







Bahasa
Halaman
Hukum


RESEARCH ARTICLE
Morphology and Histology of the Tongue and Oral Chamberof Eublepharis macularius (Squamata: Gekkonidae), with SpecialReference to the Foretongue and its Role in Fluid Uptakeand Transport
Heather A. Jamniczky • Anthony P. Russell •
Megan K. Johnson • Stephane J. Montuelle •
Vincent L. Bels
Received: 16 October 2009 / Accepted: 22 October 2009
� Springer Science+Business Media, LLC 2009
Abstract Detailed descriptions of tongue morphology of
members of Squamata that refer to functional implications
other than food processing are rare. Herein we focus on the
morphology of the dorsal epithelium and internal structure
of the tongue of the Leopard Gecko, Eublepharis macu-
larius, emphasizing the foretongue and its relation to fluid
uptake. We employ both scanning electron microscopy and
serial histology to examine the morphology of the entire
tongue, its component regions, and its situation in the oral
chamber. We recognize five distinct morphological regions
of the dorsal tongue surface, each of which is distinctive
both morphologically and histologically. The foretongue
bears papillae quite different in structure and spacing from
those of all other tongue regions, and these non-glandular
structures are involved in gathering and transporting fluid
from the environment. Fluid unloaded from the foretongue
in the region of the vomeronasal sinus is channeled through
the network of cuboidal papillae and directed towards a
pair of compartments lateral to the tongue in which fluid
pools during a drinking bout. This allows the dorsal surface
of the mid- and hind-tongue, which are involved in food
processing and manipulation, to be largely segregated from
the pathway of fluid flow. We relate our findings to
descriptions of the tongue of other taxa, and propose
functional hypotheses for the observed morphology. This
study provides new anatomical information upon which
future studies of the functional morphology of the buccal
apparatus in the Gekkota can be based.
Keywords Tongue � Epithelium � Histology �Eublepharis macularius � Gekkonidae � Drinking
Introduction
The morphology of the tongue of members of Squamata is
known from a relatively large number of studies (e.g. Seiler
1891, 1892; de Rooij 1915; Camp 1923; Gnanamuthu
1937; Iwasaki and Miyata 1985; Iwasaki et al. 1985;
Schwenk 1985, 1986, 2000; Rabinowitz and Tandler 1986;
Smith 1986, 1988; Iwasaki 1990, 2002; Delheusy et al.
1994; Toubeau and Bels 1994; Herrel et al. 2005). Inves-
tigations of the gross anatomy of the squamate tongue,
supplemented by light and electron microscopy, across a
broad spectrum of subordinate taxa, have demonstrated
considerable morphological variation in the structure of the
dorsal lingual epithelium. Much attention has been paid to
describing the features of the tongue surface, and the
phylogenetic implications of these features have been
extensively discussed (e.g. Schwenk 1985; Smith 1988).
Several authors have also offered functional explanations
for the morphological diversity evident in squamate tongue
epithelia (e.g. Beisser et al. 1998, 2004; Schwenk 2000;
Iwasaki 2002; Herrel et al. 2005), mostly in relation to
adaptations for feeding in a terrestrial environment.
H. A. Jamniczky (&) � A. P. Russell � M. K. Johnson
Vertebrate Morphology and Palaeontology Research Group,
Department of Biological Sciences, University of Calgary,
2500 University Drive NW, Calgary, AB T2N 1N4, Canada
e-mail: [email protected]
Present Address:H. A. Jamniczky
Department of Cell Biology and Anatomy, Faculty of Medicine,
University of Calgary, 3330 Hospital Drive NW, Calgary,
AB T2N 4N1, Canada
S. J. Montuelle � V. L. Bels
Museum National d’Histoire Naturelle, USM 302, 75005 Paris,
France
123
Evol Biol
DOI 10.1007/s11692-009-9072-9

As part of a larger project (Bels et al. unpublished)
investigating the biomechanics of oral processing in the
Leopard Gecko (Eublepharis macularius), we undertook a
scanning electron and light microscopical investigation of
the morphology of the tongue of E. macularius. We here
document its morphology in the context of drinking
behavior, in order to correlate the features observed with
their function in fluid collection and transport.
Water is a vital metabolic resource and it is anticipated
that the tongue, in those taxa that use this structure in the
drinking process, will be configured in such a way as to
promote effective and efficient transfer of water from the
environment to the oral chamber. Sherbrooke et al. (2007)
demonstrate this for the jaw angle region of the rain-har-
vesting Phrynosoma cornutum, implicating tongue, jaw,
and hypobranchial movements in the transfer of water from
the integument to the oral cavity. Furthermore, the oral
chamber acts as a conduit for this water between the tongue
and the glottis, and its structure is likely to be reflective of
the passage of water through it. In lizards such as gekko-
tans in which food prehension is jaw- rather than tongue-
based (Delheusy and Bels 1999; Schwenk 2000; Bels 2003;
Reilly and McBrayer 2007) we hypothesize that the tongue
is regionally specialized to accommodate both fluid and
food transport and processing.
To test this prediction, we investigate the structure of the
tongue in the Leopard gecko to determine whether func-
tional subdivision of the tongue, in relation to drinking and
food processing, is evident. We compare our results to
those that exist for other members of Gekkota (Gekko
gecko [pers. obs.], Gekko japonicus [Iwasaki 1990], Gon-
atodes antillensis, and Coleonyx variegatus [Schwenk
1985]), and suggest future research directions aimed at
furthering our understanding of the form and function of
the gekkotan and squamate buccal apparatus.
Methods
Three adult female (SVL 131–136 mm), one juvenile
female (SVL 70 mm), and one hatchling (SVL 53 mm)
female Eublepharis macularius were sacrificed by oral or
intracardiac administration of T-61 Euthanasia Solution
(Intervet Canada), according to a protocol approved by the
University of Calgary Animal Care Committee (protocol
#BI2005-22), following Canada Council on Animal Care
guidelines. Preliminary observations revealed that the
tongues of freshly sacrificed adult individuals were pro-
fusely coated with mucus, which remains in place after
fixation. To eliminate this problem in our experimental
sample so as to enable scanning electron microscopic
(SEM) visualization of tongue structure, we pretreated the
tongue as follows: the lower jaw and associated tongue
were excised from each individual immediately following
sacrifice, affixed with fine entomological pins to a paraffin
wax-lined, small, sealable plastic container and flooded
with pharmacological decongestant nasal spray (active
ingredient 0.1% xylometazoline hydrochloride). This was
allowed to sit on the tongue surface for several minutes,
and was then washed off by irrigating the tongue vigor-
ously with distilled water from a plastic wash bottle. Fol-
lowing several rounds of aqueous irrigation, the tongues
were fixed by immersion in 10% neutral buffered formalin
for 24 h, and stored in 70% ethanol.
Macroscopic structure of the tongue of E. macularius
was examined via macrophotography, photomicrography,
and SEM. For SEM observations, two adult tongues were
examined whole and one was sliced longitudinally prior to
examination. These were dehydrated using a graded etha-
nol series from 70% to 100%, critical point dried in a
Seevac CO2 Critical Point Dryer, mounted on � inch Zeiss
aluminum SEM mounts using colloidal silver paste, and
sputter coated with gold using a Hummer II Sputter Coater.
They were examined in the standard high vacuum mode of
a Philips/FEI ESEM XL30, and images were taken at
magnifications of 1259 and 5009 for each of the different
regions of the tongue surface. The tongue of the juvenile
female was photographed in situ after fixation and removal
of the lower jaw from the remainder of the head. Its tip was
subsequently removed and subjected to SEM.
Microscopic structure of the tongue of E. macularius
was visualized using serial histology. Adult tongues were
sliced sagittally, and each half was embedded separately in
paraffin wax following standard technique (Humason
1979) and oriented either for orthogonal or longitudinal
sectioning. Serial sections were cut at 10 lm and stained
with Masson’s Trichrome (Witten and Hall 2003, substi-
tuting Gill’s Haematoxylin for Mayer’s Haematoxylin).
Selected sections were photographed using a Nikon DS-L1
digital camera mounted on a Nikon 50i compound micro-
scope, integrated using a Nikon DS-5M camera control
unit. Images shown were not subjected to any post-capture
processing.
The tongue of the hatchling individual was examined in
situ by sectioning the entire head in the transverse plane.
This enabled the establishment of both fine structure and
the placement of the tongue in relation to other oral fea-
tures. Staining procedures were identical to those for the
sectioned adult tongues (see above).
Measurements of tongue structures were taken from the
histological longitudinal and cross sections of the adult
tongues. Slides were examined using the above-mentioned
microscope system. Measurements were taken directly
from the camera control unit using the measurement tool
calibrated to the appropriate magnification. Measurements
of papilla height, width, and spacing were taken from both
Evol Biol
123

longitudinal sections and cross sections of the tongues for
each of the five structural regions identified. Since the
absolute size of the structures may be influenced by the size
of the animal, the measurements were converted into two
ratios: height:width and spacing:width.
The relatively small body size of E. macularius makes it
unsuitable for vascular study using conventional vascular
infiltration techniques. Instead, a specimen of Gekko gecko,
injected with red Microfil (FlowTech Inc, Carver, MA) for
another study (Russell et al. 1988), was examined com-
paratively in relation to vascular structure, and is figured
herein. Complete details on preparation of this specimen
are available elsewhere (Russell et al. 1988).
Three adult male specimens of E. macularius were
filmed drinking in lateral, dorsal, and frontal views using
an NAC HSV-500 C3 high-speed video system at 250 Hz.
Lizards were filmed under two conditions: (i) water
delivered on the substrate in front of the head (sometimes
labeled with barium) and (ii) water drops sprayed on a
sheet of glass arranged vertically in front of them. Lizards
were deprived of water for 1 week before filming, but were
fed crickets and mealworms ad libitum during this period.
During filming, lizards were placed in a small plexiglas
corridor and water was provided using a pipette. Record-
ings were transmitted to a computer for analysis of tongue
movement and deformation during each drinking cycle.
Quantitative analysis of the mechanics of drinking will be
presented elsewhere (Bels et al. in preparation).
Results
Anatomy
Schwenk (1985: Fig. 1) divided the lepidosaur tongue into
three regions: the tip, which consists of the bifurcated
anterior portion and a small area immediately posterior to
the bifurcated portion; and the remainder of the tongue,
which was subdivided into fore- and hindtongue by a line
placed halfway between the tip and the posterior end of the
tongue. Herein, we divide the tongue on the basis of ana-
tomical features rather than proportional dimensions, and
demonstrate that there are five distinctive morphological
regions on the dorsum of the tongue of E. macularius
(Fig. 1), four of which exhibit morphologically distinct
types of papillae on the dorsal surface. Papillae in each
region are differentiable on the basis of height, width, and
the density of their occurrence (Table 1). We designate
these regions (Fig. 1b) as follows: (1) extreme tongue tip
(no papillae); (2) tongue edges (feather-like papillae); (3)
foretongue (cuboidal papillae); (4) midtongue (leaf-like
papillae); and (5) hindtongue (two subregions, containing
long fungiform (5a) and short fungiform (5b) papillae
respectively). Each region is differently involved in oral
transport and is discussed separately below. Figure 1b
maps these regions and provides orientation and location
(Fig. 1a) information relating to each of the subsequent
figures.
The extreme tongue tip (Plate 1A–C) is bifid, covered
by keratinized squamous epithelium, and bears no papillae
(Plate 1A, B). Its thickened epidermal covering is smooth
and projects ventrally (Plate 1A) for a considerable dis-
tance before forming a sharp junction with the pleated
ventral surface of the more posterior region, that constitutes
Fig. 1 a Scanning electron micrograph of the dorsal surface of the
tongue of Eublepharis macularius. Scale bar = 2 mm, magnification
179. HIND, MID, and FORE refer to tongue divisions described by
Schwenk (1985); white boxes indicate sampled tongue regions shown
in subsequent SEM images. b Outline of tongue shown in Fig. 1a.
Numbered regions refer to tongue divisions described herein.
Abbreviations: gl glottis, pf papilla-free zone at base of tongue
Table 1 Relative dimensions of papillary structures on the dorsal
lingual epithelium of Eublepharis macularius. The extreme tongue tip
is not included, as papillae are absent from this region. Values given
are ratios, to remove the effects of differences in absolute size of the
specimens examined. Original measurements were recorded in lm
Papilla type/location Height:Width Spacing:Width
Feather-like/tongue edge 2.374 ± 0.18 0.228 ± 0.04
Cuboidal/foretongue 1.711 ± 0.09 0.195 ± 0.02
Leaf-like/midtongue 4.874 ± 0.30 0.337 ± 0.03
Long fungiform/hindtongue 2.293 ± 0.16 0.261 ± 0.05
Short fungiform/hindtongue 1.823 ± 0.19 0.281 ± 0.08
Evol Biol
123

the extensible, protrusible portion of the part of the tongue
that lies free in the oral cavity. This thickened epithelium
wraps around the distal extremity of the tongue and extends
for a very short distance on its dorsal surface (Plate 1B)
before forming a sharp junction with the cuboidal papillate
region (see below). The extreme tongue tip is underlain by
two extensions of the hyoglossus muscle, one on either side
of, and close to, the midline. The ventral part of the tongue
tip (Plate 1A) bears a midline sulcus that interdigitates
with a process rising from the floor of the oral chamber
(Plate 1C). This tongue-in-groove association between the
tongue tip and oral floor provides the guiding and trans-
lation mechanism across which the tongue tip slides during
protrusion when the mouth is minimally open during
drinking, resulting in a pivoting and inversion of the
anterior part of the tongue during fluid collection (descri-
bed further below).
The foretongue (Plate 1D–G) exhibits cuboidal papillae
that are of uniform shape (Plate 1D) and are the most
densely packed of all types of papillae described. They are
approximately 1.7 times as high as wide (Table 1). The
surface of each papilla is covered by keratinized stratified
squamous epithelial cells (Plate 1E). No goblet clusters or
glandular epithelium are present. In this region of the
tongue the hyoglossus muscle is divided into four longi-
tudinal, finger-like processes on each side (Plate 1F) that
span from just lateral to the midline to the region of
juncture of the smooth ventral surface of the tongue and the
papillate tongue edge. The four, finger-like processes align
with a series of longitudinal pleats on the ventral surface of
the tongue (Plate 1A) in the region posterior to the
smoothly-clad extreme tongue tip.
Lying dorsal to the hyoglossus in the foretongue is a
mass of interwoven intrinsic tongue musculature and
Plate 1 (A) Tongue of Eublepharis macularius, tip visualized using
SEM, ventral view, scale bar 500 lm, magnification 789; (B) Tongue
of Eublepharis macularius, tip visualized using Masson’s Trichrome/
light microscopy, longitudinal section, scale bar 100 lm, magnifica-
tion 1009; (C) Tongue tip visualized using Masson’s Trichrome/light
microscopy, transverse section, scale bar 1 mm, magnification 109;
(D) Tongue of Eublepharis macularius, foretongue visualized using
SEM, scale bar 100 lm, magnification 5009, showing cuboid
papillae; (E) Tongue of Eublepharis macularius, foretongue visual-
ized using Masson’s Trichrome/light microscopy, anterior cross-
section, showing cuboid papillae and musculature, scale bar 100 lm,
magnification 1009; (F) Tongue of Eublepharis macularius,
foretongue visualized using Masson’s Trichrome/light microscopy,
anterior cross-section, scale bar 1 mm, magnification 409; (G)
Tongue of Eublepharis macularius, foretongue visualized using
macroscopic photograph of tongue in dorsal view showing tongue tip
engorged with blood; (H) Tongue of Gekko gecko, injected with
Microfil to visualize vasculature; (I) and (J) show a detailed view of
the vasculature of the fore- and hindtongue, respectively, of Gekkogecko. Abbreviations: cp capillary, ct connective tissue sheath,
ft foretongue, ht hindtongue, hy hyoglossus, in invagination, lc lat-
eral buccolingual chamber, lg longitudinalis, mt midtongue, pc pos-
terior buccolingual chamber, pl pleat, tb non-papillate tongue base,
tr transversalis, ve verticalis
Evol Biol
123

intervening connective tissue, organized into transverse,
vertical and longitudinal components. The longitudinalis
component is less distinct at the distalmost extremity of the
tongue and merges with the deeper verticalis component,
such that there is no real distinction between them
(Plate 1E). The intrinsic transversalis muscle strands are
highly interwoven with the verticalis strands, and the latter
extend from deep within the tongue, dorsal to the hyo-
glossus bundles (Plate 1F). They spread dorsolaterally to
intersect and merge with the transversalis strands that lie
deep to the cuboidal papillae (Plate 1F). The intrinsic
muscle strands connect with connective tissue sheaths
lying at the base of the papillae (Plate 1E). There are no
spaces within the tongue body in this region (Plate 1F).
The cuboidal papillae in this region are highly vascu-
larized (Plate 1E, H–J), with their core containing expan-
ded tufts of vessels that render the tongue tip red in life
(Plate 1G), visibly distinguishing it from more posterior
regions. The visible evidence of such vascularity is likely
due to the relatively thinner keratinized tips of these
papillae compared with tips of those situated more
posteriorly on the tongue (see below), as the degree of
vascularity itself is not greater in this region (Plate 1H–J),
although the terminal vascular tufts are more pronounced
(the intrinsic vasculature of the more posterior papillae as
more attenuated, in association with the longer and more
gracile form of these papillae—see below). These cuboidal
papillae receive extensions of the verticalis component of
the underlying intrinsic muscle feltwork (Plate 1E). Con-
siderably more such musculature intrudes into the lateral
papillae of this region of the tongue (see below). There is
no central skeletal core in the foretongue (Plate 1F).
An invagination that courses from dorsolateral to ven-
trolateral along the length of the tongue is situated dorso-
laterally in the extreme foretongue region (Plate 1C) and
reorients to a ventrolateral position as the posterior part of the
foretongue is approached (Plate 1F). This invagination
demarcates boundary between the papillate dorsal portion of
the tongue and the ventral tongue surface. It is bounded on its
dorsal border by epithelium that is heavily keratinized
(Plate 1F). Examination of the foretongue in situ (Plate 1A)
reveals that it is dished dorsally and makes up the ventral part
of a shallow anterior buccolingual chamber. Further poste-
riorly a pair of lateral buccolingual chambers are present,
demarcated by the juxtaposition of the papillate tongue
margin with the medial palatal shelves of the maxillae in the
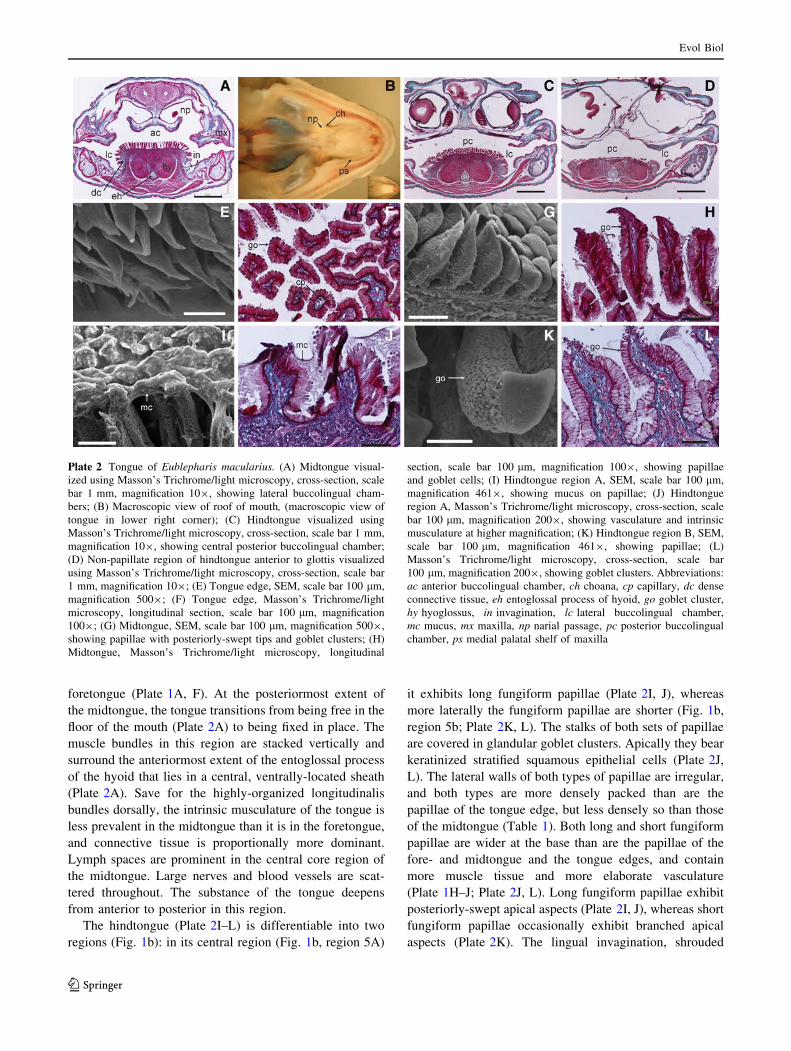
roof of the mouth (Plate 2A, B). The dorsal portion of the
mid-tongue (Fig. 1a), with its much taller papillae (see
below), lying ventral to the narial passages (Plate 2A, B), can
be employed to seal off the narial passages and isolate them
from the lateral buccolingual chambers. This junction of the
tongue with the maxillary shelves occurs at the position of
the lingual invagination and the dense connective tissue that
underlies it. More posteriorly still, in the glottal region
posterior to the narial passages (Plate 2B), a central posterior
buccolingual chamber (Plate 2C, D) lies dorsal to another
dished area of the tongue.
The remainder of the tongue, whose role in fluid transport
is less pronounced, also exhibits a variety of papilla types
and extensive vascularity. The tongue edges (Plate 2E, F),
with the exception of the extreme tongue tip and the fore-
tongue, exhibit long, slender, feather-like papillae with wide
bases and narrower tips (Plate 2E) that are not keratinized.
These papillae demarcate the junction between the papillate
dorsal surface and smooth ventral surface of the tongue. The
marginal papillae bear small, orthogonally-projecting
glandular goblet clusters (Plate 2F), discussed further
below, are invaded by capillaries (Plate 1J), contain both
muscle and connective tissue, and are relatively widely
spaced at their tips but more closely packed at their bases
(Plate 2E, F; Table 1). They are approximately 2.3 times as
high as wide (Table 1).
The midtongue (Plate 2G, H) exhibits irregular, tall
papillae with leaf-like, posteriorly-swept apical surfaces.
These are less densely packed than those of the foretongue,
but more densely packed than those of the tongue edge
(Table 1). The stalk of each papilla is covered by pro-
jecting glandular goblet clusters (Plate 2H), whereas the
apical surface is covered by strongly keratinized, stratified,
squamous epithelial cells (Plate 2H). Indeed, the exposed
basal stems of all papilla types, except the cuboidal
structures of the foretongue region, exhibit densely-packed
populations of goblet clusters that produce a thick mucus
that coats the tongue (Plate 2I). They are particularly
prolific in the mid- and hindtongue regions. Histological
sections reveal that these cells are largely devoid of con-
tents, and only the cell membranes stain, producing a
ghost-like effect (Plate 2H, L). The papillae of the mid-
tongue are relatively and absolutely taller than any of the
other types described (Table 1), vascularized (Plate 1J),
and contain both muscle and connective tissue (Plate 2H).
In the free region of the midtongue, the papillae are present
on the lateral surface of the tongue but do not extend more
ventrally than this. More posteriorly the papillae extend
further ventrolaterally (Plate 2A) and the lingual invagi-
nation assumes a more ventrolateral location, and is lined
by a thick, keratinized, simple columnar epithelium sur-
rounded by connective tissue.
The anterior region of the midtongue is free of the floor
of the mouth and houses four bundles of the hyoglossus
muscle, arrayed ventrally and ventrolaterally on either side
of the ventral midline of the tongue. The left and right
hyoglossus components are segregated by a strong, trian-
gular connective tissue core (Plate 2A). The ventral surface
of the tongue is girdled by strongly-developed connective
tissue and is pleated in a fashion similar to that seen in the
Evol Biol
123
foretongue (Plate 1A, F). At the posteriormost extent of
the midtongue, the tongue transitions from being free in the
floor of the mouth (Plate 2A) to being fixed in place. The
muscle bundles in this region are stacked vertically and
surround the anteriormost extent of the entoglossal process
of the hyoid that lies in a central, ventrally-located sheath
(Plate 2A). Save for the highly-organized longitudinalis
bundles dorsally, the intrinsic musculature of the tongue is
less prevalent in the midtongue than it is in the foretongue,
and connective tissue is proportionally more dominant.
Lymph spaces are prominent in the central core region of
the midtongue. Large nerves and blood vessels are scat-
tered throughout. The substance of the tongue deepens
from anterior to posterior in this region.
The hindtongue (Plate 2I–L) is differentiable into two
regions (Fig. 1b): in its central region (Fig. 1b, region 5A)
it exhibits long fungiform papillae (Plate 2I, J), whereas
more laterally the fungiform papillae are shorter (Fig. 1b,
region 5b; Plate 2K, L). The stalks of both sets of papillae
are covered in glandular goblet clusters. Apically they bear
keratinized stratified squamous epithelial cells (Plate 2J,
L). The lateral walls of both types of papillae are irregular,
and both types are more densely packed than are the
papillae of the tongue edge, but less densely so than those
of the midtongue (Table 1). Both long and short fungiform
papillae are wider at the base than are the papillae of the
fore- and midtongue and the tongue edges, and contain
more muscle tissue and more elaborate vasculature
(Plate 1H–J; Plate 2J, L). Long fungiform papillae exhibit
posteriorly-swept apical aspects (Plate 2I, J), whereas short
fungiform papillae occasionally exhibit branched apical
aspects (Plate 2K). The lingual invagination, shrouded
Plate 2 Tongue of Eublepharis macularius. (A) Midtongue visual-
ized using Masson’s Trichrome/light microscopy, cross-section, scale
bar 1 mm, magnification 109, showing lateral buccolingual cham-
bers; (B) Macroscopic view of roof of mouth, (macroscopic view of
tongue in lower right corner); (C) Hindtongue visualized using
Masson’s Trichrome/light microscopy, cross-section, scale bar 1 mm,
magnification 109, showing central posterior buccolingual chamber;
(D) Non-papillate region of hindtongue anterior to glottis visualized
using Masson’s Trichrome/light microscopy, cross-section, scale bar
1 mm, magnification 109; (E) Tongue edge, SEM, scale bar 100 lm,
magnification 5009; (F) Tongue edge, Masson’s Trichrome/light
microscopy, longitudinal section, scale bar 100 lm, magnification
1009; (G) Midtongue, SEM, scale bar 100 lm, magnification 5009,
showing papillae with posteriorly-swept tips and goblet clusters; (H)
Midtongue, Masson’s Trichrome/light microscopy, longitudinal
section, scale bar 100 lm, magnification 1009, showing papillae
and goblet cells; (I) Hindtongue region A, SEM, scale bar 100 lm,
magnification 4619, showing mucus on papillae; (J) Hindtongue
region A, Masson’s Trichrome/light microscopy, cross-section, scale
bar 100 lm, magnification 2009, showing vasculature and intrinsic
musculature at higher magnification; (K) Hindtongue region B, SEM,
scale bar 100 lm, magnification 4619, showing papillae; (L)
Masson’s Trichrome/light microscopy, cross-section, scale bar
100 lm, magnification 2009, showing goblet clusters. Abbreviations:
ac anterior buccolingual chamber, ch choana, cp capillary, dc dense
connective tissue, eh entoglossal process of hyoid, go goblet cluster,
hy hyoglossus, in invagination, lc lateral buccolingual chamber,
mc mucus, mx maxilla, np narial passage, pc posterior buccolingual
chamber, ps medial palatal shelf of maxilla
Evol Biol
123

by connective tissue, partially underlies the hindtongue
(Plate 2C).
Anteriorly, the hindtongue exhibits stout, vertically
stacked, paired subdivisions of the hyoglossus muscle
ventrolaterally, flanking, but at some distance from, the
central niche in which the lingual process of the hyoid
apparatus sits (Plate 2C). At the very base of the tongue
there is a central, papilla-free zone (Fig. 1, Plate 2D). The
hyoglossus muscle is much less discrete at this point, and
no subdivision into bundles is evident.
Behaviour
High-speed cinematography reveals that E. macularius
gathers fluid by cyclical protrusive-retractive motions of
the tongue, resulting in a fluid bolus being gathered by the
foretongue and eventually conveyed to the stomach by
head-tossing behavior and deglutition. In fluid gathering,
the extreme tongue tip acts as a fulcrum over which the
papillate foretongue pivots and inverts (Fig. 2a, b, d, e).
This pivot mechanism results in presentation of the dorsal
surface of the foretongue directly to the fluid as it rolls
out of the mouth (Fig. 2c, f, g). The foretongue is
greatly expanded on the substrate as it touches the water
(Fig. 2f, g), maximizing the surface area presented to the
water. Following several such cycles, emersion is achieved
by the head being raised. The protraction-retraction cycles
of the tongue continue briefly.
Radiographic images of E. macularius drinking barium-
labelled water (Bels et al. unpublished) reveal that the
tongue tip is loaded with fluid, and that water is unloaded
from the tongue into the anterior buccolingual chamber in
the region of the vomeronasal sinus. In the following cycle
fluid is removed from this region and transported to the pair
of lateral buccolingual chambers (Plate 2C), at the same
time that a new bolus of fluid is being gathered by the
foretongue and transported to the anterior buccolingual
chamber. The original bolus of fluid moves laterally from
the anterior buccolingual chamber to the lateral buccolin-
gual chambers by traversing the tongue laterally in the
region of the junction of the foretongue and midtongue.
Fluid is thus kept segregated from the mucus-rich part of
the tongue (Plate 2I) and is ultimately guided to the val-
vular glottal area, posterior to the region of air flow, where
it enters a third, central chamber (Plate 2C, D) prior to
being swallowed, assisted by an upward tilting of the head.
The hindtongue moves very little in the protraction-
retraction cycles of the foretongue, and remains essentially
Fig. 2 Frames from high-speed cinematography of E. macularius during drinking in frontal (a–c) and dorsal (d–g) views. The foretongue pivots
over the extreme tongue tip and expands laterally as its dorsal surface is presented to the liquid. See text for further details
Evol Biol
123

fixed relative to the skull, with only the mid- and foretongue
being displaced appreciably. The hindtongue is involved in
fluid transportation only inasmuch as it forms the medial and
dorsal boundaries of the internal oral compartments.
Discussion
Our observations allow us to relate the macroscopic
structure of the tongue and oral chamber of E. macularius
to the process of fluid uptake. The tongue is generally
similar, both in macroscopic and histological morphology,
to the tongue of other lizards. Iwasaki (1990) described
three types of papillae on the tongue of Gekko japonicus:
dome-shaped, which correspond to the cuboidal papillae
described here in the foretongue region (Plate 1D–G);
fan-shaped, which correspond to the leaf-like papillae
described in the present study in the midtongue region
(Plate 2G, H); and scale-like, which correspond to the short
and long fungiform papillae observed in the present study
in the hindtongue region (Plate 2I–L).
The lack of papillae on the extreme tongue tip was also
noted by Delheusy et al. (1994) for Oplurus cuvieri.
Although they reported similar foretongue papillae for that
taxon, the remainder of the tongue of O. cuvieri differs
noticeably from that of E. macularius. Papillae present in
the fore, mid- and hindtongue of O.cuvieri are generally
cylindrical in shape, and their tips are far less elaborate
than the leaf-like and fungiform papillae observed herein.
We describe the presence of glandular goblet clusters on
the exposed stalks of papillae in the mid- and hindtongue
regions. Iwasaki (1990) noted a large number of cells
containing secretory granules on the tongue of G. japoni-
cus, and Delheusy et al. (1994) reported abundant mucus
secretions in O. cuvieri. Rabinowitz and Tandler (1986)
described the morphology of papillae on the tongue of
Anolis carolinensis and designated these as ‘plumose’, on
the basis of the presence of ‘plume cells’. Comparison of
these cells (Rabinowitz and Tandler 1986: Fig. 7) to the
goblet clusters observed in E. macularius (Plate 2E–L)
suggests a structural similarity, although there are mor-
phological features that seemingly preclude their homol-
ogy. The goblet clusters we describe are more cuboidal and
exhibit few cellular contents. They invest the stems of the
papillae and extend from the papillary base to the kera-
tinized tip, whereas the plume cells of Rabinowitz and
Tandler (1986) are restricted to the apical portion of the
papillae. It is possible that the clusters we describe repre-
sent a more elaborately expressed form of those observed
by Rabinowitz and Tandler (1986), but further study is
required to test this hypothesis.
Interestingly, we failed to establish the presence of taste
buds in E. macularius. Schwenk (1985) found evidence for
widespread presence of taste buds across Lepidosauria, but
among the Gekkota he noted their absence from the tongue
of Gonatodes antillensis, and their presence only on the
hindtongue of Coleonyx variegatus. Our results provide
further evidence that the Gekkota may largely lack taste
buds. Although the explanation for this phenomenon may
be phylogenetic, there may also be a functional or adaptive
significance to it.
Our findings allow us to demonstrate the morphological
basis for fluid transport in relation to the structure of the
tongue and the associated oral regions of E. macularius.
This functional complex consists of papillae of particular
shapes and orientations; the cross-sectional morphology of
the tongue in its various regions; the association of cross-
sectional morphology with the morphology of the roof of
the mouth and the narial passageways; the form of the
lateral regions of the buccal cavity and their segregation
from the dorsal region by maxillary flanges; the muscular
organization of the tongue facilitating the interaction of
these features; and the form and pattern of distribution of
the lingual papillae.
When placed into the context of its in situ location, the
foretongue is dished dorsally and makes up the ventral part
of an anterior buccolingual chamber (Plate 1C), lying
anterior to the non-olfactory portions of the narial pas-
sageways. The cuboidal papillate region of the foretongue
in this location lies medial to the opening of the nares into
the oral chamber (Plate 1C), thus segregating the water-
laden portion of the tongue from the olfactory area. The
spacing between the papillae is wide (Plate 1D, E), and,
upon tongue extension, creates capillary channels for fluid
pickup as the dorsal surface is stretched and expanded
(Fig. 2e, f, g) by being rolled out of the mouth (with the
extreme tip remaining within the confines of the buccal
cavity; Fig. 2c). During protraction, the tongue moves over
the mandibular symphysis and expands laterally as it does
so—a function attributable to the longitudinalis compo-
nents of the intrinsic tongue muscles that lie in bundles at
the base of the papillae (Plate 1E, F). The tines of the
foretongue remain in the buccal cavity, employing the cleft
in the tongue and the process that rises from the floor of the
mouth beneath it, as a fulcrum-and-anchor device. The
rounded, laterally spread dorsal surface of the foretongue is
extended and presented to the fluid (Fig. 2b, c). During
retraction fluid is held in the interpapillary pockets and no
bead of fluid is evident on the tongue surface during
retraction (Bels et al. unpublished). The curled tongue is
straightened as it is drawn into the mouth (Fig. 2). The
absence of mucus glands from the foretongue is compatible
with its fluid transportation function, allowing fluid to enter
deep into the interpapillary spaces. Contraction of the
transversalis muscles (Plate 1E) will narrow the foreton-
gue, reducing the spaces between the papillae, and the
Evol Biol
123

verticalis components (Plate 1E) will shorten the papillae.
Collectively these two muscle masses are likely responsi-
ble for the change of configuration of the foretongue during
retraction, resulting in compression of the interpapillary
spaces and the forcing of fluid onto the dorsal surface of the
tongue, enabling its collection in the anterior buccolingual
chamber (Plate 2A–D), ventral to the maxillary shelves and
lateral to the non-olfactory portions of the narial passage-
ways. The form of the tongue and the roof of the mouth are
configured such that fluid travels into the lateral chambers
(Plate 2A) and remains segregated from the narial pas-
sages. Interestingly, Herrel et al. (2005) report the presence
of papillae on the ventral surface of the foretongue of
Lacerta spp. (which they term ‘fungiform’) that are very
similar in shape to the cuboidal papillae described herein.
These species are known to use the ventral aspect of the
tongue to gather water when drinking.
The presence of muscle fibers within papillae, as noted
for A. carolinensis by Rabinowitz and Tandler (1986),
allows neurological control of the movements of these
structures. Such control would permit the lizard to alter-
nately open and close the spaces between papillae, and to
alter their dimensions. This would facilitate both collection
and retention of water in the foretongue region during
pickup, and release at the time of transfer, and would also
allow differential control of the amount of mucus present
on the tongue. We suggest, given the cloying nature of
mucus secretions, that papillae would be relatively
expanded in order to increase the amount of mucus present
on the tongue during feeding, and would be relatively
contracted in order to decrease the amount of mucus
present on the tongue during drinking. Further experi-
mentation is required to test this hypothesis.
Previous authors (e.g. Schwenk 1985; Smith 1988;
Beisser et al. 1998, 2004; Iwasaki 2002; Herrel et al. 2005)
have postulated that the presence of papillae on the tongue of
sauropsids is explained either phylogenetically, or by the
need for adaptation for food procurement and processing in a
terrestrial environment. Food prehension is jaw- rather than
tongue-based in Gekkotans (Delheusy and Bels 1999;
Schwenk 2000; Bels 2003; Reilly and McBrayer 2007),
leaving the tongue free for extra-oral functions. Herein we
present the first evidence of a fluid procurement and trans-
portation function for the papillate Gekkotan tongue, and
relate this to the overall form and configuration of the oral
chamber. Spectacle-wiping (Bustard 1963) by the tongue may
also involve the use of the non-glandular part of the tongue,
allowing for cleaning without the deposition of mucus on the
spectacle surface. Although not a feature of eublepharid ge-
kkotans, because they have eyelids, this behaviour would
likely have co-opted the non-glandular part of the tongue as an
exaptation in all gekkotans that have a spectacle (see Cogger
[1967: Plate 7] for an example of this behaviour).
Acknowledgments We thank K. Kardong for discussion, and
P. Wise for providing specimens. This research was supported in part
by an NSERC Discovery Grant to APR.
References
Beisser, C. J., Lemell, P., & Weisgram, J. (2004). The dorsal lingual
epithelium of Rhinoclemmys pulcherrima incisia (Chelonia,
Cryptodira). The Anatomical Record, 277A, 227–235.
Beisser, C. J., Weisgram, J., Hilgers, H., & Splechtna, H. (1998). Fine
structure of the dorsal lingual epithelium of Trachemys scriptaelegans (Chelonia: Emydidae). The Anatomical Record, 250,
127–135.
Bels, V. L. (2003). Evaluating the complexity of the trophic system in
Reptilia. In V. L. Bels, J. P. Gasc, & A. Casinos (Eds.),
Vertebrate biomechanics and evolution (pp. 185–202). Oxford:
BIOS Scientific.
Bustard, H. R. (1963). Gekko behavioral trait: Tongue wiping
spectacle. Herpetologica, 19, 217–218.
Camp, C. L. (1923). Classification of the lizards. Bulletin of theAmerican Museum of Natural History, 48, 289–481.
Cogger, H. G. (1967). Australian reptiles in colour. Sydney: AH &
AW Reed.
Delheusy, V., & Bels, V. L. (1999). Feeding kinematics of Phelsumamadagascariensis (Reptilia: Gekkonidae): Testing differences
between Iguania and Scleroglossa. Journal of ExperimentalBiology, 202, 3715–3730.
Delheusy, V., Toubeau, G., & Bels, V. L. (1994). Tongue structure
and function in Oplurus cuvieri (Reptilia: Iguanidae). TheAnatomical Record, 238, 263–276.
de Rooij, N. (1915). The reptiles of the Indo-Australian archipelago(Vol. 1). Lacertilia, Chelonia, Emydosauria. Leiden: EJ Brill.
Gnanamuthu, C. P. (1937). Comparative study of the hyoid and
tongue of some typical genera of reptiles. Proceedings Zoolog-ical Society of London, 107B, 1–63.
Herrel, A., Canbek, M., Ozelmas, U., Uyanoglu, M., & Karakaya, M.
(2005). Comparative functional analysis of the hyolingual
anatomy in lacertid lizards. The Anatomical Record, 284A,
561–573.
Humason, G. L. (1979). Animal tissue techniques. San Francisco:
W.H. Freeman.
Iwasaki, S. (1990). Fine structure of the dorsal lingual epithelium of
the lizard, Gekko japonicus (Lacertilia, Gekkonidae). TheAmerican Journal of Anatomy, 187, 12–20.
Iwasaki, S. (2002). Evolution of the structure and function of the
vertebrate tongue. Journal of Anatomy, 201, 1–13.
Iwasaki, S., & Miyata, K. (1985). Scanning electron microscopy of the
lingual dorsal surface of the Japanese lizard, Takydromus tachy-dromoides. Okajimas Folia Anatomica Japonica, 62, 15–26.
Iwasaki, S., Miyata, K., & Kobayaski, K. (1985). Fine structure of the
oral epithelial cell surface in the Japanese lizard, Takydromustachromoides. Japanese Journal of Oral Biology, 27, 956–964.
Rabinowitz, T., & Tandler, B. (1986). Papillary morphology of the
tongue of the American Chameleon: Anolis carolinensis. TheAnatomical Record, 216, 483–489.
Reilly, S. M., & McBrayer, L. (2007). Prey capture and prey
processing behavior and the evolution of lingual and sensory
characteristics: Divergences and convergences in lizard feeding
biology. In S. M. Reilly, L. D. McBrayer, & D. B. Miles (Eds.),
The evolutionary consequences of foraging mode (pp. 302–333).
Cambridge: Cambridge University Press.
Russell, A. P., Walker, R. L., & Bauer, A. M. (1988). A technique for
visualization of the circulatory system in small lizards. Copeia,1988, 797–800.
Evol Biol
123

Schwenk, K. (1985). Occurrence, distribution and functional signif-
icance of taste buds in lizards. Copeia, 1985, 91–101.
Schwenk, K. (1986). Morphology of the tongue in the Tuatara,
Sphenodon punctatus (Reptilia: Lepidosauria), with comments
on function and phylogeny. Journal of Morphology, 188,
129–156.
Schwenk, K. (2000). Feeding in lepidosaurs. In K. Schwenk (Ed.),
Feeding: Form, function and evolution in tetrapod vertebrates(pp. 175–291). San Diego: Academic Press.
Sherbrooke, W. C., Scardino, A. J., de Nys, R., & Schwartzkopf, L.
(2007). Functional morphology of the scale hinges used to
transport water: Convergent drinking adaptations in desert
lizards (Moloch horridus and Phrynosoma cornutum). Zoomor-phology, 126, 89–102.
Smith, K. K. (1986). Morphology and function of the tongue and
hyoid apparatus in Varanus (Varanidae, Lacertilia). Journal ofMorphology, 187, 261–287.
Smith, K. K. (1988). Form and function of the tongue in agamid
lizards with comments on its phylogenetic significance. Journalof Morphology, 196, 157–171.
Toubeau, G., & Bels, V. L. (1994). Morphological and kinematic
study of the tongue and buccal cavity in the lizard Anguis fragilis(Reptilia: Anguidae). The Anatomical Record, 240, 423–433.
von Seiler, R. F. (1891). Ueber die Zungendrusen von Anguis,
Pseudopus und Lacerta. Arch Fur Mikrosk Anat, 38, 177–265.
von Seiler, R. F. (1892). Die Zungendriisen von Lacerta. Festschrift
fur R. Leuckart, pp. 250–258.
Witten, P. E., & Hall, B. K. (2003). Seasonal changes in the lower jaw
skeleton in male Atlantic salmon (Salmo salar L.): Remodeling
and regression of the kype after spawning. Journal of Anatomy,203, 435–450.
Evol Biol
123
Top Related
Copyright © 2022 FDOKUMEN

