







Bahasa
Halaman
Hukum


Published Ahead of Print 31 March 2008. 10.1128/MCB.00091-08.
2008, 28(11):3767. DOI:Mol. Cell. Biol. Vikrant Singh and Madhulika Srivastava Barrier Establishment
Is Independent of ChromatinH19-ICRat Enhancer Blocking Activity of the Insulator
http://mcb.asm.org/content/28/11/3767Updated information and services can be found at:
These include:
REFERENCEShttp://mcb.asm.org/content/28/11/3767#ref-list-1at:
This article cites 40 articles, 21 of which can be accessed free
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
on A
ugust 8, 2014 by guesthttp://m
cb.asm.org/
Dow
nloaded from

MOLECULAR AND CELLULAR BIOLOGY, June 2008, p. 3767–3775 Vol. 28, No. 110270-7306/08/$08.00�0 doi:10.1128/MCB.00091-08Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Enhancer Blocking Activity of the Insulator at H19-ICR IsIndependent of Chromatin Barrier Establishment�
Vikrant Singh and Madhulika Srivastava*National Institute of Immunology, Aruna Asaf Ali Road, New Delhi 110067, India
Received 17 January 2008/Returned for modification 6 February 2008/Accepted 18 March 2008
Transcriptional insulators are cis regulatory elements that organize chromatin into independently regulateddomains. At the imprinted murine Igf2/H19 locus, the H19-ICR insulator prevents the activation of the Igf2promoter on the maternal allele by enhancers that activate H19 on the same chromosome. Given the well-demonstrated role of H19-ICR as an enhancer blocker, we investigated its ability to define a chromatin barrier,as the two activities are coincident on several insulators and may act in concert to define a functionalchromatin boundary between adjacent genes with distinct transcriptional profiles. Allele-specific association ofposttranslationally modified histones, reflecting the presence of active or inactive chromatin, was analyzed inthe region encompassing H19-ICR using chromatin immunoprecipitation. The existence of differential histonemodifications upstream and downstream of H19-ICR specifically on the maternal chromosome was observed,which is suggestive of a chromatin barrier formation. However, H19-ICR deletion analysis indicated thatdistinct chromatin states exist despite the absence of an intervening “barrier.” Also, the enhancers can activatethe Igf2 promoter despite some parts of the intervening chromatin being in the silent state. Hence, H19-ICRinsulator activity is not dependent on preventing the enhancer-mediated alteration of the histone modificationsin the region between the Igf2 promoter and the cognate enhancers.
Transcription is regulated by appropriate interactions of var-ious cis- and trans-acting factors and hence is influenced bychromatin structure and organization. It is intriguing indeedthat the cis regulatory elements like enhancers can exert theirinfluence on promoters and functionally interact with themspecifically, despite being separated by several kilobases ofDNA. In this context, insulators, the cis-acting elements thatcan curtail the activity of enhancers and/or prevent the hetero-chromatin spread in a position-dependent manner, are be-lieved to play a crucial role in gene regulation (11). Insulatorshave been identified in the genomes of several organisms,including Saccharomyces cerevisiae, Drosophila, chickens, andmammals. Genome-wide analysis has predicted the presenceof nearly 14,000 sites in the human genome that could poten-tially act as insulators (16, 37). However, the mechanisms un-derlying insulator, enhancer, and silencer activities and theirinteractions are only partially understood.
Several mechanisms have been proposed for the enhancer-blocking activity of the insulators based on analysis of thechicken � globin insulator (cHS4) and Drosophila insulatorsgypsy and scs-scs�. Considering the looping model of enhanceraction, insulators might create chromatin loops due to protein-mediated tethering of insulator regions to specific subnuclearstructures like the nucleolus (39) or the nuclear periphery, thusrestricting the enhancer activity to promoters that share thesame loop (11). In view of the tracking model of enhanceractivity, insulators may prevent the spreading of some positiveactivating signals that are initiated at the enhancer andprogress till the cognate promoter, leading to its activation. In
support of this hypothesis, the cHS4 insulator from the chicken� globin locus prevented the spread of histone acetylation andinterfered with RNA polymerase II transfer between the en-hancer and the promoter in an analysis relying on minichro-mosomes (40). Thus, it has been suggested that activatingmodifications may be blocked by enhancer blockers just asheterochromatin spread is stopped by barrier elements (33). Inthis context, it is interesting to note that several insulators havea combination of enhancer-blocking and chromatin barrieractivities when tested in vitro (36). Importantly, the proposedmechanisms are not mutually exclusive and may be contextdependent (11).
An insulator regulates imprinted monoallelic expression ofthe mammalian Igf2 gene. A differentially methylated region(DMR), a complex regulatory region located upstream of theH19 gene (Fig. 1), governs several aspects of the parentalallele-specific expression of the Igf2 and H19 genes that areactivated by the shared enhancers. The DMR acts as an im-print control region (ICR) and harbors an insulator. MurineH19-ICR has four methylation-sensitive sites that bind CTCF,a Zn finger protein important for insulator function (2, 13).The unmethylated DMR on the maternal allele binds CTCFand organizes a functional insulator responsible for preventingmaternal Igf2 expression. Methylation of the DMR on thepaternal chromosome abrogates CTCF binding and insulatorfunction (9), leading to transcription of paternal Igf2. Deletionof the insulator carrying DMR (14, 30, 31) and site-specificmutagenesis of the CTCF sites (26) lead to activation of ma-ternal Igf2, demonstrating a crucial role of enhancer blockingby the insulator for Igf2 regulation. The DMR also harbors asilencer active only in its methylated state on the paternal alleleand responsible for silencing downstream H19 (30). In thepaucity of CTCF binding due to mutagenesis, the maternalallele acquires a paternal epigenotype which is hypermethyl-
* Corresponding author. Mailing address: National Institute of Im-munology, Aruna Asaf Ali Road, New Delhi 110067, India. Phone:91-11-2670-3782. Fax: 91-11-2671-7104. E-mail: [email protected].
� Published ahead of print on 31 March 2008.
3767
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

ated at the ICR, loses its insulator function, and leads tochromatin alterations at the H19 and Igf2 promoters (9, 12),leading to altered transcriptional profiles for the two genes.
Insulators have been proposed to organize the eukaryoticgenome into independently regulated domains. About 14,000sites in the human genome have been predicted that couldpotentially act as insulators (16, 37) because of their ability tobind CTCF. Also, CTCF sites have been found close to chro-matin transition points in the genome-wide analysis of histonemodifications (1) and at boundaries of several genes that es-cape X inactivation (10), implicating CTCF in barrier functiondirectly or indirectly. However, detailed analysis of the chro-matin structure at the � globin loci of mice and humans dem-onstrates that, unlike the chicken � globin locus, CTCF sitesflanking the mammalian � globin loci do not act as sites formajor histone modification transitions (5, 15) even though theyare capable of enhancer-blocking function in vitro. Also, de-spite the deletion of HS62.5 and 3�HS1 regions that bindCTCF and flank the mouse � globin locus, no significantchanges were observed in either the chromatin structure ortranscriptional profile (3) compared to those of the wild type,raising doubts about the endogenous role of CTCF sites forenhancer blocking as well as chromatin barrier formation atthis locus in mammals. In order to dissect the interdependenceof enhancer blocking and chromatin barrier formation, it maybe useful to investigate the chromatin barrier function of aninsulator which has a demonstrated role in CTCF-dependentenhancer blocking.
Since the H19-ICR insulator has a well-established role as aCTCF-dependent enhancer blocker that regulates Igf2 expres-sion in an endogenous context, we used this system to test the
relevance of chromatin barrier establishment in conjunctionwith enhancer blocking for defining functionally distinct chro-matin domains in vivo. Our studies indicate remarkable allele-specific differences in the DMR suggestive of a chromatinbarrier on the maternal chromosome that carries the func-tional insulator. However, the analysis of DMR deletion mu-tants (DMRdelG), a definitive test for the endogenous barrierfunction, indicated the absence of a functional chromatin bar-rier. Our results also suggested that the enhancers can activatepromoters despite the presence of silent chromatin in the in-tervening region, in accordance with the looping model ofenhancer action, although the position dependence of insula-tor action is expected to involve some form of tracking mech-anism also.
MATERIALS AND METHODS
Experiments using mice were conducted as approved by the InstitutionalAnimal Ethics Committee.
Analysis of Igf2 and H19 expression. Total mRNA was isolated from primarymouse embryonic fibroblasts (PMEF) derived from embryos of the requiredgenotypes, and Igf2 and H19 were detected by Northern hybridization usingstandard protocols. Elongation factor (EF) was used as the loading control.
Allele-specific Igf2 expression was analyzed using single-nucleotide primerextension (Snupe) assays in RNA isolated from PMEF. For paternal inheritanceof the DMR deletion, interspecific matings were set up such that the Igf2/H19locus had a paternally inherited domesticus allele (wild type or DMRdelG mu-tant) and a maternally inherited castaneus allele (wild type). For affecting ma-ternal inheritance of the DMR deletion, reverse crosses were set up such that thematernally inherited allele was domesticus (wild type or DMRdelG mutant) whilethe paternal allele was castaneus (wild type). PMEF were generated from theresulting embryos at embryonic day 13.5. Total mRNA of the PMEF was con-verted to cDNA, which was then subjected to Snupe analysis as described pre-viously (30) to estimate the allele-specific contribution of Igf2 to total mRNA.Control reactions with genomic DNA were used to verify the incorporation of[�-32P]dATP by castaneus DNA, of [�-32P]dGTP by domesticus DNA, and ofboth nucleotides by DNA of F1 progeny of the interspecific cross.
ChIP. Chromatin immunoprecipitation (ChIP)-grade antibodies anti-dimethylH3K4 (catalogue number 07-030), anti-acetyl H3K9 (catalogue number 07-352),and anti-dimethyl H3K9 (catalogue number 07-441) were from Upstate Biotech-nology.
For ChIP with anti-dimethyl-H3K4 and anti-acetyl-H3K9 antibodies, the Up-state Biotechnology protocol was followed with some modifications. Briefly, 107
PMEF were cross-linked in 1% formaldehyde in Dulbecco’s phosphate-bufferedsaline for 5 min at 25°C. Similarly, a single-cell suspension from about 100 mg ofneonatal (postnatal day 4) liver was cross-linked. In either case, the reaction wasquenched by glycine (0.125 M). Sonication was carried out in lysis buffer (1%sodium dodecyl sulfate [SDS], 10 mM EDTA, 50 mM Tris-Cl, pH 8.0) to shearthe chromatin to an average DNA fragment size of 200 to 250 bp. Preclearedchromatin was incubated overnight with the antibody. Chromatin from approx-imately 2 � 106 cells and 10 �l of the antibody was used for each ChIP reaction.The immune complexes were collected on protein A Sepharose beads andwashed appropriately before elution. The cross-links were reversed, and theimmunoprecipitated DNA was purified for analysis.
For ChIP with anti-dimethyl-histone H3K9 antibody, cells were cross-linked asabove and sonicated to an average DNA fragment size of 200 to 250 bp in lysisbuffer (0.1% SDS, 50 mM Tris-Cl, pH 8.0, 140 mM NaCl, 1 mM EDTA, 1%Triton X-100, 0.1% sodium deoxycholate) as per the ChIP protocol by Kouskoutiand Kyrmizi (http://www.epigenome-noe.net/researchtools/protocols.php; PROT11).Subsequent steps of a typical ChIP were followed using buffers as described in theprotocol. Immune complexes were eluted, cross-links were reversed, and DNA waspurified for analysis.
For each ChIP, a fraction of the “input” chromatin was also processed forDNA purification. A mock immunoprecipitation without antibody was carriedout in parallel (no antibody control). The results shown are representative of twoto three independent ChIPs for each antibody.
Real-time quantitative PCR for quantitation of immunoprecipitated DNA.Real-time PCRs were carried out on an ABI PRISM SDS 7000 system (AppliedBiosystems) using SYBR green as the reporter dye. All PCRs were done intriplicate. The crossover threshold value (CT) was used for quantitation using a
FIG. 1. Map of the Igf2/H19 locus. (A) Schematic map of the locusshowing relative positions of genes and regulatory elements.(B) Genomic regions deleted in the mouse mutants used in this study.The region deleted in mutant alleles DMRdelG and H19del13 isshown as the gray box in each case in reference to the wild-type allele(top line). Four black columns above the DMR represent the CTCFbinding sites. (C) Relative positions of the amplicons used for ChIPanalysis (primer sequences in Table 1). Restriction sites EcoRI (E),BamHI (B), HindIII (H), XbaI (X), and SalI (S) are depicted. TheH19 transcription start site is shown as a forward arrow above H19.
3768 SINGH AND SRIVASTAVA MOL. CELL. BIOL.
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

comparative CT method. The enrichment of an amplicon in the immunoprecipi-tated sample was calculated using the following expression: (DNA after ChIP)/(DNA in input fraction) � 2�[CT(ChIP)�CT(input)] � 2�CT.
The enrichment in the test amplicon was normalized to the enrichment ob-tained in the same ChIP experiment for the � actin amplicon. All the values inFig. 3, 4, and 5 are expressed as enrichment relative to the enrichment of the �actin amplicon. Primer sequences used for PCR are given in Table 1.
RESULTS
The H19-ICR insulator, located upstream of H19 and about80 kb downstream of Igf2 (Fig. 1), is crucial for preventingmaternal Igf2 expression in several tissues, including neonatalliver, heart, and skeletal muscle. Based on deletion analysis,H19-ICR has been inferred to be a cis-acting regulatory ele-ment such that its deletion from the paternal chromosomedoes not alter the monoallelic expression pattern of Igf2 but itsdeletion from the maternal chromosome leads to activation ofmaternal Igf2, thus rendering the Igf2 expression biallelic (14,30, 31). We considered PMEF to be amenable for ChIP anal-ysis and hence ascertained the role of the H19-ICR insulator incontrolling the imprinted expression of Igf2 in PMEF. Similarto the expression in other tissues, Igf2 was expressed in wild-type PMEF and in PMEF derived from the mutants inheritingthe H19-ICR-deleted DMRdelG allele paternally (Fig. 2A).The contributions of paternal and maternal alleles in the totalIgf2 RNA of PMEF were analyzed in DMRdelG mutants,semiquantitatively, using Snupe reactions. The Igf2 expressionwas observed to be from the paternal allele in the wild type andin cases where the DMRdelG mutation was inherited pater-nally, but it was biallelic upon maternal inheritance of theDMRdelG mutation (Fig. 2C), indicating that the imprintedmonoallelic expression in PMEF is regulated by the H19-ICR
insulator in cis as in other tissues examined previously (14,30, 31).
Also, H19-ICR is responsible for regulating imprinted ex-pression of H19, and deletion of H19-ICR from the paternalallele leads to robust activation of normally silent paternal H19(14, 30, 31). However, in PMEF the paternal H19 activation
FIG. 2. Expression analysis of Igf2 and H19 in PMEF of wild type andDMRdelG mutants. (A) Northern blot to detect Igf2 expression in thewild type (lanes 1 and 2, genotype �/�) and in mutants inheritingDMRdelG on the paternal allele (lanes 3 and 4, genotype �/DMRdelG).(B) Northern blot to detect H19 expression from the wild-type maternalallele (lanes 1 and 2, genotype �/H19del13), wild-type paternal allele(lanes 3 and 4, genotype H19del13/�), DMR-deleted maternal allele(lanes 5 and 6, genotype DMRdelG/H19del13), and DMR-deleted pater-nal allele (lanes 7 and 8, genotype H19del13/DMRdelG). Elongationfactor (EF) was used as a loading control for the analysis, and the numbersbelow each lane depict the ratio of Igf2 to EF in panel A and H19 to EFin panel B. (C) Effect of DMRdelG mutation on allele-specific expressionof Igf2 analyzed by Snupe analysis. The top panel depicts the controlreactions with genomic DNA. Incorporation of [�-32P]dATP by castaneusDNA (lane 1) and [�-32P]dGTP by domesticus DNA (lane 2) and of bothnucleotides by DNA of F1 progeny of the interspecific cross (lanes 3, 4,and 5). The bottom left panel shows the absence of any effect on the alleliccontribution to Igf2 mRNA due to paternal inheritance of DMRdelGanalyzed in the PMEF of embryos derived from interspecific mating suchthat the Igf2/H19 locus has a paternally inherited domesticus allele (wildtype or DMRdelG mutant) and a maternally inherited castaneus allele(wild type). Wild-type cDNA (lane 6, genotype �/�) incorporated pri-marily [�-32P]dGTP due to monoallelic expression of Igf2 from the pa-ternal domesticus allele. In the paternal mutant cDNA (lane 7, genotype�/DMRdelG), the allelic contribution was not altered. The bottom rightpanel shows the activation of maternal Igf2 upon inheritance ofDMRdelG maternally analyzed in the PMEF of embryos derived frominterspecific mating such that the Igf2/H19 locus has a maternally inher-ited domesticus allele (wild type or DMRdelG mutant) and a paternallyinherited castaneus allele (wild type). Wild-type cDNA (lane 8, genotype�/�) incorporated primarily [�-32P]dATP due to monoallelic paternalexpression of Igf2 while maternal mutant cDNA (lane 9, genotypeDMRdelG/�) incorporated both [�-32P]dATP and [�-32P]dGTP, dem-onstrating biallelic expression of Igf2. The genotypes of the PMEF result-ing from interspecific matings for each analysis are mentioned below eachlane: cas, castaneus; dom, domesticus.
TABLE 1. Sequences of the primers used for ChIP analysis
Amplicona Primer Primer sequence
PrA M239 5� GCTCCCTATTCCCCGATGAG 3�M240r 5� GAGCCACAAGATGGCAGGA 3�
PrB M241 5� GCTCTCACTTGGAGGGCTTGT 3�M242r 5� GCCTTGTGGATTGTTCCTGG 3�
PrC M255 5� ACTCATACCAGCTGCGGGTC 3�M256r 5� AGCCTTCCCAAGAGTTCTGCT 3�
PrD M261 5� TGGGTAATACCTCTGAGCCTGC 3�M262r 5� CCCAGTGGTGTCTGTAATCTGG 3�
PrE M225 5� CAGGGCTCAGACTAGGCTCTCA 3�M226r 5� TGAGGACTAGGTGATGGCTAGCA 3�
PrF M237 5� CTGGTGATTTGCGCTTTCGT 3�M238r 5� CCCCATTGAGAGAGAATGCAGT 3�
PrG M233 5� TCGGTTATAGGCGGGAGACATA 3�M234r 5� CCTAGGTTACCTGGGACATTGC 3�
PrH M231 5� CGCAGAGATAGCCACAACCA 3�M232r 5� CTGACGGACACCTCCAATTTG 3�
PrI M251 5� GAAAAGGGACGGTGAGGAGTG 3�M252r 5� AACCCCCAGCCTTTGTCCTA 3�
PrJ M265 5� CACCTCGGCATCTGGAGTCT 3�M266r 5� CAGCTTCACTCCGATGTTTTCC 3�
� actin M431 5� CCTAAGGCCAACCGTGAAAAG 3�M432r 5� GCTGAGAAGCTGGCCAAAGA 3�
Ig M433 5� AGCCTGCCCTAGACAAACCTT 3�M434r 5� CCCTTCACTCAACCCCCATAC 3�
a Amplicon positions relative to the H19 transcription start site are shown inFig. 1.
VOL. 28, 2008 ABSENCE OF CHROMATIN BARRIER AT THE H19-ICR INSULATOR 3769
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

due to H19-ICR deletion was hardly detectable (Fig. 2B). Sur-prisingly, the expression of H19 was almost entirely abrogatedin the PMEF of deletion mutants when the deletion was in-herited maternally (Fig. 2B). This is in contrast to observationsmade earlier in neonatal liver, skeletal muscle, and heart,where the H19-ICR deletion from the maternal allele does notabolish H19 expression. This suggests that some aspects of theregulation of H19 expression in PMEF are distinct from ex-pression in other tissues.
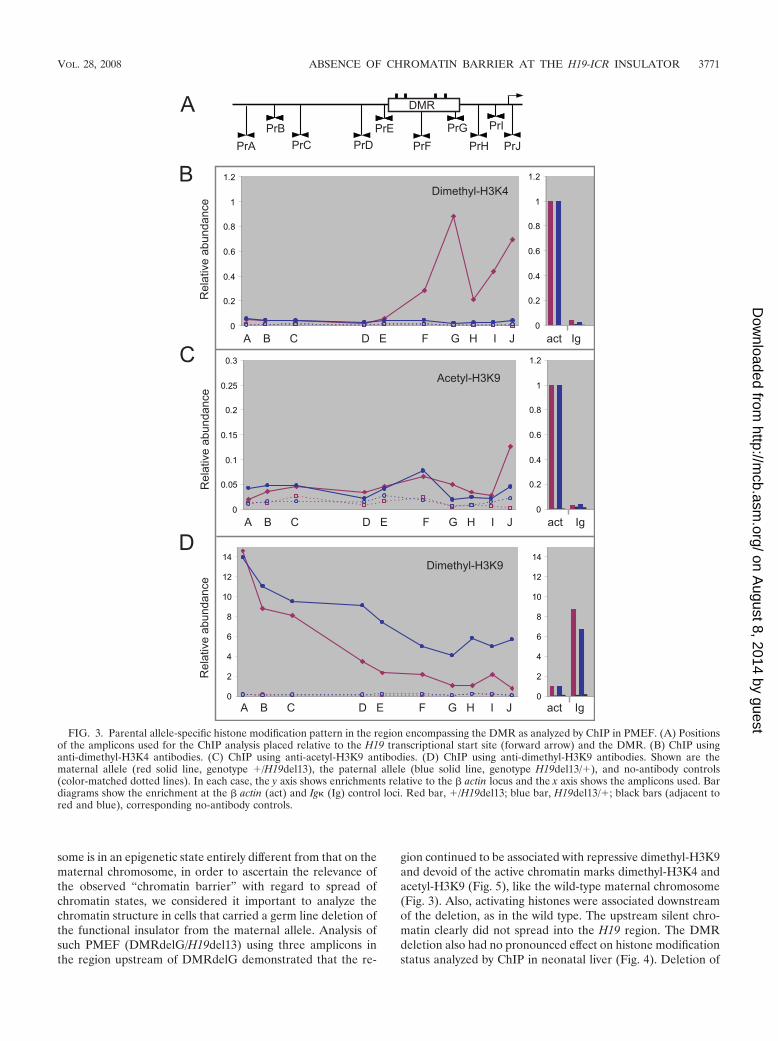
To investigate the ability of the DMR to create a chromatinbarrier, high-resolution ChIP (17) against posttranslationalmodifications of histones was carried out in PMEF and neo-natal liver. Associations of dimethyl-H3K4 and acetyl-H3K9were chosen to reflect the presence of active chromatin, whileassociation of dimethyl-H3K9 was used to reflect the presenceof a repressive chromatin region (4). We analyzed the chro-matin structure of the region encompassing the DMR thatharbors the H19-ICR insulator (Fig. 1) in an allele-specificmanner, since the H19-ICR insulator is functional only on thematernally inherited chromosome. PMEF were derived from�/H19del13 embryos to analyze the maternally inherited chro-mosome. The 13-kb deletion of H19 and the upstream region(19) on the paternal allele ensured that the information aboutthe chromatin structure is derived specifically from the mater-nal allele. Similarly, PMEF from H19del13/� embryos wereused to analyze the histone modifications on the paternal al-lele. The enrichment of the target sequences in the ChIP sam-ples was analyzed by real-time PCR, and � actin and Ig lociwere used as controls for the transcriptionally active and tran-scriptionally silent chromatin, respectively.
Chromatin structure on the maternally inherited chromo-some in PMEF. We focused our attention on the maternallyinherited insulator which is known to block the access of theenhancer to the Igf2 promoter. The region upstream to theDMR was practically devoid of activating histone modifica-tions dimethyl-H3K4 and acetyl-H3K9 (Fig. 3A and B). How-ever, the DMR and the region downstream of it, including theH19 gene, showed a significantly high enrichment in ChIP withdimethyl-H3K4. This correlates well with the active transcrip-tion of the H19 gene from the maternal chromosome. Acetyl-H3K9 presence was generally quite low through the entireregion except within the actively transcribing H19 gene (Fig.3B). The peak of activating histone modifications downstreamof the fourth CTCF site suggests recruitment of activatingfactors by the DMR which may be important for the activationof downstream H19. Consistent with this, deletion of the DMRleads to a variable degree of loss in H19 expression (32).Alternatively, the transition in chromatin structure may sug-gest the inability of the enhancer-initiated histone modifica-tions to spread past the insulator region, as observed for cHS4in minichromosome-based assays (40).
Analysis of the maternal DMR and its flanking regions forthe presence of dimethyl-H3K9 (Fig. 3C) exhibited an enrich-ment pattern complementary to the one observed with di-methyl-H3K4. The region upstream of the DMR was enrichedin the ChIP samples, while regions encompassing the DMRand downstream of it were practically devoid of this repressivechromatin mark. Together, these results are suggestive of aputative “chromatin barrier” organized by the DMR on thematernal chromosome.
Chromatin structure on the paternally inherited chromo-some in PMEF. Igf2 is expressed specifically from the paternalchromosome. On the paternal allele, neither the DMR nor theregions flanking it were enriched in ChIP with dimethyl-H3K4and acetyl-H3K9 (Fig. 3A and B) despite a high immunopre-cipitation efficiency as judged by the enrichment of � actin. Thesilent chromatin structure of the DMR and its flanking regionson the paternal chromosome was further confirmed by ChIPagainst dimethyl-H3K9 (Fig. 3C). Here the region upstream ofthe DMR was highly enriched, while the DMR and the regiondownstream of it had lower but significant enrichment, consis-tent with the known transcriptionally inactive status of the H19gene due to the presence of a silencer in the DMR responsiblefor MeCP2-dependent silencing of H19 (7). The degree ofenrichment was similar to that of Ig, a gene expected to betranscriptionally silent in PMEF. The nonfunctional insulatorregion of the paternal allele was also not associated with aputative “chromatin barrier” and was entirely distinct from thesame region on the maternal allele.
Importantly, our analysis of the paternal chromatin structureindicated that the activation of Igf2 by the downstream enhanc-ers is brought about despite a significantly large interveningregion (at least 10 kb) being in the repressed state based onhistone modification status. Not only the H19 gene and theDMR but also the region examined upstream of these is in arepressed state. Since the activation of paternal Igf2 by theenhancers is possible despite a large region being associatedwith silent chromatin modifications, it seems that the enhanceris not responsible for altering the intervening chromatin struc-ture in terms of histone modifications on the paternal chromo-some. Therefore, the differential domains of histone modifica-tion status, as seen on the maternal chromosome, cannot beattributed to stalled or paused enhancer-mediated chromatinalterations.
Allele-specific chromatin structure analysis in neonatal liver.The absence of the activating histone modifications from theregion upstream of the DMR on the paternal chromosome wasinteresting. It suggested the absence of a progressive spread ofactivating histone modifications, as might be expected if thetracking mechanism of enhancer-based activation is consid-ered. We decided to analyze the status of these modificationsin neonatal liver, a tissue which expresses both H19 and Igf2 atreasonably high levels due to promoter activation by well-characterized endodermal enhancers. As shown in Fig. 4A,dimethyl-H3K4 was associated with the maternal chromosomein the H19 gene and the region upstream of it, as in the PMEF.Also, in contrast to the PMEF chromatin, acetyl-H3K9 (Fig.4B) exhibited a high degree of association with H19 and itsupstream region in complete accordance with a higher level ofH19 expression in liver. The region upstream of the insulatorwas found not to be enriched on either the maternal or thepaternal chromosome upon ChIP with dimethyl-H3K4 andacetyl-H3K9, suggesting the absence of these modified his-tones from both the alleles. The absence of activating histonemodifications from the paternal chromosome both in PMEFand liver indicates that enhancer-mediated tracking, if opera-tional at this locus, does not involve the dimethyl-H3K4 andacetyl-H3K9 histones.
Chromatin structure in DMR-deleted chromosomes in PMEFand neonatal liver. Since the DMR on the paternal chromo-
3770 SINGH AND SRIVASTAVA MOL. CELL. BIOL.
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
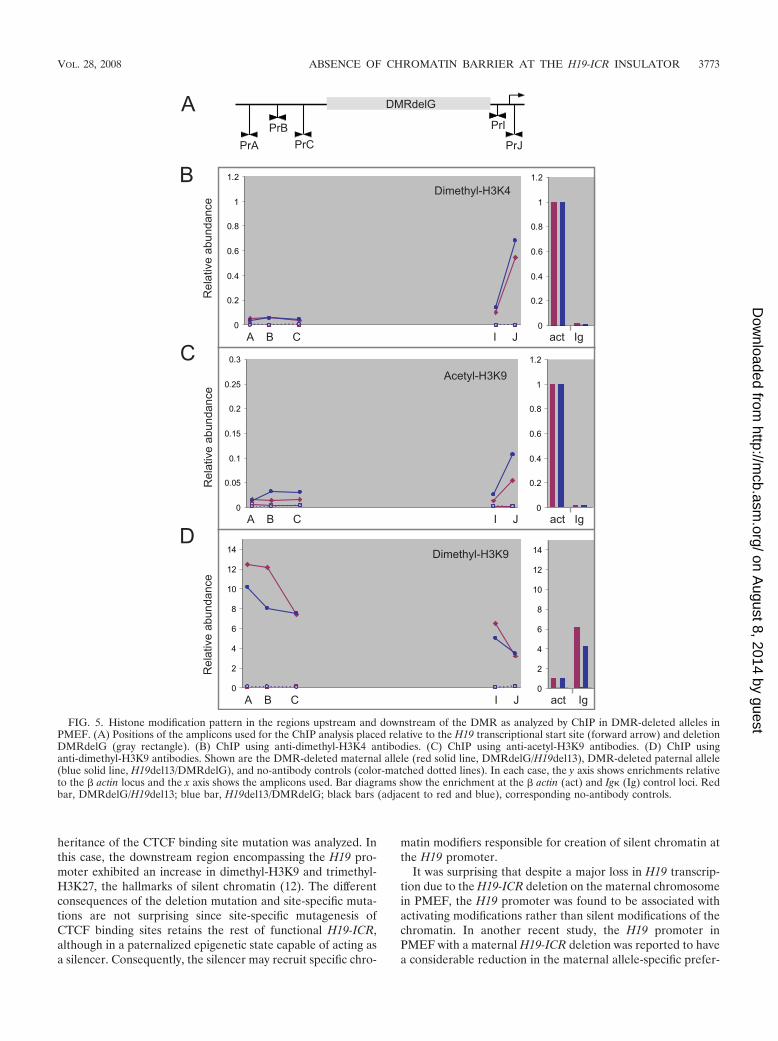
some is in an epigenetic state entirely different from that on thematernal chromosome, in order to ascertain the relevance ofthe observed “chromatin barrier” with regard to spread ofchromatin states, we considered it important to analyze thechromatin structure in cells that carried a germ line deletion ofthe functional insulator from the maternal allele. Analysis ofsuch PMEF (DMRdelG/H19del13) using three amplicons inthe region upstream of DMRdelG demonstrated that the re-
gion continued to be associated with repressive dimethyl-H3K9and devoid of the active chromatin marks dimethyl-H3K4 andacetyl-H3K9 (Fig. 5), like the wild-type maternal chromosome(Fig. 3). Also, activating histones were associated downstreamof the deletion, as in the wild type. The upstream silent chro-matin clearly did not spread into the H19 region. The DMRdeletion also had no pronounced effect on histone modificationstatus analyzed by ChIP in neonatal liver (Fig. 4). Deletion of
FIG. 3. Parental allele-specific histone modification pattern in the region encompassing the DMR as analyzed by ChIP in PMEF. (A) Positionsof the amplicons used for the ChIP analysis placed relative to the H19 transcriptional start site (forward arrow) and the DMR. (B) ChIP usinganti-dimethyl-H3K4 antibodies. (C) ChIP using anti-acetyl-H3K9 antibodies. (D) ChIP using anti-dimethyl-H3K9 antibodies. Shown are thematernal allele (red solid line, genotype �/H19del13), the paternal allele (blue solid line, genotype H19del13/�), and no-antibody controls(color-matched dotted lines). In each case, the y axis shows enrichments relative to the � actin locus and the x axis shows the amplicons used. Bardiagrams show the enrichment at the � actin (act) and Ig (Ig) control loci. Red bar, �/H19del13; blue bar, H19del13/�; black bars (adjacent tored and blue), corresponding no-antibody controls.
VOL. 28, 2008 ABSENCE OF CHROMATIN BARRIER AT THE H19-ICR INSULATOR 3771
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
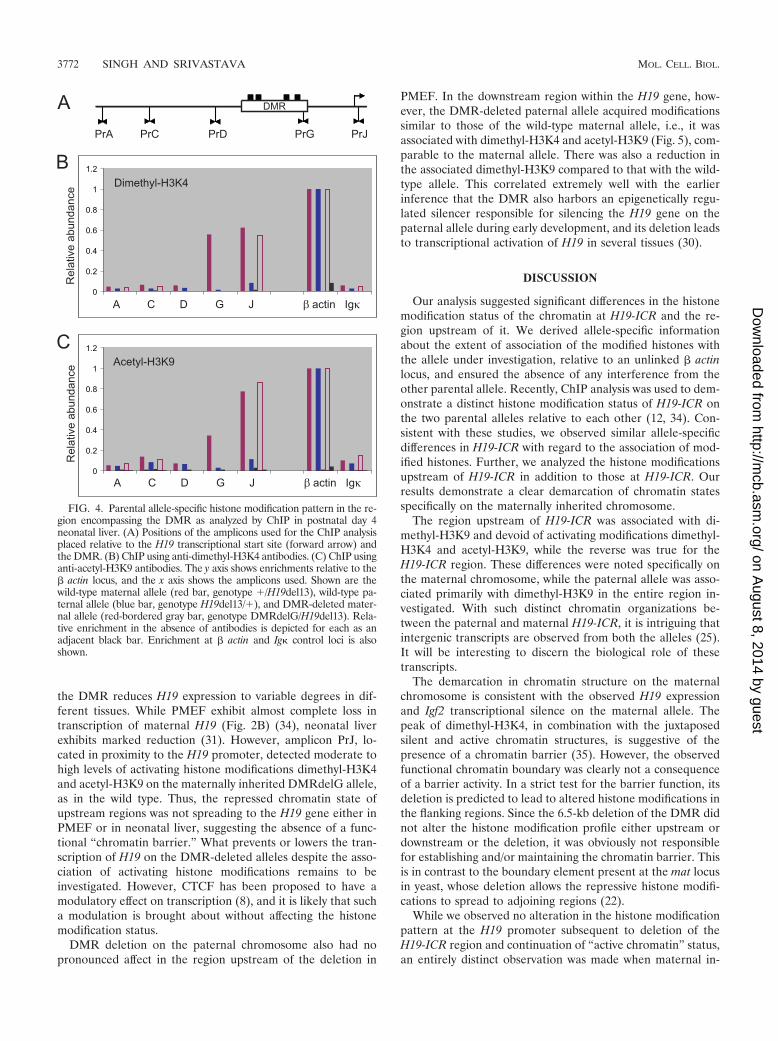
the DMR reduces H19 expression to variable degrees in dif-ferent tissues. While PMEF exhibit almost complete loss intranscription of maternal H19 (Fig. 2B) (34), neonatal liverexhibits marked reduction (31). However, amplicon PrJ, lo-cated in proximity to the H19 promoter, detected moderate tohigh levels of activating histone modifications dimethyl-H3K4and acetyl-H3K9 on the maternally inherited DMRdelG allele,as in the wild type. Thus, the repressed chromatin state ofupstream regions was not spreading to the H19 gene either inPMEF or in neonatal liver, suggesting the absence of a func-tional “chromatin barrier.” What prevents or lowers the tran-scription of H19 on the DMR-deleted alleles despite the asso-ciation of activating histone modifications remains to beinvestigated. However, CTCF has been proposed to have amodulatory effect on transcription (8), and it is likely that sucha modulation is brought about without affecting the histonemodification status.
DMR deletion on the paternal chromosome also had nopronounced affect in the region upstream of the deletion in
PMEF. In the downstream region within the H19 gene, how-ever, the DMR-deleted paternal allele acquired modificationssimilar to those of the wild-type maternal allele, i.e., it wasassociated with dimethyl-H3K4 and acetyl-H3K9 (Fig. 5), com-parable to the maternal allele. There was also a reduction inthe associated dimethyl-H3K9 compared to that with the wild-type allele. This correlated extremely well with the earlierinference that the DMR also harbors an epigenetically regu-lated silencer responsible for silencing the H19 gene on thepaternal allele during early development, and its deletion leadsto transcriptional activation of H19 in several tissues (30).
DISCUSSION
Our analysis suggested significant differences in the histonemodification status of the chromatin at H19-ICR and the re-gion upstream of it. We derived allele-specific informationabout the extent of association of the modified histones withthe allele under investigation, relative to an unlinked � actinlocus, and ensured the absence of any interference from theother parental allele. Recently, ChIP analysis was used to dem-onstrate a distinct histone modification status of H19-ICR onthe two parental alleles relative to each other (12, 34). Con-sistent with these studies, we observed similar allele-specificdifferences in H19-ICR with regard to the association of mod-ified histones. Further, we analyzed the histone modificationsupstream of H19-ICR in addition to those at H19-ICR. Ourresults demonstrate a clear demarcation of chromatin statesspecifically on the maternally inherited chromosome.
The region upstream of H19-ICR was associated with di-methyl-H3K9 and devoid of activating modifications dimethyl-H3K4 and acetyl-H3K9, while the reverse was true for theH19-ICR region. These differences were noted specifically onthe maternal chromosome, while the paternal allele was asso-ciated primarily with dimethyl-H3K9 in the entire region in-vestigated. With such distinct chromatin organizations be-tween the paternal and maternal H19-ICR, it is intriguing thatintergenic transcripts are observed from both the alleles (25).It will be interesting to discern the biological role of thesetranscripts.
The demarcation in chromatin structure on the maternalchromosome is consistent with the observed H19 expressionand Igf2 transcriptional silence on the maternal allele. Thepeak of dimethyl-H3K4, in combination with the juxtaposedsilent and active chromatin structures, is suggestive of thepresence of a chromatin barrier (35). However, the observedfunctional chromatin boundary was clearly not a consequenceof a barrier activity. In a strict test for the barrier function, itsdeletion is predicted to lead to altered histone modifications inthe flanking regions. Since the 6.5-kb deletion of the DMR didnot alter the histone modification profile either upstream ordownstream or the deletion, it was obviously not responsiblefor establishing and/or maintaining the chromatin barrier. Thisis in contrast to the boundary element present at the mat locusin yeast, whose deletion allows the repressive histone modifi-cations to spread to adjoining regions (22).
While we observed no alteration in the histone modificationpattern at the H19 promoter subsequent to deletion of theH19-ICR region and continuation of “active chromatin” status,an entirely distinct observation was made when maternal in-
FIG. 4. Parental allele-specific histone modification pattern in the re-gion encompassing the DMR as analyzed by ChIP in postnatal day 4neonatal liver. (A) Positions of the amplicons used for the ChIP analysisplaced relative to the H19 transcriptional start site (forward arrow) andthe DMR. (B) ChIP using anti-dimethyl-H3K4 antibodies. (C) ChIP usinganti-acetyl-H3K9 antibodies. The y axis shows enrichments relative to the� actin locus, and the x axis shows the amplicons used. Shown are thewild-type maternal allele (red bar, genotype �/H19del13), wild-type pa-ternal allele (blue bar, genotype H19del13/�), and DMR-deleted mater-nal allele (red-bordered gray bar, genotype DMRdelG/H19del13). Rela-tive enrichment in the absence of antibodies is depicted for each as anadjacent black bar. Enrichment at � actin and Ig control loci is alsoshown.
3772 SINGH AND SRIVASTAVA MOL. CELL. BIOL.
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
heritance of the CTCF binding site mutation was analyzed. Inthis case, the downstream region encompassing the H19 pro-moter exhibited an increase in dimethyl-H3K9 and trimethyl-H3K27, the hallmarks of silent chromatin (12). The differentconsequences of the deletion mutation and site-specific muta-tions are not surprising since site-specific mutagenesis ofCTCF binding sites retains the rest of functional H19-ICR,although in a paternalized epigenetic state capable of acting asa silencer. Consequently, the silencer may recruit specific chro-
matin modifiers responsible for creation of silent chromatin atthe H19 promoter.
It was surprising that despite a major loss in H19 transcrip-tion due to the H19-ICR deletion on the maternal chromosomein PMEF, the H19 promoter was found to be associated withactivating modifications rather than silent modifications of thechromatin. In another recent study, the H19 promoter inPMEF with a maternal H19-ICR deletion was reported to havea considerable reduction in the maternal allele-specific prefer-
FIG. 5. Histone modification pattern in the regions upstream and downstream of the DMR as analyzed by ChIP in DMR-deleted alleles inPMEF. (A) Positions of the amplicons used for the ChIP analysis placed relative to the H19 transcriptional start site (forward arrow) and deletionDMRdelG (gray rectangle). (B) ChIP using anti-dimethyl-H3K4 antibodies. (C) ChIP using anti-acetyl-H3K9 antibodies. (D) ChIP usinganti-dimethyl-H3K9 antibodies. Shown are the DMR-deleted maternal allele (red solid line, DMRdelG/H19del13), DMR-deleted paternal allele(blue solid line, H19del13/DMRdelG), and no-antibody controls (color-matched dotted lines). In each case, the y axis shows enrichments relativeto the � actin locus and the x axis shows the amplicons used. Bar diagrams show the enrichment at the � actin (act) and Ig (Ig) control loci. Redbar, DMRdelG/H19del13; blue bar, H19del13/DMRdelG; black bars (adjacent to red and blue), corresponding no-antibody controls.
VOL. 28, 2008 ABSENCE OF CHROMATIN BARRIER AT THE H19-ICR INSULATOR 3773
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

ential enrichment of activating histone modifications com-pared to that of the wild type (34). The observed discrepancyin the PMEF chromatin structure between the two studies maybe due to different experimental designs; our experiments re-ported the histone modification status quantitatively, specifi-cally from the allele under investigation, by ensuring that theother allele could not contribute toward the analysis. Promot-ers that are transcriptionally silent despite having activatinghistone modifications have been reported in some other stud-ies, and it has been suggested that these promoters are poisedfor transcription although not transcribing actively (24, 27). Itis noteworthy that deletion of H19-ICR on the paternal alleleleads to activation of H19 in several tissues, including neonatalliver, skeletal muscle, and heart (14, 30), but activation inPMEF is hardly detectable (Fig. 2B). However, deletion on thepaternal allele also led to the presence of activating modifica-tions on the H19 promoter in PMEF (Fig. 5). Thus, it appearsthat the H19 promoter, even when associated with activatinghistone modifications, is unable to transcribe effectively in theabsence of H19-ICR from either maternal or paternal alleles. Itis plausible that the mechanism for regulation of H19 expres-sion in PMEF is slightly different than in other tissues andrequires some additional regulatory factor dependent onH19-ICR.
Our analysis of the wild type and deletion mutants providesevidence that at the Igf2/H19 locus, silent and active chromatincan be maintained despite the absence of a chromatin barrierelement actively protecting the distinct domains, suggestingthat the silent chromatin state does not necessarily spread.Active and inactive chromatin have been reported to be inter-spersed without the ability to spread (23) at the Igf2r locus also.The generality of our observation regarding the absence of achromatin barrier at the Igf2/H19 locus needs to be tested inother genomic contexts. Since CTCF does not contribute tochromatin barrier function at H19-ICR, its presence at a largenumber of sites in the genome might suggest that for definingfunctionally distinct chromatin domains, the enhancer-block-ing function of CTCF and/or its ability to organize a higher-order chromatin structure, rather than barrier formation, ismore important.
With chromosome conformation capture (3C) analysis,CTCF has also been shown to organize the higher-order chro-matin structure, which may be relevant for gene regulation ina context-dependent manner (29). In accordance with this,the DMR organizes the higher-order chromatin structure atthe Igf2/H19 locus. Distinct looped domains, established by theinteraction of the DMR with other cis elements like DMR1,DMR2, and MAR3, influence enhancer-promoter interactionsand thus regulate Igf2 expression in an allele-specific manner(18, 21). Subsequent studies show that the H19-ICR insulatorcan effectively interfere with promoter-enhancer interactioneven at heterologous positions by forming associations with thepromoter and the enhancer (38). The 3C analysis providedevidence for the final stable complex formation by the en-hancer and the Igf2 promoter, but it could not explain theposition dependence of the insulator for preventing these in-teractions.
The position dependence of insulator activity intuitively sug-gests interference with some form of tracking signal initiated atthe enhancer. A few enhancers like E� and E� at the TCR�
and IgH loci, respectively, are responsible for creating a largechromatin domain with activating histone modifications (6, 20,28), in accordance with the proposed tracking model of en-hancer-based activation. In this context, the enhancers at theIgf2/H19 locus appeared different mechanistically. Our resultsdemonstrate that a significant part of the intervening region,between the active Igf2 promoter and enhancers at the Igf2/H19 locus, has a silent chromatin structure. A region of at least5 kb upstream of the H19-ICR insulator was observed to berich in repressive histones and devoid of activating histonemodifications. An identical pattern was seen on the wild-typeand DMR-deleted maternal and paternal alleles, i.e., irrespec-tive of the activation status of the Igf2 promoter. Thus, theenhancer-based activation of the Igf2 promoter was possibledespite a significant part of the intervening chromatin being ina silent state as judged by histone modification status. Surpris-ingly, the DMRs on both the maternal and paternal alleles dogenerate sense and antisense transcripts (25). In view of thebelief that the position dependence of insulator action mustinclude some form of tracking or linking signal for promoter-enhancer interaction, we infer that the tracking of an enhancer-based signal at the Igf2 locus does not involve chromatin mod-ifications investigated in our study, and hence the H19-ICRinsulator does not act by preventing this form of trackingsignal. What else may constitute a tracking signal and/or canexplain the position-dependent activity of the insulators re-mains to be investigated. It will be interesting to investigate ifenhancer-based loading of RNA polymerase on the Igf2 pro-moter is affected by the presence of the insulator, as observedfor the cHS4 insulator in minichromosome-based assays (40),despite the absence of histone modifications along the inter-vening chromatin domain.
Thus, the H19-ICR region, responsible for maintaining in-dependent expression profiles of Igf2 and H19, acts as thetransition point for distinct chromatin states on the maternalchromosome. However, there is no chromatin barrier elementassociated with the CTCF-dependent H19-ICR insulator.
ACKNOWLEDGMENTS
We thank Shirley Tilghman for the H19del13 mutant mice. We alsothank Karl Pfeifer and Sweety Shrimali for critically reading the manu-script, K. Natarajan for useful discussions, and Ramesh Yadav fortechnical assistance.
The work was supported by a grant from the Department of Bio-technology, India. Vikrant Singh is a recipient of SRF from the Coun-cil of Scientific and Industrial Research, India.
REFERENCES
1. Barski, A., S. Cuddapah, K. Cui, T. Y. Roh, D. E. Schones, Z. Wang, G. Wei,I. Chepelev, and K. Zhao. 2007. High-resolution profiling of histone methyl-ations in the human genome. Cell 129:823–837.
2. Bell, A. C., and G. Felsenfeld. 2000. Methylation of a CTCF-dependentboundary controls imprinted expression of the Igf2 gene. Nature 405:482–485.
3. Bender, M. A., R. Byron, T. Ragoczy, A. Telling, M. Bulger, and M. Groudine.2006. Flanking HS-62.5 and 3� HS1, and regions upstream of the LCR, are notrequired for beta-globin transcription. Blood 108:1395–1401.
4. Berger, S. L. 2007. The complex language of chromatin regulation duringtranscription. Nature 447:407–412.
5. Bulger, M., D. Schubeler, M. A. Bender, J. Hamilton, C. M. Farrell, R. C.Hardison, and M. Groudine. 2003. A complex chromatin landscape revealedby patterns of nuclease sensitivity and histone modification within the mousebeta-globin locus. Mol. Cell. Biol. 23:5234–5244.
6. Cobb, R. M., K. J. Oestreich, O. A. Osipovich, and E. M. Oltz. 2006.Accessibility control of V(D)J recombination. Adv. Immunol. 91:45–109.
7. Drewell, R. A., C. J. Goddard, J. O. Thomas, and M. A. Surani. 2002.
3774 SINGH AND SRIVASTAVA MOL. CELL. BIOL.
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from

Methylation-dependent silencing at the H19 imprinting control region byMeCP2. Nucleic Acids Res. 30:1139–1144.
8. Engel, N., J. L. Thorvaldsen, and M. S. Bartolomei. 2006. CTCF bindingsites promote transcription initiation and prevent DNA methylation on thematernal allele at the imprinted H19/Igf2 locus. Hum. Mol. Genet. 15:2945–2954.
9. Engel, N., A. G. West, G. Felsenfeld, and M. S. Bartolomei. 2004. Antago-nism between DNA hypermethylation and enhancer-blocking activity at theH19 DMD is uncovered by CpG mutations. Nat. Genet. 36:883–888.
10. Filippova, G. N., M. K. Cheng, J. M. Moore, J. P. Truong, Y. J. Hu, D. K.Nguyen, K. D. Tsuchiya, and C. M. Disteche. 2005. Boundaries betweenchromosomal domains of X inactivation and escape bind CTCF and lackCpG methylation during early development. Dev. Cell 8:31–42.
11. Gaszner, M., and G. Felsenfeld. 2006. Insulators: exploiting transcriptionaland epigenetic mechanisms. Nat. Rev. Genet. 7:703–713.
12. Han, L., D.-H. Lee, and P. E. Szabo. 2008. CTCF is the master organizer ofdomain-wide allele-specific chromatin at the H19/Igf2 imprinted region. Mol.Cell. Biol. 28:1124–1135.
13. Hark, A. T., C. J. Schoenherr, D. J. Katz, R. S. Ingram, J. M. Levorse, andS. M. Tilghman. 2000. CTCF mediates methylation-sensitive enhancer-blocking activity at the H19/Igf2 locus. Nature 405:486–489.
14. Kaffer, C. R., M. Srivastava, K. Y. Park, E. Ives, S. Hsieh, J. Batlle, A.Grinberg, S. P. Huang, and K. Pfeifer. 2000. A transcriptional insulator atthe imprinted H19/Igf2 locus. Genes Dev. 14:1908–1919.
15. Kim, A., C. M. Kiefer, and A. Dean. 2007. Distinctive signatures of histonemethylation in transcribed coding and noncoding human beta-globin se-quences. Mol. Cell. Biol. 27:1271–1279.
16. Kim, T. H., Z. K. Abdullaev, A. D. Smith, K. A. Ching, D. I. Loukinov, R. D.Green, M. Q. Zhang, V. V. Lobanenkov, and B. Ren. 2007. Analysis of thevertebrate insulator protein CTCF-binding sites in the human genome. Cell128:1231–1245.
17. Kuo, M. H., and C. D. Allis. 1999. In vivo cross-linking and immunoprecipi-tation for studying dynamic protein:DNA associations in a chromatin envi-ronment. Methods 19:425–433.
18. Kurukuti, S., V. K. Tiwari, G. Tavoosidana, E. Pugacheva, A. Murrell, Z.Zhao, V. Lobanenkov, W. Reik, and R. Ohlsson. 2006. CTCF binding at theH19 imprinting control region mediates maternally inherited higher-orderchromatin conformation to restrict enhancer access to Igf2. Proc. Natl. Acad.Sci. USA 103:10684–10689.
19. Leighton, P. A., R. S. Ingram, J. Eggenschwiler, A. Efstratiadis, and S. M.Tilghman. 1995. Disruption of imprinting caused by deletion of the H19gene region in mice. Nature 375:34–39.
20. Morshead, K. B., D. N. Ciccone, S. D. Taverna, C. D. Allis, and M. A.Oettinger. 2003. Antigen receptor loci poised for V(D)J rearrangement arebroadly associated with BRG1 and flanked by peaks of histone H3 dimeth-ylated at lysine 4. Proc. Natl. Acad. Sci. USA 100:11577–11582.
21. Murrell, A., S. Heeson, and W. Reik. 2004. Interaction between differentiallymethylated regions partitions the imprinted genes Igf2 and H19 into parent-specific chromatin loops. Nat. Genet. 36:889–893.
22. Noma, K., C. D. Allis, and S. I. Grewal. 2001. Transitions in distinct histoneH3 methylation patterns at the heterochromatin domain boundaries. Science293:1150–1155.
23. Regha, K., M. A. Sloane, R. Huang, F. M. Pauler, K. E. Warczok, B. Melikant,
M. Radolf, J. H. Martens, G. Schotta, T. Jenuwein, and D. P. Barlow. 2007.Active and repressive chromatin are interspersed without spreading in an im-printed gene cluster in the mammalian genome. Mol. Cell 27:353–366.
24. Roh, T. Y., S. Cuddapah, K. Cui, and K. Zhao. 2006. The genomic landscapeof histone modifications in human T cells. Proc. Natl. Acad. Sci. USA103:15782–15787.
25. Schoenfelder, S., G. Smits, P. Fraser, W. Reik, and R. Paro. 2007. Non-coding transcripts in the H19 imprinting control region mediate gene silenc-ing in transgenic Drosophila. EMBO Rep. 8:1068–1073.
26. Schoenherr, C. J., J. M. Levorse, and S. M. Tilghman. 2003. CTCF maintainsdifferential methylation at the Igf2/H19 locus. Nat. Genet. 33:66–69.
27. Schones, D. E., and K. Zhao. 2008. Genome-wide approaches to studyingchromatin modifications. Nat. Rev. Genet. 9:179–191.
28. Sen, R., and E. Oltz. 2006. Genetic and epigenetic regulation of IgH geneassembly. Curr. Opin. Immunol. 18:237–242.
29. Splinter, E., H. Heath, J. Kooren, R. J. Palstra, P. Klous, F. Grosveld, N.Galjart, and W. de Laat. 2006. CTCF mediates long-range chromatin loop-ing and local histone modification in the beta-globin locus. Genes Dev.20:2349–2354.
30. Srivastava, M., S. Hsieh, A. Grinberg, L. Williams-Simons, S. P. Huang, andK. Pfeifer. 2000. H19 and Igf2 monoallelic expression is regulated in twodistinct ways by a shared cis acting regulatory region upstream of H19. GenesDev. 14:1186–1195.
31. Thorvaldsen, J. L., K. L. Duran, and M. S. Bartolomei. 1998. Deletion of theH19 differentially methylated domain results in loss of imprinted expressionof H19 and Igf2. Genes Dev. 12:3693–3702.
32. Thorvaldsen, J. L., A. M. Fedoriw, S. Nguyen, and M. S. Bartolomei. 2006.Developmental profile of H19 differentially methylated domain (DMD) de-letion alleles reveals multiple roles of the DMD in regulating allelic expres-sion and DNA methylation at the imprinted H19/Igf2 locus. Mol. Cell. Biol.26:1245–1258.
33. Valenzuela, L., and R. T. Kamakaka. 2006. Chromatin insulators. Annu.Rev. Genet. 40:107–138.
34. Verona, R. I., J. L. Thorvaldsen, K. J. Reese, and M. S. Bartolomei. 2008.The transcriptional status but not the imprinting control region determinesallele-specific histone modifications at the imprinted H19 locus. Mol. Cell.Biol. 28:71–82.
35. West, A. G., and P. Fraser. 2005. Remote control of gene transcription.Hum. Mol. Genet. 14:R101–R111.
36. West, A. G., M. Gaszner, and G. Felsenfeld. 2002. Insulators: many functions,many mechanisms. Genes Dev. 16:271–288.
37. Xie, X., T. S. Mikkelsen, A. Gnirke, K. Lindblad-Toh, M. Kellis, and E. S.Lander. 2007. Systematic discovery of regulatory motifs in conserved regionsof the human genome, including thousands of CTCF insulator sites. Proc.Natl. Acad. Sci. USA 104:7145–7150.
38. Yoon, Y. S., S. Jeong, Q. Rong, K.-Y. Park, J. H. Chung, and K. Pfeifer. 2007.Analysis of the H19ICR insulator. Mol. Cell. Biol. 27:3499–3510.
39. Yusufzai, T. M., H. Tagami, Y. Nakatani, and G. Felsenfeld. 2004. CTCFtethers an insulator to subnuclear sites, suggesting shared insulator mecha-nisms across species. Mol. Cell 13:291–298.
40. Zhao, H., and A. Dean. 2004. An insulator blocks spreading of histoneacetylation and interferes with RNA polymerase II transfer between anenhancer and gene. Nucleic Acids Res. 32:4903–4919.
VOL. 28, 2008 ABSENCE OF CHROMATIN BARRIER AT THE H19-ICR INSULATOR 3775
on August 8, 2014 by guest
http://mcb.asm
.org/D
ownloaded from
Top Related
Copyright © 2022 FDOKUMEN

