







Bahasa
Halaman
Hukum


www.PRSJournal.com340e
Since their first description in 1997,1 evidence has accumulated to suggest that circulating endothelial progenitor cells play an impor-
tant role in the maintenance of vasculature2 and recovery of ischemic tissue.3 In murine models, endothelial progenitor cells have been shown to traffic to ischemic tissues4 and participate in new blood vessel formation, a process termed vascu-logenesis.5 However, there is controversy in the literature regarding the mechanism by which endothelial progenitor cells elicit their angiogenic
effects. Endothelial progenitor cells may either differentiate into mature endothelium and incor-porate directly into neovessels6 or, as more recent studies suggest, stimulate neovascularization by means of paracrine mechanisms.7,8
Central to their role in ischemic neovascular-ization is the ability of endothelial progenitor cells to respond to ischemic events, which is believed to occur through the activity of the chemokine stro-mal cell–derived factor-1α. Stromal cell–derived factor-1α mediates the mobilization and trafficking of stem or progenitor cells expressing the recep-tor CXCR4.9–12 We have previously demonstrated that induction of stromal cell–derived factor-1α is dependent on stabilization of hypoxia-inducible
Disclosure: The authors have no financial interest to declare in relation to the content of this article.Copyright © 2015 by the American Society of Plastic Surgeons
DOI: 10.1097/PRS.0000000000000917
Edwin Chang, Ph.D.Josemaria Paterno, M.D.Dominik Duscher, M.D.
Zeshaan N. Maan, M.B.B.S., M.S.
Jerry S. Chen, M.D.Michael Januszyk, M.D.
Melanie Rodrigues, Ph.D.Robert C. Rennert, B.A.
Sarah Bishop, M.D.,Arnetha J. Whitmore, B.S.
Alexander J. Whittam, B.S.Michael T. Longaker, M.D.,
M.B.A.Geoffrey C. Gurtner, M.D.
Stanford, Calif.
Background: Endothelial progenitor cells have been shown to traffic to and incorporate into ischemic tissues, where they participate in new blood vessel formation, a process termed vasculogenesis. Previous investigation has dem-onstrated that endothelial progenitor cells appear to mobilize from bone marrow to the peripheral circulation after exercise. In this study, the authors investigate potential etiologic factors driving this mobilization and investigate whether the mobilized endothelial progenitor cells are the same as those pres-ent at baseline.Methods: Healthy volunteers (n = 5) performed a monitored 30-minute run to maintain a heart rate greater than 140 beats/min. Venous blood samples were collected before, 10 minutes after, and 24 hours after exercise. Endothelial progenitor cells were isolated and evaluated.Results: Plasma levels of stromal cell–derived factor-1α significantly increased nearly two-fold immediately after exercise, with a nearly four-fold increase in circulating endothelial progenitor cells 24 hours later. The endothelial progen-itor cells isolated following exercise demonstrated increased colony formation, proliferation, differentiation, and secretion of angiogenic cytokines. Postex-ercise endothelial progenitor cells also exhibited a more robust response to hypoxic stimulation.Conclusions: Exercise appears to mobilize endothelial progenitor cells and augment their function by means of stromal cell–derived factor 1α–dependent signaling. The population of endothelial progenitor cells mobilized following exercise is primed for vasculogenesis with increased capacity for proliferation, differentiation, secretion of cytokines, and responsiveness to hypoxia. Given the evidence demonstrating positive regenerative effects of exercise, this may be one possible mechanism for its benefits. (Plast. Reconstr. Surg. 135: 340e, 2015.)
From the Hagey Laboratory for Pediatric Regenerative Medi-cine, Division of Plastic and Reconstructive Surgery, Depart-ment of Surgery, Stanford University School of Medicine.Received for publication May 2, 2014; accepted July 8, 2014.
Exercise Induces Stromal Cell–Derived Factor-1α–Mediated Release of Endothelial Progenitor Cells with Increased Vasculogenic Function
ExpErimEntal

Volume 135, Number 2 • Endothelial Progenitor Cell Mobilization
341e
factor-1α, a transcription factor that serves as a cen-tral regulator in the tissue response to hypoxia.4 Stromal cell–derived factor-1α concentrations are therefore greatest in the most hypoxic regions of ischemic tissue or cutaneous wounds, promoting the incorporation of endothelial progenitor cells into healing tissue.3,5,13–15 However, the charac-teristics of newly mobilized endothelial progeni-tor cells and the mechanisms that promote their release from bone marrow in response to hypoxia are poorly understood.
Physical exercise has been shown to increase the prevalence of monocytes and proangiogenic circulating cells.16,17 However, a link between phys-ical activity and changes in endothelial progeni-tor cell functionality after mobilization has never been investigated. In this study, we aimed to exam-ine the effects of exercise on endothelial progeni-tor cell mobilization and function at the cellular and molecular levels. A single bout of exercise was used to investigate the prevalence and func-tion of circulating endothelial progenitor cells in young healthy subjects. Characteristics of endo-thelial progenitor cells harvested before, immedi-ately after exercise, and 1 day later were assessed by prolonged culture in hypoxia. Using this experimental approach, we measured endothe-lial progenitor cell functionality by determining colony-forming capacity and recording changes in angiogenic cytokine profiles. We observed that exercise induced changes in the phenotype and prevalence of circulating cells. Furthermore, these changes appear to be driven by differences in hypoxia-inducible factor-1α stabilization after exercise.
PATIENTS AND METHODS
Exercise ProtocolFive male nonsmokers without any significant
medical history were recruited for this study. The average age of the subjects was 29.8 years (range, 26 to 36 years) and the average body mass index was 24.48 kg/m2 (range, 22.7 to 26.5 kg/m2). Demo-graphic details of test subjects are summarized in Table 1. All volunteers were screened by interview, and those with cardiovascular comorbidities or undergoing pharmacologic treatment for cardio-vascular disease were excluded from the study. A cohort with comparable fitness levels was selected, and informed consent was obtained from all par-ticipants. Subjects were instructed to refrain from excessive physical activity for a 72-hour period before the study. Excessive physical activity was
defined as any exercise or other strenuous actions that would induce sweating. Volunteers then per-formed a single monitored 30-minute treadmill run to maintain a heart rate above 140 beats/min. Blood samples (20 to 30 ml of venous blood) were collected by means of antecubital venipunc-ture before, 10 minutes after, and 24 hours after exercise. All blood samples were processed for complete blood count, circulating progenitor cell quantification, endothelial progenitor cell cul-ture, and plasma collection. Exercise and blood collection protocols were approved by the Stan-ford University Institutional Review Board.
Endothelial Progenitor Cell CultureEndothelial progenitor cell colonies were
cultured as described previously.18 Briefly, mono-nuclear cells were isolated from peripheral blood samples using Ficoll-Paque PLUS density gra-dient centrifugation (GE Healthcare, Newark, N.J.). Isolated cells were then seeded in Medium 199 (Gibco Life Technologies, Carlsbad, Calif.) supplemented with 20% fetal bovine serum onto fibronectin-coated six-well plates (BD Biosci-ences, San Jose, Calif.) at 5 × 106 cells/well. After 2 days in culture, nonadherent cells were trans-ferred onto 24-well fibronectin-coated plates at a density of 1 × 106 cells/well. Seeded cells were then incubated in either normoxic (21% oxygen) or hypoxic (1% oxygen) conditions at 37°C and 5% carbon dioxide for an additional 5 days, after which endothelial progenitor cell colony counts were performed. Colonies were counted manually in a minimum of four wells by blinded observers.
Endothelial Progenitor Cell Quantification by Flow Cytometry
Primary mononuclear cells and cultured endothelial progenitor cells were analyzed by flow cytometry as described previously.3,4 Viable cell populations were analyzed for CD11b-PE, CD34-APC, VEGFR2-FITC (BD Biosciences), and AC133-PE-Cy7 (Miltenyi Biotec, Inc., Auburn, Calif.). VEGFR2+/CD11b−/CD34+/AC133+ cells
Table 1. Subject Demographics
PatientAge (yr)
Weight (kg)
Height (m)
BMI (kg/m2)
1 26 70 1.67 25.12 28 65 1.65 23.93 31 80 1.82 24.24 36 68 1.73 22.75 28 83 1.77 26.5BMI, body mass index.

342e
Plastic and Reconstructive Surgery • February 2015
were defined as endothelial progenitor cells. Iso-type-identical antibodies (BD Biosciences) served as controls in each experiment. Samples were run on an LSRI analyzer (BD Biosciences) with a mini-mum of 200,000 events for mononuclear cells and 50,000 events for cultured endothelial progenitor cells. Data were analyzed using FLOWJO software (Tree Star, Inc., Ashland, Ore.).
Thymidine Incorporation AssayCultured endothelial progenitor cells (1 × 103
in 100 μl of media) were seeded per well onto a 96-well plate and incubated at 37°C overnight to allow attachment. Then, 5 μCi/ml of 3H-thymi-dine was added to each well and incubated for 6 hours. After collection of cell lysate, counts were determined by liquid scintillation.
Cytokine QuantificationVascular endothelial growth factor (VEGF),
erythropoietin, and stromal cell–derived factor-1α levels were determined with corresponding Quan-tikine enzyme-linked immunosorbent assay kits (R&D Systems, Minneapolis, Minn.) according to the manufacturer’s instructions. Assays were performed on donor plasma and conditioned media collected from endothelial progenitor cell cultures.
ImmunocytochemistryCells were washed with phosphate-buffered
saline and fixed with 4% paraformaldehyde. After multiple wash steps with phosphate-buffered saline and blocking of nonspecific binding with Powerblock (Biogenex, San Ramon, Calif.), anti-gen detection was performed with mouse anti-human CD31-PE and VEGFR2-FITC conjugated antibodies (BD Biosciences). Cell nuclei were counterstained with 4′,6-diamidino-2-phenylin-dole mounting medium (Vector Laboratories, Burlingame, Calif.). Standard immunofluores-cence imaging was performed on a Zeiss Axioplan 2 microscope (Carl Zeiss Microimaging, Thorn-wood, N.Y.). Images were prepared for publica-tion using Photoshop (Adobe Systems, San Jose, Calif.).
Quantitative Real-Time Reverse Transcription Polymerase Chain Reaction
Quantitative real-time reverse transcrip-tion polymerase chain reaction was performed with the Roche Light Cycler Sequence Detec-tion System (PE Biosystems, Foster City, Calif.). Primers for hypoxia-inducible factor-1α were
5-TTACCCACCGCTGAAACG-3 and 5-TGCTTC-CATCGGAAGGAC-3. Primers for the control housekeeping gene glyceraldehyde 3-phosphate dehydrogenase were 5-AACATCATCCCTGCCTC-TAC-3 and 5-CCCTGTTGCTGTAGCCAAAT-3. Transcription levels for hypoxia-inducible factor-1α were calculated as relative units to glyceral-dehyde 3-phosphate dehydrogenase. All primers were synthesized on request by the Peptide and Nucleic Acid Core Facility at Stanford University.
Hypoxia-Inducible Factor-1α Western BlotTwenty micrograms of total protein extracted
with radioimmunoprecipitation assay buffer (Sigma-Aldrich, St. Louis, Mo.) were separated on 4% to 12% sodium dodecyl sulfate polyacryl-amide gel electrophoresis gels (Invitrogen, Carls-bad, Calif.) and transferred to polyvinylidene difluoride membranes. Protein detection was per-formed with rabbit anti-human primary antibodies against hypoxia-inducible factor-1α (Novus, Lit-tleton, Colo.) and β-actin (Lab Vision, Fremont, Calif.). Corresponding goat anti-rabbit horserad-ish peroxidase–linked antibodies were used as the secondary antibody (Santa Cruz Biotechnologies, Santa Cruz, Calif.). Blots were developed with ECL reagent (Amersham, Piscataway, N.J.) and exposed on BioMax-MS film (Carestream Health, Inc., Rochester, N.Y.).
Statistical AnalysisUnless stated otherwise, all statistics are pre-
sented as mean ± SEM. Where applicable, univari-ate analyses were completed using an unpaired t test with Excel 2010 (Microsoft, Redmond, Wash.). Multivariate analyses were accomplished using one-way analysis of variance with Bonferroni posttest by GraphPad Prism version 4.00 (Graph-Pad Software, Inc., San Diego, Calif.). Values of p < 0.05 were considered significant.
RESULTS
Exercise Increases Plasma Stromal Cell–Derived Factor-1α and Erythropoietin Levels
A moderate exercise session of 30 minutes was chosen to evaluate the effects of an efficient and healthy level of activity that could be performed on a daily basis.19 Plasma samples collected at the three time points were assayed for the presence of the hypoxia-inducible factor-1α–inducible cyto-kines stromal cell–derived factor-1α and erythro-poietin. Stromal cell–derived factor-1α levels were present in the preexercise circulation (421 ± 98.2

Volume 135, Number 2 • Endothelial Progenitor Cell Mobilization
343e
pg/ml) and significantly increased 1.7-fold in the immediate postexercise period (717 ± 135 pg/ml; p < 0.01). Plasma stromal cell–derived factor-1α concentrations returned to baseline levels after 24 hours (489 ± 84.4 pg/ml) (Fig. 1, above, left). erythropoietin concentrations did not elevate sig-nificantly until 24 hours after exercise, increasing from 2.72 ± 0.002 mIU/ml to 3.54 ± 0.029 mIU/ml (p < 0.01) (Fig. 1, above, right).
Exercise Increases Circulating Progenitor Cell Populations
Although a universally accepted cell-surface marker profile has yet to be established, previ-ous studies have suggested that endothelial pro-genitor cells can be defined as VEGR2+/CD34+/AC133+.20,21 We have added the negative selection of CD11b to exclude monocytes that have been demonstrated to be a nonvasculogenic popula-tion present under earlier endothelial progenitor cell definitions.8 Using this panel of cell-surface markers, basal levels of endothelial progenitor cells were found to exist at a prevalence of 0.10 ± 0.02 percent of total circulating mononuclear
cells. There was no significant change in endothe-lial progenitor cell prevalence immediately after exercise (0.11 ± 0.04 percent); however, circulat-ing endothelial progenitor cell counts increased 3.6-fold 24 hours after exercise (0.36 ± 0.09 per-cent; p < 0.05) (Fig. 1, below).
Exercise Increases Endothelial Progenitor Cell Colony Formation and Proliferation
Endothelial progenitor cell colonies were quantified by counting colony-forming units after plated mononuclear cells were expanded in culture for 7 days.18 Endothelial progenitor cell colony counts were 2.1 ± 0.25 colonies/million mononuclear cells in preexercise samples. Colony counts increased 5.3-fold (11 ± 1.2 colonies/mil-lion mononuclear cells) when samples were col-lected 24 hours after exercise were compared to preexercise controls. No difference was observed in cells collected immediately after exercise (Fig. 2, left).
Cellular proliferation of endothelial pro-genitor cells in culture was quantified by a 3H-thymidine incorporation assay. Cells isolated
Fig. 1. plasma cytokine levels before and after a 30-minute exercise session. (Above, left) stromal cell–derived factor-1α (SDF-1α) levels are significantly increased immediately after exercise (p < 0.01) and return to baseline levels after 24 hours. (Above, right) Erythropoietin (EPO) remains at preexercise levels immediately after exercise and increases after 24 hours (p < 0.05). (Below) prevalence of circulating endothelial progenitor cells before and after a 30-minute exercise session. Endothelial progenitor cells were defined as VEGFr2+/CD11b−/CD34+/aC133+ and enumerated by flow cytometry. Circulating levels were unchanged immediately after exercise and increased 3.6-fold when assessed after 24 hours (p < 0.05). *p < 0.05 versus preexercise levels.

344e
Plastic and Reconstructive Surgery • February 2015
immediately after exercise demonstrated a 1.7-fold increase in proliferation compared with pre-exercise samples, whereas a 4.5-fold increase was observed in endothelial progenitor cells isolated 24 hours after exercise under normoxic condi-tions (p < 0.01) (Fig. 2, right).
We repeated these assays under hypoxic con-ditions (1% oxygen) to assess endothelial pro-genitor cell function in a setting similar to tissue ischemia. Colony counts increased over 19-fold in samples collected 24 hours after exercise (23 ± 3.8 colonies/million mononuclear cells) compared with preexercise controls (1.2 ± 0.05 colonies/mil-lion mononuclear cells; p < 0.05) when grown in hypoxia. Again, no difference in colony-forming capacity was found in cells collected immediately after exercise. We also compared colony forma-tion in hypoxia versus normoxia in cells from the three time points. Only endothelial progeni-tor cells isolated 24 hours after exercise demon-strated significantly increased colony formation in response to hypoxia (Fig. 2, left).
In the setting of hypoxia, endothelial pro-genitor cells isolated immediately after exercise demonstrated a 2.4-fold increase in proliferation compared with baseline endothelial progenitor cells (p < 0.01). An even greater 7.5-fold increase
was observed in the endothelial progenitor cells collected 24 hours after exercise (p < 0.01). Both immediate postexercise and 24-hour postexercise endothelial progenitor cell cultures demonstrated significantly increased proliferation in hypoxia versus normoxia, more so in the latter group (1.6- versus 2.2-fold increase). Preexercise endothelial progenitor cells did not demonstrate increased proliferation in hypoxia (Fig. 2, right).
Exercise Augments Endothelial Progenitor Cell Differentiation
Differentiation of endothelial progenitor cells into cells with a mature endothelial-like pheno-type was evaluated by performing flow cytometry to assess for acquisition of the endothelial-specific markers VEGFR2 and CD31 on cells after 7, 14, and 21 days in culture. VEGFR2+/CD31+ cells became more prevalent after 14 days in all sam-ples but increased significantly more in samples collected 24 hours after exercise. After 21 days, all cells again displayed increased acquisition of the endothelial cell markers. Both immediate postex-ercise and 24-hour samples displayed significantly greater differentiation than the preexercise cells; however, there was no statistical difference between the two conditions (Fig. 3, left). The
Fig. 2. Endothelial progenitor cell colony-forming ability and proliferation in vitro in response to exercise and hypoxic stimulation. (Left) Endothelial progenitor cells (EPC) collected 24 hours after exercise exhibited signifi-cantly greater colony formation in both normoxic (21% oxygen) and hypoxic (1% oxygen) culture conditions (p < 0.05). Exposure to hypoxia elicited a 2.1-fold increase in colony formation in endothelial progenitor cells collected 24 hours after exercise (p < 0.05). Cells from other time points did not demonstrate a significant response. MNCs, mononuclear cells. (Right) Cellular proliferation assessed by tritiated thymidine incorporation assay closely par-allels colony-forming data. Both postexercise and 24-hour endothelial progenitor cells demonstrated increased proliferation under normoxic and hypoxic conditions compared with the preexercise cells (p < 0.01). postexercise endothelial progenitor cells demonstrated a 1.6-fold increase in proliferation in response to hypoxia, whereas endothelial progenitor cells collected 24 hours after exercise had a 2.2-fold increase (p < 0.05). Endothelial pro-genitor cells collected before exercise did not exhibit any increased response to hypoxia. CPM, counts per minute. *p < 0.05 versus preexercise cells; ^p < 0.05 versus normoxia.

Volume 135, Number 2 • Endothelial Progenitor Cell Mobilization
345e
presence of VEGFR2+/CD31+ cells in culture was verified by immunofluorescence (Fig. 3, right).
Exercise Increases Endothelial Progenitor Cell Paracrine Activity
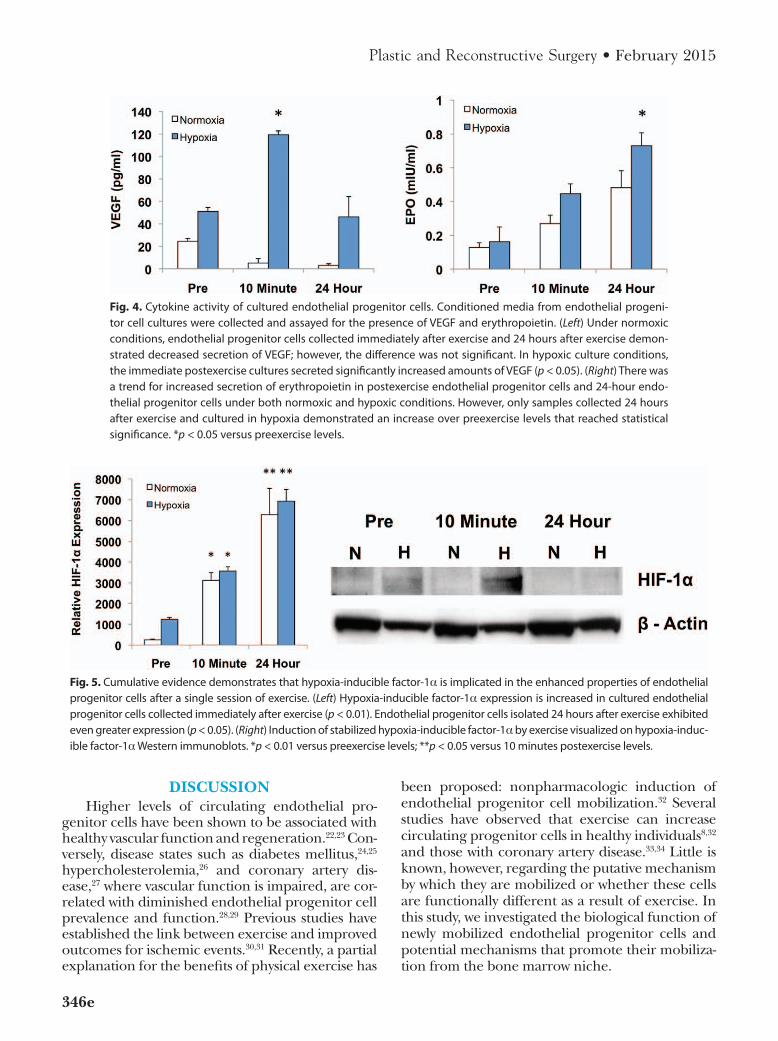
Evidence is accumulating that endothelial progenitor cells may at least in part contribute to neovascularization by means of paracrine mechanisms. Accordingly, we assessed the secre-tory activity of endothelial progenitor cells by measuring cytokine levels in conditioned media from cultures grown in normoxic and hypoxic conditions. Under normoxic conditions, there were no significant differences in VEGF pro-duction from endothelial progenitor cells col-lected at the three different time points. Within cultures exposed to hypoxia, samples collected immediately after exercise produced more than twice the amount of VEGF as the preexercise samples (119 ± 3.4 versus 51 ± 3.6 pg/ml; p < 0.05). Secreted VEGF levels from cultured endo-thelial progenitor cells harvested 1 day after exercise were statistically equivalent to the pre-exercise levels (Fig. 4, left).
There was a trend for increased secretion of erythropoietin from cells isolated immediately and 24 hours after exercise when cultured under normoxic conditions; however, this trend did not reach significance. With hypoxic stimulation, cells
collected 1 day after exercise demonstrated a 4.5-fold increase in secreted erythropoietin compared with preexercise levels (Fig. 4, right).
Exercise Increases Hypoxia-Inducible Factor-1α Expression and Stabilization
Hypoxia-inducible factor-1α expression by endothelial progenitor cells in culture was mea-sured using quantitative real-time reverse tran-scription polymerase chain reaction. Under both normoxic and hypoxic conditions, total RNA iso-lated from cultured endothelial progenitor cells demonstrated increased transcription of hypoxia-inducible factor-1α immediately after exercise compared with preexercise levels (normoxia, 3120 ± 371 versus 252.7 ± 36.2; hypoxia, 3568 ± 203 versus 1246 ± 87.6; p < 0.01). Endothelial pro-genitor cells collected 1 day after exercise exhib-ited even greater expression of hypoxia-inducible factor-1α than those harvested immediately after exercise (normoxia, 6276 ± 1266; hypoxia, 6940 ± 557; p < 0.05) (Fig. 5, left). As hypoxia-inducible factor-1α also undergoes posttranslational regula-tion by means of degradation/stabilization, these data were confirmed at the protein level using standard Western blot technique. Hypoxia-induc-ible factor-1α protein levels were increased under hypoxic stress in cells that were collected after exercise (Fig. 5, right).
Fig. 3. Endothelial differentiation of endothelial progenitor cells in culture. Endothelial progenitor cells were grown in normoxia for 21 days. (Left) Flow cytometry was used to assess cells for the endothelial-specific markers VEGFr2 and CD31. the numbers of double-positive cells at days 14 and 21 relative to day 7 are shown. VEGFr2+/CD31+ cells increased in preexercise, postexercise, and 24-hour endothelial progenitor cells at days 14 and 21. at day 14, endothelial progenitor cells isolated 1 day after exercise had a significantly greater number of differentiated cells than at the other two time points (p < 0.05). at day 21, both postexercise and day-1 endothelial progenitor cells displayed significantly greater differentiation than the preexercise endothelial progenitor cells (p < 0.05). (Right) representative image of endothelial progenitor cells labeled by immunofluorescence for VEGFr2 (green) and CD31 (red). *p < 0.05 versus preexercise cells.
346e
Plastic and Reconstructive Surgery • February 2015
DISCUSSIONHigher levels of circulating endothelial pro-
genitor cells have been shown to be associated with healthy vascular function and regeneration.22,23 Con-versely, disease states such as diabetes mellitus,24,25 hypercholesterolemia,26 and coronary artery dis-ease,27 where vascular function is impaired, are cor-related with diminished endothelial progenitor cell prevalence and function.28,29 Previous studies have established the link between exercise and improved outcomes for ischemic events.30,31 Recently, a partial explanation for the benefits of physical exercise has
been proposed: nonpharmacologic induction of endothelial progenitor cell mobilization.32 Several studies have observed that exercise can increase circulating progenitor cells in healthy individuals8,32 and those with coronary artery disease.33,34 Little is known, however, regarding the putative mechanism by which they are mobilized or whether these cells are functionally different as a result of exercise. In this study, we investigated the biological function of newly mobilized endothelial progenitor cells and potential mechanisms that promote their mobiliza-tion from the bone marrow niche.
Fig. 4. Cytokine activity of cultured endothelial progenitor cells. Conditioned media from endothelial progeni-tor cell cultures were collected and assayed for the presence of VEGF and erythropoietin. (Left) Under normoxic conditions, endothelial progenitor cells collected immediately after exercise and 24 hours after exercise demon-strated decreased secretion of VEGF; however, the difference was not significant. in hypoxic culture conditions, the immediate postexercise cultures secreted significantly increased amounts of VEGF (p < 0.05). (Right) there was a trend for increased secretion of erythropoietin in postexercise endothelial progenitor cells and 24-hour endo-thelial progenitor cells under both normoxic and hypoxic conditions. However, only samples collected 24 hours after exercise and cultured in hypoxia demonstrated an increase over preexercise levels that reached statistical significance. *p < 0.05 versus preexercise levels.
Fig. 5. Cumulative evidence demonstrates that hypoxia-inducible factor-1α is implicated in the enhanced properties of endothelial progenitor cells after a single session of exercise. (Left) Hypoxia-inducible factor-1α expression is increased in cultured endothelial progenitor cells collected immediately after exercise (p < 0.01). Endothelial progenitor cells isolated 24 hours after exercise exhibited even greater expression (p < 0.05). (Right) induction of stabilized hypoxia-inducible factor-1α by exercise visualized on hypoxia-induc-ible factor-1α Western immunoblots. *p < 0.01 versus preexercise levels; **p < 0.05 versus 10 minutes postexercise levels.

Volume 135, Number 2 • Endothelial Progenitor Cell Mobilization
347e
We first examined the temporal relationship between systemic cytokine signaling, endothe-lial progenitor cell mobilization, and endothe-lial progenitor cell activity after a brief session of aerobic exercise. A 30-minute period of exertion was shown to be sufficient to elicit a transient up-regulation of plasma stromal cell–derived factor-1α immediately after exertion followed by a series of downstream effects on endothelial progenitor cells and their biological function. Although this study did not specifically investigate the origin of this increased plasma stromal cell–derived factor-1α, the ischemic skeletal muscle environment induced by exercise may serve as a possible source of the chemokine.35 In fact, recent work has shown that hypoxia-inducible factor-1α expression and stabilization are increased in skeletal muscle after exercise36–38 and that muscle satellite cells and fibroblasts are capable of secreting stromal cell–derived factor-1α to help recruit CD34+ progeni-tor cells to ischemic tissue.35,39 Accordingly, the circulating population of endothelial progenitor cells was observed to increase significantly 1 day after exercise, suggesting a stromal cell–derived factor-1α–mediated mobilization from the bone marrow niche.
We further analyzed the effects of exercise on the functional capacity of endothelial progenitor cells, finding that both cellular proliferation and colony-forming capacity, important properties of any progenitor cell, were enhanced by aerobic activity. Previous work investigating the effects of endothelial progenitor cell therapy on endothe-lial progenitor cell function demonstrated that endothelial progenitor cells exposed to erythro-poietin exhibit PI3K/Akt-dependent increases in proliferation, colony formation, and adhesion.40,41 Our study shows that plasma levels of erythropoi-etin significantly increase in parallel with changes in endothelial progenitor cell proliferative capac-ity following exercise, suggesting a possible mech-anism for these functional changes.
The literature suggests that endothelial pro-genitor cells are capable of augmenting neovas-cularization in both animal models and humans. Currently, it remains to be elucidated whether these progenitors promote new blood vessel for-mation by means of differentiation into endothe-lial cells and engraftment into neovessels,42,43 by the release of proangiogenic soluble factors,7,8,44,45 or by a combination of both. Endothelial progeni-tor cells isolated 10 minutes and 24 hours after exercise displayed similarly increased propensi-ties for endothelial differentiation compared with preexercise samples. Endothelial progenitor cells
collected after exercise also demonstrated both increased secretion of the growth factors VEGF and erythropoietin and augmented responses to hypoxia. These findings are independent of the proliferation data, as cells were seeded in equal numbers for these assays. As circulating endo-thelial progenitor cell numbers do not increase until 1 day after exercise, the immediate systemic cytokine changes induced by exercise appear to stimulate existing circulating endothelial progen-itor cells toward a more provascular phenotype. These findings are consistent with results of ex vivo priming of endothelial progenitor cells with stromal cell–derived factor-1α, which have dem-onstrated that incubation of endothelial progeni-tor cells with recombinant stromal cell–derived factor-1α for as little as 1 minute results in dose-dependent increases in differentiation, transcrip-tion of VEGF, and enhanced therapeutic potential in a hind-limb ischemia model.46
Other work investigating the effects of exer-cise on circulating endothelial progenitor cells in healthy subjects has demonstrated immediate increases in peripheral endothelial progenitor cell levels, whereas no difference was observed in our study until 24 hours after exercise.32,47 This discrepancy between findings may be explained by differing definitions of endothelial progenitor cells. In one study, AC133+/CD144+ cells were termed endothelial progenitor cells, whereas CD34+/VEGFR2+ cells were defined as endothelial progenitor cells in another. To avoid including monocytes with an angiogenic phenotype, which have been confused with endothelial progenitor cells but which are a dis-tinct cell population,48–50 we have chosen to use VEGFR2+/CD11b−/CD34+/AC133+ as the cell-surface profile in this investigation. By depleting monocytes, we may be excluding the early-mobi-lized cells seen in other studies, but are focus-ing on a more specific endothelial progenitor cell population. Our in vitro colony-formation data closely correspond to circulating endothe-lial progenitor cell numbers, according to our definition, suggesting that this combination is a representative marker profile.
Investigators have demonstrated that aerobic exercise can lead to increased circulating levels of endothelial progenitor cells.16,32 However, the mechanism behind this mobilization and the biological characteristics of exercise-induced endothelial progenitor cells have not been exam-ined. We propose that exercise induces an acute elevation of systemic stromal cell–derived factor-1α levels that has both immediate and delayed

348e
Plastic and Reconstructive Surgery • February 2015
beneficial effects. Existing circulating endothe-lial progenitor cells become “primed” by expo-sure to this chemokine, leading to cells with increased function and expression of hypoxia-responsive genes. Later, endothelial progenitor cell numbers increase as a function of stromal cell–derived factor-1α/CXCR4–mediated mobi-lization. These newly mobilized endothelial progenitor cells are characterized by erythropoi-etin-mediated increased proliferative capacity and differentiation potential and hypoxia-induc-ible factor-1α stabilization (Fig. 6). These find-ings build on previous work by establishing that exercise is not only capable of mobilizing endo-thelial progenitor cells to the circulation, but induces progenitor cells with increased proan-giogenic characteristics. This suggests a possible mechanism for the regenerative effects seen by physical exercise and also supports the impor-tance of ongoing research in exploring ex vivo modulation of endothelial progenitor cells to maximize their potential for cell-based therapy. However, it is to be determined how baseline
fitness of different individuals and increasing lev-els of exercise would affect endothelial progeni-tor cell mobilization and function. More intense levels or longer periods of physical exercise could potentially influence these results. Further studies are needed to define exercise regimens yielding an ideal ratio of physical activity and endothelial progenitor cell mobilization that are feasible for a majority of patients.
Geoffrey C. Gurtner, M.D.Department of Surgery
Division of Plastic and Reconstructive SurgeryStanford University School of Medicine
257 Campus Drive WestHagey Building GK-201
Stanford, Calif. [email protected]
ACKNOWLEDGMENTSThis work was supported by grant no. RO1-
DK-074095 from the National Institute of Diabetes and Digestive and Kidney Diseases and grant no. RO1-AG-025016 from the National Institute on Aging.
Fig. 6. Schematic of the downstream effects of physical exercise. Exercise initiates an immediate release of stromal cell–derived factor-1α (SDF-1α) from ischemic skeletal muscle, which has both immediate and delayed effects. in the immediate postexercise period, systemic stromal cell–derived factor-1α “primes” circulating endothelial progenitor cells, augmenting their paracrine activity. a delayed effect is also observed after 24 hours with mobilization of an endothelial progenitor cell (EPC) population with increased proliferative, differentiation, and secretory capacity. HIF-1α, hypoxia-inducible factor-1α; EPO, erythropoietin.

Volume 135, Number 2 • Endothelial Progenitor Cell Mobilization
349e
REFERENCES 1. Asahara T, Murohara T, Sullivan A, et al. Isolation of puta-
tive progenitor endothelial cells for angiogenesis. Science 1997;275:964–967.
2. Rumpold H, Wolf D, Koeck R, Gunsilius E. Endothelial pro-genitor cells: A source for therapeutic vasculogenesis? J Cell Mol Med. 2004;8:509–518.
3. Tepper OM, Capla JM, Galiano RD, et al. Adult vasculogen-esis occurs through in situ recruitment, proliferation, and tubulization of circulating bone marrow-derived cells. Blood 2005;105:1068–1077.
4. Ceradini DJ, Kulkarni AR, Callaghan MJ, et al. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nat Med. 2004;10:858–864.
5. Ceradini DJ, Kulkarni AR, Callaghan MJ, et al. Progenitor cell trafficking is regulated by hypoxic gradients through HIF-1 induction of SDF-1. Nat Med. 2004;10:858–864.
6. Crosby JR, Kaminski WE, Schatteman G, et al. Endothelial cells of hematopoietic origin make a significant contribution to adult blood vessel formation. Circ Res. 2000;87:728–730.
7. Urbich C, Aicher A, Heeschen C, et al. Soluble factors released by endothelial progenitor cells promote migration of endothelial cells and cardiac resident progenitor cells. J Mol Cell Cardiol. 2005;39:733–742.
8. Rehman J, Li J, Orschell CM, March KL. Peripheral blood “endothelial progenitor cells” are derived from mono-cyte/macrophages and secrete angiogenic growth factors. Circulation 2003;107:1164–1169.
9. Chavakis E, Urbich C, Dimmeler S. Homing and engraft-ment of progenitor cells: A prerequisite for cell therapy. J Mol Cell Cardiol. 2008;45:514–522.
10. Yin Y, Huang L, Zhao X, et al. AMD3100 mobilizes endothe-lial progenitor cells in mice, but inhibits its biological func-tions by blocking an autocrine/paracrine regulatory loop of stromal cell derived factor-1 in vitro. J Cardiovasc Pharmacol. 2007;50:61–67.
11. Hristov M, Zernecke A, Liehn EA, Weber C. Regulation of endothelial progenitor cell homing after arterial injury. Thromb Haemost. 2007;98:274–277.
12. Wojakowski W, Tendera M. Mobilization of bone marrow-derived progenitor cells in acute coronary syndromes. Folia Histochem Cytobiol. 2005;43:229–232.
13. Yin Y, Huang L, Zhao X, et al. AMD3100 mobilizes endothe-lial progenitor cells in mice, but inhibits its biological func-tions by blocking an autocrine/paracrine regulatory loop of stromal cell derived factor-1 in vitro. J Cardiovasc Pharmacol. 2007;50:61–67.
14. Hristov M, Zernecke A, Liehn EA, Weber C. Regulation of endothelial progenitor cell homing after arterial injury. Thromb Haemost. 2007;98:274–277.
15. Wojakowski W, Tendera M. Mobilization of bone marrow-derived progenitor cells in acute coronary syndromes. Folia Histochem Cytobiol. 2005;43:229–232.
16. Laufs U, Werner N, Link A, et al. Physical training increases endothelial progenitor cells, inhibits neointima formation, and enhances angiogenesis. Circulation 2004;109:220–226.
17. Rehman J, Li J, Parvathaneni L, et al. Exercise acutely increases circulating endothelial progenitor cells and mono-cyte-/macrophage-derived angiogenic cells. J Am Coll Cardiol. 2004;43:2314–2318.
18. Hill JM, Zalos G, Halcox JP, et al. Circulating endothelial progenitor cells, vascular function, and cardiovascular risk. N Engl J Med. 2003;348:593–600.
19. Gram AS, Bønnelycke J, Rosenkilde M, et al. Compliance with physical exercise: Using a multidisciplinary approach
within a dose-dependent exercise study of moderately over-weight men. Scand J Public Health 2014;42:38–44.
20. Gehling UM, Ergün S, Schumacher U, et al. In vitro differen-tiation of endothelial cells from AC133-positive progenitor cells. Blood 2000;95:3106–3112.
21. Peichev M, Naiyer AJ, Pereira D, et al. Expression of VEGFR-2 and AC133 by circulating human CD34(+) cells identifies a population of functional endothelial precursors. Blood 2000;95:952–958.
22. Kawamoto A, Asahara T. Role of progenitor endothelial cells in cardiovascular disease and upcoming therapies. Catheter Cardiovasc Interv. 2007;70:477–484.
23. Povsic TJ, Goldschmidt-Clermont PJ. Endothelial progeni-tor cells: Markers of vascular reparative capacity. Ther Adv Cardiovasc Dis. 2008;2:199–213.
24. Tepper OM, Galiano RD, Capla JM, et al. Human endothe-lial progenitor cells from type II diabetics exhibit impaired proliferation, adhesion, and incorporation into vascular structures. Circulation 2002;106:2781–2786.
25. Loomans CJ, de Koning EJ, Staal FJ, et al. Endothelial pro-genitor cell dysfunction: A novel concept in the pathogen-esis of vascular complications of type 1 diabetes. Diabetes 2004;53:195–199.
26. Chen JZ, Zhang FR, Tao QM, Wang XX, Zhu JH, Zhu JH. Number and activity of endothelial progenitor cells from peripheral blood in patients with hypercholesterolaemia. Clin Sci (Lond.) 2004;107:273–280.
27. Vasa M, Fichtlscherer S, Aicher A, et al. Number and migra-tory activity of circulating endothelial progenitor cells inversely correlate with risk factors for coronary artery dis-ease. Circ Res. 2001;89:E1–E7.
28. Rauscher FM, Goldschmidt-Clermont PJ, Davis BH, et al. Aging, progenitor cell exhaustion, and atherosclerosis. Circulation 2003;108:457–463.
29. Xiao Q, Kiechl S, Patel S, et al. Endothelial progenitor cells, cardiovascular risk factors, cytokine levels and atherosclero-sis: Results from a large population-based study. PLoS One 2007;2:e975.
30. Blumenthal JA, Sherwood A, Babyak MA, et al. Effects of exercise and stress management training on markers of car-diovascular risk in patients with ischemic heart disease: A randomized controlled trial. JAMA 2005;293:1626–1634.
31. Gordon NF, Gulanick M, Costa F, et al.; American Heart Association Council on Clinical Cardiology, Subcommittee on Exercise, Cardiac Rehabilitation, and Prevention; the Council on Cardiovascular Nursing; the Council on Nutrition, Physical Activity, and Metabolism; and the Stroke Council. Physical activity and exercise recommendations for stroke survivors: An American Heart Association scien-tific statement from the Council on Clinical Cardiology, Subcommittee on Exercise, Cardiac Rehabilitation, and Prevention; the Council on Cardiovascular Nursing; the Council on Nutrition, Physical Activity, and Metabolism; and the Stroke Council. Stroke 2004;35:1230–1240.
32. Rehman J, Li J, Parvathaneni L, et al. Exercise acutely increases circulating endothelial progenitor cells and mono-cyte-/macrophage-derived angiogenic cells. J Am Coll Cardiol. 2004;43:2314–2318.
33. Steiner S, Niessner A, Ziegler S, et al. Endurance training increases the number of endothelial progenitor cells in patients with cardiovascular risk and coronary artery disease. Atherosclerosis 2005;181:305–310.
34. Adams V, Lenk K, Linke A, et al. Increase of circulating endothelial progenitor cells in patients with coronary artery disease after exercise-induced ischemia. Arterioscler Thromb Vasc Biol. 2004;24:684–690.

350e
Plastic and Reconstructive Surgery • February 2015
35. Ratajczak MZ, Majka M, Kucia M, et al. Expression of func-tional CXCR4 by muscle satellite cells and secretion of SDF-1 by muscle-derived fibroblasts is associated with the presence of both muscle progenitors in bone marrow and hematopoietic stem/progenitor cells in muscles. Stem Cells 2003;21:363–371.
36. Gustafsson T, Puntschart A, Kaijser L, Jansson E, Sundberg CJ. Exercise-induced expression of angiogenesis-related transcription and growth factors in human skeletal muscle. Am J Physiol. 1999;276:H679–H685.
37. Ameln H, Gustafsson T, Sundberg CJ, et al. Physiological activation of hypoxia inducible factor-1 in human skeletal muscle. FASEB J. 2005;19:1009–1011.
38. Lundby C, Gassmann M, Pilegaard H. Regular endurance training reduces the exercise induced HIF-1alpha and HIF-2alpha mRNA expression in human skeletal muscle in nor-moxic conditions. Eur J Appl Physiol. 2006;96:363–369.
39. De Falco E, Porcelli D, Torella AR, et al. SDF-1 involvement in endothelial phenotype and ischemia-induced recruitment of bone marrow progenitor cells. Blood 2004;104:3472–3482.
40. George J, Goldstein E, Abashidze A, et al. Erythropoietin pro-motes endothelial progenitor cell proliferative and adhesive properties in a PI 3-kinase-dependent manner. Cardiovasc Res. 2005;68:299–306.
41. Bahlmann FH, DeGroot K, Duckert T, et al. Endothelial pro-genitor cell proliferation and differentiation is regulated by erythropoietin. Kidney Int. 2003;64:1648–1652.
42. Hur J, Yoon CH, Kim HS, et al. Characterization of two types of endothelial progenitor cells and their different contri-butions to neovasculogenesis. Arterioscler Thromb Vasc Biol. 2004;24:288–293.
43. Yoon CH, Hur J, Park KW, et al. Synergistic neovasculariza-tion by mixed transplantation of early endothelial progenitor cells and late outgrowth endothelial cells: The role of angio-genic cytokines and matrix metalloproteinases. Circulation 2005;112:1618–1627.
44. Di Santo S, Yang Z, Wyler von Ballmoos M, et al. Novel cell-free strategy for therapeutic angiogenesis: In vitro generated conditioned medium can replace progenitor cell transplan-tation. PLoS One 2009;4:e5643.
45. Gnecchi M, Zhang Z, Ni A, Dzau VJ. Paracrine mecha-nisms in adult stem cell signaling and therapy. Circ Res. 2008;103:1204–1219.
46. Zemani F, Silvestre JS, Fauvel-Lafeve F, et al. Ex vivo prim-ing of endothelial progenitor cells with SDF-1 before trans-plantation could increase their proangiogenic potential. Arterioscler Thromb Vasc Biol. 2008;28:644–650.
47. Van Craenenbroeck EM, Vrints CJ, Haine SE, et al. A maximal exercise bout increases the number of circulat-ing CD34+/KDR+ endothelial progenitor cells in healthy subjects: Relation with lipid profile. J Appl Physiol (1985) 2008;104:1006–1013.
48. Vaughan EE, O’Brien T. Isolation of circulating angiogenic cells. Methods Mol Biol. 2012;916:351–356.
49. Zhang Q, Kandic I,Barfield JT, Kutryk MJ. Coculture with late, but not early, human endothelial progenitor cells up regulates IL-1 beta expression in THP-1 monocytic cells in a paracrine manner. Stem Cells Int. 2013;2013:859643.
50. Deak E, Göttig S, Rüster B, et al. Bone marrow derived cells in the tumour microenvironment contain cells with primitive haematopoietic phenotype. J Cell Mol Med. 2010;14:1946–1952.
Top Related
Copyright © 2022 FDOKUMEN

