







Bahasa
Halaman
Hukum


Eur. J. Biochem. 236, 171-179 (1996) 0 FEBS 1996
Identification of a functional cAMP response element in the secretogranin I1 gene Giuseppe CIBELLI’, Susanne JUNGLING’, Susanne SCHOCH I, Hans-Hermann GERDES’ and Gerald THIEL’
’ Institute for Genetics, University of Cologne, Germany * lnstitute for Neurobiology, University of Heidelberg, Germany
(Received 3 July/30 October 1995) ~ EJB 95 1073/2
Secretogranin I1 is an acidic secretory protein with a widespread distribution in secretory granules of neuronal and endocrine cells. The secretogranin I1 gene contains, like other members of the granin family, a cAMP response element (CRE) in its upstream region. To investigate the functional significance of this motif, intracellular cAMP levels were increased in a neuronal cell line derived from the septal region of the brain and the level of secretogranin I1 gene expression was analysed. It was found that increased cAMP levels did, in fact, induce secretogranin I1 gene expression. To analyse the cis-acting sequence responsible for this induction, a hybrid gene containing the upstream region of the mouse secretogranin I1 gene fused to P-globin as a reporter was constructed. Transfection analysis revealed that CAMP-induced transcription of the secretogranin I1 promoterlP-globin gene in septal and insulinoma cells. DNA-protein binding assays showed that recombinant CRE-binding protein (CREB), produced in bacteria or human cells, bound in a sequence-specific manner to the secretogranin I1 promoter CRE. Moreover, deletion mutagenesis revealed that the CRE motif is a bifunctional genetic regulatory element in that it mediates basal as well as CAMP-stimulated transcription. Interestingly, cAMP had no effect upon secretogranin TI gene transcription in PC12 and neuroblastoma cells. An increase in the intracellular cAMP concentration activated a GAL4-CREB fusion protein upon transcription in neuroblastoma cells indicating the integrity of the cAMP signaling pathway to the nucleus. Basal as well as CAMP-stimulated transcription, directed from the secretogranin I1 promoter was, however, impaired in insulinoma cells by overexpression of CREB-2, a negative-acting CRE-binding protein. These results indicate that competitive effects are likely to occur between CRE-bound transcriptional activators and repressors. We conclude that CAMP-stim- ulated induction of secretogranin I1 gene transcription is mediated by the CRE motif in a cell-type-specific manner, and is likely to depend on the balance between positive and negative CRE-binding proteins in a particular cell type.
Keywords: secretogranin I1 ; chromogranin B ; synapsin I ; CAMP-response-element-binding protein ; CAMP; transcription.
One of the most studied intracellular signaling pathways uses cAMP as a second messenger. Despite the regulation of multiple cellular processes by CAMP, the effects of increased cAMP con- centrations upon gene regulation has attracted much attention. In mammalian cells, the activation of the CAMP-dependent protein kinase (PKA) is believed to transduce all known effects of CAMP. Binding of cAMP to the regulatory subunit of PKA re- leases its catalytic subunit, which subsequently translocates into the nucleus, where it can phosphorylate substrates on serine and threonine residues. In particular, PKA phosphorylates and acti- vates a family of basic regionlleucine zipper transcription factors that are bound to the palindromic sequence 5’-TGACGTCA-3’ known as the CAMP-response element (CRE). The most promi- nent member of this transcription factor family is termed CRE- binding protein (CREB ; Montminy and Bilezikjian, 1987 ; Hoef-
Correspondence to G. Thiel, Institute for Genetics, University of
Fax: +49 221 470 5172. Abbreviations. Bt,cAMP N6,2’-O-dibutyryladenosine 3’,5’-mono-
phosphate; CRE, cAMP response element; CREB, CRE binding protein; GST, glutathione S-transferase; iBuMeXan, 3-isobutyl-1 -methyl-xan- thine; PKA, protein kinase A.
Cologne, Zhlpicher Str. 47, D-50674 Cologne, Germany
Enzyme. Protein kinase A (EC 2.7.1.37).
fler et al., 1988). Many CRE binding proteins have been discov- ered in recent years, exhibiting positive as well as negative tran- scriptional activities (for review, see Meyer and Habener, 1993 ; Lalli and Sassone-Corsi, 1994; Vallejo and Habener, 1994) sug- gesting that complex, often cell-type-specific mechanisms, con- trol the effects of cAMP upon transcription.
While cAMP and the CAMP-dependent protein kinase are present in virtually all cell types, the CAMP-mediated signaling pathway appears to play a particularly important role in regulat- ing brain function through both short-term and long-term effects. Short-term effects are triggered by a direct phosphorylation reaction of effector molecules such as ion channels or synaptic vesicle proteins by the CAMP-dependent protein kinase. Long- term effects, also mediated by protein phosphorylation, require changes in the gene expression repertoire of a particular neuron, mediated by the CRE and the transcription factor CREB.
Here an analysis of the regulation of the secretogranin I1 gene by cAMP is described. Secretogranin 11, one member of the granin family, is an acidic secretory protein found in secretory granules of most peptidergic endocrine cells and neurons. In the brain, the highest secretogranin I1 immunoreactivity is found in the hippocampus, the hypothalamus and the amygdaldpiriform cortex (Weiler et al., 1990). It has been suggested that secreto-

172 Cibelli et al. ( E M J . Biochem. 236)
granin 11, together with other members of the granin family such as chromogranin A and chromogranin B, plays a role in the sorting process to secretory granules (for reviews, see Scammell, 1993; Rosa and Gerdes, 1994; Huttner and Natori, 1995). In addition, a role as a precursor molecule for the generation of secretoneurin has been proposed for secretogranin IT (Kirchmair et al., 1994).
The genes encoding the granin proteins contain CRE motifs in their 5’-upstream regions. To date, there are conflicting results upon CAMP-mediated transcriptional regulation of the secreto- granin I1 gene. Forskolin, an activator of adenylyl cyclase, increased secretogranin 11 mRNA levels in chromaffin cells (Fi- scher-Colbrie et al., 1990), yet caused a decrease in PC12 pheochromocytoma cells (Thompson et al., 1992, 1994). A series of transfection experiments described here revealed that secretogranin I1 transcription i s induced by CAMP in some, but not all cell types tested. Cell-type-specific mechanisms appear to play a critical role in the ability of CAMP to induce transcription mediated by the CRE element of the secretogranin IT promoter, thus explaining the conflicting results of previous reports. Fur- ther, our data indicate that it is the balance between positive and negative-acting CRE-binding proteins in a given cell type which is likely to determine whether CRE-mediated transcription i s ac- tivated through CAMP. Additionally, we have found that the pro- moter context of a particular CRE element is critical for CAMP- mediated transcriptional regulation.
EXPERIMENTAL PROCEDURES
Reporter constructs. Plasmids OVEC, SP6/ffS and ICPOref were described recently (Thiel et al., 1994). Fragments of the 5’- upstream region of the secretogranin I1 gene were excised from plasmid pBS-mSgIIpro which contains a 4.5-kb fragment of the promoter region of the mouse secretogranin 11 gene (Schimmel et al., 1992). The secretogranin IT promoterlp-globin reporter plasmids pSgIIOVEC-1, pSgIIOVEC-2, and pSgIIOVEC-3, were constructed by subcloning the following restriction frag- ments of the secretogranin 11 promoter into OVEC: a XmnI- EheI fragment (sequence -330 to -39): a SphI-EheI fragment (sequence -160 to -39); a XmnI-DdeI fragment (sequence -330 to -73). Plasmid pSgIIOVEC-ld, containing a 4-bp dele- tion of the CRE core motif, was constructed using AatII and T4 DNA polymerase as described (Knepel et al., 1990). Plasmids pCgBOVEC-2 and pSyOVEC-3 have been previously described (Jungling et al., 1994).
Expression plasmids. Plasmids pCMVCa and pCMVCREB-2 were described recently (Jungling et al., 1994). pGAL4-CREB was constructed by inserting the entire human CREB cDNA (Hoeffler et al., 1988) into pM1 (Sadowski et al., 1992). The GAL4 fusion vector pMI was a kind gift of I. Sadowski (Univer- sity of British Columbia, Vancouver).
Cell culture. The origins of various clones of PC12 pheo- chromocytoma cells are indicated. The clone PC12/NY was used in previous experiments (Thiel et al., 1991; Jungling et al., 1994). The clones PC12/SF (Shackleford et al., 1993) and PC12/ CT were provided by K. Willert and Harold E. Varmus (Univer- sity of California, San Francisco) and Eric Schaeffer (Pfizer Inc., Groton CT), respectively. PC12-B9 cells contain an integrated retrovirus encoding the v-src gene under control of the met- allothionein promoter (Cox and Maness, 1991). PC12/Wnt-l cells express the mouse wnt-1 cDNA under the control of the long terminal repeat of the Moloney murine leukemia virus (Shackleford et al., 1993). All PC12 cell clones were cultured in 85% Dulbecco’s modified Eagle medium, 10% horse serum, 5 % fetal calf serum, 100 units/ml of penicillin, and 100 mg/ml
of streptomycin. The cell lines SN56, NS20Y, NG108-15, NS26. HIT-TlS, and 293T/17 were cultured as described (Thiel et al., 1991, 1994: Jungling et al., 1994). Differentiation was induced in PC12 and SN56 cells by the addition of 1 mM P,2’-O-dibu- tyryladenosine 3’,5’-monophosphate (Bt,cAMP) and 0.5 mhl 3- isobutyl-1-methyl xanthine (iBuMeXan). In those experiments in which neurite outgrowth was examined, cells were plated on tissue-culture dishes coated with collagen. For transfection ex- periments, cells were plated on uncoated tissue-culture plates. For visualizing neurite outgrowth, cells were fixed and stained with May-Grunwald solution and Giemsa (Merck).
Northern blots. Northern blots were performed as described (Jungling et al., 1994). To detect the secretogranin IT mRNA, a =1.7-kb XhoI fragment derived from the rat secretogranin 11 cDNA (Gerdes et al., 1988) was used as a hybridization probe.
Transfections. Cells were transfected by the calcium phos- phate coprecipitation procedure (Thiel et al., 1991). In general, 5 pg p-globin reporter plasmid and 2.5-5 pg internal refercnce plasmid ICPOref were added to each 100-mm plate. The expres- sion plasmids were used at a concentration of 3.5 pg (pCMVCa), 5 pg (pGAL4-CREB) or 10 pg (pCMVCREB-2)/ plate. All transfections were normalized for the amount of plas- mid and cytomegalovirus IE promoter by the addition of plas- mids pCB6 and pCMVS, respectively.
RNase protection mapping. RNase protection assays v:ere performed as described (Thiel et al., 1994) using a rabbit /)- globin cRNA probe derived from plasmid SP6flS. The pro- tected fragments generated by the test and reference plasmids were quantified using the BAS 1000 Bio-imaging analyzer (Fuji).
Expression of CREB in Escherichia coli. To express the DNA binding and dimerization domains of CREB as a fusion protein with Schistosoma japonicum glutathione S-transferase (GST), a BglII-EcoRI fragment of plasmid Rc/RSV-CKEB (Hoeffler et al., 1988) was cloned into pGEX-3X (Pharmacia) thus generating plasmid pGEX-CREB. This plasmid was i n tro- duced into the protease-deficient E. coli strain NB42 (Maytx et al., 1992). The GST-CREB fusion protein was expressed and purified as described (Smith and Johnson, 1988).
Expression of CREB in 293 human kidney cells. 293T/t 7 cells were transfected with pRc/RSV-CREB as indicated ahove and nuclear extracts were prepared as described (Thiel et al., 1994).
Electrophoretic mobility shift assay. Binding assays were performed as described (Thiel et al., 1994: Jungling et al., 1994). The radiolabeled secretogranin I1 CRE probe was obtained by annealing oligonucleotides S-TCGAGAGCCGGTGACGTCA- CCGTGGAAG-3’ and 5’-TCGACTTCCACGCTGACGTC4C- CGGCTC-3’. As an unrelated competitor, the polyoma enhancer activator 3 (PEA3) binding site of the human synapsin I1 pro- moter was used as described (Thiel et al., 1994).
DNase I footprinting. Binding reactions (in S O pl) con- tained 25 mM Hepes, pH7.6, 50mM KC1, 6.25 mM MgCI,, 0.5 mM EDTA, 0.5 mM dithiothreitol, 10% glycerol, 20 pg/ml poly(d1-dC), 20000-30000 cpm of radiolabeled probe and 12.5 pg recombinant GST or GST-CREB fusion protein. The samples were incubated on ice for 10 min and shifted to room temperature for 2 min; 50 1-11 of a room-temperature solution containing 5 mM CaCI, and 10 mM MgC1, was then added and the incubation continued for an additional 1 min. DNaie I (Boehringer Mannheim) was added for 1 min. The reaction was terminated by adding 100 pl DNase I stop solution (0.2 M NaCl, 0.03 M EDTA, 1 % SDS, 100 pg/ml yeast RNA). The samples were incubated for 30 min at 37”C, extracted with phenol/chlo- roform, precipitated with ethanol and finally separated on ii se- quencing gel.
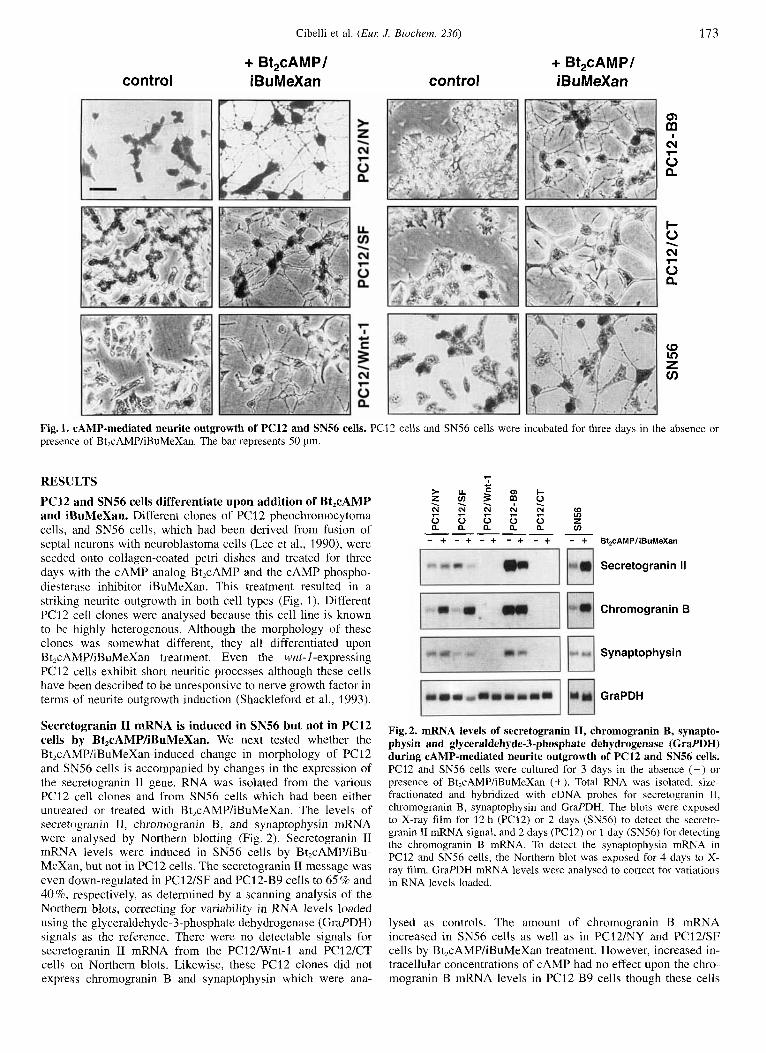
Cibelli et al. ( E m J. Biochem. 236) 173
control + Bt,cAMP/ + Bt,cAMP/ iBuMeXan control iBuMeXan
Q,
cv m
E n
I
Fig.1. CAMP-mediated neurite outgrowth of PC12 and SN56 cells. PC12 cells and SN56 cells were incubated for three days in the absence or presence of Bt,cAMP/iBuMeXan. The bar represents 50 pm.
RESULTS
PC12 and SN56 cells differentiate upon addition of Bt,cAMP and iBuMeXan. Different clones of PC12 pheochromocytoma cells, and SN56 cells, which had been derived from fusion of septa1 neurons with neuroblastoma cells (Lee et al., 1990), were seeded onto collagen-coated petri dishes and treated for three days with the cAMP analog Bt,cAMP and the cAMP phospho- diesteraye inhibitor iBuMeXan. This treatment resulted in a striking neurite outgrowth in both cell types (Fig. 1). Different PC12 cell clones were analysed because this cell line is known to be highly heterogenous. Although the morphology of these clonec was somewhat different, they all differentiated upon Bt2cAMP/iBuMeXan treatment. Even the wnt-I-expressing PC12 cells exhibit short neuritic processes although these cells have been described to be unresponsive to nerve growth factor in terms of neurite outgrowth induction (Shackleford et al., 1993).
Secretogranin I1 mRNA is induced in SN56 but not in PC12 cells by Bt,cAMP/iBuMeXan, We next tested whether the Bt,cAMP/iBuMeXan-induced change in morphology of PC12 and SN56 cells is accompanied by changes in the expression of the secretogranin I1 gene. RNA was isolated from the various PC12 cell clones and from SN56 cells which had been either untreated or treated with Bt,cAMP/iBuMeXan. The levels of secretogranin 11, chromogranin B, and synaptophysin mRNA were analysed by Northern blotting (Fig. 2). Secretogranin I1 mRN4 levels were induced in SN56 cells by Bt,cAMP/iBu- MeXan, but not in PC12 cells. The secretogranin I1 message was even down-regulated in PC12/SF and PCl2-B9 cells to 65% and 40%, respectively, as determined by a scanning analysis of the Northern blots, correcting for variability in RNA levels loaded using the glyceraldehyde-3-phosphate dehydrogenase (Graf DH) signals as the reference. There were no detectable signals for secretogranin I1 mRNA from the PC12Nnt-1 and PC12ICT cells on Northern blots. Likewise, these PC12 clones did not express chromogranin B and synaptophysin which were ana-
----- - - + - + - + - + - + - + Bt,cAMPliBuMeXan
Secretogranin II
Chromogranin B
Synaptophysin
GraPDH
Fig. 2. mRNA levels of secretogranin 11, chromogranin B, synapto- physin and glyceraldehyde-3-phosphate dehydrogenase (GraPDH) during CAMP-mediated neurite outgrowth of PC12 and SN56 cells. PC12 and SNS6 cells were cultured for 3 days in the absence (-) or presence of Bt,cAMP/iBuMeXan (+). Total RNA was isolated, size- fractionated and hybridized with cDNA probes for secretogranin 11, chromogranin B, synaptophysin and GraPDH. The blots were exposed to X-ray film for 12 h (PC12) or 2 days (SN56) to detect the secreto- granin I1 mRNA signal, and 2 days (PC12) or 1 day (SN56) for detecting the chromogranin B mRNA. To detect the synaptophysin mRNA in PC12 and SNS6 cells, the Northern blot was exposed for 4 days to X- ray film. GraPDH mRNA levels were analysed to correct for variations in RNA levels loaded.
lysed as controls. The amount of chromogranin B mRNA increased in SN56 cells as well as in PC12NY and PC12/SF cells by Bt,cAMP/iBuMeXan treatment. However, increased in- tracellular concentrations of cAMP had no effect upon the chro- mogranin B mRNA levels in PC12-B9 cells though these cells
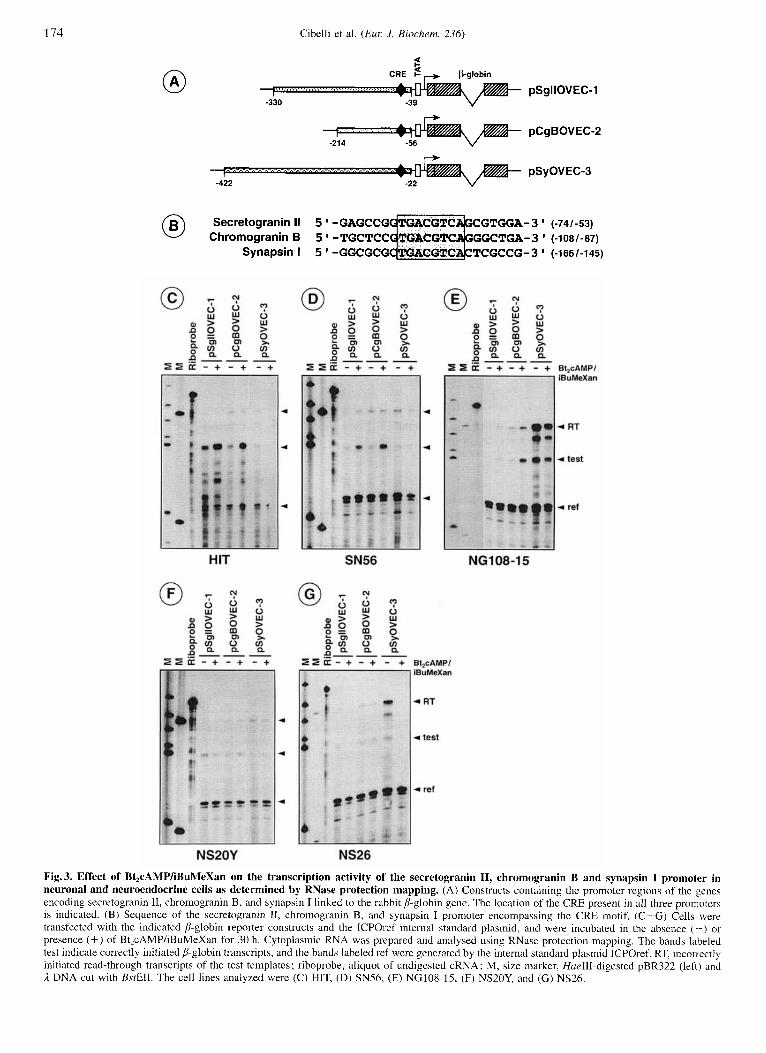
174 Cibelli et al. (Eur: J . Biochem. 236)
PSgIIOVEC-1 -330
pCg BOVEC-2
pSyOVEC-3
Secretogranin 11 5 -GAGCC TGGA- 3 (-74 I - 53) @ Chromogranin B 5 I -TGCTC GCTGA-3 (-1081-87) Synapsin I 5 I -GGCGC GCCG-3 (-1661-145)
Fig. 3. Effect of Bt,cAMP/iBuMeXan on the transcription activity of the secretogranin 11, chromogranin B and synapsin I promoter in neuronal and neuroendocrine cells as determined by RNase protection mapping. (A) Constructs containing the promoter regions of the genes encoding secretogranin 11, chromogranin B, and synapsin I linked to the rabbit b-globin gene. The location of the CRE present in all three promoters is indicated. (B) Sequence of the secretogranin 11, chromogranin B, and synapsin I promoter encompassing the CRE motif. (C-G) Cells were trdnsfected with the indicated P-globin reporter constructs and the ICPOref internal standard plasmid, and were incubated in the absence ( - - ) or presence (+) of BtZcAMP/iBuMeXan for 30 h. Cytoplasmic RNA was prepared and analysed using RNase protection mapping. The bands labeled test indicate correctly initiated ,!-globin transcripts, and the bands labeled ref were generated by the internal standard plasmid ICPOref. RT, incori.ectly initiated read-through transcripts of the test templates; riboprobe, aliquot of undigested cRNA; M, size marker, HueIII-digested pBR322 (left) and A DNA cut with BsfEII. The cell lines analyzed were (C) HIT, (D) SN56, (E) NG108-15, (F) NS20Y, and (G) NS26.
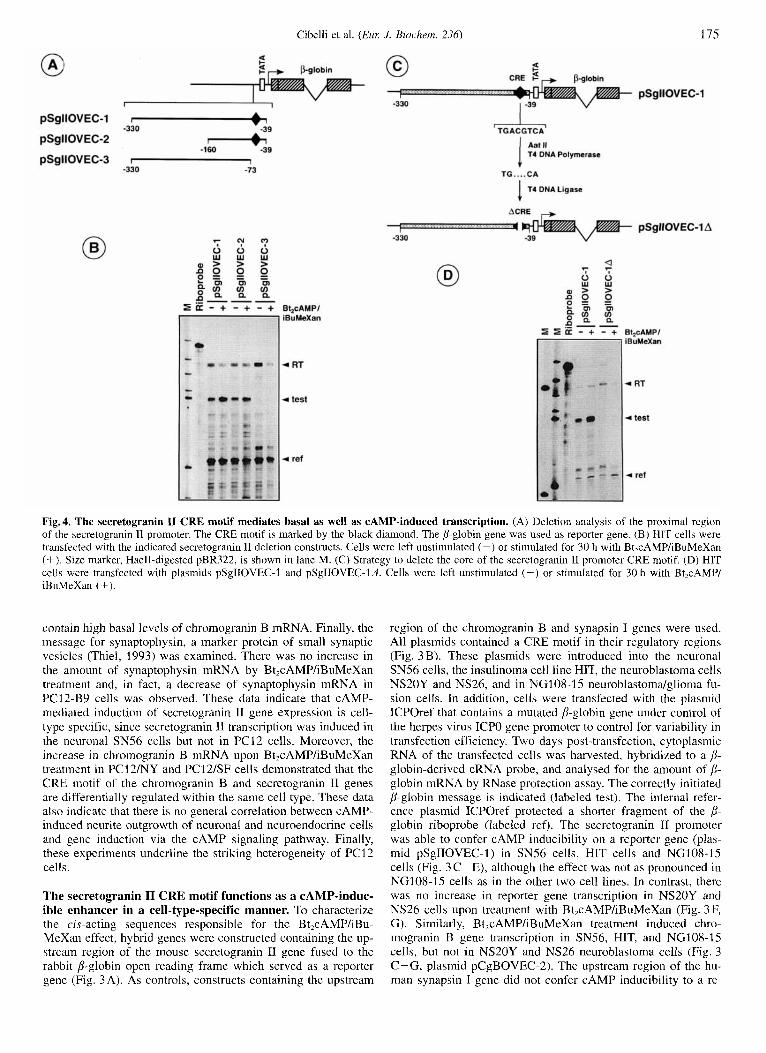
Cibelli et al. ( E m J. Biochern. 236) 175
Fig. 4. The secretogranin I1 CRE motif mediates basal as well as CAMP-induced transcription. (A) Deletion analysis of the proximal region of the secretogranin I1 promoter. The CRE motif is marked by the black diamond. The P-globin gene was used as reporter gene. (B) HIT cells were transfected with the indicated secretogranin I1 deletion constructs. Cells were left unstimulated (-) or stimulated for 30 h with Bt,cAMP/iBuMeXan (+). Size marker, HaeII-digested pBR322, is shown in lane M. (C) Strategy to delete the core of the secretogranin I1 promoter CRE motif. (D) HIT cells were transfected with plasmids pSgIIOVEC-I and pSgIIOVEC-Id iBuMeXan (+).
contain high basal levels of chromogranin B mRNA. Finally, the message for synaptophysin, a marker protein of small synaptic vesicles (Thiel, 1993) was examined. There was no increase in the amount of synaptophysin mRNA by Bt,cAMP/iBuMeXan treatment and, in fact, a decrease of synaptophysin mRNA in PC12-B9 cells was observed. These data indicate that CAMP- mediated induction of secretogranin I1 gene expression is cell- type specific, since secretogranin 11 transcription was induced in the neuronal SN56 cells but not in PC12 cells. Moreover, the increase in chromogranin B mRNA upon Bt,cAMP/iBuMeXan treatment in PC12/NY and PC12/SF cells demonstrated that the CRE motif of the chromogranin B and secretogranin I1 genes are differentially regulated within the same cell type. These data also indicate that there is no general correlation between CAMP- induced neurite outgrowth of neuronal and neuroendocrine cells and gene induction via the cAMP signaling pathway. Finally, these experiments underline the striking heterogeneity of PC12 cells.
The secretogranin I1 CRE motif Functions as a CAMP-induc- ible enhancer in a cell-type-specific manner. To characterize the cis-acting sequences responsible for the Bt,cAMP/iBu- MeXan effect, hybrid genes were constructed containing the up- stream region of the mouse secretogranin I1 gene fused to the rabbit P-globin open reading frame which served as a reporter gene (Fig. 3 A). As controls, constructs containing the upstream
Cells were left unstimulated (-) or stimulated for 30 h with Bt,cAMP/
region of the chromogranin B and synapsin I genes were used. All plasmids contained a CRE motif in their regulatory regions (Fig. 3B). These plasmids were introduced into the neuronal SNS6 cells, the insulinoma cell line HIT, the neuroblastoma cells NS20Y and NS26, and in NG108-15 neuroblastomdglioma fu- sion cells. In addition, cells were transfected with the plasmid ICPOref that contains a mutated p-globin gene under control of the herpes virus ICPO gene promoter to control for variability in transfection efficiency. Two days post-transfection, cytoplasmic RNA of the transfected cells was harvested, hybridized to a B- globin-derived cRNA probe, and analysed for the amount of 8- globin mRNA by RNase protection assay. The correctly initiated P-globin message is indicated (labeled test). The internal refer- ence plasmid ICPOref protected a shorter fragment of the 8- globin riboprobe (labeled ref). The secretogranin I1 promoter was able to confer CAMP inducibility on a reporter gene (plas- mid pSgTIOVEC-1) in SN56 cells, HIT cells and NG108-15 cells (Fig. 3C-E), although the effect was not as pronounced in NG108-15 cells as in the other two cell lines. In contrast, there was no increase in reporter gene transcription in NS20Y and NS26 cells upon treatment with Bt,cAMP/iBuMeXan (Fig. 3 F, G). Similarly, Bt,cAMP/iBuMeXan treatment induced chro- mogranin B gene transcription in SN56, HIT, and NG108-15 cells, but not in NS20Y and NS26 neuroblastoma cells (Fig. 3 C-G, plasmid pCgBOVEC-2). The upstream region of the hu- man synapsin I gene did not confer cAMP inducibility to a re-
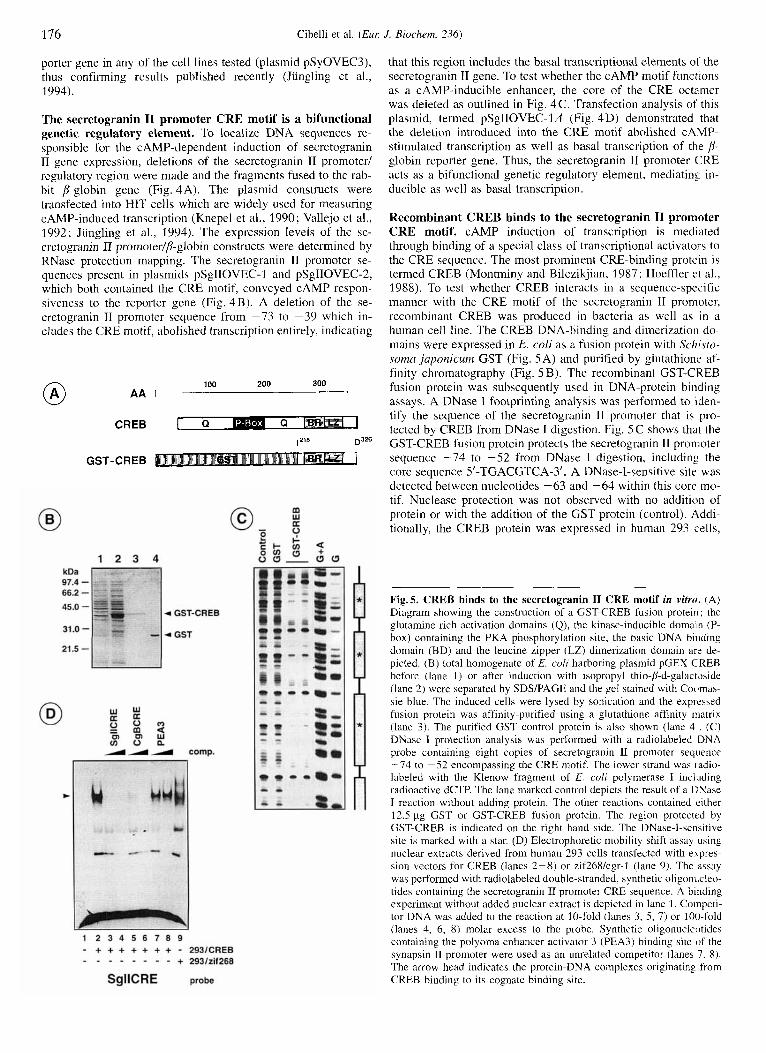
176 Cibelli et al. ( E m J. Biochem. 236)
porter gene in any of the cell lines tested (plasmid pSyOVEC3), thus confirming results published recently (Jungling et al., 1994).
The secretogranin I1 promoter CRE motif is a bifunctional genetic regulatory element. To localize DNA sequences re- sponsible for the CAMP-dependent induction of secretogranin I1 gene expression, deletions of the secretogranin I1 promoter/ regulatory region were made and the fragments fused to the rab- bit B-globin gene (Fig. 4A). The plasmid constructs were transfected into HIT cells which are widely used for measuring CAMP-induced transcription (Knepel et al., 1990; Vallejo et al., 1992; Jungling et al., 1994). The expression levels of the se- cretogranin I1 promoter/,!-globin constructs were determined by RNase protection mapping. The secretogranin 11 promoter se- quences present in plasmids pSgIIOVEC-1 and pSgIIOVEC-2, which both contained the CRE motif, conveyed cAMP respon- siveness to the reporter gene (Fig. 4B). A deletion of the se- cretogranin TI promoter sequence from -73 to -39 which in- cludes the CRE motif, abolished transcription entirely, indicating
100 200 300 . -, AA I
CREB [ Q Q l B R m 1 ,218 ~ 3 2 6
that this region includes the basal transcriptional elements of the secretogranin I1 gene. To test whether the cAMP motif functions as a CAMP-inducible enhancer, the core of the CRE octamer was deleted as outlined in Fig. 4C. Transfection analysis of this plasmid, termed pSgIIOVEC-ld (Fig. 4D) demonstrated that the deletion introduced into the CRE motif abolished CAMP- stimulated transcription as well as basal transcription of the p- globin reporter gene. Thus, the secretogranin I1 promoter (:RE acts as a bifunctional genetic regulatory element, mediating in- ducible as well as basal transcription.
Recombinant CREB binds to the secretogranin I1 promoter CRE motif. cAMP induction of transcription is mediated through binding of a special class of transcriptional activators to the CRE sequence. The most prominent CRE-binding protein is termed CREB (Montminy and Bilezikjian, 1987; Hoeffler et al., 1988). To test whether CREB interacts in a sequence-specific manner with the CRE motif of the secretogranin I1 promoter, recombinant CREB was produced in bacteria as well as in a human cell line. The CREB DNA-binding and dimerization do- mains were expressed in E. coli as a fusion protein with Schisto- soma japonicum GST (Fig. 5A) and purified by glutathione af- finity chromatography (Fig. 5 B). The recombinant GST-CKEB fusion protein was subsequently used in DNA-protein binding assays. A DNase I footprinting analysis was performed to iden- tify the sequence of the secretogranin I1 promoter that is pro- tected by CREB from DNase 1 digestion. Fig. 5 C shows that the GST-CREB fusion protein protects the secretogranin I1 pron-ioter sequence -74 to -52 from DNase I digestion, including the core sequence 5'-TCACGTCA-3'. A DNase-I-sensitive site was detected between nucleotides -63 and -64 within this core mo- tif. Nuclease protection was not observed with no addition of protein or with the addition of the CST protein (control). Addi- tionally, the CREB protein was expressed in human 293 cells,
Fig.5. CREB binds to the secretogranin I1 CRE motif in vitro. (A) Diagram showing the construction of a GST-CREB fusion protein ; the glutamine-rich activation domains (Q), the kinase-inducible domain (P- box) containing the PKA phosphorylation site, the basic DNA binding domain (BD) and the leucine zipper (LZ) dimerization domain arc de- picted. (B) total homogenate of E. coli harboring plasmid pGEX-CREB before (lane 1) or after induction with isopropyl thio-D-d-galactJside (lane 2) were separated by SDS/PAGE and the gel stained with Cocfmas- sie blue. The induced cells were lysed by sonication and the expressed fusion protein was affinity-purified using a glutathione affinity matrix (lane 3). The purified GST control protein is also shown (lane 4, . (C) DNase I protection analysis was performed with a radiolabeled DNA probe containing eight copies of secretogranin 11 promoter sequence -74 to -52 encompassing the CRE motif. The lower strand was radio- labeled with the Klenow fragment of E. coli polymerase I inchiding radioactive dCTP. The lane marked control depicts the result of a DNase I reaction without adding protein. The other reactions contained either 12.5 pg GST or GST-CREB fusion protein. The region protected by GST-CREB is indicated on the right hand side. The DNase-I-sensitive site is marked with a star. (D) Electrophoretic mobility shift assay using nuclear extracts derived from human 293 cells transfected with expres- sion vectors for CREB (lanes 2-8) or zif268/egr-l (lane 9). The assay was performed with radiolabeled double-stranded, synthetic oligonucleo- tides containing the secretogranin I1 promoter CRE sequence. A binding experiment without added nuclear extract is depicted in lane 1. Competi- tor DNA was added to the reaction at 10-fold (lanes 3, 5 , 7) or 100-fold (lanes 4, 6, 8) molar excess to the probe. Synthetic oligonucleotides containing the polyoma enhancer activator 3 (PEA3) binding site of the synapsin I1 promoter were used as an unrelated competitor (lanes 7, 8). The arrow head indicates the protein-DNA complexes originating from CREB binding to its cognate binding site.
Cibelli et al. ( E m J. Biochem. 236)
@ Reporter construct
177
5 x GAL40VEC
@ Expression constructs
pM1 1 147
pGAL4-CREB CREB 1
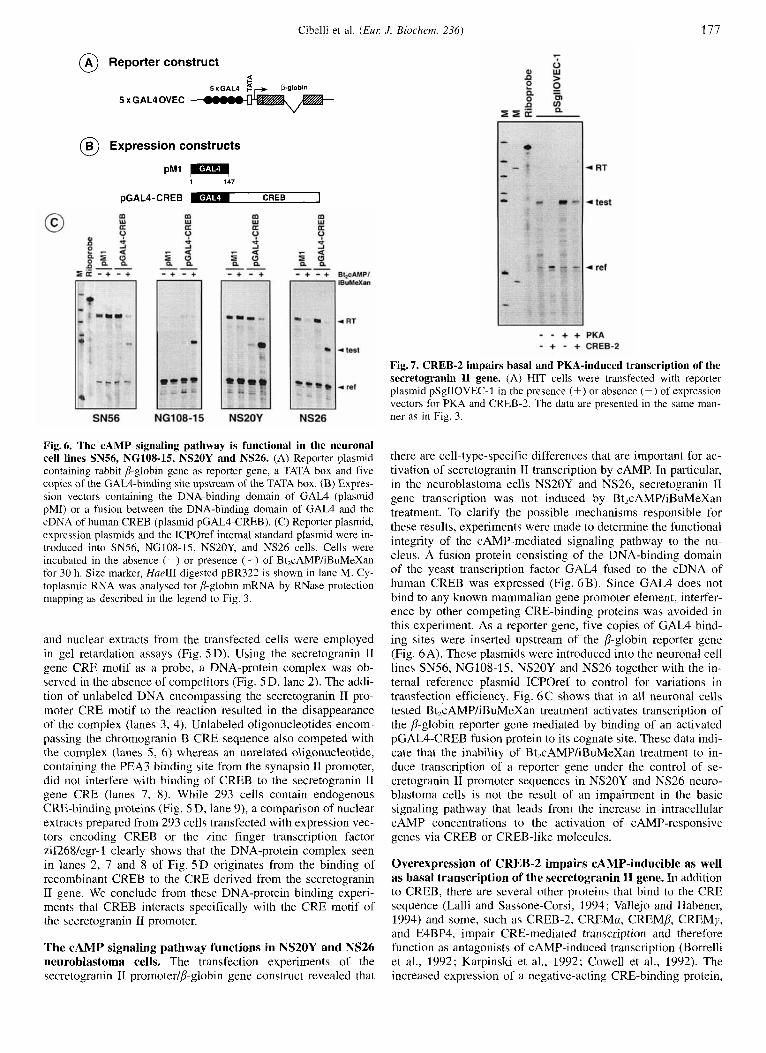
Fig. 7. CREB-2 impairs basal and PKA-induced transcription of the secretogranin I1 gene. (A) HIT cells were transfected with reporter plasmid pSgIIOVEC-1 in the presence (+) or absence (-) of expression vectors for PKA and CREB-2. The data are presented in the same man- ner as in Fig. 3.
Fig. 6. The cAMP signaling pathway is functional in the neuronal cell lines SN56, NGlOS-19, NS20Y and NS26. (A) Reporter plasmid containing rabbit jj-globin gene as reporter gene, a TATA box and five copies of the GAL4-binding site upstream of the TATA box. (B) Expres- sion vectors containing the DNA-binding domain of GAL4 (plasmid pMI) or a fusion between the DNA-binding domain of GAL4 and the cDNA of human CREB (plasmid pGAL4-CREB). (C) Reporter plasmid, expression plasmids and the ICPOref internal standard plasmid were in- troduced into SN56, NG108-15, NS20Y, and NS26 cells. Cells were incubated in the absence (-) or presence (+) of Bt,cAMP/iBuMeXan for 30 h. Size marker, HueIII-digested pBR322 is shown in lane M. Cy- toplasmic RNA was analysed for jj-globin mRNA by RNase protection mapping as described in the legend to Fig. 3.
and nuclear extracts from the transfected cells were employed in gel retardation assays (Fig. 5 D). Using the secretogranin TI gene CRE motif as a probe, a DNA-protein complex was ob- served in the absence of competitors (Fig. 5 D, lane 2) . The addi- tion of unlabeled DNA encompassing the secretogranin I1 pro- moter CRE motif to the reaction resulted in the disappearance of the complex (lanes 3 , 4). Unlabeled oligonucleotides encom- passing the chromogranin B CRE sequence also competed with the complex (lanes 5 , 6) whereas an unrelated oligonucleotide, containing the PEA3 binding site from the synapsin I1 promoter, did not interfere with binding of CREB to the secretogranin I1 gene CRE (lanes 7, 8). While 293 cells contain endogenous CRE-binding proteins (Fig. 5 D, lane 9), a comparison of nuclear extracts prepared from 293 cells transfected with expression vec- tors encoding CREB or the zinc finger transcription factor zif26Wegr-1 clearly shows that the DNA-protein complex seen in lanes 2, 7 and 8 of Fig. 5D originates from the binding of recombinant CREB to the CRE derived from the secretogranin I1 gene. We conclude from these DNA-protein binding experi- ments that CREB interacts specifically with the CRE motif of the secretogranin 11 promoter.
The cAMP signaling pathway functions in NS20Y and NS26 neuroblastoma cells. The transfection experiments of the cecretogranin I1 promoterlp-globin gene construct revealed that
there are cell-type-specific differences that are important for ac- tivation of secretogranin I1 transcription by CAMP. In particular, in the neuroblastoma cells NS20Y and NS26, secretogranin 11 gene transcription was not induced by Bt,cAMP/iBuMeXan treatment. To clarify the possible mechanisms responsible for these results, experiments were made to determine the functional integrity of the CAMP-mediated signaling pathway to the nu- cleus. A fusion protein consisting of the DNA-binding domain of the yeast transcription factor GAL4 fused to the cDNA of human CREB was expressed (Fig. 6B). Since GAL4 does not bind to any known mammalian gene promoter element, interfer- ence by other competing CRE-binding proteins was avoided in this experiment. As a reporter gene, five copies of GAL4 bind- ing sites were inserted upstream of the p-globin reporter gene (Fig. 6A). These plasmids were introduced into the neuronal cell lines SN56, NG108-15, NS20Y and NS26 together with the in- ternal reference plasmid ICPOref to control for variations in transfection efficiency. Fig. 6C shows that in all neuronal cells tested Bt,cAMP/iBuMeXan treatment activates transcription of the 8-globin reporter gene mediated by binding of an activated pGAL4-CREB fusion protein to its cognate site. These data indi- cate that the inability of Bt,cAMP/iBuMeXan treatment to in- duce transcription of a reporter gene under the control of se- cretogranin I1 promoter sequences in NS20Y and NS26 neuro- blastoma cells is not the result of an impairment in the basic signaling pathway that leads from the increase in intracellular CAMP concentrations to the activation of CAMP-responsive genes via CREB or CREB-like molecules.
Overexpression of CREB-2 impairs CAMP-inducible as well as basal transcription of the secretogranin I1 gene. In addition to CREB, there are several other proteins that bind to the CRE sequence (Lalli and Sassone-Corsi, 1994; Vallejo and Habener, 1994) and some, such as CREB-2, CREMa, CREMP, CREMy, and E4BP4, impair CRE-mediated transcription and therefore function as antagonists of CAMP-induced transcription (Borrelli et al., 1992; Karpinski et al., 1992; Cowell et al., 1992). The increased expression of a negative-acting CRE-binding protein,

178 Cibelli et al. ( E m J. Biochem. 236)
such as CREB-2, should inhibit transcription of CREB-depen- dent genes through competition for the CRE-binding site with positive-acting CRE-binding proteins such as CREB. To test this hypothesis, plasmid pSglIOVEC-1 was transfected into HIT cells together with expression vectors for PKA and CREB-2 (Fig. 7). The data show that the overexpressed CREB-2 protein completely inhibits the basal transcription activity of this con- struct. In addition, CREB-2 overexpression impaired PKA-in- duced transcription, since coexpression of PKA and CREB-2 resulted in a transcriptional signal similar or lower than that in the control experiment.
DISCUSSION
In the present study the function of the CRE motif in the secretogranin II promoter has been analysed. The secretogranin I1 promoter CRE sequence was shown to function as a CAMP- inducible enhancer in a neuronal cell line derived from the sep- tum (SN56), a fusion cell line originated from a neuroblastoma and a glioma cell (NG108-15), and an insulinoma cell line (HIT). Transcription was directly monitored by an analysis of the RNA transcripts of the /I-globin reporter gene. Deletion of the CRE motif abolished the effect of cAMP upon transcription, thus indicating that this DNA sequence is crucial for CAMP- mediated transcription induction. Through gel retardation and DNase I protection assays, the CRE binding protein CREB was demonstrated to interact in a sequence-specific manner with the secretogranin I1 promoter CRE in vitro. Moreover, experiments using deletion mutants of the secretogranin I1 promoter as well as overexpression of inhibitory CREB-2 transcription factor, re- vealed that the secretogranin II CRE is a bifunctional genetic regulatory element in that it mediates basal and CAMP-inducible transcription. Similar observations have been reported for the chromogranin B gene (Jiingling et al., 1994).
The fact that the CRE motif in the secretogranin I1 gene functions as a CAMP-inducible enhancer element in some, but not all, cell types tested indicates that cell-type-specific mecha- nisms play a role in CRE-mediated transcriptional regulation. Similar observations have been reported for the somatostatin gene, the prototype of CAMP-regulated transcriptional induction. While CAMP activates somatostatin gene transcription in PC12 cells and primary diencephalic cultures (Montminy et al., 1986; Montminy and Bilezikjian, 1987), it was unable to stimulate so- matostatin gene transcription in the somatostatin-producing rat insulinoma cell line RIN-1027-B2 (Vallejo et al., 199.5). This fact could be explained by the presence of a heat-stable inhibitor of PKA in these cells that prevented activation of PKA and sub- sequently' CREB phosphorylation (Vallejo et al., 1995). Studies were made to determine whether PKA-mediated transcription is impaired in neuroblastoma cells, which did not show a CAMP- induction of secretogranin 11 transcription. Using pGAL4- CREB, an expression vector encoding a fusion protein contain- ing the DNA binding domain of GAL4 fused to the CREB cDNA, CAMP-induced transcription was demonstrated in the neuroblastoma cells NS20Y and NS26 and the cell lines NG108- 1.5 and SN.56. Thus, it was shown that the CAMP-mediated path- way of gene transcription via CREB phosphorylation is intact in these cells and that an impairment of this pathway is not the reason for the inability of cAMP analogues to induce transcrip- tion of secretogranin in neuroblastoma cells.
It has been reported that undifferentiated F9 embryonic car- cinoma cells are refractory to cAMP induction of the CRE-con- raining somatostatin and vasoactive intestinal peptide genes (Masson et al., 1992). This was explained 'by the presence of negative regulators that block the activity of endogenous tran-
scriptional activators of the cAMP signaling pathway. Mean- while, several negative regulators of CAMP-regulated transcrip- tion have been discovered. The proteins CREMa, CREMP, and CREMy bind to the CRE sequence with a similar efficiency and specificity as CREB, however they act as down-regulator, of CAMP-mediated transcription (Foulkes et al., 1991 ; for rel'iew, see Lalli and Sassone-Corsi, 1994) by either competing with CREB for DNA binding or by forming inactive heterodiraers with CREB. Efficient repression, however, requires DNA bind- ing, thereby inhibiting the DNA binding of the transcriptional activators such as CREB (Laoide et al., 1993). It has been shown that different CREM isoforms, including both activator and re- pressor proteins, are coexpressed in a large number of cell types (Laoide et al., 1993; Mellstrom et al., 1993), suggesting that the combination of CREM isoforms in a particular cell type is a critical factor for the response to CAMP.
Another negative regulator of CAMP-mediated transcription is the basic region/leucine zipper protein CREB-2 (Karpinski et al., 1992). CREB-2 does not heterodimerize with CREB (VaUejo and Habener, 1994); thus, the mechanism for impairing CAMP- mediated gene transcription is most likely executed via displace- ment of CREB from its binding site through competitive DNA binding. Data presented here show that overexpressed CREB-2 strikingly impaired the basal and CAMP-mediated transcription of the secretogranin I1 gene in HIT cells, suggesting that CREB- 2 competes with CRE-bound transcriptional activators. This ex- periment indicates that a disturbance of the balance of transzrip- tional activators and repressors can have significant effects upon CAMP-stimulated transcription.
Additionally, the data presented here demonstrate the signifi- cance of the promoter context on the function of a particular CRE element. Most strikingly, the CRE in the synapsin I pro- moter does not confer CAMP inducibility in all cell lines tested, although this element is identical with the CRE motif in the secretogranin I1 or chromogranin B gene. Moreover. while the secretogranin I1 and chromogranin B genes respond similarly to increased intracellular cAMP concentration in many cell types, only the chromogranin B gene is induced in PC12 cells by Bt,cAMP/iBuMeXan treatment. The specific promoter context of a given CRE thus allows for an additional measure of cotn- plexity in the transcriptional response to stimulation by CAMP.
The finding that secretogranin II transcription is induced by cAMP in a variety of neuronal and neuroendocrine cells indi- cates that regulation of the secretogranin I1 gene is part of the physiological signaling pathways mediated via cAMP in the ner- vous system. It will be of interest to determine the physiological stimuli regulating secretogranin TI transcription in different pop- ulations of neurons in order to more fully understand the regula- tion of the secretogranin II gene as well as the mechanism of the cAMP signaling pathway in neurons.
We are grateful to Thomas C. Sudhof, Wieland B. Huttner and Hans- Dieter Soling for their support; we thank W. Knepel, P. Mane\s, M. Nirenberg, I. Sadowski, E. Schaeffer, H. E. Varmus, B. H. Wainer. and K. Willert for plasmids and cell lines. We thank Matthias Cramrr and Piera Cicchetti for suggestions and critical reading of the manuscript and Udo Ringeisen for preparing the figures. This work was supported by grants from the Deiitsche Fnrschuizgsgemeinschaft through SFB 274 and the Bayer AG (Bayer International Investigatorship) to G. T., the Euro- pean Union (Human Capital and Mobility Program) to G. C. arid the Deutsche For.schungs~emei~i.scl~aft to H.-H. G. (Ge 317lC-7).
REFERENCES Borrelli, E., Montmayeur, J.-P., Foulkes, N. S. & Sassone-Corsi, P.
(1992) Signal transduction and gene control : the cAMP pathway, Crif. Rev. Oncogenesis 3, 322 -338.

Cibelli et al. (EUK J. Biochem. 236) 179
Cowell, I. G., Skinner, A. & Hurst, H. C. (1992) Transcriptional repres- sion by a novel member of the bZIP family of transcription factors, Mol. Cell. Biol. 12, 3070-3077.
Cox, M. E. & Maness, P. F. (1991) Neurite extension and protein tyro- sine phosphorylation elicited by inducible expression of the v-src oncogene in a PC12 cell line, Exp. Cell. Res. 195, 423-431.
Fischer-Colbrie, R., Gutierrez, J., Hsu, C.-M., Iacangelo, A. & Eiden, L. E. (1990) Sequence analysis, tissue distribution and regulation by cell depolarizationn, and second messengers of bovine secretogranin IT (chromogranin C) mRNA, J. B i d . Chem. 265, 9208-9213.
Foulkes, N. S., Borelli, E. & Sassone-Corsi, P. (1991) CREM gene: use of alternative DNA-binding domains generates multiple antagonists of CAMP-induced transcription, Cell 64, 739-749.
Gerdes, H.-H., Phillips, E. & Huttner, W. B. (1988) The primary struc- ture of rat secretogranin TI deduced from a cDNA clone, Nucleic A&ls Res. 16, 11 811.
Hoeffler, J. P., Meyer, T. E., Yun, Y., Jameson, J. L. & Habener, J. F. (1988) Cyclic AMP- responsive DNA-binding proteins : structure based on a cloned placental cDNA, Science 242, 1430-1433.
Huttner, W. B. & Natori, S. (1995) Helper proteins for neuroendocrine secretion, Curt Biol. 5, 242-245.
Jiingling, S., Cibelli, G., Czardybon, M., Gerdes, H.-H. & Thiel, G. (1994) Differential regulation of chromogranin B and synapsin I gene promoter activity by cAMP and CAMP-dependent protein ki- nase, Eur J. Biochem. 226, 925-935.
Karpinski, B. A., Morle, G. D., Huggenvik, J., Uhler, M. D. & Leiden, J. M. (1992) Molecular cloning of human CREB-2: an ATF/CREB transcription factor that can negatively regulate transcription from the cAMP response element, Proc. Natl Acad. Sci. USA 89, 4820- 4824.
Kirchmair, R., Benzer, A., Troger, J., Miller, C., Marksteiner, J., Saria, A., Gasser, R. W., Hogue-Angeletti, R., Fischer-Colbrie, R. & Wink- ler. H. (1 994) Molecular characterization of immunoreactivities of peptides derived from chromogranin A (GE-25) and from secreto- granin I1 (secretoneurin) in human and bovine cerebrospinal fluid, Neuroscience 63, 1179-1187.
Knepel, W., Chafitz, J. & Habener, J . F. (1990) Transcriptional activation of the rat glucagon gene by the cyclic AMP-responsive element in pancreatic islet cells, Mol. Cell. B id . 10, 6799-6804.
Lalli, E. & Sassone-Corsi, P. (1994) Signal transduction and gene regula- tion: the nuclear response to CAMP, J. Biol. Chem. 269, 17359- 17 362.
Laoide, B. M., Foulkes, N. S., Schlotter, F. & Sassone-Corsi, P. (1993) The functional versatility of CREM is determined by its modular structure, EMBO J . 12, 1179-1191.
Lee, H. J., Hammond, D. N., Large, T. H. & Wainer, B. H. (1990) Im- mortalized young adult neurons from the septa1 region : generation and characterization, Dev. Brain Res. 52, 219-228.
Masson, N., Ellis, M., Goodbourn, S. & Lee, K. A. W. (1992) Cyclic AMP response element-binding protein and the catalytic subunit of protein kinase A are present in F9 embryonal carcinoma cells but are unable to activate the somatostatin promoter, Mol. Cell. B i d . 12,
Mayer, B. J., Jackson, P. K., Van Etten, R. A. & Baltimore, D. (1992) Point mutations in the ah1 SH2 domain coordinately impair phospho- tyrosine binding in vitro and transforming activity in vivo, Mol. Cell. B i d . 12, 609-618.
Mellstriim, B., Naranjo, J. R., Foulkes, N. S., Lafarga, M. & Sassone- Corsi, P. (1993) Transcriptional response to CAMP in brain: specific distribution and induction of CREM antagonists, Neurun 10, 655- 665.
Meyer, T. E. & Habener, J. F. (1993) Cyclic adenosine 3’,5’-monophos- phate response element binding protein (CREB) and related tran-
1096 - 1 106.
scription-activating deoxyribonncleic acid binding proteins, En- docrine Rev. 14, 269-290.
Montminy, M. R. & Bilezikjian, L. M. (1987) Binding of a nuclear pro- tein to the cyclic-AMP response element of the somatostatin gene, Nature 328, 175- 178.
Montminy, M. R., Low, M. J., Tapia-Arancibia, L., Reichlin, S., Mandel, G. & Goodman, R. H. (1986) Cyclic AMP regulates somatostatin mRNA accumulation in primary diencephalic cultures and in transfected fibroblast cells, J. Neurosci. 6 , 1171 - 1176.
Rosa, P. & Gerdes, H.-H. (1994) The granin protein family: Markers for neuroendocrine cells and tools for the diagnosis of neuroendocrine tumors, J. Endocrinol. Invest. 17, 207-225.
Sadowski, I., Bell, B., Broad, P. & Hollis, M. (1992) GAL4 fusion vec- tors for expression in yeast or mammalian cells. Gene 118, 137- 141.
Scammell, J. G. (1993) Granins - Marker of the regulated secretory pathway, Trends Endocrinul. Metabol. 4 , 14- 18.
Schimmel, A., Brawling, O., Ruther, U., Huttner, W. B. & Gerdes, H.- H. (1 992) The organization of the mouse secretogranin I1 gene, FEBS Lett. 314, 375-380.
Shackleford, G. M., Willert, K., Wang, J. & Varmus, H. E. (1993) The Wnt-1 proto- oncogene induces changes in morphology, gene expres- sion, and growth factor responsiveness in PC12 cells, Neuron I / ,
Smith, D. B. & Johnson, K. S. (1988) Single-step purification of poly- peptides expressed in Escherichia coli as fusions with glutathione S- transferase, Gene 67, 31 -40.
Thiel, G., Greengard, P. & Siidhof, T. C. (1991) Characterization of tis- sue-specific transcription by the human synapsin I gene promoter, Proc. Natl Acad. Sci. USA 88, 3431 -3435.
Thiel, G. (1993) Synapsin I, synapsin 11, and synaptophysin: marker proteins of synaptic vesicles, Brain Pathol. 3 , 87-95.
Thiel, G., Schoch, S. & Petersohn, D. (1994) Regulation of synapsin I gene expression by the zinc finger transcription factor zif268/egr-1, J . Biol. Chem. 269, IS 294- 15 301.
Thompson, M. E., Zimmer, W. E., Wear, L. B., MacMillan, L. A,, Thompson, W. J., Huttner, W. B., Hidaka, H. & Scammell, J. G. (1992) Differential regulation of chromogranin Bkecretogranin I and secretogranin I1 by forskolin in PC12 cells, ,4401. Bruin Res.12. 195- 202.
Thompson, M. E., Valentine, D. L., Strada, S. J., Wagner, J. A. & Scam- mell, J. G. (1994) Transcriptional regulation of secretogranin I1 and chromogranin B by cyclic AMP in a rat pheochromocytoma cell line, Mol. Pharmacol. 46, 880-889.
Vallejo, M., Miller, C. P. & Habener, J. F. (1992) Somatostatin gene transcription regulated by a bipartite pancreatic islet-D-cell-specific enhancer coupled synergetically to a CAMP response element, J. Biol. Chem. 267, 12868-12875.
Vallejo, M. & Habener, J. F. (1994) Mechanisms of transcriptional regu- lation by CAMP, in Transcription: mechanisms and regulation (Con- away, R. C. & Conaway, J. W., eds) pp. 353-368, Raven Press. New York.
Vallejo, M., Gosse, M. E., Beckman, W. & Habener, J. F. (1995) Im- paired cyclic AMP-dependent phosphorylation renders CREB a re- pressor of C/EBP-induced transcription of the somatostatin gene in an insulinoma cell line, Mol. Cell. Biol. 15, 415-424.
Weiler, R., Marksteiner, J., Bellmann, R., Wohlfarter, T., Schober, M., Fischer-Colbrie, R., Sperk, G. & Winkler, H. (1990) Chromogranins in rat brain: characterization, topographical distribution and regula- tion of synthesis, Brain Res. 532, 87-94.
865 -875.
Top Related
Copyright © 2022 FDOKUMEN

