







Bahasa
Halaman
Hukum


Elsevier Editorial System(tm) for The Journal of Steroid Biochemistry and Molecular Biology Manuscript Draft Manuscript Number SBMB-D-13-00199R1 Title Gonadal development and transcript profiling of steroidogenic enzymes in response to 17α-methyltestosterone in the rare minnow Gobiocypris rarus Article Type Full Length Article Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology Gobiocypris rarus Corresponding Author Dr Zaizhao Wang Corresponding Authors Institution Northwest AampF University First Author Shaozhen Liu Order of Authors Shaozhen Liu Lihong Wang Fang Qin Yao Zheng Meng Li Yingying Zhang Cong Yuan Zaizhao Wang Abstract It is well known that natural and anthropogenic chemicals interfere with the hormonal system of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis remains to be determined The main objective of this study was to evaluate the effects of 17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of steroidogenic enzymes with effects on higher levels of biological organization and on VTG one remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes in a time- and concentration-dependent manner The hepatic vtg mRNA level was highly inhibited in the females and slightly induced by MT Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic genes in MT-treated groups were much distinct with the EE2-treated group Suggested Reviewers Paola Irato paolairatounipdit Montserrat Soleacute msoleicmcsices
Response to Reviewers
Dear editor
Thank you very much for your letter and the comments from the reviewers about our paper
submitted to Steroid Biochemistry amp Molecular Biology (Ms Ref No SBMB-D-13-00199)
We have checked and revised the manuscript carefully according to the comments We submit here
the revised manuscript as well as the responses to reviews
If you need any other information please contact me immediately by email My email account is
zzwangnwsuafeducn and Tel is +86-29-87092139 and Fax is +86-29-87092164
Sincerely yours
Zaizhao Wang
Response to reviewer
Dear reviewer
Thanks a lot for your comments on our manuscript According to those helpful comments we have
revised this manuscript carefully point-by-point The comments need to be explained are listed as
following
1 General comment Despite the amount of results in the current paper the research
hypothesis on the effects of the synthetic androgen 17-MT in terms of histological and
steroidogenic gene expressions has been largely demonstrated in many fish species So in
the introductory section of the present study the authors should better justify the reasons
for testing low levels of MT and EE as a further confirmation of the known responses to
Detailed Response to Reviewers
MT exposure in rare minnow in the ecological context of their region (case study)
Response Thank you very much for your suggestion In the present study we detected the
effect of MT (25 50 and 100 ngL) and EE2 (25 ngL) on rare minnow EE2 exposure group as a
control group appeared in the present study The aim of present study was to evaluate the effects
of MT at low concentrations (25-100ngL) on gene expression profiles of six key steroidogenic
genes There was no manuscript study the effects of MT at low concentrations on six key
steroidogenic genes simultaneously Our purpose was to study the mechanism of MT in gonads of
fish And someone else in our research group studied the effects of MT in brain or liver of rare
minnow
According your suggestion we plan to test the effect of MT with environment concentration
on Chinese freshwater cyprinid rare minnow
2 Introduction and discussion should be updated There are at least five references between
2010-2012 while there is rich and timely information on the MT and EE response in
various fish species
Response Thank you very much for your suggestion We indeed neglected the time of
references in the present study According your suggestion we carefully consulted a lot of
literature about MT and EE2 Meanwhile the introduction and discussion in the present study
have been updated
3 Pg2 L39-39 The hepatic vtg mRNA level was highly inhibited in the females and slightly
induced by MT What does mean slightly induced while only inhibition or suppression
by MT was observed for females May be this is stated for males but also a decrease was
observed for 21-d exposure
Response Thanks for your suggestion This error should be attributed to our careless We
have replaced the sentence with ldquoThe hepatic vtg mRNA level was highly inhibited in
females and slightly altered in males by MTrdquo (Pg2 L36 ) in the revised manuscript
4 Pg5-L107 The choice of testing low levels of MT (25-100ngL) and EE (25ngL) should be
justified in regard to the levels in the environment
Response Thanks for your suggestion The answer of this question is similar to the
question one Our choice of testing low levels of MT (25-100ngL and EE2 (25ngL) is
referred to previous study in zebrafish Error Reference source not found In the present
study other than detected the effect of MT and EE2 on vtg and steroidogenic enzymes we
attempted to make certain the metabolic pathway of steroid influenced by MT So we chose
these testing levels We will choose testing level in regard to that of environment in next
study
5 Pg7-L134-135 Half of the exposure solution was changed every day On what basis water
was changed every day while the frac12 time for the targeted hormonal compounds are longer
Response Thanks for your suggestion In the present study half of the exposure solution
was changed every day The main reason is fish excrement and remnant foods should be
cleared out timely When we cleared out the excrement and remnant foods part of exposure
solution would be taken away from the tanks To add water and hormonal compounds
expediently half of the exposure solution was changed every day
The frac12 time for the targeted hormonal compounds are longer than one day (Barel-Cohen
et al 2006 Homklin et al 2011)
6 P7-L137 All exposure experiments were conducted in triplicate separate tanks at the same
time How water quality was managed to avoid any false related among triplicates of the
same treatment or among treatments
Response Thanks for your suggestion In the present study the tap water in big tanks
were dechlorinated before used in exposure tests Water quality was often managed by
measuring physicochemical parameters such as temperature pH hardness dissolved oxygen
All the water in experiment from above-mentioned big tanks So the water quality should be
the same in our study
In the present study measurements of temperature and pH in exposure tanks were done
everyday to determine the quality of water further
7 Pg7-L146 pg10-L210 Through materials and methods the number of sampled fish and
related analyses is not well described in a triplicate tank 10 fish were chosen for histology
+ the number of fish sampled for RNA isolation and RT is not specified + 6 fish were
sampled for VTG and gonadal transcripts A line time with steps and types of samples is
recommended for more explanation of the sampling protocol
Response Thanks for your suggestion The number of fish sampled for RNA isolation and
RT is not specified in para 24 because in this section we aimed to introduce the sampling
protocol In this study the fish sampled for RNA isolation and RT consist of two independent
sections fish sampled for investigating tissue distribution of 11β-HSD2 (10 male fish and 10
female fish which were not exposure to MT and EE2) and for detecting mRNA expressions
of hepatic vtg and gonadal steroidogenic genes following MT and EE2 exposure (18 male fish
and 18 female fish control MT or EE2 groups) Thus the number of fish sampled for RNA
isolation and RT was divided into two sections and shown in para 28 (pg10 line 204-205)
and 29 (pg10 line 210-211) respectively
The line time with steps and types of samples as follows
a Mar 2012 Ten male and 10 female adult fish were sampled for investigating tissue
distribution of 11β-HSD2
b May 2012 For detecting mRNA expressions of hepatic vtg and gonadal steroidogenic
genes following MT and EE2 exposure we sampled 18 fish (6 per group in triplicate) in each
group (control EE2 and MT groups in males and females) in which 12 gonads of 10 fish
were chosen for histological examination
8 Pg10-L214 What was the statistical unit and model How the time effect was included in
the model
Response Thanks for your suggestion In the present study the statistical unit is Ct value
of each detected gene (StAR cyp11a1 cyp17a1 cyp19a1a 3β-HSD and 11β-HSD) Statistical
differences were first tested with two-way ANOVA with treatment and time as two factors
Treatment was further analyzed with one-way ANOVA followed by LSD multiple comparison
(Pg7-L194-197)
As there is no way to rule out the effects of growth from day 7 to day 21 on the mRNA
expression of the detected genes the data of two-way ANOVA were not put in our manuscript We
added the sentence bdquoIn the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and
cyp19a1a the interaction of treatment and time was statistically significant (P lt 005)‟ in the
revised manuscript (Pg18-L297-298) The two-way ANOVA analyzed results are as follows
Table 1 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in female G ruras exposed to control MT and EE2 a
a Gene expression is expressed as the mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
Table 2 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in male G ruras exposed to control MT and EE2 a
a Gene expression is expressed as mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

Response to Reviewers
Dear editor
Thank you very much for your letter and the comments from the reviewers about our paper
submitted to Steroid Biochemistry amp Molecular Biology (Ms Ref No SBMB-D-13-00199)
We have checked and revised the manuscript carefully according to the comments We submit here
the revised manuscript as well as the responses to reviews
If you need any other information please contact me immediately by email My email account is
zzwangnwsuafeducn and Tel is +86-29-87092139 and Fax is +86-29-87092164
Sincerely yours
Zaizhao Wang
Response to reviewer
Dear reviewer
Thanks a lot for your comments on our manuscript According to those helpful comments we have
revised this manuscript carefully point-by-point The comments need to be explained are listed as
following
1 General comment Despite the amount of results in the current paper the research
hypothesis on the effects of the synthetic androgen 17-MT in terms of histological and
steroidogenic gene expressions has been largely demonstrated in many fish species So in
the introductory section of the present study the authors should better justify the reasons
for testing low levels of MT and EE as a further confirmation of the known responses to
Detailed Response to Reviewers
MT exposure in rare minnow in the ecological context of their region (case study)
Response Thank you very much for your suggestion In the present study we detected the
effect of MT (25 50 and 100 ngL) and EE2 (25 ngL) on rare minnow EE2 exposure group as a
control group appeared in the present study The aim of present study was to evaluate the effects
of MT at low concentrations (25-100ngL) on gene expression profiles of six key steroidogenic
genes There was no manuscript study the effects of MT at low concentrations on six key
steroidogenic genes simultaneously Our purpose was to study the mechanism of MT in gonads of
fish And someone else in our research group studied the effects of MT in brain or liver of rare
minnow
According your suggestion we plan to test the effect of MT with environment concentration
on Chinese freshwater cyprinid rare minnow
2 Introduction and discussion should be updated There are at least five references between
2010-2012 while there is rich and timely information on the MT and EE response in
various fish species
Response Thank you very much for your suggestion We indeed neglected the time of
references in the present study According your suggestion we carefully consulted a lot of
literature about MT and EE2 Meanwhile the introduction and discussion in the present study
have been updated
3 Pg2 L39-39 The hepatic vtg mRNA level was highly inhibited in the females and slightly
induced by MT What does mean slightly induced while only inhibition or suppression
by MT was observed for females May be this is stated for males but also a decrease was
observed for 21-d exposure
Response Thanks for your suggestion This error should be attributed to our careless We
have replaced the sentence with ldquoThe hepatic vtg mRNA level was highly inhibited in
females and slightly altered in males by MTrdquo (Pg2 L36 ) in the revised manuscript
4 Pg5-L107 The choice of testing low levels of MT (25-100ngL) and EE (25ngL) should be
justified in regard to the levels in the environment
Response Thanks for your suggestion The answer of this question is similar to the
question one Our choice of testing low levels of MT (25-100ngL and EE2 (25ngL) is
referred to previous study in zebrafish Error Reference source not found In the present
study other than detected the effect of MT and EE2 on vtg and steroidogenic enzymes we
attempted to make certain the metabolic pathway of steroid influenced by MT So we chose
these testing levels We will choose testing level in regard to that of environment in next
study
5 Pg7-L134-135 Half of the exposure solution was changed every day On what basis water
was changed every day while the frac12 time for the targeted hormonal compounds are longer
Response Thanks for your suggestion In the present study half of the exposure solution
was changed every day The main reason is fish excrement and remnant foods should be
cleared out timely When we cleared out the excrement and remnant foods part of exposure
solution would be taken away from the tanks To add water and hormonal compounds
expediently half of the exposure solution was changed every day
The frac12 time for the targeted hormonal compounds are longer than one day (Barel-Cohen
et al 2006 Homklin et al 2011)
6 P7-L137 All exposure experiments were conducted in triplicate separate tanks at the same
time How water quality was managed to avoid any false related among triplicates of the
same treatment or among treatments
Response Thanks for your suggestion In the present study the tap water in big tanks
were dechlorinated before used in exposure tests Water quality was often managed by
measuring physicochemical parameters such as temperature pH hardness dissolved oxygen
All the water in experiment from above-mentioned big tanks So the water quality should be
the same in our study
In the present study measurements of temperature and pH in exposure tanks were done
everyday to determine the quality of water further
7 Pg7-L146 pg10-L210 Through materials and methods the number of sampled fish and
related analyses is not well described in a triplicate tank 10 fish were chosen for histology
+ the number of fish sampled for RNA isolation and RT is not specified + 6 fish were
sampled for VTG and gonadal transcripts A line time with steps and types of samples is
recommended for more explanation of the sampling protocol
Response Thanks for your suggestion The number of fish sampled for RNA isolation and
RT is not specified in para 24 because in this section we aimed to introduce the sampling
protocol In this study the fish sampled for RNA isolation and RT consist of two independent
sections fish sampled for investigating tissue distribution of 11β-HSD2 (10 male fish and 10
female fish which were not exposure to MT and EE2) and for detecting mRNA expressions
of hepatic vtg and gonadal steroidogenic genes following MT and EE2 exposure (18 male fish
and 18 female fish control MT or EE2 groups) Thus the number of fish sampled for RNA
isolation and RT was divided into two sections and shown in para 28 (pg10 line 204-205)
and 29 (pg10 line 210-211) respectively
The line time with steps and types of samples as follows
a Mar 2012 Ten male and 10 female adult fish were sampled for investigating tissue
distribution of 11β-HSD2
b May 2012 For detecting mRNA expressions of hepatic vtg and gonadal steroidogenic
genes following MT and EE2 exposure we sampled 18 fish (6 per group in triplicate) in each
group (control EE2 and MT groups in males and females) in which 12 gonads of 10 fish
were chosen for histological examination
8 Pg10-L214 What was the statistical unit and model How the time effect was included in
the model
Response Thanks for your suggestion In the present study the statistical unit is Ct value
of each detected gene (StAR cyp11a1 cyp17a1 cyp19a1a 3β-HSD and 11β-HSD) Statistical
differences were first tested with two-way ANOVA with treatment and time as two factors
Treatment was further analyzed with one-way ANOVA followed by LSD multiple comparison
(Pg7-L194-197)
As there is no way to rule out the effects of growth from day 7 to day 21 on the mRNA
expression of the detected genes the data of two-way ANOVA were not put in our manuscript We
added the sentence bdquoIn the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and
cyp19a1a the interaction of treatment and time was statistically significant (P lt 005)‟ in the
revised manuscript (Pg18-L297-298) The two-way ANOVA analyzed results are as follows
Table 1 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in female G ruras exposed to control MT and EE2 a
a Gene expression is expressed as the mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
Table 2 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in male G ruras exposed to control MT and EE2 a
a Gene expression is expressed as mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

MT exposure in rare minnow in the ecological context of their region (case study)
Response Thank you very much for your suggestion In the present study we detected the
effect of MT (25 50 and 100 ngL) and EE2 (25 ngL) on rare minnow EE2 exposure group as a
control group appeared in the present study The aim of present study was to evaluate the effects
of MT at low concentrations (25-100ngL) on gene expression profiles of six key steroidogenic
genes There was no manuscript study the effects of MT at low concentrations on six key
steroidogenic genes simultaneously Our purpose was to study the mechanism of MT in gonads of
fish And someone else in our research group studied the effects of MT in brain or liver of rare
minnow
According your suggestion we plan to test the effect of MT with environment concentration
on Chinese freshwater cyprinid rare minnow
2 Introduction and discussion should be updated There are at least five references between
2010-2012 while there is rich and timely information on the MT and EE response in
various fish species
Response Thank you very much for your suggestion We indeed neglected the time of
references in the present study According your suggestion we carefully consulted a lot of
literature about MT and EE2 Meanwhile the introduction and discussion in the present study
have been updated
3 Pg2 L39-39 The hepatic vtg mRNA level was highly inhibited in the females and slightly
induced by MT What does mean slightly induced while only inhibition or suppression
by MT was observed for females May be this is stated for males but also a decrease was
observed for 21-d exposure
Response Thanks for your suggestion This error should be attributed to our careless We
have replaced the sentence with ldquoThe hepatic vtg mRNA level was highly inhibited in
females and slightly altered in males by MTrdquo (Pg2 L36 ) in the revised manuscript
4 Pg5-L107 The choice of testing low levels of MT (25-100ngL) and EE (25ngL) should be
justified in regard to the levels in the environment
Response Thanks for your suggestion The answer of this question is similar to the
question one Our choice of testing low levels of MT (25-100ngL and EE2 (25ngL) is
referred to previous study in zebrafish Error Reference source not found In the present
study other than detected the effect of MT and EE2 on vtg and steroidogenic enzymes we
attempted to make certain the metabolic pathway of steroid influenced by MT So we chose
these testing levels We will choose testing level in regard to that of environment in next
study
5 Pg7-L134-135 Half of the exposure solution was changed every day On what basis water
was changed every day while the frac12 time for the targeted hormonal compounds are longer
Response Thanks for your suggestion In the present study half of the exposure solution
was changed every day The main reason is fish excrement and remnant foods should be
cleared out timely When we cleared out the excrement and remnant foods part of exposure
solution would be taken away from the tanks To add water and hormonal compounds
expediently half of the exposure solution was changed every day
The frac12 time for the targeted hormonal compounds are longer than one day (Barel-Cohen
et al 2006 Homklin et al 2011)
6 P7-L137 All exposure experiments were conducted in triplicate separate tanks at the same
time How water quality was managed to avoid any false related among triplicates of the
same treatment or among treatments
Response Thanks for your suggestion In the present study the tap water in big tanks
were dechlorinated before used in exposure tests Water quality was often managed by
measuring physicochemical parameters such as temperature pH hardness dissolved oxygen
All the water in experiment from above-mentioned big tanks So the water quality should be
the same in our study
In the present study measurements of temperature and pH in exposure tanks were done
everyday to determine the quality of water further
7 Pg7-L146 pg10-L210 Through materials and methods the number of sampled fish and
related analyses is not well described in a triplicate tank 10 fish were chosen for histology
+ the number of fish sampled for RNA isolation and RT is not specified + 6 fish were
sampled for VTG and gonadal transcripts A line time with steps and types of samples is
recommended for more explanation of the sampling protocol
Response Thanks for your suggestion The number of fish sampled for RNA isolation and
RT is not specified in para 24 because in this section we aimed to introduce the sampling
protocol In this study the fish sampled for RNA isolation and RT consist of two independent
sections fish sampled for investigating tissue distribution of 11β-HSD2 (10 male fish and 10
female fish which were not exposure to MT and EE2) and for detecting mRNA expressions
of hepatic vtg and gonadal steroidogenic genes following MT and EE2 exposure (18 male fish
and 18 female fish control MT or EE2 groups) Thus the number of fish sampled for RNA
isolation and RT was divided into two sections and shown in para 28 (pg10 line 204-205)
and 29 (pg10 line 210-211) respectively
The line time with steps and types of samples as follows
a Mar 2012 Ten male and 10 female adult fish were sampled for investigating tissue
distribution of 11β-HSD2
b May 2012 For detecting mRNA expressions of hepatic vtg and gonadal steroidogenic
genes following MT and EE2 exposure we sampled 18 fish (6 per group in triplicate) in each
group (control EE2 and MT groups in males and females) in which 12 gonads of 10 fish
were chosen for histological examination
8 Pg10-L214 What was the statistical unit and model How the time effect was included in
the model
Response Thanks for your suggestion In the present study the statistical unit is Ct value
of each detected gene (StAR cyp11a1 cyp17a1 cyp19a1a 3β-HSD and 11β-HSD) Statistical
differences were first tested with two-way ANOVA with treatment and time as two factors
Treatment was further analyzed with one-way ANOVA followed by LSD multiple comparison
(Pg7-L194-197)
As there is no way to rule out the effects of growth from day 7 to day 21 on the mRNA
expression of the detected genes the data of two-way ANOVA were not put in our manuscript We
added the sentence bdquoIn the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and
cyp19a1a the interaction of treatment and time was statistically significant (P lt 005)‟ in the
revised manuscript (Pg18-L297-298) The two-way ANOVA analyzed results are as follows
Table 1 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in female G ruras exposed to control MT and EE2 a
a Gene expression is expressed as the mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
Table 2 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in male G ruras exposed to control MT and EE2 a
a Gene expression is expressed as mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

by MT was observed for females May be this is stated for males but also a decrease was
observed for 21-d exposure
Response Thanks for your suggestion This error should be attributed to our careless We
have replaced the sentence with ldquoThe hepatic vtg mRNA level was highly inhibited in
females and slightly altered in males by MTrdquo (Pg2 L36 ) in the revised manuscript
4 Pg5-L107 The choice of testing low levels of MT (25-100ngL) and EE (25ngL) should be
justified in regard to the levels in the environment
Response Thanks for your suggestion The answer of this question is similar to the
question one Our choice of testing low levels of MT (25-100ngL and EE2 (25ngL) is
referred to previous study in zebrafish Error Reference source not found In the present
study other than detected the effect of MT and EE2 on vtg and steroidogenic enzymes we
attempted to make certain the metabolic pathway of steroid influenced by MT So we chose
these testing levels We will choose testing level in regard to that of environment in next
study
5 Pg7-L134-135 Half of the exposure solution was changed every day On what basis water
was changed every day while the frac12 time for the targeted hormonal compounds are longer
Response Thanks for your suggestion In the present study half of the exposure solution
was changed every day The main reason is fish excrement and remnant foods should be
cleared out timely When we cleared out the excrement and remnant foods part of exposure
solution would be taken away from the tanks To add water and hormonal compounds
expediently half of the exposure solution was changed every day
The frac12 time for the targeted hormonal compounds are longer than one day (Barel-Cohen
et al 2006 Homklin et al 2011)
6 P7-L137 All exposure experiments were conducted in triplicate separate tanks at the same
time How water quality was managed to avoid any false related among triplicates of the
same treatment or among treatments
Response Thanks for your suggestion In the present study the tap water in big tanks
were dechlorinated before used in exposure tests Water quality was often managed by
measuring physicochemical parameters such as temperature pH hardness dissolved oxygen
All the water in experiment from above-mentioned big tanks So the water quality should be
the same in our study
In the present study measurements of temperature and pH in exposure tanks were done
everyday to determine the quality of water further
7 Pg7-L146 pg10-L210 Through materials and methods the number of sampled fish and
related analyses is not well described in a triplicate tank 10 fish were chosen for histology
+ the number of fish sampled for RNA isolation and RT is not specified + 6 fish were
sampled for VTG and gonadal transcripts A line time with steps and types of samples is
recommended for more explanation of the sampling protocol
Response Thanks for your suggestion The number of fish sampled for RNA isolation and
RT is not specified in para 24 because in this section we aimed to introduce the sampling
protocol In this study the fish sampled for RNA isolation and RT consist of two independent
sections fish sampled for investigating tissue distribution of 11β-HSD2 (10 male fish and 10
female fish which were not exposure to MT and EE2) and for detecting mRNA expressions
of hepatic vtg and gonadal steroidogenic genes following MT and EE2 exposure (18 male fish
and 18 female fish control MT or EE2 groups) Thus the number of fish sampled for RNA
isolation and RT was divided into two sections and shown in para 28 (pg10 line 204-205)
and 29 (pg10 line 210-211) respectively
The line time with steps and types of samples as follows
a Mar 2012 Ten male and 10 female adult fish were sampled for investigating tissue
distribution of 11β-HSD2
b May 2012 For detecting mRNA expressions of hepatic vtg and gonadal steroidogenic
genes following MT and EE2 exposure we sampled 18 fish (6 per group in triplicate) in each
group (control EE2 and MT groups in males and females) in which 12 gonads of 10 fish
were chosen for histological examination
8 Pg10-L214 What was the statistical unit and model How the time effect was included in
the model
Response Thanks for your suggestion In the present study the statistical unit is Ct value
of each detected gene (StAR cyp11a1 cyp17a1 cyp19a1a 3β-HSD and 11β-HSD) Statistical
differences were first tested with two-way ANOVA with treatment and time as two factors
Treatment was further analyzed with one-way ANOVA followed by LSD multiple comparison
(Pg7-L194-197)
As there is no way to rule out the effects of growth from day 7 to day 21 on the mRNA
expression of the detected genes the data of two-way ANOVA were not put in our manuscript We
added the sentence bdquoIn the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and
cyp19a1a the interaction of treatment and time was statistically significant (P lt 005)‟ in the
revised manuscript (Pg18-L297-298) The two-way ANOVA analyzed results are as follows
Table 1 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in female G ruras exposed to control MT and EE2 a
a Gene expression is expressed as the mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
Table 2 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in male G ruras exposed to control MT and EE2 a
a Gene expression is expressed as mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

expediently half of the exposure solution was changed every day
The frac12 time for the targeted hormonal compounds are longer than one day (Barel-Cohen
et al 2006 Homklin et al 2011)
6 P7-L137 All exposure experiments were conducted in triplicate separate tanks at the same
time How water quality was managed to avoid any false related among triplicates of the
same treatment or among treatments
Response Thanks for your suggestion In the present study the tap water in big tanks
were dechlorinated before used in exposure tests Water quality was often managed by
measuring physicochemical parameters such as temperature pH hardness dissolved oxygen
All the water in experiment from above-mentioned big tanks So the water quality should be
the same in our study
In the present study measurements of temperature and pH in exposure tanks were done
everyday to determine the quality of water further
7 Pg7-L146 pg10-L210 Through materials and methods the number of sampled fish and
related analyses is not well described in a triplicate tank 10 fish were chosen for histology
+ the number of fish sampled for RNA isolation and RT is not specified + 6 fish were
sampled for VTG and gonadal transcripts A line time with steps and types of samples is
recommended for more explanation of the sampling protocol
Response Thanks for your suggestion The number of fish sampled for RNA isolation and
RT is not specified in para 24 because in this section we aimed to introduce the sampling
protocol In this study the fish sampled for RNA isolation and RT consist of two independent
sections fish sampled for investigating tissue distribution of 11β-HSD2 (10 male fish and 10
female fish which were not exposure to MT and EE2) and for detecting mRNA expressions
of hepatic vtg and gonadal steroidogenic genes following MT and EE2 exposure (18 male fish
and 18 female fish control MT or EE2 groups) Thus the number of fish sampled for RNA
isolation and RT was divided into two sections and shown in para 28 (pg10 line 204-205)
and 29 (pg10 line 210-211) respectively
The line time with steps and types of samples as follows
a Mar 2012 Ten male and 10 female adult fish were sampled for investigating tissue
distribution of 11β-HSD2
b May 2012 For detecting mRNA expressions of hepatic vtg and gonadal steroidogenic
genes following MT and EE2 exposure we sampled 18 fish (6 per group in triplicate) in each
group (control EE2 and MT groups in males and females) in which 12 gonads of 10 fish
were chosen for histological examination
8 Pg10-L214 What was the statistical unit and model How the time effect was included in
the model
Response Thanks for your suggestion In the present study the statistical unit is Ct value
of each detected gene (StAR cyp11a1 cyp17a1 cyp19a1a 3β-HSD and 11β-HSD) Statistical
differences were first tested with two-way ANOVA with treatment and time as two factors
Treatment was further analyzed with one-way ANOVA followed by LSD multiple comparison
(Pg7-L194-197)
As there is no way to rule out the effects of growth from day 7 to day 21 on the mRNA
expression of the detected genes the data of two-way ANOVA were not put in our manuscript We
added the sentence bdquoIn the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and
cyp19a1a the interaction of treatment and time was statistically significant (P lt 005)‟ in the
revised manuscript (Pg18-L297-298) The two-way ANOVA analyzed results are as follows
Table 1 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in female G ruras exposed to control MT and EE2 a
a Gene expression is expressed as the mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
Table 2 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in male G ruras exposed to control MT and EE2 a
a Gene expression is expressed as mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

protocol In this study the fish sampled for RNA isolation and RT consist of two independent
sections fish sampled for investigating tissue distribution of 11β-HSD2 (10 male fish and 10
female fish which were not exposure to MT and EE2) and for detecting mRNA expressions
of hepatic vtg and gonadal steroidogenic genes following MT and EE2 exposure (18 male fish
and 18 female fish control MT or EE2 groups) Thus the number of fish sampled for RNA
isolation and RT was divided into two sections and shown in para 28 (pg10 line 204-205)
and 29 (pg10 line 210-211) respectively
The line time with steps and types of samples as follows
a Mar 2012 Ten male and 10 female adult fish were sampled for investigating tissue
distribution of 11β-HSD2
b May 2012 For detecting mRNA expressions of hepatic vtg and gonadal steroidogenic
genes following MT and EE2 exposure we sampled 18 fish (6 per group in triplicate) in each
group (control EE2 and MT groups in males and females) in which 12 gonads of 10 fish
were chosen for histological examination
8 Pg10-L214 What was the statistical unit and model How the time effect was included in
the model
Response Thanks for your suggestion In the present study the statistical unit is Ct value
of each detected gene (StAR cyp11a1 cyp17a1 cyp19a1a 3β-HSD and 11β-HSD) Statistical
differences were first tested with two-way ANOVA with treatment and time as two factors
Treatment was further analyzed with one-way ANOVA followed by LSD multiple comparison
(Pg7-L194-197)
As there is no way to rule out the effects of growth from day 7 to day 21 on the mRNA
expression of the detected genes the data of two-way ANOVA were not put in our manuscript We
added the sentence bdquoIn the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and
cyp19a1a the interaction of treatment and time was statistically significant (P lt 005)‟ in the
revised manuscript (Pg18-L297-298) The two-way ANOVA analyzed results are as follows
Table 1 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in female G ruras exposed to control MT and EE2 a
a Gene expression is expressed as the mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
Table 2 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in male G ruras exposed to control MT and EE2 a
a Gene expression is expressed as mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

As there is no way to rule out the effects of growth from day 7 to day 21 on the mRNA
expression of the detected genes the data of two-way ANOVA were not put in our manuscript We
added the sentence bdquoIn the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and
cyp19a1a the interaction of treatment and time was statistically significant (P lt 005)‟ in the
revised manuscript (Pg18-L297-298) The two-way ANOVA analyzed results are as follows
Table 1 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in female G ruras exposed to control MT and EE2 a
a Gene expression is expressed as the mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
Table 2 Transcriptional profiles of ovarian steroidogenic enzyme genes of two-way ANOVA
analysis results in male G ruras exposed to control MT and EE2 a
a Gene expression is expressed as mean plusmn SD followed two-way ANOVA analysis P-values in
bold show significant difference (P lt 005)
9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

9 Table 1 only GSI values are of importance other data should be deleted
Response Thanks for your suggestion We have deleted other data as your suggestion in the
revised manuscript Revised table is showed below
Table 3 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a
Exposure time
(days)
Exposure
concentrations
GSI ()
Female Male
7
control 938 plusmn 005 283 plusmn 001
25 ngL EE2 366 plusmn 002darr 230 plusmn 001
25 ngL MT 580 plusmn 004 273 plusmn 001
50 ngL MT 550 plusmn 003darr 299 plusmn 002
100 ngL MT 406 plusmn 002darr 234 plusmn 001
14
control 587 plusmn 004 253 plusmn 001
25 ngL EE2 233 plusmn 001darr 195 plusmn 001
25 ngL MT 820 plusmn 005 254 plusmn 008
50 ngL MT 778 plusmn 006 428 plusmn 001uarr
100 ngL MT 566 plusmn 003 337 plusmn 003
21
control 541 plusmn 003 326 plusmn 002
25 ngL EE2 636 plusmn 003 141 plusmn 001darr
25 ngL MT 488 plusmn 003 380 plusmn 004
50 ngL MT 661 plusmn 004 241 plusmn 001
100 ngL MT 547 plusmn 004 414 plusmn004
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control
groups ( P lt 005) The arrows bdquouarr‟ or bdquodarr‟ indicate the significant up-regulation or
down-regulation of the morphological data Absence of the arrows bdquouarr‟ or bdquodarr‟ indicates no
significant difference between exposure groups and control group
10 Pg12-L245-247 The number of spz decreased in the testes as exposure time went on
from 7 to 21 days The authors should precise if such time effect was significant and how such
decrease was related to the MT concentration
Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

Response Thanks very much We have read references about analysis of gonads histological
results (Zha et al 2007 Kang et al 2008) After our close analysis we think that the
developmental stages of the germ cells of the gonads in our histological pictures are hardly
distinguishable So the diverse germ cells can‟t be counted and not statistical analysis can‟t be
conducted So we have changed the sentence ldquoIn MT exposures at 25 50 and 100 ngL the tissue
vacuolation increased and the number of spermatozoa decreased in the testes as exposure time
went on from 7 to 21 daysrdquo into ldquoIn MT exposures at 25 50 and 100 ngL for 7 to 21 days
macroscopic alterations (compared to controls) that the tissue vacuolation increased and the
number of spermatozoa decreased were found in the testesrdquo (Pg11 line 226-229)
11 The statistical conclusions concerning the time effect in relation to MT concentrations are
not clearly specified in the text so the authors should specify such conclusions for each studied
variable adding f and p values
Response Thanks for your suggestion In the present study we discussed the different
concentration of MT on rare minnow gonads steroidogenic genes and hepatic vtg According your
suggestion we have added the f and p values in the line 277 281 284 and 319 (Pg15-L277-278
Pg15-L281-283 Pg15-L284-285 Pg17-L319-321)
12 Pg12-L248 Features in para 32 have been described in the methodology section and this
section may not be considered as a result This is the same for the stability of reference genes in
para 33
Response Thanks for your suggestion We found the corresponding contents that ldquoThe
full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) was obtained by RT-PCR and RACE
methods (Fig S1)rdquo in para 32 and ldquoThe geNorm Comparative Delta CT method BestKeeper
and NormFinder analysis results are shown in Table S2 β-actin showed most stable among the
four candidate reference genes by four methods of geNorm Delt CT analysis BestKeeper and
NormFinderrdquo in para 33 According your suggestion we have recomposed these sentences in the
revised manuscript (Pg 14-L261 Pg 14-L266)
References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

References
M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular
cloning and characterization of amh dax1 and cyp19a1a genes and their response to
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013)
372ndash381
S Oumlrn H Holbech TH Madsen L Norrgren GI Petersen Gonad development and vitellogenin
production in zebrafish (Danio rerio) exposed to ethinylestradiol and methyltestosterone
Aquatic Toxicology 65 (2003) 397-411
Y Zheng L Wang M Li H Liang F Qin S Liu H Wang T Wu Y Zhang Z Wang
Molecular characterization of five steroid receptors from pengze crucian carp and their
expression profiles of juveniles in response to 17α-ethinylestradiol and 17α-methyltestosterone
General and comparative endocrinology 191 (2013) 113-122
IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias
latipes) Aquat Toxicol 87 (2008) 37ndash46
J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol
Chemosphere 66 (2007) 488ndash495
S Homklin SK Ong T Limpiyakorn Biotransformation of 17a-methyltestosterone in sediment
under different electron acceptor conditions Chemosphere 82 (2011) 1401ndash1407
K Barel-Cohen LS Shore M Shemesh A Wenzel J Mueller N Kronfeld-Schor Monitoring
of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

of natural and synthetic hormones in a polluted river Journal of Environmental Management
78 (2006) 16ndash23
Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

Highlights
The full-length cDNA of 11β-HSD2 was isolated and characterized successfully
Histological data demonstrated exposure of MT impaired gonadal development
MT disturbed mRNA expressions of hepatic vtg and gonadal steroidogenic genes
Highlights (for review)
1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

1
Gonadal development and transcript profiling of steroidogenic enzymes in response to 1
17α-methyltestosterone in the rare minnow Gobiocypris rarus 2
Shaozhen Liua Lihong Wang
b Fang Qin
a Yao Zheng
a Meng Li
a Yingying Zhang
a Cong Yuan
a 3
Zaizhao Wanga
4
5
a College of Animal Science and Technology Northwest AampF University Shaanxi Key Laboratory 6
of Molecular Biology for Agriculture Yangling Shaanxi 712100 China 7
b The Hospital of Northwest AampF University Yanglin Shaanxi 712100 China 8
9
10
11
Corresponding Author 12
Zaizhao Wang PhD 13
College of Animal Science and Technology 14
Northwest AampF University 15
22 Xinong Road 16
Yangling Shaanxi 712100 China 17
Tel +86-29-87092139 18
Fax +86-29-87092164 19
E-mail zzwangnwsuafeducn 20
21
22
ManuscriptClick here to view linked References
2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

2
Abstract 23
It is well known that natural and anthropogenic chemicals interfere with the hormonal system 24
of vertebrate and invertebrate organisms How these chemicals regulate gonadal steroidogenesis 25
remains to be determined The main objective of this study was to evaluate the effects of 26
17α-methyltestosterone (MT) a synthetic model androgen on gene expression profiles of six key 27
steroidogenic genes in adult rare minnow The full-length cDNA encoding 11β-hydroxysteroid 28
dehydrogenase-2 (11β-HSD2) was firstly isolated and characterized by RT-PCR and RACE 29
methods The gonadal transcript changes of StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and 30
cyp19a1a in 6-month adult G rarus exposed to MT and 17α-ethinylestradiol (EE2) for 7 14 and 31
21 days were detected by qRT-PCR To make an effort to connect the transcriptional changes of 32
steroidogenic enzymes with effects on higher levels of biological organization and on VTG one 33
remarkable sensitive target of steroids body and gonad weights histology of gonads and hepatic 34
vtg mRNA level were measured MT caused varying degree of abnormalities in ovaries and testes 35
The hepatic vtg mRNA level was highly inhibited in females and slightly altered in males by MT 36
Transcripts of several steroidogenic genes including StAR cyp17a1 and cyp11a1 showed high 37
responsiveness to MT exposure in G rarus The gene expression profiles of these steroidogenic 38
genes in MT-treated groups were much distinct with the EE2-treated group 39
Keywords steroidogenic genes 17α-methyltestosterone 17α-ethinylestradiol histology 40
Gobiocypris rarus 41
42
43
44
3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

3
1 Introduction 45
It is well known that a wide range of natural and anthropogenic chemicals interfere with the 46
hormonal system of vertebrate and invertebrate organisms [1 2] The field studies showed 47
physiologic and developmental abnormalities in wild fish such as increased vitellogenin (VTG) 48
concentrations and gonadal intersexuality of male fish in polluted aquatic environment [3-5] 49
Anthropogenic chemicals have several distinct effects including estrogenic androgenic 50
anti-estrogenic and anti-androgenic effects [6-8] Natural and man-made androgenic chemicals 51
are attracting more attention increasingly 52
The undetermined androgenic substances from water collected downstream of a kraft mill of 53
Fen Holloway River is associated with masculinization of female mosquitofish (Gambusia affinis 54
holbrooki) [9] The 17α-methyltestosterone (MT) is widely applied in aquaculture to control 55
sexual differentiation and induce sex-reversal of genetic females to phenotypic males [10-12] 56
MT was detected (133 ngL) in samples of effluent from a chemical factory [13] Concentrations 57
of 61 ngL testosterone and 45 ngL androstenedione were reported in final effluent of 58
municipal wastewater treatment plant [14] The MT could be detected in levels ranging from 59
41ndash70 ngL in waste water obtained from the Beijing area [15] MT has been recommended as a 60
reference chemical in tests for evaluating chemicals for endocrine disruption [16] In the present 61
study we chose the synthetic androgen MT as a test chemical 62
In the laboratory experiments MT demonstrates both androgenic and estrogenic effects in 63
several model fish MT exposure caused masculinization of female adult fathead minnow 64
(Pimephales promelas) exhibited development of nuptial tubercle which is a secondary sex 65
characteristic structure normally found in sexually mature males [17 18] In adult male medaka 66
4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

4
(Oryzias latipes) both full life-cycle MT exposure at 2775 ngL and 3-week MT exposure at 380 67
ngL induced the development of testis-ova within the testicular tissues [19 20] Several studies 68
reported the VTG response in fish exposed to MT In both fathead minnow and medaka MT 69
exposure induced significant increase of VTG in adult males [17ndash19] In goldfish and fathead 70
minnow MT can induce VTG synthesis in their juveniles [21 22] However MT also resulted in 71
inhibition of VTG synthesis in female medaka and had no effect on the VTG level in the males 72
[20] 73
Xenoandrogens might affect the enzymes involved in steroidogenesis The changes of 74
steroidogenic enzyme activities resulted from xenoandrogens will most likely elicit alteration of 75
endogenous steriod hormone levels Several studies showed changes in estrogen and androgen 76
concentrations in fish exposed to MT MT can modulate 17β-estradiol (E2) and testosterone (T) 77
levels in a concentration- and time-dependent manner in cultured previtellogenic oocytes of 78
Atlantic cod (Gadus morhua) in vitro [23] The levels of 11-ketotestosterone (11-KT) and T 79
decreased in a concentration-dependent manner in male zebrafish exposed to MT at 45 to 623 80
ngL for 7 days [24] 81
Steroidogenesis involves a variety of enzymes and potential biochemical pathways Among 82
these biochemical pathways the regulatory step in acute steroid production is the transfer of 83
cholesterol across the mitochondrial membrane by steroidogenic acute regulatory protein (StAR) 84
and the subsequent conversion to pregnenolone by Cytochrome P450-mediated side-chain 85
cleavage enzyme (P450scc cyp11a1) [25ndash27] Cytochrome P450 aromatase (cyp19a1) is another 86
key steroidogenic enzyme responsible for the conversion of a range of androgens into oestrogen 87
[28] It has been demonstrated that teleost fish have two distinct isoforms of cyp19a1 genes 88
5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

5
cyp19a1a and cyp19a1b Cyp19a1a is predominantly expressed in the ovary and involved in 89
sexual differentiation and ovarian development whereas cyp19a1b mainly expressed in the brain 90
[29] The 3β-hydroxysteroid dehydrogenase (3β-HSD) is a key enzyme catalyzing the synthesis of 91
the potent steroid hormones progesterone 17α-hydroxyprogesterone androstenedione and 92
testosterone from their much less hormonally active precursors pregnenolone 93
17α-hydroxypregnenolone dehydroepiandrosterone respectively [30 31] Hydroxylase and lyase 94
activity of cytochrome 17α-hydroxylase 17 20-lyase 1 (CYP17A1) are required for the synthesis 95
of testosterone [32] The functions of 11β-hydroxysteroid dehydrogenase 2 (11β-HSD2) is to 96
convert 11β-hydroxytestosterone to 11-ketotestosterone (11-KT) the main androgen found in the 97
majority of fish species [33 34] 98
The present study was aimed to evaluate the effects of MT at low concentrations (25-100ngL) 99
on gene expression profiles of six key steroidogenic genes In addition we investigated the effects 100
of MT on the transcriptional level of hepatic VTG and gonadal development Rare minnow 101
Gobiocypris rarus was used as model fish in the present study G rarus is a Chinese freshwater 102
cyprinid endemic to the upstream of Yangze River Sichuan Province China It is an appropriate 103
ecological test organism for the assessment of endocrine disrupting chemicals due to its small size 104
(30-80 mm in total length) ease of culture short life cycle (maturation within six to eight weeks) 105
and high sensitivity to aquatic pollutants [35] We deduce that MT exposure will produce 106
differential gene expression profiles of these steroidogenic genes and subsequently change the 107
levels of endogenous steroid hormones with significant effects on gonad development in Chinarsquos 108
indigenous model fish G rarus 109
2 Materials and methods 110
6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

6
21 Animals and chemicals 111
Fertilized eggs were obtained from the female fish spawned in our laboratory After hatching 112
the larvae were fed Artemia nauplii (lt 24 h after hatching) twice a day After 30 days post 113
fertilization (dpf) fish were fed chironomid larvae once a day The adult fish were raised in 125 L 114
glass tanks with dechlorinated tap water at 25 plusmn 2 degC The photoperiod was 14 h 10 h lightdark 115
EE2 (purity gt 98) and MT (purity gt 990) were purchased from Sigma Chemicals Inc (St 116
Louis MO USA) MT and EE2 were dissolved in ethanol (AR analytical reagent) 117
22 MT and EE2 exposure 118
Adult rare minnow at 184 dpf (113 plusmn 033 g in weight 48 plusmn 05 cm in total length) were 119
exposed to MT and EE2 or solvent control (0001 ethylalcohol vv) in 45 liters glass tanks (36 120
liter water per tank) for 7 14 and 21 days (approximately 1 gram fish per liter water) The nominal 121
MT concentrations were 25 50 and 100 ngL As a potent synthetic estrogen EE2 served as a 122
reference estrogen control in the present exposure experiment The nominal EE2 concentration 123
was 25 ngL [35] Half of the exposure solution was changed every day Following exposure the 124
fish were immediately dissected and gonad tissues were frozen in liquid nitrogen and kept 125
individually in frozen tubes at -80 oC until use All exposure experiments were conducted in 126
triplicate in separate tanks at the same time 127
23 Morphometry and gonadal histological examination 128
The body weight and body length of fish were firstly measured Following sampling the 129
gonad tissues were weighed And the gonadosomatic index was calculated using the formula of 130
GSI = 100 times gonad weightbody weight [17] These data are expressed as mean plusmn SD and 131
statistical differences were tested by Analysis of Variance (ANOVA) and least significant 132
7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

7
difference (LSD) Then the gonads were immersed in paraformaldehyde (4) solution over 24 133
hours at 4 degC and prepared for histological examination We chose 10 fish in each group (control 134
EE2 and MT groups) Then formalin-fixed gonads were dehydrated and processed for paraffin 135
wax embedding Serial longitudinal sections with thickness of 7 μm were cut with a rotary 136
microtome (Leica RM2235 Leica Microsystems) and stained using hematoxylin-eosin (HE) 137
Microscopic examination was carried out using an Olympus CHC binocular microscope and 138
photography was done with a Motic Digital Microscope 139
24 RNA isolation and reverse transcription (RT) 140
The tissues of gonad brain liver muscle eye intestines and gill were sampled from adult 141
male and female G rarus to assay the tissue distribution of 11β-HSD2 mRNA Tissue samples for 142
cDNA cloning and quantitative real-time PCR (qRT-PCR) were homogenized in TRIZOL reagent 143
(Invitrogen) and the total RNAs were extracted as described previously [36] cDNAs were 144
synthesized from total RNA with M-MLV reverse transcriptase (Invitrogen) and oligo (dT)18 145
primer in 20 μL final volume 146
25 Complementary DNA cloning of 11β-HSD2 147
The full-length cDNA sequence of G rarus 11β-HSD2 gene was generated in three 148
consecutive steps using reverse transcription polymerase chain reaction (RT-PCR) and rapid 149
amplification of cDNA ends (RACE) strategies [37] 11β-HSD2 was isolated from the testis The 150
cDNA fragment of 11β-HSD2 was obtained by PCR using primers designed from the consensus 151
sequences of teleosts (Table S1) The methods of TA cloning and RACE were described in the 152
previous study [38] 153
26 Sequence analysis 154
8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

8
The full-length cDNA of 11β-HSD2 was assembled by alignment of the internal core fragment 155
5rsquo- and 3rsquo-RACE fragment using SeqMan program of Lasergene software (DNASTAR Inc) 156
Amino acid multiple alignments were performed using the Megalign program of Lasergene 157
software To establish phylogenetic tree for the G rarus 11β-HSD2 we aligned diverse vertebrate 158
11β-HSD2 gene at the level of amino acid sequences by Clustal X (183) sequences alignment 159
program respectively [39] The neighbor-joining algorithms method of Mega 40 program 160
(Molecular Evolutionary Genetic Analysis) was used to construct phylogenetic tree [4041] 161
Bootstrap analyses were conducted using 1000 replicates 162
27 Reference gene Screening 163
To obtain a reliable reference gene for the normalization of qRT-PCR data four candidate 164
reference genes were selected to validate their expression stability These genes are β-actin 165
elongation factor 1-alpha (ef1a) glyceraldehyde-3-phosphate dehydrogenase (gapdh) and tubulin 166
alpha 1 (tuba1) qRT-PCR was performed using total RNA from the samples for mRNA tissue 167
distribution and exposure (including MT EE2 and control groups) qRT-PCR was performed by 168
CFX96 real-time PCR detection systems (Bio-Rad) thermocycler and SYBR Premix ExTaq II kit 169
(TaKaRa) The detailed procedures of qRT-PCR were the same as described previously [42] The 170
qRT-PCR primers are listed in Table S1 qRT-PCR efficiency (E) was calculated from the given 171
slops in CFX Manager software by a 10-fold diluted cDNA sample series with five dilution points 172
measured in triplicate E was determined by the equation E=10 (minus1slope)
[43] The programs of 173
geNorm [44] Comparative Delta CT method [45] BestKeeper [46] and NormFinder [47] were 174
used to calculate gene expression stability (M) value of reference genes 175
(httpwwwleonxiecomreferencegenephp) The M value is defined as the average pairwise 176
9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

9
variation of a certain gene with the remaining genes tested The lowest M value indicates the most 177
stable reference gene whereas the highest M value indicates the least stable reference gene 178
28 Tissue distribution of 11β-HSD2 179
The distribution of 11β-HSD2 transcript was determined in the tissues of gonad brain liver 180
muscle eye intestine and gill of adult fish by qRT-PCR The qRT-PCR primers are listed in Table 181
S1 To verify the specificity of the primers they were tested in normal PCR amplification and the 182
PCR products were visualized on a 15 agarose gel before qRT-PCR qRT-PCR data were 183
obtained as Ct The amounts of 11β-HSD2 mRNA were normalized to the most reliable reference 184
gene The relative change in mRNA expression levels of the four genes in different tissues were 185
calculated by 2minusΔΔCt
method [48] The analysis was performed on tissues of 10 males and 10 186
females All data are expressed as mean plusmn SD 187
29 Detecting mRNA expressions of hepatic vtg and gonadal steroidogenic genes following MT 188
and EE2 exposure 189
The gene expression patterns of hepatic vtg and gonadal StAR cyp11a1 3β-HSD cyp17a1 190
11β-HSD2 and cyp19a1a were detected for the fish exposed to MT and EE2 by qRT-PCR Each 191
transcript was analyzed on 18 individuals (6 per group in triplicate) The relative changes of 192
mRNA levels of these genes after MT and EE2 exposure were calculated using 2minusΔΔCt
method with 193
the formula F=2minusΔΔCt
ΔΔCt=(Ct target gene - Ct reference gene)MT or EE2-(Ct target gene - Ct reference gene)control 194
[48] Data are expressed as the fold change compared to control groups Statistical differences 195
were first tested with two-way ANOVA with treatment and time as two factors Treatment was 196
further analyzed with one-way ANOVA followed by LSD multiple comparison 197
3 Results 198
10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

10
31 GSI and gonadal histology 199
In the present study the data of GSI were shown in Table 1 In the female fish exposure for 7 200
days MT at 50 and 100 ngL significantly decreased the GSI compared to the control groups 201
Meanwhile EE2 exposure for 7 and 14 days significantly reduced the GSI compared to the control 202
groups in female fish Following exposure for 21 days EE2 significantly reduced GSI compared 203
to the control groups respectively in the male fish 204
The histological results of EE2 and MT exposure showed inhibition of oogenesis and 205
spermatogenesis (Fig 1 and 2) The ovaries of female fish exposed to 25 ngL EE2 for 7 days 206
contained a high proportion of perinucleolar oocytes (Poc the most immature oocyte) a small 207
number of cortical alveolus stage oocytes (Coc the less mature oocyte) and no vitellogenic 208
oocyte (Voc the most mature oocyte) (Fig 1B1) The prolonged exposure time of 14 and 21 days 209
elicited the most immature oocyte Poc as the only component of the ovaries (Fig 1B2 1B3) In 210
the MT exposures at 25 and 50 ngL the number of immature oocytes (Coc and Poc) increased 211
however the number of mature oocyte (Voc) decreased as treatment time went on from 7 to 21 212
days (Fig 1Cs and 1Ds) MT exposure at 100 ngL for 7 days strongly inhibited the oocyte 213
development with the most immature oocyte Poc as the sole component in the ovaries (Fig 1E1) 214
However as exposure time prolonged (14 and 21 days) Poc and Coc were observed in the ovaries 215
(Fig 1E2 and 1E3) 216
In the male G rarus no abnormal testicular development was found in the control groups (Fig 217
2A1 2A2 and 2A3) In the male fish of EE2 groups more interstitial tissue was observed in testes 218
compared to control groups (Fig 2B1 and B2) EE2 exposure for 14 days more serious 219
vacuolation and extremely abnormal connective tissue were detected with few spermatozoa or 220
11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

11
spermatocytes observed in the testes (Fig 2B2) Meanwhile abnormal connective tissues with few 221
spermatozoa were present in all three EE2 exposure groups In MT exposures at 25 50 and 100 222
ngL for 7 to 21 days macroscopic alterations (compared to controls) that the tissue vacuolation 223
increased and the number of spermatozoa decreased were found in the testes (Fig 2C1 C2 and 224
C3) 225
32 Molecular characterization of 11β-HSD2 226
The full-length cDNA of 11β-HSD2 (GeneBank ID KC454276) is 1965 bp including 1242 bp 227
ORF encoding a putative protein of 413 amino acids with a theoretical pI of 794 and a calculated 228
molecular weight of 460 kDa a 238 bp 5rsquo-UTR and a 485 bp 3rsquo-UTR (Fig S2) Polyadenylation 229
signal (AATAAA) was found in the 3rsquo-UTR of 11β-HSD2 (Fig S2) 230
33 Homology and phylogenetic analysis of putative amino acid sequences for 11β-HSD2 231
The amino acid sequence of G rarus 11β-HSD2 shows the highest homology with zebrafish 232
and the conserved regions are NAD-binding domain 11β-HSD2 Conserved sequence and 233
Catalytic-site (Fig S3) To better understand the position of G rarus 11β-HSD2 in the 234
evolutionary history of the respective protein phylogenetic tree was constructed from the amino 235
acid alignments for 11β-HSD2 of teleosts and mammals using the neighbor-joining method G 236
rarus 11β-HSD2 was more close to their counterparts of zebrafish than that of other teleosts (Fig 237
S4) 238
34 Expression stability analysis of reference genes 239
The analysis of reference gene results is shown in Table S2 β-actin showed most stable 240
among the four candidate reference genes The recommended comprehensive ranking indicated 241
β-actin as the most stable reference gene Therefore β-actin was selected as the reference gene in 242
12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

12
the present study 243
35 Tissue distribution of 11β-HSD2 244
The tissue distribution of 11β-HSD2 mRNA is shown in Fig 3 11β-HSD2 was predominantly 245
expressed in liver The transcript of 11β-HSD2 in the testis was 4173-fold higher than that in the 246
ovary In females 11β-HSD2 transcript in the liver was 68154-fold higher than that in the ovary 247
However in males the 11β-HSD2 transcript in the liver was 201 times of that in testis The 248
transcript of 11β-HSD2 in the male brain was 165-fold higher than those in the female brain 249
36 Expression profiles of hepatic vtg in adult G rarus under MT and EE2 exposures 250
The expression of hepatic vtg in adult female and male G rarus was detected in the present 251
study (Table 2) In the female fish transcript of hepatic vtg significantly increased for 929 252
3119 and 740 fold (P=001 P=002 and P=005 respectively) following 7- 14- and 21-day EE2 253
exposure compared to the control respectively Seven-day exposure of MT at 50 ngL caused 254
extremely significant decrease of the hepatic vtg transcript in the female fish compared to control 255
groups (16937 fold P=004) The 14-day MT treatments at 25 to 100 ngL significantly 256
suppressed expression of hepatic vtg for 1552 to 369 fold in the female fish As time went on (7 257
14 and 21 days) MT treatment at 100 ngL significantly decreased the expression of hepatic vtg 258
for 1137 3161 and 78350 fold (P=005 P=005 and P=005 respectively) in the female fish 259
In the male fish EE2 exposure for 7 to 21 days caused significant increase of hepatic vtg 260
transcript with huge magnitude orders (760 230 and 106 thousand-fold Plt001 P=002 and 261
P=005 respectively) In the male fish the vtg expression was significantly stimulated for 39 to 262
22 fold by 7- and 14-day MT exposures at 50 and 100 ngL compared to control However 21-day 263
MT exposures at 25 and 50 ngL caused 1911- and 44-fold (P=005 and P=005) significant 264
13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

13
decrease of vtg transcript in male fish respectively 265
37 Expression profiles of steroidogenic genes in adult G rarus under MT and EE2 exposure 266
In the female and male fish for gonadal StAR cyp17a1 11β-HSD2 and cyp19a1a the 267
interaction of treatment and time was statistically significant (P lt 005) The fold changes of gene 268
expression for gonadal StAR cyp11a1 3β-HSD cyp17a1 11β-HSD2 and cyp19a1a under MT and 269
EE2 exposure compared to controls are shown in Table 3 and 4 In the female G rarus EE2 270
exposure at 25 ngL for 7 14 and 21 days caused extremely significant decrease of the 271
steroidogenic enzyme genes except for 3β-HSD and 11β-HSD2 In the female G rarus the effects 272
of MT on 3β-HSD and 11β-HSD2 were weaker than that on the other 4 genes (StAR cyp11a1 273
cyp17a1 and cyp19a1a) Seven-day MT exposure at 25 ngL caused 222-fold significant decrease 274
of transcript for ovarian cyp11a1 and respective 184- 157-fold increase of ovarian cyp17a1 and 275
11β-HSD2 compared to controls (Table 3) Seven-day exposure to MT at 50 ngL caused 276
extremely significant decrease the ovarian transcript of StAR (824-fold) while significant increase 277
the ovarian transcript of cyp17a1 compared to controls (653-fold Table 3) Seven-day MT 278
exposure at 100 ngL significantly down-regulated expressions of ovarian StAR cyp11a1 3β-HSD 279
and cyp17a1 (84 to 5731 fold) Fourteen-day exposure to MT at 25 and 50 ngL significantly 280
down-regulated the ovarian transcripts of cyp11a1 and cyp17a1 compared to controls (367 to 281
1372 fold respectively Table 3) MT exposure at 50 ngL for 14 days significantly up-regulated 282
expression of ovarian cyp19a1a (603-fold Table 3) MT exposure at 100 ngL for 14 days 283
significantly down-regulated expression of all six ovarian steroidogenic enzyme genes compared 284
to controls (gt 232 fold Table 3) MT exposure at 25 ngL for 21 days significantly increased the 285
expression of StAR and cyp19a1a (323- and 247-fold respectively) However 21-day MT 286
14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

14
exposure at 25 ngL significantly decreased the expression of cyp17a1 compared to controls 287
(396-fold Table 3) MT exposure at 50 ngL for 21 days extremely significantly stimulated the 288
expression of StAR cyp17a1 11β-HSD2 and cyp19a1a for 422 163 888 and 286 times in the 289
female fish respectively MT exposure at 100 ngL for 21 days significantly up-regulated the 290
expression of ovarian StAR and cyp19a1a (497 and 225 fold Table 3) The expression of 291
cyp17a1 was extremely significantly down-regulated for 1061 (Plt001) 6075 (Plt001) and 194 292
(Plt001) times following 7- 14- and 21-day exposure to MT at 100 ngL (Table 3) 293
In male G rarus EE2 exposure at 25 ngL in three durations caused generally significant 294
decrease of all transcripts of steroidogenic enzyme genes except for cyp19a1a compared to 295
controls (Table 4) Seven-day exposure to MT at 25 ngL significantly down-regulated the 296
expression of testicular StAR cyp11a1 and 11β-HSD2 for 53 159 and 359 fold respectively 297
(Table 4) MT exposure at 50 ngL for 7 days significantly suppressed the testicular transcripts of 298
StAR and 11β-HSD2 (239 and 180 fold Table 4) Seven-day exposure to MT at 100 ngL caused 299
extremely significant decrease of testicular transcripts for StAR cyp11a1 3β-HSD cyp17a1 and 300
11β-HSD2 (237 to 2117 fold Table 4) Fourteen-day MT treatment at 25 ngL significantly 301
down-regulated the expression of 3β-HSD for 244 times MT exposure at 50 ngL for 14 days 302
resulted in 5091-fold extremely significant increase of testicular cyp17a1 transcript (Table 4) MT 303
exposure at 100 ngL for 14 days significantly up-regulated expression of cyp17a1 and 11β-HSD2 304
for 7782 and 220 fold respectively In 21-day exposure duration MT at 25 ngL extremely 305
significantly up-regulated the expression of 11β-HSD2 (239 fold) MT at 50 ngL resulted in 168- 306
and 208-fold significant increase of testicular 11β-HSD2 and cyp17a1 transcripts (Table 4) MT at 307
100 ngL resulted in extremely significant decrease of testicular StAR cyp11a1 and cyp17a1 308
15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

15
transcript (532 to 3425 fold Table 4) MT at 100 ngL exposure for 7 14 and 21 days the mRNA 309
expression of gene StAR were all decreased (873 236 and 532 fold Plt001 P=005 and Plt001 310
respectively) 311
4 Discussion 312
In the present study gonadal development gonadosomatic index (GSI) and vtg mRNA were 313
employed as biological endpoints for detecting the MT effects EE2 treatment at 25 ngL caused 314
degeneration of ovaries and testes reduced GSI of female and male fish which is in line with the 315
findings of experiment on rare minnow [35] EE2 treatment at relatively low concentrations (1-25 316
ngL) induced testes-ova in adult rare minnow [35] However in the present study no testis-ova 317
were observed in EE2-treated fish The previous study demonstrated that the Betta splendens had 318
smaller gonads after EE2 (100 ngL) exposure for 4 weeks compared to the control [49] Similarly 319
in our study EE2 had inhibited the rare minnow gonads The discrepancy of exposure duration 320
andor developmental status of testis of the subject male fish in the two experiments could be the 321
cause of the difference MT treatment caused reduced GSI in 7-day exposure and degeneration of 322
oocytes in the ovaries of the female fish in three exposure durations (7 14 and 21 days) at three 323
concentrations (25 50 and 100 ngL) which is consistent with the results of experiments on 324
MT-treated female fathead minnow and medaka [20 50] MT exposure at 50 and 100 ngL also 325
resulted in histological abnormalities in testes of the adult male fish However MT at 01-50 μgL 326
had no effect on the testicular structure and amount of sperm in fathead minnow [50] In 327
MT-treated male medaka the testes in the 225-188 ngL groups had no histological aberrations 328
and only 380 ngL MT caused the occurance of testis-ova [20] Therefore in the present study the 329
testis of rare minnow showed higher responsiveness to MT compared to fathead minnow and 330
16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

16
medaka The mechanism by which MT induced these phenomena in female and male rare minnow 331
is undetermined Further investigations are needed to address the mechanism underlying abnormal 332
gonadal development in rare minnow exposed to MT 333
It is well established that EE2 is a potent inducer of VTG in fish acting via the hepatic ERs 334
So EE2 was chosen as a positive estrogenic control in the experiment The response of vtg mRNA 335
to estrogenic chemicals is more sensitive than the response of VTG protein For example the 336
induction of vtg mRNA expression was two-order magnitude greater than that of plasma VTG 337
protein level in the male fathead minnow exposed to EE2 at 10 ngL [51] So in the present study 338
vtg mRNA levels were detected in liver where VTG is synthesized As expected EE2 induced 339
hepatic mRNA expression of vtg gene in both male and female rare minnow and the response in 340
treated male fish was much more dramatic than the EE2-treated females which is consistent with 341
the observations for fathead minnow in both field and laboratory EE2 exposures [51 52] 342
In the male fish MT exposure for 7 and 14 days induced the transcription of hepatic vtg gene 343
albeit not significantly in some cases Interestingly the 21-day MT exposure inhibited mRNA 344
expression of hepatic vtg in male fish The present findings suggest that androgens may involve in 345
the regulation of VTG synthesis Several previous studies have been demonstrated the ability of 346
MT to influence VTG synthesis Seven-day exposure of MT at measured concentration of 45 ngL 347
increased the VTG concentration in whole-body homogenates of adult male fish [24] Furthermore 348
in full life cycle test of MT exposure on Japanese medaka the hepatic VTG level was increased in 349
F1 generation male at 60 days posthatch exposed to 998 ngL MT [19] In fathead minnow MT at 350
higher concentrations (02-200 microgL) also induced VTG synthesis in the male fish [1718] The 351
cause of induced VTG synthesis for MT probably is that MT can be aromatized into 352
17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

17
methylestradiol (ME2) and ME2 with estrogenic effect subsequently up-regulate vtg expression 353
via the hepatic estrogen receptor [18] 354
MT exposures at 25-100 ngL in three durations nearly all significantly suppressed expression 355
of hepatic vtg gene in female rare minnow The response of vtg transcript upon MT in female fish 356
is in line with several previous studies In full life cycle of MT exposure in Japanese medaka the 357
hepatic VTG level was significantly decreased in F1 generation female at 60 days posthatch 358
exposed to MT at 035-998 ngL [19] Significant decrease of hepatic vtg mRNA showed in the 359
hermaphrodite fish (Kryptolebias marmoratus) treated with MT [53] However 21-day MT 360
exposure at high concentrations (02 and 2 mgL) induced plasma VTG in adult female fathead 361
minnow [17] We speculate that there are species and sexual differences in VTGvtg mRNA 362
response to MT for fish 363
In the present study we analyzed the transcriptional responses of main steroidogenic genes to 364
MT and EE2 Our investigations demonstrated that the treatment of the potent estrogen EE2 365
(25ngL) for 7 to 21 days pronouncedly disturbed mRNA expressions of the main steroidogenic 366
genes in gonads of both male and female fish In testis almost the three durations of EE2 exposure 367
(7 14 and 21 day) markedly suppressed the expressions of StAR cyp11a1 cyp17a1 and 11β-HSD2 368
but did not affect mRNA levels of 3β-HSD and cyp19a1a EE2 exposure of the three durations 369
caused much greater inhibition of ovarian StAR cyp11a1 and cyp17a1 than those in testis The 370
EE2 exposures resulted in significant decrease of cyp19a1a mRNA and no effect on the transcript 371
of 11β-HSD2 in ovary Similar to the situation of 3β-HSD in testis the ovarian 3β-HSD transcript 372
was almost not affected by EE2 The results in the present study are consistent with the inhibitory 373
effects of EE2 on cyp17 and 11β-HSD in male fathead minnow [51] In testis of rainbow trout 374
18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

18
(Oncorhynchus mykiss) EE2 also inhibited the expression of four genes coding for enzymes 375
involved in androgen production including cyp11a1 cyp17 3β-HSD and 11β-HSD2 [54] In the 376
zebrafish A clear down-regulation of StAR mRNA in gonads were found in the EE2 groups [55] 377
The transcripts of ovarian StAR cyp11a1 cyp17a1 and cyp19a1a were strongly inhibited by 25 378
ngL EE2 for 7 14 and 21 days All these genes code for important enzymes for estrogen 379
production in female fish For example StAR and CYP11A1 are the key proteins involved in the 380
early steroidogenic pathway The CYP19A1 is a crucial steroidogenic enzyme catalyzing the final 381
step in the conversion of androgens to estrogens in fish [29] The marked decrease of mRNA 382
levels for these key steroidogenic genes in response to EE2 implies the reduced estrogen in ovary 383
of female fish which is in line with the significant decrease of plasma E2 in zebrafish exposed to 384
EE2 at 15-100 ngL for 2 and 7 days [56] The concurrent transcriptional responses of several 385
steroidogenic genes in testis and ovary could result from the inhibition of a common 386
transcriptional factor such as the SF-1 gene In fathead minnow EE2 (10ngL) exposure for 21 387
days significantly inhibited mRNA expression of SF-1 in both male and female fish [51] In the 388
present study the strong inhibition of steroidogenic enzymes at transcriptional level in the female 389
exposed to EE2 suggests that it is the exogenous EE2 but not the endogenous E2 that significantly 390
induces the expression of hepatic vtg 391
In the present study several genes of steroidogenic enzymes in ovary of female fish were 392
inhibited by MT Cyp17a1 mRNA expression was strongly repressed by 7- 14- and 21-day MT 393
treatment at 25 to 100 ngL MT exposures at 50 ngL for 7 day and at 100 ngL for both 7 and 14 394
days significantly inhibited StAR mRNA level MT treatment for duration of 7 days at 25 and 100 395
ngL and 14 days at 25 to 100 ngL caused marked decrease of cyp11a1 transcript 3β-HSD was 396
19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

19
strongly repressed by MT (100 ngL) for 7 and 14 days The down-regulation of genes encoding 397
steroid enzymes by androgens has already been reported in fish species including rainbow trout 398
[57 58] and the Nile tilapia [59] In mammal the mRNA expression of StAR cyp11a1 cyp17a1 399
and 3β-HSD was also inhibited by androgens [60 61] A few regulatory factors have been studied 400
on this down-regulation of androgens SF-1 is a transcription factor which has been shown to 401
stimulate the mRNA expression of StAR cyp11a1 cyp17a1 and 3β-HSD in mammal [62 63] In 402
female rainbow trout three-month treatment of another androgen 11β-hydroxyandrostenedione 403
(11βOHΔ4) down-regulated sf-1 expression in ovary [57] So SF-1 may be one of regulators 404
involved in the transcriptional inhibition of steroidogenic enzyme genes The prolonged MT 405
treatment (21 days) at 25-100 ngL resulted in significant increase of transcripts for StAR and 406
cyp19a1a which both code for enzymes involving in key steps of steroid production in ovary The 407
up-regulation of the two key steroidogenic genes which imply the increase of the enzymes could 408
counteract the continuing complete block of ovarian steroidogenesis Our previous study found 409
100 ngL MT significantly increased the mRNA expression of cyp19a1a in Pengze crucian carp 410
[64] In addition the up-regulation of cyp19a1a upon MT treatment for 21 days could be 411
conducive to the enzymatic conversion of MT to 17α-methylestradiol (ME2) by CYP19A1 412
aromatase In female fathead minnow ME2 was detected in plasma samples following the 7-day 413
MT exposure which confirms the conversion of MT to ME2 [18] 414
Our experiment demonstrates that MT treatment of male rare minnow resulted in transcriptional 415
down-regulation of testicular steroidogenic enzymes genes in the short duration (7 days) and as 416
the treatment time prolonged some genes had a decelerating trend of down-regulation or reversed 417
to up-regulation For example 7-day MT exposure at 25-100 ngL all strongly inhibited StAR 418
20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

20
expression in testis (239- to 873-fold P lt 001) whereas only the highest concentration (100 419
ngL) of MT caused significant decrease of StAR transcript in the longer exposure (236-fold P = 420
005 and 532-fold P lt 001 in 14- and 21-day exposure respectively) For cyp17a1 gene 7-day 421
MT treatment at 100 ngL markedly suppressed (2117-fold P lt 001) its expression however 422
14-day MT exposures at 50 and 100 ngL strongly up-regulated its expression (5091- and 423
7782-fold P lt 001) Similar to cyp17a1 gene the 11β-HSD2 expression also had a reversed 424
regulation with the down-regulation upon MT exposure of the short duration (7 days) at 25-100 425
ngL (180- to 1182-fold P le 001) and up-regulation upon MT exposure for longer duration 426
(220-fold for 14-day exposure at 100 ngL 239- and 168-fold for 21-day exposure at 25-50 ngL 427
P le 001) The data of the present study is in line with the regulatory effects of another androgen 428
11βOHΔ4 on the transcripts of several steroidogenic enzymes in male rainbow trout [54] The 429
11βOHΔ4 treatment for 4 to 10 days caused significant decrease of mRNA levels for cyp11a1 430
3β-HSD and 11β-HSD2 in male rainbow trout For cyp17a1 gene it expression was 431
down-regulated by short term of exposure (4-10 days) of 11βOHΔ4 and up-regulated by long term 432
of treatment (16 days) on the contrary The transcriptional modulation of steroidogenic enzymes in 433
response to MT could be triggered by factors in HPG-axis In Leydig cells of male rat treated with 434
testosterone for 2 months the transcriptional down-regulation of steroidogenic enzymes (StAR 435
cyp11 and cyp17) coupled with significantly decreased LH levels in circulation suggest that MT 436
could regulate androgen production through LH-LHR-cAMP signaling [65] In the present study 437
MT at 25 to 100 ngL barely had effects on mRNA level of cyp19a1a which is consistent with 438
previous findings in male zebrafish [24] 439
In summary MT treatment induced abnormalities of ovaries and testes in adult rare minnow 440
21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

21
which can not be distinguished with the observation in the fish treated with EE2 The hepatic vtg 441
as a well-known sensitive biomarker was barely weakly stimulated in male fish and markedly 442
inhibited in female upon MT exposure which hardly leads to conclusion that MT has estrogenic 443
effects in adult rare minnow as in the MT exposure tests on other teleosts such as fathead minnow 444
The transcripts of several steroidogenic enzymes genes such as StAR cyp17a1 and cyp11a1 445
showed high responsiveness to MT exposure in rare minnow Moreover the MT- and EE2-treated 446
fish demonstrated distinct gene expression profiling for these steroidogenic enzymes genes 447
suggesting that the similar phenotypic effects in the higher levels of biological organization for 448
MT and EE2 could result from distinct molecular mechanisms The mechanisms underlying the 449
adverse effects of exogenous androgens can not be determined by the studies merely limited to the 450
gene expression of steroidogenic enzymes and future investigations needs to be performed in wide 451
spectrum of systems such as transcription regulation nuclear receptor signaling and 452
LH-LHR-cAMP signaling 453
Acknowledgments 454
This study was supported by grants from the National Natural Science Foundation of China 455
(31270547) the Natural Science Foundation of Shaanxi Province China (2011JM3009) and the 456
Fundamental Research Funds for the Central Universities in Northwest AampF University 457
(QN2011062) 458
Appendix A Supplementary data 459
Supplementary data associated with this article can be found in the online version 460
461
462
22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

22
Reference 463
[1] P Matthiesen P Gibbs Critical appraisal of the evidence for tributyltin-mediated endocrine 464
disruption in mollusks Environ Tox Chem 17 (1997) 37ndash43 465
[2] M Tillmann U Schulte-Oehlmann M Duft B Markert J Oehlmann Effects of endocrine 466
disruptors on prosobranch snails (Mollusca Gastropoda) in the laboratory Part III 467
Cyproterone acetate and vinclozolin as antiandrogens Ecotoxicology 10 (2001) 373ndash388 468
[3] S Hashimoto H Bessho A Hara M Nakamura T Iguchi K Fujita Elevated serum 469
vitellogenin levels and gonadal abnormalities in wild male flounder (Pleuronectes yokohamae) 470
from Tokyo Bay Japan Mar Environ Res 49 (2000) 37ndash53 471
[4] R Van Aerle M Nolan S Jobling LB Christiansen JP Sumpter CR Tyler Sexual 472
disruption in a second species of wild cyprinid fish (the gudgeon Gobio gobio) in united 473
kingdom freshwaters Environ Toxicol Chem 20 (2001) 2841ndash2847 474
[5] JH Writer LB Barber GK Brown HE Taylor RL Kiesling ML Ferrey ND Jahns 475
SE Bartell HL Schoenfuss Anthropogenic tracers endocrine disrupting chemicals and 476
endocrine disruption in Minnesota lakes Sci Total Environ 409 (2010) 100ndash111 477
[6] JS Fisher Environmental anti-androgens and male reproductive health focus on phthalates 478
and testicular dysgenesis syndrome Reproduction 127 (2004) 305ndash315 479
[7] JP Sumpter Endocrine disrupters in the aquatic environment an overview Acta Hydrochim 480
Hydrobiol 33 (2005) 9ndash16 481
23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

23
[8] AM Al-Ansari SK Atkinson JR Doyle VL Trudeau JM Blais Dynamics of uptake and 482
elimination of 17α-ethinylestradiol in male goldfish (Carassius auratus) Aquat Toxicol 483
132ndash133 (2013) 134ndash140 484
[9] LG Parks CR Lambright EF Orland LJ Guillette Jr GT Ankley LE Gray Jr 485
Masculinization of female mosquito fish in Kraft mill effluent-contaminated Fenholloway 486
River water is associated with androgen receptor agonist activity Toxicol Sci 62 (2001) 487
257ndash267 488
[10] K Cailleaud H Budzinski S Lardy S Augagneur S Barka S Souissi J Forget-Leray 489
Uptake and elimination and effect of estrogen-like contaminants in estuarine copepods an 490
experimental study Environ Sci Pollut Res 18 (2011) 226ndash236 491
[11] T Haugen E Andersson B Norberg GL Taranger The production of hermaphrodites of 492
Atlantic cod (Gadus morhua) by masculinization with orally administered 493
17α-methyltestosterone and subsequent production of all-female cod populations Aquaculture 494
311 (2011) 248-254 495
[12] El-Greisy ZA and AE El-Gamal Monosex production of tilapia Oreochromis niloticus 496
using different doses of 17α-methyltestosterone with respect to the degree of sex stability after 497
one year of treatment Egyptian Journal of Aquatic Research 38 (2012) 59-66 498
[13] BMG Blankvoort RJT Rodenburg AJ Murk JH Koeman R Schilt JMMJG Aarts 499
Androgenic activity in surface water samples detected using the AR-LUX assay indication for 500
mixture effects Environ Toxicol Pharmacol 19 (2005) 263ndash272 501
[14] EP Kolodziej JL Gray DL Sedlak Quantification of steroid hormones with pheronmonal 502
24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

24
properties in municipal wastewater effluent Environ Toxicol Chem 22 (2003) 2622ndash2629 503
[15] L Sun Y Liu X Chu J-M Lin Trace Analysis of Fifteen Androgens in Environmental 504
Waters by LC-ESI-MS-MS Combined with Solid-Phase Disk Extraction Cleanup 505
Chromatographia 9-10 (2010) 867ndash873 506
[16] Organization for Economic Cooperation and Development Final report from the OECD 507
expert consultation meeting London UK 28ndash29th October 1998 Report 9906 Environmental 508
Health and Safety Division Paris (1999) France 509
[17] GT Ankley KM Jensen MD Kahl JJ Korte EA Makynen Description and evaluation 510
of a short-term reproductive test with the fathead minnow (Pimephales promelas) Environ 511
Toxicol Chem 20 (2001) 1276ndash1290 512
[18] MW Hornung KM Jensen JJ Korte MD Kahl ED Durhan JS Denny TR Henry 513
GT Ankley Mechanistic basis for estrogenic effects in fathead minnow (Pimephales 514
promelas) following exposure to the androgen MT conversion of 17α-methyltestosterone to 515
17α-methylestradiol Aquat Toxicol 66 (2004) 15ndash23 516
[19] M Seki H Yokota H Matsubara M Maeda H Tadokoro K Kobayashi Fish full 517
life-cycle testing for androgen methyltestosterone on medaka (Oryzias latipes) Environ 518
Toxicol Chem 23 (2004) 774ndash781 519
[20] IG Kang H Yokota Y Oshimaa Y Tsuruda Y Shimasaki T Honjo The effects of 520
methyltestosterone on the sexual development and reproduction of adult medaka (Oryzias 521
latipes) Aquat Toxicol 87 (2008) 37ndash46 522
[21] SH Hori T Kodama K Tanahashi Induction of vitellogenin synthesis in goldfish by 523
massive doses of androgens Gen Comp Endocrinol 37 (1979) 306ndash320 524
25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

25
[22] M Zerulla R Laumlnge T Steger-Hartmann G Panter T Hutchinson DR Dietrich 525
Morphological sex reversal upon short-term exposure to endocrine modulators in juvenile 526
fathead minnow (Pimephales promelas) Toxicol Lett 131 (2002) 51ndash63 527
[23] TM Kortner A Arukwe Effects of 17α-methyltestosterone exposure on steroidogenesis and 528
cyclin-B mRNA expression in previtellogenic oocytes of Atlantic cod (Gadus morhua) Comp 529
Biochem Physiol Part C 146 (2007) 569ndash580 530
[24] L Andersen R Goto-Kazeto JM Trant JP Nash B Korsgaard P Bjerregaard Short-term 531
exposure to low concentrations of the synthetic androgen methyltestosterone affects 532
vitellogenin and steroid levels in adult male zebrafish (Danio rerio) Aquat Toxicol 76 (2006) 533
343ndash352 534
[25] BJ Clark DM Stocco StAR-A tissue specific acute mediator of steroidogenesis Trends 535
Endocrin Met 7 (1996) 227ndash233 536
[26] DM Stocco X Wang Y Jo PR Manna Multiple signaling pathways regulating 537
steroidogenesis and steroidogenic acute regulatory protein expression more complicated than 538
we thought Mol Endocrinol 19 (2005) 2647ndash2659 539
[27] SY Skolness EJ Durhan N Garcia-Reyero KM Jensen MD Kahl EA Makynen D 540
Martinovic-Weigelt E Perkins DL Villeneuve GT Ankley Effects of a short-term 541
exposure to the fungicide prochloraz on endocrine function and gene expression in female 542
fathead minnows (Pimephales promelas) Aquat Toxicol 103 (2011) 170ndash178 543
[28] ER Simpson MS Mahendroo GD Means MW Kilgore MM Hinshelwood S 544
Graham-Lorence B Amarneh Y Ito CR Fisher MD Michael CR Mendelson SE 545
26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

26
Bulun Aromatase cytochrome P450 the enzyme responsible for estrogen biosynthesis Endocr 546
Rev 15 (1994) 342ndash355 547
[29] GV Callard AV Tchoudakova M Kishida E Wood Differential tissue distribution 548
developmental programming estrogen regulation and promoter characteristics of cyp19 genes 549
in teleost fish J Steroid Biochem Mol Biol 1ndash5 (2001) 305ndash314 550
[30] A Arukwe Steroidogenic acute regulatory (StAR) protein and cholesterol side-chain 551
cleavage (P450scc)-regulated steroidogenesis as an organ-specific molecular and cellular 552
target for endocrine disrupting chemicals in fish Cell Biol Toxicol 24 (2008) 527ndash540 553
[31] N Sakai M Tanaka M Takahashi S Fukada JI Mason Y Nagahama Ovarian 3 554
beta-hydroxysteroid dehydrogenasedelta 5-4-isomerase of rainbow trout its cDNA cloning 555
and properties of the enzyme expressed in a mammalian cell FEBS Lett 350 (1994) 309-313 556
[32] RS Kumar S Ijiri JM Trant Changes in the expression of genes encoding steroidogenic 557
enzymes in the channel catfish (Ictalurus punctatus) ovary throughout a reproductive cycle 558
Biol Reprod 63 (2000) 1676ndash1682 559
[33] JQ Jiang DS Wang B Senthilkumaran T Kobayashi HK Kobayashi A Yamaguchi W 560
Ge G Young Y Nagahama Isolation characterization and expression of 11β-hydroxysteroid 561
dehydrogenase type 2 cDNAs from the testes of Japanese eel (Anguilla japonica) and Nile 562
tilapia (Oreochromis niloticus) J Mol Endocrinol 31 (2003) 305ndash315 563
[34] ME Baker 11β-Hydroxysteroid dehydrogenase-type 2 evolved from an ancestral 564
17β-Hydroxysteroid dehydrogenase-type 2 Biochem Biophys Res Commun (2010) 565
215ndash220 566
[35] J Zha Z Wang N Wang C Ingersoll Histological alternation and vitellogenin induction in 567
27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

27
adult rare minnow (Gobiocypris rarus) after exposure to ethynylestradiol and nonylphenol 568
Chemosphere 66 (2007) 488ndash495 569
[36] H Wang J Wang T Wu F Qin X Hu L Wang Z Wang Molecular characterization of 570
estrogen receptor genes in Gobiocypris rarus and their expression upon endocrine disrupting 571
chemicals exposure in juveniles AquatToxicol 101 (2011) 276-287 572
[37] MA Frohman MK Dush GR Martin Rapid production of full-length cDNAs from rare 573
transcripts Amplification using a single gene-specific oligonucleotide primer Proc Natl 574
Acad Sci USA 89 (1988) 8998ndash9002 575
[38] T Wu H Wang F Qin SZ Liu M Li P Xu Z Wang Expression of zona pellucida B 576
proteins in juvenile rare minnow (Gobiocypris rarus) exposed to 17α-ethinylestradiol 577
4-nonylphenol and bisphenol A Comp Biochem Physiol C 155 (2012) 259ndash268 578
[39] JD Thompson TJ Gibson F Plewniak F Jeanmougin DG Higgins The ClustalX 579
Windows interface flexible strategies for multiple sequence alignment aided by quality 580
analysis tools Nucleic Acids Res 25 (1997) 4876ndash4882 581
[40] N Saitou M Nei The Neighbor-Joining method-a new method for reconstructing 582
phylogenetic trees Mol Biol Evol 4 (1987) 406ndash425 583
[41] K Tamura J Dudley M Nei S Kumar MEGA4 Molecular Evolutionary Genetics 584
Analysis (MEGA) software version 40 Mol Biol Evol 24 (2007) 1596ndash1599 585
[42] SZ Liu F Qin HP Wang TT Wu YY Zhang Y Zheng M Li ZZ Wang Effects of 586
17α-ethinylestradiol and bisphenol A on steroidogenic messenger ribonucleic acid levels in the 587
gonads of the rare minnow Gobiocypris rarus Aquat Toxicol 10 (2012) 19ndash27 588
[43] R Rasmussen Quantification on the Light Cycler In Meuer S Wittwer C Nakagawara K 589
28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

28
(Eds) Rapid cycle real-time PCR Methods and Applications Springer Press Heidelberg 590
2001 pp 21ndash34 591
[44] J Vandesompele K De Preter F Pattyn B Poppe N Van Roy A De Paepe F Speleman 592
Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of 593
multiple internal control genes Genome Biol 3 (2002) RESEARCH0034 594
[45] N Silver S Best J Jiang SL Thein Selection of housekeeping genes for gene expression 595
studies in human reticulocytes using real-time PCR BMC mol biol 7 (2006) 33 596
[46] MW Pfaffl A Tichopad C Prgomet TP Neuvians Determination of stable housekeeping 597
genes differentially regulated target genes and sample integrity BestKeeper-Excel-based tool 598
using pair-wise correlations Biotechnol Lett 26 (2004) 509ndash515 599
[47] CL Andersen JL Jensen TF Orntoft Normalization of real-time quantitative reverse 600
transcription-PCR data A model-based variance estimation approach to identify genes suited 601
for normalization applied to bladder and colon cancer data sets Cancer Res 64 (2004) 602
5245ndash5250 603
[48] KJ Livak TD Schmittgen Analysis of relative gene expression data using real-time 604
quantitative PCR and the 2(-Delta Delta C(T)) method Methods 25 (2001) 402ndash408 605
[49] TM Montgomery AC Brown HK Gendelman M Ota ED Clotfelter Exposure to 606
17a-Ethinylestradiol Decreases Motility and ATP in Sperm of Male Fighting Fish Betta 607
splendens Environ Toxicol (2012) httpdxdoiorg101002tox21752 Advance online 608
publication 609
[50] S Pawlowski A Sauer JA Shears CR Tyler T Braunbeck Androgenic and estrogenic 610
effects of the synthetic androgen 17α-methyltestosterone on sexual development and 611
29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

29
reproductive performance in the fathead minnow (Pimephales promelas) determined using the 612
gonadal recrudescence assay Aquat Toxicol 68 (2004) 277ndash291 613
[51] AL Filby KL Thorpe G Maack CR Tyler Gene expression profiles revealing the 614
mechanisms of anti-androgen- and estrogen-induced feminization in fish Aquat Toxicol 81 615
(2007) 219ndash231 616
[52] KA Kidd PJ Blanchfield KH Mills VP Palace RE Evans JM Lazorchak RW Flick 617
Collapse of a fish population after exposure to a synthetic estrogen Proc Natl Acad Sci U S 618
A 104 (2007) 8897ndash8901 619
[53] C-B Park K Soyano S Kiros T Kitamura M Minamiyama YSuzuki Transient effects of 620
methyltestosterone injection on different reproductive parameters of the hermaphrodite fish 621
Kryptolebias marmoratus Ecotoxicology 22 (2013) 1145ndash1154 622
[54] D Baron A Fostier B Breton Y Guiguen Androgen and estrogen treatments alter steady 623
state messengers RNA (mRNA) levels of testicular steroidogenic enzymes in the rainbow trout 624
Oncorhynchus mykiss Mol Reprod Dev 71 (2005) 471ndash479 625
[55] R Urbatzka E Rocha B Reis C Cruzeiro RAF Monteiro MJ Rocha Effects of 626
ethinylestradiol and of an environmentally relevant mixture of xenoestrogens on steroidogenic 627
gene expression and specific transcription factors in zebrafish Environ Pollut 164 (2012) 628
28ndash35 629
[56] JL Hoffmann SP Torontali RG Thomason DM Lee JL Brill BB Price GJ Carr DJ 630
Versteeg Hepatic gene expression profiling using Genechips in zebrafish exposed to 631
17α-ethynylestradiol Aquat Toxicol 79 (2006) 233ndash246 632
[57] M Govoroun OM Mcmeel H DrsquoCotta MJ Ricordel T Smith A Fostier Y Guiguen 633
30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

30
Steroid enzyme gene expressions during natural and androgen-induced gonadal differentiation 634
in the rainbow trout Oncorhynchus mykiss J Exp Zool 290 (2001) 558-566 635
[58] D Baron R Houlgatte A Fostier Y Guiguen Expression profiling of candidate genes 636
during ovary-to-testis trans-differentiation in rainbow trout masculinized by androgens Gen 637
Comp Endocrinol 156 (2008) 369ndash378 638
[59] RK Bhandari M Nakamura T Kobayashi Y Nagahama Suppression of steroidogenic 639
enzyme expression during androgen-induced sex reversal in Nile tilapia (Oreochromis 640
niloticus) Gen Comp Endocrinol 145 (2006) 20ndash24 641
[60] M Burgos-Trinidad GL Youngblood MR Maroto A Scheller DM Robins AH Payne 642
Repression of cAMP-induced expression of the mouse P450 17 alpha-hydroxylaseC17-20 643
lyase gene (cyp17) by androgens Mol Endocrinol 11 (1997) 87ndash96 644
[61] Q Zhou JE Shima R Nie PJ Friel MD Griswold Androgen-regulated transcripts in the 645
neonatal mouse testis as determined through microarray analysis Biol Reprod 72 (2005) 646
1010ndash1019 647
[62] KL Parker BP Schimmer Genes essential for early events in gonadal development Ann 648
Med 34 (2002) 171ndash178 649
[63] LJ Luense A Veiga-Lopez V Padmanabhan Lane K Christenson Developmental 650
Programming Gestational Testosterone Treatment Alters Fetal Ovarian Gene Expression 651
Endocrinology December 12 (2011 ) 4974ndash4983 652
[64] M Li L Wang H Wang H Liang Y Zheng F Qin S LiuY Zhang Z Wang Molecular 653
cloning and characterization of amh dax1 and cyp19a1a genes and their response to 654
17α-methyltestosterone in Pengze crucian carp Comp Biochem Physiol Part C 157 (2013) 655
31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

31
372ndash381 656
[65] TS Kostic NJ Stojkov MM Bjelic AI Mihajlovic MM Janjic SA Andric 657
Pharmacological doses of testosterone upregulated androgen receptor and 658
3-beta-hydroxysteroid dehydrogenasedelta-5-delta-4 isomerase and impaired leydig cells 659
steroidogenesis in adult rats Toxicol Sci 121 (2011) 397ndash407 660
661
662
663
664
665
666
667
668
669
670
671
672
673
674
675
676
677
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887
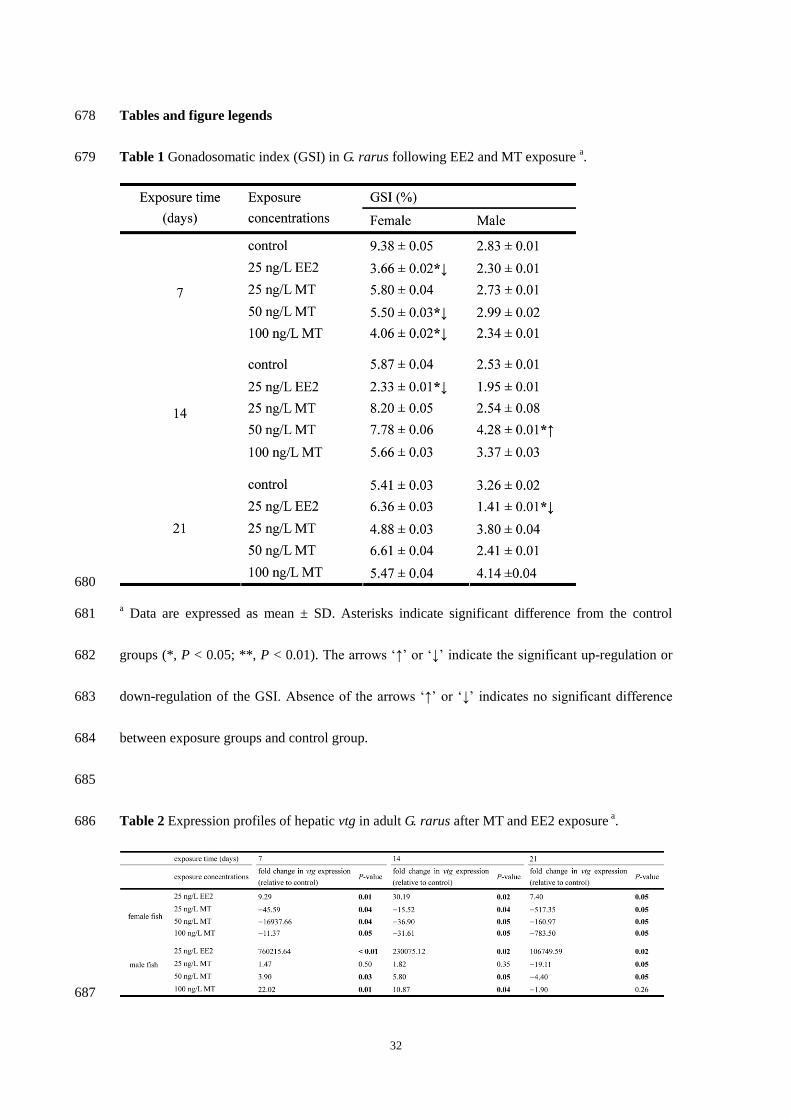
32
Tables and figure legends 678
Table 1 Gonadosomatic index (GSI) in G rarus following EE2 and MT exposure a 679
680
a Data are expressed as mean plusmn SD Asterisks indicate significant difference from the control 681
groups ( P lt 005 P lt 001) The arrows lsquouarrrsquo or lsquodarrrsquo indicate the significant up-regulation or 682
down-regulation of the GSI Absence of the arrows lsquouarrrsquo or lsquodarrrsquo indicates no significant difference 683
between exposure groups and control group 684
685
Table 2 Expression profiles of hepatic vtg in adult G rarus after MT and EE2 exposure a 686
687
33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

33
a The number indicates fold induction or repression (-) from control groups of hepatic vtg in 688
female and male G ruras P-values in bold show significant difference compared to control 689
groups (P lt 005) 690
691
Table 3 Transcriptional profiles of ovarian steroidogenic enzyme genes in female G ruras 692
exposed to MT and EE2 a 693
694
a Gene expression is expressed as the fold change compared to control groups The number 695
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 696
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in female G ruras P-values in bold show 697
significant difference compared to control groups (P lt 005) 698
699
Table 4 Transcriptional profiles of testicular steroidogenic enzyme genes in male G ruras 700
exposed to MT and EE2 a 701
702
34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

34
a Gene expression is expressed as the fold change compared to control groups The number 703
indicates fold induction or repression (-) from control groups of the six genes (StAR cyp11a1 704
3β-HSD cyp17a1 11β-HSD2 and cyp19a1a) in male G ruras P-values in bold show significant 705
difference compared to control groups (P lt 005) 706
707
Fig 1 Photomicrographs of transverse ovary sections of adult G rarus unexposed and exposed to 708
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 150 μm A1-A3 the ovary 709
of control female fish (7 14 and 21 days) B1-B3 the ovary of EE2 (25 ngL) exposure for 7 14 710
and 21 days C1-C3 the ovary of MT (25 ngL) exposure for 7 14 and 21 days D1-D3 the ovary 711
of MT (50 ngL) exposure for 7 14 and 21 days E1-E3 the ovary of MT (100 ngL) exposure for 712
7 14 and 21 days Voc vitellogenic oocyte Coc cortical alveolus stage Poc perinucleolar oocyte 713
Do degenerating oocyte 714
715
Fig 2 Photomicrographs of transverse testis sections of adult G rarus unexposed and exposed to 716
EE2 and MT using hematoxylin and eosin stain are shown Scale bars 50 μm A1-A3 the testis of 717
control group (7 14 and 21 days) B1 and B2 the testis of EE2 exposure for 7 14 days B3 the 718
testis of MT at 50 ngL for 21 days C1 and C2 the testis of MT at 50 ngL exposure for 7 14 719
days C3 the testis of MT at 100 ngL exposure for 21 days SZ spermatozoa S spermatocyte 720
AC abnormal connective V vacuolation 721
722
Fig 3 The mRNA tissue distribution of 11β-HSD2 was in G rarus different tissues (brain eye 723
gill intestine liver muscle and gonad) The results were represented the mean plusmn SD of 10 male 724
35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

35
and 10 female fish 725
726
727
728
729
730
731
732
733
734
735
736
737
738
739
740
741
742
743
744
745
746
36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

36
Fig 1 747
748
749
750
751
752
753
754
755
756
757
758
759
760
761
762
37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

37
Fig 2 763
764
765
766
767
768
769
770
771
772
773
774
775
38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

38
Fig 3 776
777
778
779
780
781
782
783
784
785
786
787
788
789
790
791
792
39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

39
Supplement tables and figure legends 793
Table S1 Primers used for fragments cloning RACE and qRT-PCR of StAR cyp11a1 3β-HSD 794
cyp17a1 11β-HSD2 and cyp19a1a 795
796
797
Table S2 Ranking output of the four reference genes according to their expression stability by 798
geNorm Comparative Delta CT method BestKeeper and NormFinder programs in G rarus 799
800
801
40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

40
Fig S1 The cloning strategy for the cDNA of 11β-HSD2 gene 802
803
Fig S2 Nucleotide sequence and deduced amino acid sequence of G rarus 11β-HSD2 Boxes 804
indicated initiator codon and stop codon Polyadenylation signal shaded of light gray 805
806
Fig S3 Comparision of 11β-HSD2 amino acid sequences among vertebrates GenBank IDs of the 807
sequences used for comparision of amino acid residues are shown as follows NP_997885 808
(zebrafish D rerio) ADI60062 (North African catfish C gariepinus) NP_001117690 (rainbow 809
trout O mykiss) BAF35260 (Japanese eel A japonica) NP_001086062 (African clawed frog X 810
laevis) AAH36780 (human H sapiens) and CAA62219 (house mouse M musculus) The 811
conserved regions of 11β-HSD2 are in boxes (I NAD-binding domain II Conserved sequence III 812
Catalytic-site) The identical highly conserved and less conserved amino acid residues were 813
indicated by () () and () respectively with the aid of an internet tool at 814
httpwwwebiacukToolsmsa muscle 815
816
Fig S4 Phylogenetic tree for G rarus 11β-HSD2 using the putative amino acid sequences The 817
phylogenetic tree were constructed using Mega 40 program [41] with neighbor-joining method 818
and bootstrap re-sampling (1000 replications) The numbers in this tree indicate the bootstrap 819
values () GenBank accession numbers of the 11β-HSD2 used in the alignment are NP_997885 820
(zebrafish D rerio) ADI60062 (North African catfish Clarias gariepinus) NP_001117690 821
(rainbow trout O mykiss) BAF35260 (Japanese eel Anguilla japonica) NP_001086062 (African 822
clawed frog Xenopus laevis) AAH36780 (human Homo sapiens) and CAA62219 (house mouse 823
41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

41
Mus musculus) 824
825
826
827
828
829
830
831
832
833
834
835
836
837
838
839
840
841
842
843
844
845
42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

42
Fig S1 846
847
848
849
850
851
852
853
854
855
856
857
858
859
860
861
862
863
864
43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

43
Fig S2 865
866
867
868
869
870
44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

44
Fig S3 871
872
873
874
875
876
877
878
879
880
881
882
883
884
885
45
Fig S4 886
887

45
Fig S4 886
887
Top Related
Copyright © 2022 FDOKUMEN

