombinant Streptococcus gordonii for mucosal delivery of a
scFv microbicidal antibody. Int. Rev. Immunol. 20, 275–287.
O’Hagan, D. T. (1994). Oral immunization and the common mucosal immune system. In
‘‘Novel Delivery Systems for Oral Vaccines’’ (D. T. O’Hagan, ed.), pp. 1–24. Boca
Raton, Florida: CRC Press.
Oliveira, M. L., Monedero, V., Miyaji, E. N., Leite, L. C., Lee Ho, P., and Perez-Martinez,
G. (2003). Expression of Streptococcus pneumoniae antigens, PsaA (pneumococcal
surface antigen A) and PspA (pneumococcal surface protein A) by Lactobacillus
casei. FEMS Microbiol. Lett. 227, 25–31.
Osset, J., Bartolome, R. M., Garcia, E., Andreu, A., Boris, S., Suarez, J. E., Vazquez, F.,
Barbes, C., Reid, G., Chan, R. C., Bruce, A. W., Costerton, J. W., McGroarty, J. A.,
Oggioni, M. R., Medaglini, D., Maggi, T., Pozzi, G., and Russell, M. W. (2001).
Assessment of the capacity of Lactobacillus to inhibit the growth of uropathogens
and block their adhesion to vaginal epithelial cells. J. Infect. Dis. 183, 485–491.
Pavan, S., Hols, P., Delcour, J., Geoffroy, M. C., Grangette, C., Kleerebezem, M., and
Mercenier, A. (2000). Adaptation of the nisin-controlled expression system in
Lactobacillus plantarum: A tool to study in vivo biological effects. Appl. Environ.
Microbiol. 66, 4427–4432.
Pearson, B. M., Pin, C., Wright, J., I’Anson, K., Humphrey, T., and Wells, J. M. (2003).
Comparative genome analysis of Campylobacter jejuni using whole genome DNA
microarrays. FEBS Lett. 554, 224–230.
Pedersen, M. B., Jensen, P. R., Janzen, T., and Nilsson, D. (2002). Bacteriophage resistance
of a �thyA mutant of Lactococcus lactis blocked in DNA replication. Appl. Environ.
Microbiol. 68, 3010–3023.
Perdigon, G., Alvarez, S., Medina, M., Vintini, E., and Roux, E. (1999). Influence of the
oral administration of lactic acid bacteria on IgA producing cells associated to
bronchus. Int. J. Immunopathol. Pharmacol. 12, 97–102.
Perner, A., and Rask-Madsen, J. (1999). Review article: The potential role of nitric oxide
in chronic inflammatory bowel disorders. Aliment. Pharmacol. Ther. 13, 135–144.
Pessi, T., Sutas, Y., Saxelin, M., Kallioinen, H., and Isolauri, E. (1999). Antiproliferative
effects of homogenates derived from five strains of candidate probiotic bacteria.
Appl. Environ. Microbiol. 65, 4725–4728.
Platteeuw, C., van Alen-Boerrigter, I., van Schalkwijk, S., and de Vos, W. M. (1996). Food-
grade cloning and expression system for Lactococcus lactis. Appl. Environ. Microbiol.
62, 1008–1013.
Porwollik, S., Wong, R. M., and McClelland, M. (2002). Evolutionary genomics of
Salmonella: Gene acquisitions revealed by microarray analysis. Proc. Natl. Acad.
Sci. USA 99, 8956–8961.
Posno, M., Heuvelmans, P. T., van Giezen, M. J., Lokman, B. C., Leer, R. J., and Pouwels,
P. H. (1991). Complementation of the inability of Lactobacillus strains to utilize
D-xylose with D-xylose catabolism-encoding genes of Lactobacillus pentosus.
Appl. Environ. Microbiol. 57, 2764–2766.
Pouwels, P. H., Leer, R. J., and Boersma, W. J. (1996). The potential of Lactobacillus as a
carrier for oral immunization: Development and preliminary characterization of
vector systems for targeted delivery of antigens. J. Biotechnol. 44, 183–192.
Pouwels, P. H., Vriesema, A., Martinez, B., Tielen, F. J., Seegers, J. F., Leer, R. J., Jore, J.,
and Smit, E. (2001). Lactobacilli as vehicles for targeting antigens to mucosal tissues
by surface exposition of foreign antigens. Methods Enzymol. 336, 369–389.

USE OF RECOMBINANT LAB IN HUMAN HEALTH 59
Prantera, C., Scribano, M. L., Falasco, G., Andreoli, A., and Luzi, C. (2002). Ineffective-
ness of probiotics in preventing recurrence after curative resection for Crohn’s
disease: A randomised controlled trial with Lactobacillus GG. Gut 51, 405–409.
Ramos, J. L., Andersson, P., Jensen, L. B., Ramos, C., Ronchel, M. C., Diaz, E., Timmis, K. N.,
and Molin, S. (1995). Suicide microbes on the loose. Biotechnology (N Y) 13, 35–37.
Rath, H. C. (2003). The role of endogenous bacterial flora: Bystander or the necessary
prerequisite? Eur. J. Gastroenterol. Hepatol. 15, 615–620.
Rautava, S., and Isolauri, E. (2002). The development of gut immune responses and gut
microbiota: Effects of probiotics in prevention and treatment of allergic disease. Curr.
Issues Intest. Microbiol. 3, 15–22.
Ravn, P., Arnau, J., Madsen, S. M., Vrang, A., and Israelsen, H. (2000). The development
of TnNuc and its use for the isolation of novel secretion signals in Lactococcus lactis.
Gene 242, 347–356.
Ravn, P., Arnau, J., Madsen, S. M., Vrang, A., and Israelsen, H. (2003). Optimization of
signal peptide SP310 for heterologous protein production in Lactococcus lactis.
Microbiology 149, 2193–2201.
Reid, G. (2002). The potential role of probiotics in pediatric urology. J. Urol. 168,1512–1517.
Reid, G., and Bruce, A. W. (2001). Selection of lactobacillus strains for urogenital probi-
otic applications. J. Infect. Dis. 183, S77–S80.
Reid, G., and Burton, J. (2002). Use of Lactobacillus to prevent infection by pathogenic
bacteria. Microbes Infect. 4, 319–324.
Reid, G., Chan, R. C., Bruce, A. W., and Costerton, J. W. (1985). Prevention of urinary tract
infection in rats with an indigenous Lactobacillus casei strain. Infect. Immun. 49,320–324.
Reid, G., Charbonneau, D., Erb, J., Kochanowski, B., Beuerman, D., Poehner, R., and
Bruce, A. W. (2003). Oral use of Lactobacillus rhamnosus GR-1 and L. fermentum
RC-14 significantly alters vaginal flora: Randomized, placebo-controlled trial in 64
healthy women. FEMS Immunol. Med. Microbiol. 35, 131–134.
Rembacken, B. J., Snelling, A. M., Hawkey, P. M., Chalmers, D. M., and Axon, A. T.
(1999). Non-pathogenic Escherichia coli versus mesalazine for the treatment of
ulcerative colitis: A randomised trial. Lancet 354, 635–639.
Renault, P. (2002). Genetically modified lactic acid bacteria: Applications to food or
health and risk assessment. Biochimie 84, 1073–1087.
Repa, A., Grangette, C., Daniel, C., Hochreiter, R., Hoffmann-Sommergruber, K.,
Thalhamer, J., Kraft, D., Breiteneder, H., Mercenier, A., and Wiedermann, U.
(2003). Mucosal co-application of lactic acid bacteria and allergen induces
counter-regulatory immune responses in a murine model of birch pollen allergy.
Vaccine 22, 87–95.
Resta-Lenert, S., and Barrett, K. E. (2003). Live probiotics protect intestinal epithelial
cells from the effects of infection with enteroinvasive Escherichia coli (EIEC). Gut
52, 988–997.
Reuter, M. A., Hanniffy, S., and Wells, J. M. (2003). Expression and delivery of heterolo-
gous antigens using lactic acid bacteria. Methods Mol. Med. 87, 101–114.
Reveneau, N., Geoffroy, M. C., Locht, C., Chagnaud, P., and Mercenier, A. (2002). Com-
parison of the immune responses induced by local immunizations with recombinant
Lactobacillus plantarum producing tetanus toxin fragment C in different cellular
locations. Vaccine 20, 1769–1777.
Ribeiro, L. A., Azevedo, V., Le Loir, Y., Oliveira, S. C., Dieye, Y., Piard, J. C., Gruss, A.,
and Langella, P. (2002). Production and targeting of the Brucella abortus antigen L7/

60 HANNIFFY et al.
L12 in Lactococcus lactis: A first step towards food-grade live vaccines against
brucellosis. Appl. Environ. Microbiol. 68, 910–916.
Ring, J. (1997). Allergy and modern society: Does ‘Western life style’ promote the devel-
opment of allergies? Int. Arch. Allergy. Immunol. 113, 7–10.
Rio, M. E., Zago Beatriz, L., Garcia, H., and Winter, L. (2002). [The nutritional status
change the effectiveness of a dietary supplement of lactic bacteria on the emerging of
respiratory tract diseases in children]. Arch. Latinoam. Nutr. 52, 29–34.
Roberts, M., Chatfield, S. N., and Dougan, G. (1994). Salmonella as carriers of heterolo-
gous antigens. In ‘‘Novel Delivery Systems for Oral Vaccines’’ (D.T. O’Hagan, ed.),
pp. 27–58. Boca Raton, Florida: CRC Press.
Robinson, K., Chamberlain, L. M., Schofield, K. M., Wells, J. M., and Le Page, R. W.
(1997). Oral vaccination of mice against tetanus with recombinant Lactococcus lactis.
Nat. Biotechnol. 15, 653–657.
Romagnani, S. (1994). Regulation of the development of type 2 T-helper cells in allergy.
Curr. Opin. Immunol. 6, 838–846.
Romero, D. A., and Klaenhammer, T. R. (1991). Construction of an IS946-based composite
transposon in Lactococcus lactis subsp. lactis. J. Bacteriol. 173, 7599–7606.
Ronchel, M. C., and Ramos, J. L. (2001). Dual system to reinforce biological containment
of recombinant bacteria designed for rhizoremediation. Appl. Environ. Microbiol. 67,2649–2656.
Rosenfeldt, V., Michaelsen, K. F., Jakobsen, M., Larsen, C. N., Moller, P. L., Pedersen, P.,
Tvede, M., Weyrehter, H., Valerius, N. H., and Paerregaard, A. (2002a). Effect of
probiotic Lactobacillus strains in young children hospitalized with acute diarrhea.
Pediatr. Infect. Dis. J. 21, 411–416.
Rosenfeldt, V., Michaelsen, K. F., Jakobsen, M., Larsen, C. N., Moller, P. L., Tvede, M.,
Weyrehter, H., Valerius, N. H., and Paerregaard, A. (2002b). Effect of probiotic
Lactobacillus strains on acute diarrhea in a cohort of nonhospitalized children
attending day-care centers. Pediatr. Infect. Dis. J. 21, 417–419.
Ross, P., O’Gara, F., and Condon, S. (1990). Thymidylate synthase gene from Lactococcuslactis as a genetic marker: An alternative to antibiotic resistance genes. Appl. Envi-
ron. Microbiol. 56, 2164–2169.
Ruseler-van Embden, J. G., van Lieshout, L. M., Gosselink, M. J., and Marteau, P. (1995).
Inability of Lactobacillus casei strain GG, L. acidophilus, and Bifidobacterium bifi-dum to degrade intestinal mucus glycoproteins. Scand. J. Gastroenterol. 30, 675–680.
Rush, C., Hafner, L., and Timms, P. (1997). Protein A as a fusion partner for the expres-
sion of heterologous proteins in Lactobacillus. Appl. Microbiol. Biotechnol. 47,537–542.
Russell, M. W. (2002). Immunization for protection of the reproductive tract: A review.
Am. J. Reprod. Immunol. 47, 265–268.
Salama, N., Guillemin, K., McDaniel, T. K., Sherlock, G., Tompkins, L., and Falkow, S.
(2000). A whole-genome microarray reveals genetic diversity among Helicobacter
pylori strains. Proc. Natl. Acad. Sci. USA 97, 14668–14673.
Sanders, J. W., Venema, G., and Kok, J. (1997). A chloride-inducible gene expression
cassette and its use in induced lysis of Lactococcus lactis. Appl. Environ. Microbiol.63, 4877–4882.
Scheiner, O., and Kraft, D. (1995). Basic and practical aspects of recombinant allergens.
Allergy 50, 384–391.
Scheppler, L., Vogel, M., Zuercher, A. W., Zuercher, M., Germond, J. E., Miescher, S. M.,
and Stadler, B. M. (2002). Recombinant Lactobacillus johnsonii as a mucosal vaccine
delivery vehicle. Vaccine 20, 2913–2920.

USE OF RECOMBINANT LAB IN HUMAN HEALTH 61
Schoolnik, G. K. (2002). Functional and comparative genomics of pathogenic bacteria.
Curr. Opin. Microbiol. 5, 20–26.
Schultz, M., Veltkamp, C., Dieleman, L. A., Grenther, W. B., Wyrick, P. B., Tonkonogy,
S. L., and Sartor, R. B. (2002). Lactobacillus plantarum 299V in the treatment and
prevention of spontaneous colitis in interleukin-10-deficient mice. Inflamm.
Bowel Dis. 8, 71–80.
Seegers, J. F. (2002). Lactobacilli as live vaccine delivery vectors: Progress and prospects.
Trends Biotechnol. 20, 508–515.
Seepersaud, R., Hanniffy, S. B. P., Sizer, P., LePage, R. W. F., and Wells, J. M. (2004).
Characterisation of a novel leucine-rich repeat protein antigen from Streptococcus
agalactiae that elicits protective immunity. Submitted for publication.
Sepp, E., Julge, K., Vasar, M., Naaber, P., Bjorksten, B., and Mikelsaar, M. (1997).
Intestinal microflora of Estonian and Swedish infants. Acta Paediatr. 86, 956–961.
Shah, N. P. (2000). Probiotic bacteria: Selective enumeration and survival in dairy foods.
J. Dairy Sci. 83, 894–907.
Shanahan, F. (2001). Probiotics in inflamatory bowel disease. Gut 48, 609.
Shiba, T., Aiba, Y., Ishikawa, H., Ushiyama, A., Takagi, A., Mine, T., and Koga, Y. (2003).
The suppressive effect of bifidobacteria on Bacteroides vulgatus, a putative patho-
genic microbe in inflammatory bowel disease. Microbiol. Immunol. 47, 371–378.
Shida, K., Takahashi, R., Iwadate, E., Takamizawa, K., Yasui, H., Sato, T., Habu, S.,
Hachimura, S., and Kaminogawa, S. (2002). Lactobacillus casei strain Shirota sup-
presses serum immunoglobulin E and immunoglobulin G1 responses and systemic
anaphylaxis in a food allergy model. Clin. Exp. Allergy 32, 563–570.
Shu, Q., and Gill, H. S. (2001). A dietary probiotic (Bifidobacterium lactis HN019)
reduces the severity of Escherichia coli O157:H7 infection in mice. Med. Microbiol.
Immunol. (Berl) 189, 147–152.
Shu, Q., Lin, H., Rutherfurd, K. J., Fenwick, S. G., Prasad, J., Gopal, P. K., and Gill, H. S.
(2000). Dietary Bifidobacterium lactis (HN019) enhances resistance to oral Salmonel-
la typhimurium infection in mice. Microbiol. Immunol. 44, 213–222.
Simhon, A., Douglas, J. R., Drasar, B. S., and Soothill, J. F. (1982). Effect of feeding on
infants’ faecal flora. Arch. Dis. Child. 57, 54–58.
Sirard, J. C., Niedergang, F., and Kraehenbuhl, J. P. (1999). Live attenuated Salmonella:
A paradigm of mucosal vaccines. Immunol. Rev. 171, 5–26.
Sorensen, K. I., Larsen, R., Kibenich, A., Junge, M. P., and Johansen, E. (2000). A food-
grade cloning system for industrial strains of Lactococcus lactis. Appl. Environ.
Microbiol. 66, 1253–1258.
Stahl, S., Samuelson, P., Hansson, M., Andreoni, C., Goetsch, L., Libon, C., Liljeqvist, S.,
Gunneriusson, E., Binz, H., Nguyen, T. N., and Uhlen, M. (1997). Development of
non-pathogenic Staphylococci as vaccine delivery vehicles. In ‘‘Gram-Positive Bac-
teria as Vaccine Vehicles for Mucosal Immunisation’’ (G. Pozzi and J. M. Wells, eds.),
pp. 61–81. Springer-Verlag, Berlin.
Steidler, L., Hans, W., Schotte, L., Neirynck, S., Obermeier, F., Falk, W., Fiers, W., and
Remaut, E. (2000). Treatment of murine colitis by Lactococcus lactis secreting inter-
leukin-10. Science 289, 1352–1355.
Steidler, L., Neirynck, S., Huyghebaert, N., Snoeck, V., Vermeire, A., Goddeeris, B., Cox,
E., Remon, J. P., and Remaut, E. (2003). Biological containment of genetically mod-
ified Lactococcus lactis for intestinal delivery of human interleukin 10.
Nat. Biotechnol. 21, 785–789.
Steidler, L., Robinson, K., Chamberlain, L., Schofield, K. M., Remaut, E., Le Page, R. W.,
and Wells, J. M. (1998). Mucosal delivery of murine interleukin-2 (IL-2) and IL-6 by

62 HANNIFFY et al.
recombinant strains of Lactococcus lactis coexpressing antigen and cytokine. Infect.
Immun. 66, 3183–3189.
Strachan, D. P. (1989). Hay fever, hygiene, and household size. BMJ 299, 1259–1260.
Strober, W., Fuss, I. J., and Blumberg, R. S. (2002). The immunology of mucosal models of
inflammation. Annu. Rev. Immunol. 20, 495–549.
Strus, M., Pakosz, K., Gosciniak, H., Przondo-Mordarska, A., Rozynek, E., Pituch, H.,
Meisel-Mikolajczyk, F., and Heczko, P. B. (2001). [Antagonistic activity of Lactoba-cillus bacteria strains against anaerobic gastrointestinal tract pathogens (Helicobacter
pylori, Campylobacter coli, Campylobacter jejuni, Clostridium difficile)]. Med. Dosw.
Mikrobiol. 53, 133–142.
Szafranski, P., Mello, C. M., Sano, T., Smith, C. L., Kaplan, D. L., and Cantor, C. R. (1997).
A new approach for containment of microorganisms: Dual control of streptavidin
expression by antisense RNA and the T7 transcription system. Proc. Natl. Acad. Sci.
USA 94, 1059–1063.
Szajewska, H., and Mrukowicz, J. Z. (2001). Probiotics in the treatment and prevention of
acute infectious diarrhea in infants and children: A systematic review of published
randomized, double-blind, placebo-controlled trials. J. Pediatr. Gastroenterol. Nutr.
33(Suppl. 2), S17–S25.
Taha, T. E., Hoover, D. R., Dallabetta, G. A., Kumwenda, N. I., Mtimavalye, L. A., Yang,
L. P., Liomba, G. N., Broadhead, R. L., Chiphangwi, J. D., and Miotti, P. G. (1998).
Bacterial vaginosis and disturbances of vaginal flora: Association with increased
acquisition of HIV. AIDS 12, 1699–1706.
Takala, T. M., and Saris, P. E. (2002). A food-grade cloning vector for lactic acid bacteria
based on the nisin immunity gene nisI. Appl. Microbiol. Biotechnol. 59, 467–471.
Takala, T. M., Saris, P. E., and Tynkkynen, S. S. (2003). Food-grade host/vector expres-
sion system for Lactobacillus casei based on complementation of plasmid-associated
phospho-beta-galactosidase gene lacG. Appl. Microbiol. Biotechnol. 60, 564–570.
Tejada-Simon, M. V., Lee, J. H., Ustunol, Z., and Pestka, J. J. (1999). Ingestion of yogurt
containing Lactobacillus acidophilus and Bifidobacterium to potentiate immuno-
globulin A responses to cholera toxin in mice. J. Dairy Sci. 82, 649–660.
Thole, J. E., van Dalen, P. J., Havenith, C. E., Pouwels, P. H., Seegers, J. F., Tielen, F. D.,
van der Zee, M. D., Zegers, N. D., and Shaw, M. (2000). Live bacterial delivery
systems for development of mucosal vaccines. Curr. Opin. Mol. Ther. 2, 94–99.
Threlfall, E. J. (2002). Antimicrobial drug resistance in Salmonella: Problems and per-
spectives in food- and water-borne infections. FEMS Microbiol. Rev. 26, 141–148.
Tlaskalova-Hogenova, H., Tuckova, L., Lodinova-Zadnikova, R., Stepankova, R.,
Cukrowska, B., Funda, D. P., Striz, I., Kozakova, H., Trebichavsky, I., Sokol, D.,
Rehakova, Z., Sinkora, J., Fundova, P., Horakova, D., Jelinkova, L., and Sanchez,
D. (2002). Mucosal immunity: Its role in defense and allergy. Int. Arch. Allergy
Immunol. 128, 77–89.
van de Guchte, M., Serror, P., Chervaux, C., Smokvina, T., Ehrlich, S. D., and Maguin, E.
(2002). Stress responses in lactic acid bacteria. Antonie Van Leeuwenhoek 82,187–216.
van Deventer, S. J., Elson, C. O., and Fedorak, R. N. (1997). Multiple doses of intravenous
interleukin 10 in steroid-refractory Crohn’s disease. Crohn’s Disease Study Group.
Gastroenterology 113, 383–389.
van Ginkel, F. W., Nguyen, H. H., and McGhee, J. R. (2000). Vaccines for mucosal
immunity to combat emerging infectious diseases. Emerg. Infect. Dis. 6, 123–132.
Venturi, A., Gionchetti, P., Rizzello, F., Johansson, R., Zucconi, E., Brigidi, P., Matteuzzi,
D., and Campieri, M. (1999). Impact on the composition of the faecal flora by a new

USE OF RECOMBINANT LAB IN HUMAN HEALTH 63
probiotic preparation: Preliminary data on maintenance treatment of patients with
ulcerative colitis. Aliment. Pharmacol. Ther. 13, 1103–1108.
Vesa, T., Pochart, P., and Marteau, P. (2000). Pharmacokinetics of Lactobacillus plantar-um NCIMB 8826, Lactobacillus fermentum KLD, and Lactococcus lactis MG 1363 in
the human gastrointestinal tract. Aliment. Pharmacol. Ther. 14, 823–828.
Villa-Komaroff, L., Efstratiadis, A., Broome, S., Lomedico, P., Tizard, R., Naber, S. P.,
Chick, W. L., and Gilbert, W. (1978). A bacterial clone synthesizing proinsulin. Proc.Natl. Acad. Sci. USA 75, 3727–3731.
Vinderola, C. G., Medici, M., and Perdigon, G. (2004). Relationship between interaction
sites in the gut, hydrophobicity, mucosal immunomodulating capacities and cell
wall protein profiles in indigenous and exogenous bacteria. J. Appl. Microbiol. 96,230–243.
Vitini, E., Alvarez, S., Medina, M., Medici, M., de Budeguer, M. V., and Perdigon, G.
(2000). Gut mucosal immunostimulation by lactic acid bacteria. Biocell 24,223–232.
von der Weid, T., Bulliard, C., and Schiffrin, E. J. (2001). Induction by a lactic acid
bacterium of a population of CD4(þ) T cells with low proliferative capacity that
produce transforming growth factor beta and interleukin-10. Clin. Diagn. Lab.Immunol. 8, 695–701.
von Mutius, E., Martinez, F. D., Fritzsch, C., Nicolai, T., Roell, G., and Thiemann, H. H.
(1994). Prevalence of asthma and atopy in two areas of West and East Germany.
Am. J. Respir. Crit. Care. Med. 149, 358–364.
von Wright, A., and Bruce, A. W. (2003). Genetically modified microorganisms and their
potential effects on human health and nutrition. Trends Food Sci. Technol. 14,264–276.
von Wright, A., Wessels, S., Tynkkynen, S., and Saarela, M. (1990). Isolation of a replica-
tion region of a large lactococcal plasmid and use in cloning of a nisin resistance
determinant. Appl. Environ. Microbiol. 56, 2029–2035.
Walsh, C. (2000). Molecular mechanisms that confer antibacterial drug resistance. Nature406, 775–781.
Walter, J., Heng, N. C., Hammes, W. P., Loach, D. M., Tannock, G. W., and Hertel, C.
(2003). Identification of Lactobacillus reuteri genes specifically induced in the mouse
gastrointestinal tract. Appl. Environ. Microbiol. 69, 2044–2051.
Walter, J., Hertel, C., Tannock, G. W., Lis, C. M., Munro, K., and Hammes, W. P. (2001).
Detection of Lactobacillus, Pediococcus, Leuconostoc, and Weissella species in hu-
man feces by using group-specific PCR primers and denaturing gradient gel electro-
phoresis. Appl. Environ. Microbiol. 67, 2578–2585.
Walter, J., Tannock, G. W., Tilsala-Timisjarvi, A., Rodtong, S., Loach, D. M., Munro, K.,
and Alatossava, T. (2000). Detection and identification of gastrointestinal Lactobacil-
lus species by using denaturing gradient gel electrophoresis and species-specific PCR
primers. Appl. Environ. Microbiol. 66, 297–303.
Wells, J. M., and Bennik, M. H. J. (2003). Impact of genomics on foodborne pathogens.
Nutri. Res. Rev. 16, 21–35.
Wells, J. M., and Mercenier, A. (2003). Lactic acid bacteria as mucosal delivery vehicles.
In ‘‘The Lactic Acid Bacteria,’’ Vol III, pp. 260–291. Kluwer Academic/Plenum
Publishers, New York.
Wells, J. M., and Pozzi, G. (1997). An overview of gram-positive bacteria as vaccine
vehicles for musocal immunization. In ‘‘Gram-Positive Bacteria as Vaccine Vehicles
for Mucosal Immunisation’’ (G. Pozzi and J. M. Wells, eds.), pp. 1–8. Springer-Verlag,
Berlin.

64 HANNIFFY et al.
Wells, J. M., Robinson, K., Chamberlain, L. M., Schofield, K. M., and Le Page, R. W.
(1996). Lactic acid bacteria as vaccine delivery vehicles. Antonie Van Leeuwenhoek
70, 317–330.
Wells, J. M., and Schofield, K. M. (1996). Cloning and expression systems for lactococci.
Nato Asi. Ser. H. 98, 37–63.
Wells, J. M., Wilson, P. W., Norton, P. M., Gasson, M. J., and Le Page, R. W. (1993a).
Lactococcus lactis: High-level expression of tetanus toxin fragment C and protection
against lethal challenge. Mol. Microbiol. 8, 1155–1162.
Wells, J. M., Wilson, P. W., Norton, P. M., and Le Page, R. W. (1993b). A model system for
the investigation of heterologous protein secretion pathways in Lactococcus lactis.
Appl. Environ. Microbiol. 59, 3954–3959.
Wheeler, A. W., and Woroniecki, S. R. (2001). Immunological adjuvants in allergy
vaccines: Past, present and future. Allergology International 50, 295–301.
Wiedermann, U. (2003). Mucosal immunity–mucosal tolerance. A strategy for treatment
of allergic diseases. Chem. Immunol. Allergy 82, 11–24.
Wiedermann, U., Jahn-Schmid, B., Fritsch, R., Bauer, L., Renz, H., Kraft, D., and Ebner, C.
(1998). Effects of adjuvants on the immune response to allergens in a murine model
of allergen inhalation: Cholera toxin induces a Th1-like response to Bet v 1, the major
birch pollen allergen. Clin. Exp. Immunol. 111, 144–151.
Wiedermann, U., Jahn-Schmid, B., Lindblad, M., Rask, C., Holmgren, J., Kraft, D., and
Ebner, C. (1999). Suppressive versus stimulatory effects of allergen/cholera toxoid
(CTB) conjugates depending on the nature of the allergen in a murine model of type I
allergy. Int. Immunol. 11, 1131–1138.
Wiesenfeld, H. C., Hillier, S. L., Krohn, M. A., Landers, D. V., and Sweet, R. L. (2003).
Bacterial vaginosis is a strong predictor of Neisseria gonorrhoeae and Chlamydia
trachomatis infection. Clin. Infect. Dis. 36, 663–668.
Wu, H. Y., Abdu, S., Stinson, D., Russell, M. W., Moldoveanu, Z., Huang, W. Q.,
Compans, R. W., and Mestecky, J. (2000). Generation of female genital tract antibody
responses by local or central (common) mucosal immunization: Compartmentaliza-
tion within the common mucosal immune system. Infect. Immun. 68, 5539–5545.
Xin, K. Q., Hoshino, Y., Toda, Y., Igimi, S., Kojima, Y., Jounai, N., Ohba, K., Kushiro, A.,
Kiwaki, M., Hamajima, K., Klinman, D., and Okuda, K. (2003). Immunogenicity and
protective efficacy of orally administered recombinant Lactococcus lactis expressing
surface-bound HIV Env. Blood 102, 223–228.
Zegers, N. D., Kluter, E., van Der Stap, H., van Dura, E., van Dalen, P., Shaw, M., and
Baillie, L. (1999). Expression of the protective antigen of Bacillus anthracis by
Lactobacillus casei: Towards the development of an oral vaccine against anthrax.
J. Appl. Microbiol. 87, 309–314.
Zoetendal, E. G., Akkermans, A. D., and De Vos, W. M. (1998). Temperature gradient gel
electrophoresis analysis of 16S rRNA from human fecal samples reveals stable and
host-specific communities of active bacteria. Appl. Environ. Microbiol. 64,3854–3859.
Zoetendal, E. G., Akkermans, A. D. L., Akkermans-van Vliet, W. M., de Visser, J. A. G. M.,
and de Vos, W. M. (2001). The host genotype affects the bacterial community in the
human gastrointestinal tract. Microbial Ecol. Health Dis. 13, 129–134.
Zoetendal, E. G., von Wright, A., Vilpponen-Salmela, T., Ben-Amor, K., Akkermans, A. D.,
and de Vos, W. M. (2002). Mucosa-associated bacteria in the human gastrointestinal
tract are uniformly distributed along the colon and differ from the community
recovered from feces. Appl. Environ. Microbiol. 68, 3401–3407.
Top Related

Copyright © 2022 FDOKUMEN

