







Bahasa
Halaman
Hukum


1 23
EuphyticaInternational Journal of PlantBreeding ISSN 0014-2336Volume 178Number 2 Euphytica (2010) 178:203-214DOI 10.1007/s10681-010-0298-5
Pollen morphology as fertility predictor inhybrid tea roses

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
Science+Business Media B.V.. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

Pollen morphology as fertility predictor in hybrid tea roses
Luca Pipino • Marie-Christine Van Labeke •
Andrea Mansuino • Valentina Scariot •
Annalisa Giovannini • Leen Leus
Received: 23 February 2010 / Accepted: 1 November 2010 / Published online: 15 November 2010
� Springer Science+Business Media B.V. 2010
Abstract Fertility of hybrid tea roses is often
reduced due to their interspecific origin but also to
intensive inbreeding. New genotypes used as pollen
donors represent an economic risk for a breeding
programme, as their influence on seed production is
unknown. In this study 11 cut rose genotypes were
selected from a company database as high fertile or
low fertile male parents, according to the number of
seeds per hybridisation. Pollen morphology and in
vitro germination of the selected genotypes were
characterised. Pollen was either small (mean diameter
\30 lm), shrunken, and irregular (abnormal), or
large (mean diameter[30 lm), elliptical and crossed
by furrows (normal). High correlations were found
between the number of seeds produced per hybrid-
isation and the pollen diameter (r = 0.94) or the
percentage of normal pollen (r = 0.96). In order to
evaluate the predictive power of the models, we
conducted regression analyses and performed a
validation experiment on genotypes not present in
the database and without background information on
fertility. Pollen diameter and percentage of normal
pollen were characterised and fitted in the regression
models for seed set predictions. Validation with an
independent dataset gave a good prediction for 83.3%
of the data. This indicates that using either the mean
pollen diameter or the percentage of normal pollen
resulted in effective fertility prediction. This tool
could enhance the genetic variability in crossings
between hybrid tea roses, thus creating possibilities
for less economically risky exploitation of new
tetraploid genotypes as male parents.
Keywords Breeding � Hybridisation �Pollen diameter � Rosa � Seed production
Introduction
Cut rose breeding programmes in the twentieth
century focused mainly on introduction of new flower
colours, thornless stems, higher production and a
L. Pipino � V. Scariot
Department of Agronomy, Forest and Land Management,
University of Turin, Via Leonardo da Vinci 44,
10095 Grugliasco, TO, Italy
L. Pipino � M.-C. Van Labeke
Department of Plant Production, Ghent University,
Coupure Links 653, 9000 Ghent, Belgium
A. Mansuino
NIRP International, Az. Agricola di Ghione L. & Figli,
via San Rocco 1, Fraz. Bevera, 18039 Ventimiglia,
IM, Italy
A. Giovannini
CRA-FSO, Research Unit for Floriculture and Ornamental
Species, Corso Inglesi 508, 18038 Sanremo, IM, Italy
L. Pipino � L. Leus (&)
Plant Sciences Unit, ILVO, Institute for Agricultural and
Fisheries Research, Caritasstraat 21, 9090 Melle, Belgium
e-mail: [email protected]
123
Euphytica (2011) 178:203–214
DOI 10.1007/s10681-010-0298-5
Author's personal copy

good post-harvest performance. Yet, the fertility of cut
roses, typically tetraploid (2n = 2x = 28) hybrid tea
roses, is frequently reduced probably due to their
interspecific origin but also to intensive inbreeding (de
Vries and Dubois 1996; Debener et al. 2000). To avoid
the risk of very low seed production, breeders hybri-
dise using high pollen loads of a limited number of
male parents with the desired characteristics (Zlesak
2006), chosen for their known fertility (i.e., number of
seeds produced per hybridisation). New pollen donors
represent an economic risk for a breeding programme,
as their influence on seed production is unknown.
However, generating wider populations in terms of
number of seedlings is only one of the aspects that
contribute to the overall efficiency of a breeding
programme. The successful introduction of a new
parent into a breeding programme depends on its
potential to transmit the desired traits to the offspring.
It may be more efficient in some cases to choose a
parent with moderate fertility if it produces offspring
with a higher rate of advanced selections and,
ultimately, cultivars. In any case, recognising param-
eters related to fertility could increase the overall
efficiency of a breeding programme.
Since the end of the 1970s, several studies
concerning pollen quality in vitro (germination and
pollen tube elongation) or in vivo (considering the
hip set or the mean number of seeds per hip) were
carried out with the aim of improving sexual
reproduction of roses (Visser et al. 1977a, b; de
Vries and Dubois 1987; Gudin and Arene 1991, 1992;
Gudin et al. 1991a, b; Zlesak et al. 2007). These
studies mainly focused on the environmental effects
which influenced pollen quality, in order to define the
best conditions to pollinate or to store the pollen. The
integration of this knowledge on pollen quality
increased the number of seedlings obtained per
hybridisation by more than 200% (Gudin 1995).
Visser et al. (1977b) observed that pollen viability of
hybrid tea roses is influenced by the season and is
positively correlated to seedling production. Gudin
et al. (1991a) observed seasonal influence on pollen
fertility and also on pollen size. Temperatures
between 23 and 30�C and a relative humidity of
about 60–65% are the best conditions for in vitro
pollen germination and pollen tube elongation (Gudin
et al. 1991b). De Vries and Dubois (1987) observed
how in vivo temperatures between 22 and 26�C
improve pollination results.
In the past, pollen size has been widely studied by
means of pollen diameter. This morphological char-
acteristic has been correlated to several other param-
eters but not to seed set. Darwin (1884) measured the
pollen diameter of 44 species, observing how pollen
size was positively correlated to the distance between
the surface of the stigma and the transmission tissue of
the style. More recently, Cruden (2009) confirmed this
correlation, suggesting that after germination pollen
grows autotrophically through the stigma to reach the
resources in the style. Erlanson (1931a) studied the
relation between percentage of empty pollen grains and
the ploidy level in Rosa species; the same author
proposed a positive relation between microspore
diameter and ploidy (1931b). In roses, 2n pollen has
a diameter 1.2–1.3 times larger than n pollen
(Darlington 1937). Crespel et al. (2006) used this
information to confirm the presence of 2n pollen in a
population of diploid interspecific hybrids. Most
studies on fertility have been performed on cut roses,
as they are the most important commercial group.
Nevertheless, it can be expected that this data on
fertility can be useful for other types of roses. For
instance, Zlesak (2009) created a model of ploidy
prediction measuring the pollen diameter of a broad
range of different rose cultivars, species and breeding
lines.
No studies carried out on roses have focused on the
screening of breeding databases to aid the individu-
ation of fertility related markers, and therefore no
reliable model of fertility prediction has yet been
published. The aim of our study was to investigate the
relationship between fertility and pollen characteris-
tics. We selected genotypes of hybrid tea roses from a
cut rose breeding database based on the number of
seeds produced per hybridisation. We then investi-
gated pollen morphology and germination of the
selected genotypes and correlated this data with
fertility parameters, i.e. seeds per hybridisation, mean
seed germination and mean seed germination per hip.
Materials and methods
Genotype selection, plant material and pollen
sampling
Hybrid tea roses were selected among 213 genotypes
from a database, consisting of 14,109 hybridisations
204 Euphytica (2011) 178:203–214
123
Author's personal copy

performed from 1994 to 2007 at the company NIRP
International (Cuers, France; Ventimiglia, Italy). The
ploidy level of a subset of 30 genotypes was assessed
by flow cytometry according to De Schepper et al.
(2001), recognising all genotypes as tetraploid.
Considering the origin of hybrid tea roses, the general
knowledge on their ploidy level and the fact that most
breeders choose to work with tetraploid genotypes
(Tackholm 1923; Erlanson 1938; Krussmann 1981;
de Vries and Dubois 1996; Crespel et al. 2002; Zlesak
2006), we assumed that the cut roses used at NIRP
International were tetraploid. We introduced three
criteria to reduce the bias created by cross combina-
tions using different seed parents on the character-
isation of the fertility of a genotype. Namely, we
excluded male parents from the database screening if
they were used with less than 20 different seed
parents, in less than 100 crossings and during less
than six crossing years. These restrictions provided a
subset of 107 tetraploid genotypes, used as male
parent in the breeding programmes. For these rose
genotypes, the number of seeds produced per hybrid-
isation was calculated. Four of the most fertile
genotypes (mean number of seeds produced per
hybridisation [15) and seven of the least fertile
(mean number of seeds produced per hybridisation
\7) were chosen. These two groups were called
‘‘high fertile’’ and ‘‘low fertile’’, respectively
(Table 1). For each of the 11 selected genotypes
two other parameters were calculated: mean seed
germination (%) meaning the germination per total
number of obtained seeds (values originating from
less than 10 hips per female parent were not
considered) and the mean seed germination (%) per
hip, calculated as the ratio between the seed germi-
nation (%) and the number of hips originated from
the male parent. Flow cytometry analysis confirmed
the tetraploidy of the 11 selected genotypes.
Twenty plants of each genotype were cultivated in
single pots with perlite at NIRP International (Cuers,
France) and fertilised according to the company’s
practises. The glasshouses were unheated but the
monthly mean outside day/night temperatures were
never less than 5.9�C. During June 2008, the monthly
mean outside temperatures were 24 ± 7.1�C (data of
the meteorological station of Cuers, Chambre d’Agri-
culture du Var, France). Pollen of the selected geno-
types was obtained as a bulk of at least 20 flowers (each
one collected from a different plant of the same
genotype) by gathering the anthers during June 2008.
Following the pollination method used at NIRP, the
anthers were placed at room temperature in open Petri
Table 1 Fertility parameters of the 11 male parents selected
from the database (number of seeds per hybridisation, mean
seed germination, mean seed germination per hip) and pollen
characteristics (in vitro germination, diameter, diameter and
percentage of normal pollen)
Fertility Pollen
Total Normal
Group Genotype
code
Seeds/hyb. Seed germ. (%) Seed germ.
(%)/hip
Germ. (%) Diam. (lm) Diam. (lm) (%)
High fertile 1848 21.3 ± 10.7A aB 29.9 ± 16.9 a 5.9 ± 9.8 a 31.8 ± 1.6 b 40.8 ± 8.6 a 43.1 ± 6.9 a 73.1
2518 20.0 ± 12.6 a 24.3 ± 13.7 a 2.7 ± 1.7 a 46.5 ± 2.6 a 40.2 ± 9.6 a 43.6 ± 7.3 a 68.8
2756 15.5 ± 14.8 a 37.1 ± 12.4 a 3.7 ± 2.3 a 22.9 ± 1.5 b 39.4 ± 10.2 ab 43.3 ± 8.1 a 59.7
2491 16.0 ± 17.9 a 36.3 ± 14.5 a 4.2 ± 5.5 a 22.8 ± 3.7 b 35.1 ± 9.3 b 40.1 ± 6.2 b 63.5
Low fertile 2376 4.4 ± 5.2 b 24.7 ± 18.6 a 1.2 ± 1.3 a 14.6 ± 2.3 c 29.3 ± 10.1 c 39.6 ± 6.8 b 27.5
1331 6.3 ± 8.4 b 19.3 ± 15.1 a 1.8 ± 3.0 a 18.9 ± 0.9 c 28.3 ± 9.0 c 38.0 ± 5.4 c 24.4
1999 1.1 ± 2.3 b 22.8 ± 16.8 a 1.0 ± 1.8 b 18.4 ± 1.5 c 28.0 ± 10.0 c 38.9 ± 6.5 c 25.5
2695 2.7 ± 3.2 b 36.0 ± 19.5 a 2.6 ± 2.1 a 0 d 27.4 ± 8.6 cd 37.1 ± 5.6 c 19.3
1985 5.1 ± 6.3 b 23.0 ± 13.3 a 2.0 ± 2.7 a 13.6 ± 0.2 c 26.6 ± 8.6 d 37.2 ± 5.0 c 26.7
2145 3.8 ± 6.1 b 18.5 ± 12.8 a 2.4 ± 5.6 a 5.8 ± 1.6 d 24.8 ± 7.0 e 35.4 ± 4.4 c 10.2
1627 1.9 ± 3.9 b 15.4 ± 16.7 b 1.3 ± 5.5 a 4.6 ± 0.5 d 21.3 ± 7.3 f 36.2 ± 4.8 c 9.5
A Mean ± standard deviationB Different letter indicates significant differences at the 0.05 level
Euphytica (2011) 178:203–214 205
123
Author's personal copy

dishes in order to favour release and drying of the
pollen (Crespel and Mouchotte 2003). After two days
the Petri dishes were sealed and stored at -20�C.
Pollen was sent to ILVO (Belgium) for analysis.
Pollen germination
In vitro pollen germination of the 11 selected genotypes
was assessed using germination medium (40 mg l-1
H3BO3; 152 mg l-1 CaCl2(H2O); 150 g l-1 sucrose;
7 g l-1 agar; pH 5.6; Leus 2005) using one Petri dish
(diameter 9 cm) for each replicate. Three replicates
were performed for each rose genotype. Pollen was
spread on the medium with a small paint brush.
Germination was evaluated after 24 h incubation at
22 ± 2�C in dark conditions using a Leitz Laborlux D
Binocular Phase Contrast microscope (Leica Micro-
systems GmbH, Wetzlar, Germany) at a magnification
of 1009. Pollen was considered germinated when a
pollen tube reached a length of at least 1.5 times the
pollen diameter (Leus 2005). The mean pollen
germination (%) was calculated as the germination
per total number of observed pollen grains (200) on
the Petri dish. Pollen tube length was also observed
for each genotype.
Pollen diameter
Dry pollen was dusted onto a glass slide without a
cover slip. No hydration or embedding was per-
formed, because imbibed pollen swells, resulting in
significant changes in pollen diameter and a globular
shape (data not shown). Microscopic observations
(1609) were made using a Leica DMIRB microscope
(Leica Microsystems GmbH, Wetzlar, Germany).
Stored digital images of the pollen were used to
analyse the pollen size with LAS software (Leica
Application Suite). To measure the pollen diameter,
the major axis (transverse diameter) of the ellipsoidal
dry pollen grain was considered. In the text below,
the term ‘diameter’ describes ‘the major axis’. The
diameter of at least 400 pollen grains of the collected
bulk of each genotype was measured.
Paternal effect on seed set and offspring
germination
One high fertile and one low fertile female parent
(genotypes 1985 and 1627, respectively) as well as
one high fertile and one low fertile male parent
(genotypes 1848 and 1999, respectively) were selected
from the database. The same pollen samples used for
the in vitro germination experiment and the diameter
measurement were used for the crossings. The high
fertile seed parent 1985 was crossed with the high
fertile pollen parent 1848 (334 pollinations) and the
low fertile pollen parent 1999 (60 pollinations). The
same pollen donors were used on the low fertile seed
parent 1627, leading to 198 and 213 pollinations,
respectively. The crossings were carried out during
the first 10 days of June 2008, at the NIRP Interna-
tional glasshouses (Cuers, France). Seeds were gath-
ered and counted at the beginning of October 2008.
Afterwards seeds were stratified in sand at 18 ± 2�C
for 22 days and then transferred at 5 ± 1�C for
40 days. Per cross combination, four replicates of one
hundred seeds were sown in perlite beds at a
germination density of 209 seeds m-2. The experi-
mental set-up of the germination experiment was a
randomized complete block design. Germinations
were counted after 90 days.
Statistical analysis
Pollen germination and diameter measurements of
the genotypes were analysed using a one-way
ANOVA and Tukey’s HSD test for multiple com-
parisons. Student’s t-test was used to compare the
means of the high and low fertility group. The
frequency distribution of normal and abnormal pollen
was studied using a Chi-square test (Pearson).
Correlations (Pearson) were calculated between
the pollen characteristics (pollen diameter (lm),
normal pollen (%), normal pollen diameter (lm),
pollen germination (%)) and fertility parameters
(seeds/hybridisation, mean seed germination (%)
and mean seed germination (%)/hip). If correlations
were higher than r = 0.90 regression analysis was
conducted on the data. Analyses were performed with
SPSS 15.0 (SPSS Inc., Chicago, IL).
Validation experiment
To evaluate the predictive power of the regression
analysis, 18 genotypes not present in the database and
without background information on fertility were
introduced in the breeding programme, using random
seed parents for the crossings. The pollen of these
206 Euphytica (2011) 178:203–214
123
Author's personal copy

genotypes, gathered during June 2008, was dried and
analysed as described above. In this experiment the
diameter of at least 100 pollen grains per genotype
was measured. Seeds were gathered and counted in
October to evaluate the actual number of seeds
produced per hybridisation by each genotype. The
pollen data of these genotypes were then used as
predictors for the number of seeds produced per
hybridisation using the regression equation. The
predictions were validated by checking which values
of actual number of seeds produced per hybridisation
exceeded the prediction interval (95%).
Results
Genotype selection
For the 107 genotypes used as pollen donors and
matching the minimal hybridisation standards in the
database analysis, the mean number of seeds produced
per hybridisation was 9.13 ± 9.30. The fertility
parameters for the selected subset of 11 genotypes
are given in Table 1. For the two groups established
(‘‘high fertile’’ and ‘‘low fertile’’) the number of seeds
per hybridisation differed significantly (F10,325 =
17.79, P \ 0.001) (t = 1.98, df = 111, P =
9.27E-16). Mean seed germination, as a percentage
of the obtained seeds, was only significantly lower for
genotype 1627 compared to the other genotypes
(F10,193 = 4.65, P \ 0.001). The mean seed germina-
tion per hip, calculated as the ratio between the seed
germination (%) and the number of hips originated
from the same male parent, was only significantly
lower for genotype 1999 (F10,374 = 2.96, P = 0.001)
(Table 1). If data were clustered for fertility, the high
fertile group showed a significantly higher amount of
germinated seeds (mean 32.1 ± 15.2%) than those of
the low fertile group (mean 22.4 ± 17.2%) (t = 1.97,
df = 202, P = 3.8E-05). Also for mean seed germi-
nation per hip, the genotypes of the high fertile group
had a significantly higher percentage of germinated
seeds per hybridisation (mean 4.1 ± 5.8%) compared
to the low fertile group (mean 1.7 ± 4.3%) (t = 1.98,
df = 131, P = 0.0002).
Pollen germination
The mean pollen germination varied significantly
between the 11 genotypes and ranged from 0%
(genotype 2695) to 46.5% (genotype 2518) (F10,22 =
52.96, P \ 0.001) (Table 1). A repeat experiment
(June 2009) confirmed that pollen of genotype 2695
does not germinate (data not shown). The high fertile
group was characterised by a significantly higher mean
pollen germination (31.02 ± 10.73%) compared to the
mean of the low fertile group (10.85 ± 7.22%)
(t = 2.10, df = 17, P = 2.12E-05).
Pollen diameter
The mean pollen diameter varied significantly among
the 11 genotypes studied (F10,10147 = 530.83,
P \ 0.001) (Table 1). The group of high fertile
genotypes was characterised by larger diameters
(mean 38.43 ± 9.79 lm) than low fertile genotypes
(mean 26.15 ± 9.13 lm) (t = 1.96, df = 10156,
P = 0). Microscopic observations revealed two types
of pollen grains: undeveloped with a diameter smaller
than 30 lm and ellipsoidal with a diameter larger
than 30 lm (Fig. 1). Therefore the distribution of the
pollen in two classes, i.e., abnormal pollen (diameter
smaller than 30 lm) and normal pollen (diameter
larger than 30 lm), was calculated for both the high
and low fertile group (Table 2). A significantly
higher percentage of normal pollen was found for
Fig. 1 Pollen of a low
fertile (a) and a high fertile
(b) genotype
Euphytica (2011) 178:203–214 207
123
Author's personal copy

the high fertile group (Pearson Chi-square = 2075.1,
P \ 0.001). Only 20.6% of the pollen of the low
fertile group was normal, while the high fertile group
had 65.6% normal pollen.
The mean diameter of the normal pollen was
calculated for each genotype as well as its percentage
of the whole population (Table 1). The 11 genotypes
were characterised by percentages of normal pollen
varying between 9.5 and 73.1%. When the genotypes
were analysed for their mean normal pollen diameter,
three significantly different groups could be distin-
guished: one composed of three high fertile genotypes
(1848, 2518 and 2756), a second composed of one
high and one low fertile genotype (2491 and 2376)
and a third composed of six low fertile genotypes
(1331, 1999, 2695, 1985, 2145 and 1627) (F10,4873 =
76.04, P \ 0.001) (Table 1). When data were clus-
tered for fertility, the diameter of the normal pollen of
the high fertile group (mean 42.37 ± 7.29 lm) was
also significantly higher than the normal pollen of the
low fertile group (mean 37.79 ± 5.79 lm) (t = 1.96,
df = 4504, P = 5.21E-122).
Paternal effect on seed set and offspring
germination
In order to strengthen the database observation by
evaluating the paternal effect on seed set and
germination, a crossing experiment was performed
with four genotypes selected from the database
(Table 3). According to the database, the mean number
of seeds per hybridisation of the two female parents
1985 and 1627 was 19.6 ± 18.7 and 2.1 ± 3.0,
respectively. The mean number of seeds per hybrid-
isation of the two male parents 1848 and 1999 was
21.3 ± 10.7 and 1.1 ± 2.3, respectively. When pollen
of the high fertile male parent (1848) was used on both
female parents the number of seeds produced per
pollination was very high compared to the low fertile
male parent (1999) and to its value in the database. In
crosses with pollen donor 1999, the number of seeds
produced per pollination was very low for genotype
1985 and almost equal to zero for the low fertile seed
parent 1627 (Table 3). Seeds of the high fertile
genotype 1985 9 1848 reached 45.8% of germination,
while no germination occurred among the progeny
sired from pollen donor 1999. Only 13.6% of the seeds
of 1627 9 1848 germinated, and no seeds of the
crossing with 1999 germinated (Table 3).
Data analysis
The correlations (Pearson) for the variables pollen
diameter (lm), normal pollen (%), normal pollen
diameter (lm), pollen germination (%), seeds/hybrid-
isation, mean seed germination (%) and mean seed
germination (%)/hip are presented in Table 4. Seeds
per hybridisation correlated strongly with the mean
pollen diameter and percentage of normal pollen
(r [ 0.90); a strong correlation was also obtained
with the mean diameter of normal pollen (r = 0.87).
The correlations of mean seed germination per hip
with pollen diameter, percentage of normal pollen
and seeds per hybridisation were still high (respec-
tively r = 0.75, r = 0.78 and r = 0.83). The
Table 2 Frequency distribution of normal pollen among the
genotypes sorted into fertility groups
Group Normal pollen Total (counts)
counts %
High fertile 2,607 65.6 3,974
Low fertile 1,274 20.6 6,184
Table 3 Paternal effect on seed set (number of seeds per pollination) and offspring germination (mean seed germination): a crossing
experiment
Seed parent Pollen donor
High fertile (1848) Low fertile (1999)
Seeds/pollination Seed germ. (%) Seeds/pollination Seed germ. (%)
High fertile (1985) 60.2 45.8 ± 4.6A 1.80 0
Low fertile (1627) 9.0 13.6 ± 4.9 0.05 0
A Mean ± standard deviation
208 Euphytica (2011) 178:203–214
123
Author's personal copy
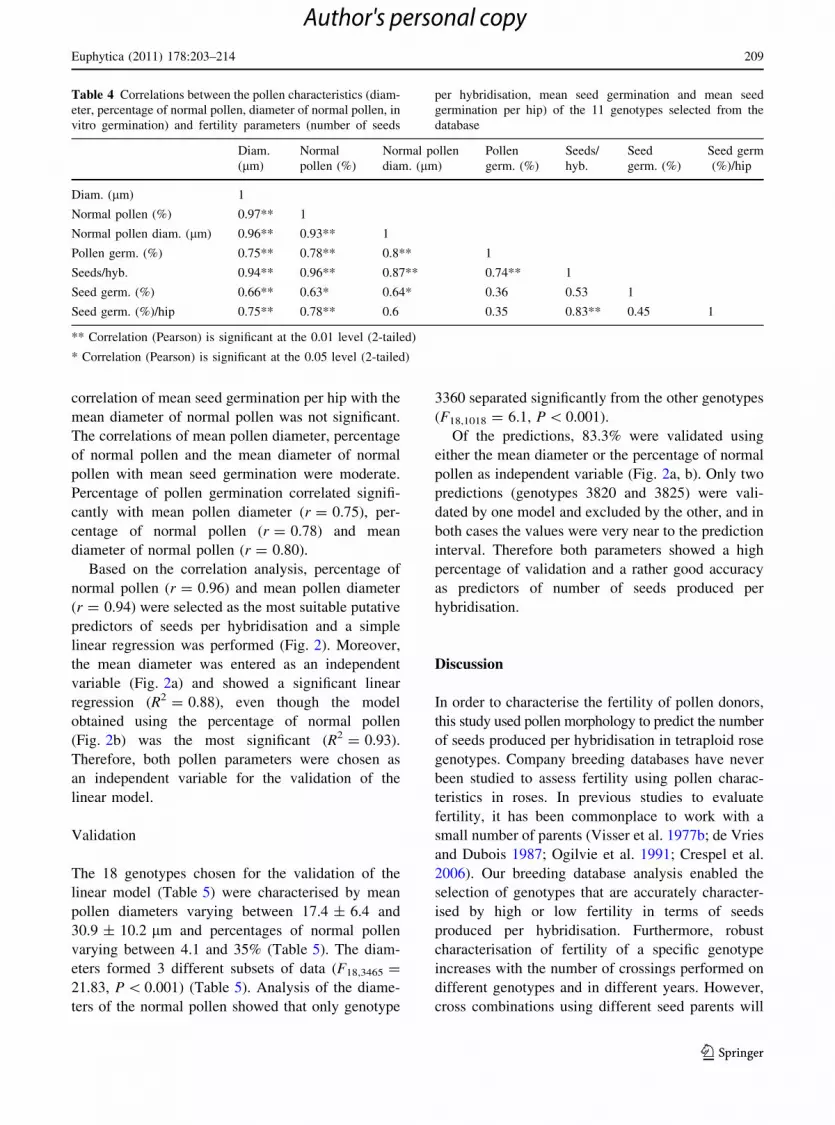
correlation of mean seed germination per hip with the
mean diameter of normal pollen was not significant.
The correlations of mean pollen diameter, percentage
of normal pollen and the mean diameter of normal
pollen with mean seed germination were moderate.
Percentage of pollen germination correlated signifi-
cantly with mean pollen diameter (r = 0.75), per-
centage of normal pollen (r = 0.78) and mean
diameter of normal pollen (r = 0.80).
Based on the correlation analysis, percentage of
normal pollen (r = 0.96) and mean pollen diameter
(r = 0.94) were selected as the most suitable putative
predictors of seeds per hybridisation and a simple
linear regression was performed (Fig. 2). Moreover,
the mean diameter was entered as an independent
variable (Fig. 2a) and showed a significant linear
regression (R2 = 0.88), even though the model
obtained using the percentage of normal pollen
(Fig. 2b) was the most significant (R2 = 0.93).
Therefore, both pollen parameters were chosen as
an independent variable for the validation of the
linear model.
Validation
The 18 genotypes chosen for the validation of the
linear model (Table 5) were characterised by mean
pollen diameters varying between 17.4 ± 6.4 and
30.9 ± 10.2 lm and percentages of normal pollen
varying between 4.1 and 35% (Table 5). The diam-
eters formed 3 different subsets of data (F18,3465 =
21.83, P \ 0.001) (Table 5). Analysis of the diame-
ters of the normal pollen showed that only genotype
3360 separated significantly from the other genotypes
(F18,1018 = 6.1, P \ 0.001).
Of the predictions, 83.3% were validated using
either the mean diameter or the percentage of normal
pollen as independent variable (Fig. 2a, b). Only two
predictions (genotypes 3820 and 3825) were vali-
dated by one model and excluded by the other, and in
both cases the values were very near to the prediction
interval. Therefore both parameters showed a high
percentage of validation and a rather good accuracy
as predictors of number of seeds produced per
hybridisation.
Discussion
In order to characterise the fertility of pollen donors,
this study used pollen morphology to predict the number
of seeds produced per hybridisation in tetraploid rose
genotypes. Company breeding databases have never
been studied to assess fertility using pollen charac-
teristics in roses. In previous studies to evaluate
fertility, it has been commonplace to work with a
small number of parents (Visser et al. 1977b; de Vries
and Dubois 1987; Ogilvie et al. 1991; Crespel et al.
2006). Our breeding database analysis enabled the
selection of genotypes that are accurately character-
ised by high or low fertility in terms of seeds
produced per hybridisation. Furthermore, robust
characterisation of fertility of a specific genotype
increases with the number of crossings performed on
different genotypes and in different years. However,
cross combinations using different seed parents will
Table 4 Correlations between the pollen characteristics (diam-
eter, percentage of normal pollen, diameter of normal pollen, in
vitro germination) and fertility parameters (number of seeds
per hybridisation, mean seed germination and mean seed
germination per hip) of the 11 genotypes selected from the
database
Diam.
(lm)
Normal
pollen (%)
Normal pollen
diam. (lm)
Pollen
germ. (%)
Seeds/
hyb.
Seed
germ. (%)
Seed germ
(%)/hip
Diam. (lm) 1
Normal pollen (%) 0.97** 1
Normal pollen diam. (lm) 0.96** 0.93** 1
Pollen germ. (%) 0.75** 0.78** 0.8** 1
Seeds/hyb. 0.94** 0.96** 0.87** 0.74** 1
Seed germ. (%) 0.66** 0.63* 0.64* 0.36 0.53 1
Seed germ. (%)/hip 0.75** 0.78** 0.6 0.35 0.83** 0.45 1
** Correlation (Pearson) is significant at the 0.01 level (2-tailed)
* Correlation (Pearson) is significant at the 0.05 level (2-tailed)
Euphytica (2011) 178:203–214 209
123
Author's personal copy

still bias the characterisation of the fertility of a
genotype. To reduce this bias, thresholds of exclusion
were introduced to select the genotypes. In our study
the results of the fertility analysis of the database with
an overall mean number of seeds per hybridisation of
9.13 ± 9.30 are in agreement with the number of
seeds per hybridisation reported in literature. Ogilvie
et al. (1991) hybridised 17 hybrid tea roses as seed
parents with two winter hardy cultivars used as male
parents, reporting for male parents a mean value of
3.83 and 10.83 seeds obtained per hybridisation. De
Vries and Dubois (1987) obtained 8.6 seeds per hip
(with a single pollination) hybridising two hybrid tea
roses. In agreement with the values reported for the
fertility of the low fertile genotypes, Werlemark et al.
(2009) consider 3.0–5.2 seeds per hip a low fertility
value for progenies sired from an interspecific
crossing between the pentaploid R. rubiginosa used
as seed parent and several tetraploid garden roses.
Breeding programmes with other types of roses, such
as garden roses, include a broader range of genotypes
for crossing, e.g., interploidy crosses and (re)intro-
duction of species. Despite their greater genetic
diversity, data on pollen fertility would likely be a
very valuable addition for these breeding programs
and validation experiments to confirm the utility of
pollen diameter and germination on fecundity are
warranted.
In our study, pollen grains with a diameter up to
30 lm almost always appeared shrunken with either
an irregular or spherical shape, with no furrows on
the surface-typical of aborted pollen. Pollen grains
with a diameter greater than 30 lm always looked
ellipsoidal, with furrows on the surface typical of
normal pollen. In contrast with Crespel et al. (2006)
and Zlesak (2009), who used immersed pollen, our
pollen size analysis was performed on dry pollen as
used for pollination in breeding. Immersed pollen
changes shape from ellipsoidal to spherical and
increases in size (Cruden 2009). In preliminary
experiments we observed that pollen grains imbibed
in water showed a significantly lower variance in the
diameter measurements (results not shown). Normal
pollen (diameter more than 30 lm) clearly swells to a
spherical shape, while small abnormal pollen main-
tains more or less the original shrunken and irregular
shape in most cases. Moreover, we found a significant
positive correlation between percentage of normal
pollen and mean percentage of pollen germination
(r = 0.78). Therefore, immersion in water could be a
simple test to evaluate germinability or calculate the
amount of normal pollen. This test is comparable to
what happens after pollination when rose pollen
comes in contact with the exudates of the stigma
surface: pollen grains absorb water and swell to start
the germination process (Jacob and Ferrero 2003).
Normal pollen (%)0 10 20 30 40 50 60 70 80
seed
s / h
ybrid
isat
ion
-10
0
10
20
30
High fertile genotypeLow fertile genotypeLinear regressionConfidence interval (95%)Prediction interval (95%)Validation
(b)
474.2307.0 −= xy
(a) Diameter (µm)
20 30 40
seed
s / h
ybrid
isat
ion
-10
0
10
20
30
High fertile genotypeLow fertile genotypeLinear regressionConfidence interval (95%)Prediction interval (95%)Validation
839.25184.1 −= xy
Fig. 2 Linear regression analyses of the genotypes selected
from the database. Graphs show the models created fitting the
mean diameter (a) and the percentage of normal pollen (b).
Black triangles in the graphs represent the actual fertility of the
genotypes used for the validation of the model
210 Euphytica (2011) 178:203–214
123
Author's personal copy

Several authors report that abnormal pollen does not
stain during viability tests or significantly changes
shape when immersed (Cole 1917; Erlanson 1931a;
Calvino Mameli 1951; Visser et al. 1977a, b; Crespel
et al. 2006). In our study pollen of the low fertile
genotype 2695 never germinated in vitro. However,
this genotype is able to produce offspring. Therefore,
it should be remarked that in some cases in vitro
pollen germination does not completely reflect ger-
mination capacity in vivo.
To more accurately relate the pollen characteristics
to the fertility parameters of the database, we
conformed as much as possible to the conditions of
gathering, storage and use of pollen as listed in the
company’s protocols. This created robust and high
correlations between the number of seeds per hybrid-
isation and the measured pollen diameters, the
percentage of normal pollen and the diameter of
normal pollen. In this study, mean diameter and
percentage of normal pollen are highly correlated
parameters (r = 0.97). Within the normal pollen, the
mean diameter of normal pollen grains also differed
for the high and low fertile genotypes. The more
fertile genotypes were characterised by larger pollen
grains. Low fertile genotypes produce more abnormal
pollen, which is shrunken and in some cases clearly
aborted. The high fertile genotypes showing larger
diameters had in some observations also longer
pollen tubes than the low fertile genotypes (results
not shown). Cole (1917) studied the pollen of 37
species belonging to the genus Rosa and observed,
though not determining the ploidy level of the
species, that a large amount of pollen is abortive or
sterile and has a great variability in size already
before the anthers dehiscence. Cole assumed that the
amount of abnormal pollen is correlated to a certain
degree of hybridism within these species. Calvino
Mameli (1951) studied the pollen of 48 genotypes of
roses (18 were diploid species and 30 were tetraploid
hybrid teas), also highlighting differences in the grain
Table 5 Hybridisation performances (number of crossings,
number of seed parents, and number of seeds per hybridisation)
and pollen characteristics (mean diameter, mean diameter and
percentage of normal pollen) of the genotypes used for the
validation experiment
Hybridisations Pollen
Total Normal
Genotype code Pollinations (n) Seed parents (n) Seeds/hyb. Diam. (lm) Diam. (lm) (%)
3820 15 1 2.6 17.4 ± 6.4A cB 36.9 ± 6.4 a 4.1
3823 29 2 0.7 20.8 ± 7.2 c 34.8 ± 5.6 a 4.3
3360 234 11 1.8 23.8 ± 7.1 b 34 ± 3 b 6.6
2817 289 14 1.5 21.1 ± 7.6 c 39.7 ± 7.2 a 6.8
4083 66 3 0.7 26.2 ± 6.7 b 34.3 ± 3.9 a 8.2
3518 138 8 2.1 24.1 ± 8.3 b 35.5 ± 5.2 a 9.0
4096 20 1 0.5 22.4 ± 7.3 c 34.9 ± 4.9 a 9.1
3817 116 8 0.3 24.4 ± 8.1 b 34.6 ± 3.6 a 9.9
3816 43 6 10.7 24.2 ± 8.9 b 36.7 ± 5.9 a 11.1
3741 33 3 0.1 27.3 ± 10.1 b 36 ± 5.9 a 19.7
3742 132 7 13.1 27.4 ± 10.3 a 39.7 ± 6.9 a 20.6
4092 37 2 0.3 25.3 ± 7.4 b 35.4 ± 3.6 a 21.5
4087 31 2 2.1 27.9 ± 9.3 a 36 ± 5.4 a 21.8
3826 137 9 4.9 26.2 ± 12.5 b 39 ± 7.6 a 24.5
3824 144 11 4.3 28.1 ± 10.2 a 38.7 ± 5.8 a 26.8
3740 261 11 4.1 28.6 ± 10.2 a 36.8 ± 5.3 a 26.9
4100 309 21 9.1 29 ± 9.3 a 38.2 ± 6.8 a 30.6
3825 175 15 14.3 30.9 ± 10.2 a 39.2 ± 6.3 a 35.0
A Mean ± standard deviationB Different letter indicates significant differences at the 0.05 level
Euphytica (2011) 178:203–214 211
123
Author's personal copy

sizes before the anthers dehiscence, and reported that
the length of the major axis of normal pollen grains of
hybrid tea roses, when immersed in bergamot oil
(which does not alter the pollen shape for the first 2 h
after immersion), ranges between 32.2 and 66.4 lm.
More recently, other authors related the co-existence
of sub-populations of pollen grains with higher sizes
to the occurrence of 2n pollen in a dihaploid rose
(El Mokadem et al. 2002), in three tetraploid hybrid
tea roses (Crespel and Gudin 2003) and in 10
individuals of a population of 53 diploid rose hybrids
derived from a dihaploid rose and R. wichurana
(Crespel et al. 2006). The diameter values that are
related to the occurrence of 2n pollen refer to
immersed pollen, and are not comparable with our
values that refer to dry pollen. Therefore, we cannot
compare our diameter values with those reported in
literature. No subpopulations differing in pollen size
were observed. It should be remarked that pollen size
and shape also changes in relation to the genotype
and to environmental conditions (Gudin et al. 1991a;
Crespel et al. 2006; Zlesak 2009). The evidence in the
genus Rosa of a positive correlation between pollen
grain size and ploidy level was proved by Jacob and
Pierret (2000) and by Zlesak (2009).
In our study only moderate correlations of pollen
diameter and percentage of normal pollen with mean
seed germination per hip were found. This could be
related to maternal effects on seed germination. For
example, the progeny’s germination value of male
parent 1848 was higher when sired from a high fertile
female parent (1985) than a low fertile one (1627).
Empty seeds or seeds with desiccated embryos are
found in hybrid tea roses, in which germination
capacity is also related to seed density (Gudin et al.
1992). These results are in agreement with the
general knowledge of predominant maternal influ-
ence on seed development, germination and early
seedling growth in plants (Roach and Wulff 1987;
Schmid and Dolt 1994; Thiede 1998). Our crossings
were not performed as a full diallel scheme, and
therefore the maternal effect cannot be excluded. In
addition, all the genotypes showed high standard
deviation values for mean seed germination, probably
due to the possible maternal effect as well as to the
annual environmental influences on seed germina-
tion. This also explains why the correlation between
mean seed germination and seeds per hybridisation
was not high. Nevertheless, our results clearly
indicate that high fertile male parents have a positive
influence on seed germination. Ogilvie et al. (1991)
also found that the number of seeds produced per
pollination varied according to the male parent. The
authors suggest that the use of most distantly related
cultivars in rose breeding would probably give a
higher seed set, but also suggest that there are
probably other factors that may influence pollination
success. Rose hybridisation is a part of a process of
unnatural selection, in which the application on the
surface of the stigma of very large amounts of pollen
of only one genotype could completely suppress
sexual selection among male parents. Breeders of
hybrid tea roses compensate low male parent perfor-
mances by adding more pollen during the pollination
process. De Vries and Dubois (1987) showed that the
number of seeds obtained per pollinated flower
(pollination index) did not further increase after five
pollinations between hybrid teas. Moreover, if the
ratios between the number of seeds per hip and
the number of pollinations are calculated, these
values already started to decrease as of the second
pollination.
Given the parameters that might influence pollen
diameter besides male genotype, we judged the
validation results to be good. In this study 83.3% of
the predicted seeds per hybridisation were within the
95% prediction intervals of both regression models.
The mean pollen diameter explained 87.9%
(R2 = 0.879) of the variation in seed set while the
percentage of normal pollen explained 93.2%
(R2 = 0.932). This indicates that either the mean
pollen diameter or the percentages of normal pollen
can be used as fertility predictors for hybrid tea roses.
Both parameters showed that all the roses used for the
validation are low fertile genotypes. The effective-
ness of the predictive tool for these low fertile
genotypes has been demonstrated and can success-
fully increase the efficiency of breeding programmes
with new tetraploid genotypes. Creating a prediction
of the number of seeds per hybridisation for new
genotypes by calculating the percentage of normal
pollen grains or measuring the mean pollen diameter
is reliable, but should be carefully standardised. Our
model for pollen fertility prediction is built on the
basis of hybridisations performed for 14 years at the
same location, by the same breeders and standard-
ised protocols of storage and manipulation of the
pollen. A different location, pollen manipulation and
212 Euphytica (2011) 178:203–214
123
Author's personal copy

pollination conditions could affect the effectiveness
of the model. Nevertheless, the efficiency of the
equation was validated for genotypes which were
never used in the company breeding programmes and
were therefore not present in the database. For this
reason breeders could apply the method developed on
the basis of their own database information and create
a prediction model valid for their location and their
protocols of gathering, storage and usage of pollen.
Uncomplicated pollen observations can improve the
efficiency of breeding programmes. Our proposal for
narrowing down the pool of potential male parents in
a breeding programme is to indicate the fecundity
level of a male parent by measuring percentage of
normal pollen and pollen diameter and selecting
those with the greatest likelihood of fertility. The
quality of the offspring raised from the male parent
and the prioritisation of traits will then determine
whether the male will be suitable for continued use in
the breeding programme.
Acknowledgments The authors wish to thank all the
technical staff of NIRP International, Azienda Agricola di
Ghione Luciano & Figli s.s. (Ventimiglia, Italy).
References
Calvino Mameli E (1951) Ricerche sul polline del genere Rosa.
Ann Sperim Agr 5:377–407
Cole RD (1917) Imperfection of pollen and mutability in the
genus Rosa. Bot Gaz 63:110–123
Crespel L, Gudin S (2003) Evidence for the production of
unreduced gametes by tetraploid Rosa hybrida L.
Euphytica 133:65–69. doi:10.1023/A:1025640405827
Crespel L, Mouchotte J (2003) Methods of cross-breeding. In:
Roberts AV, Debener T, Gudin S (eds) Encyclopedia of
rose science, vol 1. Elsevier Academic Press, Oxford,
pp 30–33
Crespel L, Gudin S, Meynet J, Zhang D (2002) AFLP-based
estimation of 2n gametophytic heterozygosity in two
parthenogenetically derived dihaploids of Rosa hybrida L.
Theor Appl Genet 104:451–456
Crespel L, Ricci S, Gudin S (2006) The production of 2n pollen
in rose. Euphytica 151:155–164. doi:10.1007/s10681-
006-9136-1
Cruden RW (2009) Pollen grain size, stigma depth, and style
length: the relationships revisited. Plant Syst Evol
278:223–238. doi:10.1007/s00606-008-0142-8
Darlington CD (1937) Recent advances in cytology, 2nd edn.
P. Blakinston’s son and Co, London
Darwin C (1884) The different forms of flowers on plants of
the same species, 2nd edn. J Murray, London
De Schepper S, Leus L, Mertens M, Heursel J, Van Bockstaele
E, De Loose M (2001) Flow cytometric analysis of ploidy
in Rhododendron subgenus Tsutsusi. HortScience 36:
125–127
de Vries DP, Dubois LAM (1987) The effect of temperature on
fruit set, seed set and seed germination in ‘Sonia’ 9
‘Hadley’ hybrid tea-rose crosses. Euphytica 36:117–120.
doi:10.1007/BF00730654
de Vries DP, Dubois LAM (1996) Rose breeding: past, present,
prospects. Acta Hortic 420:241–248
Debener T, Janakiram T, Mattiesch L (2000) Sports and
seedlings of rose varieties analyzed with molecular
markers. Plant Breed 119:71–74
El Mokadem H, Crespel L, Meynet J, Gudin S (2002) The
occurrence of 2n-pollen and the origin of sexual polyp-
loids in dihaploid roses (Rosa hybrida L.). Euphytica
125:169–177
Erlanson EW (1931a) Sterility in wild roses and in some
species hybrid. Genetics 16:76–96
Erlanson EW (1931b) A group of tetraploid roses in Oregon.
Bot Gaz 91:55–64
Erlanson EW (1938) Phylogeny and polyploidy in Rosa. New
Phytol 37:72–81
Gudin S (1995) Rose improvement: a breeder’s experience.
Acta Hortic 420:125–128
Gudin S, Arene L (1991) Influence of the pH of the stigmatic
exudate on male-female interaction in Rosa hybrida L.
Sex Plant Reprod 4:110–112. doi:10.1007/BF00196496
Gudin S, Arene L (1992) Putrescine increases effective polli-
nation period in roses. HortTechnology 2:211–213
Gudin S, Arene L, Bulard C (1991a) Influence of season on
pollen quality. Sex Plant Reprod 4:113–117
Gudin S, Arene L, Pellegrino C (1991b) Influence of temper-
ature and hygrometry on rose pollen germination. Adv
Hortic Sci 5:96–98
Gudin S, Arene L, Chavagnat A (1992) Relation entre
imbibition, densite, taux de remplissage et faculte ger-
minative chez l’akene de Rosa hybrida L. Agronomie
12:123–126
Jacob Y, Ferrero F (2003) Pollen grains and tubes. In: Roberts
AV, Debener T, Gudin S (eds) Encyclopedia of rose sci-
ence, vol 1. Elsevier Academic Press, Oxford, pp 518–523
Jacob Y, Pierret V (2000) Pollen size and ploidy level in the
genus Rosa. Acta Hortic 508:289–292
Krussmann G (1981) The complete book of roses. Timber
Press, Portland
Leus L (2005) Resistance breeding for powdery mildew
(Podosphaera pannosa) and black spot (Diplocarpon ro-sae) in roses. PhD Thesis, Faculty of Bioscience Engi-
neering, Ghent University, Belgium
Ogilvie I, Cloutier D, Arnold N, Jui PY (1991) The effect of
gibberellic acid on fruit and seed set in crosses of garden
and winter hardy Rosa accessions. Euphytica 52:119–123
Roach DA, Wulff RD (1987) Maternal effects in plants. Annu
Rev Ecol Syst 18:209–235. doi:10.1146/annurev.es.18.
110187.001233
Schmid B, Dolt C (1994) Effects of maternal and paternal
environment and genotype on offspring phenotype in
Solidago altissima L. Evolution 48:1525–1549
Tackholm G (1923) Zytologische Studien uber die Gattung
Rosa. Acta Hortic Berg 7:97–381
Thiede DA (1998) Maternal inheritance and its effect on
adaptive evolution: a quantitative genetic analysis of
Euphytica (2011) 178:203–214 213
123
Author's personal copy

maternal effects in a natural plant population. Evolution
52:998–1015
Visser T, de Vries DP, Scheurink JAM, Welles GWH (1977a)
Hybrid tea-rose pollen I. Germination and storage.
Euphytica 26:721–728. doi:10.1007/BF00021697
Visser T, de Vries DP, Welles GWH, Scheurink JAM
(1977b) Hybrid tea-rose pollen II. Inheritance of pollen
viability. Euphytica 26:729–732. doi:10.1007/BF0002
1698
Werlemark G, Carlson-Nisson U, Esselink GD, Nybom H
(2009) Studies of intersectional crosses between penta-
ploid dogrose species (Rosa sect. Caninae L.) as seed
parents and tetraploid garden roses as pollen donors. In:
Taxeira da Silva JA (ed) Floriculture and ornamental
biotechnology 3, special issue 1. Global Science Books,
London, pp 21–27
Zlesak DC (2006) Rose Rosa 9 hybrida. In: Anderson NO
(ed) Flower breeding and genetics: issues, challenges and
opportunities for the 21st century. Springer, Dordrecht,
pp 695–738. doi:10.1007/978-1-4020-4428-1_26
Zlesak DC (2009) Pollen diameter and guard cell length as
predictors of ploidy in diverse rose cultivars, species and
breeding lines. In: Taxeira da Silva JA (ed) Floriculture
and ornamental biotechnology 3, special issue 1. Global
Science Books, London, pp 53–70
Zlesak DC, Zuzek K, Hokanson SC (2007) Rose pollen via-
bility over time at varying storage temperatures. Acta
Hortic 751:337–343
214 Euphytica (2011) 178:203–214
123
Author's personal copy
Top Related
Copyright © 2022 FDOKUMEN

