







Bahasa
Halaman
Hukum


Om
JCMa
b
c
d
a
ARR2AA
K(CAHR
1
taboir[
bdFH3Pps
0d
Chemico-Biological Interactions 183 (2010) 369–379
Contents lists available at ScienceDirect
Chemico-Biological Interactions
journa l homepage: www.e lsev ier .com/ locate /chembio int
xidative stress induction by (+)-cordiaquinone J triggers bothitochondria-dependent apoptosis and necrosis in leukemia cells
osé Delano B. Marinho-Filhoa, Daniel P. Bezerrab, Ana J. Araújoa, Raquel C. Montenegroa,laudia Pessoaa, Jaécio C. Dinizc, Francisco A. Vianac, Otília D.L. Pessoad, Edilberto R. Silveirad,anoel O. de Moraesa, Letícia V. Costa-Lotufoa,∗
Departamento de Fisiologia e Farmacologia, Faculdade de Medicina, Universidade Federal do Ceará, Rua Cel. Nunes de Melo, 1127, 60430-270 Fortaleza, Ceará, BrazilDepartamento de Fisiologia, Centro de Ciências Biológicas e da Saúde, Universidade Federal de Sergipe, Aracaju, Sergipe, BrazilDepartamento de Química, Universidade do Estado do Rio Grande do Norte, Mossoró, Rio Grande do Norte, BrazilDepartamento de Química Orgânica e Inorgânica, Centro de Ciências, Fortaleza, Ceará, Brazil
r t i c l e i n f o
rticle history:eceived 10 August 2009eceived in revised form7 November 2009ccepted 30 November 2009vailable online 4 December 2009
a b s t r a c t
(+)-Cordiaquinone J is a 1,4-naphthoquinone isolated from the roots of Cordia leucocephala that has anti-fungal and larvicidal effects. However, the cytotoxic effects of (+)-cordiaquinone J have never beingexplored. In the present study, the effect of (+)-cordiaquinone J on tumor cells viability was investi-gated, showing IC50 values in the range of 2.7–6.6 �M in HL-60 and SF-295 cells, respectively. Studiesperformed in HL-60 leukemia cells indicated that (+)-cordiaquinone J (1.5 and 3.0 �M) reduces cell via-bility and 5-bromo-2-deoxyuridine incorporation after 24 h of incubation. (+)-Cordiaquinone J showed
eywords:+)-Cordiaquinone JytotoxicitypoptosisL-60 cells
rapid induction of apoptosis, as indicated by phosphatidylserine externalization, caspase activation, DNAfragmentation, morphologic changes, and rapid induction of necrosis, as indicated by the loss of mem-brane integrity and morphologic changes. (+)-Cordiaquinone J altered the redox potential of cells byinducing the depletion of reduced GSH intracellular content, the generation of reactive oxygen speciesand the loss of mitochondrial membrane potential. However, pre-treatment of cells with N-acetyl-l-
f theecro
eactive oxygen species cysteine abolished most oinvolving apoptosis and n
. Introduction
Quinones are biologically active compounds found widelyhroughout nature that have led to the development of manynticancer drugs, such as anthracyclines (daunorubicin, doxoru-icin), mitomycin and mitoxantrone [1,2]. The general mechanism
f quinone toxicity may be correlated with the chemical behav-ors of these compounds, such as redox cycling and conjugationeactions with bionucleophiles, which act as Michael acceptors2]. These properties may contribute to the cytotoxic mech-Abbreviations: 7-AAD, 7-amino-actinomycin; AO/EB, acridine orange/ethidiumromide; BrdU, 5-bromo-2′-deoxyuridine; DCF, dichlorofluorescein; DMSO,imethyl sulfoxide; Dox, doxorrubicin; EC50, half-maximal effective concentration;SC, forward light scatter; H2-DCF-DA, 2′ ,7′-dichlorodihydrofluorescein diacetate;TS, high-throughput screening; IC50, half-maximal inhibitory concentration; MTT,-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium; NAC, N-acetyl-l-cysteine;BMC, peripheral blood mononuclear cells; PBS, phosphate-buffered saline; PE,hycoeritin; PS, phosphatidylserine; ROS, reactive oxygen species; SSC, side lightcattering; JNK, c-Jun N-terminal kinase.∗ Corresponding author. Tel.: +55 85 3366 8255; fax: +55 85 3366 8333.
E-mail address: [email protected] (L.V. Costa-Lotufo).
009-2797/$ – see front matter © 2009 Elsevier Ireland Ltd. All rights reserved.oi:10.1016/j.cbi.2009.11.030
observed effects related to (+)-cordiaquinone J treatment, including thosesis induction.
© 2009 Elsevier Ireland Ltd. All rights reserved.
anism of action of quinoid anticancer drugs since DNA andcrucial cellular proteins are their final targets [2,3]. Alterna-tively, ROS can activate a number of signaling pathways and arealso mediators of apoptosis via mitochondria-dependent mech-anisms [4–6]. In this context, the generation of mitochondrialROS, which occurs during quinone redox cycling, modifies mem-brane permeability, resulting in mitochondrial depolarization,Bax relocalization, cytochrome c release, and caspase activation[2,4].
Cordiaquinones are meroterpenoids from plants belonging tothe genus Cordia [7,8]. To date, twelve different cordiaquinoneshave been isolated: A–D from the roots of Cordia corymbosa [9,10],E–H from the roots of Cordia linnaei [11], J and K from the roots ofCordia curassavica [12], and L and M from Cordia leucocephala [13].These molecules show activity against Cladosporium cucumerinum,Candida albicans and the larvae of the yellow fever-transmittingmosquito Aedes aegypti [11,12,8].
The present work describes the isolation of (+)-cordiaquinoneJ from the roots of C. leucocephala and the evaluation of its cyto-toxic effects against tumor cell lines. In addition, we used HL-60leukemia cells as a model to characterize the mechanism under-lying the effects of cordiaquinone and to assess the contribution
3 Biolog
oc
2
2
iastprpF
2
idddBehpata
Fabruf
70 J.D.B. Marinho-Filho et al. / Chemico-
f ROS generation and reduced glutathione (GSH) depletion to thisytotoxicity.
. Material and methods
.1. General
Optical rotations were measured on a PerkinElmer 341 dig-tal polarimeter. IR spectra (KBr pellets) were obtained on
PerkinElmer FT-IR 1000 spectrometer. High-resolution masspectra were measured on a Shimadzu LCMS-IT-TOF (ion trap-ime-of-flight hybrid mass spectrometer). NMR experiments wereerformed on a Bruker DRX-500 spectrometer. Column chromatog-aphy was done using silica gel 60 (70–230 mesh), while TLC waserformed on precoated silica gel polyester sheets (kieselgel 60254, 0.20 mm, Merck).
.2. Isolation and structural determination of (+)-cordiaquinone J
C. leucocephala was collected in May 2006 during the flower-ng stage. The plant was harvested in Mossoró County, Rio Grandeo Norte State, Brazil, and a voucher specimen (MOSS 8827) waseposited in the Herbarium Dárdano de Andrade Lima, Universi-ade Federal Rural do Semi-Árido – UFERSA, Rio Grande do Norte,razil. Air-dried and powdered roots of C. leucocephala (650 g) werextracted at room temperature with hexane, followed by a 70%
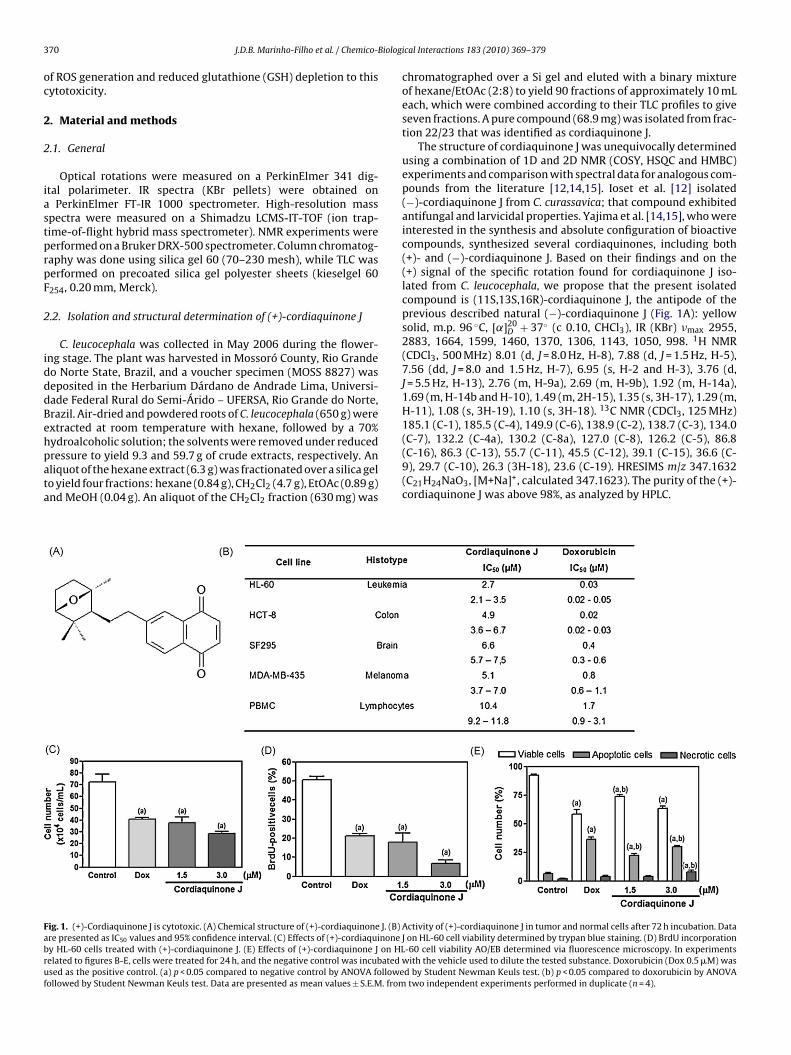
ydroalcoholic solution; the solvents were removed under reducedressure to yield 9.3 and 59.7 g of crude extracts, respectively. Anliquot of the hexane extract (6.3 g) was fractionated over a silica gelo yield four fractions: hexane (0.84 g), CH2Cl2 (4.7 g), EtOAc (0.89 g)nd MeOH (0.04 g). An aliquot of the CH2Cl2 fraction (630 mg) wasig. 1. (+)-Cordiaquinone J is cytotoxic. (A) Chemical structure of (+)-cordiaquinone J. (B)re presented as IC50 values and 95% confidence interval. (C) Effects of (+)-cordiaquinoney HL-60 cells treated with (+)-cordiaquinone J. (E) Effects of (+)-cordiaquinone J on HLelated to figures B-E, cells were treated for 24 h, and the negative control was incubatedsed as the positive control. (a) p < 0.05 compared to negative control by ANOVA followeollowed by Student Newman Keuls test. Data are presented as mean values ± S.E.M. from
ical Interactions 183 (2010) 369–379
chromatographed over a Si gel and eluted with a binary mixtureof hexane/EtOAc (2:8) to yield 90 fractions of approximately 10 mLeach, which were combined according to their TLC profiles to giveseven fractions. A pure compound (68.9 mg) was isolated from frac-tion 22/23 that was identified as cordiaquinone J.
The structure of cordiaquinone J was unequivocally determinedusing a combination of 1D and 2D NMR (COSY, HSQC and HMBC)experiments and comparison with spectral data for analogous com-pounds from the literature [12,14,15]. Ioset et al. [12] isolated(−)-cordiaquinone J from C. curassavica; that compound exhibitedantifungal and larvicidal properties. Yajima et al. [14,15], who wereinterested in the synthesis and absolute configuration of bioactivecompounds, synthesized several cordiaquinones, including both(+)- and (−)-cordiaquinone J. Based on their findings and on the(+) signal of the specific rotation found for cordiaquinone J iso-lated from C. leucocephala, we propose that the present isolatedcompound is (11S,13S,16R)-cordiaquinone J, the antipode of theprevious described natural (−)-cordiaquinone J (Fig. 1A): yellowsolid, m.p. 96 ◦C, [˛]20
D + 37◦ (c 0.10, CHCl3), IR (KBr) �max 2955,2883, 1664, 1599, 1460, 1370, 1306, 1143, 1050, 998. 1H NMR(CDCl3, 500 MHz) 8.01 (d, J = 8.0 Hz, H-8), 7.88 (d, J = 1.5 Hz, H-5),7.56 (dd, J = 8.0 and 1.5 Hz, H-7), 6.95 (s, H-2 and H-3), 3.76 (d,J = 5.5 Hz, H-13), 2.76 (m, H-9a), 2.69 (m, H-9b), 1.92 (m, H-14a),1.69 (m, H-14b and H-10), 1.49 (m, 2H-15), 1.35 (s, 3H-17), 1.29 (m,H-11), 1.08 (s, 3H-19), 1.10 (s, 3H-18). 13C NMR (CDCl3, 125 MHz)185.1 (C-1), 185.5 (C-4), 149.9 (C-6), 138.9 (C-2), 138.7 (C-3), 134.0
(C-7), 132.2 (C-4a), 130.2 (C-8a), 127.0 (C-8), 126.2 (C-5), 86.8(C-16), 86.3 (C-13), 55.7 (C-11), 45.5 (C-12), 39.1 (C-15), 36.6 (C-9), 29.7 (C-10), 26.3 (3H-18), 23.6 (C-19). HRESIMS m/z 347.1632(C21H24NaO3, [M+Na]+, calculated 347.1623). The purity of the (+)-cordiaquinone J was above 98%, as analyzed by HPLC.Activity of (+)-cordiaquinone J in tumor and normal cells after 72 h incubation. DataJ on HL-60 cell viability determined by trypan blue staining. (D) BrdU incorporation-60 cell viability AO/EB determined via fluorescence microscopy. In experiments
with the vehicle used to dilute the tested substance. Doxorubicin (Dox 0.5 �M) wasd by Student Newman Keuls test. (b) p < 0.05 compared to doxorubicin by ANOVAtwo independent experiments performed in duplicate (n = 4).

iologi
2
e1it0ttm
2
lSI1g3awwdwR2abt
2
2
H(w02wttwal2ptctata5
2
mPi2afC
J.D.B. Marinho-Filho et al. / Chemico-B
.3. Hemolytic activity
Membrane disruption was performed in 96-well plates. Briefly,ach well received 100 �l of a 0.85% NaCl solution containing0 mM CaCl2 and 100 �l of a 2% suspension of mouse erythrocytes
n the same medium. The (+)-cordiaquinone J was tested at concen-rations ranging from 12 to 500 �M. Triton X-100 (Isofar, Brazil) at.1% (in 0.85% NaCl) was used as a positive control. After incuba-ion for 60 min at room temperature, the plate was centrifuged,he supernatant was removed and the liberated hemoglobin was
easured at 540 nm (DTX-880, Beckman Coulter®) [16].
.4. Cell line and cell culture
The human cell lines used in this work were HL-60 (promye-ocytic leukemia), HCT-8 (colon), MDA-MB-435 (melanoma) andF-295 (brain), which were all obtained from the National Cancernstitute (Bethesda, MD, USA). The cells were maintained in RPMI640 medium supplemented with 10% fetal bovine serum, 2 mMlutamine, 100 U/ml penicillin, and 100 �g/ml streptomycin at7 ◦C with 5% CO2. Peripheral blood mononuclear cells (PBMC) werelso tested. Heparinized blood from healthy, non-smoking donorsho had not taken any drugs for at least 15 days prior to samplingas collected, and the PBMC were isolated via a standard method ofensity-gradient centrifugation over Ficoll–Hypaque. PBMC wereashed and re-suspended at a concentration of 3 × 105 cells/ml inPMI 1640 medium supplemented with 20% fetal bovine serum,mM glutamine, 100 U/ml penicillin, and 100 �g/ml streptomycint 37 ◦C with 5% CO2. Phytohemagglutinin (4%) was added at theeginning of the experiment. After 24 h of culture, the cells werereated with the (+)-cordiaquinone J.
.5. Proliferation assay
.5.1. Inhibition of tumor cell proliferationThe cytotoxicity of (+)-cordiaquinone J was tested against the
L-60 (promyelocytic leukemia), HCT-8 (colon), MDA-MB-435melanoma) and SF-295 (brain) cell lines. For all experiments, cellsere plated in 96-well plates (105 cells/well for adherent cells or
.5 × 105 cells/well for suspended cells in 100 �l of medium). After4 h, (+)-cordiaquinone J (0.06–77.0 �M) dissolved in 1% DMSOas added to each well using a high-throughput screening sys-
em (Biomek 3000 – Beckman Coulter, Inc. Fullerton, CA, USA), andhe cultures were incubated for 72 h. Doxorubicin (0.5 �M, Zodiac)as used as a positive control. Control groups received the same
mount of DMSO. Tumor cell growth was quantified by the ability ofiving cells to reduce the yellow dye 3-(4,5-dimethyl-2-thiazolyl)-,5-diphenyl-2H-tetrazolium bromide (MTT) to a purple formazanroduct [17]. At the end of the incubation, the plates were cen-rifuged and the medium was replaced with fresh medium (150 �l)ontaining MTT (0.5 mg/ml). Three hours later, the plates were cen-rifuged, the MTT formazan product was dissolved in 150 �l DMSO,nd the absorbance was measured using a multiplate reader (Spec-ra Count, Packard, Ontario, Canada). The drug effect was quantifieds the percentage of the control absorbance of the reduced dye at95 nm.
.5.2. Alamar blue assayTo investigate the selectivity of (+)-cordiaquinone J toward nor-
al proliferating cells, an Alamar Blue assay was performed withBMC after 72 h of drug exposure [18]. Briefly, PBMC were plated
n 96-well plates (2 × 104 cells/well in 100 �l of medium). After4 h, (+)-cordiaquinone J (0.06–77.0 �M) dissolved in 1% DMSO wasdded to each well using the HTS, and the cells were incubatedor 72 h. Doxorubicin (0.02–8.6 �M) was used as a positive control.ontrol groups received the same amount of DMSO. Twenty-fourcal Interactions 183 (2010) 369–379 371
hours before the end of the incubation, 10 �l of a stock solution(0.436 mg/ml) of Alamar Blue (resazurin; Sigma–Aldrich Co., St.Louis, MO, USA) was added to each well. The absorbance was mea-sured using a multiplate reader (DTX 880 Multimode Detector,Beckman Coulter, Inc. Fullerton, California, USA). The drug effectwas quantified as the percentages of the control absorbances at570 nm and 600 nm [18].
2.6. Analysis of the mechanisms involved in the cytotoxic activity
The following experiments were performed to elucidate themechanisms involved in the cytotoxic action of (+)-cordiaquinoneJ (1.5 and 3.0 �M) in HL-60 cells. Doxorubicin (0.5 �M) was used asa positive control.
2.6.1. Trypan blue exclusionCell viability was determined using the trypan blue dye exclu-
sion test after incubation of HL-60 cells (3 × 105 cells/ml) with(+)-cordiaquinone J. Aliquots were removed from cultures after24 h, and the cells that excluded trypan blue were counted in aNeubauer chamber.
2.6.2. Measurement of DNA synthesisHL-60 cells (3 × 105 cells/ml) were plated onto 24-well tissue
culture plates (2 ml/well) and treated with (+)-cordiaquinone J.Doxorubicin (0.5 �M) was used as a positive control. Ten micro-liters of 5-bromo-2′-deoxyuridine (BrdU, 10 mM) was added toeach well, and the cultures were incubated for 3 h at 37 ◦Cbefore completing the 24-h period of drug exposure. To assaythe amount of BrdU incorporated into the DNA, cells were har-vested, transferred to cytospin slides, and allowed to dry for 2 hat room temperature. Cells that incorporated BrdU were labeledby direct peroxidase immunocytochemistry using the chromogendiaminobenzidine. Slides were counterstained with hematoxylin,mounted, and coverslipped. Evaluation of BrdU positivity was per-formed via light microscopy (Olympus, Tokyo, Japan). Two hundredcells were counted per sample to determine the percentage of pos-itive cells.
2.6.3. Morphological analysis with May–Grunwald–Giemsastaining
Untreated and treated HL-60 cells were examined for mor-phological changes via light microscopy (Olympus, Tokyo, Japan).To evaluate cell morphology, cells were harvested, transferred tocytospin slides, fixed with methanol for 10 s and stained withMay–Grunwald–Giemsa (Bioclin, Brazil).
2.6.4. Morphological analysis with fluorescence microscopyHL-60 cells were treated with (+)-cordiaquinone J for 24 h.
Then, cells were pelleted and re-suspended in 25 �l PBS. After-wards, 1 �l of an aqueous solution of acridine orange/ethidiumbromide (AO/EB, 100 �g/ml) was added, and the cell types wereobserved under a fluorescence microscope (Olympus, Tokyo,Japan). Three hundred cells were counted per sample using thefollowing classification: viable cells, apoptotic cells and necroticcells.
2.6.5. Cell membrane integrity and viabilityHL-60 cells were incubated with the test compounds for 0.5, 3, 6
and 24 h before evaluating cell membrane integrity and cell viabil-
ity by the exclusion of propidium iodide (50 �g/ml, Sigma AldrichCo., St. Louis, MO, USA). Briefly, 100 �l of treated and untreated cellswere incubated with propidium iodide (50 �g/ml). The cells werethen incubated for 5 min. Fluorescence was measured in a GuavaEasyCyte Mine using Guava Express Plus software, and cell mor-
3 Biolog
pf
2a
E1ip1c3
2
odrta12ltcs
2
sAt5TtaAiowAol
2
pw(icswa
2
w(reDtiw
72 J.D.B. Marinho-Filho et al. / Chemico-
hology, granularity and membrane integrity [19] were analyzedor three independent experiments.
.6.6. Internucleosomal DNA fragmentation and cell cyclenalysis
DNA fragmentation was analyzed by flow cytometry (GuavaasyCyte Mine) after 0.5, 3, 6 and 24 h of incubation. Briefly,00 �l of treated and untreated cells were incubated for 30 min
n the dark with a hypotonic solution containing 50 �g/mlropidium iodide, 0.1% sodium citrate, and 0.1% Triton X-00. Fluorescence was measured and DNA fragmentation andell cycle were analyzed using ModFit LT for Win32 version.1.
.6.7. Mitochondrial transmembrane potentialMitochondrial depolarization was evaluated by incorporation
f Rhodamine 123 (Sigma Aldrich Co., St. Louis, MO, USA). Rho-amine 123 is a cell-permeable, cationic, fluorescent dye that iseadily sequestered by active mitochondria without inducing cyto-oxic effects. Briefly, cells were centrifuged at 2000 rpm for 5 minnd the pellet was re-suspended in 500 �l of 1 �g/ml of Rhodamine23 for 15 min in the dark at 37 ◦C. After incubation (0.5, 3, 6 and4 h), the cells were centrifuged at 2000 rpm for 5 min and the pel-
et was re-suspended in 500 �l of PBS and incubated for 30 min inhe dark. Fluorescence was measured and the percentage of mito-hondrial depolarization was analyzed using Guava Express Plusoftware.
.6.8. Phosphatidylserine (PS) externalizationPS externalization was analyzed by flow cytometry after PS
taining with Annexin V after 3 h of incubation [20]. A Guava Nexinssay Kit was used to determine early apoptosis. Cells were washed
wice with cold PBS and then re-suspended in 135 �l of PBS with�l of 7-amino-actinomycin D (7-AAD) and 10 �l of Annexin V–PE.he cells were gently vortexed and incubated for 20 min at roomemperature (20–25 ◦C) in the dark. Afterwards, the cells werenalyzed by flow cytometry (EasyCyte from Guava Technologies).nnexin V is a phospholipid-binding protein that has a high affin-
ty for PS. 7-AAD, a cell-impermeant dye, is used as an indicatorf membrane structural integrity. Fluorescence of Annexin V–PEas measured as yellow fluorescence at 583 nm, and that of 7-AD was measured as red fluorescence at 680 nm. The percentagesf early and late apoptotic cells and viable cells were then calcu-ated.
.6.9. Caspase 3/7 activationCaspase 3/7 activity was analyzed by flow cytometry using a cas-
ase 3/7 FAM Kit after 3 h of incubation. The cells (3 × 105 cells/ml)ere incubated with Fluorescent Labeled Inhibitor of Caspases
FLICATM) and maintained for 1 h at 37 ◦C in a CO2 incubator. Afterncubation, 80 �l of wash buffer was added and the cells wereentrifuged at 2000 rpm for 5 min. The resulting pellet was re-uspended in 200 �l of wash buffer and centrifuged again. The cellsere then re-suspended in the working solution (propidium iodide
nd wash buffer) and analyzed immediately using flow cytometry.
.6.10. Measurement of reactive oxygen species generationIntracellular reactive oxygen species (ROS) accumulation
as monitored using 2′,7′-dichlorodihydrofluorescein diacetateH2-DCF-DA), which is converted to the highly fluorescent dichlo-ofluorescein (DCF) in the presence of intracellular ROS [21]. At the
nd of the treatments (1 and 3 h), cells were loaded with 20 �M H2-CF-DA and incubated for 30 min in the dark at 37 ◦C. Cells werehen harvested, washed and re-suspended in PBS and analyzedmmediately via flow cytometry using excitation and emission
avelengths of 490 and 530 nm, respectively. �-Lapachone (2 �M)
ical Interactions 183 (2010) 369–379
and doxorubicin (0.5 �M) were used as positive controls. Addition-ally, the cells were pre-incubated with 5 mM N-acetyl-l-cysteine(NAC) before cordiaquinone treatment to evaluate the participationof ROS in the observed effects.
2.6.11. Measurement of intracellular reduced glutathione (GSH)content
At the end of treatment (15 min, 1 and 3 h), cells(7 × 106 cells/ml) were pelleted by centrifugation at 400 × gfor 5 min, re-suspended in 400 �l of lysis buffer containing 0.2%Triton X-100. The proteins were precipitated by 100 �l (0.5 M)of perchloric acid, then removed by centrifugation at 500 × g for5 min. The level of reduced GSH in the supernatant was measuredaccording to the method of Silber et al. [22]. Briefly, 200 �l ofthe cell lysate supernatant, 200 �l phosphate EDTA buffer, pH8, and 100 �l of 5,5 dithiobis-2-nitrobenzoic acid (DTNB) wereadded. DTNB oxidizes the sulfhydryl group in GSH to GSSG, withstochimetric formation of 5-thio-2-nitrobenzoic acid (TNB), whichhas a yellow color. The concentration of reduced GSH was quan-tified spectrophotometrically at 412 nm against a GSH standardcurve. �-Lapachone (2 �M) and doxorubicin (0.5 �M) were usedas positive controls. Protein concentration was determined bythe method of Bradford [23] and the results were expressed as�moles/mg protein.
2.7. Statistical analysis
Data are presented as mean ± S.E.M. The IC50 values and their95% confidence intervals (CI 95%) were obtained by non-linearregression using the GRAPHPAD program (Intuitive Software forScience, San Diego, CA). Differences between experimental groupswere compared by one-way analysis of variance (ANOVA) followedby Student Newman Keuls (p < 0.05).
3. Results
3.1. (+)-Cordiaquinone J inhibits the cell viability of humantumor cell lines and normal PBMC, but does not have non-specificlytic activity on mouse erythrocyte membranes
MTT analysis showed that (+)-cordiaquinone J exhibited cyto-toxic activity against all tumor cell lines, with IC50 values after 72 hof incubation that ranged from 2.7 to 6.6 �M (in HL-60 and SF-295cells, respectively) (Fig. 1B). For normal PBMC, the IC50 value for(+)-cordiaquinone J was 10.4 �M. Doxorubicin, the positive con-trol, presented an IC50 of 1.7 �M for PBMC, and values in tumorcell lines ranging from 0.02 to 0.8 �M (in HCT-8 and MDA-MB435,respectively) (Fig. 1B). The absence of hemolytic activity with miceerythrocytes (EC50 > 500 �M, data not shown) suggests that thecytotoxicity of (+)-cordiaquinone J is not related to non-specificmembrane damage.
Since HL-60 cells were especially sensitive to the antiprolifera-tive effect of (+)-cordiaquinone J, further studies were performedwith this cell line using concentrations corresponding to the IC50(3 �M) and IC50/2 (1.5 �M).
3.2. (+)-Cordiaquinone J displays antiproliferative effects inhuman leukemia cells with morphological features associatedwith apoptosis and necrosis induction
To characterize the cordiaquinone J-induced antiproliferative
effects in the HL-60 cell line, cell viability, BrdU incorporation andcellular morphology were evaluated after a 24-h treatment. At bothconcentrations (1.5 and 3 �M), (+)-cordiaquinone J reduced cellviability and BrdU incorporation, corroborating the MTT analysisresults (p < 0.05, Fig. 1C–E).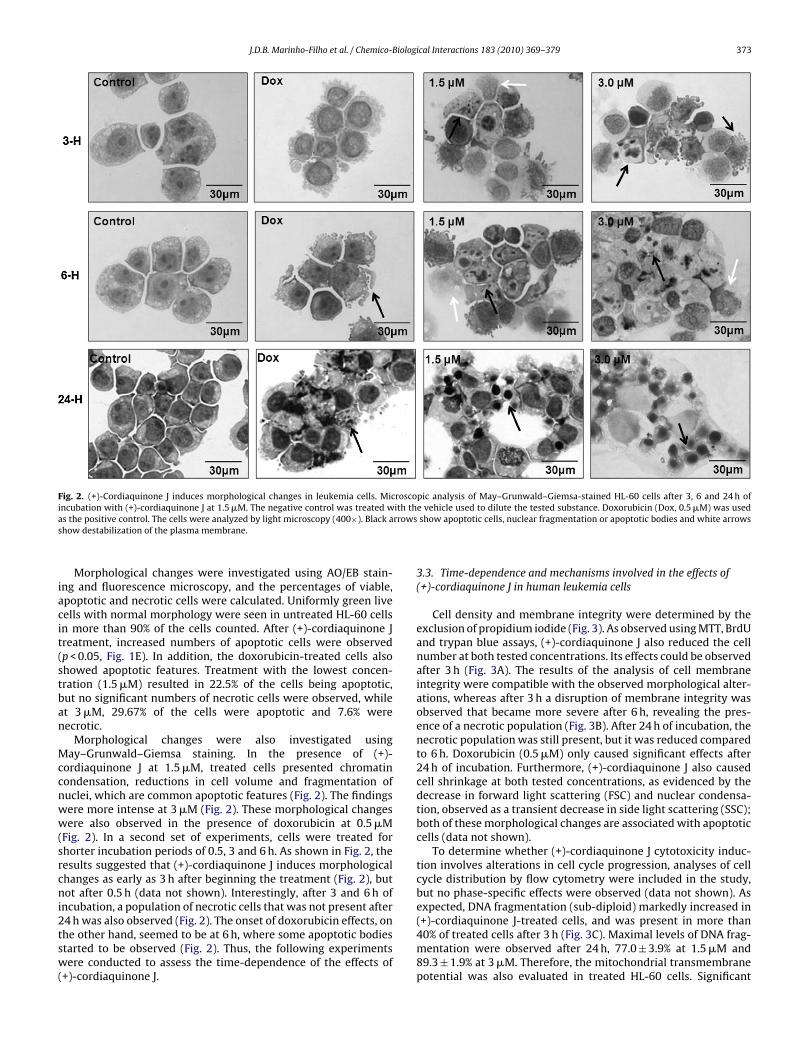
J.D.B. Marinho-Filho et al. / Chemico-Biological Interactions 183 (2010) 369–379 373
Fig. 2. (+)-Cordiaquinone J induces morphological changes in leukemia cells. Microscopic analysis of May–Grunwald–Giemsa-stained HL-60 cells after 3, 6 and 24 h ofi ith tha rrowss
iacit(stban
Mccnww(srcni2tsw(
ncubation with (+)-cordiaquinone J at 1.5 �M. The negative control was treated ws the positive control. The cells were analyzed by light microscopy (400×). Black ahow destabilization of the plasma membrane.
Morphological changes were investigated using AO/EB stain-ng and fluorescence microscopy, and the percentages of viable,poptotic and necrotic cells were calculated. Uniformly green liveells with normal morphology were seen in untreated HL-60 cellsn more than 90% of the cells counted. After (+)-cordiaquinone Jreatment, increased numbers of apoptotic cells were observedp < 0.05, Fig. 1E). In addition, the doxorubicin-treated cells alsohowed apoptotic features. Treatment with the lowest concen-ration (1.5 �M) resulted in 22.5% of the cells being apoptotic,ut no significant numbers of necrotic cells were observed, whilet 3 �M, 29.67% of the cells were apoptotic and 7.6% wereecrotic.
Morphological changes were also investigated usingay–Grunwald–Giemsa staining. In the presence of (+)-
ordiaquinone J at 1.5 �M, treated cells presented chromatinondensation, reductions in cell volume and fragmentation ofuclei, which are common apoptotic features (Fig. 2). The findingsere more intense at 3 �M (Fig. 2). These morphological changesere also observed in the presence of doxorubicin at 0.5 �M
Fig. 2). In a second set of experiments, cells were treated forhorter incubation periods of 0.5, 3 and 6 h. As shown in Fig. 2, theesults suggested that (+)-cordiaquinone J induces morphologicalhanges as early as 3 h after beginning the treatment (Fig. 2), butot after 0.5 h (data not shown). Interestingly, after 3 and 6 h of
ncubation, a population of necrotic cells that was not present after
4 h was also observed (Fig. 2). The onset of doxorubicin effects, onhe other hand, seemed to be at 6 h, where some apoptotic bodiestarted to be observed (Fig. 2). Thus, the following experimentsere conducted to assess the time-dependence of the effects of+)-cordiaquinone J.
e vehicle used to dilute the tested substance. Doxorubicin (Dox, 0.5 �M) was usedshow apoptotic cells, nuclear fragmentation or apoptotic bodies and white arrows
3.3. Time-dependence and mechanisms involved in the effects of(+)-cordiaquinone J in human leukemia cells
Cell density and membrane integrity were determined by theexclusion of propidium iodide (Fig. 3). As observed using MTT, BrdUand trypan blue assays, (+)-cordiaquinone J also reduced the cellnumber at both tested concentrations. Its effects could be observedafter 3 h (Fig. 3A). The results of the analysis of cell membraneintegrity were compatible with the observed morphological alter-ations, whereas after 3 h a disruption of membrane integrity wasobserved that became more severe after 6 h, revealing the pres-ence of a necrotic population (Fig. 3B). After 24 h of incubation, thenecrotic population was still present, but it was reduced comparedto 6 h. Doxorubicin (0.5 �M) only caused significant effects after24 h of incubation. Furthermore, (+)-cordiaquinone J also causedcell shrinkage at both tested concentrations, as evidenced by thedecrease in forward light scattering (FSC) and nuclear condensa-tion, observed as a transient decrease in side light scattering (SSC);both of these morphological changes are associated with apoptoticcells (data not shown).
To determine whether (+)-cordiaquinone J cytotoxicity induc-tion involves alterations in cell cycle progression, analyses of cellcycle distribution by flow cytometry were included in the study,but no phase-specific effects were observed (data not shown). Asexpected, DNA fragmentation (sub-diploid) markedly increased in
(+)-cordiaquinone J-treated cells, and was present in more than40% of treated cells after 3 h (Fig. 3C). Maximal levels of DNA frag-mentation were observed after 24 h, 77.0 ± 3.9% at 1.5 �M and89.3 ± 1.9% at 3 �M. Therefore, the mitochondrial transmembranepotential was also evaluated in treated HL-60 cells. Significant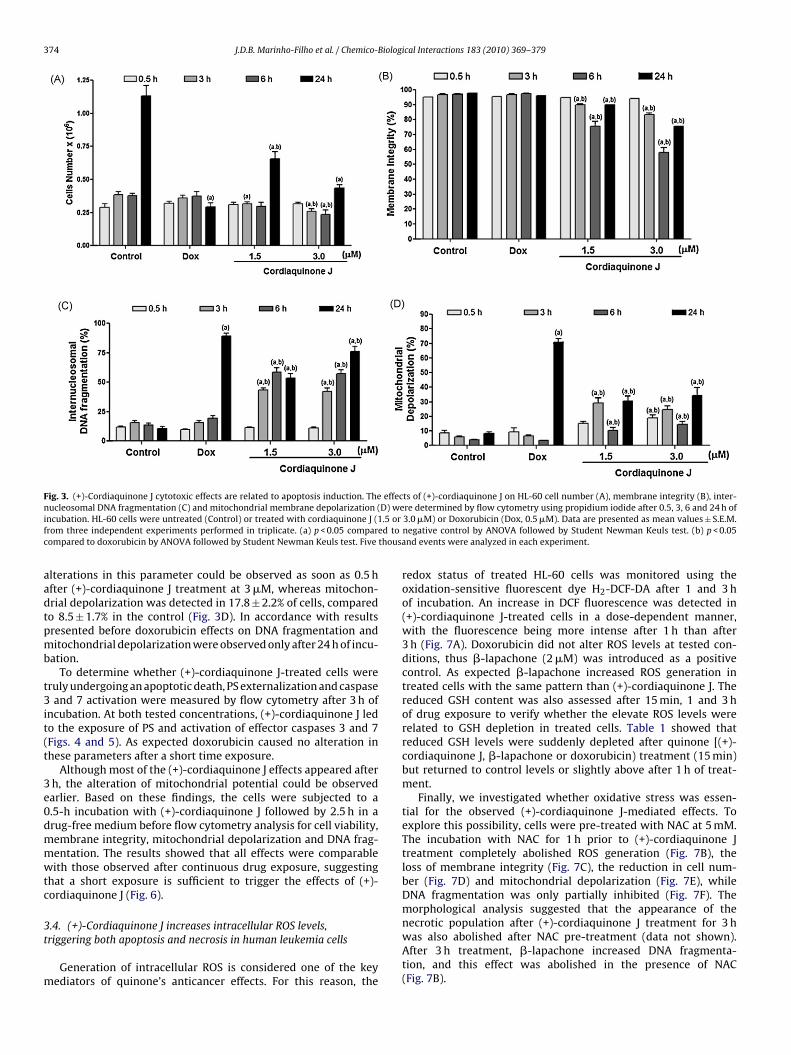
374 J.D.B. Marinho-Filho et al. / Chemico-Biological Interactions 183 (2010) 369–379
Fig. 3. (+)-Cordiaquinone J cytotoxic effects are related to apoptosis induction. The effects of (+)-cordiaquinone J on HL-60 cell number (A), membrane integrity (B), inter-n D) wei .5 orf ed toc thousa
aadtpmb
t3it(t
3e0dmmwtc
3t
m
ucleosomal DNA fragmentation (C) and mitochondrial membrane depolarization (ncubation. HL-60 cells were untreated (Control) or treated with cordiaquinone J (1rom three independent experiments performed in triplicate. (a) p < 0.05 comparompared to doxorubicin by ANOVA followed by Student Newman Keuls test. Five
lterations in this parameter could be observed as soon as 0.5 hfter (+)-cordiaquinone J treatment at 3 �M, whereas mitochon-rial depolarization was detected in 17.8 ± 2.2% of cells, comparedo 8.5 ± 1.7% in the control (Fig. 3D). In accordance with resultsresented before doxorubicin effects on DNA fragmentation anditochondrial depolarization were observed only after 24 h of incu-
ation.To determine whether (+)-cordiaquinone J-treated cells were
ruly undergoing an apoptotic death, PS externalization and caspaseand 7 activation were measured by flow cytometry after 3 h of
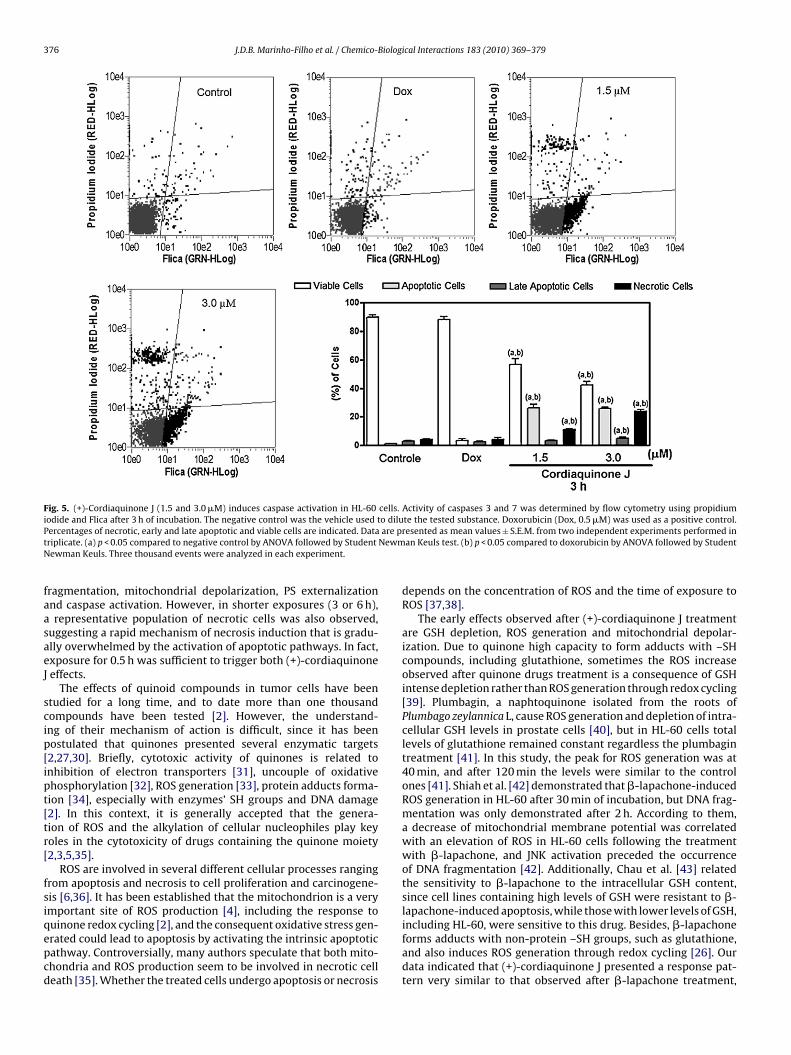
ncubation. At both tested concentrations, (+)-cordiaquinone J ledo the exposure of PS and activation of effector caspases 3 and 7Figs. 4 and 5). As expected doxorubicin caused no alteration inhese parameters after a short time exposure.
Although most of the (+)-cordiaquinone J effects appeared afterh, the alteration of mitochondrial potential could be observedarlier. Based on these findings, the cells were subjected to a.5-h incubation with (+)-cordiaquinone J followed by 2.5 h in arug-free medium before flow cytometry analysis for cell viability,embrane integrity, mitochondrial depolarization and DNA frag-entation. The results showed that all effects were comparableith those observed after continuous drug exposure, suggesting
hat a short exposure is sufficient to trigger the effects of (+)-ordiaquinone J (Fig. 6).
.4. (+)-Cordiaquinone J increases intracellular ROS levels,riggering both apoptosis and necrosis in human leukemia cells
Generation of intracellular ROS is considered one of the keyediators of quinone’s anticancer effects. For this reason, the
re determined by flow cytometry using propidium iodide after 0.5, 3, 6 and 24 h of3.0 �M) or Doxorubicin (Dox, 0.5 �M). Data are presented as mean values ± S.E.M.negative control by ANOVA followed by Student Newman Keuls test. (b) p < 0.05nd events were analyzed in each experiment.
redox status of treated HL-60 cells was monitored using theoxidation-sensitive fluorescent dye H2-DCF-DA after 1 and 3 hof incubation. An increase in DCF fluorescence was detected in(+)-cordiaquinone J-treated cells in a dose-dependent manner,with the fluorescence being more intense after 1 h than after3 h (Fig. 7A). Doxorubicin did not alter ROS levels at tested con-ditions, thus �-lapachone (2 �M) was introduced as a positivecontrol. As expected �-lapachone increased ROS generation intreated cells with the same pattern than (+)-cordiaquinone J. Thereduced GSH content was also assessed after 15 min, 1 and 3 hof drug exposure to verify whether the elevate ROS levels wererelated to GSH depletion in treated cells. Table 1 showed thatreduced GSH levels were suddenly depleted after quinone [(+)-cordiaquinone J, �-lapachone or doxorubicin) treatment (15 min)but returned to control levels or slightly above after 1 h of treat-ment.
Finally, we investigated whether oxidative stress was essen-tial for the observed (+)-cordiaquinone J-mediated effects. Toexplore this possibility, cells were pre-treated with NAC at 5 mM.The incubation with NAC for 1 h prior to (+)-cordiaquinone Jtreatment completely abolished ROS generation (Fig. 7B), theloss of membrane integrity (Fig. 7C), the reduction in cell num-ber (Fig. 7D) and mitochondrial depolarization (Fig. 7E), whileDNA fragmentation was only partially inhibited (Fig. 7F). Themorphological analysis suggested that the appearance of the
necrotic population after (+)-cordiaquinone J treatment for 3 hwas also abolished after NAC pre-treatment (data not shown).After 3 h treatment, �-lapachone increased DNA fragmenta-tion, and this effect was abolished in the presence of NAC(Fig. 7B).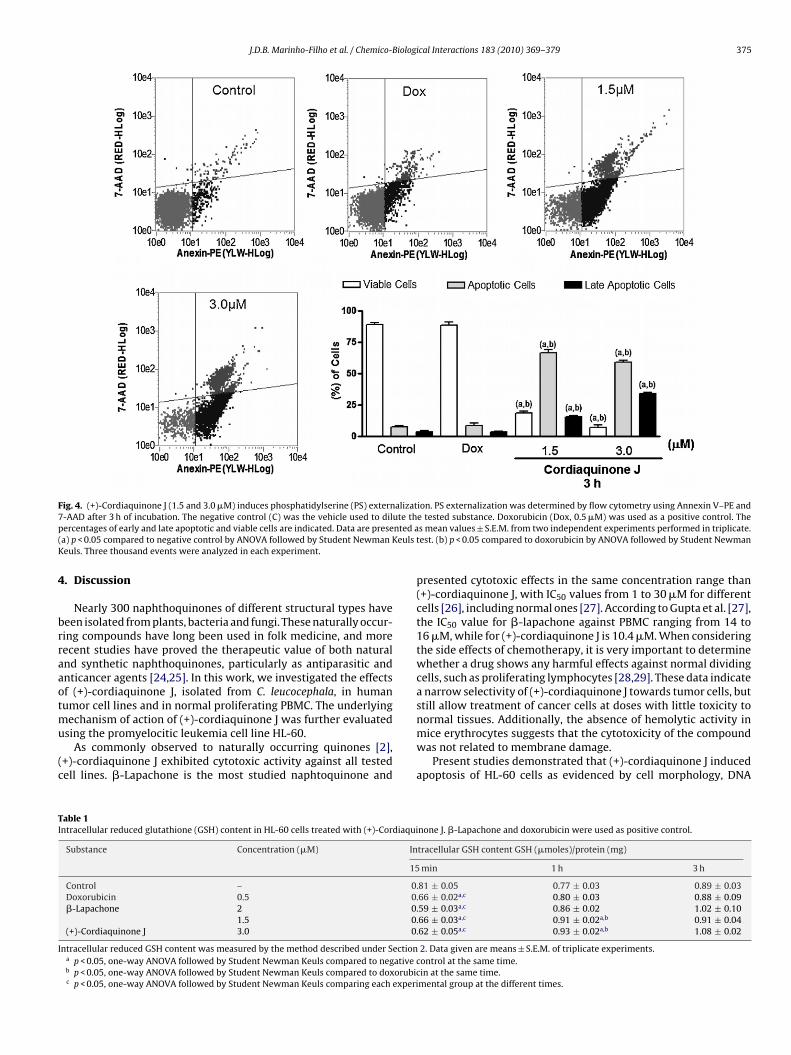
J.D.B. Marinho-Filho et al. / Chemico-Biological Interactions 183 (2010) 369–379 375
Fig. 4. (+)-Cordiaquinone J (1.5 and 3.0 �M) induces phosphatidylserine (PS) externalization. PS externalization was determined by flow cytometry using Annexin V–PE and7 ute thp nted a( euls tK
4
brraaotmu
(c
TI
I
-AAD after 3 h of incubation. The negative control (C) was the vehicle used to dilercentages of early and late apoptotic and viable cells are indicated. Data are presea) p < 0.05 compared to negative control by ANOVA followed by Student Newman Keuls. Three thousand events were analyzed in each experiment.
. Discussion
Nearly 300 naphthoquinones of different structural types haveeen isolated from plants, bacteria and fungi. These naturally occur-ing compounds have long been used in folk medicine, and moreecent studies have proved the therapeutic value of both naturalnd synthetic naphthoquinones, particularly as antiparasitic andnticancer agents [24,25]. In this work, we investigated the effectsf (+)-cordiaquinone J, isolated from C. leucocephala, in humanumor cell lines and in normal proliferating PBMC. The underlying
echanism of action of (+)-cordiaquinone J was further evaluated
sing the promyelocitic leukemia cell line HL-60.As commonly observed to naturally occurring quinones [2],+)-cordiaquinone J exhibited cytotoxic activity against all testedell lines. �-Lapachone is the most studied naphtoquinone and
able 1ntracellular reduced glutathione (GSH) content in HL-60 cells treated with (+)-Cordiaqui
Substance Concentration (�M) In
15
Control – 0Doxorubicin 0.5 0�-Lapachone 2 0
1.5 0(+)-Cordiaquinone J 3.0 0
ntracellular reduced GSH content was measured by the method described under Sectiona p < 0.05, one-way ANOVA followed by Student Newman Keuls compared to negative cb p < 0.05, one-way ANOVA followed by Student Newman Keuls compared to doxorubicc p < 0.05, one-way ANOVA followed by Student Newman Keuls comparing each experi
e tested substance. Doxorubicin (Dox, 0.5 �M) was used as a positive control. Thes mean values ± S.E.M. from two independent experiments performed in triplicate.est. (b) p < 0.05 compared to doxorubicin by ANOVA followed by Student Newman
presented cytotoxic effects in the same concentration range than(+)-cordiaquinone J, with IC50 values from 1 to 30 �M for differentcells [26], including normal ones [27]. According to Gupta et al. [27],the IC50 value for �-lapachone against PBMC ranging from 14 to16 �M, while for (+)-cordiaquinone J is 10.4 �M. When consideringthe side effects of chemotherapy, it is very important to determinewhether a drug shows any harmful effects against normal dividingcells, such as proliferating lymphocytes [28,29]. These data indicatea narrow selectivity of (+)-cordiaquinone J towards tumor cells, butstill allow treatment of cancer cells at doses with little toxicity tonormal tissues. Additionally, the absence of hemolytic activity in
mice erythrocytes suggests that the cytotoxicity of the compoundwas not related to membrane damage.Present studies demonstrated that (+)-cordiaquinone J inducedapoptosis of HL-60 cells as evidenced by cell morphology, DNA
none J. �-Lapachone and doxorubicin were used as positive control.
tracellular GSH content GSH (�moles)/protein (mg)
min 1 h 3 h
.81 ± 0.05 0.77 ± 0.03 0.89 ± 0.03
.66 ± 0.02a,c 0.80 ± 0.03 0.88 ± 0.09
.59 ± 0.03a,c 0.86 ± 0.02 1.02 ± 0.10
.66 ± 0.03a,c 0.91 ± 0.02a,b 0.91 ± 0.04
.62 ± 0.05a,c 0.93 ± 0.02a,b 1.08 ± 0.02
2. Data given are means ± S.E.M. of triplicate experiments.ontrol at the same time.in at the same time.mental group at the different times.
376 J.D.B. Marinho-Filho et al. / Chemico-Biological Interactions 183 (2010) 369–379
Fig. 5. (+)-Cordiaquinone J (1.5 and 3.0 �M) induces caspase activation in HL-60 cells. Activity of caspases 3 and 7 was determined by flow cytometry using propidiumiodide and Flica after 3 h of incubation. The negative control was the vehicle used to dilute the tested substance. Doxorubicin (Dox, 0.5 �M) was used as a positive control.P are pt NewmN
faasaeJ
scip[ipt[tr[
fsiqepcd
ercentages of necrotic, early and late apoptotic and viable cells are indicated. Datariplicate. (a) p < 0.05 compared to negative control by ANOVA followed by Studentewman Keuls. Three thousand events were analyzed in each experiment.
ragmentation, mitochondrial depolarization, PS externalizationnd caspase activation. However, in shorter exposures (3 or 6 h),representative population of necrotic cells was also observed,
uggesting a rapid mechanism of necrosis induction that is gradu-lly overwhelmed by the activation of apoptotic pathways. In fact,xposure for 0.5 h was sufficient to trigger both (+)-cordiaquinoneeffects.
The effects of quinoid compounds in tumor cells have beentudied for a long time, and to date more than one thousandompounds have been tested [2]. However, the understand-ng of their mechanism of action is difficult, since it has beenostulated that quinones presented several enzymatic targets2,27,30]. Briefly, cytotoxic activity of quinones is related tonhibition of electron transporters [31], uncouple of oxidativehosphorylation [32], ROS generation [33], protein adducts forma-ion [34], especially with enzymes’ SH groups and DNA damage2]. In this context, it is generally accepted that the genera-ion of ROS and the alkylation of cellular nucleophiles play keyoles in the cytotoxicity of drugs containing the quinone moiety2,3,5,35].
ROS are involved in several different cellular processes rangingrom apoptosis and necrosis to cell proliferation and carcinogene-is [6,36]. It has been established that the mitochondrion is a verymportant site of ROS production [4], including the response to
uinone redox cycling [2], and the consequent oxidative stress gen-rated could lead to apoptosis by activating the intrinsic apoptoticathway. Controversially, many authors speculate that both mito-hondria and ROS production seem to be involved in necrotic celleath [35]. Whether the treated cells undergo apoptosis or necrosisresented as mean values ± S.E.M. from two independent experiments performed inan Keuls test. (b) p < 0.05 compared to doxorubicin by ANOVA followed by Student
depends on the concentration of ROS and the time of exposure toROS [37,38].
The early effects observed after (+)-cordiaquinone J treatmentare GSH depletion, ROS generation and mitochondrial depolar-ization. Due to quinone high capacity to form adducts with –SHcompounds, including glutathione, sometimes the ROS increaseobserved after quinone drugs treatment is a consequence of GSHintense depletion rather than ROS generation through redox cycling[39]. Plumbagin, a naphtoquinone isolated from the roots ofPlumbago zeylannica L, cause ROS generation and depletion of intra-cellular GSH levels in prostate cells [40], but in HL-60 cells totallevels of glutathione remained constant regardless the plumbagintreatment [41]. In this study, the peak for ROS generation was at40 min, and after 120 min the levels were similar to the controlones [41]. Shiah et al. [42] demonstrated that �-lapachone-inducedROS generation in HL-60 after 30 min of incubation, but DNA frag-mentation was only demonstrated after 2 h. According to them,a decrease of mitochondrial membrane potential was correlatedwith an elevation of ROS in HL-60 cells following the treatmentwith �-lapachone, and JNK activation preceded the occurrenceof DNA fragmentation [42]. Additionally, Chau et al. [43] relatedthe sensitivity to �-lapachone to the intracellular GSH content,since cell lines containing high levels of GSH were resistant to �-lapachone-induced apoptosis, while those with lower levels of GSH,
including HL-60, were sensitive to this drug. Besides, �-lapachoneforms adducts with non-protein –SH groups, such as glutathione,and also induces ROS generation through redox cycling [26]. Ourdata indicated that (+)-cordiaquinone J presented a response pat-tern very similar to that observed after �-lapachone treatment,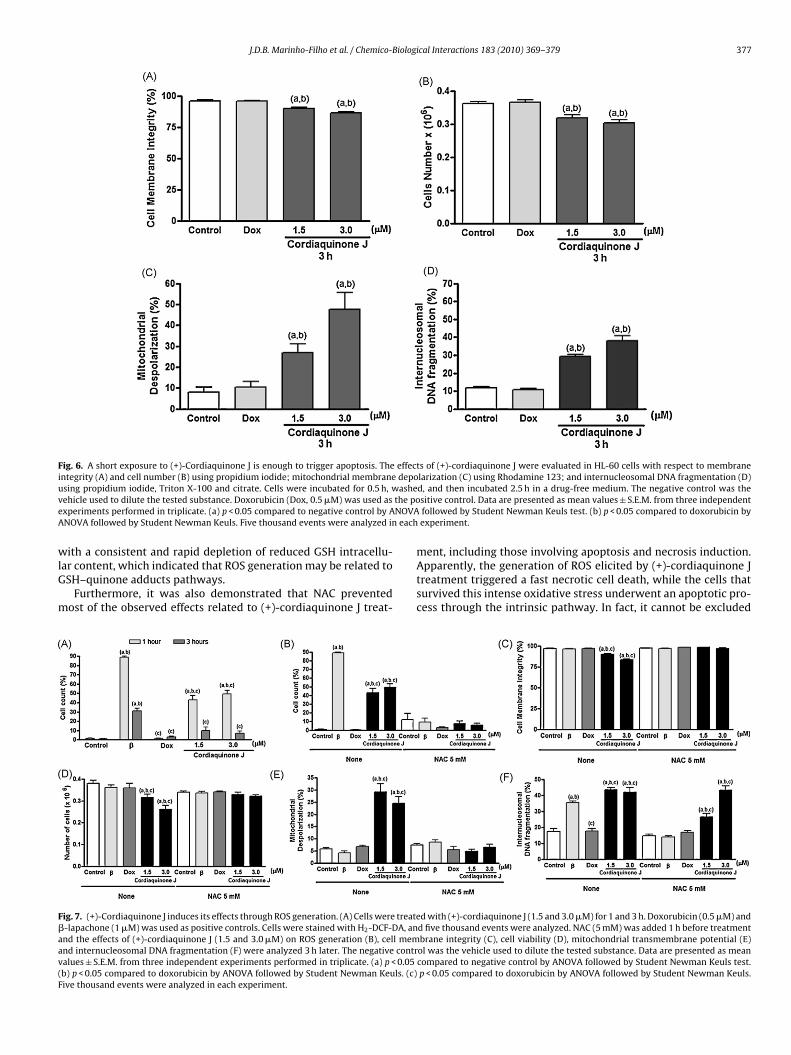
J.D.B. Marinho-Filho et al. / Chemico-Biological Interactions 183 (2010) 369–379 377
Fig. 6. A short exposure to (+)-Cordiaquinone J is enough to trigger apoptosis. The effects of (+)-cordiaquinone J were evaluated in HL-60 cells with respect to membraneintegrity (A) and cell number (B) using propidium iodide; mitochondrial membrane depolarization (C) using Rhodamine 123; and internucleosomal DNA fragmentation (D)u ashev the poe NOVAA each
wlG
m
F�aav(F
sing propidium iodide, Triton X-100 and citrate. Cells were incubated for 0.5 h, wehicle used to dilute the tested substance. Doxorubicin (Dox, 0.5 �M) was used asxperiments performed in triplicate. (a) p < 0.05 compared to negative control by ANOVA followed by Student Newman Keuls. Five thousand events were analyzed in
ith a consistent and rapid depletion of reduced GSH intracellu-ar content, which indicated that ROS generation may be related toSH–quinone adducts pathways.
Furthermore, it was also demonstrated that NAC preventedost of the observed effects related to (+)-cordiaquinone J treat-
ig. 7. (+)-Cordiaquinone J induces its effects through ROS generation. (A) Cells were treate-lapachone (1 �M) was used as positive controls. Cells were stained with H2-DCF-DA, annd the effects of (+)-cordiaquinone J (1.5 and 3.0 �M) on ROS generation (B), cell memnd internucleosomal DNA fragmentation (F) were analyzed 3 h later. The negative contralues ± S.E.M. from three independent experiments performed in triplicate. (a) p < 0.05b) p < 0.05 compared to doxorubicin by ANOVA followed by Student Newman Keuls. (c)ive thousand events were analyzed in each experiment.
d, and then incubated 2.5 h in a drug-free medium. The negative control was thesitive control. Data are presented as mean values ± S.E.M. from three independentfollowed by Student Newman Keuls test. (b) p < 0.05 compared to doxorubicin byexperiment.
ment, including those involving apoptosis and necrosis induction.Apparently, the generation of ROS elicited by (+)-cordiaquinone Jtreatment triggered a fast necrotic cell death, while the cells thatsurvived this intense oxidative stress underwent an apoptotic pro-cess through the intrinsic pathway. In fact, it cannot be excluded
d with (+)-cordiaquinone J (1.5 and 3.0 �M) for 1 and 3 h. Doxorubicin (0.5 �M) andd five thousand events were analyzed. NAC (5 mM) was added 1 h before treatmentbrane integrity (C), cell viability (D), mitochondrial transmembrane potential (E)ol was the vehicle used to dilute the tested substance. Data are presented as meancompared to negative control by ANOVA followed by Student Newman Keuls test.p < 0.05 compared to doxorubicin by ANOVA followed by Student Newman Keuls.

3 Biolog
taq
clsJDsnbs
(Rbiaet
A
BaIuae6
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[[
[
[
[
[
78 J.D.B. Marinho-Filho et al. / Chemico-
hat NAC might forms nucleophilic monoaddition or diadditiondducts with quinones, which also possible contribute to reduceuinone toxicity [44,45].
The presence of DNA fragmentation at the highest testedoncentration (3.0 �M), even in the absence of mitochondrial depo-arization, ROS generation and loss of membrane integrity, coulduggest that despite mediation of the effects of (+)-cordiaquinoneby oxidative stress, the compound may have direct effects onNA, as has been observed with other quinoid antitumor agents
uch as anthracyclines [2,35]. Other possible mechanism of actionot based on ROS generation, such as sulphydryl arylation ofiomolecules and topoisomerase inhibition, also need to be con-idered in this regard.
In conclusion, the results presented herein demonstrate that+)-cordiaquinone J presents antiproliferative effects related toOS generation. Moreover, the resulting oxidative stress inducesoth necrosis and apoptosis in tumor cells. Further studies are
n progress to evaluate whether (+)-cordiaquinone J also inter-cts directly with DNA and whether its electrochemical propertiesxplain the additional mechanisms involved in the cytotoxicity ofhis compound.
cknowledgments
We wish to thank CNPq, Instituto Claude Bernard, FUNCAP,NB/FUNDECI, and FINEP for financial support in the form of grantsnd fellowship awards. The authors also thank the National Cancernstitute (Bethesda, MD, USA) for donation of the tumor cell linessed in this study. The authors thank Silvana Franca dos Santosnd Rogéria Montenegro for technical assistance. The English wasdited by American Journal Experts (key # 0774-93A6-AD85-C591-C9E).
eferences
[1] W.O. Foye (Ed.), Cancer Chemotherapeutic Agents, American Chemical Society,Washington, DC, 1995.
[2] C. Asche, Antitumor quinones, Mini Rev. Med. Chem. 5 (2005) 449–467.[3] A. Kawiak, J. Piosik, G. Stasilojc, A. Gwizdek-Wisniewska, L. Marczak, M. Sto-
biecki, J. Bigda, E. Lojkowska, Induction of apoptosis by plumbagin throughreactive oxygen species-mediated inhibition of topoisomerase II, Toxicol. Appl.Pharmacol. 223 (2007) 267–276.
[4] C. Fleury, B. Mignotte, J.L. Vayssière, Mitochondrial reactive oxygen species incell death signaling, Biochimie 84 (2002) 131–141.
[5] D. Bironaite, A.V. Kalvelyte, A. Imbrasaite, A. Stulpinas, The intracellular antiox-idant balance of HL-60 cells and its implication in the apoptosis induced byquinoidal compounds, Biologija 1 (2004) 48–51.
[6] J.M. Matés, J.A. Segura, F.J. Alonso, J. Márquez, Intracellular redox status andoxidative stress: implications for cell proliferation, apoptosis, and carcinogen-esis, Arch. Toxicol. 82 (2008) 273–299.
[7] A.Y.F. Saitou, M. Sekimoto, S. Maetoko, G. Yabuta, Synthesis and absolute con-figuration of cordiaquinone K, antifungal and larvicidal meroterpenoid isolatedfrom the Panamanian plant Cordia curassavica, Tetrahedron Lett. 44 (2003)6915–6918.
[8] E. Arkoudis, M. Stratakis, Synthesis of cordiaquinones B, C, J, and K on the basisof a bioinspired approach and the revision of the relative stereochemistry ofcordiaquinone C, J. Org. Chem. 73 (2008) 4484–4490.
[9] L.W. Bieber, I. Messana, S.C.N. Lins, A.A. da Silva Filho, A.A. Chiappeta, J.F. deMello, Meroterpenoid naphthoquinones from Cordia corymbosa, Phytochem-istry 29 (1990) 1955–1959.
10] L.W. Bieber, H.C. Krebs, W. Schafer, Further meroterpenoid naphthoquinonesfrom Cordia corymbosa, Phytochemistry 35 (1994) 1027–1028.
11] J.R. Ioset, A. Marston, M.P. Gupta, K. Hostettmann, Antifungal and larvicidalmeroterpenoid naphthoquinones and a naphthoxirene from the roots of Cordialinnaei, Phytochemistry 47 (1998) 729–734.
12] J.R. Ioset, A. Marston, M.P. Gupta, K. Hostettmann, Antifungal and larvicidalcordiaquinones from the roots of Cordia curassavica, Phytochemistry 53 (2000)613–617.
13] J.C. Diniz, F.A. Viana, O.F. Oliveira, M.A. Maciel, M.C. Torres, R. Braz-Filho, E.R.
Silveira, O.D. Pessoa, 1H and 13C NMR assignments for two new cordiaquinonesfrom roots of Cordia leucocephala, Magn. Reson. Chem. 47 (2009) 190–193.14] A. Yajima, F. Saitou, M. Sekimoto, S. Maetoko, G. Yabuta, Synthesis and abso-lute configuration of cordiaquinone K, antifungal and larvicidal meroterpenoidisolated from the Panamanian plant Cordia curassavica, Tetrahedron Lett. 44(2003) 6915–6918.
[
ical Interactions 183 (2010) 369–379
15] A. Yajima, F. Saitou, M. Sekimoto, S. Maetoko, T. Nukada, G. Yabuta, Synthesisof cordiaquinone J and K via B-alkyl Suzuki–Miyaura coupling as a key step anddetermination of the absolute configuration of natural products, TetrahedronLett. 61 (2005) 9164–9172.
16] P.C. Jimenez, S.C. Fortier, T.M.C. Lotufo, C. Pessoa, M.E.A. Moraes, M.O. Moraes,L.V. Costa-Lotufo, Biological activity in extract of ascidians (Tunicata, Ascidi-acea) from northeastern Brazilian coast, J. Exp. Mar. Biol. Ecol. 287 (2003)93–101.
17] T.J. Mosmann, Rapid colorimetric assay for cellular growth and survivor: appli-cation to proliferation and cytotoxicity assays, J. Immunol. Methods 65 (1983)55–63.
18] S.A. Ahmed, R.M. Gogal, J.E. Walsh, A new rapid and simple non-radioactiveassay to monitor and determine the proliferation of lymphocytes an alterna-tive to [3H]thymidine incorporation assay, J. Immunol. Methods 170 (1994)211–224.
19] Z. Darzynkiewicz, S. Bruno, G. Del Bino, W. Gorczyca, M.A. Hotz, P. Lassota, F.Traganos, Features of apoptotic cells measured by flow cytometry, Cytometry13 (1992) 795–808.
20] I. Vermes, C. Haanen, H. Steffens-Nakken, C. Reutelingsperger, A novel assayfor apoptosis. Flow cytometric detection of phosphatidylserine expression onearly apoptotic cells using fluorescein labelled Annexin V, J. Immunol. Methods184 (1995) 39–51.
21] C.P. LeBel, H. Ischiropoulos, S.C. Bondy, Evaluation of the probe2′ ,7′dichlorofluorescin as an indicator of reactive oxygen species formationand oxidative stress, Chem. Res. Toxicol. 5 (1992) 227–231.
22] R. Silber, M.C. Farber, E. Papdopaulos, Glutathione depletion in chronic lym-phocytic leukemia B lymphocytes, Blood 80 (1992) 2038–2043.
23] M.M. Bradford, A rapid and sensitive method for the quantification of micro-gram quantities of protein utilizing the principle of protein–dye binding, Anal.Biochem. 72 (1976) 248–254.
24] R. Munday, B.L. Smith, C.M. Munday, Comparative toxicity of 2-hydroxy-3-alkyl-1,4-naphthoquinones in rats, Chem. Biol. Interact. 98 (1995) 185–192.
25] V.K. Tandon, R.B. Chhor, R.V. Singh, S. Rai, D.B. Yadav, Design, synthesis and eval-uation of novel 1,4-naphthoquinone derivatives as antifungal and anticanceragents, Bioorg. Med. Chem. Lett. 14 (2004) 1079–1083.
26] A.B. Pardee, Y.Z. Li, C.J. Li, Cancer therapy with beta-lapachone, Curr. CancerDrug Targets 2 (2002) 227–242.
27] D. Gupta, K. Podar, Y.T. Tai, B. Lin, T. Hideshima, M. Akiyama, R. LeBlanc, L.Catley, N. Mitsiades, C. Mitsiades, D. Chauhan, N.C. Munshi, K.C. Anderson,�-Lapachone, a novel plant product, overcomes drug resistance in human mul-tiple myeloma cells, Exp. Hematol. 30 (2002) 711–720.
28] V. Zuco, R. Supino, S.C. Righetti, L. Cleris, E. Marchesi, C. Passerine-Gambacorti,F. Formelli, Selective cytotoxicity of betulinic acid on tumor cell lines, but noton normal cells, Cancer Lett. 175 (2002) 17–25.
29] M.C. Anazetti, P.S. Melo, N. Duran, M. Haun, Comparative cytotoxicity ofdimethylamide-crotonin in the promyelocytic leukemia cell line (HL-60) andhuman peripheral blood mononuclear cells, Toxicology 188 (2003) 261–274.
30] H.Q. Wu, Z.S. Huang, X.Z. Bu, Y.D. Shen, Z.L. Zhang, B.F. Xie, Z.C. Liu, L.Q. Gu,A.S.C. Chan, The molecular mechanisms involved in the cytotoxicity of alkanninderivatives, Eur. J. Med. Chem. 40 (2005) 1341–1345.
31] J.L. Vennerstrom, J.W. Eaton, Oxidants, oxidant drugs and malaria, J. Med. Chem.31 (1988) 1269–1277.
32] J.L. Howland, Phosphorylation coupled to the oxidation of tetramethyl-p1-phenylenediamine in rat-liver mitochondria, Biochim. Biophys. Acta 77 (1963)419–429.
33] T.J. Monks, P. Hanzlik, G.M. Cohen, D. Ross, D.G. Graham, Quinones chemistryand toxicity, Toxicol. Appl. Pharmacol. 112 (1992) 2–16.
34] H.E. Kleiner, M.I. Rivera, N.R. Pumford, T.J. Monks, S.S. Lau, Immunochemicaldetection of quinol-thioether-derived protein adducts, Chem. Res. Toxicol. 11(1998) 1282–1290.
35] E.A. Hillard, F.C. Abreu, C.C.M. Ferreira, G. Jaouen, M.O.F. Goulart, C. Ama-tore, Electrochemical parameters and techniques in drug development, withan emphasis on quinone and related compounds, Chem. Commun. (2008)2612–2628.
36] J. Wang, J. Yi, Cancer cell killing via ROS, Cancer Biol. Ther. 7 (2008) 1875–1884.37] T. Ishikawa, J.J. Bao, Y. Yamane, K. Acimaru, K. Fridrich, C.D. Wright, M.T.
Kuo, Coordinated induction of MRP/GS-X pump and �-glutamylcysteine syn-thetase by heavy metals in human leukemia cells, J. Bio. Chem. 271 (1996)14981–14988.
38] G. Bartosz, Reactive oxygen species: destroyers or messengers? Biochem. Phar-macol. 77 (2009) 1303–1305.
39] C.C.C. Wang, Y.M. Chiang, S.C. Sung, Y.L. Hsu, J.K. Chang, P.L. Kuo, Plumbagininduces cell cycle arrest and apoptosis through reactive oxygen species/c-JunN-terminal kinase pathways in human melanoma A375.S2 cells, Cancer Lett.259 (2008) 82–98.
40] A.A. Powolny, S.V. Singh, Plumbagin-induced apoptosis in human prostate can-cer cells is associated with modulation of cellular redox status and generationof reactive oxygen species, Pharm. Res. 25 (2008) 2171–2180.
41] A. Kawiak, J. Piosik, G. Stasilojc, A. Gwizdek-Wisniewska, L. Marczak, M. Sto-biecki, J. Bigda, E. Lojkowska, Induction of apoptosis by plumbagin through
reactive oxygen species-mediated inhibition of topoisomerase II, Toxicol.Applied Pharmacol. 223 (2007) 267–276.42] S.G. Shiah, S.E. Chuang, Y.P. Chau, S.C. Shen, M.L. Kuo, Activation of c-Jun NH2-terminal kinase and subsequent CPP32/Yama during topoisomerase inhibitor�-lapachone-induced apoptosis through an oxidation-dependent pathway,Cancer Res. 59 (1999) 391–398.

iologi
[
[
J.D.B. Marinho-Filho et al. / Chemico-B
43] Y.P. Chau, S.G. Shiah, M.J. Don, M.L. Kuo, Involvement of hydrogen peroxide intopoisomerase inhibitor �-lapachone-induced apoptosis and differentiation inhuman leukemia cells, Free Rad. Biol. Med. 24 (1998) 660–670.
44] M.B. Bagh, A.K. Maiti, S. Jana, K. Banerjee, A. Roy, S. Chakrabarti, Quinone andoxyradical scavenging properties of N-acetylcysteine prevent dopamine medi-ated inhibition of Na+, K+-ATPase and mitochondrial electron transport chain
[
cal Interactions 183 (2010) 369–379 379
activity in rat brain: implications in the neuroprotective therapy of Parkinson’sdisease, Free Radic. Res. 42 (2008) 574–581.
45] C.P. Fall, J.P. Bennett Jr., Characterization and time course of MPP1-inducedapoptosis in human SH-SY5Y neuroblastoma cells, J. Neurosci. Res. 55 (1999)620–628.
Top Related
Copyright © 2022 FDOKUMEN

